dynamics of hierarchy formation: the …aris.ss.uci.edu/~lin/22.pdf · behaviour, volume 80,...
TRANSCRIPT

Behaviour, volume 80, 218-240/ 1982
DYNAMICS OF HIERARCHY FORMATION: THE SEQUENTIAL DEVELOPMENT OF DOMINANCE
RELATIONSHIPS
by IVAN D. CHASE1
(Department of Sociology, State University of New York at Stony Brook, Stony Brook, N.Y. 11794, U.S.A.)
(With 3 Figures) (Acc 5-IV-I982)
A central unresolved problem in the study of dominance concerns dynamics: what are the behavioral processes leading to hierarchy formation? For example, an experimenter brings a group of previously unacquainted animals together and a short while later observes a stable dominance hierarchy based upon agonistic behavior. What are the patterns of interaction used by these newly assembled animals in "deciding" their dominance relationships? How did they go from not knowing one another to clearly recognizing members to which they were dominant or subordinate?
The goal of this paper is to investigate the dynamics of hierarchy formation and start answering questions such as these. This paper develops a general method for examining the theoretically possible sequences of relationship formation for animals "winning" dominance encounters, "losing" dominance encounters, and bystanders to the dominance encounters of others. Some of these sequences are congruent with the kinds
1 This study was supported by grants from the Harry Frank Guggenheim Foundation, the U.S. Public Health Service (Biomedical Research Support Grant 5 S07 RR 07067-13 to SUNY at Stony Brook), the University Awards Program, and the Research Development Fund of the State University of New York at Stony Brook. I thank Jack HAILMAN for his advice and encouragement during t h e first pan of this research and Jeffrey BAYI.IS, Shelly COHEN, Scott FELD, Mark GRANOVETTER, Erica JEN, Bob NORMAN, Gene WEINSTEIN, and Bill WIMSATT for providing criticisms of earlier papers and suggesting a variety of helpful ideas. I would like to thank Paul SIEGEL for help with my in i t ia l efforts in the observation of dominance behavior; Gordon STEPHENSON for aid with the SSR System; Phil REESE and Jo Ann COMITO for assistance with the Apple microcomputer system; Gary SIMON for statistical consultation; Erica JEN for aid in data analysis; and Shelly COHEN, Sara EDWARDS. and Nancy RAFFETTO for assistance in data collection. Part of this study was completed at the Department of Zoology, University of Wisconsin-Madison, and I thank the department for its support.

DYNAMICS OF HIERARCHY FORMATION 219
of hierarchy structures commonly found in many species while others are not. The method is illustrated by application to experiments on hierarchy formation in chickens. CHASE (1980) hypothesized typical strategies for winners, losers, and bystanders in dominance encounters and developed a method for the sequential analysis of successive, individual aggressive acts. This paper extends the method to the sequential development of dominance relationships and tests empirical support for the hypotheses. Part I of this paper introduces the method and uses it to analyze the dynamics of dominance relationship formation in groups of three chickens. Part II generalizes the method, uses it to analyze hierarchy formation in groups of four chickens, points out the implications for explaining typical hierarchy structures in chickens and other species, and reviews the available data in other species.
Part I Animals and methods. Animals.
I used 27 two-year old white Leghorn hens, 24 hens in the original design and three to replace animals that died or became ill during the course of the experiment. The hens were purchased at one week of age from a commercial hatchery, housed in a common cage until approximately four months old, and then kept in individual cages (74 x 77 x 43 cm). The hens had met in pairs for a previous dominance experiment, but had been separated for several months before this experiment. Hens separated for two or three weeks appear to forget a previous dominance relationship (SCHJELDERUP-EBBE, 1935; GUHL, 1975; MAIER, 1964 but cf. SMITH & HALE, 1959 for chickens with relationships modified through aversive s t imuli) . Experimental apparatus.
I tested the hens in a separate observation room using a neutral cage 1 m in diameter and 71 cm high. Partitions divided the cage in to three sectors of equal area and were used to keep the animals apart until the experiment began. The cage had one food and one-water dispenser and each was available ad libitum. Experimental procedure.
After approximately 10 minutes in the observation cage, I removed the partitions separating the chickens and began to record data. After four hours of observation, the chickens were returned to their home cages. The author and a research assistant took alternate one hour shifts of data collection; the pretest interobserver reliability was 89%. (formula: number of interobserver agreements/number of interobserver agreements + number of interobserver disagreements.) The observer sat in a low chair at chicken-eye level and recorded data with an SSR Keyboard. (See STEPHENSON el at., 1975 and STEPHENSON, 1979 for a description of the SSR Keyboard.) All occurrences of the following four aggressive contact behaviors were recorded: (1) Peck: one hen strikes or grabs another with its beak or pulls another's leathers; (2)Jump On: one chicken jumps into the air s t r i k ing another chicken with one or both of her feet; (3) Claw: one chicken scratches another with the claws of one foot while keeping the other foot on the substrate; (4) Nib-

220 IVAN D. CHASE ble: one chicken lightly brushes the feathers or wing identification badge of another with its beak (any beak contact to the head or neck, no matter how light in intensity, was coded as a Peck). The SSR Keyboard automatically provided the real time of each action. A combined total of 2801 aggressive acts were recorded in 96 hours of observation time; the average rate of aggression was 29.2 acts per hour or 116.7 acts per triad.
Results Dominance c r i t e r i a .
I used two criteria to determine the formation of dominance relationships. One animal was considered to dominate another if she: (1) delivered any combination of three strong aggressive contact actions (pecks, jump ons, and claws) to the other and (2) there was a 30 minute period following the third action during which the receiver of the actions did not attack the initiator. The involvement of either the initiator or the receiver with other chickens or the length of time necessary to complete three actions was not considered in these criteria. Once formed, a dominance relationship could be reversed if a subordinate fulfilled the criteria over a previous superior. Although somewhat arbitrary, these criteria have support as a valid indicator of stable dominance relationships; only two out of 52 dominance relationships init ially formed were later reversed. The average time required to fulfill the criteria in the 52 initial relationships was 82.6 minutes; only two relationships were formed less than 30 minutes before the end of an observation period. There was, then, a sufficient time period after the formation of most relationships for reversals to occur.
Sequences of dominance r e l a t i o n s h i p formation. There are four and only four possible configurations for the first two
dominance relationships formed in a triad. By convention, I will call the chicken becoming dominant in the first relationship A, the initial subordinate B, and the bystander C. After the initial dominance relationship: (1) the initial dominant can dominate the bystander (Double Dominance or DD), (2) the bystander can dominate the initial subordinate (Double Subordinance or DS), (3) the bystander can dominate the i n i t i a l dominant (Bystander Dominates In i t i a l Dominant or BDID), or (4) the i n i t i a l subordinate can dominate the bystander (Initial Subordinate Dominates Bystander or ISDB). These four possible sequences are shown in Fig. 1.
If the dominance relationships in a triad are formed randomly, i.e., if there is a .5 probability of either animal dominating in a pair, then each sequence has an equal .25 probability of occurrence. Although each pat-

