effects of sedimentation on subtidal macroalgal assemblages: an experimental study from a...
TRANSCRIPT

LJournal of Experimental Marine Biology and Ecology,215 (1997) 269–288
Effects of sedimentation on subtidal macroalgal assemblages:an experimental study from a mediterranean rocky shore
*Laura Airoldi , Francesco Cinelli`Dipartimento di Scienze dell ’Uouoe dell’Ambiente, Universita di Pisa, Via A. Volta 6, I-56126 Pisa, Italy
Received 24 April 1996; revised 16 October 1996; accepted 13 November 1996
Abstract
The effects of sedimentation on the development and structure of macroalgal assemblages wereinvestigated from June 1992 to December 1993 on a rocky subtidal shore South of Livorno(Ligurian Sea). Experimental procedures were specifically designed to address: (1) which speciesrecruit in patches of bare rock under different sedimentation rates; (2) whether small-scalevariations of sediment deposition influence the local diversity of the assemblage; (3) whether theeffects of sedimentation are independent from the time of the year when succession begins.Depositional environment was manipulated in the field by means of transparent Plexiglas panelswhich on average reduced the amount of sediment sinking over the algal assemblage by 35%.Possible effects on the penetration of light and flow microenvironment were assessed. Small-scalevariations of sedimentation rates significantly influenced the local structure and diversity of thealgal assemblage. Such effects were particularly evident during the recolonisation of patches ofbare rock and were dependent upon the time at which succession was initiated. The erect algaewere the ones more affected by depositional environment and showed different responses to theexperimental reduction of sediment supplies. All the plots were quantitatively dominated by afilamentous turf mostly consisting of Polysiphonia setacea, whose growth was potentiallyenhanced by reduced sedimentation rates. At the end of the experiments, the overall diversity andevenness of the assemblage were lower in treatments with lessened sediment supplies. It issuggested that small-scale spatial variability of the depositional environment may affect thewithin-habitat diversity of algal assemblages either through direct effects on individual species oron their propagules, and through indirect effects mediated by competitive outcomes. 1997Elsevier Science B.V.
Keywords: Sedimentation; Diversity; Recruitment; Macroalgae; Turf; Subtidal; Rocky shore
*Corresponding author. Tel.: (139-50) 500943; fax: (139-50) 49694; e-mail: [email protected]
0022-0981/97/$17.00 1997 Elsevier Science B.V. All rights reservedPII S0022-0981( 96 )02770-0

270 L. Airoldi, F. Cinelli / J. Exp. Mar. Biol. Ecol. 215 (1997) 269 –288
1. Introduction
The worldwide increase of water turbidity and sediment load in coastal areas as aconsequence of anthropogenic activities has stirred interest into the impact of sedi-mentation upon rocky shore organisms. Such interest is reflected by an extensiveliterature (Moore, 1977; Seapy and Littler, 1982; Taylor, 1983; Rogers, 1990; Engledowand Bolton, 1994; Saiz Salinas and Isasi Urdangarin, 1994; Shaffer and Parks, 1994;Airoldi et al., 1995). However, so far, few quantitative experiments have examined inthe field the effects of this important physical process on the composition and structureof macroalgal communities (Neushul et al., 1976; Kendrick, 1991).
Sedimentation is believed to influence the distribution and diversity of seaweeds(Devinny and Volse, 1978; Robles, 1982; Stewart, 1983; D’Antonio, 1986; Shiel andFoster, 1986; Santos, 1993). However, most studies have been limited to descriptions ofspecies typically associated with sediment influenced habitats (Lewis, 1964; Mathieson,1967; Markham and Newroth, 1972; Mathieson and Fralick, 1972; Markham, 1973;Daly and Mathieson, 1977), and the causal mechanisms that shape the local structure ofalgal assemblages are poorly understood. The prevalent opinion is that high rates ofsediment deposition and movement are detrimental to the overall richness and diversityof the community through exclusion of less tolerant species (Daly and Mathieson, 1977;Little and Smith, 1980; Engledow and Bolton, 1994; Saiz Salinas and Isasi Urdangarin,1994). On the other hand sediment disturbance may promote diversity throughpreventing monopolisation of space by competitively dominant species and throughmaintaining patchiness (Foster, 1975; Taylor and Littler, 1982; Littler et al., 1983;McQuaid and Dower, 1990).
The purpose of the present study was to investigate the effects of sedimentation on thedevelopment and structure of a macroalgal assemblage growing on a rocky subtidalshore (south of Livorno, Western Mediterranean Sea). This tract of coast is characterizedby high sedimentation rates (Airoldi et al., 1996), and local patterns of sedimentdeposition are variable on a scale of meters (Airoldi, unpubl. data), potentiallyinfluencing the within-habitat diversity of the algal assemblage. Experimental pro-cedures, including manipulations of the depositional environment, were specificallydesigned to address (1) which species recruit on patches of bare rock under differentrates of sediment deposition, (2) whether small-scale variations of depositional environ-ment influence the local diversity of the assemblage and (3) whether the effects ofsedimentation are independent from the time of the year when succession begins. Thequantitative characterisation and the temporal variations of the depositional environmentof the study area and possible influences on patterns of abundance of the algal speciesare reported elsewhere (Airoldi et al., 1995, 1996).
2. Methods
2.1. Study site
The experiments were carried out from June 1992 to December 1993 on an exposed

