ecohab: a regional comparison of upwelling and coastal...
TRANSCRIPT

ECOHAB: A Regional Comparison of Upwelling and Coastal Land Use Patterns on the Development of HAB Hotspots Along the California Coast
Lead PI:Raphael M. Kudela
Professor, Ocean Sciences
1156 High Street
University of California Santa Cruz
Santa Cruz, CA 95064
831-459-3290; 831-459-4882 (FAX); [email protected]
David Caron and Burton Jones University of Southern California, Los Angeles, CA Meredith Howard Southern California Coastal Water Research Project, Costa Mesa, CA G. Jason Smith Moss Landing Marine Laboratories, Moss Landing, CA Chris Scholin Monterey Bay Aquarium Research Institute, Moss Landing, CA Yi Chao University of California, Los Angeles, Los Angeles, CA Greg Doucette NOAA

A Regional Comparison of Upwelling and Coastal Land Use Patterns on the Development of HAB Hotspots Along the California Coast
1
I. Proposed Research Narrative 1. Statement of the Problem: The coast of California represents approximately 1,000 miles of the Pacific coastline of the U.S. and major fractions of the coastal economy and environmental resources of our nation. Alarmingly, blooms of harmful and toxic algae have increased in frequency and severity along this coast during the past few decades. Harmful blooms of dinoflagellates (e.g. Alexandrium, Lingulodinium, Cochlodinium) have become increasingly common (Jester et al. 2009), particularly in summer and autumn. Spring blooms of toxic Pseudo-nitzschia are now a recurring problem, first reported in central California during the 1990s and on an annual basis in southern California since 2003 (c.f. Trainer et al. 2000; Scholin et al. 2000; Busse et al. 2006; Schnetzer et al. 2007; Anderson et al. 2008). Many explanations have been proposed and investigated vis-à-vis the possible environmental driving forces for the observed patterns of harmful blooms in this region. The possibilities include hydrographic features that affect macro- and micronutrient availability (coastal upwelling, the proximity to canyons, the existence of retention/incubation areas, river and sewage effluent discharge) and biological aspects of the algae (e.g. light and nutrient competition, vertical migration, mixotrophy). Notable similarities and some marked differences exist in the relative importance of these forcing factors between central and southern California that might provide valuable insights into the similarities and differences in HAB initiation, taxonomic composition and persistence. The same HA species occur along the entire coastline yet the appearance of blooms appears to be staggered in time. While central California experiences a strong influence from the California Current and is strongly influenced by intense agricultural use of coastal land, southern California typically experiences milder coastal upwelling due to seaward diversion of the California Current and presence of the Channel Islands. Inputs of nutrients from land runoff in southern California also reflect contributions from and modification by the large urbanized expanses of this region. These influences are certainly tempered by interannual climate shifts such as ENSO and NPGO, among others (e.g. Kahru et al. 2009). The close proximity of these two regions, resulting in shared mesoscale forcing, but also regional differences in potential HAB forcing mechanisms yielding a natural experimental system. The contrasting oceanographic settings and anthropogenic influences form the basis of our comparative study to understand the environmental factors controlling the initiation, bloom formation, and toxicity of HABs, focusing primarily on Pseudo-nitzschia (domoic acid producer) and secondarily on Alexandrium catenella (Paralytic Shellfish Poisoning) and other dinoflagellates. No single study has yet attempted to directly compare these environments, and therefore no comprehensive theory or model exists regarding the development and progression of these harmful events along the California coast. 1.1. Goal: Our primary objective is to develop a better understanding of the ecophysiological conditions leading to bloom and toxin initiation for Pseudo-nitzschia, by simultaneously comparing two “hot spots”, Monterey Bay and San Pedro, California. Better understanding of these factors will lead to improved understanding of how bloom dynamics change in response to shifting environmental conditions, why “hot spots” exist, and will ultimately provide improved monitoring, predictive modeling, and management decisions. Why a Regional, Comparative Proposal? The experimental and comparative methods have been called “the two great methods of science” (Mayr, 1982) although direct comparative approaches are relatively less common than experimental manipulation within a single system. Both the Monterey and Southern California regions are well characterized for harmful algal events; both regions have MERHAB Regional projects ending in 2010, and the two regional observing systems (SCCOOS and CeNCOOS) support HAB monitoring as a priority, while the recently initiated California HABMAP program supports state-wide efforts to facilitate and coordinate HAB research and monitoring, facilitate knowledge transfer to resource managers. Moreover, the region hosts two exceptional research groups in underwater robotics and sensor networks at MBARI and USC. Despite this wealth of historical information and ongoing experiments and monitoring, we have limited understanding of how offshore (defined as not

A Regional Comparison of Upwelling and Coastal Land Use Patterns on the Development of HAB Hotspots Along the California Coast
2
sampled by shore-based monitoring programs) bloom dynamics influence California. By bringing this team together, we combine the wealth of knowledge from each region, together with the developing expertise in autonomous networks and ocean observing. We feel strongly that this approach is the critical next step to bring greater understanding to HAB and toxin initiation, and to link offshore and nearshore processes. 1.2. Hypotheses: We propose that HA events in US coastal waters exhibit broadly similar patterns related to physical forcing. HA events in the Gulf of Maine, West Florida Shelf, and Pacific Northwest all have a strong advection/transport component to major bloom events (c.f. McGillicuddy et al. 2008; Olascoaga et al. 2008; Trainer et al. 2009). In the Gulf of Maine, this is primarily caused by the interplay of cyst beds, environmental conditions conducive to offshore blooms (temperature, light), and subsequent transport and accumulation against the coast (McGillicuddy et al. 2008). In Florida, onshore propagation of Karenia may be linked to a complex interplay between offshore populations, iron, nitrogen fixation, transport, and the dynamics associated with urbanization and runoff in the coastal ocean (Walsh et al. 2006). In the Pacific Northwest, there is strong evidence for a bloom “incubator” in the Juan de Fuca Eddy region, with subsequent onshore transport dependent on physical conditions (MacFadyen et al. 2008; Trainer et al. 2009). In California, persistent bloom regions also exist, seemingly related to transport and retention. Monterey Bay is an open bay with a persistent cyclonic eddy circulation and the presence of a “bloom incubator” region in the NE corner (an upwelling shadow). Santa Barbara Channel also exhibits a persistent eddy structure, and is a high-biomass, high-toxin region (this region is being investigated by an ongoing NSF-ECOHAB program; we have access to these data, but will not focus on that region for this project). The San Pedro shelf region has, since 2003, become one of the biggest wildlife intoxication hotspots in California, and is characteristic of the Southern California Bight, with retentive circulation and long residence times compared to the open coast (e.g. Monterey). Many bloom events in California “appear” suddenly, and the first manifestation of widespread toxicity is frequently the presence of intoxicated marine birds and mammals with no corresponding increase in toxicity or high cell abundances at shore-based stations (e.g. Scholin et al. 2000). These bloom events span hundreds of kilometers of coastline over a period of weeks to months and may initiate offshore, or in the subsurface where monitoring is sparse, allowing them to seemingly “appear.” Hypothesis 1: blooms initiate as subsurface features (subsurface maxima) and eventually manifest as surface blooms.
H1 Corollary: subsurface conditions are unique and substantially different from subsequent surface bloom manifestation, making it difficult or impossible to predict bloom initiation from shore-based or surface-only sampling.
Hypothesis 2: blooms are predominantly the result of advective processes and retention in eddy-like circulation; subsurface maxima are less important. H2: Corollary: surface blooms can be tracked and predicted based primarily on
knowledge of advection and retention, and conducive surface ecophysiological conditions are broadly similar along the coast of California.
Hypothesis 3: there are a unique set of environmental conditions leading from bloom initiation to toxicity that can be identified through a comparative approach, allowing us to contrast potential factors (such as stratification, nutrient load, nutrient type) between regions.
H3 Corollary: There are underlying climatological conditions that make some years/regions more (less) susceptible to these events. For example, there has been an apparent southward shift (within California) in both DA and PSP events, and a northward shift in true red tide events. These patterns are predictable, and can explain why some years are more (less) susceptible to large-scale toxicity events.
The alternate hypotheses are that HAB events in California are truly stochastic and/or the underlying ecophysiological responses are so complex or varied over modest geographical scales that relatively simple conceptual models such as proposed here cannot adequately explain bloom and toxin initiation.

A Regional Comparison of Upwelling and Coastal Land Use Patterns on the Development of HAB Hotspots Along the California Coast
3
1.3. Summary of Work Plan: We propose a 5-year effort comprised of 3 intensive field-sampling years (3 months per year) combined with physical and statistical models informed by a network of sensing/sampling methodologies. Studies will be conducted to directly compare and contrast two well-studied and well-documented coastal ‘hot spots’ of toxic algal blooms along the California coast. Monterey Bay in central California represents an open embayment that is strongly affected by upwelling, and also where river outflow and land runoff are influenced by extensive agricultural use of coastal land. The San Pedro Shelf area (~300 miles south) experiences some degree of coastal upwelling (with significant eddying around the Channel Islands) but is situated near a highly urbanized region of the country (greater Los Angeles) and
thus is influenced by very different chemical composition of runoff, river output and sewage discharges. Both regions are well characterized, and we take advantage of the many previous and ongoing programs in the region to greatly leverage the requested funds. We will conduct a field-based program to document spring algal blooms in these two areas, coupled with extensive oceanographic observations and directed shipboard studies to characterize the physical/chemical setting and the algal ecology. We will focus on spring because we expect to capture both DA and PSP events in Monterey (Figure 1), and to date, toxic events primarily occur in the spring (Schnetzer et al. 2007; Figure 2). The project is unique in that it brings together investigators from three highly complementary
research areas (HAB ecology, underwater robotics, modeling) to
characterize bloom dynamics, and to use the information to ultimately develop and test predictive models of HAB events along the California coast. Robotics and sensor network studies provide state-of-the-art approaches to document the emergence of HAB events and provide the trigger and contextual information for biological studies (i.e. shipboard work) that will characterize bloom composition and toxin occurrence. Together, these multiscale observational datasets will document specific HAB evolution within the context of local ecosystem and oceanographic dynamics, which in turn will guide development and assessment of models incorporating these observational data, and be used to demonstrate the utility of adaptive sampling and real-time model tuning for HAB applications. The modeling effort will focus on a diagnostic and reanalysis mode, serving to provide a consistent framework for interpretation of the field data and assessment of collection needs, with testing of real-time model tuning and adaptive sampling as part of the field programs, laying the groundwork for future MERHAB and/or PCMHAB efforts. Greater detail is provided in the following sections, but a brief summary of our plans (see also Milestone Chart) follows: Year 1: Establish the realtime operation of the Regional Ocean Model System (ROMS) for the two regions, set up data distribution server via OPeNDAP/THREDDS. Integrate existing statistically-driven transport and bloom/toxin prediction models (Anderson et al. 2009; Lane et al. 2009) with ROMS. Purchase Webb Slocum glider for Monterey (one exists for San Pedro). Standardize shore-based protocols. Year 2: Continue modeling effort, use output as part of decision matrix for identifying potential bloom initiation sites. Deploy gliders for 12 weeks (roughly February-May), ESP cell/toxin detection moorings, and utilize boat time to identify
Figure 1. Climatological HA events from Santa Cruz Wharf (5 year composite) showing presence of both groups during spring months.

A Regional Comparison of Upwelling and Coastal Land Use Patterns on the Development of HAB Hotspots Along the California Coast
4
possible bloom features, focusing on Hypothesis 1. Maintain (with collaborators) shore-based HAB monitoring efforts to compare subsurface observations with existing pier-based programs. Examine ecophysiology of subsurface populations (H2) versus subsurface maxima. Year 3: Continue as for Year 2, but explore role of advective offshore-onshore transport versus subsurface maxima. Refine modeling efforts based on years 1-2. Year 4: Based on our understanding of bloom initiation (H1 versus H2), increase effort to understand ecophysiological triggers for bloom and toxin initiation in the dominant physical regimes. Modeling in maintenance and update mode. Year 5: Reduce field effort; synthesize results in comparison with other regions (Santa Barbara Channel, Pacific Northwest), utilize model hindcasts to integrate field observations and evaluate hypotheses using a comparative approach between the two systems. 2. State of Knowledge: California has the longest history of HAB monitoring in the U.S. In the early 1930’s a state-funded shellfish monitoring program was initiated in response to the presence of saxitoxin (STX), the causative agent of paralytic shellfish poisoning (PSP). In 1991, a major mortality event of marine birds off central California signaled the presence of another neurotoxin, domoic acid, produced by several species of the pennate diatom Pseudo-nitzschia (Work et al. 1993). The California Department of Public Health (CDPH) maintains a shellfish monitoring program to cover both PSP and domoic acid (DA). Presently at least four major toxin syndromes are documented in California, including PSP, ASP, Diarrheic Shellfish Poisoning (DSP) and yessotoxin poisoning (YTX; Kudela et al. 2005; Howard et al. 2008; Anderson et al. 2008). Coastal California has also increasingly been experiencing true red tides caused by various dinoflagellates (Jester et al. 2009; Jessup et al. 2009; Ryan et al. 2008, 2009). Our primary goal is to develop a better understanding of the bloom initiation and toxin induction of Pseudo-nitzschia, with a secondary goal of developing similar understanding at a regional scale for Alexandrium and PSP toxins. The sampling and modeling required to meet these goals allow us to characterize extreme bloom (red tide) events. Although of less concern for human resource managers, these events pose significant problems for proposed future desalination operations in the region (Caron et al., in press). 2.1. Amnesic Shellfish Poisoning Pseudo-nitzschia spp. are ubiquitous in California waters, but major toxin events often occur over large spatial and temporal scales (e.g. Southern California Bight to Monterey Bay, Trainer et al., 2000). Prior to 2000, toxigenic blooms were considered both rare and unusual in southern California (Lange et al., 1994); however, in recent years ASP has become increasingly important in the southern regions of the state (e.g. Trainer et al., 2000; Anderson et al., 2006; Busse et al., 2006, Schnetzer et al., 2007). The apparent synchrony of ASP events in California suggests that there must be large-scale forcing responsible for the otherwise coincidental timing of major DA outbreaks. Multiple factors have been shown to trigger the production of DA by Pseudo-nitzschia (cf. reviews by Bates 1998, Bates et al. 1998, Bates 2000; Bates & Trainer, 2007), but the most thoroughly characterized are macronutrient limitation by either phosphate or silicate in cultures (Pan et al. 1996a,b,c). Pseudo-nitzschia has previously been associated with both eutrophication and a reduction in the ratio of N:Si (c.f. review by Bates et al. 1998; Kudela 2008), but in California there is only circumstantial evidence, linked to (e.g.) El Niño runoff (Scholin et al. 2000). Recently, Anderson et al. (2006) reported a correspondence between limiting Si concentrations, the ratios of Si(OH)4:NO3 and Si(OH)4:PO4
3- (consistent with sewage discharge in the SCB) and the concentrations of Pseudo-nitzschia and particulate DA, but concluded that the relationship is complex, with added variability caused by mesoscale circulation (c.f. Kudela, 2008), while Schnetzer et al. (2007) showed an inverse correlation between DA and Si, N, P, and nutrient ratios. Lane et al. (2009) suggested that nutrients from runoff may be important seasonally/episodically, perhaps explaining the conflicting evidence to date for a strong and convincing link to nutrients or nutrient ratios. DA production by Pseudo-nitzschia spp. has also been linked to iron and copper stress, and indirectly to anthropogenic changes through excess availability of Cu associated with runoff (Ladizinsky, 2003) or decreasing Fe associated with modifications to river flow (Johnson et al., 2001). Iron limitation directly modulates Si:N ratios in diatoms, and DA may serve in Fe-acquisition directly (Rue & Bruland 2001, Maldonado et al. 2002), or through a Cu-mediated