DYNAMICS OF HIERARCHY FORMATION 221
tern is equally probable in theory, the Double Dominance sequence occurred in the great majority (17 out of 23 triads or 74%) of triads (dominance relationships were not formed in one triad and it was dropped from this analysis). Of the remaining three sequences, Double Subordinance occurred four times (17%) and BDID and ISDB were quite rare indeed occurring once each (4%). (χ2 = 30.4, d.f. =3, p<.001).
Double Dominance Double Subordinance
Bystander Dominates
Initial Dominant Initial Subordinate Dominates Bystander
Fig. 1. The four possible sequences in the formation of dominance relationships in triads. Relationships are numbered in order of formation.
Discussion Sequence usage.
In more intuitive terms these results indicate that the most common sequence is for an initial dominant to subsequently win against the bystander — winning appears to follow winning. The next most common result is for the bystander to gain dominance over the initial subordinate — losing sometimes follows losing. It is very rare indeed for an initial subordinate to dominate a bystander or for a bystander to dominate an in i t ia l dominant. Or to put it yet another way, initially dominant chickens usually go on to dominate bystanders, initially subordinate

222 IVAN D. CHASE
chickens usually are unsuccessful in subsequent dominance encounters, and if bystanders win dominance contests it is almost always against initial subordinates.
I m p l i c a t i o n for hierarchy formation. This distribution of sequences has a crucial implication for hierarchy structure
in groups larger than three. In order to show the implication, some background information is needed at this point. The dominance hierarchies found in small groups of chickens, having fewer than about ten members, are frequently linear or near linear (for chickens see Guhl, 1958, 1968, 1975 and for jungle fowl see BANKS, 1956). By definition, the dominance relationships in all possible triads making up a linear hierarchy are transitive. In a triad with transitive dominance relationships, if A dominates B and B dominates C, then A also dominates C (Figure 2).
Fig. 2. The configuration of relationships in transitive and intransitive relationships
triads. If a hierarchy structure is not linear, then it necessarily contains at least one intransitive triad, and the more intransitive triads, the further from linearity. In an intransitive triad, if A dominates B and B dominates C, then C dominates, rather than is dominated by, A (Fig. 2). In the dominance literature, intransitive triads are sometimes called "triangles" or "pecking triangles." The hierarchy represented in Table 1 has one intransitive triad involving B, C, and D; hierarchies having more intransitive triads and further from linearity are also possible.
The four sequences of dominance relationships formation have differing implications for the formation of transitive and intransitive triads. DD and DS sequences guarantee triads with transitive relationships, while BDID and ISDB sequences can produce either transitive or intransitive triads. For example, when the third relationship is formed, the DD sequence results in either A dominates B, B dominates C, and A dominates C, or A dominates C, C dominates B, and A dominates B,

DYNAMICS OF HIERARCHY FORMATION 223
both transitive triads. In the Bystander Dominates Initial Dominant sequence, if B later dominates C, there is an intransitive triad, or conversely, if C later dominates B, there is a transitive triad.
The results from experiment 1 indicate that 91% of the triads used the two sequences guaranteeing transitivity. These results suggest that linear and near linear hierarchies are produced by usage of the same two sequences in the component triads of larger groups of chickens. And, in a like manner, these results suggest that hierarchies far from linearity are avoided because BDID and ISDB sequences are rarely used. I designed the experiment described below in order to test this possibility.
TABLE 1 Dominance matrix for one possible example of a non-linear hierarchy
Dominant Number Animal Dominated animal dominated
A B C D E A - 1 0 1 1 3 B 0 - 1 1 1 3 C 1 0 - 1 1 3 D 0 0 0 - 1 1 E 0 0 0 0 - 0
Part 2 In this experiment I assembled tetrads or groups of four chickens. Although these
groups only have one more member than the groups of three used in the previous experiment, the combinatorial complexities in tetrads are considerably greater. For example, while there is of course only one triad in a group of three, there are four possible component triads making up a group of four. Groups of size four are, then, the first crucial test of the hypothesis suggested by the results of the first experiment. Animals and methods. Animals.
I used 21 of the hens taking part in Experiment 1. After a short period of group living the hens were housed individually in 76 x 51 x 61 cm cages between the two experiments. Experimental apparatus.
I tested the hens in a separate observation room in a 1 52×102×81 cm cage. Removable partitions separated the cage into four sections of equal area and were used to keep the chickens apart when data were not being collected. Food and water were

