effect of dl-nebivolol, its enantiomers and metabolites on the intracellular production of...
TRANSCRIPT

A
iaatAompsodRbait©
K
1
friiwtl
1d
Pharmacological Research 55 (2007) 303–309
Effect of dl-nebivolol, its enantiomers and metabolites on the intracellularproduction of superoxide and nitric oxide in human endothelial cells
Stefano Evangelista a,∗, Ulisse Garbin b, Anna Fratta Pasini b, Chiara Stranieri b,Veronica Boccioletti b, Luciano Cominacini b
a Preclinical Development Department, Menarini Ricerche spa, Via Sette Santi 1, 50131 Firenze, Italyb Department of Biomedical and Surgical Sciences (Internal Medicine D), University of Verona, Verona, Italy
Accepted 22 December 2006
bstract
Nebivolol, a third generation selective �1-adrenoceptor (�1-AR) antagonist, has been reported to reduce intracellular oxidative stress and tonduce the release of nitric oxide (NO) from the endothelium. Nebivolol is also subjected to a complex metabolic process where glucuronidation,romatic and alicyclic hydroxylation are the major pathways leading to several metabolites. We have studied the effect of nebivolol, its enantiomersnd metabolites on intracellular oxidative stress and NO availability in human umbilical vein endothelial cells (HUVECs). Furthermore, sincehe receptors involved in this endothelial effect of nebivolol remain controversial, we have studied this matter by the use of antagonists of �-R. dl-Nebivolol, d-nebivolol and l-nebivolol significantly reduced the formation of reactive oxygen species (ROS) and superoxide induced byxidized-low density lipoprotein (ox-LDL), although the racemic and l-form were significantly more active than d-nebivolol in this activity. Aarked decrease in the availability of intracellular NO was found in HUVECs exposed to ox-LDL and this parameter was normalized by the
rior incubation with dl-nebivolol, d-nebivolol and l-nebivolol; the effect of racemate was mainly mimicked by its l-enantiomer. eNOS activityignificantly increased by a 5-min contact of HUVECs with dl-nebivolol and l-nebivolol, but not with d-nebivolol, and a similar pattern wasbserved when the intracellular calcium increase was measured. The metabolites A2, A3′, A12 and A14 but not A1, A3 and R 81,928, derived fromifferent metabolic pathways, retained the antioxidant activity of the parent racemic compound dl-nebivolol, reducing the intracellular formation ofOS and superoxide. The effects of dl-nebivolol on intracellular formation of NO, eNOS activity and intracellular Ca2+ were partially antagonized
y the antagonists of �1-2-AR nadolol or by the �3-AR antagonist SR59230A and further antagonized by their combination or by (�1-2-3-ARntagonist bupranolol. In conclusion, this study shows that the NO releasing effect of nebivolol is mainly due to its l-enantiomer; the racemate andts enantiomers possess a remarkable antioxidant activity that contributes to its effect on the cellular NO metabolism and the activation of �3-ARhrough a calcium dependent pathway is involved in the mechanisms leading to the NO release.2007 Elsevier Ltd. All rights reserved.
ss; R
etdctta
eywords: �-Adrenergic receptor; Nebivolol; Nitric oxide (NO); Oxidative stre
. Introduction
Nebivolol is a third generation beta-blocker, highly selectiveor the �1-adrenoceptors (AR) and endowed with the ability toelease nitric oxide from the cardiovascular endothelium [1]. Its a racemic mixture of two enantiomers, d- and l-nebivolol, andts antihypertensive activity is mainly ascribed to d-nebivolol,
hich shows an over 100-fold greater affinity for �1-ARs thanhe l-enantiomer [2]. Both enantiomers [3], but particularly the-form [4,5], possess an endothelium-dependent vasorelaxant
∗ Corresponding author. Tel.: +39 055 5680519; fax: +39 055 5680510.E-mail address: [email protected] (S. Evangelista).
b�oi[oo
043-6618/$ – see front matter © 2007 Elsevier Ltd. All rights reserved.oi:10.1016/j.phrs.2006.12.008
eactive oxygen species (ROS)
ffect that can be antagonized by NO synthase inhibitors. Owingo these characteristics nebivolol possesses a peculiar pharmaco-ynamic profile producing vasodilation as observed in severalonditions and species (see Ref. [1] for review). Some recep-ors have been described as responsible for the activation ofhe endothelial effect of nebivolol, among them the �2- [6,7]nd �3-[8–11] ARs, the estrogen receptors of the plasma mem-rane [12], 5HT1A receptors [13] as commonly found for otherblockers, and the P2Y purinoceptor [14]. A further mechanism
f action is due to the antioxidant properties of nebivolol that can
ncrease the release of NO by reducing its oxidative inactivation15]. Reactive oxygen species (ROS) are reactive derivativesf O2 metabolism, including superoxide anion, hydrogen per-xide, hydroxyl radical and nitric oxide. It is well known that
3 logica
spi
nste
waAdhw
oos(arco
2
2
i2BMsBmSSUTCouesmwoca
letbw
m8imasc5ts
2
ucFst(CdvBlt[
2
fat[nfiAof(5dbtan(
2
04 S. Evangelista et al. / Pharmaco
uperoxide, can inactivate NO in a chemical reaction producingeroxynitrite, and nebivolol has been found to interfere with thisnactivation by enhancing the bioavailability of NO [15,16].