L. Airoldi, F. Cinelli / J. Exp. Mar. Biol. Ecol. 215 (1997) 269 –288 271
rocky shore located about 10 km south of Livorno (Ligurian Sea). A detailed descriptionof the characteristics of the area can be found in Airoldi et al. (1995, 1996). Theexperimental site was a gently sloping ( | 308, 13–17 m in depth) field of sandstoneboulders colonised by an algal assemblage strongly dominated by a dense turf mostlyconsisting of Polysiphonia setacea Hollenberg (Airoldi et al., 1995). This species, whichoccurs on rocks and as epiphyte on other organisms, develops dense mats of uprightfilaments up to 1 cm high from prostrate branches firmly attached to the substratum byrhizoids (Verlaque, 1989). The calcareous alga Halimeda tuna (Ellis et Solander)Lamouroux represented the only abundant erect species. The abundance of grazers waslow, and these were mainly represented by specimens of the herbivorous fish Sarpasalpa (Linnaeus).
A total of 48 boulders, selected at random among 80 that had an horizontal top surfacelarger than 35 cm 3 40 cm, were used in the experiments described below. Theminimum distance between boulders was 1.5 m.
2.2. Manipulation of depositional environment
In the following experiments sediment deposition over the algal assemblage wasreduced in treatment plots by means of 34 cm 3 39 cm transparent Plexiglas panels.These were permanently anchored onto the top surface of the boulders, at a height ofapproximately 15 cm from the algal assemblage, by means of four stainless steel bars(diameter 4 mm) cemented with epoxy putty (Subcoat S, Veneziani) into holes producedin the rock by hammer and chisel (Fig. 1). The panels were fixed at the upper threadedextremities of the bars by means of nuts, in order to allow their removal and deploymentby divers during monitoring of the plots.
The effectiveness of the panels in reducing the amount of sediment sinking over thealgal assemblage was assessed from February to July 1993 by comparing sedimentationrates over six treatment and six control plots established at random in the area. Smallcylindrical sediment traps (diameter 20 mm and aspect: ratio 2) were deployed verticallystable on the horizontal top surface of the boulders by means of lead weights during 11periods ranging from 5 to 28 days. Every time, however, some traps were lost, probablyboth as a consequence of severe storms and vandals. Only eight periods, when at leastfour traps could be retrieved from both treatment and control plots, were included in theanalysis. Traps were closed with plastic stoppers before retrieval. The sediment collectedin each trap was filtered through ignited and preweighed 25 mm Whatman GF/F glassmicrofiber filters, rinsed with distilled water to remove salts and dried at 608C for 24 hbefore weighing. The data were analysed using a two-way mixed model ANOVA withpanel (treatment vs. control) as fixed factor and time (eight levels) as random factor. Thenumber of replicates for each combination of panel and time was balanced by choosingat random a constant smaller number of four traps from those retrieved (Underwood,1981). In this and in the following experiments, before running the analysis theassumption of homogeneity of variances was examined using Cochran’s C or Hartley’sF tests (Winer, 1971). Opportune transformations were applied when necessarymax
(Sokal and Rohlf, 1981). Student–Newman–Keuls tests were used for a posteriorimultiple comparisons of means.

272 L. Airoldi, F. Cinelli / J. Exp. Mar. Biol. Ecol. 215 (1997) 269 –288
Fig. 1. Panels used in treatment plots to reduce the amount of sediment sinking over the algal community on arocky subtidal shore South of Livorno (see text for further explanations).
To avoid shading of the algal assemblage as a consequence of the establishment of athin film of sediment and fouling over the transparent panels, these were carefullycleaned in situ at least once a week throughout the experiments, except on six occasionswhen bad weather conditions made diving unsafe for 14, 14, 17, 18, 20 and 22 daysrespectively. The effectiveness of such cleaning was tested by comparing the irradiancebelow the panels and in the control plots after a short ( , 10) or a long ( . 10) intervalof days had elapsed between two successive cleaning. Measurements were carried out bymeans of a quantometer (Biospherical Instrument Irradiancemeter) and were repeatedthroughout the research four times for each interval of days. Every time irradiance wasmeasured in six treatment and six control plots selected at random among those used forthe experiments described below. Data were analysed using a three-way mixed modelanalysis of variance with panel (treatment vs. control) and interval ( , 10 days vs. . 10days) as fixed factors and time (eight levels) as random variable nested in interval.
An additional set of measurements was taken in order to assess the flow mi-croenvironment near the rocky–seawater interface. The rate of dissolution of initiallydried (608C, 24 h) and weighed gypsum balls (diameter 2.3 mm, average weightincluding steel bars and plastic labels 11 g) was measured on six occasions from July1992 to June 1993 in the same six treatment and six control plots used to assess theeffectiveness of the panels in reducing rates of sediment deposition. Gypsum balls wereprepared following Gambi et al. (1989) and mounted on lead weights. Lead weights withtwo to four balls were positioned in the field for periods variable from 24 to 48 h.

L. Airoldi, F. Cinelli / J. Exp. Mar. Biol. Ecol. 215 (1997) 269 –288 273
Additional balls closed in a transparent, non hermetic box were used each time tocalibrate the measurements (Bailey-Brock, 1979). After recovery, gypsum balls wererinsed lightly in freshwater to remove salts, dried at 608C for 24 h and weighed. Thedissolution (weight loss) of each ball was converted to an estimate of flux (cm/sec)according to the following function obtained by Bailey-Brock (1979)
Flux 5 3.65 (Me /Mc 2 1)
where Me is the weight loss of each gypsum ball during field exposure and Mc is theweight loss of calibration balls. Fluxes estimated with balls mounted on the same leadweight were pooled, and the mean was used as the dependent variable. Data wereanalysed using a two-way mixed model ANOVA with panel (treatment vs. control) asfixed factor and time (six levels) as random factor.
2.3. Effects of sedimentation on macroalgae
A first experiment was set up in June 1992 in order to determine the effects ofreduced sedimentation rates on the composition and the structure of the algal assem-blage. Twelve 13 cm317 cm plots were established on the horizontal top surface of asmany boulders, permanently marked at their edges with epoxy putty and then assigned atrandom to treatments (reduced sediment deposition by means of Plexiglas panels) andcontrols (unmanipulated plots) with six replicates each. The abundance of algae wasmonitored monthly by visual surveys and photographs taken with a 1:7 extension tubeon a Nikonos III underwater camera equipped with an electronic flash and Fujichrome100 Asa films. In the laboratory the transparencies were projected onto a grid of 100equally spaced dots, and the percent cover values of each species or group of specieswas expressed as the number of hits after correction for those points which laid overunidentifiable substratum. Species present in quadrats but not abundant enough to bescored from intercepts with the grid were assigned an arbitrary cover value of 0.1%. Thephotographic sampling did not allow quantification of the abundance of those specimenscovered by the algal turf. A major characterisation of the composition and the structureof the assemblage was carried out at the end of the experiment, in June 1993, bydestructively sampling the plots with hammer and chisel. The samples were analysed inthe laboratory as described by Airoldi et al. (1995), and the abundance of species wasexpressed in terms of biomass (dry weight) per unit area. For each sample theShannon–Weaver diversity index (H9) and the species evenness (J) were calculated. Thedata from the photographic sampling were analysed using Student’s t-tests on thepercent cover values of each species in each plot averaged over the sampling dates. Thedata obtained from the destructive sampling (number of species, biomass, H9, J) wereanalysed using Student’s t-tests.
A second 232 factorial experiment was conducted to examine the effects ofsedimentation on patterns of development of the algal assemblage following completeremoval of the biota at different times of the year. In June 1992, 12 experimental plots(16 cm320 cm) were scraped clean of resident organisms by means of hammer andchisel and then assigned at random to treatments (reduced sediment deposition by means

