effects acetate, propionate, and butyrate the anaerobic...
TRANSCRIPT

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Apr. 1993, p. 1003-10110099-2240/93/041003-09$02.00/0Copyright ©) 1993, American Society for Microbiology
Effects of Acetate, Propionate, and Butyrate on the ThermophilicAnaerobic Degradation of Propionate by Methanogenic
Sludge and Defined CulturesJULES B. VAN LIER,1* KATJA C. F. GROLLE,1 CARLA T. M. J. FRIJTERS,l
ALFONS J. M. STAMS,2 AND GATZE LETTINGA1
Department ofEnvironmental Technology, Wageningen Agricultural University, Bomenweg 2,6703 HD Wageningen, 1 and Department ofMicrobiology, Wageningen Agricultural
University, 6703 CT Wageningen,2 The Netherlands
Received 21 September 1992/Accepted 8 January 1993
The effects of acetate, propionate, and butyrate on the anaerobic thermophilic conversion of propionate bymethanogenic sludge and by enriched propionate-oxidizing bacteria in syntrophy with Methanobacteriumthermoautotrophicum AH were studied. The methanogenic sludge was cultivated in an upflow anaerobic sludgebed (UASB) reactor fed with propionate (35 mM) as the sole substrate for a period of 80 days. Propionatedegradation was shown to be severely inhibited by the addition of 50 mM acetate to the influent of the UASBreactor. The inhibitory effect remained even when the acetate concentration in the effluent was below the levelof detection. Recovery of propionate oxidation occurred only when acetate was omitted from the influentmedium. Propionate degradation by the methanogenic sludge in the UASB reactor was not affected by theaddition of an equimolar concentration (35 mM) of butyrate to the influent. However, butyrate had a stronginhibitory effect on the growth of the propionate-oxidizing enrichment culture. In that case, the conversion ofpropionate was almost completely inhibited at a butyrate concentration of 10 mM. However, addition of a
butyrate-oxidizing enrichment culture abolished the inhibitory effect, and propionate oxidation was evenstimulated. All experiments were conducted at pH 7.0 to 7.7. The thermophilic syntrophic culture showed a
sensitivity to acetate and propionate similar to that of mesophilic cultures described in the literature. Additionsof butyrate or acetate to the propionate medium had no effect on the hydrogen partial pressure in the biogasof an UASB reactor, nor was the hydrogen partial pressure in propionate-degrading cultures affected by thetwo acids. Our results indicate that the level of hydrogen in the biogas of thermophilic anaerobic reactors is notthe appropriate control parameter for propionate degradation. High concentrations of NaCl, sodium acetate,or sodium propionate (100 mM) did not affect the growth of M. thermoautotrophicum AH on hydrogen.
Acetate, propionate, and butyrate are intermediate prod-ucts in the anaerobic bioconversion of organic matter tomethane and carbon dioxide (5, 16, 29). Under balancedmethanogenic conditions propionate and butyrate are furtheroxidized to acetate, hydrogen, and bicarbonate, the mainprecursors of methanogenesis (Table 1). For thermodynamicreasons propionate and butyrate can be degraded only whenacetate and especially hydrogen are effectively eliminated bythe methanogens (1, 2, 6, 7, 12, 17, 18, 23). At elevatedtemperatures the thermodynamics for the acetogenic con-
versions are somewhat more favorable (Table 1). Neverthe-less, propionate often is the first volatile fatty acid (VFA)which accumulates during thermophilic anaerobic treatmentof waste and wastewater (14, 25, 28). The contribution ofpropionate to the total amount of VFA in the effluent ofthermophilic upflow anaerobic sludge bed (UASB) reactorsis much higher under thermophilic conditions than undermesophilic conditions (27). The reason for the high sensitiv-ity of propionate oxidation in thermophilic methanogenicsystems is not clear yet.
Recently we have described the enrichment of thermo-philic, spore-forming, propionate-oxidizing bacteria in syn-trophy with Methanobacterium thermoautotrophicum AH or
Methanobacterium thernoformicicum Z245 (23). Growth ofthe propionate-oxidizing bacteria was possible only after
* Corresponding author.
inoculation into a hydrogen-pregrown culture of M. ther-moautotrophicum AH cells which were embedded in FeSprecipitates. In that study, results were obtained which showthat the decay rate of M. thermoautotrophicum AH underunfed conditions is extremely high. Therefore, starvation ofhydrogenotrophic methanogens might play an important rolein the sensitivity of propionate oxidation in thermophilicmethanogenic ecosystems. Growth of the propionate-de-grading enrichment was stimulated additionally by the pres-ence of aceticlastic methanogens. Studies on propionateconversion under mesophilic conditions have demonstratedthat the anaerobic oxidation of propionate is influenced byboth acetate and propionate concentrations (10, 11, 14, 15,19). Furthermore, deterioration of propionate degradation isaccompanied by increased concentrations of acetate both inthe mesophilic range and the thermophilic range (14, 25, 27).In this paper we describe the effects of acetate, propionate,and butyrate on the thermophilic oxidation of propionate incontinuous-flow methanogenic sludge bed systems and indefined propionate-oxidizing cultures.