224 IVAN D. CHASE available ad libitum in each section of the cage. The observation cage occupied the far end of the room and a blind separated the cage from the rest of the room. The observer sat behind the blind and recorded data through a small view hole. The light-dark schedule in the observation room was identical to that where the chickens were housed. Experimental procedure.
To prepare for an observation session, I took four chickens from their own cages and placed them in the observation cage (with partitions in place) on the afternoon before the day of data collection. The hens had approximately two hours to adjust to the cage before lights out at 1800. Data collection began the next day approximately three hours after lights on at 0600. The partitions and all but one each of the food and water containers were removed under low intensity light (so that the chickens would not have sufficient light for interaction). As soon as the observer was in place, the lighting was switched back to normal and data recording began. The first observer recorded data for one and a half hours, before being relieved by the second observer. Each observer took two hour and a half shifts a day for a total of six continuous hours of observation a day (pretest interobserver reliability = 87%). At the end of observation, the observer replaced the partitions and food and water containers and the chickens remained in the observation cage overnight. The chickens were again observed the next day for six hours following the procedure of the first day. Experimental design.
A balanced incomplete block design was used to assemble the chickens (HALL, 1967). This type of design provides a method for assembling smaller groups from a larger pool such that no two individuals meet more than once, all individuals are in an equal number of groups, and all individuals in a group have previously been in an equal number of experimental groups. A total of 14 groups took part in the experiment, and each chicken had at least three days rest between tests. Data collection.
An Apple II microcomputer system was used for data collection. All occurrences of the same four contact behaviors used in Experiment 1 were recorded in the tetrads with times of occurrence automatically provided by an on-board clock. A combined total of 7,402 aggressive acts were recorded in 168 total hours of observation time for a rate of 44.1 acts per hour or 528.7 per tetrad.
Results Structure of hierarchies.
I used the two dominance criteria described above to determine the resultant hierarchy structures in the 14 tetrads. This examination revealed an overwhelming tendency toward linear hierarchies in all groups. In order to determine if a hierarchy is linear or not, all of the possible pairwise dominance relationships in a group must be settled. In tetrads there are six possible pairwise dominance relationships. At the end of the first day of observation, all possible pairwise relationships had been decided in five groups, eight groups were missing one relationship, and one group was missing two relationships. The five groups with all six

DYNAMICS OF HIERARCHY FORMATION 225
relationships each had a linear hierarchy. In the nine groups with missing relationships, six were guaranteed to have linear hierarchies regardless of the direction of the missing relationship, and the remaining three groups could have had either linear or non-linear hierarchies depending upon the direction of the missing relationship(s). For these latter groups, an intransitive triad was possible if the missing relationship formed in one direction and a transitive triad if it formed in the other direction.
On Day 2 of data collection, five of the six groups guaranteed to have linear hierarchies completed their missing relationship, and, of course, all formed linear hierarchies. One group did not complete its missing relationship and continued to have a hierarchy guaranteed to be linear. Also on Day 2 the three groups which could have had either linear or non-linear hierarchies each resolved their missing relationships, and each formed a linear hierarchy. In sum, five tetrads had linear hierarchies on the first day of observation, and nine tetrads had missing relationships and could not be characterized. Of these nine groups all but one developed a linear hierarchy by the second day of observation; the one remaining group had a hierarchy guaranteed to be linear regardless of the direction of the missing relationship. This strong tendency toward linear hierarchies corroborates the findings of other studies in chickens mentioned above (χ2 = 23.3, d.f. = 1, p.<.001). Sequence usage. Determining sequence usage in the component triads of groups larger than three requires a small modification of technique. This modification entails breaking a group down into all possible component triads and noting the sequence patterns of the first two dominance relationships formed in each triad. Fig. 3 illustrates this procedure. The figure shows the order of relationship formation in two hypothetical tetrads and the resulting sequence patterns in each component triad. In Tetrad 1 there are two DD and two DS sequences. All four sequences ensure transitive triads, and the resulting group hierarchy is necessarily linear. In Tetrad 2 there are three BDID sequences and one ISDB pattern. None of these four sequences ensure transitive triads, and the resulting group hierarchy is, in fact, non-linear with triads B, C, D and A, C, D being intransitive. At least two dominance relationships were formed during the first day of observation in 55 out of 56 possible component triads in the 14 groups. As in the isolated triads, the most common pattern was Double Dominance (33 or 60%) with Double Subordinance a somewhat distant second (15 or 21%). BDID (3 or 6%) and ISDB (4 or 7%) were again
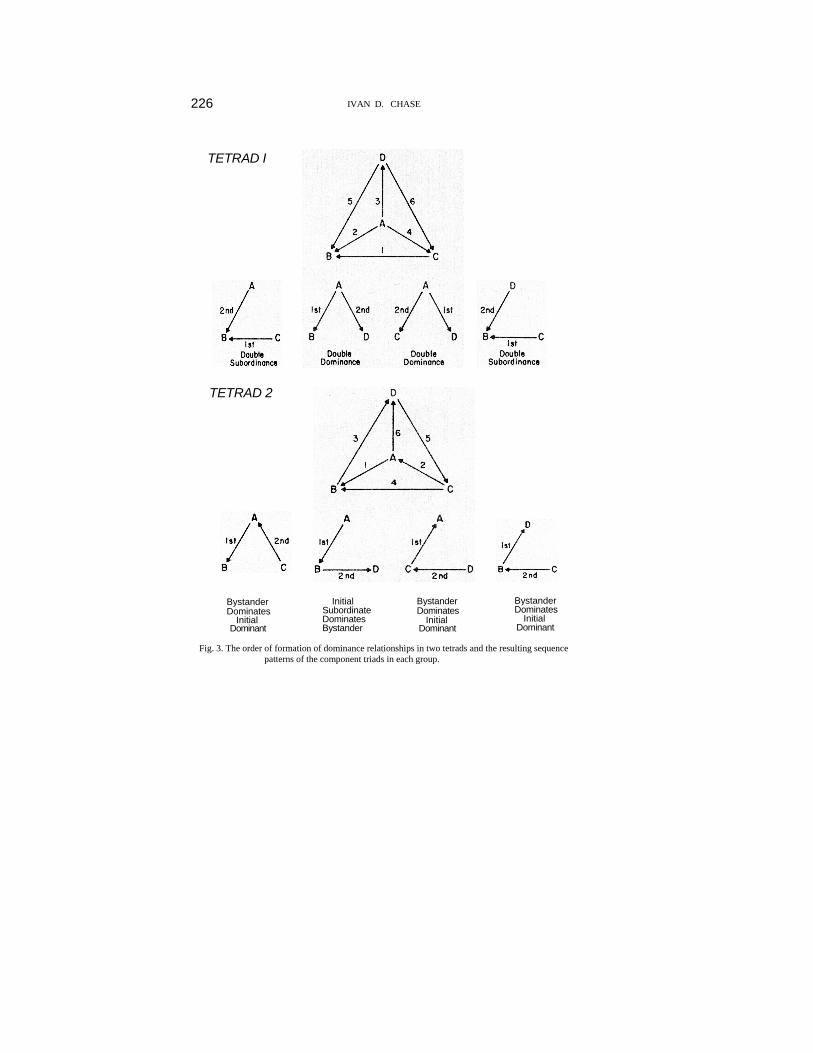
226 IVAN D. CHASE
TETRAD I
TETRAD 2
Bystander Dominates
Initial Dominant
Initial Subordinate Dominates Bystander
Bystander Dominates
Initial Dominant
Bystander Dominates
Initial Dominant
Fig. 3. The order of formation of dominance relationships in two tetrads and the resulting sequence patterns of the component triads in each group.
DYNAMICS OF HIERARCHY FORMATION 227
relatively rare (χ2 = 42.4, d.f. = 3, p<.001).1) Together the two sequences guaranteeing transitivity accounted for 87% of all sequences. I also examined the data record for Day 2 in order to determine the order in which dominance relationships previously established on Day 1 were re-enacted and integrated with those newly forming on Day 2. The results mirrored those of Day 1 (Table 2): DD and DS were again most frequent and accounted for 86% of all sequence usage (χ2 = 30.4, d.f. =3, p<.001).
TABLE 2 Distribution of sequences of dominance relationship formation in
component triads of tetrads by day
Dominance sequences Day of Double Double Bystander dominates Initial subordinate
observation dominance subordinance initial dominant dominates bystander Day 1 60.0%* 27.3% 5.5% 7.3% Day 2 48.2%** 37.5% 3.6% 10.7%
* N-55, ** N = 56.
Real versus "constructed" triads. The results presented so far demonstrate that the involvement of the bystander with
the two other chickens in a triad is not random, i.e., the sequences involving the bystander which guarantee transitive triads are much more common than those that do not. This finding raises an important question: would the structure of dominance relationships in a triad be the same if the bystander was not present when the first relationship was formed? In other words, what would the structure of dominance relationships look life if animals making up a triad met only as pairs?
In order to test whether the structure of dominance relationships in triads depended upon the animals all being together at the same time, I compared real versus "constructed" triads. The structure of both kinds of triads was determined by the data collected in Experiment 2. A real triad is formed by hens that actually met as part of one of the 14 tetrads; a
1) Although it can be shown that the sequence patterns in any two component triads in a tetrad are statistically independent, the set of all four component triads is not always so. However, the fact that the triads are independent pairwise allows obtaining the first two moments for the normal approximation to the multinomial, and consequently the chi-squared approximation still holds.