Indeed the increase in endothelial NO release induced byebivolol is highly dependent on the tissue/cell system andpecies examined, being possibly related to the different recep-or type and/or coupled transduction pathway activated in eachxperimental setting.
Nebivolol is also subjected to a complex metabolic processhere glucuronidation, aromatic and alicyclic hydroxylation
re the major pathways leading to several metabolites [17,18].mong the latter, it has been recently reported that the 4-ketoerivative metabolite produced from the alicyclic hydroxylation,as a similar pattern of activity on NO availability as comparedith nebivolol itself [15].Therefore, the aim of the present study is to evaluate the effect
f nebivolol, its enantiomers and metabolites on intracellularxidative stress and NO availability and the interrelation-hips between them in human umbilical vein endothelial cellsHUVECs), chosen as primary human culture and as knownnd widespread model to study NO [19]. Furthermore, since theeceptors involved in this endothelial effect of nebivolol remainontroversial, we have studied this matter by using antagonistsf �-AR.
. Materials and methods
.1. Cell culture and treatment
HUVECs were isolated from human umbilical veins accord-ng to the method of Jaffe et al. [20] and used at passage–4. The cells were grown in 75 cm2 culture flasks (Falcon,ecton Dickinson, Lincoln Park, USA), filled with 10 ml of-199 (Sigma, St. Louis, USA) containing 10% fetal calf
erum (Seromed, Berlin, Germany), 2 mM glutamine (Seromed,erlin, Germany), 30 �g ml−1 endothelial cell growth supple-ent (Sigma, St. Louis, USA), 100 �g ml−1 heparin (Sigma,t. Louis, USA), 100 U ml−1 penicillin–streptomycin (Sigma,t. Louis, USA), 100 �g ml−1 streptomycin (Sigma, St. Louis,SA) and 2.5 �g ml−1 anphotericin (Sigma, St. Louis, USA).he flasks were incubated at 37 ◦C, 100% humidity and 5% ofO2. The medium was refreshed every 2 days. At the beginningf each experiment the cells were harvested by trypsinisation,sing 0.05% trypsin (Sigma Co, St. Louis, USA) and 0.537 mMthylenediamine tetraacetic acid (EDTA) in phosphate bufferedaline without calcium and magnesium (Seromed, Berlin, Ger-any). The trypsin was inactivated by dilution, and the cellsere washed and counted. Cells were plated at a concentrationf 40,000 cells cm−2 on a multiwell plate (9.6 cm2 well−1) (Fal-on, Becton Dickinson, Lincoln Park, USA), grown for 2 daysnd then used for the incubations.
HUVECs were harvested and characterized as to acety-ated low-density lipoprotein (LDL) binding and factor VIII
xpression, according to established and previously describedechniques [21]. To assess cell survival, hexosaminidase, a sta-le cytosolic enzyme released by cells when they undergo lysis,as measured according to the method of Landegren [22].i[dp
l Research 55 (2007) 303–309
dl-Nebivolol, d-nebivolol and l-nebivolol along with someetabolites (metabolites A1, A2, A3, A3′, A12 and A14, and R
1,928 from Berlin-Chemie, Berlin, Germany) were dissolvedn ethanol and then diluted at the final concentration in culture
edium M199. Identical dilutions of the solvent were preparednd used as control. Since racemic nebivolol was previouslyhown to reduce in a concentration-dependent manner the intra-ellular increase of ROS and superoxide and to reduce them by0% at 10 �M [15], this concentration was chosen to comparehe racemic nebivolol, its enantiomers and metabolites in thistudy.
.2. LDL isolation and oxidation
Whole blood, obtained by venipuncture from healthy vol-nteers (with informed consent) after 12 h of fasting wasollected into vacutainer tubes (Becton Dickinson, Meylan,rance) containing EDTA (1 mg ml−1), and processed for LDLeparation in 1 day by sequential flotation in NaBr solu-ion containing 1 mg ml−1 EDTA [23]. Cu2+ modified LDL1.7 mg protein ml−1) was prepared by exposure of LDL to 5 �MuSO4 for 18 h at 37 ◦C, and the extent of LDL oxidation wasetermined by thiobarbituric acid-reactive substances, as pre-iously described [24]. Protein was measured by the PierceCA protein assay reagent [25]. Oxidative stress in endothe-
ial cells was induced by the addition of oxidized (ox)-LDL athe concentration of 50 �g protein ml−1 as previously reported15].
.3. ROS and superoxide measurement
Intracellular ROS and superoxide levels were measured byollowing the oxidation of 2′,7′-dichlorofluorescein diacetatend hydroethidine by flow cytometry (Coulter Corpora-ion, Hialeah, FL, USA) as described by Cominacini et al.15]. HUVECs were first preincubated with dl-nebivolol, d-ebivolol, l-nebivolol (experiments performed in triplicate onve separate occasions), metabolites A1, A2, A3, A3′, A12,14, and R 81,928 (experiments performed in quadruplicaten three separate occasions) at the concentration of 10 �Mor 30 min at 37 ◦C. The cells were then exposed to ox-LDL50 �g protein ml−1) for 10 min at 37 ◦C in the presence ofmM arginine (Sigma, St. Louis, MO, USA) and 3 �M tetrahy-robiopterin (TB4; Sigma) to prevent superoxide formationy endothelial nitric oxide synthase (eNOS). Furthermore, toest the contribution of �-AR on ROS and superoxide levelsfter preincubation with dl-nebivolol, the (�1-2-AR antagonistadolol (10 �M; Sigma) and the �3-AR antagonist SR59230A2 �M; Tocris, Bristol, UK) were used.