A Regional Comparison of Upwelling and Coastal Land Use Patterns on the Development of HAB Hotspots Along the California Coast
5
high affinity transport system (Wells et al., 2005). Recent laboratory and field data demonstrate that Pseudo-nitzschia may also increase toxicity when growing on urea as a nitrogen source largely of anthropogenic origin (Howard et al. 2007; Kudela et al. 2008a; Kudela et al. accepted). Thus, cultural eutrophication may potentially have the unanticipated consequence of selecting for Pseudo-nitzschia spp. and promoting toxin production in this organism. However, others (Thessen et al. 2009) have pointed out that there is substantial strain- and species-level variability in Pseudo-nitzschia, and the nutrient form may be less important than growth status for predicting toxicity. Clearly there is still a lack of consensus on what, exactly, triggers toxicity in this genus. In general, blooms of Pseudo-nitzschia in California occur during anomalously weak (but not absent) upwelling conditions, typically during a transition from excess to limiting macronutrients (Kudela et al 2004). Similar patterns have been reported for the coast of Oregon (P. Strutton, pers. comm.), while coastal Washington ASP events appear to be much more closely tied to mesoscale circulation patterns associated with upwelling/downwelling conditions, and the presence of the Juan de Fuca Eddy, which acts as an “incubator” for Pseudo-nitzschia (McFadyen et al., 2008; Trainer et al. 2009). The apparent latitudinal trend in bloom events in coastal California waters is consistent with large-scale physical forcing (independent of runoff), suggesting that the spatial pattern is indicative of an equally large-scale change in environmental conditions, perhaps related to proposed floristic shifts in the region (Jester et al. 2009). 2.2. Paralytic Shellfish Poisoning The first link between shellfish poisoning and marine phytoplankton was discovered in the late 1920s after a PSP event caused over 100 human poisonings along the coast from San Francisco to Monterey Bay (Sommer & Meyer 1937. Human poisonings have been chronicled and a PSP database maintained by CDPH indicates that PSP toxin events occur most years (Price et al., 1991, Langlois pers. comm.). Large-scale outbreaks of PSP toxicity appear to exhibit a northward temporal trend, similar to the reported decadal expansion of major ASP events along the US west coast, with PSP events often initiating near San Francisco and extending as far north as the California-Oregon border (Langlois et al., 2001). Although primarily a northern California phenomenon, PSP outbreaks were first reported in 1918 from San Diego County (Price et al. 1991), and STX is routinely detected throughout California. Over shorter time periods (last 5+ years), PSP has been more problematic in Southern California, again similar to ASP. A. catenella is a strong swimmer, in part due to the formation of long chains of cells (Fraga et al., 1989), and PSP outbreaks typically initiate on the open coast, and only then move into bays and estuaries (Langlois, 2002). A consistent pattern of PSP events during relaxation of upwelling is apparent with a rapid increase of A. catenella, followed by onshore transport during relaxation-favorable winds, with subsequent toxicity of shellfish (Price et al., 1991; Langlois and Smith, 2001). Thus PSP events also appear to be correlated to large-scale oceanographic features, in particular the upwelling-relaxation cycle and the onshore transport of toxic cells (c.f. Anderson et al. 2005; Kudela et al. 2005). 2.3. Red Tides Coastal upwelling systems host diverse dinoflagellate species, many of which can form HABs (Horner et al., 1997; Smayda, 1997; Kudela et al., 2005). While largely ignored in California, red tides have become increasingly problematic. Yessotoxin was detected in mussel and seawater samples during the 2005 Lingulodinium polyedrum bloom in southern California (Howard et al., 2008). The fish and shellfish killing Cochlodinium (Curtiss et al. 2008; Kudela et al. 2008c) has also become problematic in California waters, while emerging HAB threats include organisms such as Akashiwo sanguinea, previously thought to be benign, but recently linked to large scale marine wildlife mortality (Jessup et al. 2009). At least some of these events have again been linked to meso- or large-scale forcing (Cloern et al., 2005), although Coclodinium increases have also been linked to significant sewage inputs into the nearsurface ocean (Reifel, 2009). 2.4. Bloom Initiation: The link between biology and physics For all of the HA groups (Pseudo-nitzschia, Alexandrium, other red tides) there is strong evidence for “hot spots” along the California coast where blooms and/or toxin accumulation are recurrent features (Horner et al., 1997; Trainer et al., 2000; Hickey and Banas, 2003; Kudela et al. 2005; Schnetzer et al. 2007). In other parts of the continental US, these hotspots have been associated with bloom transport

A Regional Comparison of Upwelling and Coastal Land Use Patterns on the Development of HAB Hotspots Along the California Coast
6
mechanisms and bloom retentive features (e.g. MacFadyen et al. 2008; Trainer et al. 2009). Similar mechanisms have been suggested for California, but there has been relatively little effort to identify bloom initiation and concentration mechanisms in these hotspots (other than red tides; Ryan et al., 2008, 2009). For at least one region, the Santa Barbara Channel, current efforts to predict HAB events are hampered by the occasional horizontal advection of water masses into the study region (Anderson et al., 2006, 2009), highlighting the critical need to better understand bloom initiation and transport. For this proposal we cannot focus on all possible hotspots in California, but instead focus on two regions with markedly different hydrography and forcing, the Monterey Bay and San Pedro regions. 2.4.1. Monterey Bay During the past 25 years, a number of studies investigating the hydrography of Monterey Bay and its adjacent shelf have been conducted (Strub et al. 1987, Noble et al. 1992, Breaker and Broenkow 1994, Rosenfeld et al. 1994, Paduan and Rosenfeld 1996). Remotely sensed and in situ data from these studies have provided a picture of the surface and internal circulation patterns. The flows in Monterey Bay can be characterized by two regimes: spring-summer and fall-winter. The spring-summer regime, which occurs from mid-March to mid-October, is characterized by strong northwesterly winds produced by an offshore region of high pressure (the California High). These northwesterly winds drive the intense periods of upwelling of cold, nutrient-rich waters outside Monterey Bay, producing a band of cold water often crossing the entrance of Monterey Bay. During this time, the interior of the Bay is characterized by a cyclonic gyre, formed as a dynamic response to active upwelling. This warm, re-circulating gyre is highly stratified and has a calculated residence time of roughly 8-12 days (Graham and Largier 1997). Although winds are predominantly from the northwest during the spring-summer regime, there are periods where winds decrease substantially or reverse direction. These periods are termed ‘relaxation events’. The water column has been observed to warm, mix and de-stratify in response to these relaxation events. The fall-winter regime, which occurs from mid-October to mid-March, is characterized by southwesterly winds produced by an offshore region of low pressure (the Aleutian Low). The fall-winter regime is also characterized by intense storms that cause the water column to mix and de-stratify. Plankton biologists have long known that organisms are not distributed homogeneously in the water column (Cassie 1963, Wiebe and Holland 1968). Distributions vary both horizontally and vertically across a continuum of time and space scales. An extensive literature addressing horizontal "patchiness" results from decades of oceanographic research (e.g., Haury et al. 1978, Mackas and Boyd 1979). Within the Monterey Bay, subsurface harmful algal cell maxima are prevalent, producing “thin layers” (McManus et al. 2008), thicker subsurface maxima following the pycnocline, possibly related to bloom subduction (Ryan et al. 2002), and layers of vertically migrating dinoflagellates (Kudela et al. 2008c). Surface concentrations of both red tides (Ryan et al. 2005b) and Pseudo-nitzschia at frontal regions are frequently observed (Rienecker et al. 2008), highlighting the potential importance of both vertical and horizontal aggregation. Ryan et al. (2005a) speculated that the identified subsurface layer of Pseudo-nitzschia may have been influenced by transport of resuspended iron in a front. Iron concentrations are a potential trigger for toxin production (Rue and Bruland 2001). More recently, Loureiro et al. (2009a,b) showed that P. delicatissima utilizes DOM in both the lab and field, consistent with previous assertions that osmotrophy may provide enhanced survival of this genus in the dark, such as in subsurface layers (Mengelt and Prézelin, 2004). Regardless of the mechanisms, we know from limited preliminary sampling that “offshore” (few km) dynamics can be completely different from the picture provided by traditional shore-based sampling. The well-documented 1998 event (Scholin et al. 2000) exhibited little to no toxicity in sentinel mussels analyzed by the California Department of Public Health, while over 400 California sea lions died. More recently, a toxic Pseudo-nitzschia bloom was remotely identified less than 10 km from the Santa Cruz Wharf sampling site (Fig. 6); during this period, no toxigenic Pseudo-nitzschia were identified during weekly sampling (using whole cell probes), and mussels were negative for DA, highlighting the potentially serious disconnect between our understanding of bloom dynamics, which is largely informed by historical near-shore or shore-based sampling, and reality.

A Regional Comparison of Upwelling and Coastal Land Use Patterns on the Development of HAB Hotspots Along the California Coast
7
April 26,
0.01
– 1
1.01
–
2.5
2.51
–
5.0
5.01
–
μg particulate DA
2.4.2. Southern California Although HABs have been documented in the Southern California Bight (SCB) for more than a century (Torrey, 1902), only within the last decade have several toxic events in the region been attributed to outbreaks of DA resulting from blooms of Pseudo-nitzschia species. Extensive field sampling since 2003 has documented recurring blooms of Pseudo-nitzschia seasonally. In particular, blooms during 2006 and 2007 revealed high concentrations of domoic acid, particularly in the region of the San Pedro Shelf and Channel (Fig.2). HAB studies in this region prior to 2007 were largely restricted to sampling surface waters, and only recently has the possibility been documented that these high concentrations of DA might be surface manifestations of blooms taking place at depth and then transported into surface waters. Subsurface maxima in phytoplankton biomass can contain substantial accumulations of toxic Pseudo-nitzschia species (unpublished data). The RAPDALERT (MERHAB) project in this region has effectively used gliders to obtain 2-dimensional (Fig. 3) and 3-dimensional tracking of phytoplankton blooms in the San Pedro Shelf region. Glider surveillance provides near-real time, telemetered data on the presence of subsurface blooms, depth-specific information on their location within the water column, and geographical extent. The latest glider missions have been particularly useful because they have provided an explanation of significant mortality events among sea lions and seabirds prior to any substantive manifestation of domoic acid in the surface
plankton. These results, coupled with the demonstrated importance of subsurface phytoplankton maxima in Monterey Bay (previous section), provide the observational underpinning of our primary hypothesis to be tested in this project. The SCB is anomalous in its response to some of the large scale processes that affect the California coastal ocean. Because the Bight is relatively protected from the wind forcing, chlorophyll biomass in the SCB shows a neutral or slightly negative response to the upwelling index (Thomas, 2009), but the Bight may be consistent with other large scale trends such as increasing chlorophyll (Kahru and Mitchell, 2008) and productivity (Kahru et al., 2009). The SCB is unique because of its overall response to larger scale upwelling, and also because it is a region of some of the most intensive anthropogenic inputs along the west coast. The current population of southern California is more than 21 million and is expected to grow at a rate of about 1% per year exceeding 31 million by 2050 (State of California, 2007). Sixty point sources discharge over 4.7 million cubic meters of treated effluent per day into the coastal ocean (Lyon et al., 2006; Lyon and Stein, 2008, 2009). The major point sources of pollutant discharge into the
Fig. 2. Spatial distribution of particulate DA concentrations in surface waters of the San Pedro Shelf area during 2006 and 2007. Maximal values measured inside the harbor during 2006 exceeded 14 µg DA L-1. Maximal values during 2007 just outside the harbor exceeded the 2006 values (>25 µg DA L-1).
A Regional Comparison of Upwelling and Coastal Land Use Patterns on the Development of HAB Hotspots Along the California Coast
8
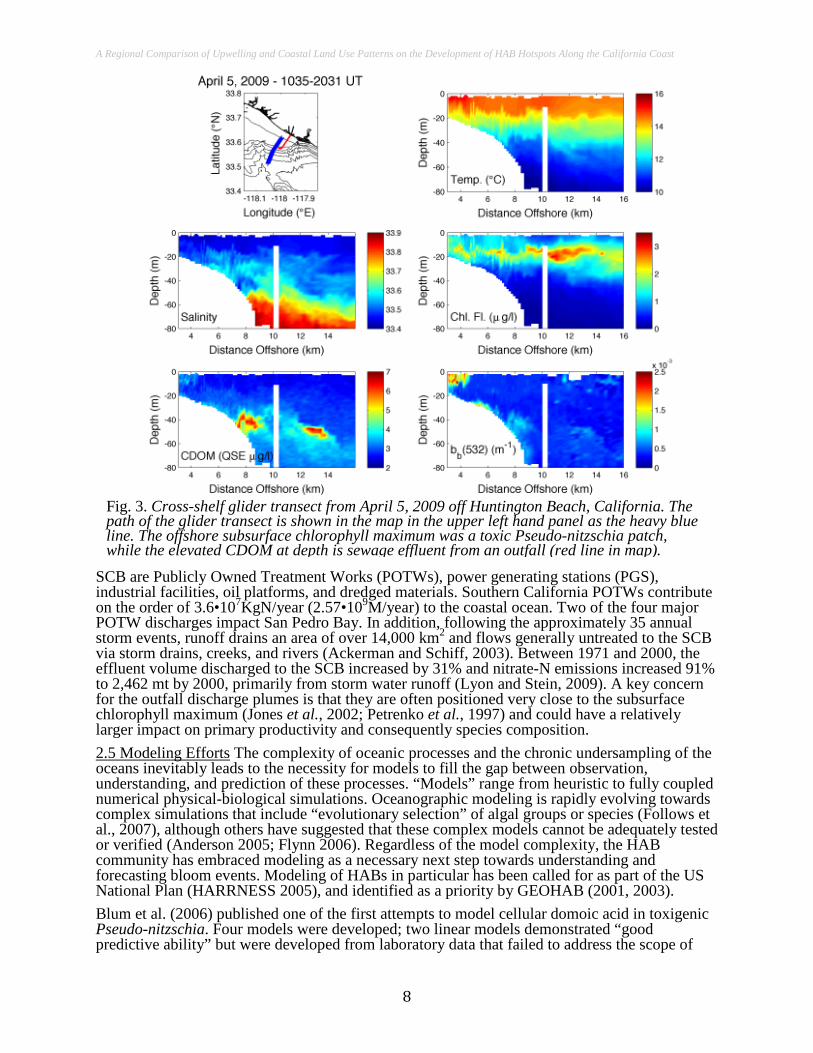
Fig. 3. Cross-shelf glider transect from April 5, 2009 off Huntington Beach, California. The path of the glider transect is shown in the map in the upper left hand panel as the heavy blue line. The offshore subsurface chlorophyll maximum was a toxic Pseudo-nitzschia patch, while the elevated CDOM at depth is sewage effluent from an outfall (red line in map). SCB are Publicly Owned Treatment Works (POTWs), power generating stations (PGS),
industrial facilities, oil platforms, and dredged materials. Southern California POTWs contribute on the order of 3.6•107KgN/year (2.57•109M/year) to the coastal ocean. Two of the four major POTW discharges impact San Pedro Bay. In addition, following the approximately 35 annual storm events, runoff drains an area of over 14,000 km2 and flows generally untreated to the SCB via storm drains, creeks, and rivers (Ackerman and Schiff, 2003). Between 1971 and 2000, the effluent volume discharged to the SCB increased by 31% and nitrate-N emissions increased 91% to 2,462 mt by 2000, primarily from storm water runoff (Lyon and Stein, 2009). A key concern for the outfall discharge plumes is that they are often positioned very close to the subsurface chlorophyll maximum (Jones et al., 2002; Petrenko et al., 1997) and could have a relatively larger impact on primary productivity and consequently species composition. 2.5 Modeling Efforts The complexity of oceanic processes and the chronic undersampling of the oceans inevitably leads to the necessity for models to fill the gap between observation, understanding, and prediction of these processes. “Models” range from heuristic to fully coupled numerical physical-biological simulations. Oceanographic modeling is rapidly evolving towards complex simulations that include “evolutionary selection” of algal groups or species (Follows et al., 2007), although others have suggested that these complex models cannot be adequately tested or verified (Anderson 2005; Flynn 2006). Regardless of the model complexity, the HAB community has embraced modeling as a necessary next step towards understanding and forecasting bloom events. Modeling of HABs in particular has been called for as part of the US National Plan (HARRNESS 2005), and identified as a priority by GEOHAB (2001, 2003). Blum et al. (2006) published one of the first attempts to model cellular domoic acid in toxigenic Pseudo-nitzschia. Four models were developed; two linear models demonstrated “good predictive ability” but were developed from laboratory data that failed to address the scope of