228 IVAN D. CHASE
real triad is then simply a component triad of one of the experimental tetrads. In contrast, a constructed triad is composed of hens that have never been assembled all together in one tetrad at the same time. Instead, the members of a constructed triad have met each other only as pairs within different tetrads. An example will help to make the distinction between real and constructed triads clear. Chickens A, B, C, and D meet in a tetrad. One of the component triads in this group is A, B, and C; this is a real triad. Other tetrads in the experiment are (A, E, F, and G) and (B, E, H, and I). Chickens A, B, and E meet only as pairs within tetrads and they form a constructed triad.
A structural comparison of the two kinds of triads indicates that all real triads were transitive, but that some constructed triads were intransitive. Out of 56 possible real triads 54 completed all three dominance relationships by Day 2 and all were transitive (the remaining two triads were guaranteed to be transitive when the third relationship was formed). A total of 63 constructed triads could be assembled; 54 (86%) were transitive and nine (14%) were intransitive. If the direction of dominance relationships is determined randomly, then there is a .75 probability of a transitive triad and .25 probability of an intransitive triad in independent triads. Therefore, in 63 triads about 47 would be expected to be transitive by chance alone, and about 16 would be expected to be intransitive. Although nine intransitive triads are certainly fewer than expected by chance alone, it is a relatively large number compared to the random expectation of 16. These results suggest that the structure of dominance relationships in triads is often influenced by the actual presence of the bystander. When the bystander is actually present, transitive triads appear virtually certain, but when the bystander is not present, there is a tendency toward intransitive triads. Constructed triads, of course, prohibit the use of the Double Dominance and Double Subordinance sequences, and without these sequences intransitive triads appear to be more frequent. Discussion 1. Bystanders and dominance r e l a t i o n s h i p formation.
In tetrads, as in isolated triads, sequences of dominance relationships involving bystanders are non-random. The most common pattern is for an ini t ial dominant in a component t r iad to subsequently dominate the bystander (Double Dominance), and the next most common pattern is for the bystander to subsequently dominate the ini t ial subordinate (Double Subordinancc). Together these two patterns compose the great ma-

DYNAMICS OF HIERARCHY FORMATION 229
jority of all sequences. Only infrequently does the chicken losing one dominance encounter become dominant over the bystander (ISDB) or does the bystander become dominant over the initial dominant (BDID).
Expressed in another way, these results indicate that "winners" of dominance encounters usually defeat bystanders rather than vice versa — winning begets winning. When a bystander does win it is usually against the "loser" of a dominance encounter — losing frequently begets losing if winning does not beget winning first. Previous work by MCBRIDE (1958) has indicated the tendency of winners to keep winning and losers to keep losing in successive pairwise contests in chickens. But when animals are together simultaneously in groups of three or greater the bystander is present vis-a-vis both the winner and the loser, and both tendencies can operate at the same time. Thus, the presence of the bystander, plus the tendency toward continued winning and losing, can help explain why the great majority of sequences are those guaranteeing transitivity. Additional support for this interpretation is given by the comparison of real and constructed triads. In real triads, where bystanders were present, all groups had transitive dominance relationships, but in the constructed triads, where bystanders were not present, there were intransitive triads.
Three differences between the tetrad results and those from the isolated triads are worthy of note. First, although the DD sequence composed the majority of sequences in both experiments, it was not as common in tetrads as in isolated triads (74% for isolated triads versus 60% for Day 1 tetrads). Second, the DS sequence was considerably more common in tetrads than isolated triads (27% for Day 1 tetrads versus 17% for isolated triads). Third, although the two sequences which do not guarantee transitivity were rare in both isolated triads and tetrads, they were somewhat more common in tetrads (13% for Day 1 tetrads versus 8% for isolated triads). These differences would appear to result from some combination of differences in group size, length of observation, and cage size. Differential sequence frequency, as noted, can imply differences in hierarchy structure, and a next step in research is to investigate if sequence occurrence continues to change as group size grows larger (see below).
2. Do dominance sequences e x p l a i n hierarchy structure in l a r g e r groups of chickens?
The results reported here support what might be termed a "jigsaw puzzle" model of hierarchy formation where the "pieces" are interaction