274 L. Airoldi, F. Cinelli / J. Exp. Mar. Biol. Ecol. 215 (1997) 269 –288
of Plexiglas panels) and controls (unmanipulated plots) with six replicates each. Thegreat anfractuosity of the substratum made cleaning operation rather difficult, andprostrate axes of those specimens living in the deepest crevices were probably notcompletely removed. A second series of cleared areas was produced in December 1992.The time at which succession was initiated was not replicated within season: anydifference among the plots produced in different times may, therefore, reflect chancetemporal variation (in either patterns of recruitment and environmental factors) ratherthan predictable seasonal trends. Patterns of colonisation of cleared areas exposed todifferent sedimentation rates were sampled monthly for 1 yr by photographs and visualobservations as described above. The abundance of algae was estimated over a smallersurface (13 cm317 cm) in order to avoid edge effects. The reproductive phenology ofthe dominant alga Polysiphonia setacea was also studied throughout the research bybiweekly to monthly samples, in order to assess its reproductive status. Percent coverdata were analysed using a two-way ANOVA on the average values of each species overtime; both sedimentation (reduced vs. natural) and time of beginning (June vs.December) were fixed factors. The plots produced in June were also sampled destruc-tively after 1 yr, and the data analysed as described in the above experiment.
3. Results
3.1. Effectiveness of panels
On average, the panels used in the experiments significantly reduced the amount ofsediment sinking on the algal assemblage by about 35% (Fig. 2, Table 1). Althoughsedimentation rates varied significantly among sampling periods, the differencesobserved between treatment and control plots generally persisted over time, as indicatedby the lack of significant panel3time interaction (Table 1).
The weekly cleaning of the transparent panels prevented the establishment of a film ofsediment and fouling and the consequent shading of the organisms below. In fact, when10 days at maximum had elapsed between cleaning, the irradiance measured below thepanels in treatment plots was not significantly different from that measured in thecontrols (Fig. 3 Table 2). On the contrary, as expected, when bad weather conditionsprevented cleaning procedures for periods longer than 10 days, the irradiance below thepanels significantly decreased down to 37% of that measured in the control plots. Themagnitude of such differences, however, sharply lessened when environmental ir-radiance was low (Fig. 3 Table 2). Moreover, it may be reasonably presumed that thedifferences between treatment and control plots arose gradually, becoming relevant onlyat the end of the intervals.
The presence of the panels over the algal assemblage did not affect the flowmicroenvironment at the rocky–seawater interface. The rates of dissolution of thegypsum balls were, in fact, similar in treatment and control plots during all thesamplings (Fig. 4 Table 3).

L. Airoldi, F. Cinelli / J. Exp. Mar. Biol. Ecol. 215 (1997) 269 –288 275
Fig. 2. Effectiveness of panels in reducing the sedimentation rate on the algal assemblage. Sedimentation rates22 21(mg m d ) over the experimental plots (treatment panels vs controls) were measured by means of sediment
traps during eight periods from February to July 1993. Data are means (11 S.E.) over four replicates.
3.2. Structure of the algal assemblage
3.2.1. Photographic samplingThroughout the first experiment, both treatment and control plots were quantitatively
dominated by a dense filamentous turf (mainly consisting of Polysiphonia setacea)whose percent cover values were never lower than 78% (Fig. 5 Table 4). The erectspecies were mostly represented by the articulated calcareous alga Halimeda tuna (Fig.5). This species was generally less abundant in plots with reduced sedimentation rates,but differences observed were not statistically significant (Table 4). Dictyota dichotoma(Hudson) Lamouroux and Giffordia sp. were also present in spring and summer monthsespecially in plots with reduced sedimentation rates (Fig. 5). However, their distribution
Table 1Analysis of the effectiveness of panels in reducing rates of sediment deposition on the algal assemblage
Source of variation df MS F P Cochran’s C test(transformation)aPanel (5P) 1 4.296 11.18 0.012 C50.292 P.0.05aTime (5T) 7 3.841 9.775 ,0.001 (logarithmic)
P3T 7 0.384 0.977 0.458Error 48 0.393
Sedimentation rates in treatment (with panel) and control plots were compared at different times of the year.Panel is a fixed factor, while time is random.a Significant P values (P,0.05).
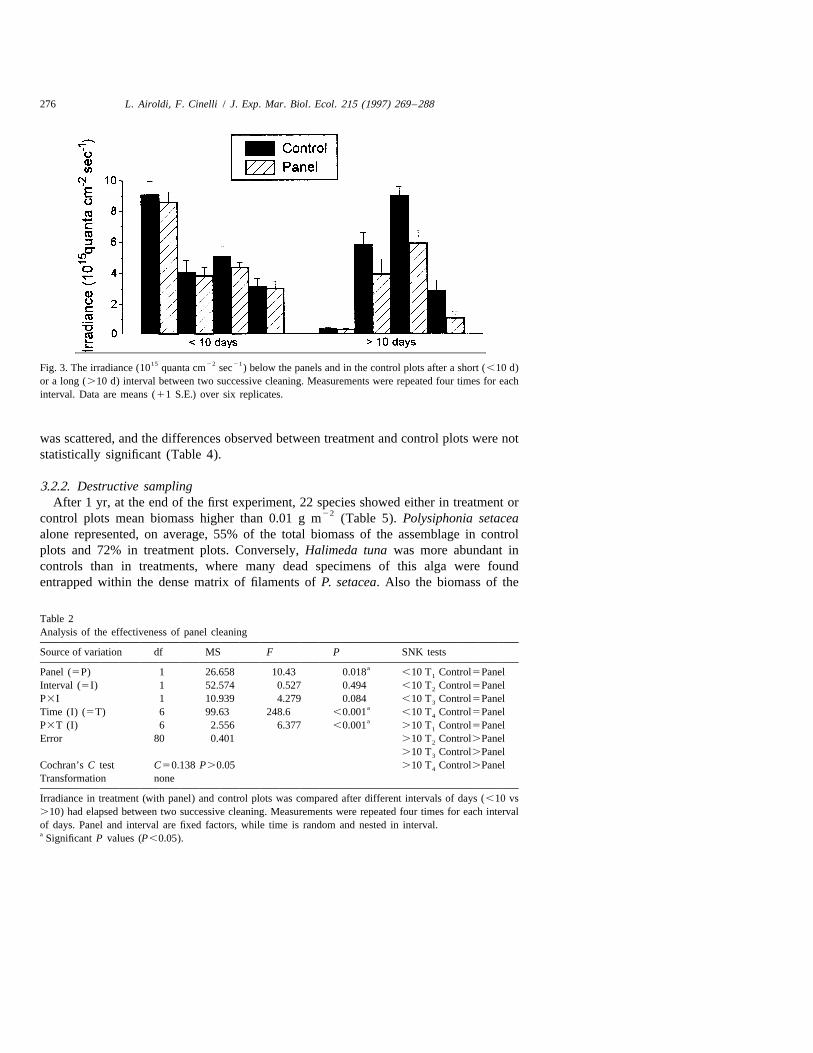
276 L. Airoldi, F. Cinelli / J. Exp. Mar. Biol. Ecol. 215 (1997) 269 –288
15 22 21Fig. 3. The irradiance (10 quanta cm sec ) below the panels and in the control plots after a short (,10 d)or a long (.10 d) interval between two successive cleaning. Measurements were repeated four times for eachinterval. Data are means (11 S.E.) over six replicates.
was scattered, and the differences observed between treatment and control plots were notstatistically significant (Table 4).
3.2.2. Destructive samplingAfter 1 yr, at the end of the first experiment, 22 species showed either in treatment or
22control plots mean biomass higher than 0.01 g m (Table 5). Polysiphonia setaceaalone represented, on average, 55% of the total biomass of the assemblage in controlplots and 72% in treatment plots. Conversely, Halimeda tuna was more abundant incontrols than in treatments, where many dead specimens of this alga were foundentrapped within the dense matrix of filaments of P. setacea. Also the biomass of the
Table 2Analysis of the effectiveness of panel cleaning
Source of variation df MS F P SNK testsaPanel (5P) 1 26.658 10.43 0.018 ,10 T Control5Panel1
Interval (5I) 1 52.574 0.527 0.494 ,10 T Control5Panel2
P3I 1 10.939 4.279 0.084 ,10 T Control5Panel3aTime (I) (5T) 6 99.63 248.6 ,0.001 ,10 T Control5Panel4aP3T (I) 6 2.556 6.377 ,0.001 .10 T Control5Panel1
Error 80 0.401 .10 T Control.Panel2
.10 T Control.Panel3
Cochran’s C test C50.138 P.0.05 .10 T Control.Panel4
Transformation none
Irradiance in treatment (with panel) and control plots was compared after different intervals of days (,10 vs.10) had elapsed between two successive cleaning. Measurements were repeated four times for each intervalof days. Panel and interval are fixed factors, while time is random and nested in interval.a Significant P values (P,0.05).