A Regional Comparison of Upwelling and Coastal Land Use Patterns on the Development of HAB Hotspots Along the California Coast
9
nutrient concentrations and ratios encountered in the field. A third linear model and a logistic regression were developed from combined laboratory data and field data. Split-sample validations of these models demonstrated their “adequate reliability”, but the limited amount of field data (N = 46) and the predominance of laboratory data left the applicability of these models as an open question. Anderson et al (2009) present the results of a model development study for Pseudo-nitzschia blooms in the Santa Barbara Channel. Pseudo-nitzschia abundance and DA concentration were quantified, respectively, from water samples collected along a seven-station, cross-channel transect from November 2004-June 2006. A statistical analysis of hydrographic and optical data measured concurrently with phytoplankton and toxins was used to understand what factors would lead to positive predictions of toxigenic Pseudo-nitzschia blooms and DA events. Using a stepwise regression approach, this study identified several variables as good predictors of Pseudo-nitzschia abundance and high toxin concentrations. Ultimately, six models were developed that relate a set of predictor variables to three different response variables: Pseudo-nitzschia cell abundance, particulate DA or cellular DA. Lane et al. (2009) similarly developed predictive logistic models of toxigenic Pseudo-nitzschia blooms in Monterey Bay, California, from a multi-project dataset. Models were developed for year-round (Annual model) or seasonal use (Spring and Fall-Winter models). The consideration of seasonality was significant: chlorophyll a and silicic acid were predictors in all models, but period-specific inclusions of temperature, upwelling index, river discharge, and/or nitrate provided significant model refinement. Predictive power for ‘unknown’ (future) bloom cases was demonstrated at ≥75% for all models, out-performing a chlorophyll a anomaly model.
Figure 4 illustrates the predictor variables that are shared and not shared between the models. While the regions of interest, and in some cases, the dependent variable, differ between these studies, the similarities and differences shared between the models provide insight into universal patterns of Pseudo-nitzschia ecophysiology, and indicate which variables may be fundamental to understanding bloom and toxin initiation. Where upwelling was directly evaluated as a predictor variable, a weak positive relationship was identified between upwelling and Pseudo-nitzschia bloom incidence throughout the year. Silicic acid and nitrate both emerged as significant predictor variables in models developed for Pseudo-nitzschia toxicity (Blum et al. 2006) and in the models developed by Lane et al. (2009). In both studies, the relational patterns agree: association to the dependent variable is negative for silicic acid and positive for nitrate. While neither of these variables emerged as individual predictors in the
models developed by Anderson et al. (2009), a negative relationship was demonstrated between the ratio of silicic acid to nitrate and blooms of Pseudo-nitzschia, indicating a possibly confounded negative and positive relationship between blooms and silicic acid and nitrate, respectively. Anderson et al. (2009) chose not to evaluate river discharges as model variables, but presented a Pseudo-nitzschia bloom and cellular toxicity model that included particle absorption and CDOM absorption, variables associated with river discharge events (Warrick et al. 2004, 2007). In both models, high particulate absorption was negatively associated with the dependent variable, suggesting a direct negative relationship between high river discharge and Pseudo-nitzschia blooms. The Fall-Winter model from Lane et al. (2009) which addresses the time period in which ‘first flush’ and high discharge events generally occur, also demonstrates a
Figure 4. Venn diagrams illustrating the shared and unique variables included in models of Pseudo-nitzschia ecophysiology presented within Lane et al. (2009), Anderson et al. (2009), and Blum et al. (2006)

A Regional Comparison of Upwelling and Coastal Land Use Patterns on the Development of HAB Hotspots Along the California Coast
10
direct negative relationship between river discharge and bloom incidence. By applying a comparative approach between Monterey and Southern California, the relative importance of these variables will be directly assessed. For example, we expect strikingly different patterns of upwelling, nutrient load, nutrient ratios, and riverine discharge between systems, facilitating identification of (dis)similar patterns between the two regions. 3. Approach/Activities 3.1. Overall Strategy Our overall approach will be the study of HAB related processes, focusing on Pseudo-nitzschia using simultaneous data collection from two regions to facilitate a direct comparison. We rely on a sophisticated tiered sampling methodology: Slocum gliders, existing shore-based monitoring efforts (see related studies section), deployment of 2 ESP sensors for cell/toxin detection (1 in each region), satellite imagery, and data assimilation into ROMS will provide a rich background data set for the identification of potential cell/toxin initiation events. Statistical models will be used as another data input, with the recognition that these models were developed primarily for identification of surface expressions (large blooms) and are biased towards the regions and datasets used to parameterize them. When a feature of interest is identified, or if a bloom is identified by another means (e.g. sudden increase in reported marine bird or mammal strandings; increase in toxicity or cell density from the CDPH network), we will adaptively sample the region if it is within our working domains, and re-task assets (such as the gliders) to further interrogate the potential event. Ship-based sampling to verify the species composition and presence of toxin will commence. If the “event” is not of interest (not a HAB event), we will use the data to refine the statistical models and our heuristic view of the ecophysiological conditions necessary to initiate an event. When a potential bloom is detected, we will more aggressively sample the feature (in space and time), including ship-based field experiments to assess species assemblage, growth conditions, response to fundamental parameters such as temperature, light, nutrients, and adjust accordingly as the bloom evolves. 3.2. Modeling (statistical and ROMS) For the California coast, the high-resolution ocean model is based on the Regional Ocean Modeling System (ROMS) framework, a community-based model designed for regional applications (Shchepetkin and McWilliams, 2005). The model uses a vertical coordinate following bottom topography (Song and Haidvogel, 1994). Chao has developed a number of ROMS configurations in the Pacific and Atlantic oceans. One of the earliest ROMS configurations is centered around the Monterey Bay and has been used to support a well-described 2003 field experiment (Chao et al., 2008; Chao et al., 2009; Doyle et al., 2009; Wang et al., 2009) as well as additional experiments in 2006 and 2008. Another ROMS configuration over the southern California coastal ocean with a 1-km spatial resolution has been running in real-time since April 2007 (http://ourocean.jpl.nasa.gov/SCB). For both configurations, we have developed a 3-dimensional variational data assimilation (3DVAR) method into ROMS (Li et al., 2008, 2009). Our ROMS 3DVAR is designed to assimilate all available observations (including satellite) simultaneously and reliably. A key component that is currently missing from the SBC and Monterey Bay ROMS is the coupling with a biogeochemical/ecosystem model capable of generating nutrient fields for use with empirical (statistical) models to predict HAB events (Anderson et al., 2009; Lane et al., 2009). We are not including a request for this capability, because we are in negotiations with the California Ocean Protection Council to support this effort for development of near-real time prediction of nearshore HAB events. While complementary, these two efforts have fundamentally different goals. We wish to understand the onshore-offshore coupling and bloom initiation of toxic events, which will ultimately inform the statistical modeling efforts supported by OPC. The OPC effort is focused on direct management application and real-time prediction (i.e. there is little interest in the underlying ecology of blooms, except to the extent that it improves the management tools, and relatively little interest in blooms not impacting the shoreline). While we are not directly interested in the statistical models for this effort, we will utilize output from and iteratively refine them and to identify potential bloom initiation in concert with the adaptive sampling network. These models were developed from surface blooms; if subsurface

A Regional Comparison of Upwelling and Coastal Land Use Patterns on the Development of HAB Hotspots Along the California Coast
11
bloom initiation is important and ecophysiological conditions are fundamentally different, we would expect the statistical models to fare poorly. If bloom initiation is directly related to mesoscale conditions (upwelling/downwelling, nutrient supply, temperature), then the statistical models may be robust. To date, the statistical models have been developed for specific regions. An important component of our effort will be to apply these models in a comparative approach, to see if the same parameters capture bloom dynamics across the California coastline. 3.3. Shore-based sampling Our approach heavily leverages existing field monitoring programs. The CDPH maintains a statewide HAB monitoring network consisting of plankton and shellfish sampling, in combination with targeted sample collection from commercial harvesters. Cal-PReEMPT augmented this sampling network at three locations, Marin County, Santa Cruz Wharf, and the Cal Poly Pier and later added commercial shellfish growing areas in Santa Barbara and San Luis Obispo (Morro Bay) where sampling has occurred at least weekly, including molecular probe methods for HABs, testing of field toxin kits (Jellett and Mercury Science), and comparison of “volunteer” methods versus rigorous lab-based phytoplankton and toxin identification. RAPDALERT has similarly augmented the traditional shore-based (CDPH) network in Southern California with advanced sampling methods, including remote vehicles, satellite, rapid response boat surveys, and enhanced time-series using (e.g.) cELISA for rapid toxin detection, particularly from wildlife samples. These MERHAB programs are nearing the end of funding, and key, albeit modest, components have been picked up by the Regional Coastal Ocean Observing Systems, CeNCOOS and SCCOOS. Through HABMAP, these efforts have become better organized and more consistent in terms of data collected, methods used, and data access/dissemination. CeNCOOS has developed a pier-based observation system, with HAB monitoring at Avila Beach, Monterey Wharf II, and Santa Cruz Wharf, while other programs (see Associated Work) also provide relevant data. The SCCOOS HAB program involves sampling at 6 piers from San Luis Obispo to San Diego. We will utilize the shore network as a “test” for our ability to predict large-scale bloom initiation. If a major toxin event occurs that we do not detect in our study regions, this will inform our decisions to focus on subsurface versus advective/retentive processes (H1 vs. H2). 3.4. Glider Operations A central component of this proposal (and budget) involves the simultaneous, comparative deployment of Slocum electric gliders. In 2009, the RAPD-ALERT program demonstrated the utility of this approach, using glider observations to identify a subsurface chlorophyll maximum and targeting boat sampling to detect a toxic Pseudo-nitzchia event (Figure 3). For our project, gliders are the most cost-effective method for identifying potential blooms (a fraction of the cost of the MBARI AUV, for example). A recent analysis, using satellite fluorescence line height (FLH) as a proxy for biology, demonstrated that a single Slocum electric glider provided as much information as six optimally placed moorings for a region such as Monterey Bay (Figure 5), whereas our field site at Santa Cruz Wharf captured only about 20% of the variance in the surface FLH for the same region; obviously the glider also provides the critical depth dimension.
Figure 5. Comparison of statistically reconstructed fluorescence line height (FLH) climatology using 6 optimally placed moorings (left), the Santa Cruz Wharf sampling site (middle), or a Slocum electric glider covering the indicated black line over 6 days (right). Colors indicate local R^2 compared to the original FLH data. Adapted from Frolov et al. 2009.

A Regional Comparison of Upwelling and Coastal Land Use Patterns on the Development of HAB Hotspots Along the California Coast
12
Figure 6. (a) ESP network experiment design in Monterey Bay, California. (b) Vertical sections of temperature and chlorophyll concentration (2 of 10 measured parameters) within the ESP network on October 15 and 21, 2008. (c) Time-series of probe array images relative to nitrate concentrations (1 of 5 measured parameters) at ESP1. Quantifiable variation in Pseudo-nitzschia species abundance is evident in probe array shading; probe-quantified DA concentrations are noted in red (Doucette et al., 2009).
We propose to deploy the gliders for 12 weeks (each region) in Years 2-4. We will seek additional funding to maintain these operations through Year 5, and to use the gliders for HAB monitoring during other time periods (e.g. autumn). Each glider will be identically equipped with CTD and two WET Labs ECO Triplet sensors providing chlorophyll, CDOM, phycoerythrin, and 3 backscatter channels (for identifying non-biological particles and potentially providing information about the size spectrum, index of refraction, and relative bio-available iron concentrations (Kudela et al. 2006, 2008a). We will rely on the experience gained by the RAPDALERT program for optimal use of gliders in HAB event detection. USC will continue to
maintain the southern glider made available through MERHAB. UCSC will purchase a second glider, to be maintained and operated by MBARI/CeNCOOS for this project (see letter). 3.5. ESP Deployments The application of molecular analytical techniques is pervasive in the ocean sciences, yet the vast majority of work typically occurs in shore-based laboratories after return of discrete sets of samples collected at sea. Scholin and co-workers (2008, 2009) have worked to overcome that limitation through development and deployment of the Environmental Sample Processor (ESP). The ESP allows for application of molecular probe technology in situ to enable remote detection of water-borne microorganisms, their genes and gene products. The ESP allows real-time detection chemistries and can be used to archive samples for laboratory analyses after the instrument is recovered. DNA probe arrays reveal target species based on ribosomal RNA (rRNA) sequences using a sandwich hybridization assay (SHA) format for the simultaneous detection of various organisms in a single sample, including a variety of harmful and toxic algae (e.g., Scholin et al. 2003, Ayers et al. 2005, Haywood et al. 2007, 2009, Mikulski et al. 2008) invertebrates (Goffredi et al. 2006, Jones et al. 2007) and marine bacteria (Paul et al. 2007, Preston et al. 2009). The protein arrays utilize a competitive ELISA (cELISA) technique for detecting target substances such as the algal toxin domoic acid (DA; Doucette et al. 2009). Detection of DA onboard the ESP is currently accomplished using a membrane-based array similar to that used to detect rRNA (Doucette et al. 2009). The assay is highly sensitive, with a detection limit to low
ng/mL DA in seawater for a 0.5 L sample volume acquired by the ESP, far below what would be considered a public health concern. DA cELISA results generated during month-long deployments have corresponded closely to lab values for water samples collected concurrently at the ESP mooring.

A Regional Comparison of Upwelling and Coastal Land Use Patterns on the Development of HAB Hotspots Along the California Coast
13
Multi-node ESP networks have been fielded within a larger coastal ocean observing system comprised of satellite remote sensing, regional long-term moorings and autonomous underwater vehicles (AUVs). During network deployments in fall of 2007 and 2008, ESP sensors detected significant variability in populations of HAB species and toxins, and AUV surveys effectively captured bay-wide environmental and biological changes linked to this variability This is a powerful approach for linking environmental forcing to HAB response. For example, during the October 2008 study, the maximum ESP signals for Pseudo-nitzschia spp and DA measured on October 15 coincided with elevated nitrate concentrations. Variability was clearly linked to the larger upwelling system dynamics, as a pulse of aging upwelled waters, relatively cold and chlorophyll-rich, dominated the study region. In contrast, conditions observed on October 21 showed that waters throughout the study region were warmer and contained lower chlorophyll concentrations, and Pseudo-nitzschia spp declined along with DA (Figure 6). We propose to establish an ESP network spanning hundreds of km of the California coast, simultaneously monitoring Pseudo-nitzschia and DA variability in Monterey Bay and on the San Pedro shelf. Both of these regions are known to be hotspots for the development of toxic Pseudo-nitzschia blooms (Scholin et al., 2000; Schnetzer et al. 2007; Sekula-Wood et al., 2009), and both may form dense subsurface layers (McManus et al., 2007, Caron et al., 2009). The coastal waters of these two regions experience significantly different natural and anthropogenic forcing, providing ideal contrast for the comparative ecosystems approach espoused by GEOHAB. 3.6. Decision Support System Conventional ways of sampling and observing the ocean have relied on ship-based observations that often require on station presence and provide limited spatio-temporal coverage. Fixed observing infrastructure based on moorings cannot capture the dynamic scales of processes that extend over large horizontal scales, vary in depth, or evolve in space with typical advection rates of ~1 m/s. Satellite observations, although capable of capturing the spatial evolution of some biological events, are limited to the sea surface when there it is visible. Autonomous Underwater Vehicles (AUVs) in this context provide a valuable platform for coverage and cost-effective operation. One principal problem continues to be where to position them to be able to resolve key ecological questions, and how to derive optimal path positioning so as to be sampling “at the right place and time” based on science needs.
We have designed and will construct an approach that captures data from a diverse set of sources to target AUV deployment. Our efforts include several complementary approaches: (1) given an ocean circulation model and using points on a convex hull as proxies for hotspots for biological activity, generate an optimal path traversal (Smith et al. 2009); (2) use a range of data sources including satellite observations, mooring data and HF radar as well as the glider data, to project patch advection using a statistical approach (Das et al. submitted); (3) build decision making capability using Artificial
Intelligence techniques onboard AUVs to adaptively sample dynamic coastal phenomenon (Rajan et al. 2009). Systematic application of these three capabilities will provide a shore-side Decision Support System (DSS) tool (see schematic) to adaptively target the placement of gliders within bloom patches. Such a DSS will enable scientists on shore to seek advice and generate “what if” scenarios in deciding potential changes to glider track lines, projecting sudden changes to environmental conditions impacting a slow moving glider or generating alternatives