MATERIALS AND METHODS
Inoculum. The upflow anaerobic sludge bed (UASB) reac-tor was seeded with mesophilic granular sludge in theamount of 22.6 g of volatile suspended solids per liter ofreactor volume, which corresponds to a total volume of 2liters of wet sludge. The inoculum originated from a 1,700-mi3
1003
Vol. 59, No. 4
on May 31, 2018 by guest
http://aem.asm
.org/D
ownloaded from

1004 vAN LIER ET AL.
TABLE 1. Acetogenic and methanogenic reactions involved inpropionate oxidation and the Gibbs free-energy changesa
AGO' AG'Reaction (U) at (U) at
25°C 550C
HCO32+ H+ + 4H2CH4 + -135.6 -122.53H20
Acetate- + H20-*HCO3 + CH4 -31.0 -34.7Propionate- + 3H2O--HCO03 +76.1 +62.3+ acetate- + H+ + 3H2
Butyrate- + 2H20--2 acetate +48.1 +37.9+ H+ + 2H2
a Gibbs free-energy changes were calculated by using the van't Hoffequation, standard enthalpy values for compounds (8), and Gibbs free-energychanges (AGO') at 25'C (24).
UASB reactor treating potato-processing wastewater at36°C. The sludge was kindly provided by the Aviko factoryat Steenderen, The Netherlands.
Media. The UASB reactor was fed with a concentratedstock solution which contained (per liter) 67.5 g of sodiumpropionate, 3.7 g of NH4Cl, 1.5 g of MgSO4. 7H20, 13.8 g ofNaH2PO4. 2H20, 10.6 g of K2HPO4, 0.2 g of CaCl2- 2H20,and 0.3 g of yeast extract. The pH of the feed stock was 6.5,which resulted in a reactor pH of 7.0 to 7.7. To each 1 literof stock solution 13.3 ml of a trace element solution (25) wasadded. Before it was fed into the UASB reactor, the stocksolution was diluted 20 times with hot tap water containing30 mg of Ca2+ per liter. Sodium acetate and sodium butyratewere added during short time intervals in the ratio describedbelow. Defined culture studies were performed by using thesame medium and culture conditions as described previously(23).UASB reactor. The continuous-flow experiments were
performed at 55'C in a polyacrylate (perspex) UASB reactorwith a working volume of 5.75 liters. The experimental setupwas the same as that described previously (25). The reactorwas operated for more than 80 days with propionate as thesole carbon source with a hydraulic retention time of 8 h.Day 70 was defined as zero time for the butyrate inhibitionexperiment. Before the experiment was started, the propi-onate load was raised to a level of 100 mmol of propionateper liter of reactor volume per day, corresponding to achemical oxygen demand of 11.2 kg/m3 of reactor volumeper day. After 10 days of acclimatization butyrate was addedas the second carbon source in the influent at a molar ratio ofpropionate to butyrate of 1:0.4. Two weeks after the firstaddition, the supply of butyrate was increased to a molarratio of propionate to butyrate of 1:1 for 8 days. Thereafter,butyrate was left out of the medium, and the reactor wasoperated for 2 months with propionate as the sole substrate.The end of this period was defined as zero time for theacetate inhibition experiment. Acetate was added to theinfluent for a period of 2 weeks at a molar ratio of propionateto acetate of 1:1.7 at a propionate concentration of 27 mM.This procedure was repeated after a 3-week period duringwhich the system was fed only propionate as the carbonsource. Then the supply of acetate was increased to a molarratio of propionate to acetate of 1:2.6 for about 1 week; thenthe reactor was operated again with propionate as the solecarbon source for 15 days.
Defined cultures. In a previous paper we described theenrichment of thermophilic, spore-forming, propionate-oxi-dizing bacteria from thermophilic (55°C) methanogenic
sludge by repeated transfer in hydrogen-pregrown culturesof M. thermoautotrophicum AH (23). This culture did notcontain aceticlastic methanogens. In the present study thehighly enriched propionate-oxidizing culture was used toassess the effects of VFAs on the thermophilic propionateconversion. In all experiments the propionate-oxidizing cul-ture was inoculated into hydrogen-pregrown cultures of M.thermoautotrophicum AH by using the same medium andculture conditions described previously (23). To investigatethe effect of acetate, propionate, or butyrate on the growthof the hydrogenotrophic methanogens, M. thennoau-totrophicum /H was pregrown either in the presence or inthe absence of the fatty acid salt. In the latter experiment thefatty acid salt was added after growth of M. thermoau-totrophicum AH on H2-CO2. In both experiments the finalstep was inoculation of 10% of the dense propionate-oxidiz-ing culture into the media. This was defined as zero time forthe experiment. However, no differences were found be-tween the two different experiments. Therefore, the resultsof these experiments are presented as duplicate results (seeFig. 3 and 4).
