effects of a gulf stream warm-core ring on summer seabird distribution in the northwest atlantic...
TRANSCRIPT

Effects of a Gulf Stream Warm-Core Ring on Summer Seabird Distribution in the NorthwestAtlantic OceanAuthor(s): J. Christopher HaneySource: Limnology and Oceanography, Vol. 32, No. 3 (May, 1987), pp. 665-673Published by: American Society of Limnology and OceanographyStable URL: http://www.jstor.org/stable/2836755 .
Accessed: 17/06/2014 21:02
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
American Society of Limnology and Oceanography is collaborating with JSTOR to digitize, preserve andextend access to Limnology and Oceanography.
http://www.jstor.org
This content downloaded from 62.122.79.81 on Tue, 17 Jun 2014 21:02:55 PMAll use subject to JSTOR Terms and Conditions

NOTES
Limnol. Oceanogr., 32(3), 1987, 665-673 C 1987, by the American Society of Limnology and Oceanography, Inc.
Effects of a Gulf Stream warm-core ring on summer seabird distribution in the northwest Atlantic Ocean'
Abstract-Species composition, distribution, and abundance of seabirds at a Gulf Stream warm- core ring differed from water masses of the north- west Atlantic slope, shelf, and Sargasso Sea dur- ing August 1984. Mean seabird abundance within the 2-month-old ring (12.3 birds km-2) was sim- ilar to Slope Water (13.0 birds kmi-2). Mean bio- mass of the seabird assemblage within the ring (0.85 kg km-2 wet wt) was intermediate between biomass values in Sargasso Sea and Slope Water. The seabird assemblage within the ring was com- prised of one nekton-feeding species (Puffinus Iherminieri) displaced from the parent water mass (Gulf Stream), and the storm-petrels Oceanites oceanicus and Oceanodroma leucorhoa common in adjacent Slope Water.
Rings formed from western boundary currents provide unique opportunities for examining the effects of altered environ- ments on species and communities of oceanic plankton and nekton. Ring forma- tion initiates a meso-scale invasion of one oceanic community by another, with con- current intercommunity interaction (Wiebe et al. 1976). As anomalous features in the surrounding water masses, rings provide sharp contrasts in physical and biological regimes, allowing temporal and spatial changes in organism distribution, biomass, or species composition to be studied (Ortner et al. 1978, 1979; Hitchcock et al. 1985; Romanetal. 1985).
I here describe seabird distribution, abundance, biomass, and species compo- sition at a Gulf Stream warm-core ring. Warm-core rings (WCRs) are formed in northwest Atlantic Slope Water when an- ticyclonic meanders of the Gulf Stream de-
1 This work was supported by fellowships from the University of Georgia with logistic support from NSF grant OCE 84-15963 to L. S. Shapiro.
tach from the main current body. WCRs have central cores derived from the Sargas- so Sea and are surrounded by a remnant of the Gulf Stream rotating clockwise at 0.5- 2.0 m s- I (Joyce et al. 1 983). Newly formed WCRs are typically 100-200 km in diam- eter, average 1,000 m in depth, and migrate to the southwest at 3-5 km d- 1 (Joyce and Wiebe 1983). The hydrographic and bio- logical characteristics of WCRs are at first similar to the Sargasso Sea, but may be sub- sequently altered by interactions with shelf, slope, or Gulf Stream water. The seabird assemblage at WCR 84-51 was compared to assemblages in slope, shelf, and Sargasso Sea water masses. Results from these com- parisons are examined in the context of WCR biology and discussed in light of re- cent hypotheses on the factors thought to influence seabird interactions with marine environments.
I thank L. S. Shapiro, W. K.-Bellows, T. Boynton, and the crew of the RV Cape Hat- teras for technical support, and S. McIntosh for drafting the figures. Comments from two anonymous reviewers improved earlier drafts of the manuscript.
Counts of seabirds were made in the northwest Atlantic Ocean between 360 and 43?N during a research cruise from Bigelow Laboratory for Ocean Science to Bermuda and back (Fig. 1). Counts were conducted in shelf (SH), slope (SL), Sargasso Sea (SS), and WCR water masses between 21 and 31 August 1984.