L. Airoldi, F. Cinelli / J. Exp. Mar. Biol. Ecol. 215 (1997) 269 –288 277
21Fig. 4. Intensity of flow microenvironment (cm sec ) below the panels and in the control plots estimated bymeans of gypsum balls on six occasions from July 1992 to June 1993. Data are means (11 S.E.) over sixreplicates (see text for further explanations).
prostrate species, which were commonly covered by the turf, was generally higher incontrol plots. Overall, however, the variations of depositional environment had nosignificant effects on the composition and structure of the assemblage, although a trendtoward a decrease of species diversity and evenness was observed in treatment plots(Table 6).
3.3. Patterns of recruitment
3.3.1. Photographic samplingIn the recruitment experiment, encrusting coralline algae were among the first
colonists of the plots produced in both periods. The recruitment of these speciesproceeded rapidly, and they reached mean percent covers up to 20% in both treatmentsand controls within two months from the beginning of the experiment. After that time,
Table 3Analysis of the effects of panels on water fluxes near the bottom surface
Source of variation df MS F P Cochran’s C test(transformation)
Panel (5P) 1 0.203 6.483 0.052 C50.186 P.0.05aTime (5T) 5 1.895 60.19 ,0.001 (logarithmic)
P3T 5 0.031 0.996 0.434Error 36 0.031
The rate of dissolution of gypsum balls in treatment (with panel) and control plots was compared at differenttimes of the year. In each plot fluxes were estimated with 2–4 balls, and the mean was used as the dependentvariable. Panel is a fixed factor while time is random.a Significant P values (P,0.05) are shown in bold type.

278 L. Airoldi, F. Cinelli / J. Exp. Mar. Biol. Ecol. 215 (1997) 269 –288
Fig. 5. Effects of small-scale spatial variation of sediment deposition on patterns of abundance of macroalgae.Data are mean percent covers (11 S.E.) on six replicates. Treatments, reduced sediment deposition; controls,natural sediment deposition.
however, they were progressively overgrown by the filamentous turf, and it was nolonger possible to follow their development until the final destructive sampling.
Patterns of recruitment of the filamentous turf were significantly influenced by thetime of beginning of the experiment (Fig. 6 Table 7). The turf rapidly colonised bothtreatment and control plots, reaching percent covers above 80% after 8 months in theplots produced in June and after 6 months in those produced in December, butrestoration of a thick mat took longer (Airoldi, pers. obs.). In both periods cleared spacewas filled by lateral vegetative encroachment of prostrate axes from the surrounding turfand, to a lesser extent, by regrowth of outlived axes not completely removed from thesubstratum (see Section 2). In spite of the frequent sampling, P. setacea was never seenin a reproductive state, suggesting that recruitment from settled propagules did notcontribute to the recovery of this species.
Table 4Analysis of the effects of sedimentation (reduced vs natural) on patterns of abundance of macroalgae
Algal species t df P Hartley’s F test Transformationmax
Filamentous turf 0.53 10 0.607 F 52.53 P.0.05 nonemax
Halimeda tuna 1.59 10 0.143 F 51.07 P.0.05 nonemax
Dictyota dichotoma 1.69 10 0.122 F 514.7 0.01,P,0.05 nonemax
Giffordia sp. 1.33 10 0.217 F 5` P,0.01 nonemax
Student’s t-tests were performed on the average percent cover values of each species trough time (see text forfurther explanations).

L. Airoldi, F. Cinelli / J. Exp. Mar. Biol. Ecol. 215 (1997) 269 –288 279
Table 522Mean biomass (g m ) and standard deviations of the most abundant macroalgae in treatment (reduced
sedimentation rates) and control (natural sedimentation rates) plots at the end of the experiment to determinethe effects of sedimentation on the composition and structure of the assemblage
Species Treatments Controls
M S.D. M S.D.
Prostrate layerEncrusting corallines 24.19 19.63 28.59 17.99Peyssonnelia bornetii Boudouresque et Denizot 1 2.44 17.03 35.47Peyssonnelia harveyana P.L. et H.M. Crouan ex J. Agardh 13.55 12.38 10.18 11.88Peyssonnelia polymorpha (Zanardini) Schmitz 4.11 10.06Peyssonnelia rubra (Greville) J. Agardh 0.68 0.61 1.25 1.14
TurfAcrothamnion preissii (Sonder) Wollaston 0.08 0.19 0.74 1.66Botryocladia boergesenii J. Feldmann 0.04 0.06 1
Chondria mairei G. Feldmann 0.07 0.14 1
Chordariacea unidentified 1 0.03 0.06Dasyopsis plana (C. Agardh) Zanardini 0.04 0.1Derbesia tenuissima (De Notaris) P.L. et H.M. Crouan 0.17 0.4Feldmannophycus rayssiae (J. et G. Feldmann) Augier etBoudouresque 1 0.47 0.79
¨Halopteris filicina (Grateloup) Kutzing 0.03 0.08Jania rubens (Linnaeus) Lamouroux 0.01 0.07 0.07Laurencia obtusa (Linnaeus) Lamouroux 0.07 0.11 0.45 0.71Laurencia pelagosae (Shiffner) Ercegovic 0.19 0.46Polysiphonia setacea (Hollenberg) 137.11 34.99 108.04 21.96
Erect layerDictyota dichotoma (Hudson) Lamouroux 0.22 0.19 0.34 0.34Flabellia petiolata (Turra) Nizamuddin 1 1.22 2.92Galaxaura oblongata (Ellis et Solander) Lamouroux 1 0.04 0.1Giffordia sp. 1 0.08 0.19Halimeda tuna (Ellis et Solander) Lamouroux 11.24 5.79 25.82 17.56
22Only species that showed a mean biomass higher than 0.01 g m either in treatments or controls are reported.22
1 indicates mean biomass ,0.01 g m ; M, mean; S.D., standard deviation (n56).
Table 6Mean values and standard deviations of the total number of species (N), total biomass (B), Shannon–Weaverdiversity index (H9) and species evenness (J) in treatment (reduced sedimentation rates) and control (naturalsedimentation rates) plots at the end of the experiment to determine the effects of sedimentation on thecomposition and structure of the algal assemblage
Variable Treatment (S.D.) Control (S.D.) t df P Hartley’s F testmax
N 13 (1.26) 13.17 (1.72) 0.191 10 0.852 F 51.854 P.0.05max
B 188.64 (26.42) 196.86 (50.17) 0.355 10 0.729 F 53.605 P.0.05max
H9 1.216 (0.4) 1.679 (0.36) 2.106 10 0.061 F 51.228 P.0.05max
J 0.33 (0.11) 0.45 (0.08) 2.115 10 0.06 F 51.791 P.0.05max
Untransformed data were analysed using Student’s t-tests.