A Regional Comparison of Upwelling and Coastal Land Use Patterns on the Development of HAB Hotspots Along the California Coast
14
to direct researchers to appropriate sites for experimental studies. The goal is to provide a tool that can identify promising areas for sampling based on the integrated data sources and generates advice. In doing so we leverage our experience of providing similar functionalities as a DSS for scientists on NASA’s Mars Exploration Rovers mission (Bresina et al. 2005). We will begin this task by integrating additional sources of data in our statistical advection model to build an initial DSS prototype for validation. Using those model outputs we will incorporate the work of Smith et al. (2009) to generate optimal glider traverses. We will integrate automated planning capability within the DSS to adaptively suggest targets for the glider to reach given available information. Based on these efforts, we will endeavor to close the control loop, from gliders generating data for the DSS to have the DSS generate a set of viable locations for the glider to traverse. During this work we will analyze the feasibility of obtaining outputs from ocean models like ROMS and ecological transport models for synthesis within the DSS (dotted red lines in schematic). The DSS, in combination with shore-based sampling and statistical HAB models, will provide event response “triggers” for shipboard sampling. 3.7. Shipboard Sampling & Experiments 3.7.1 Field collection. The overall goal of the shipboard work is to respond to and characterize HAB events as they are identified in the two regions during the period of field observations, and to conduct experimental studies at those times to test aspects of the biology of the HAB species present that might provide insight into key features explaining their success. When a potential HAB event has been identified (as described above) shipboard surveys will be initiated to characterize the feature. We need maximal flexibility to sample emerging events, and have therefore budgeted for local ship use rather than request UNOLS time (we cannot predict years in advance specifically when blooms will develop). Following the successful example of the RAPDALERT program (Schnetzer et al. 2007) and previous Bight regional monitoring surveys, a rapid spatial survey will be conducted to determine if HAB species are present in the observed chlorophyll maxima. If an HA event is potentially initiating (determined by HA presence, toxin analysis, confirmation from the ESP moorings, etc.) we will initiate a comprehensive shipboard sampling survey. For this study “HAB events” include the observational spectrum from high relative densities (regardless of toxin quota) of HAB species through low densities of cells with high toxin quotas. The stations will be selected for water collection based on bloom data from the gliders, moorings, ESP’s and in Monterey Bay, high-resolution vertical profiles from an Acrobat towed vehicle (no cost to this proposal). Representative stations will then be identified for sampling using a CTD rosette system. 3.7.2. Standard analyses. Samples to be analyzed from the grid and the selected stations include pigments, nutrients (nitrate+nitrite, ammonium, urea, silicic acid, phosphate), species composition (to functional group if no HABs are present), toxins (DA and PSP), and physical information. If HABs are present, the collected water will be maintained in deckboard incubators and transported to shore for further experiments in environmental chambers. Rapid assessment will be conducted aboard the boats using field microscopes and Mercury Science field ELISA kits (“pucks”), which we have been developing in conjunction with Mercury Science (Litaker et al. 2008). Shore-based analysis will use standard protocols for nutrients (N, P, Si using FIA; ammonium and urea measured manually following the protocols of Holmes et al., 1999 and Goeyens et al. 1998) and chlorophyll a (Welschmeyer, 1994). Cell enumeration will be conducted using microscopy with settling chambers (full phytoplankton assemblage), FlowCAM analysis (for larger nanoplankton and microplankton), whole-cell probes (Miller and Scholin 1996, 1998, 2000) and/or quantitative PCR (Hayashi and Smith 2005; Caron unpublished) for toxigenic species, and SEM as necessary to confirm Pseudo-nitzschia to species (MLML recently acquired a new SEM capability), and quantitative PCR for Alexandrium (being adapted now in the Caron lab) if PSP is present but cells abundances are too low for routine microscopic analysis. Toxin analysis for PSP toxins will utilize Abraxis ELISA kits; DA will be analyzed by Mercury Science ELISA with confirmatory LC/MS using an Agilent 6300 system maintained in the Kudela lab (Litaker et al. 2008). 3.7.3. Physiological assays. An important component of this project is to determine not only if HAB species are present, but also if these water masses at which we observe elevated

A Regional Comparison of Upwelling and Coastal Land Use Patterns on the Development of HAB Hotspots Along the California Coast
15
abundances (subsurface or surface) can serve as a source of toxic cells. To determine the competency of the community (e.g. are the cells actively growing or senescent/dead?) and to examine the specific conditions that are conducive to HAB species growth, we will examine nutrient and irradiance physiology and perform grazer perturbation experiments to determine general physiological status and growth response of targeted HAs. We will conduct a limited number of 15N (nitrate, ammonium, urea) experiments to characterize the nitrogenous physiology of these organisms, to assess potential sources of nutrients leading to the development of a bloom (e.g. runoff versus upwelled water versus wastewater effluent; Kudela et al. 2008b). Experiments will be conducted within 24 hours at in situ temperatures; all incubations will be conducted for the shortest reasonable time period (ca. 1-4 hours for 15N; see Kudela et al. 1997, Kudela and Chavez 1997, Kudela and Cochlan 2000; Kudela et al., 2008b,c; Cochlan et al., 2008 for similar experiments). In addition to these short-term experiments, we will conduct experiments in an attempt to manipulate the natural assemblage to produce a bloom; for example, if a subsurface layer is encountered, would it bloom under surface conditions (Wetz et al., 2004), or do nutrient perturbations change community structure or toxin production (Howard et al. 2007)? These “grow-out” type experiments will be conducted over 3-5 days using simulated environmental conditions (environmental chambers) or deckboard, (if the feature of interest warrants multiple ship days for a given event). These experiments will use different nitrogen sources, N:P ratios and two different light levels (corresponding to the surface versus subsurface light levels). We are not including an explicit trace-metal component, since maintaining TM-clean incubations is beyond the scope of this proposal. We can, however, assess the potential role of iron in these experiments by manipulating the available dissolved iron with desferoximine-B (Wells, 1999). Previous experiments conducted in Monterey (Kudela unpublished) demonstrate this technique to work well over a few days duration. As part of these grow-out experiments, we will perform a limited number of grazer-dilution experiments to assess HA community dynamics in relation to top-down and bottom-up pressures. These experiments can simultaneously assess light-nutrient interactions as well as the relative growth/grazing rates of HA species versus the entire community (Olson et al. 2008), and we have ample experience in the performance of grazing experiments. HA will be quantified using whole-cell probing methods and SEM (P. multiseries, P. australis, A. catenella) versus the bulk (chlorophyll) assemblage, keeping the workload to reasonable levels. Specific growth of P. multiseries and P. australis will be assayed relative to the native Pseudo-nitzschia community using ITS targeted QPCR (Hayashi et al. 2007) in order to assess specific stimulation of toxigenic species. Advances in grazer-dilution experimental methods also enable us to greatly reduce the number of dilutions, since mathematically it has been shown that 2 or 3 dilutions (1-2 highly diluted, 1 100% bottle) provide the same information as more elaborate, multiple-dilution methods (Gallegos 1989; Worden and Binder 2003, Scianni 2008), although these results must still be interpreted with some caution (Landry and Calbert 2005; Dolan and McKeon, 2005). The shipboard experimental data will be used to refine the DSS (3.6) and the ROMS and statistical models (3.2). Data on potential events will be provided to managers via the RCOOS and HABMAP networks, and ROMS will be used to estimate transport trajectories (see http://ourocean.jpl.nasa.gov/SCB/, “Drifter”, “Trajectory” for an example of the types of tools available). One metric of “success” will be defined as the ability to identify a potential event offshore, and track its transport (e.g. it stays offshore or advects onshore), coupling these responses to shore-based monitoring and wildlife impacts. A second success metric will be the refinement of these tools to better identify the next feature, while our ultimate success metric is the ability to quantitatively identify the ecophysiological conditions that lead to toxic Pseudo-nitzschia (or Alexandrium) events and incorporate that information into a predictive model. 3.8. Data Mining Year 5 will largely be devoted to data assimilation and hindcasts (using all of the available field data, ROMS, and statistical models) to evaluate whether there are consistent forcing factors driving HAB events in California. At the end of this program, we will have 10+ years of data and observations (5 years from this project, 5+ years from MERHAB and other projects). This will provide a rich data set for addressing post hoc hypotheses and extrapolating to future conditions. For example, there was an apparent shift in approximately 2004-2005 in

A Regional Comparison of Upwelling and Coastal Land Use Patterns on the Development of HAB Hotspots Along the California Coast
16
central California from diatom-dominated to a diatom/dinoflagellate community (Jester et al. 2009). We have previously linked these shifts to long-term changes in upwelling and stratification (Kudela et al., 2008d), while Pseudo-nitzschia dynamics have been linked to a common set of environmental conditions, including weak but not absent upwelling (Kudela et al., 2004). It has been difficult or impossible to directly test these hypotheses to date, because of the short data records (c.f. Lane et al. 2009). Similarly, long-term trends in the California Current suggest the coastal ocean (including HABs) is “greening,” and it is unclear what is driving these patterns (Kahru et al. 2009). We believe that if we are to understand these low-frequency oscillations and extrapolate to future conditions, our 10+ year data set, encompassing at least 1 ENSO cycle and potentially a “regime shift” in California (Chavez et al. 2003) is our best opportunity to do so. 4. Expected Results & Benefits Our primary result will be twofold: first, we will directly test our 3 hypotheses, developing a better understanding of HA initiation and bloom dynamics in California in response to physical and environmental forcing factors, enabling extrapolation to future conditions. Second, we will establish a cutting-edge HAB alert detection system and demonstrate the utility of an integrated observing system for harmful algal blooms as part of the Regional Coastal Ocean Observing System. In relation to the RFP, we also address several points: a) we will provide new monitoring tools within existing programs; b) we will conduct comprehensive data analysis and integration; c) we will develop real-time, scientific response, focusing on the critical and under-sampled nearshore environment not covered by existing shore-based monitoring programs; d) we will provide recommendations to SCCOOS, CeNCOOS, and other partners through HABMAP; and e) we will continue our outreach activities through CeNCOOS and HABMAP. 5. General Project Information 5.1. Project Team Management Our proposal brings an interdisciplinary and team together to work on a problem spanning several hundred kilometers of California coastline. Dr. Kudela will provide overall project management and will be responsible for Annual and Final Report preparation and dissemination, as well as for coordination of two open meetings, in Year 1 and 5. He is the Chair of the GEOHAB SSC, a member of the NEWS2USE SSC, and a member of the US National HAB Committee, Director of the UCSC Center for Remote Sensing, and an active participant in CeNCOOS and HABMAP. The majority of the team members have worked together previously; hence we expect a relatively smooth integration of the team components. In particular, we believe that the fruitful interactions between underwater robotics/sensor network personnel, HAB personnel and modelers gives this project a high chance for unique scientific advances. Clarissa Anderson (UCSC) developed predictive models for Pseudo-nitzschia in both California and the Chesapeake Bay (as part of an NRC postdoctoral fellowship with NOAA). She is part of the MERHAB Cal-PReEMPT project, creating preliminary hindcasts of Pseudo-nitzschia blooms and domoic acid events in the SBC using ROMS model data and satellite retrievals as inputs to current HAB prediction models. Anderson will take the lead on implementation and refinement of the statistical models, and application to the ROMS numerical model. Dave Caron (USC) is the lead PI for the Rapid Analysis of Pseudo-nitzschia and Domoic Acid, Locating Events in near-Real-Time (RAPDALERT) program, funded by NOAA-MERHAB. Caron is involved with development of HAB monitoring capabilities in Southern California, and interfaces directly with many of the Wildlife Rescue Groups. Caron is also a member of the Cal-HABMAP Steering Committee. Caron will take the lead on Southern California project activities. Yi Chao (JPL/UCLA) is the lead PI for several modeling projects off the coast of California. He has been heavily involved in the planning and implementation of ocean forecasting projects for both CeNCOOS and SCCOOS. Chao is also a participant in the modeling component of the Bight ’08 Regional Monitoring Program. Chao will be the lead on the numerical modeling component, using the ROMS model with data assimilation and forecasting capabilities. Meredith Howard (USC and SCCWRP) is a researcher at USC and at the Southern California Coastal Water Research Project (SCCWRP). She is the coordinator for both the Cal-HABMAP and Bight ’08 Regional Monitoring Programs and will be the main point

A Regional Comparison of Upwelling and Coastal Land Use Patterns on the Development of HAB Hotspots Along the California Coast
17
of contact for coordination with this project. She is also a participant in the SCCOOS HAB and the MERHAB RAPDALERT projects. Burton Jones (USC) is an interdisciplinary biological oceanographer who serves as chairman of the SCCOOS Executive Steering Committee. Jones has been involved in HAB research in California for many years, and has recently been working on the development of real-time monitoring efforts using technologies such as autonomous vehicles. Jones and Caron led the implementation of the SCOOS HAB monitoring effort and Jones will be in charge of Southern California glider operations. Heather Kerkering (CeNCOOS) is a Program Manager for the Central and Northern California Ocean Observing System. Her contributions will include coordination, outreach, education, engaging users, hosting workshops, and leading CeNCOOS staff in development of web-based products and reports for this effort. G. Jason Smith is a Research Faculty who has been studying the molecular ecophysiology of Pseudo-nitzschia and other phytoplankton for over fifteen years. His group has maintained a HA monitoring station at the Monterey Wharf II for 6 years and has developed ITS targeted QPCR assays for numerous Pseudo-nitzschia spp. As PI and Pacific Coast Technical Coordinator for NOAA ACT he has been active in evaluation of a range of monitoring technologies included HA species and toxin detection assays. Smith will be responsible for maintaining the MWII shore monitoring, coordinating and participating on small boat adaptive sampling surveys and grow out studies in Monterey Bay. Chris Scholin, John Ryan, and Jim Birch (MBARI) will lead the ESP deployments, and will be involved in field efforts for Monterey. Scholin pioneered development of the ESP and has overseen its application in coastal and deep ocean environments. Scholin will work to coordinate deployments of the ESP in Southern California and Monterey Bay. Birch has extensive instrumentation and project management experience in the areas of fluid mechanics, instrument design, and biodetection. He is the Instrumentation Group Leader at MBARI, and the project manager of the ESP program, overseeing the technical development of the ESP. Birch will oversee the preparation, deployment, and operation of the ESP instruments in both Monterey and S. California. Ryan has extensive experience conducting ecosystem process studies in Monterey Bay and analyzing multi-scale observations for studies of marine microbiology. He has led experiment design for multiple studies integrating ESP networks with intensive environmental observations from remote sensing and in situ platforms. Ryan will support ECOHAB experiment design, execution and data analysis. Kanna Rajan (MBARI) and Gaurav Sukhatme (USC) will take the lead on development of a Decision Support Tool framework for automated planning of autonomous vehicles (gliders in this case) using mooring and model data as inputs. Sukhatme is a member of both the Center for Embedded Network Sensors (CENS) and Center for Robotics and Embedded Systems (CRES) and is Director of the Robotic Embedded Systems Lab. Rajan, the Principal Researcher for Autonomy at MBARI, has developed and applied autonomy capabilities for space exploration and ocean research. His AI software has been operational on spacecraft, Mars Exploration Rovers, and autonomous underwater vehicles. Greg Doucette (NOAA) is a Research Oceanographer who has been active in HAB/Pseudo-nitzschia research in California for over ten years, primarily emphasizing measurement of domoic acid associated with natural Pseudo-nitzschia populations and examining trophic transfer of DA in local food webs. In collaboration with Scholin, the first autonomous, near real-time measurements of HAB species and toxin were made in Monterey Bay using the ESP; Doucette will be responsible for DA measurements on all ESP deployments during the project. 5.2. Relationship to Other Projects HABMAP: The California Harmful Algal Bloom Monitoring and Alert Program includes representatives from local, state, and federal research groups, academia, the Regional Coastal Ocean Observing Systems groups CeNCOOS and SCCOOS, and non-profit research centers such as MBARI and the Marine Mammal Center. A copy of the initial HABMAP report and other documentation is available at http://www.cencoos.org/sections/news/HABMAP.shtml. Since its inception, HABMAP has been actively involved with regional and national efforts to establish a coordinated HAB observing and forecasting system for the US West Coast. This includes representation from HABMAP on the West Coast HAB Summit coordinating committee that was established to develop the 2009 West Coast Regional HAB Summit, http://www.cop.noaa.gov/stressors/extremeevents/hab/current/HAB_Summit09/west_coast