Hydrographic stations consisting of 200-m CTD casts were made between 0500 and 0700 hours, before the in-transit sea- bird counts. Biological sampling at each sta- tion included vertical pump sampling of chlorophyll a fluorescence, size-fractionat- ed estimates of phytoplankton biomass, nu-
665
This content downloaded from 62.122.79.81 on Tue, 17 Jun 2014 21:02:55 PMAll use subject to JSTOR Terms and Conditions
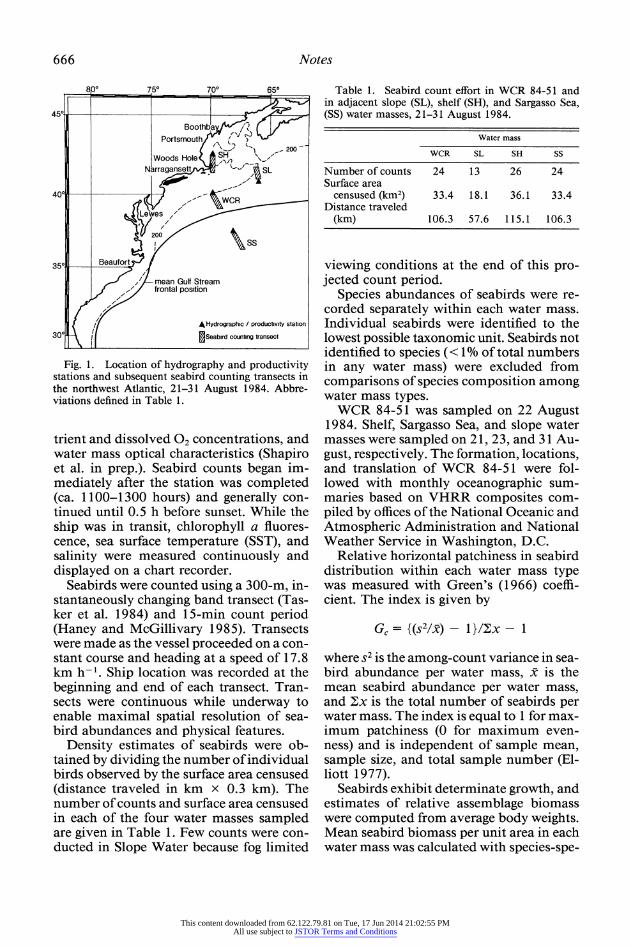
666 Notes
8O? 750 700 650
45r Boothba i '
40 t ;
35? Beauf P o
mean Gulf Stream
X X ~~~~frontal position
Z // ~~~~~~~~AHydrographic / productivity station
300 4\ I SSeabird countNng asansect
Fig. 1. Location of hydrography and productivity stations and subsequent seabird counting transects in the northwest Atlantic, 21-31 August 1984. Abbre- viations defined in Table 1.
trient and dissolved 2 concentrations, and water mass optical characteristics (Shapiro et al. in prep.). Seabird counts began im- mediately after the station was completed (ca. 1100-1300 hours) and generally con- tinued until 0.5 h before sunset. While the ship was in transit, chlorophyll a fluores- cence, sea surface temperature (SST), and salinity were measured continuously and displayed on a chart recorder.
Seabirds were counted using a 300-m, in- stantaneously changing band transect (Tas- ker et al. 1984) and 15-min count period (Haney and McGillivary 1985). Transects wera were vessel proceeded on a con- stant course and heading at a speed of 17.8 km h-1. Ship location was recorded at the beginning and end of each transect. Tran- sects were continuous while underway to enable maximal spatial resolution of sea- bird abundances and physical features.
Density estimates of seabirds were ob- tained by dividing the number of individual birds observed by the surface area censused (distance traveled in km x 0.3 km). The number of counts and surface area censused in each of the four water masses sampled are given in Table 1. Few counts were con- ducted in Slope Water because fog limited
Table 1. Seabird count effort in WCR 84-51 and in adjacent slope (SL), shelf (SH), and Sargasso Sea, (SS) water masses, 21-31 August 1984.
Water mass
WCR SL SH SS
Number of counts 24 13 26 24 Surface area
censused (kM2) 33.4 18.1 36.1 33.4 Distance traveled
(km) 106.3 57.6 115.1 106.3
viewing conditions at the end of this pro- jected count period.