280 L. Airoldi, F. Cinelli / J. Exp. Mar. Biol. Ecol. 215 (1997) 269 –288
Fig. 6. Effects of small-scale spatial variation of sediment deposition on patterns of recruitment of macroalgaeon patches of bare rock cleared in (A) June 1992 and (B) December 1992. Data are mean percent covers (11S.E.) on six replicates. Symbols as in Fig. 5.

L. Airoldi, F. Cinelli / J. Exp. Mar. Biol. Ecol. 215 (1997) 269 –288 281
Table 7Analysis of the effects of sedimentation (reduced vs natural) and time of beginning of the experiment (June1992 vs December 1992) on patterns of recruitment of the algal assemblage
Source of variation df MS F P Cochran’s C test (transformation)
Filamentous turfSedimentation (5S) 1 129.9 2.338 0.142 C50.496 P.0.05
aBeginning (5B) 1 561.6 10.11 0.047 (none)S3B 1 40.81 0.734 0.402Error 20 55.55
Halimeda tuna C50.687 P,0.01Sedimentation (5S) 1 1.743 1.680 0.209 (none)Beginning (5B) 1 3.657 3.526 0.075S3B 1 2.249 1.037 0.156Error 20 1.037
Dictyota dichotoma C50.503 P.0.05Sedimentation (5S) 1 0.106 0.018 0.895 (none)Beginning (5B) 1 11.84 1.996 0.173S3B 1 11.31 1.907 0.182Error 20 5.929
Padina pavonica C50.758 P,0.01Sedimentation (5S) 1 0.371 0.465 0.503 (none)Beginning (5B) 1 0.721 0.903 0.353S3B 1 0.354 0.443 0.513Error 20 0.798
Acetabularia acetabulum C50.567 P.0.05aSedimentation (5S) 1 35.12 5.97 0.023 (none)aBeginning (5B) 1 73.38 12.49 0.002
S3B 1 25.22 4.293 0.051Error 20 5.875
Giffordia sp. C50.510 P.0.05aSedimentation (5S) 1 81.04 18.82 ,0.001 (none)aBeginning (5B) 1 30.58 7.104 0.014aS3B 1 28.52 6.627 0.018
Error 20 4.304
ANOVAs were performed on the average percent cover values of each species through time (see text forfurther explanations).a Significant P values (P,0.05).
Recruitment of Halimeda tuna was very low both in treatment and control plots, andduring the experiment this species never reached mean percent covers higher than 3.4%(Fig. 6). A tendency towards a major abundance of H. tuna in control plots produced inJune was observed, but such differences were not statistically significant (Table 7).
Dictyota dichotoma and Padina pavonica (Linnaeus) Lamouroux showed a markedseasonal pattern, with a peak of abundance in late spring and summer (Fig. 6). Thesespecies colonised both treatment and control plots, reaching percent covers sensiblyhigher than in the adult surrounding assemblage, as also suggested by the comparison

282 L. Airoldi, F. Cinelli / J. Exp. Mar. Biol. Ecol. 215 (1997) 269 –288
with results of the first experiment (Fig. 5). No significant differences were observedamong treatments and controls (Table 7).
Also Acetabularia acetabulum (Linnaeus) Silva and Giffordia sp. showed markedtemporal patterns, colonising the cleared areas only during spring and summer months(Fig. 6). However, patterns of recruitment of these species were influenced by bothvariations of sedimentation rates and time of beginning of the experiment (Table 7). Theoutcomes of such effects were different for the two species. A. acetabulum, in fact,recruited especially in control plots produced in June, while Giffordia sp. exclusivelycolonised treatment plots produced in both periods, reaching high percent cover only inthose produced in June (Fig. 6).
3.3.2. Destructive samplingAfter 1 yr of succession, both treatment and control plots produced in June were
quantitatively dominated by the filamentous turf, which ranged from 85 to 94% of the22total biomass. Only ten other species showed mean biomass higher than 0.01 g m
(Table 8). Such a low number of species was mainly due to the negligible presence ofprostrate forms, which were almost exclusively represented by encrusting corallines withlow biomass. On the contrary, the abundance of erect species, especially Acetabulariaacetabulum, Dictyota dichotoma and Padina pavonica, was high if compared with thatof the natural assemblage in the study area (see controls of the first experiment and datareported in Airoldi et al., 1995).
The influence of depositional environment on patterns of succession of the algalassemblage was evident. The experimental reduction of sedimentation rates, in fact, not
Table 822Mean biomass (g m ) and standard deviations of the most abundant macroalgae in treatment (reduced
sedimentation rates) and control (natural sedimentation rates) plots at the end of the experiment to determinethe effects of sedimentation on the patterns of recruitment of the assemblage on cleared substrata
Species Treatments Controls
M S.D. M S.D.
Prostrate layerEncrusting corallines 0.74 0.84 1.10 0.68
TurfAcrothamnion preissii (Sonder) Wollaston 0.36 0.36 0.37 0.46Chondria mairei G. Feldmann 0.04 0.08 0.03 0.05Laurencia obtusa (Linnaeus) Lamouroux 0.04 0.1 0.44 0.56Polysiphonia setacea (Hollenberg) 102.21 63.16 85.54 32.02Sphacelaria cirrosa (Roth) C. Agardh 1 0.14 0.31
Erect layerAcetabularia acetabulum (Linnaeus) Silva 0.27 0.25 1.10 0.94Dictyota dichotoma (Hudson) Lamouroux 1.72 1.96 7.16 9.45Giffordia sp. 1.84 3.81 0.40 0.88Halimeda tuna (Ellis et Solander) Lamouroux 1.08 1.11 3.26 3.99Padina pavonica (Linnaeus) Lamouroux 1 1.17 1.32
22Only species that showed a mean biomass higher than 0.01 g m either in treatments or controls are reported.22
1 indicates mean biomasses ,0.01 g m ; M, mean; S.D., standard deviation (n56).

