effect of scaffold branch girdling and various bud-growth
TRANSCRIPT

Effect of scaffold branch girdling andvarious bud-growth stimulatory
treatments on flowering time, intensityand variance, and on inflorescence
set in Mauritius litchi
HortResearch SA, POBox 3849, Tzaneen 0850, South AfricaE-mail: [email protected]
ABSTRACTThe main scaffold branches of bearing HLH Mauritius (Tai So) litchi trees were either girdled (cinctured)or left. To girdled or non-girdled trees, low-biuret urea, potassium nitrate, or amino acid complexwere sprayed in mid-June 2003, prior to flowering in July. Furthermore, paclobutrazol was applied tothe soil under the canopy in mid-March, 2003, the roots were pruned in early June, 2003, or theterminal branches were headed back by 30 to 45 cm in mid-June, 2003. The winter conditions(temperatures) were strongly inductive, this presumably masking any treatment benefit of stimula-ting bud-development and increasing flowering intensity. Winter pruning or girdling delayed bud-development, reduced flowering intensity, and increased flowering stage variance. These effectswere pronounced following winter pruning. Girdling markedly increased the number of fruits set perinflorescence and tree yield. Headed branched terminated by more leaves showed a greater propen-sity to initiate inflorescences as opposed to a new shoots. Flowering was more intense, set perinflorescence better, and the initiation of bud-development earlier on the sun-exposed as opposed tothe more shaded side of the trees, this indicating the importance of canopy sun-exposure for in-creased tree performance. Potassium nitrate spraying increased inflorescence stage of developmentvariance. Soil paclobutrazol application was ineffective in influencing inflorescence developmentstage or variance, or flowering intensity or fruit set. An effect in this regard would be expected whenenvironmental conditions are less effective in inducing flowering. Repeating the study when winterconditions are less inductive was deemed necessary.
INTRODUCTIONHLH Mauritius (Tai So) litchi trees grown in thenorthern regions of South Africa often fail toflower or flowering is not intense, it occurringonly on portions of the canopy. Inflorescencesare often vegetative and inflorescence develop-ment accompanied by new shoot growth. Fail-ure to flower has been associated with bud-de-velopment not being synchronous, it occurringover a period only partially overlapping that whenenvironmental conditions are most inductive forflowering (Oosthuyse, 2003). The general in-capacity of trees to flower has also been as-cribed to an environment that is generally onlymarginally inductive for flowering (Oosthuyse,2003). Intense flowering occurs during colderwinters, and is coupled by concentrated bud-development. Increased fruit set is noted tooccur when full-bloom arises slightly later in theseason, when conditions are warmer(Oosthuyse, 2003).
The months during which the night tempera-tures generally fall below lOoe are consideredto be inductive, i.e., the months of May, Juneand July. During this period, day temperaturesexceeding 250e are often experienced. Inter-posed warm periods, when day and night tem-peratures increase, are also experienced. Dailyaverage temperatures increase rapidly duringAugust.
Temperatures exceeding 200e during the pe-riod of flower initiation reduce flowering (Menzeland Simpson, 1995). Temperatures of less than200e promote flowering (Menzel and Simpson,1990).
In the present study, various treatments to ef-fect concentrated bud-development during Julywere evaluated. Soil paclobutrazol application,winter branch or root pruning, scaffold branchgirdling, and spray application of an amino acidcomplex, potassium nitrate, or low biuret ureawere assessed. The effect on the time of bud-

development and flowering, flowering stage vari-ance, flowering intensity, and number of fruitsretained per inflorescence was quantified.
MATERIALS AND METHODSOn 19 March 2003, seventy bearing Mauritiuslitchi trees, in a number rows, in an orchard blockon Bender Farm (Charlie Bender, Hamawasha,Tzaneen) were selected for size-uniformity. Oneach of 10 trees, the folloWing treatments wereindividually carried out (the treatment abbrevia-tions are provided):
La - Untreated controlL1 - On March 20, 2003, 28 ml of Cultar(paclobutrazol) in 20 liters of water was appliedto the soil under the canopy (after the post-har-vest flush).