In one experiment we investigated the effect of butyrate-oxidizing bacteria on propionate conversion. Butyrate-oxi-dizing acetogens were obtained by enriching thermophilicbutyrate-degrading bacteria by repeated transfer (10%) inhydrogen-pregrown cultures of M. thermoautotrophicumAH with 20 mM butyrate as the sole carbon source. M.thermoautotrophicum AH appeared to be the most abundantor even the only syntrophic partner of the butyrate oxidiz-ers. Inocula of 2% of the dense propionate-oxidizing cultureand 1% of the butyrate-oxidizing culture were added to-gether with FeCl2 (final concentration, 1 mM) to the hydro-gen-pregrown culture of M. thermoautotrophicum AH. Priorto inoculation the gas phase was changed to 172 kPa ofN2-CO2 (80:20, vol/vol). Propionate (20 mM) and butyrate (0to 50 mM) were added to the bottles. All incubations werecarried out without shaking at 55°C.
Analysis. The VFA and gas compositions were determinedby gas chromatography as described previously (23, 25).Hydrogen was also determined by gas chromatography witha Hewlett-Packard model 5890 gas chromatograph equippedwith a thermal conductivity detector and a 25 H molecularsieve (60-80 mesh). The column size was 1.5 m by 6.4 mm.Argon was used as the carrier gas at a flow rate of 25 ml/min.The column temperature was 40°C, the injection port tem-perature was 110°C, and the detector temperature was125°C. All other analyses were performed by standard meth-ods (3).
Kinetic analysis. Kinetic constants were estimated byusing the "Nelder" nonlinear regression routine (20) forparameter estimation. Data were fitted to a noncompetitiveproduct inhibition model as described by other workers (10,19).
RESULTS
Effect of VFAs on propionate conversion by thermophilicmethanogenic sludge. The effect of VFAs on propionateconversion by methanogenic sludge was investigated in aUASB reactor which was started up with propionate as thesole carbon source. After the start-up was completed, bu-tyrate was added to the medium at a molar ratio of propi-onate to butyrate of 1:0.4, while the propionate concentra-tion was kept at 35 mM. The effects of the butyrate additionon propionate conversion are shown in Fig. 1. Propionatedegradation was affected by butyrate only during the first 3
APPL. ENVIRON. MICROBIOL.
on May 31, 2018 by guest
http://aem.asm
.org/D
ownloaded from

THERMOPHILIC ANAEROBIC DEGRADATION OF PROPIONATE 1005
0EE
0)
4-C
0EE
-t9-C_0):34-4-0)
0-o
0
E
E
0)L.
W
16
12
8
4
0
0.4
0.2
time (days)FIG. 1. Effect of butyrate additions to the influent of a UASB reactor which was started up with propionate as the sole substrate: acetate
(C1), propionate (A), and butyrate (O) concentrations in the influent (A) and effluent (B) of the reactor, as well as the absolute amounts of
butyrate and propionate removed per day (C).
B
VOL. 59, 1993
on May 31, 2018 by guest
http://aem.asm
.org/D
ownloaded from

1006 VAN LIER ET AL.
TABLE 2. Growth rates of propionate oxidizers at differentinitial concentrations of propionate, calculated from the
exponential increase in acetate concentrationduring the first 25 days of incubation
Total Undissociated p (day-') in p (day-') inpropionate propionic acid the presence the absence ofconcn (mM) concn (mM)' of propionate" propionateb
5 0.04 0.053 0.05410 0.08 0.092 0.10325 0.21 0.085 0.09650 0.42 0.098 0.071100 0.84 0.084 0.072
Calculated by using a pK, of 4.92 at 55°C (22) and pH 7.b Propionate oxidizers were inoculated into cultures of M. thermoau-
totrophicum AH pregrown on hydrogen either in the presence or absence ofpropionate.
days of exposure. During this period a very fast increase inthe butyrate-degrading activity by the methanogenic consor-tium was measured. After 3 days, butyrate removal wasalmost complete, and no inhibitory effects of this acid wereobserved. Even after the butyrate concentration was raisedto 35 mM, the propionate conversion rate remained unaf-fected. The hydrogen partial pressure in the gas phase of thereactor remained at 20 Pa throughout the whole experimen-tal period, except during the first day of butyrate addition(day 13), when the hydrogen partial pressure temporarilyincreased to 1,000 Pa; it dropped to its initial level of 20 Pathe next day. At day 34 of the inhibition experiment,butyrate was omitted from the medium, and the reactor wasoperated again with propionate as the sole carbon source. Inthis period substrate inhibition was observed after a suddenincrease in the propionate concentration from 32 to 52.5 mM(data not shown). This resulted in a decrease in the propi-onate removal rate of 20% and a concomitant decrease in theeffluent pH from 7.7 to 7.3. According to the results obtainedin the defined-culture studies (Table 2), this slight drop in pHshould not be detrimental for propionate conversion. How-ever, an influent pH of 6.5 for the feed stock solutionresulted in an undissociated propionic acid concentration of1.38 mM at the inlet point of the reactor. Previous studies onmesophilic propionate degradation revealed a considerableinhibition at this concentration (10). Stable reactor perfor-mance was obtained after the propionate concentration inthe influent was decreased to 27 mM.One week after zero time of the acetate inhibition exper-
iment, acetate was added to the propionate medium at amolar ratio of propionate to acetate of 1:1.7 for a period of 2weeks (Fig. 2). This addition was repeated during days 38 to51 and then during days 65 to 73. In the latter period a higherconcentration of acetate was used. During all of the acetateexposures the propionate concentration remained at about27 mM in the influent. Figure 2 shows the effect of acetateaddition on propionate conversion by the methanogenicsludge bed. The propionate peak at day 29 (Fig. 2A) wascaused by a disturbance in the food supply. In contrast to thebutyrate additions, acetate apparently initiated an increasein the propionate concentration in the effluent of the UASBreactor, which persisted until the supply of acetate waseliminated. This inhibitory effect of acetate remained, andeven increased, despite the very low acetate concentrationsin the effluent. The propionate conversion capacity dropped40 to 50% during the acetate additions (Fig. 2C). Because ofthe great differences between the absolute amounts of ace-tate and propionate which were removed per day, we
expressed the amounts of daily acetate and propionateremoval in Fig. 2C as percentages of the influent amount.
