dynamical behaviour of an ecological system with
TRANSCRIPT

ORIGINAL ARTICLE
Dynamical behaviour of an ecological systemwith Beddington–DeAngelis functional response
Sahabuddin Sarwardi1 • Md. Reduanur Mandal1 • Nurul Huda Gazi1
Received: 19 May 2016 / Accepted: 23 May 2016 / Published online: 18 June 2016
� Springer International Publishing Switzerland 2016
Abstract The objective of this paper is to systematically
study the dynamical behaviour of an ecological system
with Beddington–DeAngelis functional response which
avoids the criticism arising for ratio-dependent functional
response at low population densities of both species. The
essential mathematical features of the present model have
been analyzed thoroughly: local and global stability and the
bifurcations arising in some selected situations. We show
that the dynamical outcomes of the interactions among the
species are much sensitive to the system parameters and
initial population volumes. The ranges of the significant
parameters under which the system admits a Hopf bifur-
cation are investigated. The explicit formulae for deter-
mining the stability, direction and other properties of
bifurcating periodic solutions are also derived with the use
of both the normal form and the center manifold theory (cf.
Carr (1981)). Numerical illustrations are performed finally
in order to validate the applicability of the model under
consideration.
Keywords Ecological model � Stability � Hopf
bifurcation � Limit cycle � Center manifold � Numerical
simulation
Mathematics Subject Classification 92D25, 92D30,
92D40
Introduction
Mathematical models are important tools for analyzing
ecological models. The dynamic relationship between
predator and its prey is one of the dominant themes in
mathematical ecology due to its universality (cf. Anderson
and May (1981), Beretta and Kuang (1998), Freedman
(1990), Hadeler and Freedman (1989), Hethcote et al.
(2004), Ma and Takeuchi (1998), Venturino (1995), Xiao
and Chen (2001)). The most common method of modelling
ecological interactions consists of two differential equa-
tions with simple correspondence between the consumption
of prey by the predator and the populations growth. The
traditional predator–prey models have been studied exten-
sively (cf. Cantrell and Cosner (2001), Cosner et al. (1999),
Cui and Takeuchi (2006), Huo et al. (2007) and Hwang
(2003)), but these studies are questioned by several biolo-
gists. The most crucial element in these models is the
‘‘functional response’’—the expression that describes the
rate at which the prey are consumed by a predator. Models
were limited to use the Malthusian growth function, the
predator per capita consumption of prey following Holling
types II and III functional responses or density dependent
mortality rates. These functional responses depend only on
the prey volume x, but soon it became clear that the
predator volume y can influence this function by direct
interference while searching or by pseudo interference (cf.
Curds and Cockburn (1968), Hassell and Varley (1969) and
Salt (1974)). A simple way of incorporating predator
dependence in the functional response was proposed by
Arditi and Ginzburg Arditi and Ginzburg (1989), who
considered this response as a function of the ratio x/y. The
ratio-dependent response function produces richer dynam-
ics than that of all the Holling types responses, but it is
often criticized for the paradox that occurs at low densities
& Sahabuddin Sarwardi
1 Department of Mathematics, Aliah University, IIA/27,
New Town, Kolkata 700 156, West Bengal, India
123
Model. Earth Syst. Environ. (2016) 2:106
DOI 10.1007/s40808-016-0143-5

of both populations. Normally one would expect that the
population growth rate decrease when both the populations
fall bellow some critical size, because food-searching effort
becomes very high. For some ecological interactions ratio-
dependent model give negative feed back. The Lotka–
Volterra type predator–prey model with the Beddington–
DeAngelis functional response has been proposed and well
studied. This proposed model can be expressed as follows:
dx01dt
¼ rx01�1 � x01
k
�� c1x
01x
02
a1 þ x01 þ b1x02
dx02dt
¼ �d1x02 þ
c1e1x01x
02
a1 þ x01 þ b1x02
8>>><
>>>:
ð1:1Þ
with the initial conditions x01ð0Þ ¼ x010 [ 0 and
x02ð0Þ ¼ x020 [ 0. The functions x01ðtÞ; x02ðtÞ are the sizes of
prey and predator at any time t. All the system parameters
are assumed to be positive and have their usual biological
meanings. The functional responsec1x
01x0
2
a1þx01þb1x
02
in system
(1.1) was introduced by Beddington (1975) and DeAngelis
et al. (1975). It is similar to the well-known Holling type-II
functional response but has an extra term b1x2 in the
denominator which models mutual interference between
predators. It represents the most qualitative features of the
ratio-dependent models, but avoids the ‘‘low-densities
problem’’, which usually is the source of controversy. It
can be derived mechanistically from considerations of time
utilization (cf. Beddington (1975)) or spatial limits on
predation.