Species abundances of seabirds were re- corded separately within each water mass. Individual seabirds were identified to the lowest possible taxonomic unit. Seabirds not identified to species (< 1% of total numbers in any water mass) were excluded from comparisons of species composition among water mass types.
WCR 84-51 was sampled on 22 August 1984. Shelf, Sargasso Sea, and slope water masses were sampled on 21, 23, and 31 Au- gust, respectively. The formation, locations, and translation of WCR 84-51 were fol- lowed with monthly oceanographic sum- maries based on VHRR composites com- piled by offices of the National Oceanic and Atmospheric Administration and National Weather Service in Washington, D.C.
Relative horizontal patchiness in seabird distribution within each water mass type was measured with Green's (1966) coeffi- cient. The index is given by
G= {(S2/j) - 1}/ x - 1
where S2 is the among-count variance in sea- bird abundance per water mass, x is the mean seabird abundance per water mass, and Xx is the total number of seabirds per water mass. The index is equal to 1 for max- imum patchiness (0 for maximum even- ness) and is independent of sample mean, sample size, and total sample number (El- liott 1977).
Seabirds exhibit determinate growth, and estimates of relative assemblage biomass were computed from average body weights. Mean seabird biomass per unit area in each water mass was calculated with species-spe-
This content downloaded from 62.122.79.81 on Tue, 17 Jun 2014 21:02:55 PMAll use subject to JSTOR Terms and Conditions

Notes 667
200 -200~
-S 1
S40 >SX~~4
35 _ X - _
30
June July 80 75 70 65 60 55 80 75 70 65 60 55
200F 200
35 - _ __ _
____ ~~30 _ _
August 25 Septemb~r
Fig. 2. Evolution and translation of WCR 84-51 during summer and fall 1984.
cific body weights in kilograms (Cramp and Simmons 1977, 1983; Clapp et al. 1982, 1983):
n (t F,Mi)/A where Fi and MA are, respectively, the total numbers and body mass in kilograms of the ith species, n is the number of species ob- served within the water mass, and A is the surface area censused per water mass (Table 1).
WCR 84-51 was the 51st anticyclonic eddy formed during 1984 in the northwest Atlantic Ocean. WCR 84-51 originated from a large Gulf Stream meander near 39000'N, 65?30'W during June 1984 (Fig. 2). Ring diameter measured about 275 by 320 km at the time of detachment on 9 July.
By August, WCR 84-51 had translated 250 km to the northwest near Hydrogra- pher's Canyon. The ring interacted with the Gulf Stream around 1 August and was re- duced in diameter from 240 km on 26 July to 85 km on 31 August. During September the ring translated 155 km west, seaward of the continental shelf edge (200-m isobath).
By October, WCR 84-51 was entrained by the Gulf Stream.
The WCR hydrographic station (Fig. 3) was located at the northern edge of the ring. The seabird counting transect crossed the western third of the ring on a south-south- east course between 40?00'N, 70?00'W and 39007'N, 69017'W. Evidence of ring inter- action with Shelf or Slope Water was indi- cated by surface salinities of 33.9-34.6%oo (Table 2) during the transect (cf. Joyce and Wiebe 1983; Joyce and Stalcup 1984). A 20-25-m surface mixed layer (Fig. 3) was characterized by temperatures of 24?-250C at the ring's northern edge, increasing to 27.6?C near ring center, and decreasing slightly to 27.2?C near the ring's southern boundary.
Mean seabird abundance within the ring was 12.3 birds km-2, only slightly less than mean abundances in Slope Water (13.0 birds km-2; Fig. 4). Seabird abundance within the ring was 68 times greater than in the Sar- gasso Sea, but less than the mean abundance in Shelf Water (19.4 birds km-2).