L2 - On June 11, 2003, the amino acid com-plex, Maxigrow, was spray applied until run-off.500 ml of the complex and 15 ml of Citowett(adjuvant) were added per 100 I of water.
L3 - On June 11, 2003, potassium nitrate wasspray applied until run-off. 4 kg of SQM potas-sium nitrate and 15 ml of Citowett were addedper 100 I of water.
L4 - On June 11, 2003, low-biuret urea wasspray applied until run-off. 1 kg of low-biureturea and 15 ml of Citowett were added per 100I of water.
Ls- On June 3, 2003, a trench 30 cm deep and30 cm wide was made around the trunk to ef-fect root pruning (Figure 1).
L6 - On June 10 and 11, 2003, the terminalbranches were headed back by 30 to 45 cm (Fig-ure 2).
From March 24, until April 1, 2003, the mainscaffold branches (the branches arising from thetrunk), of half of the trees used were girdled(Split-plot design), the girdles being approxi-
Figure 1. Trench made around the trunkto effect root pruning. The trench wasfilled with soil a day or two after it wasdug.
Figure 2. Terminal branch heading em-ployed to stimulate and concentrate bud-development. Flowering (bud-develop-ment) commenced inearly to mid-July. Thewinter was cold, in relative terms, beingsufficiently inductive for intense flowering.The trees all flowered intensely and syn-chronously during July and August, 2003.
mately 1 mm in width (cincturing). Each branchwas entirely circumvented, one girdle (cincture)being made per branch.
The sprays were applied until run-off in thelate afternoon or early morning when theevaporative demand was reduced. The treat-ments were carried out three to four weeks priorto the commencement of flowering. On June 2,2003, before the start of flowering, 20 branchends per tree were labeled. Data concerningstage of inflorescence or shoot development, orfruit set, were collected from these branches.
Figure 3. Flowering stage in the advancedtrees on July 15, 2003.

Figure 4. A headed branch from whichdata was collected to assessthe relation-ship between subtending leaf number andthe ability of a branch to initiate inflores-cences as opposed to new shoots (photo-graph taken on July 15, 2003).
95 Perced LSDIntovals foc Facllr l'vt:an5
o 1 234 5 6Trestment
Figure 5. Differences in terminal out-growth length in relation to treatment. "0"= control; "1" = soil paclobutrazol; "2" =spray amino acid complex; "3" = spraypotassium nitrate; "4" = spray LB urea;"5" = root pruning; "6" = winter heading.
Figure 3 shows the stage of flowering of someof the advanced trees on July 15, 2003.
On August 6, 2003, when inflorescence elon-gation was general, the length of the longest newstructure, whether a shoot or inflorescence, oneach of the labeled branches was recorded. OnSeptember 23, 2003, the stage of flowering ofthe most advanced inflorescence per labeledbranch was identified according to the classifica-tions proposed by Oosthuyse (1991) for mango.Each stage was converted to an index for analy-sis, the indices varying from 0 (dormant bud) to1 (post petal-fall). On November 17, 2003, thenumber of fruits on the most distal inflorescenceon each labeled branch was counted. The loca-tion of the labeled branches, whether on the sun-exposed or more shaded half of the tree, wasrecorded. The data collected from each side wereanalyzedto detect possibledifferences in responserelating to tree-aspect.
The trees were irrigated throughout the win-ter. Fertilizer was applied at or shortly after har-vest, fertilization having been based on the re-sults of leaf analysis for the purpose of rectifyingbalance.
The tree averages were subjected to analysisof variance. Variance reductions due to La to L6,
girdling, and girdling versus treatment interac-tion, were determined. Mean separation in re-spect of treatment (La to L6) was assessedon the5% least significant difference (LSD) criterion.
The relationship between number of leavesterminating a branch, and the branch's ability toinitiate an inflorescence was explored. Varia-tion in leaf number was created by the action ofheading (Figure 4). The number of leaves within20 cm of the cut-end of 60 branches was countedon August 13, 2003. Whether an inflorescenceor new shoot terminated each branch was addi-tionally recorded.