The present study is organized as follows: The basic
assumptions and the model formation are proposed in Sect.
2. Section 3 deals with some preliminary results. The
equilibria and their feasibility are given in Sect. 4. The
local analyses of the system around the boundary as well as
interior equilibria are discussed in Sect. 5. The global
analysis of the system around the interior equilibrium is
studied at length in Sect. 6. Simulation results are reported
in Sect. 7 while a final discussion and interpretation of the
results of the present study in ecological terms appear in
the concluding Sect. 8.
Model formulation
Firstly we replaced the logistic growth function rx1ð1 � x1
kÞ
of the prey species by the modified quasi-linear growth
function rx1ð1 � x1
x1þkÞ ¼ rð k
kþx1Þx1 ¼ r0x1 ðr0\rÞ in order
to make the model free from any axial equilibrium. This
fits better some special type of ecosystems, where the
environmental carrying capacity varies w.r.t. its prey size,
i.e., the carrying capacity is always greater than its present
prey size. In the present model we introduce one more
predator species in the model (1.1) to make it closer to
reality. Thus, our final model is
dx1
dt¼ rx1ð1�
x1
x1 þ kÞ� c1x1x2
a1 þ x1 þb1x2
� c2x1x3
a2 þ x1 þb2x3
dx2
dt¼�d1x2 þ
c1e1x1x2
a1 þ x1 þb1x2
;
dx3
dt¼�d2x3 þ
c2e2x1x3
a2 þ x1 þb2x3
8>>>>>><
>>>>>>:
ð2:1Þ
where x1 is the population size of the prey and x2; x3 are
the population sizes of the predator species at any time
t. It is assumed that all the system parameters are pos-
itive constants. Here r and k are the growth rate and the
half-saturation constant for the prey species, d1; d2 are
the first and second predators death rates respectively.
c1, c2 are the respective search rates of the first and
second predator on the prey species, c1
a1, c2
a2are the
maximum number of prey that can be eaten by the first
and second predator per unit time respectively; 1a1
, 1a2
are
their respective half saturation rates while e1, e2 are the
conversion factors, denoting the number of newly born
of the first and second predator for each captured prey
species respectively ð0\e1; e2\1Þ. The parameters b1
and b2 measure the coefficients of mutual interference
among the first and second predator respectively. The
terms c1x1x2
a1þx1þb1x2and c2x1x3
a2þx1þb2x3denote the respective
predator responses.
Some preliminary results
Existence and positive invariance
Letting, x ¼ ðx1; x2; x3Þt; f : R3 ! R3, f ¼ ðf1; f2; f3Þt; the
system (2.1) can be rewritten as _x ¼ f ðxÞ. Here
fi 2 C1ðRÞ for i ¼ 1; 2; 3; where f1 ¼ rx1ð1 � x1
x1þkÞ
� c1x1x2
a1þx1þb1x2� c2x1x3
a2þx1þb2x3; f2 ¼ �d1x2 þ c1e1x1x2
a1þx1þb1x2and f3 ¼
�d2x3 þ c2e2x1x3
a2þx1þb2x3: Since the vector function f is a smooth
function of the variables ðx1; x2; x3Þ in the positive octant
X0 ¼ fðx1; x2; x3Þ : x1 [ 0; x2 [ 0; x3 [ 0g; the local exis-
tence and uniqueness of the solution of the system (2.1)
hold.
Persistence
If a compact set D � X0 ¼ fðx1; x2; x3Þ : xi [ 0; i ¼1; 2; 3g exists such that all solutions of (2.1) eventually
enter and remain in D, the system is called persistent.
106 Page 2 of 14 Model. Earth Syst. Environ. (2016) 2:106
123

Proposition 3.1 The system (2.1) is persistent if the
conditions: ðiÞ r[ d1 þ d2; ðiiÞx11[ a2d2
c2e2�d2; ðiiiÞ x12
[a1d1
c1e1�d1are satisfied.