Differences in mean seabird biomass among water masses were more pro- nounced. Biomass of seabirds in the ring
This content downloaded from 62.122.79.81 on Tue, 17 Jun 2014 21:02:55 PMAll use subject to JSTOR Terms and Conditions

668 Notes
0 5 10 15 20 25 30 35 0 5 10 15. 20 25 30 35
20 32
40 6
60 _
T ~~~~~~~28 8 0_ _ _ _ _ _ _ _ _ _ _
160
0 ~~~~~~~~~~~~~~~92 120Sea_______ Gulf wtrea a Core Rin lope 1ater
5 15 10 20 25 30 35 Co 5 10 15 20 25 30 35__
0 24 32 _
48 T64
72 96 T
96 r128 _
1 20 160
144 196 ____ ~~~~Shelf Wate Sarasso, Sea I
Temperature, T, in 0C and Salinity,S, in%o
Fig. 3. Vertical temperature-salinity profiles (?200 m)in WCR 84-51 compared to slope, shelf,and Sargasso Sea water during August 1984.
was 0.85 kg wet wt km-2-a biomass that is only 25% of that found in Slope Water and 10% of that in Shelf Water. Compared with the Sargasso Sea, however, biomass in the ring was 45 times greater. Although sea- bird biomass in WCR water was less than in Slope Water, mean abundances were sim- ilar in the two water types. Smaller bodied seabirds contributed proportionately more to total abundance within the ring (Fig. 5).
Seabird patchiness also differed among water masses (Fig. 6). The relative strength of aggregation was greatest in the ring where the G, value (0.217) was 6-10 times greater than in other water masses. Seabirds were
more evenly distributed within Shelf Water (G, = 0.020), followed by Sargasso Sea (G, = 0.025), and Slope Water (G, = 0.032).
Large aggregations of the storm-petrel Oceanites oceanicus contributed most to the patchiness at the northern edge of WCR 84- 51 (left side of Fig. 6). Seabird aggregations within ring center (Table 3) consisted pri- marily of Oceanodroma leucorhoa and the shearwater Puffinus Iherminieri, the latter species feeding near patches of the macroal- ga Sargassum (Haney 1986a).
Both the relative abundances and pro- portions of seabird species differed among water masses. The assemblage within WCR
Table 2. Ranges of sea surface temperature (SST) and surface salinities within four water mass types during northwest Atlantic seabird counting transects, August 1984.
Water mass
WCR SL SH SS
SST (?C) 24.3-27.6 18.8-21.7 22.5-23.0 27.3-29.0 Salinity (Too) 33.9-34.6 31.7-31.8 30.5-30.9 35.8-36.1
This content downloaded from 62.122.79.81 on Tue, 17 Jun 2014 21:02:55 PMAll use subject to JSTOR Terms and Conditions

Notes 669
20
15
IX10 S
0
Ss 2 -WCR
4 SL
6 -
o 8-
ix 10 - SH Fig. 4. Comparisons of mean seabird density and
biomass within WCR 84-51 (WCR), slope (SL), shelf (SH), and Sargasso Sea (SS) water masses.
1000. Warm Core Ring
100-
10-
0.
z ,oo- Slope Water
10 -
10oo Shelf Water
10- *
(U i
,o- Sargasso Sea
2 2 44 66 88 110
Distance, Km
Fig. 5. Numbers of seabirds along a horizontal transect within WCR 84-51 compared to transects within other water masses in the northwest Atlantic Ocean, August 1984. The four figure sections have equal vertical scales.
Numerical Composition Warm Core Ring Slope
Shelf Sargasso Sea
Biomass Composition
Warm Core Ring Slope
Shelf Sargasso Sea
[7] Hydrobatidae (Oceanites, Oceanodroma) E] Procellariidae (Puffinus, Calonectris)
Phaethontidae (Phaethon) m Laridae (Larus, Sterna)
Fig. 6. Faunal composition of seabirds at WCR 84- 51 and slope, shelf, and Sargasso Sea water masses. Numerical composition indicates relative proportions based on total numbers of taxa per water mass. Bio- mass composition indicates relative proportions of the assemblage comprised by taxa adjusted for body mass in kilograms.
84-51 was dominated by hydrobatids (93% of numbers; 55% of biomass), followed by procellarids (7%; 43%), and larids (</1%; 2%). Species composition within the ring was most similar to Slope Water (Fig. 5). However, WCR 84-51 contained a rela- tively higher percentage of hydrobatids and lower percentage of procellarids than Slope Water (P < 0.005, 2 x 2 contingency table).
This content downloaded from 62.122.79.81 on Tue, 17 Jun 2014 21:02:55 PMAll use subject to JSTOR Terms and Conditions

670 Notes
Table 3. Seabird species abundances within WCR 84-51. Total number (N), density or No. km-2, and percentage of total N are given for northern ring edge, ring center, and southern ring edge intervals. Each interval consisted of a 27.8-km-long series of count transects (n = 6).