L. Airoldi, F. Cinelli / J. Exp. Mar. Biol. Ecol. 215 (1997) 269 –288 283
Table 9Mean values and standard deviations of the total number of species (N), total biomass (B), Shannon–Weaverdiversity index (H9) and species evenness (J) in the experimental plots (treatment panels vs controls) at theend of the experiment to determine the effects of sedimentation on patterns of recruitment of the algalassemblage
Variable Treatment (S.D.) Control (S.D.) t df P Hartley’s F testmax
N 9.33 (1.21) 10.5 (0.84) 1.941 10 0.081 F 52.095 P.0.05max
B 109.12 (64.77) 100.69 (39.74) 0.272 10 0.791 F 52.657 P.0.05maxaH9 0.44 (0.296) 0.82 (0.26) 2.342 10 0.041 F 51.291 P.0.05maxaJ 0.14 (0.08) 0.24 (0.076) 2.307 10 0.043 F 51.221 P.0.05max
Untransformed data were analysed using Student’s t-tests.a Significant P values (P,0.05).
only affected significantly the abundance of Acetabularia acetabulum, Dictyota dich-otoma and Padina pavonica (whose total biomass was lower in treatment than in controlplots, Student’s t-test, t52.24, P,0.05), but also significantly decreased the overalldiversity and species evenness of the assemblage (Table 9). Such decrease of diversitywas partly determined by the increase of abundance of Polysiphonia setacea, whosemean biomass was 19.5% higher in plots with reduced sedimentation rates. However, theeffects of sedimentation on the development of this species were not detected assignificant due to the high variability among replicate plots (Student’s t-test, t50.576,P.0.05).
4. Discussion
4.1. Manipulation of depositional environment
Experimental manipulations of the subtidal depositional environment are still few(Neushul et al., 1976; Rogers, 1983; Kendrick, 1991). The commonest approach used insuch experiments is the increase of sediment deposition through addition of knownamounts of sediment to the experimental plots. Reduction of sedimentation rates hasbeen attempted less frequently, through removal of sediment by water motion. In bothcases, the effectiveness of the manipulation is clearly dependent on the frequency ofapplication or removal of sediment as well as on environmental conditions (e.g., naturalsedimentation rates, water flow, composition of the biological community). However, noattempt has ever been made to quantify the real effectiveness of the method employed.Moreover, the duration and intensity of the treatment may greatly affect the outcome ofthe experiments: prolonged sediment smothering, in fact, may have quite differenteffects than short term deep burial by sediments (Marszalek, 1981; Rogers, 1990). Inthis study transparent panels were used to reduce natural sediment deposition over smallportions of the algal assemblage, reducing the sedimentary flux by 35% on average. Theeffectiveness of the panels was confirmed by the visual survey of the plots: after a fewmonths from the beginning of the experiments, in fact, the algal assemblage below thepanels looked clearer of sediment than in the control plots. This suggests that, although

284 L. Airoldi, F. Cinelli / J. Exp. Mar. Biol. Ecol. 215 (1997) 269 –288
the short-term reduction of sediment deposition below the panels was not quantitativelyvery high, its persistence induced a long-term impact on the algal assemblage.
A major shortcoming of manipulative field experiments in which structures such ascages, barriers or panels are used is the risk of introducing artifacts that may bias theresults (Connell, 1974; Underwood, 1986; Peterson and Black, 1994). This concern isgenerally evaluated by integrating into the experimental design a procedural controlwhich should reproduce the method used in treatments but not affect the variable ofinterest (but see Johnson, 1992). In the present study, however, we could not find anyadequate control to unconfound potential artifacts: partial panels, in fact, do not mimicaccurately the presence of the entire panels, while introducing a partial manipulation ofsedimentation rates. As an alternative, we directly quantified in the field the effects onthose variables (i.e., penetration of light and flow microenvironment) that could be mostaffected by the presence of panels. Results have shown that the accurate and constantmaintenance of the panels kept the effects on irradiance down to a level whichpresumably had no biological relevance. Effects on the flow microenvironment were notdetected either. Artifacts on grazing pressure were not assessed, but these may bereasonably considered irrelevant due to the scarcity of grazers in the area. Possibleconfounding effects on the inputs of propagules were not quantified either. However,dispersal of propagules generally occurs through lateral transport by bottom currents(Santelices, 1990; Amsler et al., 1992), which were not significantly affected by thepresence of panels.
4.2. Influence of depositional environment
Small-scale variation of the depositional environment significantly influenced the localstructure and diversity of the algal assemblage. Such effects were especially evidentduring the colonisation of patches of bare rock and were dependent on the time ofbeginning of the succession. The erect algae (i.e., those species developing above thefilamentous turf) were the most affected by variations of the depositional environmentand showed different responses to the experimental reduction of sediment deposition.The filamentous turf quantitatively dominated all the plots, but its growth was potentiallyenhanced by reduced sediment loads. At the end of the experiments the overall diversityand evenness of the assemblage was lower in treatments subjected to reducedsedimentation rates.
The above results provide support to the hypothesis that small-scale spatial variabilityof depositional environments may affect the within-habitat diversity of algal assem-blages. In the study area such influence was exerted particularly through control ofabundance of dominant species rather than control of the overall composition andspecies richness of the assemblage. This suggests not only direct effects on individualplants or their propagules, but also indirect effects through mediation of competitiveoutcomes.
Direct effects of sediment on algae in rocky habitats include smothering and scouring.The morphological and life-history attributes that might confer adaptive ability to algaein environments with sediment deposition and movement have been discussed by severalauthors (Daly and Mathieson, 1977; Stewart, 1983; Littler et al., 1983; Airoldi et al.,