.4. NO measurement
Intracellular NO was measured by utilizing the fluorescent
ndicator 4,5 diaminofluorescein diacetate in flow cytometry15,26]. HUVECs were incubated with 10 �M of dl-nebivolol,-nebivolol and l-nebivolol or 100 nM bradykinin (Sigma), as aositive control, for 30 min at 37 ◦C. In other experiments (per-
logical Research 55 (2007) 303–309 305
fealaca((
2
nootobL(5itd��
2
wucetwa1AtSb
2
phg
3
(ow
Fig. 1. Effect of dl-nebivolol, d-nebivolol and l-nebivolol on intracellular pro-duction of reactive oxygen species (ROS) (A) or superoxide (B) induced byoxidized-LDL in HUVECs. Results are expressed as mean fluorescence inten-sity (MFI) and are means ± S.D. of experiments performed in triplicate on fivesc
aR
nNfand dl-nebivolol and l-nebivolol were significantly different(mean values ± S.D. were 81.1 ± 5.8 and 57.0 ± 3.6 respec-tively, p < 0.01). Exposure to ox-LDL (50 �g protein ml−1)
Table 1Effect of dl-nebivolol, d-nebivolol and l-nebivolol on basal reactive oxygenspecies (ROS) and superoxide concentration expressed as mean fluorescenceintensity (MFI), and intracellular Ca2+ expressed in nM in HUVECs
ROS (MFI) Superoxide (MFI) Ca2+ (nM)
Control 0.65 + 0.05 0.36 + 0.04 83 + 7dl-Nebivolol 0.61 + 0.06 0.31 + 0.03 327 + 12*
d-Nebivolol 0.59 + 0.04 0.34 + 0.04 94 + 12
S. Evangelista et al. / Pharmaco
ormed in triplicate on five separate occasions) HUVECs werexposed to ox-LDL (50 �g protein ml−1) for 10 min at 37 ◦Cfter preincubation with 10 �M of dl-nebivolol, d-nebivolol and-nebivolol for 10 min at 37 ◦C in the presence of 5 mM argininend 3 �M TB4. To test the contribution of �-AR to NO con-entration after preincubation with dl-nebivolol, the �1-2-ARntagonist nadolol (10 �M), the �3-AR antagonist SR59230A2 �M), and the unselective �1-2-3-AR antagonist bupranolol10 �M, Schwarz Pharma, Monheim, Germany) were used.
.5. eNOS activity measurement
To test if the counteracting effect of dl-nebivolol, d-ebivolol and l-nebivolol on NO reduction induced byx-LDL in HUVECs was also dependent on the activityf eNOS, the enzyme activity was measured by evaluatinghe metabolism of 3H arginine to 3H citrulline as previ-usly described [27]. The activity of eNOS was measured inasal conditions and after preincubation of HUVECs with ox-DL (50 �g protein ml−1), bradykinin (100 nM), dl-nebivolol
10 �M), d-nebivolol (10 �M) and 1-nebivolol (10 �M) formin at 37 ◦C. Furthermore, in other experiments (performed
n quadruplicate on three separate occasions) to test the con-ribution of �-AR to eNOS activity after preincubation withl-nebivolol, the �1-2-AR antagonist nadolol (10 �M), the3-AR antagonist SR59230A (2 �M), and the unselective1-2-3-AR antagonist bupranolol (10 �M) were used.
.6. Intracellular Ca2+ concentration
The changes in intracellular Ca2+ concentration in HUVECsere assessed using the fluorescent Ca2+ indicator fura2 (Molec-lar Probes) as previously described [28]. The fura2-loadedells were excited alternately at 340 nm and 380 nm and themitted fluorescence was measured at 510 nm using a spec-rofluometer (Shimadzu). The intracellular Ca2+ concentrationas estimated using the equation previously described [29]
fter the exposure of the cells to dl-nebivolol, d-nebivolol and-nebivolol at 10 �M. Furthermore, to test the contribution of �-R to Ca2+ concentration after preincubation with dl-nebivolol,
he �1-2-AR antagonist nadolol (10 �M), the �3-AR antagonistR59230A (2 �M), and the unselective �1-2-3-AR antagonistupranolol (10 �M) were used.
.7. Statistical analysis
Results are expressed as mean ± S.D. Statistical analysis waserformed by analysis of variance followed by Tukey’s test postoc analysis. All data were analyzed using the “SPSS 11” pro-ram for Macintosh.
. Results
dl-Nebivolol, d-nebivolol and l-nebivolol significantlyp < 0.001) reduced the formation of ROS (Fig. 1 A) and super-xide (Fig. 1B) induced by ox-LDL and the racemic and l-formere significantly (p < 0.01) more active than d-nebivolol in this
l
Rd
eparate occasions. *p < 0.001 as compared to vehicle (control) and #p < 0.01 asompared to d-nebivolol.
ctivity. dl-Nebivolol and its enantiomers had no effect on basalOS and superoxide concentration (Table 1).