Northern Center Southern
N Density % N Density % N Density %
Calonectris diomedea 2 0.1 50 2 0.1 50 0 0.0 0 Puffinus gravis 0 0.0 0 0 0.0 0 1 0.0 100 Puffinus Iherminieri 4 0.1 19 1 1 0.4 52 6 0.2 29 Oceanites oceanicus 337 12.1 99 4 0.1 1 0 0.0 0 Oceanodroma leucorhoa 1 0.0 3 24 0.9 75 7 0.3 22 Stercorarius sp. 0 0.0 0 1 0.0 100 0 0.0 0
The WCR assemblage had relatively fewer procellarids and larids and more hydrobat- ids than Shelf Water (P < 0.005, 2 x 3 contingency table), and no phaethontids and more hydrobatids relative to Sargasso Sea water (P < 0.005, 2 x 2 contingency table).
Seabirds within WCR 84-51 consisted primarily (96%) of individuals belonging to species common in adjacent Slope Water. Puffinus Iherminieri and a few larids com- prised the remainder of the assemblage. The Shelf Water seabird assemblage and species- poor Sargasso Sea assemblage contributed little to the ring seabird fauna.
Hydrobatids (storm-petrels) feed primar- ily on surface zooplankton (Ashmole 1971). The warm-core ring seabird assemblage thus contained relatively more zooplankton feeders and fewer nekton feeders (procel- larids: shearwaters; and larids: gulls, terns, jaegers) than did adjacent water masses.
Most species showed strong affinities for a single water type (Fig. 7). Six species were entirely restricted to one water mass. The only species restricted to the ring was Au- dubon's shearwater (P. iherminieri), a warm- water counterpart and sibling species to the Manx shearwater (Puffinus puffinus). The distributions of the two species did not overlap (Fig. 7). Puffinus Iherminieri is a subtropical species common in Gulf Stream and stream-influenced waters off the south- eastern United States (Clapp et al. 1982). At higher latitudes the species may be re- stricted to the Gulf Stream (Brown 1977) or to locations and seasons corresponding to elevated sea surface temperatures (Rowlett 1980).
Seabird distributions in WCR 84-51 in- dicate that apex consumers are not exempt
from the horizontal heterogeneity associ- ated with rings that affects other marine trophic groups. Seabirds depend directly or indirectly on several taxa influenced by rings-including euphausiids, copepods, and other zooplankton (McWilliam and Phillips 1983; Wiebe and Flierl 1983; Tranter et al. 1983; Roman et al. 1985), cephalopods (Brandt 1983), and midwater fishes (Brandt 1981; Olson and Backus 1985). Changes in seabird abundance and biomass (Fig. 4), as- semblage structure (Fig. 6), and individual species distributions (Fig. 7; Table 3) at the ring could result from variability in fau- nistic composition of zooplankton prey within the ring (Griffiths and Brandt 1983; Tranter et al. 1983), in absolute abundance of prey (McWilliam and Phillips 1983), or in size classes (Brandt 1983) of these prey.
The transition states of warm-core rings result in greater zooplankton biomass com- pared to adjacent Slope Water (Roman et al. 1985). One zooplankton-feeding storm- petrel, 0. oceanicus, was more abundant in WCR water than in Slope Water (Fig. 7). The other storm-petrel, 0. leucorhoa, was also common in ring center, although it was more abundant in Slope Water (Table 3). Variation in the distribution of physical processes and organisms within warm-core rings could affect the distributions of sea- birds. Zooplanktivorous storm-petrels were most abundant near the ring's northern edge (Fig. 6; Table 3). Roman et al. (1985) found that surface zooplankton biomass was high- est near the northeast boundary of WCR 82-B, decreasing across the ring to the southwest. Elevated zooplankton biomass at the boundary of WCR 82-B was attrib- uted to a sharpened thermocline associated
This content downloaded from 62.122.79.81 on Tue, 17 Jun 2014 21:02:55 PMAll use subject to JSTOR Terms and Conditions

Notes 671
Calonectris diomedea Puffinus gravis Puffinus griseus Puffinus puffinus
.40- .40- .40-
20- 220- .20 -
- 0 0 0 0- WCR SL SH SS WCR SL SH SS WCR SL SH SS WCR SL SH SS
rz Puff inus Iherminieri Oceanites oceanicus Oceanodroma leucorhoa Phaethon lepturus 80 12 5 .04
O .60- 4- .03-
.40-62 .02-
WCR SL SH SS WCR SL SH SS WCR SL SH SS WCR SL SH SS Larus argentatus Larus marinus Sterna hirundo
3 40
1 2- .20-
0- 0 0 WCR SL SH SS WCR SL SH SS WCR SL SH SS
Fig. 7. Relative abundances of the 1 1 most abundant seabird species observed within four water mass types. Vertical representations of abundance among species are not to scale. Abbreviations defined in Table 1.