Effect of VFAs on propionate degradation in a definedpropionate-degrading culture. The inhibition effect of lowerfatty acids on the growth of thermophilic propionate oxidiz-ers in syntrophy with M. thermoautotrophicum AH wasinvestigated by using a highly enriched propionate-oxidizingculture which was unable to degrade acetate (23). Theaddition of inorganic salts or fatty acid salts had only littleeffect on M. thennoautotrophicum AH itself. Comparablegrowth rates ofM. thermoautotrophicum AH were measuredwith hydrogen in media to which 0, 50, or 100 mM KCl,NaCl, potassium acetate, sodium acetate, or sodium propi-onate was added (data not shown). Furthermore, no differ-ences in the rates of propionate conversion were foundirrespective of whether the methanogens were pregrown inthe presence or absence of the fatty acid salts (Fig. 3 and 4).Therefore, we attribute the effects of fatty acids on thepropionate-oxidizing enrichment culture to inhibition of theacetogens.As depicted in Fig. 3, propionate oxidation was severely
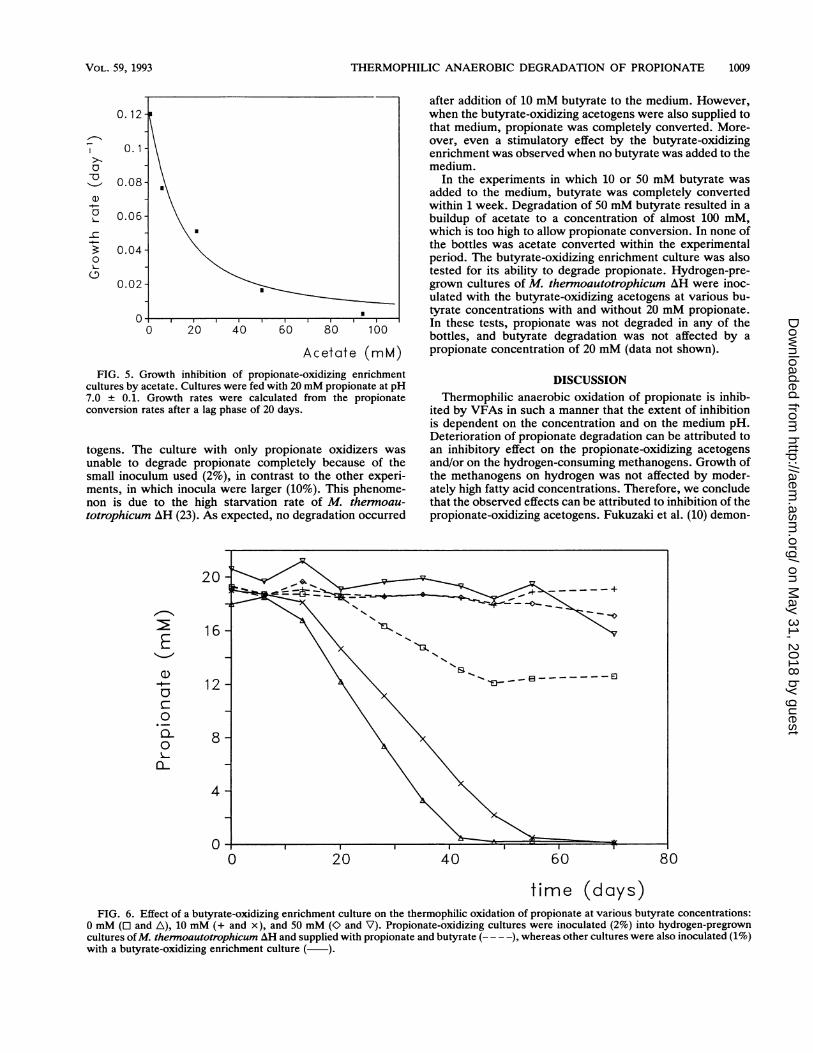
inhibited at acetate concentrations higher than 20 mM. Thisinhibition effect can be attributed to acetate as 100 mM NaClhad a less severe inhibitory effect (Fig. 3). Inhibition ofpropionate oxidation by acetate can be described by usingthe following noncompetitive inhibition equation used byother workers (10, 19): pL = {Jm/[l + (Ac/Ki)n]}, where ,. isthe specific growth rate of the acetogens; p.r is the maximumspecific growth rate; Ac is the acetate concentration; Ki isthe inhibition constant; and n is the exponent of inhibition.The kinetic constants pUm and ,u were calculated from theexponential decrease in the propionate concentration. Fig-ure 5 shows a plot of the growth rate of the propionate-oxidizing coculture versus acetate concentration. The solidline was computed statistically by using the commonly usednonlinear absolute least-square criterion as the error func-tion. The computed values for Ki and n are 12.4 mM acetateand 1.20, respectively. The Ki value equals 78 F.M undisso-ciated acetic acid, as calculated by using a pKa of 4.80 at55°C (22). Because of the heterogeneity of our datum points,application of the alternative relative least-square criterion ismore appropriate (21). When the latter error function wasused, the computed constants were somewhat higher: 18.4mM acetate (116 p.M undissociated acetic acid) for Ki and2.27 for n.The effect of propionate was studied by adding propionate
concentrations up to 100 mM to the medium. Table 2summarizes the calculated growth rates of the propionateoxidizers at the various initial concentrations of total propi-onic acid, as well as undissociated propionic acid. Surpris-ingly, propionate conversion rates were in the same rangeirrespective of the propionate concentration. The growthrates were calculated by using the exponential increase inthe acetate concentration during the first 3 to 4 weeks ofincubation. Thereafter, inhibition by acetate may have influ-enced the propionate degradation rate, especially at highinitial propionate concentrations.