RESULTSFlower inductive conditions experiencedduring thewinter months (May, June and July) were ad-equately strong, since flowering was intense andsynchronous. The desired treatment effect, beingthat of concentratedand general bud-developmentduring the period when conditions are maximallyinductive, would, expectantly, have been disguisedby the overriding effect of the environmental con-ditions. Nonetheless,differences relating to treat-ment (~ to L6) and girdling were found.
I. Growth stage and growth stage vari-ance on August 6, 2003Figures 5 and 6, and Table 1, show the differ-ences in terminal out-growth length relating totreatment or girdling.

Reduced terminal out-growth length variancemay have been associated with soil paclobutrazoltreatment in late March, otherwise, differences re-lating to treatment or girdling were not apparent.
II. Inflorescence development stage andstage-variance on September 23, 2003In considering inflorescence development stage,the labeled branched not having flowered wereexcluded from the analysis.
Figures 7 and 8 show the differences in inflo-rescence stage of development in relation totreatment and girdling.
Flowering was less advanced on the moreshaded as opposed to the sun-exposed side ofthe trees. Branch heading delayed flowering,inflorescence development being markedly lessadvanced in the headed branches. Scaffoldbranch girdling delayed flowering slightly.
Table 2 shows the inflorescence developmentstage variance in relation to treatment andgirdling.
Of the treatments, L1 - L6, increased inflores-cence stage variance was associated with sprayapplication of potassium nitrate and winter prun-
Table 1. Terminal out-growth length on August 6, 2003.
oGirdlilYJ (1)
Figure6. Difference interminal out-growthlength in relation to whether girdling wascarried out (1) or not (0).
Treatment Growth (cm)- Growth (cm)- Growth (cm)-whole tree exposed side shaded side
La -control 13.05a 13.36ab 12.69a
L1 - soil paclobutrazol 11.43a 1l.89ab 1O.38a
L2 - spray amino acid complex 13.38a 14.84a 12.20a
L3 - spray potassium nitrate 12.27a 12.76ab 12.26a
L4 - spray low biuret urea 12.34a 13.73a 11.09a
Ls - root pruning 10. 14ab 11. 16ab 9.09a
L6 - winter heading 6.18b 8.47b 3.93b
Girdling (0) - absence 14.25a 14.86a 13.75a
Girdling (1) - presence 8.26b 9.77b 6.72b
Interaction (L x G) - significance level 0.0475* 0.3094n5 0.0051 **
Bud-development was less advanced on themore shaded as opposed to the sun-exposedside of the trees. Winter heading delayed bud-development, the extent of terminal out-growthbeing markedly reduced in the headed branches.Scaffold branch girdling delayed bud-develop-ment. Interaction between treatment (La - L6)
and girdling (G) was significant due to the delayin bud-development in the headed branches be-ing similar, with respect to whether girdling wascarried out or not (data not shown).
Table 2 shows the out-growth length variance inrelation to treatment and girdling.
ing. Girdling also increased inflorescence stagevariance. Significant interaction resulted from thedifference in variance being negligible regardinggirdling in the trees sprayed with the amino acidcomplex and that in the trees whose brancheswere headed in winter (data not shown).
III. Tree flowering intensity and fruitretention (data recorded on November17,2003)Figures 9 and 10 show the differences in flow-ering branch percentage in relation to treatmentand girdling.

Treatment Growth Var. - Growth Var. - Growth Var. -
whole tree exposed side shaded side
La -control 23.52a 22.83ab 22.15a
L1 - soil paclobutrazol 19.60a 14.68a l1,43a
L2 - spray amino acid complex 27.88a 27.87ab 22.68a
L3 - spray potassium nitrate 32.26a 34.36b 22.34a
L4 - spray low biuret urea 22.72a 21,25ab 15.32a
Ls - root pruning 21,17a 20.99ab l1,57a
L6 - winter heading 34.56a 29.47ab 19.25a
Girdling (0) - absence 26.99a 24.61a 20.57a
Girdling (1) - presence 24.93a 24.38a 15.08a
Interaction (L x G) - significance level 0.5369n5 0.2541n5 0.0698n5
Table 5 shows the percentages of labeledbranches flowering in relation to tree aspect,treatment and girdling.