Proof We use the method of average Lyapunov function
(cf. Gard and Hallam (1979)), considering a function of the
form
Vðx1; x2; x3Þ ¼ xc1
1 xc2
2 xc3
3 ;
where c1; c2 and c3 are positive constants to be determined.
We define
Pðx1;x2;x3Þ¼_V
V
¼ c1
�r� rx1
x1þk� c1x2
a1þx1þb1x2
� c2x3
a2þx1þb2x3
�
þc2
��d1þ
c1e1x1
a1þx1þb1x2
�
þc3
��d2þ
c2e2x1
a2þx1þb2x3
�:
We now prove that this function is positive at each of the
boundary equilibria. Let ci¼c; for i¼1;2;3: In fact at E0;
we have Pð0;0;0Þ¼cðr�d1�d2Þ[0 from the condition
(i). Moreover, from condition (ii) and (iii), we find the
values of P at E1 and E2 respectively,
Pðx11; x21
; 0Þ ¼ c��d2 þ
e2c2x11
a2 þ x11
�[ 0;
Pðx12; 0; x32
Þ ¼ c��d1 þ
e1c1x12
a1 þ x12
�[ 0:
Hence, there always exists a positive number c such that
P[ 0 at the boundary equilibria. Hence V is an average
Lyapunov function and thus, the system (2.1) is persis-
tent. h
Since the system is uniformly persistent, there exists
r[ 0 and s[ 0 such that xiðtÞ[ r; for all t[ s;i ¼ 1; 2; 3:
Boundedness
Boundedness implies that the system is biologically con-
sistent. The following propositions ensure the boundedness
of the system (2.1).
Proposition 3.2 The prey population is always bounded
from above.
Proof Before proving that the prey population is bounded
above, we need to prove that the predator populations x2
and x3 are bounded above. To prove this result, considering
the second sub equation of the system (2.1) and one can
obtain the following differential inequality:
dx2
dt� � ðd1 � c1e1Þx2:
Integrating the above differential inequality between the
limits 0 and t, we have x2ðtÞ� x2ð0Þe�ðd1�c1e1Þt: Thus, if
ðd1 � c1e1Þ[ 0; then a positive number s1 is found and there
exist a positive constant m1 such that x2ðtÞ�m1; for all
t� s1: By using a similar argument, one can obtain that, if
ðd2 � c2e2Þ[ 0; then corresponding to a positive number s2
there exists a positive constantm2 such that x3ðtÞ�m2; for all
t� s2: Both results can be written as xi [m ¼ min ðm1;m2Þfor all t[ s3 ¼ max ðs1; s2Þ; i ¼ 2; 3; with the additional
condition min ðd1 � c1e1; d2 � c2e2Þ[ 0:
Now from the first sub-equation of (2.1), the following
inequality is found
dx1
dt��ðe1 þ e2Þr� rk
�x1
km
k�rm� ðe1 þ e2Þr
�
ðe1 þ e2Þr� rk� x1
!
:
Hence, by using simple standard arguments, we have
lim supt!þ1
x1ðtÞ�krm � ðe1 þ e2Þkrðe1 þ e2Þr� rk
¼ w; where
rm
e1 þ e2
\r\rk
e1 þ e2
:
h
Proposition 3.3 The solutions of (2.1) starting in X0 are
uniformly bounded with an ultimate bound.
Proof Considering the total environment population v ¼x1 þ x2
e1þ x3
e2; using the theorem on differential inequality
(cf. Birkhoff and Rota (1982)) and following the steps of
Haque and Venturino (2006), Sarwardi et al. (2013),
boundedness of the solution trajectories of this model is
established. In particular,
lim supt!þ1
�x1 þ
x2
e1
þ x3
e2
�� ðr þ 1Þk þ w
q¼ M; where
q ¼ min ð1; d1; d2Þ;with the last bound independent of the initial condition.
Hence, all the solutions of (2.1) starting in R3þ for any
h[ 0 evolve with respect to time in the compact region
�X ¼ ðx1; x2; x3Þ 2 R3þ : x1 þ
x2
e1
þ x3
e2
�M þ h
� �: ð3:2Þ
Equilibria and their feasibility
The equilibria of the dynamical system (2.1) are:
1.
(a) The trivial equilibrium point E0ð0; 0; 0Þ is
always feasible.