Figure 4 shows that propionate conversion is very se-verely inhibited by butyrate. Although the number of usefuldata was limited, computer analyses in which the equationgiven above was used were carried out to estimate thefollowing butyrate inhibition constants: 1.7 mM butyrate (13p.M undissociated butyric acid, calculated by using a pKa of4.90 at 55°C [22]) for Ki and 2.65 for n. Addition of butyrateoxidizers to the culture abolishes the inhibition effect causedby butyrate itself. Instead, the acetate and hydrogen formed
APPL. ENVIRON. MICROBIOL.
on May 31, 2018 by guest
http://aem.asm
.org/D
ownloaded from

THERMOPHILIC ANAEROBIC DEGRADATION OF PROPIONATE 1007
80
0EE--
0)
:34-
>
F-
LL
E
E
C.0
4-
4-
0)
.-,
a)
41-I
E
0
L
4-C
_
E4-4-
>Qc-
60
40
20
0
20-
16 -
12 -
8-
4-
0O100 -
80 -
60 -
40 -
20 -
0-
time (days)FIG. 2. Effect of acetate additions to the influent of a UASB reactor degrading propionate as the sole substrate: acetate (5) and propionate
(A) concentrations in the influent (A) and effluent (B) of the reactor, as well as the relative amounts of acetate and propionate removed,expressed as percentages of the influent amount (C).
VOL. 59, 1993
on May 31, 2018 by guest
http://aem.asm
.org/D
ownloaded from

1008 vAN LIER ET AL.
20
E
Q)
0C-
0
0-0~*L_
a-
16
12
8
4-~~ ~ ~~~I6
02o 20 40 60
time (days)FIG. 3. Propionate degradation by thermophilic propionate-oxidizing enrichment cultures at various acetate concentrations: 0 mM (V), 5
mM (x), 20 mM (A), 50 mM (O), and 100 mM (O). Cultures were inoculated (10%) into cultures of M. thermoautotrophicum AH pregrownon hydrogen. As a reference the effect of 100 mM NaCl (---- -) is also shown.
from butyrate influence propionate conversion. In order toassess the effect of the butyrate oxidizers, an M. thermoau-totrophicum AH culture pregrown on hydrogen in the pres-ence of 20 mM propionate was inoculated with the propi-onate-oxidizing enrichment culture (2% of a stock culture)
20
C)I-
00-0a
16
12
8
4-
0
and supplied with butyrate (0, 10, and 50 mM). Some of thebottles were also inoculated with a butyrate-oxidizing en-richment culture. Figure 6 shows a plot of propionateconcentration versus incubation time at the various butyrateconcentrations with and without the butyrate-oxidizing ace-
0 20 40
time (days)FIG. 4. Propionate degradation by thermophilic propionate-oxidizing enrichment cultures at various butyrate concentrations: 0 mM (l),
5 mM (O), 10 mM (A), and 20 mM (x). Cultures were inoculated (10%) into cultures of M. thermoautotrophicum AH pregrown on hydrogen.