with an intruding streamer of Shelf Water origin (Joyce and Stalcup 1984; Nelson et al. 1985). Olson and Backus (1985) found myctophid fishes aggregated by advection in the frontal boundary between WCR 82-B and Slope Water, and in the ring center. Leach's storm-petrels (0. leucorhoa) feed nocturnally on vertically migrating mycto- phids (Brown 1980; Watanuki 1985). Storm- petrels were most abundant both at the edge and center of the ring (Table 3).
Warm-core rings represent anomalous in- trusions into more temperate water masses and have been previously suspected as ve- hicles for the north- or shoreward displace- ment of tropical and subtropical seabird species (Rowlett 1980; Hunt and Schneider 1987). The occurrence of P. Iherminieri in WCR 84-51 and its absence from Slope Water is particularly significant. Individuals of this species probably originated from the Gulf Stream water mass where they were common during a brief crossing of the stream on 30 August. Other northwest Atlantic sea- bird species with similar water-mass affin- ities could also be affected by warm-core rings, including band-rumped storm-petrel (Oceanodroma castro), black-capped petrel
(Pterodroma hasitata), and bridled tern (Sterna anaethetus) (cf. Rowlett 1980; Ha- ney 1986b, 1987).
Whether the displacement of P. Ihermi- nieri at the ring was primarily due to active or passive habitat selection is unclear. If the foraging ambits of P. Iherminieri were sig- nificantly smaller than the spatial dimen- sions of the ring at the time scales of ring evolution and translation, passive displace- ment could predominate. Displacement would thus require within-ring containment and transport of the species from the parent water mass. Shearwaters could easily return to the parent water mass, however, unless foraging opportunities were similar or per- haps better within the ring. Active displace- ment would occur if foraging ambits of the species were larger than the ring. In this case, shearwater affinities for the ring would represent aggregation or extrinsic recruit- ment of individuals to a physically and bi- ologically distinct marine habitat. Although the foraging ambits of seabirds over time scales of weeks to months (the time scales of rings) are not well known, Warham et al. (1977) found that ambits for some procel- larids are at least 1,000 km, suggesting that
This content downloaded from 62.122.79.81 on Tue, 17 Jun 2014 21:02:55 PMAll use subject to JSTOR Terms and Conditions

672 Notes
active rather than passive displacement may predominate at these meso-scale features.
J. Christopher Haney2
Department of Zoology University of Georgia Athens 30602
References ASHMOLE, N. P. 1971. Sea bird ecology and the ma-
rine environment, p. 224-286. In D. S. Farner and J. R. King [eds.], Avian biology, v. 1. Academic.
BRANDT, S. B. 1981. Effects of a warm-core eddy on fish distributions in the Tasman Sea off East Aus- tralia. Mar. Ecol. Progr. Ser. 6: 19-33.
1983. Pelagic squid associations with a warm- core eddy of the East Australian Current. Aust. J. Mar. Freshwater Res. 34: 573-585.
BROWN, R. G. B. 1977. Atlas of eastern Canadian seabirds. Suppl. 1, Halifax-Bermuda transects. Can. Wildl. Serv.
1980. Seabirds as marine animals, p. 1-39. In J. Burger et al. [eds.], Behavior of marine an- imals, v. 1. Plenum.
CLAPP, R. B., R. C. BANKS, D. MORGAN-JACOBS, AND W. A. HOFFMAN. 1982. Marine birds of the southeastern United States and Gulf of Mexico. Part 1, Gaviiformes through Pelecaniformes. U.S. Fish Wildl. Serv.