The exposure of HUVECs to bradykinin, dl-nebivolol, d-ebivolol and l-nebivolol in HUVECs significantly increasedO concentration (Fig. 2A). The effect of the racemic and l-
orm was significantly higher than that of d-nebivolol (p < 0.01)
-Nebivolol 0.62 + 0.04 0.30 + 0.05 625 + 17*
esults and are means ± S.D. of three experiments each one performed in qua-ruplicate.* p < 0.001 as compared to control (vehicle).

306 S. Evangelista et al. / Pharmacologica
Fig. 2. Effect of bradykinin, dl-nebivolol, d-nebivolol and l-nebivolol onbasal intracellular nitric oxide (NO) availability (A); effect of dl-nebivolol,d-nebivolol and l-nebivolol on oxidized-LDL (ox-LDL)-induced decrease inNO (B), expressed as mean fluorescence intensity (MFI) and effect of ox-LDL,bradykinin, dl-nebivolol, d-nebivolol and l-nebivolol on endothelial NO syn-thase (eNOS) (C) in HUVECs. All results are means ± S.D. of experimentsperformed in triplicate on five separate occasions. *p < 0.05 or **p < 0.01 as com-pared to vehicle (control; A and C) or ox-LDL (B) and #p < 0.01 as comparedt
iidolsawd
o(
bat
titn(soT3ne5airbaoa
Rb(
4
o
ntmReeboip
aiwttforms. These might be endowed with similar or even more
o d-nebivolol.
nduced a marked decrease (p < 0.001) in the availability ofntracellular NO that was restored by the prior incubation withl-nebivolol, d-nebivolol and 1-nebivolol (Fig. 2B). The effectf the d-nebivolol on this parameter was again significantlyower (p < 0.01) as compared to l- and dl-nebivolol. Fig. 2Chows that eNOS activity significantly (p < 0.001) increased by
5-min contact with dl-nebivolol and l-nebivolol; this effectas of similar extent to that produced by bradykinin, while-nebivolol and ox-LDL did not affect eNOS activity (Fig. 2C).pha
l Research 55 (2007) 303–309
The exposure to dl-nebivolol or l-nebivolol induced changesf intracellular Ca2+, while the exposure to d-nebivolol did notTable 1).
As shown in Table 2, the metabolites A2, A3′, A12 and A14ut not A1, A3 and R 81,928, all tested at 10 �M, retained thentioxidant activity of their precursor dl-nebivolol in reducinghe intracellular formation of ROS and superoxide.
In order to investigate the �-AR subtype involved inhe dl-nebivolol-induced increase in NO, eNOS activity andntracellular Ca2+, the �1-2-AR antagonist nadolol (10 �M),he �3-AR antagonist SR59230A (2 �M), or their combi-ation, and the unselective �1-2-3-AR antagonist bupranolol10 �M) were used (Fig. 3A–C). All the used antagonistsignificantly (p < 0.001) reduced the effect of dl-nebivololn NO, eNOS activity and intracellular Ca2+ (Fig. 3A–C).he increase in NO induced by dl-nebivolol was reduced by9.1, 63.4, 81.4 and 81.6% with nadolol, SR59230A, bupra-olol and nadolol + SR59230A, respectively (Fig. 3A). Theffect of dl-nebivolol on eNOS activity was reduced by 33.1,8.0, 90.6 and 90.2% with nadolol, SR59230A, bupranololnd nadolol + SR59230A, respectively (Fig. 3B). Similarly thencrease in intracellular Ca2+ induced by dl-nebivolol waseduced by 31.4, 53.1, 73.3 and 71.4% with nadolol, SR59230A,upranolol and nadolol + SR59230A, respectively (Fig. 3C). Inll assays the inhibition obtained with the combined treatmentf nadolol + SR59230A was significantly (p < 0.001) differents compared to their separate treatment (Fig. 3A–C).
The inhibitory action of dl-nebivolol on the generation ofOS and superoxide induced by ox-LDL was not counteractedy �-AR antagonists nadolol, SR59230A and their combinationTable 3).
. Discussion
Present results show that the effect of the racemic nebivololn NO release (and formation) is mainly due to its l-enantiomer.
Our previous study showed that one of the mechanisms ofebivolol antioxidant activity is due to direct reduction of ROShat are produced by a NADPH oxidase system [15]. The race-
ate and the 1-enantiomer are similarly effective in reducingOS and superoxide formation induced by the exposure ofndothelial cells to a potent oxidant (ox-LDL) while the d-nantiomer was less effective. Likewise, metabolites derivedy the different pathways, such as aromatic hydroxylation andxidative n-dealkylation [17] show an antioxidant effect of sim-lar extent as compared to the racemate and its 4-keto derivativeroduced by alicyclic hydroxylation [15].
Recent data in humans [30] appear to fully confirm thentioxidant potential of nebivolol, sharply distinguished fromts �-blocking activity, as shown by the direct comparisonith atenolol. This clear-cut activity, at the low concentra-
ions reached in plasma at therapeutic doses, may be related tohe fact that nebivolol is extensively metabolized to oxidated
otent antioxidant properties than nebivolol itself [5], since aydroxylated benzopyrane moiety is common to many naturalntioxidants (e.g., tocopherols).