230 IVAN D. CHASE
patterns in component triads of larger groups. This model views hierarchy formation as a dynamic process in which sequences of dominance relationship formation is smaller groups concatenate to form hierarchies in larger groups. It argues that the structural form of a hierarchy can be explained by regularities or "building blocks" of interaction involving two individuals and a bystander. Two caveats should be noted, however, in the application of the jigsaw puzzle model. First, the model does not imply that chickens consciously strive to produce transitive dominance relationships. Instead, it points out that transitivity and intransitivily necessarily arise out of particular behavioral sequences during hierarchy formation. Second, the model is somewhat more an analytical conve-nience than a true "chicken's eye-view" of dominance interactions. The model does not imply that chickens think in terms of completing triadic relationships. However, when the actions of winners, losers, and bystanders are viewed through the triadic framework, the resultant structure of hierarchies can be analysed and explained.
An important next step in research is to investigate the dominance sequences in groups of chickens larger than four. The question here of course is: do Double Dominance and Double Subordinance patterns form the great majority of all sequences and thus explain the linear and near linear hierarchies which are often reported in chickens? There is some evidence to indicate that as chicken groups become larger, exceed, say, about ten members, hierarchies begin to depart further from linearity (see SCHJELDERUP-EBBE, 1975 and GUHL, 1975 for chickens and BANKS, 1956 for jungle fowl). So another important parallel concern for future research would be how sequence frequency changes, if it does, with group size. 3. Sequential patterns versus i n d i v i d u a l c h a r a c t e r i s t i c s as explanations of hierarchy structure.
The view presented here that dynamic patterns explain the structural form of hierarchies contrasts, at least in part, with earlier interpretations. Traditionally, an individual's rank in a hierarchy has often been explained by its "individual" characteristics — qualities that could be measured before the individual joined a group forming a hierarchy. In this view hierarchy structure is seen, in effect, as a manifestation of the differences in individual characteristics. For example, the individual ranking highest on the characteristics associated with dominance (e.g., aggressiveness, fighting ability, size, testosterone level, past dominance success, etc.) would be expected to occupy the top position in the hierar-

DYNAMICS OF HIERARCHY FORMATION 231
chy, the individual ranking next highest on the characteristics to occupy the next to the top position, and so on down to the lowest ranking individual who would occupy the lowest position in the hierarchy.
There is indeed much evidence which indicates that the characteristics of individuals help predict their positions in hierarchies. Predictive characteristics include size, hormone levels, rank in previous hierarchies, weight, seniority, relative maturity, and aggressiveness among a variety of other factors (see ALLEE et al., 1949; BROWN, 1975; COLLIAS, 1943; SCHEIN, 1975; and WILSON, 1975 for general reviews and SAVIN-WILLIAMS, 1977, 1979, 1980 for special attention to hierarchy rank in human children).
The distinction between process or dynamic and individual characteristic explanations of hierarchy structure is an important one for the discussion here. It provides a framework for assessing the results presented here, and it emphasizes the contrast between these two fundamentally different ways of accounting for hierarchy structures.
In an attempt to overlook the differences between the explanations, it might be argued that the sequence patterns reported here are merely reflections of differences in individual dominance ability. For example, the DD sequence could be explained as follows: A, the chicken highest in dominance ability, first has an encounter with B, A wins, then A goes on to have an encounter with C, and A wins again. The DS sequence could be explained similarly. Thus, the individual difference approach would explain the great proportion of the two sequences guaranteeing transitivity, and the dynamic patterns reported here would simply be the behavioral mechanisms through which individual differences produced linear and near linear hierarchies.
If it were true that the patterns reported here were simply the manifestations of individual differences, then it still would be of theoretical importance to understand the proximate behavioral mechanisms through which individual differences were translated into hierarchy position. However, a variety of results argue against the in-dividual characteristic approach. First, the individual characteristic approach cannot easily explain why the two sequences guaranteeing transi t ivi ty are much more frequent than the two sequences leading to either transitive or intransitive relationships. Reasonable scenarios using individual differences can account for DD and DS sequences, and just as reasonable scenarios can be given to account for the BDID and ISDB sequences. For example, to get ISDB pattern: A, the chicken ranking highest on dominance ability, first dominates B, the chicken ranking se-

232 IVAN D. CHASE
cond on dominance ability, and then B dominates C, the chicken lowest in ability. Therefore, although an individual characteristic approach can "explain" DD and DS, it also "explains" the other two patterns equally well and does not account for the differential usage of the four sequence patterns. A more adequate theory must, I would argue, take into account the effects of previous encounters on subsequent ones.
Second, there are additional results, both analytical and experimental that argue against the individual difference interpretation. The analytical results are based on earlier work by LANADAU (1951) and myself (CHASE, 1974). In that work individual characteristic explanations of hierarchy structure appearing in the literature were expressed in their basic core form, and that form was converted into a simple mathematical model, a canonical expression that captured the explanation. Two mathematical models proved to encompass most of the individual characteristic explanations: a correlational and a pairwise interaction model. The correlational model includes, for example, explanations that individuals' positions in hierarchies are determined by their physical attributes, their genetic endowment, their hormonal state, their past social performance, their aggressiveness, or any composite of these and similar factors. The pairwise interaction model assumes that each member of a group has an independent one-on-one contest with each other member, that the winner of a contest dominates the loser in the final group hierarchy, and that each individual has a certain probability of winning in each contest (also see MCBRIDE, 1964 for an incisive critique of the pairwise contest model).
Stringent mathematical conditions are required by both models in order to predict the empirically common linear and near linear hierarchies. These results hold regardless of the species and regardless of the factors assumed to correlate with hierarchy position or explain success in pairwise interactions. Specifically, the correlational model required correlation coefficients higher than .9, and the pairwise interaction model required one individual to have a .95 probability of beating everyone, a second individual to have a .95 probability of beating everyone but the first individual, and so on to the last individual having only a .05 probability of defeating any group member (CHASE, 1974). Analysis of the available data indicated that these stringent conditions were not met (CHASE, 1974). The conclusion of this research was that although correlational and pairwise encounter approaches give much useful information about dominance relationships, the amount of information that they give is not sufficient to account for many empirically common hierarchy structures. Although individuals in groups certainly differ, these differences