, D. MORGAN-JACOBS, AND R. C. BANKS. 1983. Marine birds of the southeastern United States and Gulf of Mexico. Part 3, Charadriiformes. U.S. Fish Wildl. Serv.
CRAMP, S., AND K. E. I. SIMMONS. 1977, 1983. The birds of the western Palearctic. V. 1 and 3. Oxford.
ELLIOTT, J. M. 1977. Some methods for the statistical analysis of samples of benthic invertebrates. Freshwater Biol. Assoc. Sci. Publ. No. 25.
GREEN, R. H. 1966. Measurement of non-random- ness in spatial distributions. Res. Pop. Ecol. Kyoto Univ. 8: 1-7.
GRIFFITHS, F. B., AND S. B. BRANDT. 1983. Meso- pelagic crustacea in and around a warm-core eddy in the Tasman Sea off eastern Australia. Aust. J. Mar. Freshwater Res. 34: 609-623.
HANEY, J. C. 1986a. Seabird patchiness in tropical oceanic waters: The influence of Sargassum "reefs." Auk 103: 141-151.
1986b. Seabird segregation at Gulf Stream frontal eddies. Mar. Ecol. Progr. Ser. 28: 279-285.
1987. Aspects of the pelagic ecology and be- havior of the black-capped petrel (Pterodroma hasitata). Wilson Bull. 99: 153-168.
2 Present address: Alaska Fish and Wildlife Research Center, U.S. Fish and Wildlife Service, 101 1 East Tu- dor Road, Anchorage 99503.
, AND P. A. McGILLIVARY. 1985. Aggregations of Cory's shearwaters (Calonectris diomedea) at Gulf Stream fronts. Wilson Bull. 97: 191-200.
HITCHCOCK, G. L., C. LANGDON, AND T. J. SMAYDA. 1985. Seasonal variations in the phytoplankton biomass and productivity of a warm-core Gulf Stream ring. Deep-Sea Res. 32: 1287-1300.
HUNT, G. L., JR., AND D. SCHNEIDER. 1987. Scale dependent processes in the physical and biological environment of marine birds, in press. In J. P. Croxall [ed.], Seabird feeding ecology: the role of seabirds in marine ecosystems. Cambridge.
JOYCE, T., R. SCHMIDT, AND M. STALCUP. 1983. In- fluence of the Gulf Stream upon short-term evo- lution of a warm-core ring. Aust. J. Mar. Fresh- water Res. 34: 515-524.
, AND M. C. STALCUP. 1984. An upper ocean current jet and internal waves in a Gulf Stream warm-core ring. J. Geophys. Res. 89: 1997-2003.
, AND P. WIEBE. 1983. Warm-core rings of the Gulf Stream. Oceanus 26: 34-44.
MCWILLIAM, P. S., AND B. F. PHILLIPS. 1983. Phyl- losoma larvae and other crustacean macrozoo- plankton associated with eddy J, a warm-core eddy off southeastern Australia. Aust. J. Mar. Fresh- water Res. 34: 653-663.
NELSON, D. M., AND OTHERS. 1985. Distribution and composition of biogenic particulate matter in a Gulf Stream warm-core ring. Deep-Sea Res. 32: 1347-1369.
OLSON, D. B., AND R. H. BACKUS. 1985. The con- centrating of organisms at fronts: A cold-water fish and a warm-core Gulf Stream ring. J. Mar. Res. 43: 113-137.
ORTNER, P. R., E. M. HULBERT, AND P. H. WIEBE. 1979. Phytohydrography, Gulf Stream rings, and herbivore habitat contrasts. J. Exp. Mar. Biol. Ecol. 39: 101-124.
, P. H. WIEBE, L. HAURY, AND S. BOYD. 1978. Variability in zooplankton biomass distribution in the northern Sargasso Sea: The contribution of Gulf Stream cold-core rings. Fish Bull. 76: 323- 334.
ROMAN, M. R., A. L. GAUZENS, AND T. J. COWLES. 1985. Temporal and spatial changes in epipelagic microzooplankton and mesozooplankton biomass in warm-core Gulf Stream ring 82-B. Deep-Sea Res. 32: 1007-1022.
ROWLETT, R. A. 1980. Observations of marine birds and mammals in the northern Chesapeake bight. U.S. Fish Wildl. Serv.