Model. Earth Syst. Environ. (2016) 2:106 Page 3 of 14 106
123

2.
(a) The first boundary equilibrium point is
E1ðx11; x21
; 0Þ: The component x11is a root
of the quadratic equation l1x211þ ðl2 þ l1k þ
rkb1e1Þx11þ l2k ¼ 0; where l1 ¼ ðd1 � c1e1Þ,
l2 ¼ a1d1: If l1\0, then the quadratic equation
in x11possesses a unique positive root and
consequently x21¼ ðc1e1�d1Þx11
�a1d1
b1d1: The feasi-
bility of the equilibrium E1 is maintained if
the condition x11[ b1d1
c1e1�d1is satisfied.
(b) The second boundary equilibrium point is
E2ðx12; 0; x32
Þ: The component x12is
the root of the quadratic equation m1x212þ
ðm2 þ m1k þ rkb2e2Þx12þ m2k ¼ 0; where
m1 ¼ ðd2 � c2e2Þ, l2 ¼ a2d2: If m1\0, the
quadratic equation in x12possesses a unique
positive root and consequently x32¼
ðc2e2�d2Þx12�a2d2
b2d2: The feasibility of the equilib-
rium E2 holds if the condition x12[ b2d2
c2e2�d2
holds.
3.
(a) The interior equilibrium point is
E�ðx1�; x2�; x3�Þ; where the first component
x1� is the root of the following quadratic
equation:
n1x21� þ ðn2 þ n1k þ rkb1b2e1e2Þx1� þ n2k ¼ 0;
ð4:1Þ
where n1 ¼ b1e1ðd2 � c2e2Þ þ b2e2ðd1 � c1e1Þ and
n2 ¼ b2e2d1a1 þ b1e1d2a2:
Case I: Let n1\0. In this case there exists exactly one
positive root of the quadratic equation (4.1)
irrespective of the sign of ðn2 þ n1k
þrkb1b2e1e2Þ:Case II: Let n1 [ 0. In this case there are two possi-
bilities: (i) if n2 þ n1k þ rkb1b2e1e2 [ 0, then
there is no positive solution and (ii) if
n2 þ n1k þ rkb1b2e1e2\0, then there exists
two positive roots or no positive root. Here we
consider the Case I. Under this assumption the
next two components of the interior equilib-
rium can be obtained as x2� ¼ðc1e1�d1Þx1��a1d1
b1d1; x3� ¼ ðc2e2�d2Þx1��a2d2
b2d2: The
feasibility of this equilibrium point E� holds
under the condition x1� [ max a1d1
c1e1�d1;
n
a2d2
c2e2�d2g: Moreover, the positivity condition
of second and third components of the interior
equilibrium ensures the impossibility of the
Case II.
Remark The feasibility and existences conditions of both
the planar equilibria E1 and E2 immediately implies the
existence of the unique feasible interior equilibrium point
E�: But the existence of the unique feasible interior equi-
librium point E� implies three possibilities: (i) E1 exists
and E2 does not exist, (ii) E2 exists and E1 does not exist,
(iii) existence of both.
Local stability and bifurcation
The Jacobian matrix J(x) of the system (2.1) at any point
x ¼ ðx1; x2; x3Þ is given by
JðxÞ3�3 ¼
rk2
ðx1 þ kÞ2� c1x2ða1 þb1x2Þða1 þ x1 þb1x2Þ2
� c2x3ða2 þb2x3Þða2 þ x1 þb2x3Þ2
� c1x1ða1 þ x1Þða1 þ x1 þb1x2Þ2
� c2x1ða2 þ x1Þða2 þ x1 þb2x3Þ2
c1e1x2ða1 þb1x2Þða1 þ x1 þb1x2Þ2
�d1 þc1e1x1ða1 þ x1Þða1 þ x1 þb1x2Þ2
0
c2e2x3ða2 þb2x3Þða2 þ x1 þb2x3Þ2
0 �d2 þc2e2x1ða2 þ x1Þða2 þ x1 þb2x3Þ2
0
BBBBBBBB@
1
CCCCCCCCA
: ð5:1Þ
106 Page 4 of 14 Model. Earth Syst. Environ. (2016) 2:106
123

Its characteristic equation is DðkÞ ¼ k3 þ k1k2 þ k2kþ
k3 ¼ 0, where k1 ¼ �trðJÞ, k2 ¼ M and k3 ¼ � detðJÞ;where M is the sum of the principal minors of order two of
J.