DYNAMICS OF HIERARCHY FORMATION 233
do not appear strong enough to explain the structural form of common hierarchies. The experimental results arguing against the primacy of individual characteristic
approaches contain three sets of findings. The first set concerns the results for real and constructed triads presented here. If "dominance ability" were a stable individual characteristic, lasting at least a week or two and not affected by group context, then constructed triads would necessarily always be transitive. The fact that intransitive constructed triads were found indicates that at least in some cases chickens do not have a stable, individual dominance ability that transfers across social contexts.
Two innovative experiments by BERNSTEIN & GORDON (1980) provide the second set of results. In the first experiment they formed two groups of male rhesus monkeys, let each group establish a dominance hierarchy, and then serially added the members of group 1 to group 2 at weekly intervals in reverse order of dominance in their home group (group 1). If dominance relationships were determined primarily by individual characteristics per se, two results would have been expected after the groups were merged: (1) the relative ranks of individuals in group 1 would be about the same before and after merger with group 2 and (2) some of the higher ranking members of group 1 would probably dominate a considerable number of the lower ranking members group 2 (unless one argued that, by chance, all the members of group 2 ranked higher on "dominance ability" than all the members of group 1). Their actual results were contrary to both these expectations. First, there was no correlation (Spearman coefficient = -.05) between the original hierarchical ranks of group 1 males and their relative ranks within the merged hierarchy (BERNSTEIN & GORDON, 1980: 1036). Second, all the members of group 1 ranked below five out of six of the group 2 members, and only two out of 11 members of group 1 ranked above the previously lowest ranked member of group 2.
In the i r second experiment, they established two separate groups each having four adult males plus females and juveniles. After the adult males formed a hierarchy in their respective groups, the alpha and beta animals of each group were introduced to the other's cage for short periods. If individual attributes were the primary determinant of hierarchy rank, the relative rank of the home and introduced alpha and beta males should be the same regardless of whose cage in which an introduction took place. Again their results were contrary to those expected with an individual characteristic explanation: the relative ranks of the two pairs of alphas

234 IVAN D. CHASE
and betas depended upon the cage in which a visit took place. The alpha and beta of group 1 were at the bottom of the resident hierarchy when they visited in group 2's cage and vice versa.
But what about cases where seniority is not a factor, when, for example, a group of animals comes together simultaneously to form a hierarchy? The third set of findings concerns cases of this sort. GUHL (1975) reports a series of experiments in which groups of chickens were assembled to form hierarchies, separated for a short period of time, and then reassembled to form second hierarchies. If individual characteristics were the main determinants of rank in hierarchies, and if these characteristics were relatively stable over short periods of time, then the rank of a chicken in the first hierarchy would be highly correlated with its rank in the second hierarchy. GUHL (1975; 161) only reports the actual correlation coefficient for one of these experiments, and it is low (.30) and not statistically significant. He mentions three other replications but does not report actual coefficients; however, he does say that the coefficients were not significant in two out of three cases.
In a related experiment KING (1965) used initial paired encounters to determine the dominance-submission relationships within three flocks of ten chickens each. In the initial paired encounters technique each chicken in a group meets each other animal in an independent, pairwise contest in a neutral arena. After the paired encounters were completed, the chickens were housed separately for three weeks and then assembled in a common cage with their previous opponents. When the chickens had been in their common cages for three weeks, the hierarchies in each cage were determined. In two out of three flocks KING (1965) found a low coefficient of repeatability between the number of fellow group members a chicken pecked in the initial paired encounters and the final hierarchy. The coefficient was relatively high in the third flock, but KING (1965) indicates that the hierarchy in this flock was unstable. In two out of three flocks, the "dominance ability" of chickens as measured by their paired encounters showed almost no correlation with their final position in the group hierarchy. WOOD-GUSH (1971) similarly compared the aggressiveness of chickens as measured by fights against a standard panel of other chickens with their places in group hierarchies. He found no correlation between aggression score and number of flock mates dominated.
In sum, although differences in individual characteristics can be partially predictive of differences in dominance ranks, there is a growing body of analytical and empirical work which indicates the limitations of individual characteristic explanations. Individual characteristics appear

DYNAMICS OF HIERARCHY FORMATION 235
not to be the primary determinants of dominance relations, and it is difficult if not impossible to explain the structural form of many empirically common hierarchies by these differences.
4. Can the jigsaw puzzle model explain hierarchy formation in o t h e r spec ies? Linear and near linear hierarchies are reported in small groups for a broad variety of
species including wasps and bumble bees (WILSON, 1971), chaffinches (MARLER, 1955), red cross bills (TORDOFF, 1954), coyotes (BEKOFF, 1976), cows (SCHEIN & FOHRMAN, 1955), ponies (TYLER, 1972), pigs (RASMUSSEN et al, 1962), rhesus monkeys (SALE, 1967), baboons (HAUSFATER, 1975), vervets (STRUHSAKER, 1967) and human children and adolescents (MISSAKIAN, 1976; SAVIN-WILLIAMS, 1977, 1979, 1980). However, some species such as wolves apparently often form hierarchies quite far from linearity (LOCKWOOD, 1979). The jigsaw puzzle approach can easily be applied across species, and there is no reason to believe, a priori, that chickens are unique in their usage of Double Dominance and Double Subordinance sequences. A reasonable hypothesis is that the linear and near linear hierarchies found in many species are also produced by a preponderance of DD and DS sequences in the component triads of larger groups. This potential for an approach to hierarchy formation with cross-species validity gives particular significance to the data presented here.
In the first application of the jigsaw puzzle model to another species, Barchas and Mendoza (in press) examined sequences of dominance relationship formation in same-sex triads for male and female rhesus monkeys. Twelve out of thirteen triads formed at least two dominance relationships during observation, and all twelve groups used either DD or DD sequences. In the rhesus triads, however, DS was more common than DD.
A set of studies has also found cross-species uniformity at a finer scale of hierarchy formation dynamics: sequences of successive aggressive acts. Transitivity and intransitivity are general properties of relationships, and sequences of successive aggressive acts can also be analyzed for their implication for transitivity and intransitivity. This analysis is developed in CHASE (1980). As in dominance relationships there are four possible sequences of aggressive acts involving first two original combatants and then a bystander, and two sequences ensure transitive attack relationships and two give rise to either transitive or intransitive attack relationships. Using this method of analysis, CHASE (1980) found that the great