S. Evangelista et al. / Pharmacological Research 55 (2007) 303–309 307
Table 2Effect of dl-nebivolol and its metabolites tested at 10 �M on the intracellular production of reactive oxygen species (ROS) and superoxide in HUVECs exposed toox-LDL
Chemical formula Metabolic pathway ROS (MFI) Superoxide (MFI)
ox-LDL – 6.25 ± 0.7 3.98 ± 0.4dl-Nebivolol (RSSS + SRRR)-[iminobis (methylene) bis
[6-fluoro-3,4-2H-1-benzopyran-2-methanol] hydrochloride– 3.01 ± 0.3* 1.48 ± 0.1*
Metabolite A1 5′,6′deF,6′OH-nebivolol Aromatic hydroxylation 5.99 ± 0.8 3.45 ± 0.4Metabolite A2 5,6deF,6OH-nebivolol Aromatic hydroxylation 3.66 ± 0.4* 1.87 ± 0.2*
Metabolite A3 7F,6deF,6OH-nebivolol Aromatic hydroxylation 4.76 ± 0.5 3.13 ± 0.3Metabolite A3′ 7′F,6′deF,6′OH-nebivolol Aromatic hydroxylation 3.11 ± 0.3* 1.65 ± 1.7*
Metabolite A12 1-(6F-2H-benzopyran-1-yl)-1,2-dihydroxy-ethane Oxidative n-dealkylation 3.32 ± 0.3* 1.93 ± 0.2*
Metabolite A14 2-(6F-2H-benzopyran-1-yl)-2-hydroxy-acetic acid Oxidative n-dealkylation 2.48 ± 0.3* 1.32 ± 0.1*
M
R . of t
tdsbtii
cpeiblCevwrWwlainw
TEvt
odddd
Rm
sHbtn
nohiiSls3abtitttA
etabolite R 81,928 2-(6F-2H-benzopyran-1-yl)-2-hydroxy-ethanamine
esults are expressed as mean fluorescence intensity (MFI), and are mean ± S.D* p < 0.001 as compared to ox-LDL.
The effect of dl- and l-nebivolol on intracellular NO forma-ion and eNOS activity was significantly greater than that of the-nebivolol. Furthermore, d-nebivolol was not able to induce aignificant eNOS activity that was increased after 5 min contacty bradykinin, dl- or l-nebivolol. As shown previously [15,16]he antioxidant activity of nebivolol and its l-form play a rolen NO availability by reducing ROS and superoxide that cannactivate NO in a chemical reaction producing peroxynitrite.
We present data supporting the evidence that in endothelialells the response to nebivolol is linked to the calcium signal, asointed out in other systems [7,10,31,32]. In particular Parentit al. [31] showed that the NO production induced by nebivololn different vascular systems (rat mesenteric vascular bed andovine coronary endothelial cells) was blunted by the phospho-ipase C (PLC) inhibitor U73122 and the endoplasmic reticuluma2+-ATPase inhibitor thapsigargin, thus indicating that thisffect was dependent on the intracellular calcium release. Con-ersely, Gosgnach et al. [8] showed in HUVECs that nebivololas not able to elicit a PLC dependent calcium increase, but
ather an activation of phospholipase A2 and adenylate cyclase.e presently do not have an explanation for this discrepancyhich might be due to the different experimental conditions uti-
ized. On the other hand, recently Dessy et al. [10] in bovineortic endothelial cells confirmed that nebivolol induces an
ncrease in calcium signal and NO production, and showed thatebivolol dephosphorylates the eNOS at a site (threonine495)ithin the calcium-calmodulin consensus binding sequence,able 3ffect of nadolol, SR59230A and nadolol + SR59230A on dl-nebivolol inducedariation in reactive oxygen species (ROS) and superoxide in HUVECs exposedo ox-LDL
ROS (MFI) Superoxide (MFI)
x-LDL 6.45 ± 0.7 4.24 ± 0.3l-Nebivolol 2.94 ± 0.5* 1.57 ± 0.4*
l-Nebivolol + nadolol 3.01 ± 0.4* 1.51 ± 0.3*
l-Nebivolol + SR59230A 2.98 ± 0.6* 1.49 ± 0.4*
l-Nebivolol + nadolol + SR59230A 2.86 ± 0.5* 1.53 ± 0.3*
esults are expressed as mean fluorescence intensity (MFI), and areeans ± S.D. of three experiments each one performed in quadruplicate.* p < 0.001 as compared to ox-LDL.
fitacGat�
irbbw
at
Oxidative n-dealkylation 4.66 ± 0.5 2.76 ± 0.3
hree experiments each one performed in quadruplicate.
uch as more activating calmodulin binds to eNOS [33]. InUVECs the endothelium-dependent NO liberation inducedy nebivolol has recently been shown to coincide with eNOSranslocation and a phosphorylation at eNOS-serine 1177 witho modification of eNOS phosphorylation at threonine495 [34].
In order to investigate which �-AR subtype mediates theebivolol activated mechanisms we have used a panel of antag-nists endowed with different selectivity for the �-ARs. Weave found that nadolol (a mixed �1-2-AR antagonist) inhib-ted the nebivolol activity on NO release, eNOS activity andntracellular calcium release and the selective �3-AR antagonistR59230A significantly reduced it at a greater extent. Neverthe-
ess nebivolol in the presence of a selective �3-AR antagonistuch as SR59230A [35] still induced a NO release of about0–40% indicating the competitive nature of the antagonismnd the multiple target activation induced by the drug. Bothupranolol (a mixed �1-2-3-AR antagonist) and the combina-ion of nadolol with SR59230A were markedly more effectiven reducing the nebivolol responses, thus indicating an addi-ive contribution by the (�2- and �3-ARs. It should pointed outhat nebivolol, like its d-enantiomer, at the concentration usedhroughout this study (10 �M) is able to bind both �2 and �3Rs [2, Meini, unpublished data].The involvement of �3-ARS in the effect of nebivolol was
rstly shown in HUVECs [8]. In this study the authors foundhat the relatively selective �3-antagonist s-(-) cyanopindololnd bupranolol, but not nadolol, inhibited nebivolol effects onyclic AMP production and nitrite formation. Furthermore, deroot et al. [9] showed that nebivolol-induced relaxation of rat
orta was inhibited by s-(-) cyanopindolol, and mimicked byhe �3-AR agonist BRL 37344 while the relatively effective2-antagonist butoxamine was inactive.