APPL. ENVIRON. MICROBIOL.
on May 31, 2018 by guest
http://aem.asm
.org/D
ownloaded from
THERMOPHILIC ANAEROBIC DEGRADATION OF PROPIONATE 1009
0.12
0-o
a)
0
0L.0D
0. 1
0.08
0.06
0.04
0.02
01 I0 20 40 60 80 100
Acetate (mM)FIG. 5. Growth inhibition of propionate-oxidizing enrichment
cultures by acetate. Cultures were fed with 20 mM propionate at pH7.0 ± 0.1. Growth rates were calculated from the propionateconversion rates after a lag phase of 20 days.
togens. The culture with only propionate oxidizers wasunable to degrade propionate completely because of thesmall inoculum used (2%), in contrast to the other experi-ments, in which inocula were larger (10%). This phenome-non is due to the high starvation rate of M. thermoau-totrophicum AH (23). As expected, no degradation occurred
20
a)E
0Q)0~o00~
16
12
8
4
0
after addition of 10 mM butyrate to the medium. However,when the butyrate-oxidizing acetogens were also supplied tothat medium, propionate was completely converted. More-over, even a stimulatory effect by the butyrate-oxidizingenrichment was observed when no butyrate was added to themedium.
In the experiments in which 10 or 50 mM butyrate wasadded to the medium, butyrate was completely convertedwithin 1 week. Degradation of 50 mM butyrate resulted in abuildup of acetate to a concentration of almost 100 mM,which is too high to allow propionate conversion. In none ofthe bottles was acetate converted within the experimentalperiod. The butyrate-oxidizing enrichment culture was alsotested for its ability to degrade propionate. Hydrogen-pre-grown cultures of M. thermoautotrophicum AH were inoc-ulated with the butyrate-oxidizing acetogens at various bu-tyrate concentrations with and without 20 mM propionate.In these tests, propionate was not degraded in any of thebottles, and butyrate degradation was not affected by apropionate concentration of 20 mM (data not shown).
DISCUSSIONThermophilic anaerobic oxidation of propionate is inhib-
ited by VFAs in such a manner that the extent of inhibitionis dependent on the concentration and on the medium pH.Deterioration of propionate degradation can be attributed toan inhibitory effect on the propionate-oxidizing acetogensand/or on the hydrogen-consuming methanogens. Growth ofthe methanogens on hydrogen was not affected by moder-ately high fatty acid concentrations. Therefore, we concludethat the observed effects can be attributed to inhibition of thepropionate-oxidizing acetogens. Fukuzaki et al. (10) demon-
0 20 40 60 80
time (days)FIG. 6. Effect of a butyrate-oxidizing enrichment culture on the thermophilic oxidation of propionate at various butyrate concentrations:
0 mM (El and A), 10 mM (+ and x), and 50 mM (O and V). Propionate-oxidizing cultures were inoculated (2%) into hydrogen-pregrowncultures ofM. thermoautotrophicum AH and supplied with propionate and butyrate (- - - -), whereas other cultures were also inoculated (1%)with a butyrate-oxidizing enrichment culture ( ).
VOL. 59, 1993
on May 31, 2018 by guest
http://aem.asm
.org/D
ownloaded from

1010 vAN LIER ET AL.
strated that the degree of inhibition was strongly dependenton the concentration of the undissociated form of the fattyacids. The neutral form of these acids diffuses more easilyacross the bacterial membrane, causing a drop in intracellu-lar pH. The increased diffusivity might be further enhancedby the interference of the apolar chain with the bacterialmembranes, which results in a higher permeability of thesemembranes. As in toxicity of alcohols, which is correlatedwith chain length (9), one might expect a higher degree ofinhibition with longer-chain-length fatty acids. In our exper-iments growth of thermophilic propionate-oxidizing bacteriawas most severely inhibited by butyrate. Strong inhibitioneven occurred at low concentrations. However, comparedwith acetate, high concentrations of propionate affected thegrowth of the thermophilic propionate-oxidizing acetogensonly slightly (Table 2). This may have been the net result ofinhibitory and stimulatory effects. Substrate inhibition prob-ably is also caused by toxicity of the undissociated form ofpropionic acid (10, 19) and its effect on the bacterial mem-branes. At high propionate concentrations the high amountof sodium may also contribute to inhibition. On the otherhand, a high propionate concentration, and thus a lowacetate/propionate ratio, lowers the Gibbs free-energychanges for propionate conversion (Table 1). A very lowratio enhances the growth of the acetogenic bacteria. Theeffects of propionate on propionate oxidation rates weresimilar to those observed by Fukuzaki et al. (10) withmesophilic propionate-acclimatized sludge. This indicatessimilar susceptibilities of mesophilic and thermophilic pro-pionate-oxidizing consortia.Addition of butyrate-oxidizing acetogens to the media