L. Airoldi, F. Cinelli / J. Exp. Mar. Biol. Ecol. 215 (1997) 269 –288 285
1995); these include tough thalli, the regeneration of upright axes from basal thallusparts that can physically resist or tolerate smothering and scouring, opportunistic cyclesof reproduction and growth and the capacity to reproduce vegetatively, which removesdependence upon spore attachment to buried substrata and allows a rapid recovery fromdamage. Adaptations of this kind can be found in all the major space occupiers of thestudy area (Airoldi et al., 1995), and are particularly noticeable in the dominant speciesPolysiphonia setacea. Our results, however, suggest that rates of growth of the turf werenegatively affected by sediment smothering. The abundance of the turf, in fact, tended toincrease under reduced sedimentation rates, although this pattern could be perceivedonly from detailed measurements of the biomass at the end of the experiments. Higherrates of growth of filamentous turfs with lower sediment supplies have been alsoobserved by Adey and Goertemiller (1987) in the Caribbean and by Kendrick (1991) inthe Galapagos. Sediment may be considered a structural constituent of algal turfs(Scoffin, 1970; Stewart, 1983; Airoldi et al., 1995), and the characteristics of deposition-al environment might directly affect their patterns of growth. Experimental inves-tigations, including manipulations of the quality and size spectrum of sedimentingmaterial as well as estimates of the turf biomass, are actually in progress to verify thishypothesis.
The results of clearing experiments suggest a possible role of competitive interactionsas a structuring factor for the algal assemblage, at least within the time scale considered.The removal of the resident organisms, in fact, enhanced the recruitment of some erectspecies which were sparse at the study site. Such effects were particularly evident in theplots produced in June and exposed to natural sedimentation rates. A possibleexplanation is that the algal turf could preclude the development of other species byexploitation of space and/or interference interactions through overgrowing, and thatlocal stress and disturbance are necessary for the persistence of some components of theassemblage. Variations of sedimentation rates might, therefore, influence indirectly thedistribution of some species and the local structure and diversity of the assemblagethrough affecting patterns of growth of the algal turf.
Similarly to observations in other sediment-impacted areas (Robles, 1982; D’Antonio,1986), grazing pressure is low during most of the year at our study site (Airoldi, pers.obs.), and disturbance is mainly related to physical factors (i.e., sediment scouring andwave action) which vary greatly through time (Airoldi et al., 1996). The effects ofmanipulation of the depositional environment varied among patches of cleared sub-stratum produced at different times of the year, possibly reflecting differences inpropagule availability and post-settlement growth and mortality of organisms (Sousa,1984; Breitburg, 1985; Menge, 1991). In particular, variations of sedimentation ratesaffected recruitment of species especially in quadrats produced in June, while no effectswere observed in plots cleared in December. In the study area maximal sedimentdeposition and movement generally occurs in the autumn, while in the summer theassemblage is free from sediment burial and scour (Airoldi et al., 1996). Predictabletemporal patterns in the disturbance regime, together with the modes of recolonisation ofcleared areas could, therefore, be important determinants of the effects of sedimentdeposition on the overall diversity of the assemblage.
In conclusion, the experimental data of the present study support the hypothesis that

286 L. Airoldi, F. Cinelli / J. Exp. Mar. Biol. Ecol. 215 (1997) 269 –288
small-scale spatial variations in the depositional environment may affect the develop-ment and local diversity of algal assemblages; the outcomes, however, are dependent ona variety of factors, including disturbance regime, successional patterns and morphologi-cal and life-history characteristics of the important space occupiers. It is suggested thatthe causal mechanisms that shape algal communities in sediment impacted areas relyeither on direct effects on individual plants or their propagules and on indirect effectsmediated by competitive interactions. Multifactorial experiments, including manipulationof both physical and biological factors, will help elucidate this hypothesis.
Acknowledgments
We wish to thank Marco Abbiati, Luigi Piazzi, Salvatore Portella, Stefano Acunto andthe Guardia di Finanza of Livorno for collaboration in the field work, Fabio Rindi forhelp with the determination of algae, Lisandro Benedetti-Cecchi and Gary A. Kendrickfor help with the statistical analysis and valuable comments on the manuscript, and twoanonymous referees for helpful revision of the manuscript. This study was completed asa part of the doctoral thesis research of Laura Airoldi at the University of Genova, Italy.
References
Adey, W.H. and T. Goertemiller, 1987. Coral reef algal turfs: master producers in nutrient poor seas.Phycologia, Vol. 26, pp. 374–386.
Airoldi, L., F. Rindi and F. Cinelli, 1995. Structure, seasonal dynamics and reproductive phenology of afilamentous turf assemblage on a sediment influenced, rocky subtidal shore. Bot. Mar., Vol. 38, pp.227–237.
Airoldi, L., M. Fabiano and F. Cinelli, 1996. Sediment deposition and movement over a turf assemblage in ashallow rocky coastal area of the Ligurian Sea. Mar. Ecol. Prog. Ser., Vol. 133, pp. 241–251.
Amsler, C.D., D.C. Reed and M. Neushul, 1992. The microclimate inhabited by macroalgal propagules. Br.Phycol. J., Vol. 27, pp. 253–270.
Bailey-Brock, J., 1979. Sediment trapping by chaetopterid polichaetes on a Hawaiian fringing reef. J. Mar.Res., Vol. 37, pp. 643–656.
Breitburg, D.L., 1985. Development of a subtidal epibenthic community: factors affecting species compositionand the mechanisms of succession. Oecologia, Vol. 65, pp. 173–184.
Connell, J.H., 1974. Ecology: field experiments in marine ecology. In, Experimental marine biology, edited byR.N. Mariscal, Academic Press, New York, pp. 21–54.
Daly, M.A. and A.C. Mathieson, 1977. The effect of sand movement on intertidal seaweeds and selectedinvertebrates at Bound Rock, New Hampshire, USA. Mar. Biol., Vol. 43, pp. 45–55.
D’Antonio, C.M., 1986. Role of sand in the domination of hard substrata by the intertidal alga Rhodomelalarix. Mar. Ecol. Prog. Ser., Vol. 27, pp. 263–275.
Devinny, J.S. and L.A. Volse, 1978. The effect of sediments on the development of Macrocystis pyriferagametophytes. Mar. Biol., Vol. 48, pp. 343–348.
Engledow, H.R. and J.J. Bolton, 1994. Seaweed a-diversity within the lower eulittoral zone in Namibia: theeffects of wave action, sand inundation, mussels and limpets. Bot. Mar., Vol. 37, pp. 267–276.
Foster, M.S., 1975. Algal succession in a Macrocystis pirifera forest. Mar. Biol., Vol. 32, pp. 313–329.Gambi, M.C., M.C. Buia, E. Casola and M. Scardi, 1989. Estimates of water movement in Posidonia oceanica
beds: a first approach. In, International workshop on Posidonia beds, edited by C.F. Boudouresque, A.Meinesz, E. Fresi and V. Gravez, GIS Posidonie, France, Vol. 2, pp. 101–112.