Note that a Hopf bifurcation occurs if there exist a
certain bifurcation parameter r ¼ rc such that C2ðrcÞ ¼k1ðrcÞk2ðrcÞ � k3ðrcÞ ¼ 0 with k2ðrcÞ[ 0 andddrð Re ðkðrÞÞÞjr¼rc
6¼ 0; where k is root of the character-
istic equation DðkÞ ¼ 0.
Local analysis of the system around E0;E1;E2
Stability The eigenvalues of the Jacobian matrix JðE0Þ are
r;�d1 and �d2. Hence E0 is unstable in nature (saddle
point). Let JðE1Þ ¼ ðnijÞ3�3 and JðE2Þ ¼ ðgijÞ3�3: Using
the Routh-Hurwitz criterion, it can be easily shown that the
eigenvalues of the matrices JðE1Þ and JðE2Þ have negative
real parts iff the conditions e1x11þ x21
[ kð1�b1e1Þ�a1
b1and
e2x12þ x32
[ kð1�b2e2Þ�a2
b2are satisfied. Hence the equilibria
E1 and E2 are locally asymptotically stable under the above
conditions (cf. Sect. 4 of Sarwardi et al. (2012)).
Bifurcation Since the equilibrium point E0 is a saddle,
there is no Hopf bifurcation around it. In order to have
Hopf bifurcation around the equilibria E1, E2, it is suffi-
cient to show that the coefficient of k in the quadratic
factor of the characteristic polynomial of JðEkÞ ðk ¼ 1; 2Þis zero and the constant term is positive. The conditions
for which annihilation of the linear terms in the quadratic
factors of the characteristic polynomials of JðE1Þ and
JðE2Þ can be made possible are n11 þ n22 ¼ 0 and g11 þg33 ¼ 0: For a detailed analysis, interested readers are
referred to Appendix A of Haque and Venturino (2006).
The parametric regions where Hopf bifurcations occur
around E1 and E2 are respectively established by the
equality constraints e1x11þ x21
¼ kð1�b1e1Þ�a1
b1and e2x12
þx32
¼ kð1�b2e2Þ�a2
b2.
Local analysis of the system around the interior
equilibrium
Proposition 5.1 The system (2.1) around E� is locally
asymptotically stable if the condition
(i) k\min fa1 þ b1x2�; a2 þ b2x3�g is satisfied.
Proof Let Jðx�Þ = ðJijÞ3�3 be the Jacobian matrix at the
interior equilibrium point E� ¼ x� of the system (2.1). The
components of Jðx�Þ are
J11 ¼c1x1�x2�
�k � ða1 þ b1x2Þ
�
ðx1� þ kÞða1 þ x1� þ b1x2�Þþ
c2x1�x3��k � ða2 þ b2x3Þ
�
ðx1� þ kÞða2 þ x1� þ b2x3�Þ;
J12 ¼ � c1x1�ða1þx1�Þða1þx1�þb1x2�Þ2 \0, J13 ¼ � c2x1�ða2þx1�Þ
ða2þx1�þb2x3�Þ2 \0,
J21 ¼ c1e1x2�ða1þb1x2�Þða1þx1�þb1x2�Þ2 [ 0, J22 ¼ � b1c1e1x1�x2�
ða1þx1�þb1x2�Þ2 \0, J23 ¼ 0,
J31 ¼ c2e2x3�ða2þb2x3�Þða2þx1�þb2x3�Þ2 [ 0, J32 ¼ 0, J33 ¼ � b2c2e2x1�x3�
ða2þx1�þb2x3�Þ2
\0:
Then the characteristic equation of the Jacobian matrix
Jðx�Þ can be written as
k3 þ k1k2 þ k2kþ k3 ¼ 0; ð5:2Þ
where k1¼�trðJÞ¼�ðJ11þJ22þJ33Þ, k2¼M11þM22 þM33
¼ðJ11J22�J21J12ÞþJ22J33þðJ11J33�J31J13Þ, k3¼�detðJÞ¼�
�J11J22J33�J12J21J33�J31J13J22
�, and C2¼k1k2�k3 ¼
�ðJ11þJ22Þ�J33ðJ11þJ22þJ33ÞþðJ11J22�J21J12Þ
�þJ13J31
ðJ11þJ33Þ:It is clear that k1 [ 0 if J11\0, i.e.,
k\min fa1 þ b1x2�; a2 þ b2x3�g and consequently
C2 [ 0. Hence the Routh–Hurwitz condition is satisfied
for the matrix J�, i.e., all the characteristic roots of J� have
negative real parts. So the system is locally asymptotically
stable around E�.