eliminated the inhibitory effect of butyrate and even stimu-lated propionate conversion. Stimulation of the growth ofthe propionate degraders in the presence of the butyrate-oxidizing acetogens might be caused by excretion of essen-tial growth factors by the butyrate oxidizers. However, it ismore likely that the effect can be attributed to an increase inthe number of hydrogenotrophic methanogens. The butyrateoxidizers were also enriched in hydrogen-pregrown culturesof M. thermoautotrophicum AH, and therefore, methano-gens were inoculated in small numbers together with theacetogens. Furthermore, growth of hydrogenotrophic meth-anogens also occurs as a result of butyrate degradation toacetate and hydrogen. These and previous results (23) showthat the number of viable hydrogenotrophic methanogens isvery important for rapid and complete conversion of propi-onate. In the UASB experiments adverse effects of butyrateon the propionate conversion were observed only during thefirst 3 days after the first addition of butyrate to the sub-strate.Growth of the propionate-oxidizing enrichment culture
was affected by acetate to a considerable degree (Fig. 5).This was also the case, although to a lesser extent, for themethanogenic consortia in the UASB reactor. In the lattersystem exposure to high acetate concentrations is possibleonly in certain compartments of the sludge. Addition ofacetate to the influent of the UASB reactor immediatelyresulted in a sharp increase in the propionate concentrationin the effluent, which was accompanied by a high acetateconcentration (Fig. 2B). The acetate conversion capacity ofthe sludge bed increased during the period when acetate wasadded to the influent. However, despite that, propionatedegradation decreased further even when almost completeacetate removal was achieved by the methanogenic consor-tium. On the basis of the substrate conversion kineticsequation given above, we might expect a recovery of propi-
onate degradation whenever the acetate concentration in thesludge bed drops. The reason that such an effect was notfound is not clear. One explanation might be that the numberof hydrogenotrophic methanogens in the sludge bed haddecreased, resulting in a lower hydrogen removal efficiency.However, also in that case, recovery of propionate removalmight be expected because of the high growth rate of thethermophilic methanogens (4). Inhibition of propionate con-version by extremely low concentrations of acetate in amesophilic methanogenic fluidized bed reactor was de-scribed by Gorris et al. (11). However, it is not clear whetherin that study a causal connection existed between theobserved effects.Hydrogen, an important intermediate of thermophilic pro-
pionate conversion, has two effects. For thermodynamicreasons a high level of hydrogen directly inhibits oxidation ofpropionate to acetate and carbon dioxide (Table 1). On theother hand, a high concentration of hydrogen stimulates thegrowth of hydrogenotrophic methanogens. The latter areessential for the development of a stable and dense propi-onate-oxidizing subpopulation in which the acetogens are inclose proximity to the methanogens. Results obtained hereindicate that propionate oxidation is stimulated by otherhydrogen-producing conversions of nontoxic intermediates(i.e., butyrate). In a previous study on thermophilic propi-onate conversion conducted in our department, it was pro-posed that a two-step digestion process should be appliedwhenever propionate is a major compound and hydrogenproduction is inevitable during the thermophilic treatment ofwastewater (28). In the second reactor of this system propi-onate would be removed. The overall better performancewas attributed to a decreased hydrogen partial pressure inthe second reactor, which for thermodynamic reasons stim-ulates propionate oxidation. Our results show that in addi-tion to a low hydrogen concentration, a low level of acetateseems to be indispensable for effective degradation of pro-pionate. It should be noted that both conditions can beachieved easily in the above-mentioned digestion processconsisting of two or more steps. In this study we foundsevere inhibition of propionate oxidation without any in-crease in the hydrogen partial pressure in the biogas. There-fore, the suitability of hydrogen as an overall control param-eter for anaerobic digestion (26) has to be reconsidered, aconclusion which was recently also drawn by other workers(13).
ACKNOWLEDGMENTS
This research was supported by the Netherlands Ministry ofHousing, Physical Planning, and Environment (VROM) Novem-grant 51120/1510, by the Royal Netherlands Academy of Arts andSciences, and by Paques Environmental Technology, Balk, TheNetherlands.
REFERENCES1. Ahring, B. K., and P. Westermann. 1987. Thermophilic anaer-
obic degradation of butyrate by a butyrate-utilizing bacterium incoculture and triculture with methanogenic bacteria. Appl.Environ. Microbiol. 53:429-433.
2. Ahring, B. K., and P. Westermann. 1988. Product inhibition ofbutyrate metabolism by acetate and hydrogen in a thermophiliccoculture. Appl. Environ. Microbiol. 54:2393-2397.
3. American Public Health Association. 1985. Standard methods forthe examination of water and wastewater, 16th ed. AmericanPublic Health Association, Washington, D.C.
4. Balch, W. E., G. E. Fox, L. J. Magrum, C. R. Woese, and R. S.Wolfe. 1979. Methanogens: reevaluation of a unique biologicalgroup. Microbiol. Rev. 43:260-296.
APPL. ENVIRON. MICROBIOL.
on May 31, 2018 by guest
http://aem.asm
.org/D
ownloaded from

THERMOPHILIC ANAEROBIC DEGRADATION OF PROPIONATE 1011
5. Boone, D. R. 1982. Terminal reactions in anaerobic digestion ofanimal waste. Appl. Environ. Microbiol. 43:57-64.
6. Boone, D. R., and M. P. Bryant. 1980. Propionate-degradingbacterium, Syntrophobacter wolinii sp. nov., gen. nov., frommethanogenic ecosystems. Appl. Environ. Microbiol. 40:626-632.