The branches on the more shaded side of thetrees were slightly less inclined to initiate inflo-rescences, the difference being greatest in theheaded trees. Winter pruning resulted in amarked reduction in the proportion of branchesinitiating inflorescences. Differences in relation
95 Perced LSDlntlw.l1s foc Factoc Ntans
/186
I I Ij /181!i /176
·r /171
Ii!:
/166
/161o 1 234 5 6
TreatrrIed
Figure 7. Differences in stage of inflores-cence development in relation to treat-ment. "0" = control; "1" = soil paclobu-trazol; "2" = spray amino acid complex; "3"= spray potassium nitrate; "4" = spray LBurea; "5" = root pruning; "6" = winterheading.
to treatments L1 to Ls were not apparent. Thebranches on the girdled trees initiated a lesserpercentage of inflorescences than the brancheson the non-girdled trees. Significant interactionresulted from an inconsistent inclination for the"more shaded" branches on the girdled trees toinitiate a lower proportion of inflorescences thanthe "more shaded" branches on the non-girdledtrees (data not shown).
Figures 12 and 13 show the differences innumber of set fruits per "dominant" inflorescencein relation to treatment and girdling.
Table 6 shows the total number of fruits set
95 Perced LSDlntlw.l1s foc Factoc Ntans
/183j!i /179
·ri!:
/175
o 1
Girdling (1)
Figure 8. Difference in stage of inflores-cencedevelopment in relation to whethergirdling was carried out {1} or not {O}.

Treatment Stage (index) - Stage (index) - Stage (index) -
whole tree exposed side shaded side
La - control 0.80a 0.78a 0.79a
L1 - soil paclobutrazol 0.82a 0.85a 0.76a
L2 - spray amino acid complex 0.84a 0.85a 0.78a
l, - spray potassium nitrate 0.79a 0.81" 0.71"L4 - spray low biuret urea 0.83a 0.86a 0.78a
Ls - root pruning 0.78a 0.81" 0.75a
L6 - winter heading 0.66b 0.59b 0.33b
Girdling (0) - absence 0.84a 0.84a 0.76a
Girdling (1) - presence 0.74b 0.75b 0.64b
Interaction (L x G) - significance level 0.3584n5 0.8337n5 0.0831n5
Treatment Stage (index) - Stage (index) - Stage (index) -
whole tree exposed side shaded side
La -control 0.014a 0.038ab 0.007a
L1 - soil paclobutrazol 0.030ab 0.024a 0.025ab
L2 - spray amino acid complex 0.006a 0.027a 0.015a
L3 - spray potassium nitrate 0.044bc 0.057ab 0.055b
L4 - spray low biuret urea O.013a 0.018a 0.008a
Ls - root pruning 0.024ab 0.026a O.014a
L6 - winter heading 0.057c 0.080b 0.114C
Girdling (0) - absence 0.010a 0.022a 0.023a
Girdling (1) - presence 0.044b 0.056b 0.045b
Interaction (L x G) - significance level 0.1609n5 0.8980n5 0.0159*
Treatment Flowering Flowering Flowering
branches (%) - branches (%) - branches (%)-
whole tree exposed side shaded side
La -control 87.0a 79.5ab 93.8a
L1 - soil paclobutrazol 89.5a 91.8a 84.2a
L2 - spray amino acid complex 90.5a 91.1" 90.3a
L3 - spray potassium nitrate 84.0a 88.1" 81.1"
L4 - spray low biuret urea 94.5a 96.1" 93.5a
Ls - root pruning 93.5a 94.6a 92.6a
L6 - winter heading 45.5b 63.0b 30.4b
Girdling (0) - absence 87.0a 90.4a 84.2a
Girdling (1) - presence 80.0b 82.3b 77.5b
Interaction (L x G) - siqnificance level 0.1860n5 0.8576n5 0.0381*

95 Percent LSDInteMl1s foc Fact<r M:ans
I I I I I I
o 1 234 5 6TreatrnerL
Figure 9. Differences in flowering branchpercentage in relation to treatment. "0"= control; "1" = soil paclobutrazol; "2" =spray amino acid complex; "3" = spraypotassium nitrate; "4" = spray LB urea;"5" = root pruning; "6" = winter heading.