Recent data reinforce this concept showing that NO releasenduced by nebivolol in BAECs as well as its vasorelaxation ofat and human coronary microarteries were inhibited again byupranolol, but not nadolol [10]. These findings were confirmedy the observation that in mice deficient to the �3-ARs, nebivolol
as unable to vasodilate their coronary microvessels [10].The inhibitory effect of nebivolol on the generation of ROSnd superoxide induced by ox-LDL was not counteracted byhe �-AR antagonists suggesting that the antioxidative effect
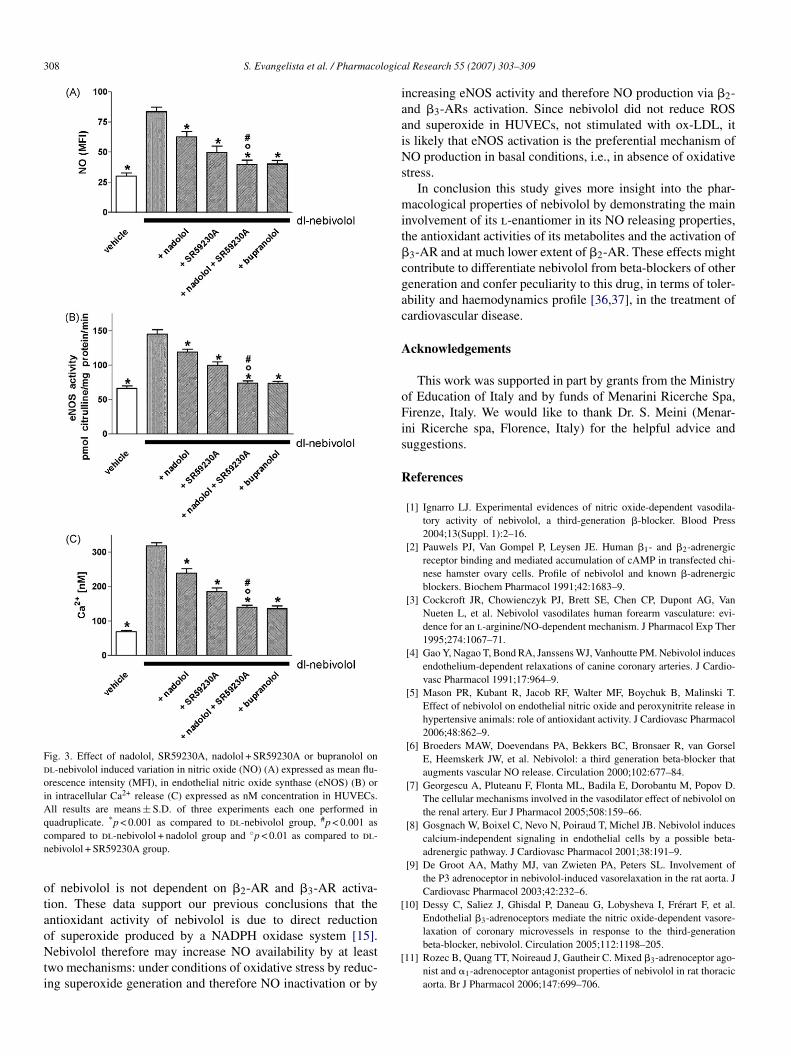
308 S. Evangelista et al. / Pharmacologica
Fig. 3. Effect of nadolol, SR59230A, nadolol + SR59230A or bupranolol ondl-nebivolol induced variation in nitric oxide (NO) (A) expressed as mean flu-orescence intensity (MFI), in endothelial nitric oxide synthase (eNOS) (B) orin intracellular Ca2+ release (C) expressed as nM concentration in HUVECs.All results are means ± S.D. of three experiments each one performed inq * #
cn
otaoNti
iaaiNs
mit�cgac
A
oFis
R
[
laxation of coronary microvessels in response to the third-generation
uadruplicate. p < 0.001 as compared to dl-nebivolol group, p < 0.001 asompared to dl-nebivolol + nadolol group and ◦p < 0.01 as compared to dl-ebivolol + SR59230A group.