A Regional Comparison of Upwelling and Coastal Land Use Patterns on the Development of HAB Hotspots Along the California Coast
18
_summit.html supported by the West Coast Governor’s Agreement; this group is currently working with NOAA to establish upcoming priorities for federally funded HAB research as part of the Harmful Algal Bloom & Hypoxia Research and Control Act (HABHRCA). NOAA MERHAB Projects: two NOAA MERHAB regional projects have been operating in California for the last 5 years (ending in 2010). These projects are central to this proposal in that they provide consistent and high quality data sets necessary for implementation of regional forecast models. Cal-PReEMPT: The California Program for Regional Enhanced Monitoring of Phyco-Toxins was established to bridge the gap between availability of new tools for cell and toxin detection and integration of those tools into State of California monitoring efforts. Representing a partnership between the California Department of Public Health, UCSC, and commercial shellfish growers, the program established pilot sites for method testing and development. Based on the initial success with the performance of field plankton identifications and the commercially-available diagnostic kits for toxins in shellfish, these tools were expanded to commercial shellfish growers in Santa Barbara and Morro Bay. This project demonstrated the critical need for consistent data to successfully develop predictive models; the Lane et al. (2009) publication describing predictive models for Monterey Bay was based nearly exclusively on Cal-PReEMPT data, because the remaining ~2000 data points available from Monterey between 1991-2008 were missing key parameters, of questionable quality, or otherwise compromised. RAPDALERT:The Rapid Analysis of Pseudo-nitzschia & Domoic Acid, Locating Events in near-Real Time Program is a collaborative research project including biologists, computer scientists and roboticists from USC, the UCLA, and SCCWRP. The overall goal of this project is the development of technology for the monitoring and study of HAB events in San Pedro Channel through the development of wirelessly networked sensor nodes (stationary sensors and autonomous underwater vehicles). This technology, linked to directed human response to collect samples, has been instrumental in documenting blooms of Pseudo-nitzschia and outbreaks of domoic acid poisoning in this region for the past several years. CeNCOOS & SCCOOS: the two Regional Coastal Ocean Observing Systems are key partners in both HABMAP and this proposal. The two programs are working closely to maintain consistency in HAB monitoring and research along the entire California coast and each can provide expertise in data management, data dissemination, and public outreach. They provide more consistent, long-term operational support not achievable by academic research programs, and are somewhat buffered from the volatile State budget, since primary funding comes from the federal government. They are actively engaged in the development and actions of the West Coast Governors Agreement on Ocean Health and in outreach to numerous user groups. SBC Project – Persistence and fate of domoic acid in the Santa Barbara Basin: This NSF-funded project is led by C. Benitez-Nelson at the University of South Carolina and got started in March 2009. The greater goal of the project is to quantify effects on domoic acid properties as it is vertically transported to depth after the demise of surface blooms since it is now well-known that domoic acid not only accumulates in benthic organisms but persists at the sediment interface beyond seasonal timescales. The study conducts field sampling of water column Pseudo-nitzschia and domoic acid observations during monthly cruises in the SBC. While we are not directly focusing on the SBC, this will be a valuable data set and project analyzed as part of our comparative approach; our project will benefit from insights gained in the SBC, and vice versa. PNWTOX--The Columbia River plume and HABs in the Pacific Northwest: bio-reactor, barrier or conduit? This is a project funded jointly by NSF and NOAA. The overall approach of this project will be to study HAB related processes with merged datasets from ECOHAB-PNW, and CoOP River Influences on Shelf Ecosystems, funded by NOAA and NSF, including growth and grazing rates, phytoplankton and microzooplankton community structure, Pseudo-nitzschia, nutrients, domoic acid time series and spatial maps, satellite imagery, as well as transport data from moored arrays and water properties over the whole domain. At the same time existing transport data will be used to develop a state of the art physical model for the region including both potential HAB initiation sites, the Columbia River plume and freshwater upwelling sources such as the entrance to the Strait of Juan de Fuca. PNWTOX is relevant to this proposal for several reasons. First, Kudela is a co-I, and will be (among other tasks) providing historical and

A Regional Comparison of Upwelling and Coastal Land Use Patterns on the Development of HAB Hotspots Along the California Coast
19
ongoing satellite data. This effort will be used as match to this proposal, since the entire west coast can be processed simultaneously. Second, the PNW is clearly dominated by physical transport; this will be another valuable comparative system for inclusion in our summary analyses, identifying similarities and differences between the PNW and California. The Bight ’08 Regional Monitoring Program The Southern California Bight (SCB) Regional Monitoring Program focuses on assessing the condition of the coastal ocean and is conducted in cooperation with over 30 organizations in southern California once every five years. The overall goals are to quantify the major nutrient sources to the Southern California Bight (atmospheric deposition, upwelling, wastewater effluent and riverine inputs) and to characterize the extent, magnitude, and ecological characteristics of Pseudo-nitzschia blooms. The relative nutrient (N, P, Si) contributions of four major sources to the SCB will be quantitatively estimated, and comparative loading for anthropogenic versus natural nutrient sources will also be determined. These data will be used to assess the timing and magnitude of nutrient delivery to the coastal ocean relative to remotely-sensed and field observations of HABs. The field season for most of the data collection is in the spring of 2010 with the intention of collecting data during pre-bloom, bloom, and post-bloom conditions. This project will be the largest spatial HAB dataset collected in California and will provide a “Year 1” dataset for our project. Controlled, Agile, and Novel Observing Network--CANON (MBARI): A number of MBARI research engineers and scientists are undertaking a feasibility study for an institute initiative that will push the limits of prediction, detection, and response in the coastal ocean using HABs as the candidate example of a phenomenon that may be linked to human activity, with significant societal importance. The first step in 2009 will be to conduct a system engineering study of which marine assets will provide the best first detection capabilities for HABs for the investment, given the known spatial and temporal scales for HAB development and the needs of resource managers for information and response. This effort will directly inform our proposal, and we are leveraging expertise and assets at reduced or no cost for this effort. California Ocean Protection Council: A separate proposal was submitted (2009) to the OPC through California Sea Grant. Although funding was awarded to a competing proposal (on salmon recovery), the OPC indicated that they will fund some aspects of the proposal. The primary objectives were to implement existing HAB models from Santa Barbara and Monterey Bay previously developed as research exercises, to test and expand these existing models in other regions, to begin developing a similar modeling effort for paralytic shellfish poisoning, and to provide consistent field monitoring and validation data to adequately assess the model results. We have taken this into consideration for this proposal, and anticipate that OPC will fund work aimed at development of a real-time and forecast capability for HABs in California through the use of statistical models. II. Application to Management 1. Coordination with End Users Members of this proposal are actively involved with ongoing research and monitoring programs as detailed in the Associated Work section, as well as with the West Coast HAB Summit, the various sanctuary programs, the National HAB Committee, the GEOHAB program, the Alliance for Coastal Technology, and many other related efforts. To coordinate activities with these other groups and disseminate information, we will build on the newly established HABMAP communication network. We will leverage the distributed databases established by CeNCOOS and SCCOOS for data management and interface with end-users. CeNCOOS and SCCOOS will communicate with their many regional partners and neighboring Regional Associations on the status and value of this effort as well as request their participation in providing feedback for product development and information distribution. To that end, we include one month salary support for Heather Kerkering, CeNCOOS Program Coodinator, in Year 1; Kerkering will provide an additional 3 weeks of unfunded support in the out years. In a separate proposal to OPC (“Statewide Alert Network for Harmful Algal Blooms, CalHABMAP: California Harmful Algal Bloom Monitoring and Alert Program”) the following organizations were identified as key members of HABMAP: NOAA, Environmental Protection

A Regional Comparison of Upwelling and Coastal Land Use Patterns on the Development of HAB Hotspots Along the California Coast
20
Agency, Food and Drug Administration, California State Water Resources Control Board, Southern California Regional Water Boards, University of California Santa Cruz, UC Santa Barbara, UC Los Angeles, UC Irvine, UC Davis, Scripps Institute of Oceanography, University of Southern California, California Institute of Technology, California State Polytechnic University, California State University Long Beach, University of Washington, University of South Carolina, Humboldt State University, San Francisco State University, Moss Landing Marine Labs, MBARI, Oregon State University, Southern California Coastal Water Research Project, California Ocean Science Trust, Heal the Bay, The Marine Mammal Center, Pacific Marine Mammal Center, Monterey Abalone Company, Fishtech, Inc., International Bird Rescue Research Center, Wetlands and Wildlife Care Center, Pacific Abalone Farms, Pacific Shellfish Institute, The Abalone Farm, Inc., Hog Island Oyster Co., California Council for Wildlife Rehabilitators, Orange County Sanitation District, California Department of Public Health, Oregon Department of Fish and Wildlife, California Department of Fish and Game. Our direct participation with CeNCOOS, SCCOOS, ACT, and HABMAP will thus ensure timely and responsive feedback between project members and end-users. 2. Expected Significance As a research proposal, our primary outcomes from this project will be peer-reviewed manuscripts, but we anticipate several products of direct use to end-users and managers: • 5 year ROMS output for the Monterey Bay Region. The ROMS model output will be freely available to other users. This is a priority for CeNCOOS, but funding has not been available to support continuous modeling efforts other than the targeted field programs in 2006 and 2008. • Improved statistical models for HAB forecasting. Our results feed directly into the proposed modeling/forecasting effort to be supported by the California Ocean Protection Council. • Continued monitoring for HABs. Other than the shore-based monitoring program maintained by the California Department of Publich Health, there are no permanent HAB observing networks in California; we rely on the continued funding of research projects such as this one to provide real-time event response and alert to managers to augment the (heavily impacted by recent budget cuts) State program. • Improved understanding of bloom/toxin initiation in two CA hotspots. This will directly inform and provide leverage for future management and mitigation efforts supported by NOAA. • The Slocum electric glider purchased for Monterey will be maintained and used for other relevant programs by CeNCOOS; funds will be sought for expanded HAB monitoring in autumn. 3. Activities Our primary objectives are research-based, so we place relatively less emphasis on management and outreach activities, but we anticipate outreach and education will continue through CeNCOOS and SCCOOS. For example, as part of Cal-PReEMPT funding, we routinely work with CeNCOOS to develop outreach and education tools via their web portals (www.cencoos.org/sections/classroom/algal_blooms.html and www.cencoos.org/sections/conditions/algal_blooms.html). We have budgeted funds in Years 1 and 5 to host open project meetings, where we anticipate inviting representatives from the human health, wildlife health, and research communities to discuss our planned activities (Year 1) and outcomes (Year 5) coordinated through SCCOOS, CeNCOOS, and HABMAP. We will use HABMAP for Years 2-4 to keep the community updated on our progress and event response. We also expect that the Alliance for Coastal Technologies will conduct a cell and toxin detection comparison, as recommended by the ACT workshop “Technologies and Methodologies for the Detection of Harmful Algae and Their Toxins”. Our team will participate (GJ Smith is the West Coast ACT Coordinator); if the ACT technology demonstration does not go forward, HABMAP anticipates finding funds for a similar exercise focused on the California region. Either (or both) demonstrations would benefit from participation and access to the data and technologies supported by this proposal. We envision, for example, that a demonstration by ACT/HABMAP in conjunction with one of our field seasons would substantially leverage NOAA funds (shiptime, ESP, glider access, cell and toxin ID as part of this project) with minimal additional effort on our part. Finally, we include training for 6 graduate students/year on a multidisciplinary project.

A Regional Comparison of Upwelling and Coastal Land Use Patterns on the Development of HAB Hotspots Along the California Coast
21
REFERENCES
Prior Support Publications—UCSC MERHAB 1. Kudela, RM, MDA Howard, B Jenkins, PE Miller, and GJ Smith. Revised, pending
acceptance. Using the molecular toolbox to compare harmful algal blooms in upwelling systems. Progress in Oceanography.
2. Kudela, RM, S Seeyave, and WP Cochlan. Revised, pending acceptance. The Role of Nutrients in Regulation and Promotion of Harmful Algal Blooms in Upwelling Systems. Progress in Oceanography.
3. Ryan, JP, A Fischer, R Kudela, J Gower, S King, R Marin III, and F Chavez. 2009. Influences of upwelling and downwelling winds on red tide bloom dynamics in Monterey Bay, California. Continental Shelf Research 29: 785-795.
4. Anderson, CR, DA Siegel, RM Kudela, and MA Brzezinski. 2009. Empirical models of toxigenic Pseudo-nitzschia blooms: potential use as a remote detection tool in the Santa Barbara Channel. Harmful Algae, 8, 478-492.
5. Howard, MDA, GJ Smith, and RM Kudela. 2009. Phylogenetic relationships of yessotoxin-producing dinoflagellates based on the large subunit and internal transcribed spacer ribosomal DNA. Applied and Environmental Microbiology, 75: 54-63, doi:10.1128/AEM.00818-08.
6. Jessup DA, MA Miller, JP Ryan, HM Nevins, HA Kerkering, A Mekebri, DB Crane, TA Johnson, and RM Kudela. 2009. Mass stranding of marine birds caused by a surfactant-producing red tide. Public Library of Science—ONE, 4(2): e4550. doi:10.1371/journal.pone.0004550.
7. Lane, JQ, P Raimondi, and RM Kudela. 2009. The development of a logistic regression model for the prediction of toxigenic Pseudo-nitzschia blooms in Monterey Bay, California. Marine Ecology Progress Series 383:37-51.
8. Ryan, JP, A Fischer, R Kudela, J Gower, S King, R Marin III, and F Chavez. 2009. Influences of upwelling and downwelling winds on red tide bloom dynamics in Monterey Bay, California. Continental Shelf Research 29:785-795.
9. Anderson, D., J. Burkholder, W. Cochlan, P. Glibert, C. Gobler, C. Heil, R. Kudela, M. Parsons, J. Rensell, D. Townsend, V. Trainer, and G. Vargo. 2008. Harmful algal blooms and eutrophication: Examples and linkages from selected coastal regions of the United States, Harmful Algae, 8, 39-53.
10. Cochlan, WP, J Herndon, and RM Kudela. 2008. Inorganic and Organic Nitrogen Uptake by the toxigenic diatom Pseudo-nitzschia australis (Bacillariophyceae). Harmful Algae 8, 111-118.
11. Curtiss, C., Langlois, G., Busse, L., Mazzillo, F., & Silver, M. 2008. The emergence of Cochlodinium along the California Coast (USA). Harmful Algae, 7, 337-346.
12. Litaker RW, Stewart TN, LeEberhardt B-TL, Wekell JC, Trainer VL, Kudela RM, Miller PE, Roberts A, Hertz C, Johnson TA, Frankfurter G, Smith GJ, Schnetzer A, Schumaker J, Bastian JL, Odell A, Gentien P, Le Gal D, Hardison DR, Tester PA. 2008. Rapid enzyme-linked immunosorbent assay for detection of the algal toxin domoic acid. Journal of Shellfish Research, 27:1301-1310
13. Glibert, PM, R Azanza, M Burford, K Furuya, E Abal, A Al-Azri, F Al-Yamani, P Andersen, DM Anderson, J Beardall, GM Berg, L Brand, D Bronk, J Brookes, JM Burkholder, A Cembella, WP Cochlan, J Collier, Y Collos, R Diaz, M Doblin, T Drennen, S Dyhrman, Y Fukuyo, M Furnas, J Galloway, E Granéli, DV Ha, G Hallegraeff, J Harrison, PJ Harrison, C

A Regional Comparison of Upwelling and Coastal Land Use Patterns on the Development of HAB Hotspots Along the California Coast
22
Heil, K Heimann, R Howarth, C Jauein, AA Kana, TM Kana, H Kim, R Kudela, C Legrand, M Mallin, M Mulholland, S Murray, J O’Neil, G Pitcher, Y Qi, N Rabalais, R Raine, S Seitzinger, PS Salomon, C Solomon, DK Stoecker, G Usup, J Wilson, K Yin, M Zhou, and M Zhu. 2008. Ocean urea fertilization for carbon credits pose high ecological risks. Marine Pollution Bulletin, 56, 1049-1056
14. Goldstein, T, Zabka, TS, DeLong, RL, Wheeler, EA, Ylitalo, G, Bargu, S, Silver, M, Leighfield, T, Van Dolah, F, Langlois, G, Sidor, I, Dunn, JL, Gulland, FMD. 2009. The role of domoic acid in abortion and premature parturition of California sea lions (Zalophus californianus) on San Miguel Island, California. J. Wildlife Diseases, 45 (1): 91-108.
15. Howard, M, Silver, M, & Kudela, R. 2008. Yessotoxin detected in mussel (Mytilus californicus) and phytoplankton samples from the US west coast. Harmful Algae, 7, 646-652, doi: 10.1016/j.hal.2008.01.003.
16. Jester, R.; Lefebvre, K.; Langlois, G.; Vigilant, V.; Baugh, K.; Silver, M.W. 2009. A shift in the dominant toxin-producing algal species in central California alters phycotoxins in food webs. Harmful Algae, 8: 291-298.
17. Kudela, R.M., JQ Lane, and WP Cochlan. 2008. The potential role of anthropogenically derived nitrogen in the growth of harmful algae in California, USA. Harmful Algae 8, 103-110.
18. Kudela, R.M., J.P. Ryan, M. Blakely, J.Q. Lane, and T.D. Peterson. 2008. Linking the physiology and ecology of Cochlodinium to better understand harmful algal bloom events: a comparative approach. Harmful Algae, 7, 278-292.
19. McManus, M, R Kudela, M Silver, G Steward, P Donaghay, and J Sullivan. 2008. Cryptic blooms: Are thin layers the missing connection? Estuaries and Coasts. 31(2), 396-401.
20. Ryan, JP, JFR Gower, SA King, WP Bissett, AM Fischer, RM Kudela, Z Kolber, FM Mazzillo, EV Rienecker, and FP Chavez. 2008. A coastal ocean extreme bloom incubator. Geophysical Research Letters 35, L12602, doi:10.1029/2008GL034081.
21. Schnetzer A., P. E. Miller, R. A. Schaffner, B. Stauffer, B. Jones, S. B. Weisberg, P. M. DiGiacomo, W. Berelson, and D. A. Caron. 2007. Pseudo-nitzschia spp. and domoic acid in the San Pedro Channel and Los Angeles harbor areas of the southern California bight, 2003-2004. Harmful Algae 6(3): 372-387.
22. Busse, L. B., E. Venrick, R. Antrobus, P. E. Miller, V. Vigilant, M. Silver, C. Mengelt, L. Mydlarz, B. Prezelin. 2006. Domoic acid in phytoplankton and fish in San Diego, CA, USA. Harmful Algae 5(1): 91-101.
23. Kudela, R.M. 2006. Linking the physiology and ecology of the harmful red tide organism Cochlodinium to better understand bloom events. Recent progress on the research and management of Cochlodinium blooms pp. 51-56, Cheju National University, Republic of Korea
24. Lundholm, N., O. Moestrup, Y. Kotaki, K. Hoef-Emden, C. Scholin, P. Miller. 2006. Inter- and intraspecific variation of the Pseudo-nitzschia delicatissima complex (Bacillariophyceae) illustrated by rRNA probes, morphological data and phylogenetic analyses. Journal of Phycology 42:464-481.
25. O'Halloran, C., M.W. Silver, T.R. Holman, and C.A. Scholin. 2006. Heterosigma akashiwo in central California waters. Harmful Algae, 5(2):124-132.
26. Silver, M.W. Protecting Ourselves from Shellfish Poisoning. American Scientist. July-. August 2006: 316-325.