95 Percent LSDInteMl1s foc Factoc M:ans
92
89
~L
i 86
'f 83
~
80
TT
0 1
Girdling (1)
Figure 10. Difference in flowering branchpercentage in relation to whether girdlingwas carried out (1) or not (0).
23456TreatrnerL
Figure 11. Differences in number of fruitsper inflorescence in relation to treatment."0" = control; "1" = soil paclobutrazol; "2"= spray amino acid complex; "3" = spraypotassium nitrate; "4" = spray LB urea;"5" = root pruning; "6" = winter heading.
95 Percent LSDIntav<is foc Fador M:ans
6.8 I~~ 58
j4.8
3.8 I0 1
Girdling (1)
Figure 12. Difference in average numberof fruits set per inflorescence in relationto whether girdling was carried out (1) ornot (0).
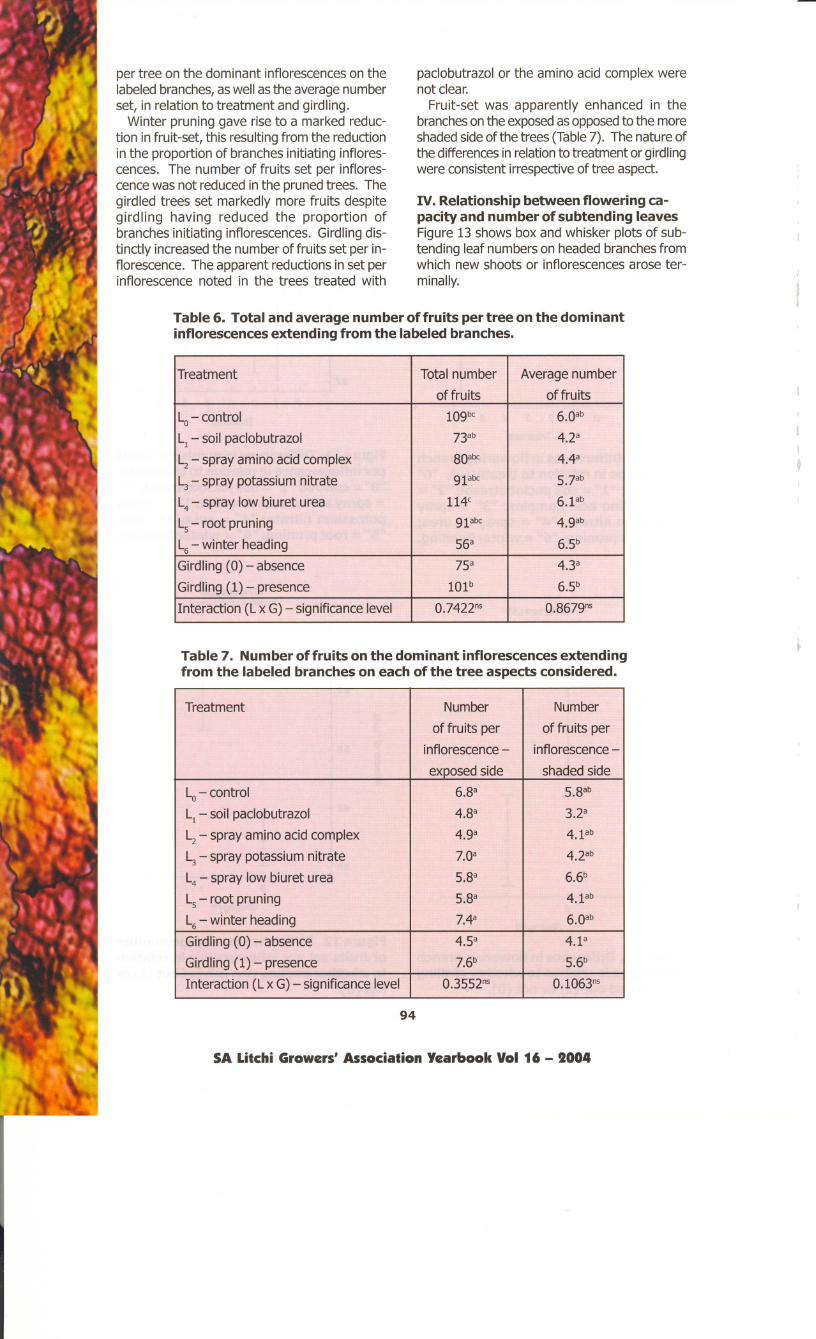
per tree on the dominant inflorescences on thelabeled branches, as well as the average numberset, in relation to treatment and girdling.