236 IVAN D. CHASE
majority (87%) of attack sequences in the experiment with isolated triads described here were of the two kinds ensuring transitivity. Additional work in preparation indicates that the sequences ensuring transitivity compose the majority of all attack sequences in the experiment with chicken tetrads.
CHASE (1980) also reanalyzed data on attack sequences in Java-monkeys presented in DE WAAL, et al. (1976). In these data the two attack sequences ensuring transitivity accounted for 62% of all sequence occurrences. These data were gathered in a large, mixed-sex group with a relatively stable dominance hierarchy rather than the small, all female groups of the chicken experiments, and perhaps the different experimental conditions as well as the species difference accounts for the lowered proportion of the attack sequences ensuring transitivity in the monkeys. Further cross-species results are found in MOUSSAS (1980); she found that 71% of the attack sequences in a group of ten two to four year old children were of the types ensuring transitivity.
Finally, the results reported here are complementary to several innovative studies of the development of social relationships in various non-human primate species (KUMMER, 1975; MASON, 1978; SEYFARTH et al., 1978; STAMMBACK, 1978; VAITL, 1978). These studies also find that interactions between the members of one pair have important implications for the interactions between other pairs and thus for explaining the overall form of group social relationships. These studies similarly investigate the formation of typical patterns of social relationships in groups, and several find that regularities or "building blocks" of interaction in smaller groups, like the jigsaw puzzle model described here, help to explain overall group patterns. Relationships of concern in these studies include dominance, grooming, and cross-sex alliances.
An exciting and important next step is to carry forward the investigation of behavioral processes used in hierarchy formation in a variety of species. Such a program should reveal both similarities and differences in species typical patterns and will help us to construct a general approach to this important behavioral phenomenon.
Summary A method for investigating the behavioral sequences used in forming dominance hierarchies is
presented. There are four possible sequences for the formation of the first two dominance relationships in groups of three individuals (triads). Designating the winner in the first relationship formed as the initial dominant, the loser as the initial subordinate, and the animal not involved as the bystander, the four possihlc sequences are: (1) the initial dominant goes on to dominate the bystander (Double Dominance), (2) the bystander

DYNAMICS OF HIERARCHY FORMATION 237
later dominates the ini t ia l subordinate (Double Subordinate), (3) the bystander later dominates the ini t ia l dominant (Bystander Dominates Ini t ia l Dominant), and (4) the initial subordinate later dominates the bystander (Initial Subordinate Dominates Bystander). Although each sequence has an equal probability of occurrence of dominance relationships are formed randomly, two of the sequences have different implications for the formation of the empirically common linear and near linear hierarchies than the other two. Linear and near linear hierarchies are, by definition, composed of all or nearly all triads with transitive dominance relationships (A dominates B, B dominates C, and A dominates C), while hierarchies far from linearity have many triads with intransitive relationships (A dominates B, B dominates C, and C dominates A). Double Dominance and Double Subordinancc sequences guarantee the formation of triads with transitive dominance relationships, but the other two sequences can give triads with either transitive or intransitive relationships. Linear and near linear hierarchies are ensured, therefore, by a preponderance of Double Dominance and Double Subordinancc sequences in (he com-ponent triads of larger groups.
In an application of the method developed, sequences of dominance relationships formed in groups of three and four chickens were analyzed. In both experiments Double Dominance and Double Subordinance composed the overwhelming majority of all sequences — 91% in triads and 87% in tetrads. A further set of results indicated that intransitive relationships were common in "constructed" triads where chickens met only as component pairs and not in "real" triads where all three met together at the same time. These results suggest that hierarchy formation can be best viewed as a developmental process where preceding dominance interactions influence succeeding ones. This view was characterized as the "jigsaw puzzle" model of hierarchy formation, and it was contrasted with the more classical approach of explaining hierarchy structures by differences in the individual characteristics of group members. Many species beside chickens form linear and near linear hierarchies, and it was hypothesized that Double Dominance and Double Subordinance sequences also compose the majority of sequence occurrences in many of these species. The data available for other species were reviewed and offered tentative support for the hypothesis.
References ALLEE, W. C, FOREMAN, D., BANKS, E. M. & HOLABIRD, C. (1955). Effects of an androgen on
dominance and subordination in six common breeds of Gallus gallus. — Physiol. Zool. 28, p. 89-115.
BANKS. E. M. (1956). Social organization in Red Jungle Fowl hens (Gallus gallus subsp.). — Ecology 37, p. 239-248.
BARCHAS, P. R. & MENDOZA, S. P. (in press). Behavioral processes leading to linear hierarchies. — In: Status hierarchies: selected issues in the study of social differentiation (P. R. BARCHAS & S. P. MENDOZA, eds) Westport, Conn.: Greenwood Press.
BEKOFF, M. (1977). Quantitative studies of three areas of classical ethology: social dominance, behavioral taxonomy, and behavioral variability. — In: Quantitative methods in the study of animal behavior (B. A. HAZLETT, ed.) p. 1-46. New York: Academic Press.
BERNSTEIN, I. S. & GORDON, T. P. (1980). The social component of dominance relationships in rhesus monkeys (Macaca mulatta). — Anim. Behav. 28, p. 1033-1039.
BROWN, J. L. (1975). The evolution of behavior. — New York: Norton. CHASE, I. D. (1974). Models of hierarchy formation in animal societies. — Behavioral Science
19. p. 374-382. -----(1980). Social process and hierarchy formation in small groups: a comparative
perspective. — American Sociological Review 45, p. 905-924.