A Regional Comparison of Upwelling and Coastal Land Use Patterns on the Development of HAB Hotspots Along the California Coast
23
27. Fire, S.E., and M.W. Silver. 2005. Domoic acid in the Santa Cruz wharf fishery. California Fish and Game, 91:179-192.
28. Kudela, R., G. Pitcher, T. Probyn, F. Figueiras, T. Moita, and V. Trainer. 2005. Harmful Algal Blooms in coastal upwelling systems. Oceanography, 18(2): 184-197.
29. Ryan, J.P., H.M. Dierssen, R.M. Kudela, C.A. Scholin, K.S. Johnson, F.P. Chavez, A.M. Fischer, E.V. Rienecker, P. McEnaney, R. Marin, and J.M. Sullivan. Coastal ocean physics and red tides. Oceanography, 18(2): 214-223.
Prior Support Publications--USC MERHAB 1. Caron DA, Garneau M-E, Seubert E, Howard MDA, Darjany D, Schnetzer A, Cetinic I,
Filteau G, Lauri P, Jones B and Trussell S (2009) Harmful algae and their potential impacts on desalination operations off southern California. Water Research. In press.
2. Caron DA (2009) New accomplishments and approaches for assessing protistan diversity and ecology in natural ecosystems. BioScience 59:287-299.
3. Caron DA (2009) Protistan biogeography: why all the fuss? J Euk Microbiol 56:105-112. 4. Caron DA, Countway PD (2009) The protistan Rare Biosphere in aquatic microbial ecology:
hypotheses on the role of diversity in a changing world. Aq Microb Ecol: In press. 5. Caron DA, Countway PD, Savai P, Gast RJ, Schnetzer A, Moorthi SD, Dennett MR, Jones
AC (2009) Defining molecular taxonomic units for microbial eukaryote ecology. Appl Environ Microbiol: In press.
6. Caron DA, Gast RJ (2008) The diversity of free-living protists: seen and unseen, cultured and uncultured. In: Zengler K (ed) Accessing uncultivated microorganisms: From the environment to organisms and genomes and back. ASM Press, Washington, DC, p 67-93.
7. Caron DA, Gast RJ, Countway PD, Heidelberg KB (2009) Microbial eukaryote ecology: questions of diversity and biogeography. Microbe 4:71-77.
8. Caron DA, Schnetzer A (2007) Protistan community structure. In: Hurst CJ, Crawford RL, Garland JL, Lipson DA, Mills AL, Stetzenbach LD (eds) Manual of environmental microbiology. ASM Press, Washington, p 454-468.
9. Caron DA, Stauffer BA, Moorthi SD, Singh A, Batalin M, Graham E, Hansen M, Kaiser WJ, Das J, Pereira A, Dhariwal A, Zhang B, Oberg C, Sukhatme GS (2008) Macro- to fine-scale spatial and temporal distributions and dynamics of phytoplankton and their environmental driving forces in a small subalpine lake in southern California, USA. Limnol Oceanogr 58:2333-2349.
10. Fitzpatrick E, Caron DA, Schnetzer A. Development and environmental application of a genus-specific quantitative PCR approach for Pseudo-nitzschia species. Submitted to Marine Biology.
11. Gast RJ, Moran DM, Dennett MR, Caron DA (2007) Kleptoplasty in an Antarctic dinoflagellate: caught in evolutionary transition? Environ Microbiol 9:39-45.
12. Lee AS, Mahapatro M, Caron DA, Requicha AAG, Stauffer B, Thompson ME, Zhou C (2006) Whole-cell sensing for a harmful bloom-forming alga by measuring antibody-antigen forces. IEEE Trans Nanobioscience 5:149-156.
13. Moorthi SD, Countway PD, Stauffer BA, Caron DA (2006) Use of quantitative real-time PCR to investigate the dynamics of the red tide dinoflagellate Lingulodinium polyedrum. Microb Ecol 52:136-150.

A Regional Comparison of Upwelling and Coastal Land Use Patterns on the Development of HAB Hotspots Along the California Coast
24
14. Pereira A, Heidarsson H, Oberg C, Caron DA, Jones B, Sukhatme GS. A communication framework for cost-effectie operation of AUVs in coastal regions. Field Service Robotics. In press.
15. Schnetzer A, Miller PE, Schnaffner RA, Stauffer BA, Jones BH, Weisberg SB, DiGiacomo PM, Berelson WM, Caron DA (2007) Blooms of Pseudo-nitzschia and domoic acid in the San Pedro Channel and Los Angeles harbor areas of the Southern California Bight, 2003-2004. Harmful Algae 6:372-387.
16. Singh A, Batalin M, Stealey MJ, Zhang B, Dhariwal A, Stauffer B, Moorthi S, Oberg C, Pereira A, Chen V, Lam Y, Caron DA, Hansen M, Kaiser WJ, Sukhatme G. 2007. Human assisted robotic team campaigns in aquatic monitoring. J Field Robotics 24:969–989.
17. Smith RN, Chao Y, Jones BH, Caron DA, Li PP, Sukhatme GS. Trajectory design for autonomous underwater vehicles based on ocean model predictions for feature tracking. Field Services Robotics. In press.
18. Sukhatme GS, Dhariwal A, Zhang B, Oberg C, Stauffer B, Caron DA. 2007. The Design and Development of a Wireless Robotic Networked Aquatic Microbial Observing System. Environ Eng Sci 24:205-215.
19. Pereira, A., H. Heidarsson, H., C. Oberg, D.A. Caron, B. Jones and G.S. Sukhatme. In press. A communication framework for cost-effective Operation of AUVs in coastal regions. The 7th International Conference on Field and Service Robotics (FSR-2009), Cambridge, MA, July 2009.
20. Smith, R., Y. Chao, B.H. Jones, D.A. Caron, PP. Li and G. S. Sukhatme. In press. Trajectory design for autonomous underwater vehicles based on ocean model predictions for feature tracking. Proceedings of the International Conference on Field and Service Robotics.
21. Smith, R.N., J. Das, H. Heidarsson, A.M. Pereira, F. Arrichiello, I. Cetinic, L. Darjany, M.-E. Garneau, M.D. Howard, C. Oberg, M. Ragan, E. Seubert, E.C. Smith, B.A. Stauffer, A. Schnetzer, G. Toro-Farmer, D.A. Caron B.H. Jones and G.S. Sukhatme. In press. The USC Center for Integrated Networked Aquatic PlatformS (CINAPS): observing and monitoring the Southern California Bight. IEEE Robotics and Automation Magazine.
22. Smith, R., J. Das, H. Heidarsson, A. de Menezes Pereira, D.A. Caron, B.H. Jones, and G.S. Sukhatme. "Implementation of an Embedded Sensor Network for the Coordination of Slocum Gliders for Coastal Monitoring and Observation", To appear in ACM International Workshop on UnderWater Networks, Berkeley, CA, Nov 2009.
23. Smith, R., B. Stauffer, J. Das, H. Heidarsson, A. de Menezes Pereira, Y. Chao, L. Darjany, I. Cetinic, D. Estrin, C. Oberg, M. Ragan, B.H. Jones, G.S. Sukhatme, and D.A. Caron. "Design and Implementation of Sensor Networks for the Observation and Research of Harmful Algal Blooms in Southern California Coastal Waters", In Proceedings of the Conference on Coastal Environmental Sensing Networks, Boston, MA, Jul 2009.
Prior Support Publications--Doucette 1. Doucette, G.J., et al. 2009. Remote, subsurface detection of the algal toxin domoic acid onboard
the Environmental Sample Processor: assay development and field trials. Harmful Algae 8:880-888.
2. Scholin, C., et al. 2009. Remote detection of marine microbes, small invertebrates, harmful algae and biotoxins using the Environmental Sample Processor (ESP). Oceanography 22:158-167

A Regional Comparison of Upwelling and Coastal Land Use Patterns on the Development of HAB Hotspots Along the California Coast
25
3. Greenfield, D., et al. 2008. Field applications of the second-generation Environmental Sample Processor (ESP) for remote detection of harmful algae: 2006-2007. Limnol. Oceanogr.: Methods 6: 667-679.
4. Greenfield, D.I., et al. 2006. Application of the Environmental Sample Processor (ESP) methodology for quantifying Pseudo-nitzschia australis using ribosomal RNA-targeted probes in sandwich and fluorescent in situ hybridization. Limnol. Oceanogr.: Methods 4: 426-435.
Prior Support Publications--MBARI 1. Doucette, G.J., Mikulski, C.M., Jones, K.L., King, K.L., Greenfield, D.I., Marin III, R.,
Jensen, S., Roman, B., Elliott, C.T., Scholin, C.A. 2009. Remote, subsurface detection of the algal toxin domoic acid onboard the Environmental Sample Processor: assay development and field trials. Harmful Algae. doi:10.1016/j.hal.2009.04.006.
2. Goffredi, S.K., W. Jones, C. Scholin, R. Marin, S. Hallam, R.C. Vrijenhoek. 2006. Molecular detection of marine larvae. Marine Biotechnology 8: 149-160.
3. Greenfield, D., et al. 2008. Field applications of the second-generation Environmental Sample Processor (ESP) for remote detection of harmful algae: 2006-2007. Limnol. Oceanogr.: Methods 6: 667-679.
4. Greenfield, D.I., et al. 2006. Application of the Environmental Sample Processor (ESP) methodology for quantifying Pseudo-nitzschia australis using ribosomal RNA-targeted probes in sandwich and fluorescent in situ hybridization. Limnol. Oceanogr.: Methods 4: 426-435.
5. Jones, W.J., et al. 2007. A Robotic Molecular Method for in situ Detection of Marine Invertebrate Larvae. Mol. Ecol. Res. 8: 540-550.
6. Marin III, R., and C. Scholin. Sandwich Hybridization. In: Microscopic and molecular methods for quantitative phytoplankton analysis (Ed. B. Edvardsson et al.). IOC Manuals and Guides (in press).
7. Metfies, K., K. Töbe, C. Scholin, and L.K. Medlin. 2006. Laboratory and field applications of ribosomal RNA probes to aid the detection and monitoring of harmful algae. In: Ecology of Harmful Algae (Granéli, E., and Turner, J.T. eds), pp. 311-325. Springer Verlag, Berlin, Heidelberg, New York.
8. Paul, J. et al. 2007. In situ Instrumentation. Oceanography 20: 58-66. 9. Preston, C., et al. 2009. Near real-time, autonomous detection of marine bacterioplankton on
a coastal mooring in Monterey Bay, California, using rRNA-targeted DNA probes. Environmental Microbiology 11: 1168-1180.
10. Roman, B., C. Scholin, S. Jensen, E. Massion, R, Marin III, C. Preston, D. Greenfield, W. Jones, and K. Wheeler. 2007. Controlling a Robotic Marine Water Sampler with the Ruby Scripting Language. Journal of American Laboratory Automation 12: 56-61.
11. Scholin, C., Doucette, G., Jensen, S., Roman, B., Pargett, D., Marin III, R., Preston, C., Jones, W., Feldman, J., Everlove, C., Harris, A., Avarado, N., Massion, E., Birch, J., Greenfield, D., Wheeler, K., Vrijenhoek, R., Mikulski, C., Jones, K. 2009. Remote detection of marine microbes, small invertebrates, harmful algae and biotoxins using the Environmental Sample Processor (ESP). Oceanography 22: 158-167.
Prior Support Publications—Anderson 1. Sekula-Wood, E.S., Schnetzer, A., Benitez-Nelson, C.R., Anderson, C.R., Berelson, W.,
Burns, J., Caron, D., Ferry, J., Morton, S.L., Thunell, R. (2009) The toxin express: Rapid

A Regional Comparison of Upwelling and Coastal Land Use Patterns on the Development of HAB Hotspots Along the California Coast
26
downward transport of the neurotoxin domoic acid in coastal waters. Nature Geoscience 2(4): 272-275, doi: 10.1038/NGEO472
2. Anderson, C.R., Siegel, D.A., Kudela, R.M., and Brzezinski, M.A. (2009) Empirical habitat models of toxigenic Pseudo-nitzschia spp.: Potential use as a remote detection tool in the Santa Barbara Channel. Harmful Algae, 8: 478-492, doi: 10.1016/j.hal.2008.10.005
3. Anderson, C.R., Sapiano, M.R.P.,Prasad, M. B.K., Long, W., Tango, P.J., Brown, C.W., Murtugudde, R. Predicting toxigenic Pseudo-nitzschia blooms in the Chesapeake Bay. Submitted, Marine Ecology Progress Series
4. Prasad, M.B.K., Sapiano, M.R.P., Long, W., Anderson, C.R., Murtugudde, R. Long-term variability of nutrients in the Chesapeake Bay: A retrospective analysis, 1990-2006. Submitted, Estuaries and Coasts
References—Main Proposal Ackerman, D., Schiff, K., (2003) Modeling storm water mass emissions to the southern
California bight. Journal of Environmental Engineering-Asce 129 (4), 308-317.
Anderson, C., M. Brzezinski, L. Washburn, and R. Kudela (2006), Circulation and environmental conditions during a toxigenic Pseudo-nitzschia australis bloom in the Santa Barbara Channel, California, Marine Ecology Progress Series, 327, 119-133.
Anderson, C., D. Siegel, R. Kudela, and M. Brzezinski (2009), Empirical models of toxigenic Pseudo-nitzschia blooms: Potential use as a remote detection tool in the Santa Barbara channel, Harmful Algae, 8, 478-492, doi:10.1016/j.hal.2008.10.005.
Anderson, D., J. Burkholder, W. Cochlan, P. Glibert, C. Gobler, C. Heil, R. Kudela, M. Parsons, J. Rensell, D. Townsend, V. Trainer, and G. Vargo (2008), Harmful algal blooms and eutrophication: Examples and linkages from selected coastal regions of the United States, Harmful Algae, 8, 39-53.
Anderson, D., D. Kulis, B. Keafer, K. Gribble, R. Marin, and C. Scholin (2005), Identification and enumeration of Alexandrium spp. from the Gulf of Maine using molecular probes, Deep-Sea Research II, 52, 2467-2490.
Ayers K, Rhodes L, Tyrrell J, Gladstone M, Scholin C. (2005) International accreditation of sandwich hybridisation assay format DNA probes for micro-algae. New Zealand J. Marine and Freshwater Res. 39: 1225–1231.
Bates, S. (1998), Ecophysiology and metabolism of ASP toxin production, in Physiological ecology of harmful algal blooms, edited by D. Anderson, et al., pp. 405-426, Springer-Verlag, Heidelberg.
Bates, S., D. Garrison, and R. Horner (1998), Bloom dynamics and physiology of domoic-acid producing Pseudo-nitzschia species, in Physiological ecology of harmful algal blooms, edited by D. Anderson, et al., pp. 267-292, Springer-Verlag, Heidelberg.
Bates, S., and V. Trainer (2006), The ecology of harmful diatoms, in Ecology of Harmful Algae, edited by E. Granéli and J. Turner, Springer-Verlag, Heidelberg.
Blum, I, DV Subba Rao, Y Pan, S Swaminathan, and NG Adams (2006) Development of statistical models for prediction of the neurotoxin domoic acid levels in the pennate diatom Pseudonitzschia pungens f. multiseries utilizing data from cultures and natural blooms. In