Winter pruning gave rise to a marked reduc-tion in fruit-set, this resulting from the reductionin the proportion of branches initiating inflores-cences. The number of fruits set per inflores-cence was not reduced in the pruned trees. Thegirdled trees set markedly more fruits despitegirdling having reduced the proportion ofbranches initiating inflorescences. Girdling dis-tinctly increased the number of fruits set per in-florescence. The apparent reductions in set perinflorescence noted in the trees treated with
paclobutrazol or the amino acid complex werenot clear.
Fruit-set was apparently enhanced in thebranches on the exposed as opposed to the moreshaded side of the trees (Table 7). The nature ofthe differences in relation to treatment or girdlingwere consistent irrespective of tree aspect.
IV. Relationship between flowering ca-pacity and number of subtending leavesFigure 13 shows box and whisker plots of sub-tending leaf numbers on headed branches fromwhich new shoots or inflorescences arose ter-minally.
Table 6. Total and average number of fruits per tree on the dominantinflorescences extending from the labeled branches. .
Treatment Total number Average number
of fruits of fruits
La -control 109be 6.0ab
L1 - soil paclobutrazol 73ab 4.2a
L2 - spray amino acid complex sOabe 4.4a
L3 - spray potassium nitrate 910be 5.7ab
L4 - spray low biuret urea 114e 6.1Ob
Ls - root pruning 910be 4.9ab
L6 - winter heading 56a 6.5b
Girdling (0) - absence 75a 4.3a
Girdling (1) - presence 101b 6.5b
Interaction (L x G) - significance level 0.7422n5 0.S679n5
Table 7. Number of fruits on the dominant inflorescences extendingfrom the labeled branches on each of the tree aspects considered.
Treatment Number Number
of fruits per of fruits per
inflorescence - inflorescence -
exposed side shaded side
La - control 6.Sa 5.Sab
L1 - soil paclobutrazol 4.Sa 3.2a
L2 - spray amino acid complex 4.9a 4.1Ob
L3 - spray potassium nitrate 7.0a 4.2ab
L4 - spray low biuret urea 5.Sa 6.6b
Ls - root pruning 5.Sa 4.1Ob
L6 - winter heading 7.4a 6.0ab
Girdling (0) - absence 4.5a 4.10
Girdling (1) - presence 7.6b 5.6b
Interaction (L x G) - significance level 0.3552n5 0.1063n5

11
9
~~ 7-I
.~~ 5'§CI).•.0
~3
i1
£be and W1isk« Plc:tsfarFa;UL=:! ilia
S F
Sh:x1 (5) or Infforescetre (FJ
Figure 13. Box and whisker plots of sub-tending leaf numbers on headed branchesfrom which new shoots or inflorescencesarose terminally.
Headed branches on which greater numbersof leaves remained after heading were more in-clined to initiate inflorescences as opposed tonew shoots.
DISCVSSIONANDCONaUSIONThe winter conditions during the period of thestudy were strongly inductive for HLH Mauritus,evidenced by uniform and intense flowering oc-curring during July, August and September, 2003.An effect of soil paclobutrazol treatment, orspray treatment with the amino acid complex,potassium nitrate or low-biuret urea, or root orbrand pruning, all carried out to stimulate andconcentrate bud-development, would expectantlyhave been overridden. Nonetheless, treatmentdifferences were found. Studies having the ob-jective of stimulating and concentrating bud-de-velopment in litchi have not been considered pre-Viously to the knowledge of the author. Refer-ence for discussion is thus limited.