238 IVAN D. CHASE
COLLIAS, N. E, (1943). Statistical analysis of factors which make for success in initial encounters between hens, — Am. Nat. 77, p. 519-538.
GUHL, A. M. (1958). The development of social organization in the domestic chick. — Anim. Behav. 6, p. 92-111.
-----(1968). Social inertia and social stability in chickens. — Anim. Behav. 16, p. 219-232.
-----(1975). Social behavior of the domestic fowl. — In: Social hierarchy and dominance (M. SCHEIN, ed.) p. 156-201. Stroudsburg, Penn.: Dowden, Hutchinson & Ross.
HALL, M. (1967). Combinatorial theory. — Waltham, Mass.: Blasidell. HAUSFATER, G. (1975). Dominance and reproduction in baboons. — Contributions to
Primatology 7, p. 1-150. KING, M. G. (1965). The effect of social context on dominance capacity of domestic
hens. — Anim. Behav. 13, p. 132-133. KUMMER, H. (1975). Rules of dyad and group formation among captive gelada baboons.
— Proceeding from the symposia of the fifth congress of the International Prima- tological Society (S. KUDO, M. KAWAI, A. EHARA, S. KAWAMURA, eds) p. 129-159. Tokyo: Japan Science Press.
LANDAU, H. G. (1951). On dominance relations and the structure of animal societies: I. Effect of inherent characteristics. — Bulletin of Mathematical Biophysics 13, p. 1-19.
LOCKWOOD, R. (1979). Dominance in wolves: useful construct or bad habit? — In: The behavior and ecology of wolves (E. KLINGHAMMER, ed.) p. 225-244. New York: Garland Press.
MAIER, R. A. (1964). The role of the dominance-submission ritual in social recognition of hens. — Anim. Behav. 12, p. 59.
MCBRIDE, G. (1958). The measurement of aggressiveness in the domestic hen. — Anim. Behav. 6, p. 87-91.
---- (1964). A general theory of social organization and behavior. — University of Queensland Papers, Faculty of Veterinary Series, 1, p. 75-110.
MARLER, P. (1955). Studies of fighting in chaffinches (1) Behaviour in relation to the social hierarchy. — Brit. J. Anim. Behav. 3, p. 111-117. MASON, W. A. (1978). Ontogeny of social systems. — In: Recent advances in primatology, Vol. 1 (D.J. CHIVERS & J. HERBERT, eds) p. 5-14. London: Academic. MISSAKIAN, E. A. (1976). Aggression and dominance relations in peer groups of children
6-45 months of age. -— Paper presented at the annual meeting of the Animal Behavior Society, Boulder, Colorado.
MOUSSAS, E. (1980). Dominance in two-year old children. — Unpublished paper, Department of Psychology, SUNY at Stony Brook, Stony Brook, NY.
RASMUSSEN, O. G., BANKS, E. M., BERRY, T. H. & BECKER, D. E. (1962). Social dominance in gilts. —J. Anim. Sci. 21, p. 520-522.
SADE, D. S. (1967). Determinant of dominance in a group of free-ranging rhesus monkeys. — In: Social communication among primates (S. A, ALTMANN, ed.) p. 99-114. Chicago: University of Chicago Press.
SAVIN-WILLIAMS, R. C. (1977). Dominance in a human adolescent group. — Anim. Behav. 25, p. 400-406. -----(1979). Dominance hierarchies in groups of early adolescents. — Child Develop ment 50, p. 923-935. -----(1980). Dominance hierarchies in groups of middle to late adolescent males. —
Journal of Youth and Adolescence 9, p. 75-85. SCHEIN, M. (Ed.). (1975). Social hierarchy and dominance. — Stroudsberg, Penn.:
Dowden, Hutchinson & Ross. ----- & FOHRMAN, M. H. (1955). Social dominance relationships in a herd of diary cattle.
— Brit. J. Anim. Behav. 3, p. 45-55. ; SCHJELDERUP-EBBE, T. (1935). Social behavior in birds. — In: Handbook of social

DYNAMICS OF HIERARCHY FORMATION 239 psychology (C. MURCHISON, ed.) p. 947-972. Worcester, Mass.: Clark University Press.
----- (1975). Contributions to the social psychology of the domestic chicken. — In: Social hierarchy and dominance (M. SCHEIN, ed.) p.35-49. Stroudsburg, Penn.: Dowden, Hutchinson & Ross.
SEYFARTH, R. M., CHENEY, D. L. & HINDE, R. A. (1978). Some principles relating social interactions and social structure among primates. — In: Recent advances in primatology, Vol. 1 (D.J. CHIVERS & J. HERBERT, eds) p. 39-51. London: Academic
Press. SMITH, W. & HALE, E. B. (1959). Modification of social rank in the domestic fowl. — J.
Comp. Physiol. Psychol. 52, p. 373-375. STAMMBACH, E. (1978). On social differentiation in groups of captive female hamadryas
baboons. — Behaviour 67. p. 322-338. STEPHENSON, G. R. (1975). PLEXYN: a computer compatible grammar for coding social
interactions. — In: Social interaction analysis (M. E. LAMB, S. J. SUOMI & G. R. STEPHENSON, eds) p. 157-184. Madison: University of Wisconsin Press.
-----. SMITH, D. P. & ROBERTS, T. W. (1975). The SSR system: an open format event recording system with computerized transcription. — Behavior Research Methods and Instrumentation 7, p. 497-515.
STRUHSAKER, T. T. (1967). Social structure among vervet monkeys (Cercopithecus aethiops). - Behaviour 29, p. 8.1-121. TORDOFF, H. B. (1954). Social organization in a dock of captive, non-breeding red cross-
bills. — Condor 56, p. 346-358. TYLER. S. J. (1972). The behaviour and social organization of the New Forest ponies. —
Anim. Behav, Monographs 5, p. 87-196. VAITI, E. (1978). Nature and implications of the complexly organised social system in
non-human primates. — In: Recent advances in primatology, Vol. I (D.J. CHIVERS & J. HERBERT, eds) p. 18-30. London: Academic Press.
WAAL, F. B. M. DE, HOOFF. J. A. R. A. M. VAN & NETTO, W. J. (1976). An ethological analysis of types of agonistic interaction in a captive group of Javamonkeys (Macaca fascicularis). — Primates 17, p. 257-290.
WILSON, E. O. (1971). The insect societies. — Cambridge: Belknap Press of Harvard University Press.
----- (1975). Sociobiology. — Cambridge: Belknap Press of Harvard University Press. WOOD-GUSH, D. G. M. (1971). The behaviour of the domestic fowl. — London: Heine-
mann Educational Books.