A Regional Comparison of Upwelling and Coastal Land Use Patterns on the Development of HAB Hotspots Along the California Coast
27
Algal cultures, analogues of blooms and applications, edited by D. Subba Rao: Science Publishers Inc.
Breaker, L. C., V. M. Krasnopolsky, D. B. Rao, and X. H. Yan (1994), The Feasibility of Estimating Ocean Surface Currents on an Operational Basis Using Satellite Feature Tracking Methods, Bull Amer Meteorol Soc, 75, 2085-2095.
Bresina, J., A.K. Jonsson, P.H. Morris and K. Rajan (2005), Activity Planning for the Mars Exploration Rovers, Intnl. Conf. on Automated Planning and Scheduling, June 2005, Monterey, CA.
Busse, L., E. Venrick, R. Antrobus, P. Miller, V. Vigilant, M. Silver, C. Mengelt, L. Mydlarz, and B. Prézelin (2006), Domoic acid in phytoplankton and fish in San Diego, CA, USA, Harmful Algae, 5, 91-101.
Caron DA, Garneau M-E, Seubert E, Howard MDA, Darjany D, Schnetzer A, Cetinic I, Filteau G, Lauri P, Jones B and Trussell S (2009) Harmful algae and their potential impacts on desalination operations off southern California. Water Research. In press.
Cassie R.M. (1963) Microdistribution in the plankton. Oceanogr. Mar. Bio. Annu. Rev. 1:223-252. Wiebe P. and W. R. Holland. 1968: Plankton patchiness: effects of repeated net tows. Limnol.
Oceanogr., 13: 315-321.
Chao, Y., Li, Z., Farrara, J.D., Moline, M., Schofield, O.M.E., Majumdar, S.J., (2008), Synergistic applications of autonomous underwater vehicles and regional ocean modeling system in coastal ocean forecasting. Limnol. Oceanogr., 53 (5), 2251–2263.
Chao, Y., Z., L., Farrara, J., McWillliams, J.C., Bellingham, J., Capet, X., Chavez, F., Choi, J.K., Davis, R., Doyle, J., Frantaoni, D., Li, P.P., Marchesiello, P., Moline, M.A., Paduan, J., Ramp, S., (2009), Development, implementation and evaluation of a data-assimilative ocean forecasting system off the central Deep-Sea Research II.
Chavez, F. P., J. Ryan, S. E. Lluch-Cota, and C. M. Niquen (2003), From anchovies to sardines and back: Multidecadal change in the Pacific ocean, Science, 299, 217-221.
Cloern, J., T. Schraga, C. Lopez, N. Knowles, R. Labiosa, and R. Dugdale (2005), Climate anomalies generate an exceptional dinoflagellate bloom in San Francisco Bay, Geophysical Research Letters, 32, L14608, doi:10.1029/2005GL023321.
Cochlan, W., J. Herndon, and R. Kudela (2008), Inorganic and organic nitrogen uptake by the toxigenic diatom Pseudo-nitzschia australis (Bacillariophyceae) Harmful Algae, 8, 111-118.
Curtiss, C., G. Langlois, L. Busse, F. Mazzillo, and M. Silver (2008), The emergence of Cochlodinium along the California Coast (USA), Harmful Algae, 7, 337-346.
Das, J., K. Rajan, S. Frolov, J. Ryan, F. Py, D. Caron, and G. Sukhatme, (2010), Towards Marine Bloom Trajectory Prediction for AUV Mission Planning, Submitted to IEEE International Conference on Robotics and Automation.
Dolan, J. R., and K. McKeon (2005), The reliability of grazing rate estimates from dilution experiments: Have we over-estimated rates of organic carbon consumption by microzooplankton?, Ocean Science, 1, 1-7.
Doucette, G.J., Mikulski, C.M., Jones, K.L., King, K.L., Greenfield, D.I., Marin III, R., Jensen, S., Roman, B., Elliott, C.T., Scholin, C.A. (2009), Remote, subsurface detection of the algal

A Regional Comparison of Upwelling and Coastal Land Use Patterns on the Development of HAB Hotspots Along the California Coast
28
toxin domoic acid onboard the Environmental Sample Processor: assay development and field trials. Harmful Algae. doi:10.1016/j.hal.2009.04.006.
Doyle, J.D., Yiang, Q., Chao, Y., Farrara, J., (2009), High-resolution real-time modeling of the marine atmospheric boundary layer in support of the AOSNII field campaign. Deep-Sea Research II.
Flynn, K. J. (2006), Castles built on sand: dysfunctionality in plankton models and the inadequacy of dialogue between biologists and modellers, J. Plankton Res, 27, 1205-1210.
Follows, M., S. Dutkiewicz, S. Grant, and S. Chisholm (2007), Emergent biogeography of microbial communities in a model ocean, Science, 315, 1843-1846, DOI: 10.1126/science.1138544
Fraga, S, S Gallager, and D Anderson. (1989) Chain-forming dinoflagellates: an adaptation to red tides. In Red Tides: Biology, Environmental Science and Toxicology, edited by T. Okaichi, D. Anderson and T. Nemoto. New York: Elsevier.
Frolov, S, J Ryan, J Bellingham, F Chavez, K Rajan, Z Kolber, S Ramp, C Scholin (2009), Optimal design of ocean observatories: designing a harmful algal bloom observatory in Monterey Bay. GEOHAB Open Science Meeting on Modeling HABs, June 2009, Galway, Ireland.
Gallegos, C. (1989), Microzooplankton grazing on phytoplankton in the Rhode River, Maryland: nonlinear feeding kinetics, Marine Ecology Progress Series, 57, 23-33.
GEOHAB (2003), Global Ecology and Oceanography of Harmful Algal Blooms, Implementation Plan, 36 pp, SCOR and IOC, Baltimore and Paris.
GEOHAB (2005), Global Ecology and Oceanography of Harmful Algal Blooms, GEOHAB Core Research Project: HABs in Upwelling Systems, 82 pp, SCOR and IOC, Paris and Baltimore.
Goeyens, L., N. Kindermans, M. Abu Yusuf, and M. Elskens (1998), A room temperature procedure for the manua determination of urea in seawater, Estuarine, Coastal and Shelf Science, 47, 415-418.
Goffredi, S.K., W. Jones, C. Scholin, R. Marin, S. Hallam, R.C. Vrijenhoek (2006), Molecular detection of marine larvae. Marine Biotechnology 8: 149-160.
Graham, W. M., and J. L. Largier (1997), Upwelling shadows as nearshore retention sites: the example of northern Monterey Bay, Cont Shelf Res, 17, 509-532.
Greenfield, D., R. Marin III, S. Jensen, E. Massion, B. Roman, J. Feldman, and S. CA (2006), Application of Environmental Sample Processor (ESP) methodology for quantifying Pseudo-nitzschia australis using ribosomal RNA-targeted probes in sandwich and fluorescent in situ hybridization formats, Limnol.Oceanogr. Meth., 4, 426-435.
HARRNESS (2005) Harmful Algal Research and Response: A National Environmental Science Strategy 2005–2015. Washington, DC: Ecological Society of America.
Haury L.R., J.A. McGowan and P.H. Wiebe. (1978) Patterns and processes in the time-space scales of plankton distributions. In: Spatial pattern in plankton communities, J.H. Steele, ed., 277-327,
Plenum Press, New York, NY.

A Regional Comparison of Upwelling and Coastal Land Use Patterns on the Development of HAB Hotspots Along the California Coast
29
Hayashi, K and GJ Smith (2005), Applications of rDNA ITS sequence analysis to assess inter- and intraspecific diversity in Pseudo-nitzschia communities of Monterey Bay, CA. 3rd Symposium on Harmful Algae in the U.S. Presentation.
Hayashi, K, N Welschmeyer, and GJ Smith (2007), Long-term, temporal variability in Pseudo-nitzschia population dynamics and domoic acid toxicity in Monterey Bay, CA. 4th Symposium on Harmful Algae in the U.S. Presentation.
Haywood, A.J., C. A. Scholin, R. Marin III, K. A. Steidinger, C. A. Heil and J. Ray. (2007) Molecular detection of the brevetoxin-producing dinoflagellate Karenia brevis (Dinophyceae) and closely related species using ribosomal RNA probes and a semi-automated sandwich hybridization assay. Journal of Phycology 43:1271–1286
Hickey, B., and N. Banas (2003), Oceanography of the U.S. Pacific Northwest coastal ocean and estuaries with application to coastal ecology, Estuaries, 26, 1010-1031.
Holmes, R. M., A. Aminot, R. Kerouel, B. A. Hooker, and B. J. Peterson (1999), A simple and precise method for measuring ammonium in marine and freshwater ecosystems, Can J Fisheries Aquat Sci, 56, 1801-1808.
Horner, R., D. Garrison, and F. Plumley (1997), Harmful algal bloom and red tide problems on the U.S. west coast, Limnol Oceanogr, 42, 1076-1088.
Howard, M., N. Ladizinsky, W. Cochlan, and R. Kudela (2007), Nitrogenous preference of toxigenic Pseudo-nitzschia australis (Bacillariophyceae) from field and laboratory experiments, Harmful Algae, 6, 206-217.
Howard, M., M. Silver, and R. Kudela (2008), Yessotoxin detected in mussel (Mytilus californicus) and phytoplankton samples from the US west coast, Harmful Algae, 7, 646-652, doi: 10.1016/j.hal.2008.01.003.
Jessup, DA, MA Miller, JP Ryan, HM Nevins, HA Kerkering, A Mekebri, DB Crane, TA Johnson, and RM Kudela (2009), Mass stranding of marine birds caused by a surfactant-producing red tide. PLOS-ONE, DOI:10.1371/journal.pone.0004550.
Jester, R., K. Lefebvre, G. Langlois, V. Vigilant, K. Baugh, and M. W. Silver (2009), A shift in the dominant toxin-producing algal species in central California alters phycotoxins in food webs, Harmful Algae, 8, 291-298, doi:10.1016/j.hal.2008.07.001.
Johnson, K. S., F. P. Chavez, V. A. Elrod, S. E. Fitzwater, J. T. Pennington, K. R. Buck, and P. M. Walz (2001), The annual cycle of iron and the biological response in central California coastal waters, Geophysical Research Letters, 28, 1247-1250.
Jones, B.H., Noble, M.A., Dickey, T.D. (2002) Hydrographic and particle distributions over the Palos Verdes Continental Shelf: spatial, seasonal and daily variability. Continental Shelf Research 22 (6-7), 945-965.
Jones, W.J., C. Preston, R. Marin III, C. Scholin, R. Vrijenhoek (2007), A Robotic Molecular Method for in situ Detection of Marine Invertebrate Larvae. Mol. Ecol. Res. 8: 540-550.
Kahru, M., Mitchell, B., (2008) Ocean Color Reveals Increased Blooms in Various Parts of the World. Eos Trans. AGU 89 (18).
Kahru, M., R. Kudela, M. Manzano-Sarabia, and B. Mitchell (2009), Trends in primary

A Regional Comparison of Upwelling and Coastal Land Use Patterns on the Development of HAB Hotspots Along the California Coast
30
production in the California Current detected with satellite data, Journal of Geophysical Research, 114, C02004, doi:02010.01029/02008JC004979.
Kudela, R. (2008), Silicon:Nitrogen Interactions in the Marine Environment, in Nitrogen in the Marine Environment, 2nd Edition, edited by D. Capone, et al., pp. 1589-1626, Elsevier, New York.
Kudela, R., N. Banas, J. Barth, E. Frame, D. Jay, J. Largier, E. Lessard, T. Peterson, and A. Vander Woude (2008a), New insights into the controls and mechanisms of plankton productivity in coastal upwelling waters of the northern California Current System, Oceanography, 21, 40-54.
Kudela, R., and F. Chavez (2004), The impact of coastal runoff on ocean color during an El Niño year in Central California, Deep-Sea Research II, 51, 1173-1185.
Kudela, RM, and WP Cochlan (2000), Nitrogen and carbon uptake kinetics and the influence of irradiance for a red tide bloom off southern California, Aquatic Microbial Ecology, 21, 31-47.
Kudela, RM, WP Cochlan, and R. C. Dugdale (1997), Carbon and nitrogen uptake response to light by phytoplankton during an upwelling event, J. Plankton Res, 19, 609-630.
Kudela, R., N. Garfield, and K. Bruland (2006), Bio-optical signatures and biogeochemistry from intense upwelling and relaxation in coastal California, Deep-Sea Research II, 53, 2999-3022.
Kudela, R., J. Lane, and W. Cochlan (2008b), The potential role of anthropogenically derived nitrogen in the growth of harmful algae in California, USA, Harmful Algae, 8, 103-110.
Kudela, R., G. Pitcher, T. Probyn, F. Figueiras, T. Moita, and V. Trainer (2005), Harmful algal blooms in coastal upwelling systems, Oceanography, 18, 184-197.
Kudela, R., J. Ryan, M. Blakely, J. Lane, and T. Peterson (2008c), Linking the physiology and ecology of Cochlodinium to better understand harmful algal bloom events: A comparative approach, Harmful Algae, 7, 278-292.
Kudela, RM, J Ryan, VL Trainer, G Pitcher, T Moita, and FG Figueiras, (2008d) Similarities between harmful algal bloom hotspots in Eastern Boundary Current Systems: the importance of retention to red tides. In Eastern Boundary Current Upwelling Systems Symposium, Las Palmas, Gran Canaria, Spain.
Kudela, RM, S Seeyave, and WP Cochlan (2010), The role of nutrients in regulation and promotion of harmful algal blooms in upwelling systems, Progress in Oceanography, Accepted.
Ladizinsky, NL. (2003), The influence of dissolved copper on the production of domoic acid by Pseudonitzschia species in Monterey Bay, California: Laboratory experiments and field observations. M.Sc., California State University, Monterey Bay.
Landry, M. R., and A. Calbert (2005), Reality checks on microbial food web interactions in dilution experiments: responses to the comments of Dolan and McKeon, Ocean Science, 1, 39-44.
Lane, J., P. Raimondi, and R. Kudela (2009), Development of a logistic regression model for the prediction of toxigenic Pseudo-nitzschia blooms in Monterey Bay, California, Marine Ecology Progress Series, 383, 37-51.

A Regional Comparison of Upwelling and Coastal Land Use Patterns on the Development of HAB Hotspots Along the California Coast
31
Lange, CB, FMH Reid, and M Vernet. (1994),Temporal distribution of the potentially toxic diatom Pseudonitzschia australis at a coastal site in Southern California. Marine Ecology Progress Series 104:309-312.
Langlois, G. (2001) Marine biotoxin monitoring in California, 1927–1999. In Harmful Algae Blooms on the North American West Coast, edited by R. RaLonde. Fairbanks, AK: University of Alaska Sea Grant College Program.
Li, Z, Y Chao, JC McWilliams, and K Ide (2008), A three-dimensional variational data assimilation scheme for the Regional Ocean Modeling System: Implementation and basic experiments. Journal of Geophysical Research 113, C05002.
Li, Z, Y Chao, JC McWilliams, and K Ide (2009), A Three-Dimensional Variational Data Assimilation Scheme for the Regional Ocean Modeling System. Journal of Atmospheric and Oceanic Technology, 25, 2074-2090.
Litaker, R., T. Stewart, B.-T. Eberhardt, J. Wekell, V. Trainer, R. Kudela, P. Miller, A. Roberts, C. Hertz, T. Johnson, G. Frankfurter, G. Smith, A. Schnetzer, J. Schumacker, J. Bastian, A. Odell, P. Gentien, D. Le Gal, D. Hardison, and P. Tester (2008), Rapid enzyme-linked immunosorbent assay for detection of the algal toxin domoic acid, Journal of Shellfish Research, 27, 1301-1310.
Loureiro, S., E. Garces, M. Fernandez-Tejedor, D. Vaqué, and J. Camp (2009a), Pseudo-nitzschia spp. (Bacillariophyceae) and dissolved organic matter (DOM) dynamics in the Ebro Delta (Alfacs Bay, NW Mediterranean Sea), Estuaries and Coasts, 83, 539-549.
Loureiro, S., C. Jauzein, E. Garces, Y. Collos, J. Camp, and D. Vaqué (2009b), The significance of organic nutrients in the nutrition of Pseudo-nitzschia delicatissima (Bacillariophyceae), J. Plankton Res, 31, 399-410.
Lyon, G.S., Petschauer, D., Stein, E.D., (2006) Effluent discharges to the Southern California Bight from large municipal wastewater treatment facilities in 2003 and 2004. Southern California Coatal Water Research Project - Annual Report 2006. Southern California Coatal Water Research Project, Costa Mesa, CA, pp. 1-15.
Lyon, G.S., Stein, E.D. (2008), Effluent discharges to the Southern California Bight from small municipal wastewater treatment facilities in 2005. Southern California Coatal Water Research Project - Annual Report 2008. Southern California Coatal Water Research Project, Costa Mesa, CA, pp. 1-14.
Lyon, G.S., Stein, E.D., (2009), How effective has the Clean Water Act been at reducing pollutant mass emissions to the Southern California Bight over the past 35 years? Environmental Monitoring and Assessment 154 (1-4), 413-426.
MacFadyen, A., B. M. Hickey, and W. P. Cochlan (2008), Influences of the Juan de Fuca Eddy on circulation, nutrients, and phytoplankton production in the northern California Current System, Journal of Geophysical Research, 113, C08008.
Mackas D.L. and C.M. Boyd. (1979) Spectral analysis of zooplankton spatial heterogeneity. Science 204: 62-64.
Maldonado, MT, MP Hughes, EL Rue, and ML Wells (2002), The effect of Fe and Cu on growth and domoic acid production by Pseudo-nitzschia multiseries. Limnol. Oceangr. 47:515-526.