7. Boone, D. R., and L. Xun. 1987. Effects of pH, temperature, andnutrients on propionate degradation by a methanogenic enrich-ment culture. Appl. Environ. Microbiol. 53:1589-1592.
8. Chang, R. 1977. Physical chemistry with applications to biolog-ical systems. Macmillan Publishing Co., New York.
9. Durre, P., H. Bahl, and G. Gottschalk. 1988. Membrane pro-cesses and product formation in anaerobes, p. 187-206. In L. E.Erickson and D. Y.-C. Fung (ed.), Handbook on anaerobicfermentations. Marcel Dekker, Inc., New York.
10. Fukuzaki, S., N. Nishio, M. Shobayashi, and S. Nagai. 1990.Inhibition of the fermentation of propionate to methane byhydrogen, acetate, and propionate. Appl. Environ. Microbiol.56:719-723.
11. Gorris, L. G. M., J. M. A. van Deursen, C. van der Drift, andG. D. Vogels. 1989. Inhibition of propionate degradation byacetate in methanogenic fluidized bed reactors. Biotechnol.Lett. 11:61-66.
12. Henson, J. M., and P. H. Smith. 1985. Isolation of a butyrate-utilizing bacterium in coculture with Methanobacterium ther-moautotrophicum from a thermophilic digester. Appl. Environ.Microbiol. 49:1461-1466.
13. Kidby, D. W., and D. B. Nedwell. 1991. An investigation into thesuitability of biogas hydrogen concentration as a performancemonitor for anaerobic sewage sludge digesters. Water Res.25:1007-1012.
14. Lin, C.-Y., K. Sato, T. Noike, and J. Matsumoto. 1986. Metha-nogenic digestion using mixed substrate of acetic, propionic,and butyric acids. Water Res. 20:385-394.
15. Mawson, A. J., R. L. Earle, and V. F. Larsen. 1991. Degradationof acetic and propionic acids in the methane fermentation.Water Res. 25:1549-1554.
16. McInerney, M. J. 1988. Anaerobic hydrolysis and fermentationof fats and proteins, p. 373-415. In A. J. B. Zehnder (ed.),Biology of anaerobic microorganisms. John Wiley, New York.
17. McInerney, M. J., M. P. Bryant, and N. Pfennig. 1979. Anaer-
obic bacterium that degrades fatty acids in syntrophic associa-tion with methanogens. Arch. Microbiol. 132:129-135.
18. Mucha, H., F. Lingens, and W. Trosch. 1988. Conversion ofpropionate to acetate and methane by syntrophic consortia.Appl. Microbiol. Biotechnol. 27:581-586.
19. Nanba, A., R. Nukada, and S. Nagai. 1983. Inhibition by aceticand propionic acids of the growth of Propionibacterium sher-manii. J. Ferment. Technol. 61:551-556.
20. Nelder, J. A., and R. Mead. 1965. A simplex method for functionminimization. Comput. J. 8:308-313.
21. Saez, P. B., and B. E. Rittman. 1992. Model-parameter estima-tion using least squares. Water Res. 26:789-796.
22. Sillen, L. G., and A. E. Martell. 1964. Stability constants ofmetal ion complexes. The Chemical Society, London, GreatBritain.
23. Stams, A. J. M., K. C. F. Grolle, C. T. M. J. Frijters, and J. B.Van Lier. 1992. Enrichment of a thermophilic propionate-oxidizing acetogenic bacterium in coculture with Methanobac-terium thermoautotrophicum or Methanobacterium thermo-formicicum. Appl. Environ. Microbiol. 58:346-352.
24. Thauer, R. K., K. Jungermann, and K. Decker. 1977. Energyconservation in chemotrophic anaerobic bacteria. Bacteriol.Rev. 41:100-180.
25. van Lier, J. B., K. C. F. Grolle, A. J. M. Stams, E. Conway deMacario, and G. Lettinga. 1992. Start-up of a thermophilicupflow anaerobic sludge bed (UASB) reactor with mesophilicgranular sludge. Appl. Microbiol. Biotechnol. 37:130-135.
26. Whitmore, T. N., D. Lloyd, G. Jones, and T. N. Williams. 1987.Hydrogen-dependent control of the continuous anaerobic diges-tion process. Appl. Microbiol. Biotechnol. 26:383-388.
27. Wiegant, W. M., J. A. Claassen, and G. Lettinga. 1985. Ther-mophilic anaerobic digestion of high strength wastewaters.Biotechnol. Bioeng. 27:1374-1381.
28. Wiegant, W. M., M. Hennink, and G. Lettinga. 1986. Separationof the propionate degradation to improve the efficiency ofthermophilic anaerobic treatment of acidified wastewaters. Wa-ter Res. 4:517-524.
29. Zehnder, A. J. B. 1978. Ecology of methane formation, p.349-376. In R. Mitchell (ed.), Water pollution microbiology,vol. 2. John Wiley, New York.
VOL. 59, 1993
on May 31, 2018 by guest
http://aem.asm
.org/D
ownloaded from