Bud-development was delayed slightly bygirdling (scaffold branch cincturing) or winterpruning. Variance in development-stage was in-creased as a result of potassium nitrate treat-ment, winter pruning, or scaffold-branch girdling.These effects were not marked, however.
Tree flowering intensity was markedly reducedin the pruned trees, the headed branches ter-minated by more leaves having a greater pro-
pensity to initiate inflorescences as opposed tonew shoots. Menzel et al. (1996), in cutting backterminal shoots of Bengal, Kwai May Pink or WaiChee trees by about 30 cm in winter or summer,noted that heading in winter seldom promotedflushing. Flowering and yield were as good af-ter winter pruning as after summer pruning. Inour study, winter heading is certain instancesgave rise to almost leafless branches. The re-duction in flowering intensity probably relatedto severer heading.
Winter pruning delayed bud-development, andincreased inflorescence stage of developmentvariance observed in late September when flow-ering was advanced. It may thus not be consid-ered a suitable treatment to stimulate and con-centrate bud-development. That the number offruits set per inflorescence was not reduced, mayrelate to a favourable leaf to fruit ratio in thepruned trees during the fruit growth and devel-opment period (Roe, et al., 1995).
The headed branches with greater numbersof subtending leaves were more prone to initi-ate inflorescences than those with lesser num-bers of subtending leaves, indicating the impor-tance of sufficient functional leaves close to thepoint of bud-development for inflorescence ini-tiation to occur when environmental conditionsare sufficiently inductive for flowering.
In general, girdling in litchi has been found toreduce root and shoot growth, to increase flow-ering intensity, to increase fruit-set, to reducefruit drop, and to improve fruit appeal (Roe, etal., 1995; Zhou et al., 1996). In our study, scaf-fold branch girdling was highly effective in in-creasing the number of fruits per inflorescence.Quantification of the effect was done in mid-No-vember, after the period of "natural" fruit drop.Tree yield was increased despite the slight re-duction in flowering intensity caused by girdling.Inconsistency with respect to increased flower-ing intensity resulting from girdling has beenfound preViously (Koo-Duang and Subhadra-bandhu, 1987). Morse and Oosthuizen (1993)advised that only healthy, vigorous trees with astrong post-harvest flush be girdled. The rateof healing of the girdle (cincture) may be of is-sue. Delayed healing, resulting from too wide acut, my be detrimental, whereas, too narrow agirdle may not render a marked effect due torapid healing. Healing rate should be consid-ered in a further study.
Girdling delayed bud-development and in-creased the variance in inflorescence stage ofdevelopment. It might thus not be considered atreatment to stimulate and concentrate bud-de-velopment.
Menzel and Simpson (1990) reported an in-

crease in flowering intensity resulting frompaclobutrazol treatment. In their study, soilpaclobutrazol treatment reduced flushing andincreased flowering in five out of eight orchards,maintained dormancy and reduced flowering inone orchard, and had variable effects in twoorchards, depending on the method of applica-tion. Soil applications were generally more ef-fective than foliar sprays at controlling vegeta-tive growth and promoting flowering. The maxi-mum level of flowering in paclobutrazol-treatedtrees occurred when the control trees floweredmoderately (40-60% of terminal branches). Theresponses were sometimes weak when the treeswere vegetative «30% flowering). Paclo-butrazol had no significant effect or reducedflowering in heavily flowering (70%-100% flow-ering) trees. Paclobutrazol had only a small ef-fect on panicle development, fruit set or fruitquality at most of the sites. Yield generally re-flected the flowering response to paclobutrazol.These findings clarify that paclobutrazol treat-ment can have a pronounced effect on litchi treephenology, particularly when environmental con-ditions do not favour intense flowering. In ourstudy, where environmental conditions wereconducive to intense flowering, paclobutrazol soiltreatment had no apparent effect on time of bud-development, time of bud-development vari-ance, or flowering intensity. A reduction innumber of fruits retained per inflorescence ap-peared likely. Further assessment of paclo-butrazol to concentrate bud-development andenhance flowering intensity may thus be seento be necessary.