A Regional Comparison of Upwelling and Coastal Land Use Patterns on the Development of HAB Hotspots Along the California Coast
32
Mayr, E. (1982), The growth of biological thought: Diversity, evolution, and inheritance, 974 pp., Harvard University Press, Cambridge, MA.
McGillicuddy Jr., D. J., D. M. Andeson, C. A. Stock, D. R. Lynch, and D. W. Townsend (2008), Modeling blooms of Alexandrium fundeyense in the Gulf of Maine, in Real-Time Costal Observing Systems for Marine Ecosystem Dynamics and Harmful Algal Blooms: Theory, Instrumentation, and Modelling, edited by M. Babin, et al., pp. 599-626, United Nations Educational, Scientific, and Cultual Organization, Paris.
McManus, M., R. Kudela, M. Silver, G. Steward, P. Donaghay, and J. Sullivan (2007), Cryptic blooms: Are thin layers the missing connection?, Estuaries and Coasts, 10.1007/s12237-007-9025-4.
Mikulski, C.M., Y.T. Park, K.L. Jones, C.K. Lee, W.A. Lim, C.A. Scholin, G.J. Doucette. (2008), Development and filed application of rRNA-targeted probes for the detection of Cochlodinium ploykrikoides in Korean coastal waters using whole cell and sandwich hybridization formats. Harmful Algae 7: 347–359.
Mengelt, C., and B. Prézelin (2004), Dark survival and subsequent light recovery for Pseudo-nitzschia multiseries paper presented at Harmful Algae 2002, Florida Fish and Wildlife Commission, Florida Institute of Oceanography, and Intergovernmental Oceanographic Commission of UNESCO, St Petersburg, FL.
Miller, P., and C. Scholin (1996), Identification of culture Pseudo-nitzschia (Bacillariophyceae) using species-specific LSU rRNA targeted fluorescent probes, J. Phycol, 32, 646-655.
Miller, P., and C. Scholin (1998), Identification and enumeration of cultured and wild Pseudo-nitzschia (Bacillariophyceae) using species-specific LSU rRNA-targeted fluorescent probes and filter-based whole cell hybridization, J. Phycol, 34, 371-382.
Miller, P., and C. Scholin (2000), n detection of Pseudo-nitzschia species using rRNA-targeted probes: sample fixation and stability, J. Phycol, 36, 238-250. Noble M.A., S.R. Ramp and K. Kinoshita (1992) Current Patterns Over the Shelf and Slope
Adjacent to the Gulf of the Farallones. Executive Summary. Prepared for the U.S. Environmental Protection Agency, Region IX, San Francisco, CA.
Olascoaga, M. J., F. J. Beron-Vera, L. E. Brand, and H. Koçak (2008), Tracing the early development of Harmful Algal Blooms on the West Florida Shelf with the aid of Lagrangian coherent structures, Journal of Geophysical Research, 113, c12014.
Olson, M. B., E. J. Lessard, W. P. Cochlan, and V. L. Trainer (2008), Intrinsic growth and microzooplankton grazing on toxigenic Pseudo-nitzschia spp. diatoms from the coastal northeast Pacific, Limnol Oceanogr, 53.
Paduan, J. D., and L. K. Rosenfeld (1996), Remotely sensed surface currents in Monterey Bay from shore-based HF radar (Coastal Ocean Dynamics Application Radar), Journal of Geophysical Research, 101, 20669-20686.
Pan, Y., D. Subba Rao, and K. Mann (1996a), Changes in domoic acid production and cellular chemical composition of the toxigenic diatom Pseudo-nitzschia multiseries under phosphate limitation, J. Phycol, 32, 371-381.
Pan, Y., D. Subba Rao, K. Mann, R. Brown, and R. Pocklington (1996b), Effects of silicate

A Regional Comparison of Upwelling and Coastal Land Use Patterns on the Development of HAB Hotspots Along the California Coast
33
limitation on production of domoic acid, a neurotoxin, by the diatom Pseudonitzschia pungens f. multiseries (Hasle). I. Batch culture studies, Marine Ecology Progress Series, 131, 225-233.
Pan, Y., D. Subba Rao, K. Mann, W. Li, and W. Harrison (1996c), Effects of silicate limitation on production of domoic acid, a neurotoxin, by the diatom Pseudonitzschia pungens f. multiseries (Hasle). II. Continuous culture studies, Marine Ecology Progress Series, 131, 235-243.
Paul, J, C Scholin, G van den Engh, MJ Perry, (2007), In situ Instrumentation. Oceanography 20: 58-66.
Petrenko, A.A., Jones, B.H., Dickey, T.D., LeHaitre, M., Moore, C., (1997) Effects of a sewage plume on the biology, optical characteristics, and particle size distributions of coastal waters. Journal of Geophysical Research-Oceans 102 (C11), 25061-25071.
Preston, C., et al. 2009. Near real-time, autonomous detection of marine bacterioplankton on a coastal mooring in Monterey Bay, California, using rRNA-targeted DNA probes. Environmental Microbiology 11: 1168-1180.
Price, DW, KW Kizer, and KH Hansgen (1991), California’s paralytic shellfish poisoning prevention program, 1927–1989. Journal of Shellfish Research 10:119-145.
Rajan, K., F. Py, C. McGann, J. Ryan, T. O'Reilly, T. Maughan and B. Roman (2009), Onboard Adaptive Control of AUVs using Automated Planning and Execution, International Symposium on Unmanned Untethered Submersible Technology (UUST) August 2009. Durham, NH.Rienecker, E., J. Ryan, M. Blum, C. Dietz, L. Coletti, R. Marin III, and W. P. Bissett (2008), Mapping phytoplankton in situ using a laser scattering sensor, Limnol.Oceanogr. Meth., 6, 153-161.
Rosenfeld, L. K., F. B. Schwing, N. Garfield, and D. E. Tracy (1994), Bifurcated flow from an upwelling center: a cold water source for Monterey Bay, Cont Shelf Res, 14, 931-954.
Rue, E., and K. Bruland (2001), Domoic acid binds iron and copper: a possible role for the toxin produced by the marine diatom Pseudo-nitzschia, Marine Chemistry, 76, 127-134.
Ryan, J., F. Chavez, and J. Bellingham (2005a), Physical-biological coupling in Monterey Bay, California: topographic influences on phytoplankton ecology, Marine Ecology Progress Series, 287, 23-32.
Ryan, J., H. Dierssen, R. Kudela, C. Scholin, K. Johnson, J. Sullivan, A. Fischer, E. Rienecker, P. McEnaney, and F. Chavez (2005b), Coastal ocean physics and red tides: An example from Monterey Bay, California, Oceanography, 18, 246-255.
Ryan, J., F. Chavez, J. Bellingham, E. Rienecker, H. Dierssen, R. Kudela, A. Vander Woude, R. Maffione, and A. Fischer (2002), Environmental processes in the Monterey Bay National Marine Sanctuary: Studies integrating AVIRIS and synoptic in situ remote sensing, paper presented at Proceedings of the 2004 JPL Airborne Earth Science Workshop. . JPL Publication 87-38., JPL, NASA, Jet Propulsion Laboratory, Pasadena, California, USA.
Ryan, J., A. Fischer, R. Kudela, J. Gower, S. King, R. Marin III, and F. Chavez (2009), Influences of upwelling and downwelling winds on red tide bloom dynamics in Monterey Bay, California, Cont Shelf Res, 29, 785-795.
Ryan, J., J. Gower, S. King, W. BIssett, A. Fischer, R. Kudela, Z. Kolber, F. Mazzillo, E. V.

A Regional Comparison of Upwelling and Coastal Land Use Patterns on the Development of HAB Hotspots Along the California Coast
34
Rienecker, and F. Chavez (2008), A coastal ocean extreme bloom incubator, Geophysical Research Letters, 35, L12602, doi:10.1029/2008GL034081.
Schnetzer, A., P. Miller, R. Schaffner, B. Stauffer, B. Jones, S. Weisberg, P. DiGiacomo, W. Berelson, and D. Caron (2007), Blooms of Pseudo-nitzschia and domoic acid in the San Pedro Channel and Los Angeles harbor areas of the Southern California Bight, 2003-2004, Harmful Algae, 6, 372-387.
Scholin, C. A., F. Gulland, G. J. Doucette, S. Benson, M. Busman, F. P. Chavez, J. Cordaro, R. DeLong, A. De Vogelaere, J. Harvey, M. Haulena, K. Lefebvre, T. Lipscomb, S. Loscutoff, L. J. Lownstine, R. Marin III, P. E. Miller, W. A. McLellan, P. D. R. Moeller, C. L. Powell, T. Rowles, P. Silvagni, M. Silver, T. Spraker, V. Trainer, and F. M. Van Dolah (2000), Mortality of sea lions along the central California coast linked to a toxic diatom bloom, Nature, 403, 80-84.
Scholin, C., Doucette, G., Jensen, S., Roman, B., Pargett, D., Marin III, R., Preston, C., Jones, W., Feldman, J., Everlove, C., Harris, A., Avarado, N., Massion, E., Birch, J., Greenfield, D., Wheeler, K., Vrijenhoek, R., Mikulski, C., Jones, K. (2009) Remote detection of marine microbes, small invertebrates, harmful algae and biotoxins using the Environmental Sample Processor (ESP). Oceanography 22: 158-167.
Scholin, C.A., E. Vrieling, L. Peperzak, L. Rhodes and P. Rublee. (2003), Detection of HAB species using lectin, antibody and DNA probes. In: G.M. Hallegraeff, D.M. Anderson A.D. Cembella (eds), Manual on Harmful Marine Microalgae, pp. 131-64. Paris, Intergovernmental Oceanographic Commission, UNESCO. (Vol. 11. Second Edition).
Scianni, C (2008), Oxygen-based validation of a new dilution protocol for determining microzooplankton grazer-dilution rates. M.S. Thesis, California State University Stanislaus, 90 pp.
Sekula-Wood, E., A. Schnetzer, C.R. Benitez-Nelson, C. Anderson, W.M. Berelson, M.A. Brzezinski, J.M. Burns, D.A. Caron, I. Cetinic, J.L. Ferry, E. Fitzpatrick, BH. Jones, P.E. Miller, S.L. Morton, R.A. Schaffner, D.A. Siegel, and R. Thunel. (2009) Rapid downward transport of the neurotoxin domoic acid in coastal waters. Nature Geosci. 2: 272-275.
Shchepetkin, A.F., McWilliams, J.C., (2005) The Regional Ocean Modeling System: A split-explicit, free-surface, topography following coordinates ocean model. Ocean Modelling 9, 347-404.
Smayda, T. (1997), Harmful Algal Blooms: Their ecophysiology and general relevance to phytoplankton blooms in the sea, Limnol Oceanogr, 42, 1137-1153.
Sommer, H, and KF Meyer (1937), Paralytic shellfish poisoning Arch. Path. 24:560-598.
Smith, R., Y. Chao, P. Li, B. Jones, D. Caron and G. Sukhatme (2009), Trajectory design for Autonomous Underwater Vehicles based on Ocean Model Predictions for Feature Tracking, 7th International Conference on Field and Service Robots, July 2009, Cambridge, Massachusetts.
Song, Y., Haidvogel, D.B., (1994), A semi-implicit ocean circulation model using a generalized topography-following coordinate system. Journal of Computational Physics 115 (1), 228-244

A Regional Comparison of Upwelling and Coastal Land Use Patterns on the Development of HAB Hotspots Along the California Coast
35
State of California, D.o.F., (2007) Population Projections for California and Its Counties 2000-2050. http://www.dof.ca.gov/research/demographic/reports/projections/p-1/.
Strub P.T., J.S. Allen, A. Huyer and R.L. Smith. (1987) Seasonal cycles of currents, temperatures, winds, and sea level over the northeast Pacific continental shelf: 35oN to 48oN. JGR 92(C2):
1507-1526.
Thomas, A.C., (2009) Interannual variability in chlorophyll concentrations in the Humboldt and California Current Systems. Progress in Oceanography In press.
Torrey, H. B. (1902), An unusual occurrence of Dinoflagellata on the California coast, American Naturalist, 36, 187-192.
Trainer, V. L., N. G. Adams, B. D. Bill, C. M. Stehr, and J. C. Wekell (2000), Domoic acid production near California coastal upwelling zones, June 1998, Limnol Oceanogr, 45, 1818-1833.
Trainer, V. L., B. M. Hickey, E. J. Lessard, W. P. Cochlan, C. G. Trick, M. L. Wells, A. MacFadyen, and S. K. Moore (2009), Variability of Pseudo-nitzschia and domoic acid in the Juan de Fuca eddy region and its adjacent shelves, Limnol Oceanogr, 54, 289-308.
Walsh, J. K. Jolliff, B. P. Darrow, J. M. Lenes, S. P. Milroy, D. A. Dieterle, K. L. Carder, F. R. Chen, G. A. Vargo, R. H. Weisberg, K. A. Fanning, F. E. Muller-Karger, K. A. Steidinger, C. A. Heil, C. R. Tomas, J. S. Prospero, T. N. Lee, G. J. Kirkpatrick, T. E. Whitledge, D. A. Stockwell, T. A. Villareal, A. E. Jochens, and P. S. Bontempi (2006), Red tides in the Gulf of Mexico: where, when and why?, Journal of Geophysical Research, 111, C11003.
Wang, X., Y. Chao, C. Dong, J. Farrara, Z. Li, J. C. McWilliams, J. D. Paduan, L. K. Rosenfeld (2009), Modeling tides in Monterey Bay, California. Deep-Sea Research II, 56, 219-231, doi:10.1016/j.dsr2.2008.08.012.
Wang, X., Christian, J.R., Murtugudde, R., Busalacchi, A.J., (2005) Ecosystem dynamics and export production in the central and eastern equatorial Pacific: A modeling study of impact of ENSO. Geophysical Research Letters 32, L2608.
Warrick, J.A., DiGiacomo, P.M., Weisberg, S.B., Nezlin, N.P., Mengel, M., Jones, B.H., Ohlmann, J.C., Washburn, L., Terrill, E.J., Farnsworth, K.L., (2007) River plume patterns and dynamics
within the Southern California Bight. Continental Shelf Research 27, 2427-2448.
Warrick, J.A., Mertes, L.A.K., Siegel, D.A., Mackenzie, C., (2004) Estimating suspended sediment concentrations in turbid coastal waters of the Santa Barbara Channel with SeaWiFS.
International Journal of Remote Sensing 25 (10), 1995-2002.
Wells, M. L. (1999), Manipulating iron availability in nearshore waters, Limnol Oceanogr, 44, 1002-1008.
Wells, M. L., C. G. Trick, W. P. Cochlan, M. P. Hughes, and V. L. Trainer (2005), Domoic acid: The synergy of iron, copper, and the toxicity of diatoms., Limnol Oceanogr, 50, 1908-1917.
Welschmeyer, N. A. (1994), Fluorometric analysis of Chlorophyll a in the presence of Chlorophyll b and pheopigments, Limnol Oceanogr, 39, 1985-1992.
Wetz, M. S., P. A. Wheeler, and R. M. Letelier (2004), Light-induced growth of phytoplankton collected during the winter from the benthic boundary layer off Oregon, USA, Marine Ecology Progress Series, 280, 95-104.

A Regional Comparison of Upwelling and Coastal Land Use Patterns on the Development of HAB Hotspots Along the California Coast
36
Worden, A., and B. Binder (2003), Application of dilution experiments for measuring growth and mortality rates among Prochlorococcus and Synechococcus populations in oligotrophic environments, Aquatic Microbial Ecology, 30, 159-174.
Work, TM, AM Beale, L Fritz, MA Quilliam, M Silver, K Buck, and JLC Wright (1993), Domoic acid intoxication of brown pelicans and cormorants in Santa Cruz, California. In Toxic phytoplankton blooms in the sea, edited by T. Smayda and Y. Shimizu. Amsterdam: Elsevier.