Theorem 5.2 The dynamical system (2.1) undergoes a
Hopf bifurcation around the interior equilibrium point E�whenever the critical parameter r attains the value r ¼ rc in
the domain
DHB ¼nrc 2 Rþ : C2ðrcÞ ¼ k1ðrcÞk2ðrcÞ � k3
ðrcÞ ¼ 0 with k2ðrcÞ[ 0 anddC2
drjr¼rc
6¼ 0o:
Proof The equation (5.2) will have a pair of purely
imaginary roots if k1k2 � k3 ¼ 0 for some set of values of
the system parameters. Let us now suppose that r ¼ rc is
the value of r satisfying the condition k1k2 � k3 ¼ 0. Here
only J11 contains r explicitly. So, we write the equation
k1k2 � k3 ¼ 0 as an equation in J11 to find rc as follows:
h1J211 þ h2J11 þ h3 ¼ 0; ð5:3Þ
where h1 ¼ J22 þ J33, h2 ¼ �J222 þ J2
33 � J13J31 � J12J21,
h3 ¼ ðJ22 þ J33ÞJ22J33 � J13J31J33 � J12J21J22.
Thus, J11 ¼ 12h1
ð�h2 ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffih2
2 � 4h1h3
pÞ ¼ J�11:
Or,
r ¼ ðx1� þ kÞ2
k2
� J�11 þc1x2�ða1 þ b1x2�Þða1 þ x1� þ b1x2�Þ2
� c2x3�ða2 þ b2x3�Þða2 þ x1� þ b2x3�Þ2
" #
¼ rc:
ð5:4Þ
Using the condition k1k2 � k3 ¼ 0; from equation (5.2) one
can obtain
ðkþ k1Þðk2 þ k2Þ ¼ 0; ð5:5Þ
which has three roots k1 ¼ þiffiffiffik
p2; k2 ¼ �i
ffiffiffik
p2; k3 ¼
�k1: Thus, there is a pair of purely imaginary eigenvalues
Model. Earth Syst. Environ. (2016) 2:106 Page 5 of 14 106
123

iffiffiffik
p2. For all values of k, the roots are, in general, of the
form
k1ðrÞ ¼ n1ðrÞ þ in2ðrÞ; k2ðrÞ ¼ n1ðrÞ � in2ðrÞ; k3ðrÞ ¼ �k1ðrÞ:
Differentiating the characteristic equation (5.2) w.r.t. r,
we have
dkdr
¼� k2 _k1 þ k _k2 þ _k3
3k2 þ 2k1kþ k2
jk¼iffiffiffik2
p
¼_k3 � k2
_k1 þ i _k2
ffiffiffiffiffik2
p
2ðk2 � ik1
ffiffiffiffiffik2
pÞ
¼_k3 � ð _k1k2 þ k1
_k2Þ2ðk2
1 þ k2Þþ i
ffiffiffiffiffik2
pðk1
_k3 þ k2_k2 � k1
_k1k2Þ2k2ðk2
1 þ k2Þ
¼ �dC2
dr
2ðk21 þ k2Þ
þ i
" ffiffiffiffiffik2
p_k2
2k2
�k1
ffiffiffiffiffik2
pdC2
dr
2k2ðk21 þ k2Þ
#
:
ð5:6Þ
Hence,
d
dr
�Re ðkðrÞÞ
�jr¼rc
¼�dC2
dr
2ðk21 þ k2Þ
jr¼rc6¼ 0: ð5:7Þ
Using the monotonicity condition of the real part of the
complex rootdðReðkðrÞÞÞ
d rjr¼rc
6¼ 0 (cf. Wiggins (2003), pp.