f nebivolol is not dependent on �2-AR and �3-AR activa-ion. These data support our previous conclusions that thentioxidant activity of nebivolol is due to direct reduction
f superoxide produced by a NADPH oxidase system [15].ebivolol therefore may increase NO availability by at leastwo mechanisms: under conditions of oxidative stress by reduc-ng superoxide generation and therefore NO inactivation or by
[
l Research 55 (2007) 303–309
ncreasing eNOS activity and therefore NO production via �2-nd �3-ARs activation. Since nebivolol did not reduce ROSnd superoxide in HUVECs, not stimulated with ox-LDL, its likely that eNOS activation is the preferential mechanism ofO production in basal conditions, i.e., in absence of oxidative
tress.In conclusion this study gives more insight into the phar-
acological properties of nebivolol by demonstrating the mainnvolvement of its l-enantiomer in its NO releasing properties,he antioxidant activities of its metabolites and the activation of3-AR and at much lower extent of �2-AR. These effects mightontribute to differentiate nebivolol from beta-blockers of othereneration and confer peculiarity to this drug, in terms of toler-bility and haemodynamics profile [36,37], in the treatment ofardiovascular disease.
cknowledgements
This work was supported in part by grants from the Ministryf Education of Italy and by funds of Menarini Ricerche Spa,irenze, Italy. We would like to thank Dr. S. Meini (Menar-
ni Ricerche spa, Florence, Italy) for the helpful advice anduggestions.
eferences
[1] Ignarro LJ. Experimental evidences of nitric oxide-dependent vasodila-tory activity of nebivolol, a third-generation �-blocker. Blood Press2004;13(Suppl. 1):2–16.
[2] Pauwels PJ, Van Gompel P, Leysen JE. Human �1- and �2-adrenergicreceptor binding and mediated accumulation of cAMP in transfected chi-nese hamster ovary cells. Profile of nebivolol and known �-adrenergicblockers. Biochem Pharmacol 1991;42:1683–9.
[3] Cockcroft JR, Chowienczyk PJ, Brett SE, Chen CP, Dupont AG, VanNueten L, et al. Nebivolol vasodilates human forearm vasculature: evi-dence for an l-arginine/NO-dependent mechanism. J Pharmacol Exp Ther1995;274:1067–71.
[4] Gao Y, Nagao T, Bond RA, Janssens WJ, Vanhoutte PM. Nebivolol inducesendothelium-dependent relaxations of canine coronary arteries. J Cardio-vasc Pharmacol 1991;17:964–9.
[5] Mason PR, Kubant R, Jacob RF, Walter MF, Boychuk B, Malinski T.Effect of nebivolol on endothelial nitric oxide and peroxynitrite release inhypertensive animals: role of antioxidant activity. J Cardiovasc Pharmacol2006;48:862–9.
[6] Broeders MAW, Doevendans PA, Bekkers BC, Bronsaer R, van GorselE, Heemskerk JW, et al. Nebivolol: a third generation beta-blocker thataugments vascular NO release. Circulation 2000;102:677–84.
[7] Georgescu A, Pluteanu F, Flonta ML, Badila E, Dorobantu M, Popov D.The cellular mechanisms involved in the vasodilator effect of nebivolol onthe renal artery. Eur J Pharmacol 2005;508:159–66.
[8] Gosgnach W, Boixel C, Nevo N, Poiraud T, Michel JB. Nebivolol inducescalcium-independent signaling in endothelial cells by a possible beta-adrenergic pathway. J Cardiovasc Pharmacol 2001;38:191–9.
[9] De Groot AA, Mathy MJ, van Zwieten PA, Peters SL. Involvement ofthe P3 adrenoceptor in nebivolol-induced vasorelaxation in the rat aorta. JCardiovasc Pharmacol 2003;42:232–6.
10] Dessy C, Saliez J, Ghisdal P, Daneau G, Lobysheva I, Frerart F, et al.Endothelial �3-adrenoceptors mediate the nitric oxide-dependent vasore-
beta-blocker, nebivolol. Circulation 2005;112:1198–205.11] Rozec B, Quang TT, Noireaud J, Gautheir C. Mixed �3-adrenoceptor ago-
nist and �1-adrenoceptor antagonist properties of nebivolol in rat thoracicaorta. Br J Pharmacol 2006;147:699–706.

logica
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
S. Evangelista et al. / Pharmaco
12] Garban HJ, Buga GM, Ignarro LJ. Estrogen receptor-mediated vascularresponsiveness to nebivolol: a novel endothelium-related mechanism oftherapeutic vasorelaxation. J Cardiovasc Pharmacol 2004;43:638–44.
13] Kakoki M, Hirata Y, Hayakawa H, Nishimatsu H, Suzuki Y, Nagata D, etal. Effects of vasodilatatory �-adrenoceptor antagonists on endothelium-derived nitric oxide release in rat kidney. Hypertension 1999;33:467–71.
14] Kalinowski L, Dobrucki LW, Szczepanska-Konkel M, Jankowski M, Mar-tyniec L, Angielski S, et al. Third-generation beta-blockers stimulate nitricoxide release from endothelial cells through ATP efflux: a novel mechanismfor antihypertensive action. Circulation 2003;107:2747–52.
15] Cominacini L, Fratta Pasini A, Garbin U, Nava C, Davoli A, CriscuoliM, et al. Nebivolol and its 4-keto derivative increase nitric oxide inendothelial cells by reducing its oxidative inactivation. J Am Coll Cardiol2003;42:1838–44.
16] Mason R, Kalinowski L, Jacob RF, Jacoby AM, Malinski T. Nebivololreduces nitroxidative stress and restores nitric oxide bioavailability inendothelium of black Americans. Circulation 2005;112:3795–801.