L. Airoldi, F. Cinelli / J. Exp. Mar. Biol. Ecol. 215 (1997) 269 –288 287
Johnson, L.E., 1992. Potential and peril of field experimentation: the use of copper to manipulate molluscanherbivores. J. Exp. Mar. Biol. Ecol., Vol. 160, pp. 251–262.
Kendrick, G.A., 1991. Recruitment of coralline crusts and filamentous turf algae in the Galapagos archipelago:effect of simulated scour, erosion and accretion. J. Exp. Mar. Biol. Ecol., Vol. 147, pp. 47–63.
Lewis, J.R., 1964. The ecology of rocky shores. English University Press, London.Little, C. and L.P. Smith, 1980. Vertical zonation on rocky shores in the Severn Estuary. Est. Coast. Shelf. Sci.,
Vol. 2, pp. 651–669.Littler, M.M., D.R. Martz and D.S. Littler, 1983. Effects of recurrent sand deposition on rocky intertidal
organisms: importance of substrate heterogeneity in a fluctuating environment. Mar. Ecol. Prog. Ser., Vol.11, pp. 129–139.
Markham, J.W., 1973. Observation on the ecology of Laminaria sinclarii on three northern Oregon beaches. J.Phycol., Vol. 9, pp. 336–341.
Markham, J.W. and P.R. Newroth, 1972. Observations on the ecology of Gymnogrongus linearis and relatedspecies. Intl. Seaweed Sym., Vol. 7, pp. 127–130.
Marszalek, D.S., 1981. Impact of dredging on a subtropical reef community, Southeast Florida, U.S.A. Proc.4th Int. Coral Reef Congr., Vol.1, pp. 147–153.
Mathieson, A.C., 1967. Morphology and life history of Phaeostrophion irregulare S. et G. Nova Hedwigia,Vol. 13, pp. 293–318.
Mathieson, A.C. and R.A. Fralick, 1972. Investigations of New England marine algae V. The algal vegetationof the Hampton-Seabrook Estuary and the open coast near Hampton, New Hampshire. Rhodora,Vol. 74, pp.406–435.
McQuaid, C. and K.M. Dower, 1990. Enhancement of habitat heterogeneity and species richness on rockyshores inundated by sand. Oecologia, Vol. 84, pp. 142–144.
Menge, B.A., 1991. Relative importance of recruitment and other causes of variation in rocky intertidalcommunity structure. J. Exp. Mar. Biol. Ecol., Vol. 146, pp. 69–100.
Moore, P.G., 1977. Inorganic particulate suspensions in the sea and their effects on marine animals. Oceanogr.Mar. Biol. Annu. Rev., Vol. 15, pp. 225–363.
Neushul, M., M.S. Foster, C.A. Coon, J.W. Woessner and B.W.W. Harger, 1976. An in situ study of recruitmentgrowth and survival of subtidal marine algae: techniques and preliminary results. J. Phycol., Vol. 12, pp.397–408.
Peterson, C.H. and R. Black, 1994. An experimentalist’s challenge: when artifacts of intervention interact withtreatments. Mar. Ecol. Prog. Ser., Vol. 111, pp. 289–297.
Robles, C., 1982. Disturbance and predation in an assemblage of herbivorous diptera and algae on rockyshores. Oecologia, Vol. 54, pp. 23–31.
Rogers, C.S., 1983. Sublethal and lethal effects of sediments applied to common Caribbean reef corals in thefield. Mar. Pollut. Bull., Vol. 14, pp. 378–382.
Rogers, C.S., 1990. Responses of coral and reef organisms to sedimentation. Mar. Ecol. Prog. Ser.,Vol. 62, pp.185–202.
Saiz Salinas, J.I. and I.I. Isasi Urdangarin, 1994. Response of sublittoral hard substrate invertebrates toestuarine sedimentation in the outer harbour of Bilbao (N. Spain). P.S.Z.N.I.: Mar. Ecol., Vol. 15, pp.105–131.
Santelices, B., 1990. Patterns of reproduction, dispersal and recruitment of seaweeds. Oceanogr. Mar. Biol.Annu. Rev., Vol. 28, pp. 177–276.
Santos, R., 1993. A multivariate study of biotic and abiotic relationships in a subtidal algal stand. Mar. Ecol.Prog. Ser., Vol. 94, pp. 181–190.
Scoffin, T.P., 1970. The trapping and binding of subtidal carbonate sediments by marine vegetation in Biminilagoon, Bahamas. J. Sedim. Petrol., Vol. 40, pp. 249–273.
Seapy, R.P. and M.M. Littler, 1982. Population and species diversity fluctuations in a rocky intertidalcommunity relative to severe aerial exposure and sediment burial. Mar. Biol., Vol. 71, pp. 87–96.
Shaffer, J.A. and D.S. Parks, 1994. Seasonal variations in and observations of landslide impacts on the algalcomposition of a Puget Sound nearshore kelp forest. Bot. Mar., Vol. 37, pp. 315–323.
Shiel, D.R. and M.S. Foster, 1986. The structure of subtidal algal stands in temperate waters. Oceanogr. Mar.Biol. Annu. Rev., Vol. 24, pp. 265–307.
Sokal, R.R. and F.J. Rohlf, 1981. Biometry, W.H. Freeman, San Francisco, CA.

288 L. Airoldi, F. Cinelli / J. Exp. Mar. Biol. Ecol. 215 (1997) 269 –288
Sousa, W.P., 1984. Intertidal mosaics: patch size, propagule availability, and spatially variable patterns ofsuccession. Ecology, Vol. 65, pp. 1918–1935.
Stewart, J.G., 1983. Fluctuations in the quantity of sediment trapped among algal thalli on intertidal rockplatforms in southern California. J. Exp. Mar. Biol. Ecol., Vol. 73, pp. 205–211.
Taylor, B.D., 1983. Sediment yields in coastal southern California, American Society of Civil Engineers, J.Hydraul. Div., Vol. 109, pp. 109–111.
Taylor, P.R. and M.M. Littler, 1982. The roles of compensatory mortality, physical disturbance, and substrateretention in the development and organization of a sand-influenced, rocky-intertidal community. Ecology,Vol. 63, pp. 135–146.
Underwood, A.J., 1981. Techniques of analysis of variance in experimental marine biology and ecology.Oceanogr. Mar. Biol. Annu. Rev., Vol. 19, pp. 513–605.
Underwood, A.J., 1986. The analysis of competition by field experiments. In, Community ecology: pattern andprocess, edited by J. Kikkawa and D.J. Anderson, Blackwell Scientific, London, pp. 240–268.
` `Verlaque, M., 1989. Contribution a la flore des algues marines de Mediterrauee: Especes rares ou nouvellesˆpour les cotes Francaises. Bot. Mar., Vol. 32, pp. 101–113.
Winer, B.J., 1971. Statistical principles in experimental designs. McGraw-Hill Kogakusha, Tokyo.