380), one can easily establish the transversality conditiondC2
drjr¼rc
6¼ 0; to ensure the existence of Hopf bifurcation
around E�:
Global analysis of the system around the interiorequilibrium
Direction of Hopf bifucation of the system (2.1)
around E�
In this Section we study on the direction of Hopf bifucation
around the interior equilibrium. From the model equations
(2.1), we have
_x ¼ f ðxÞ; ð6:1Þ
where x ¼ ðx1; x2; x3Þt, f ¼ ðf 1; f 2; f 3Þt ¼rx1ð1 � x1
x1 þ kÞ � c1x1x2
a1 þ x1 þ b1x2
� c2x1x3
a2 þ x1 þ b2x3
�d1x2 þc1e1x1x2
a1 þ x1 þ b1x2
�d2x3 þc2e2x1x3
a2 þ x1 þ b2x3
0
BBBB@
1
CCCCA:
Here, at x ¼ x�, f ¼ 0. Let y ¼ ðy1; y2; y3Þ ¼ ðx1 � x1�;x2 � x2�; x3 � x3�Þ. Putting in equation (6.1), we have
_y ¼ Jðx1�; x2�; x3�Þyþ /; ð6:2Þ
where the components of nonlinear vector function / ¼ð/1;/2;/3Þt are given by
/i ¼ f ix1x1y2
1 þ f ix2x2y2
2 þ f ix3x3y2
3 þ 2f ix2x3y2y3 þ 2f ix3x1
y3y1
þ 2f ix1x2y1y2 þ h.o.t. ; i ¼ 1; 2; 3: ð6:3Þ
The coefficients of nonlinear terms in yi; i ¼ 1; 2; 3 are
given by
f 1x1x1
¼� 2rk2
ðx1 þ kÞ3þ 2c1x2ða1 þ b1x2Þða1 þ x1 þ b1x2Þ3
þ 2c2x3ða2 þ b2x3Þða2 þ x1 þ b2x3Þ3
;
f 1x2x2
¼ 2b1c1x1ða1 þ x1Þða1 þ x1 þ b1x2Þ3
; f 1x3x3
¼ 2b2c2x1ða2 þ x1Þða2 þ x1 þ b2x3Þ3
;
f 1x2x3
¼ 0; f 1x1x2
¼�c1a1ða1 þ x1 þ b1x2Þþ 2b1c1x1x2
ða1 þ x1 þ b1x2Þ3;
f 1x3x1
¼�c2a2ða2 þ x1 þ b2x3Þþ 2b2c2x3x1
ða2 þ x1 þ b2x3Þ3;
f 2x1x1
¼�2c1e1x2ða1 þ b1x2Þða1 þ x1 þ b1x2Þ3
; f 2x2x2
¼�2b1c1e1x1ða1 þ x1Þða1 þ x1 þ b1x2Þ3
;
f 2x3x3
¼ 0; f 2x1x2
¼ a1c1e1ða1 þ x1 þ b1x2Þþ 2b1c1e1x1x2
ða1 þ x1 þ b1x2Þ3;
f 2x2x3
¼ 0; f 2x3x1
¼ 0; f 3x1x1
¼ � 2c2e2x3ða2 þ b2x3Þða2 þ x1 þ b2x3Þ3
;
f 3x2x2
¼ 0; f 3x3x3
¼ � 2b2c2e2x1ða2 þ x1Þða2 þ x1 þ b2x3Þ3
;
f 3x2x3
¼ 0; f 3x3x1
¼ a2c2e2ða2 þ x1 þ b2x3Þ þ 2b2c2e2x1x3
ða2 þ x1 þ b2x3Þ3;
f 3x1x2
¼ 0:
Let P be the matrix formed by the column vectors
ðu2; u1; u3Þ; which are the eigenvectors corresponding to
the eigenvalues k1;2 ¼ iffiffiffiffiffik2
pand k3 ¼ �k1 of Jðx1�;
x2�; x3�Þ: Then Jðx1�; x2�; x3�Þu2 ¼ iffiffiffiffiffik2
pu2; Jðx1�; x2�;
x3�Þu1 ¼ �iffiffiffiffiffik2
pu1; and Jðx1�; x2�; x3�Þu3 ¼ �k1u3:
Thus,
P ¼
0 1 1�J21
ffiffiffik2
p
J222þk2
�J21J22
J222þk2
�J21
J22þk1
�J31
ffiffiffik2
p
J233þk2
�J31J33
J233þk2
�J31
J33þk1
0
BB@
1
CCA ¼ ðpijÞ3�3:
Let us make use of the transformation y ¼ Pz; so that the
system (6.2) is reduced to the following one
_z ¼ P�1Jðx1�; x2�; x3�ÞPz þ P�1
/ ¼0 � i
ffiffiffiffiffik2
p0
iffiffiffiffiffik2
p0 0
0 0 � k1
0
B@
1
CAz þ P�1/:ð6:4Þ
106 Page 6 of 14 Model. Earth Syst. Environ. (2016) 2:106
123

Here P�1 ¼ AdjPdetP
¼ ðqijÞ3�3; where
q11 ¼ 1
detP
J21J22J31
ðJ222 þ k2ÞðJ33 þ k1Þ
� J21J33J31
ðJ222 þ k2ÞðJ22 þ k1Þ
!