17] Hendrickx J, Bockx M, Zwijsen C, Borgmans C, Mannens G, MeuldermansW, et al. Location of the hydroxyl functions in hydroxylated metabolites ofnebivolol in different animal species and human subjects as determined byon-line high-performance liquid chromatography-diode-array detection. JChromatogr A 1996;729:341–54.
18] Mangrella M, Rossi F, Fici F, Rossi F. Pharmacology of nebivolol. PharmRes 1998;38:419–31.
19] Rosenkranz-Weiss P, Sessa WC, Milstien S, Kaufman S, Watson CA, PoberJS. Regulation of nitric oxide synthesis by proinflammatory cytokines inhuman umbilical vein endothelial cells. Elevations in tetrahydrobiopterinlevels enhance endothelial nitric oxide synthase specific activity. J ClinInvest 1994;93:2236–43.
20] Jaffe EA, Nachman RL, Becker CG, Minick CR. Culture of human endothe-lial cells derived from umbilical veins. J Clin Invest 1973;52:2745–52.
21] Smith CW, Marlin SD, Rothlein R, Toman C, Anderson DC. Cooperativeinteractions of LFA-1 and Mac-1 with intercellular adhesion molecule-1 infacilitating adherence and transendothelial migration of human neutrophilisin vitro. J Clin Invest 1989;83:2008–17.
22] Landegren U. Measurement of cell numbers by means of the endogenousenzyme hexosaminidase: application to detection of lymphokines and cellsurface antigens. J Immunol Methods 1984;67:379–85.
23] Havel RJ, Eder MA, Bragdon JM. The distribution and chemical compo-sition of ultracentrifugally separated lipoproteins in human serum. J ClinInv 1955;34:1345–53.
24] Cominacini L, Fratta Pasini A, Garbin U, Davoli A, Tosetti ML, Cam-pagnola M, et al. Oxidized low density lipoproteins (ox-LDL) binding toox-LDL receptor-1 in endothelial cells induces the activation of NF-kBthrough an increased production of intracellular reactive oxygen species. JBiol Chem 2000;275:12633–8.
[
l Research 55 (2007) 303–309 309
25] Smith P, Krohn RJ, Hermanson GT, Mallia AK, Gartner FH, ProvenzanoMD, et al. Measurement of protein using bicinchoninic acid. Anal Biochem1985;105:293–9.
26] Cominacini L, Rigoni A, Fratta Pasini A, Garbin U, Davoli A, CampagnolaM, et al. The binding of oxidized low density lipoprotein (ox-LDL) to ox-LDL receptor-1 reduces the intracellular concentration of nitric oxide inendothelial cells through an increased production of superoxide. J BiolChem 2001;276:13750–5.
27] Bredt DS, Snyder SH. Isolation of nitric oxide synthetase, a calmodulin-requiring enzyme. Proc Natl Acad Sci USA 1990;87:682–5.
28] Choi J, Hammer LW, Hester R. Calcium-dependent synthesis ofprostacyclin in ATP-stimulated venous endothelial cells. Hypertension2002;39:581–5.
29] Grynkiewicz G, Poenie M, Tsien RY. A new generation of Ca2+ indi-cators with greatly improved fluorescence properties. J Biol Chem1985;260:3440–50.
30] Fratta Pasini A, Garbin U, Nava MC, Stranieri C, Davoli A, Sawamura T,et al. Nebivolol decreases oxidative stress in essential hypertensive patientsand increases nitric oxide by reducing its oxidative inactivation. J Hypert2005;23:589–96.
31] Parenti A, Filippi S, Amerini S, Granger HJ, Fazzini A, Ledda F. Inositolphosphate metabolism and nitric-oxide synthase activity in endothelial cellsare involved in the vasorelaxant activity of nebivolol. J Pharmacol Exp Ther2000;292:698–703.
32] Gryglewski RJ, Uracz W, Marcinkiewicz E, Jakubowski A, Malinski T.Role of endothelial nitric oxide in pleiotropic action of cardiovasculardrugs: nebivolol. In: Gryglewski RJ, Minuz P, editors. Nitric oxide: basicresearch and clinical applications. Amsterdam: IOS Press; 2001. p. 57–69.
33] Fleming I, Fisslthaler B, Dimmeler S, Kemp BE, Busse R. Phosphorylationof Thr495 regulates Ca2+/calmodulin-dependent endothelial nitric oxidesynthase activity. Circ Res 2001;88:e68–75.
34] Ladage D, Brixius K, Hoyer H, Steingen C, Wesseling A, Malan D,et al. Mechanisms underlying nebivolol-induced endothelial nitric oxidesynthase activation in human umbilical vein endothelial cells. Clin ExpPharmacol Physiol 2006;33:720–4.
35] De Ponti F, Gibelli G, Croci T, Arcidiaco M, Crema F, Manara L. Func-tional evidence of atypical �3-adrenoceptors in the human colon usingthe �3-selective adrenoceptors antagonist SR59230A. Br J Pharmacol1996;117:1374–6.
36] Zanchetti A. Clinical pharmacodynamics of nebivolol: new evidence ofnitric oxide-mediated vasodilating activity and peculiar haemodynamic
properties in hypertensive patients. Blood Press 2004;1:17–32.37] Flather MD, Shibata MC, Coats AJ, Van Veldhuisen DJ, Parkhomenko A,Borbola J, et al. Randomized trial to determine the effect of nebivolol onmortality and cardiovascular hospital admission in elderly patients withheart failure (SENIORS). Eur Heart J 2005;26:215–25.