TASKER, M. L., P. H. JONES, T. DIXON, AND B. F. BLAKE. 1984. Counting seabirds at sea from ships: A review of methods employed and a suggestion for a standardized approach. Auk 101: 567-577.
TRANTER, D. J., D. J. TAFE, AND R. L. SANDLAND. 1983. Some zooplankton characteristics of warm- core eddies shed by the East Australian Current, with particular reference to copepods. Aust. J. Mar. Freshwater Res. 34: 587-607.
WARHAM, J., B. R. KEELEY, AND G. J. WILSON. 1977. Breeding biology of the mottled petrel. Auk 94: 1- 17.
WATANUKI, Y. 1985. Food of breeding Leach's storm-
This content downloaded from 62.122.79.81 on Tue, 17 Jun 2014 21:02:55 PMAll use subject to JSTOR Terms and Conditions

Notes 673
petrels (Oceanodroma leucorhoa). Auk 102: 884- 886.
WIEBE, P. H., AND G. R. FLIERL. 1983. Euphausiid invasion/dispersal in Gulf Stream cold-core rings. Aust. J. Mar. Freshwater Res. 34: 625-652.
, AND OTHERS. 1976. Gulf Stream cold-core
rings: Large-scale interaction sites for open ocean plankton communities. Deep-Sea Res. 23: 695- 710.
Submitted: 4 March 1986 Accepted: 20 December 1986
Limnol. Oceanogr., 32(3), 1987, 673-680 ? 1987, by the American Society of Limnology and Oceanography, Inc.
Effect of interreplicate variance on zooplankton sampling design and data analysis'
Abstract-The variance of replicate marine and freshwater zooplankton samples is related (r2 > 0.99) to the mean population density and the volume of the sample taken. A general variance relationship calculated from 1,189 sets of repli- cate samples taken throughout the world is pre- sented. This function can predict the variance of both marine and freshwater samples taken with several types of sampling gear. Only 12% of in- dividual taxa examined showed variance rela- tionships that departed significantly from this general function. The variance function shows how the requisite number of samples decreases with increased population density, sampler vol- ume, and lowered precision requirements. The X02 power transformation is recommended on empirical grounds, although caution is advised in the use of all data transformations.
Most agree that zooplankton sampling variability is large (e.g. de Bemardi 1984; Fasham 1978; Malone and McQueen 1983; Omori and Ikeda 1984). Quantitative pre- diction of sampling variance is important for two practical reasons. First, all zoo- plankton population ecologists must decide how many samples and what volume of sample to take. The requisite number of samples is determined by the precision re- quired and the sampling variance (s2). Pre- diction of S2 would allow efficient sampling design. Second, many researchers use para- metric statistical methods because these
I A contribution to the Group d'Ecologie des Eaux Douces of l'Universite de Montreal. This research was supported by the Natural Sciences and Engineering Re- search Council of Canada, and the Department of Ed- ucation of the Province of Quebec (FCAR).
methods are powerful, familiar, and widely available in computer packages. The vari- ance of raw population estimates varies with the population density, and data must be transformed to equalize variances, alleviate skewness, and increase the probability of additivity (Snedecor and Cochran 1967; Southwood 1966). Knowledge of the rela- tive size of the variance and mean (m) of replicate zooplankton population estimates can allow effective power transformation of raw data (Taylor 1961).
The purpose of this study is to assemble information on interreplicate variance in marine and freshwater zooplankton sam- ples and to develop an equation to char- acterize this variability. Further, we show how these analyses can be used to choose sample number and size and to choose proper transformations for zooplankton data.
We thank H. Cyr, C. Plante, L. Rath, and three anonymous reviewers for criticism of the manuscript and B. Pinel-Alloul for help- ful discussion.
Data on m and S2 (n - 1 weighting) of replicate zooplankton population estimates were collected from the published literature. We tried to gather data covering a range of environments in many geographical areas, collected with a variety of sampling gear (Table 1). Population estimates were usu- ally made at the genus or species level; 161 taxa were surveyed (Table 2). We collected 1,189 sets of replicate observations (n'), covering a range of m from 5 x 10-7 to
This content downloaded from 62.122.79.81 on Tue, 17 Jun 2014 21:02:55 PMAll use subject to JSTOR Terms and Conditions