Root pruning in litchi has not been previouslyconsidered in terms of its effect in stimulatingbud-development or increasing flowering inten-sity. Its effect when conditions are less induc-tive should, in the view of the author, receiveconsideration.
Flowering was more intense, set per inflores-cence better, and the commencement of bud-development earlier on the sun-exposed as op-posed to the more shaded side of the trees. Thisindicates the importance of adequate canopy sun-expose for enhanced tree performance. Con-curring results have been found in other sub-tropical fruit or nut trees (Trochoulias, 1987;Deidda et al., 1988; Bastawros, 1993; Garcia etal., 1995).
Repeating this study when environmental con-ditions are less favourable for flowering of HLHMauritius litchi would appear to be necessary.Summer pruning, and the application of thio-urea, cytokinin and cycloheXimide in late Mayare treatments that may also hold promise. Thequantification of the effect of girdling on f1ower-
ing intensity and tree yield during warmer win-ters also requires attention in the view of theauthor.
UTERAnJRECITEDBASTAWROS,M.B. 1993. Effect of shading on vegeta-
tive growth, flowering and fruit characters of man-goes. Egyptian Journal of Agricultural Research 71:997-1006.
DEIDDA, P., FILIGHEDDU,M.R., CANU,A., FARRO,L. &BENINCASA, F. 1988. Light distribution on citruscanopy affects physiological parameters and fruitingpattern. Annali delia Facolta di Agraria dell'Universita
Sassari, Studi-Sassaresi 34: 115-122.GARCIALUIS, A., FORNES,F. & GUARDIOLA,J.L. 1995.
Leaf carbohydrates and flower formation in Citrus.Journal of the American Society for Horticultural Sci-
ence 120: 222-227.KOO DUANG,A. & SUBHADRABANDHU,S. 1987. Effects
of branch girdling on total nonstructural carbohydratecontent in relation to flowering of two Iychee (Litchi
chinensis Sonn.) cultivars. Kasetsart Journal of Natu-
ral Sciences 21: 132-141.MENZEL, CM. & SIMPSON, D.R. 1990. Effect of
paclobutrazol on growth and flowering of Iychee (Li-
tchi chinensis). Australian Journal of Experimental
Agriculture 30: 131-137.MENZEL,eM. & SIMPSON,D.R. 1990. Effect of environ-
ment on growth and flowering of Iychee (Litchi
chinensis Sonn.). Acta Hort 275: 161-166.MENZEL.eM. & SIMPSON, D.R. 1995. Temperatures
above 20°C reduceflowering in Iychee(Litchi chinensis
Sonn.) Journal of Horticultural Science 70: 981-987.MENZEL,eM., SIMPSON, D.R. & DOOGAN,VJ. 1996.
Preliminary observations on growth, flowering andyield of pruned Iychee trees. Journal of the Southern
African Society for Horticultural Sciences 6: 16-19.MORSE,R.L. & OOSTHUIZEN,J.H. 1993. Litchi flower
manipulation by girdling. South African Litchi Grow-
ers' Association Yearbook 5: 10-11.OOSTHUYSE,SA 1991. Stages of development of the
mango panicle. South African Mango Growers' Asso-
ciation Yearbook 11: 59-61.OOSTHUYSE,SA 2003. Litchi tree manipulation to en-
sure health and regular cropping. Litchi SA, 2003,
No.1, pp. 1-8.ROE, DJ., MENZEL, eM. & OOSTHUIZEN, J.H. 1995.
Litchi fruit growth depends on current CO2 assimila-tion rather than on stored reserves. South African
Litchi Growers' Association Yearbook 7: 11-16.TROCHOULIAS, T. 1987. High density plantings of
macadamia. Proceedings of the Second AustralianMacadamia Research Workshop, Bangalow PalmsResort, NSW, 15-19 Sep. pp. 155-164.
ZHOUXJ., WU D.Y.,HUANGH.B., LAI J. & HUANGD.G.1996. Manipulating growth and fruiting of young li-tchi trees by spiral girdling. Acta Horticulturae Sinica
23: 13-18.