;
q12 ¼ 1
detP
J31
J233 þ k1
� J31J33
J233 þ k2
Þ;
q13 ¼ 1
detP
�J21
J21 þ k1
þ J21J22
J222 þ k2
!
;
q21 ¼ 1
detP
J21
ffiffiffiffiffik2
pJ31
ðJ222 þ k2ÞðJ22 þ k1Þ
� J21
ffiffiffiffiffik2
pJ31
ðJ233 þ k2ÞðJ22 þ k1Þ
!
;
q22 ¼ 1
detP
ffiffiffiffiffik2
pJ31
ðJ233 þ k2Þ
; q23 ¼ 1
detP
�ffiffiffiffiffik2
pJ31
ðJ233 þ k2Þ
;
q31 ¼ 1
detP
J21
ffiffiffiffiffik2
pJ31J33
ðJ222 þ k2ÞðJ2
33 þ k2Þ� J21
ffiffiffiffiffik2
pJ31J22
ðJ233 þ k2ÞðJ2
22 þ k2Þ
!
;
q32 ¼ 1
detP
�ffiffiffiffiffik2
pJ21
ðJ222 þ k2Þ
; q33 ¼ 1
detP
ffiffiffiffiffik2
pJ21
ðJ222 þ k2Þ
:
The system (6.4) can be written as
d
dt
z1
z2
¼ 0 �
ffiffiffiffiffik2
pffiffiffiffiffik2
p0
!z1
z2
þ Fðz1; z2; z3Þ;
ð6:5Þdz3
dt¼� k1z3 þ Gðz1; z2; z3Þ: ð6:6Þ
On the center-manifold (cf. Carr (1981), Kar et al. (2012))
z3 ¼ 1
2b11z
21 þ 2b12z1z2 þ b22z
22
� �ð6:7Þ
Therefore,
_z3 ¼ b11z1 þ b12z2 b12z1 þ b22z2ð Þ 0 �ffiffiffiffiffik2
pffiffiffiffiffik2
p0
!z1
z2
!
¼ffiffiffiffiffik2
pb12z
21 þ
ffiffiffiffiffik2
pðb22 � b11Þz1z2 �
ffiffiffiffiffik2
pb12z
22
ð6:8Þ
Using (6.4) and (6.6), we have
_z3 ¼ �k1z3 þ q31/1 þ q32/2 þ q33/3 ð6:9Þ
From the Eqs. (6.8) and (6.9), we have
ffiffiffiffiffik2
pb12z
21 þ
ffiffiffiffiffik2
pðb22 � b11Þz1z2 �
ffiffiffiffiffik2
pb12z
22
¼ � 1
2k1ðb11z
21 þ 2b12z1z2 þ b22z
22Þ þ q31
�f 1x1x1
fp11z1 þ p12z2 þ p13
1
2ðb11z
21 þ 2b12z1z2 þ b22z
22Þg
2
þ f 1x2x2
fp21z1 þ p22z2 þ p23
1
2ðb11z
21 þ 2b12z1z2 þ b22z
22Þg
2 þ f 1x3x3
fp31z1 þ p32z2 þ p33
1
2ðb11z
21 þ 2b12z1z2 þ b22z
22Þg
2
þ 2f 1x3x1
fp31z1 þ p32z2 þ p33
1
2ðb11z
21 þ 2b12z1z2 þ b22z
22Þgfp11z1 þ p12z2 þ p13
1
2ðb11z
21 þ 2b12z1z2 þ b22z
22Þg
þ 2f 1x1x2
fp11z1 þ p12z2 þ p13
1
2ðb11z
21 þ 2b12z1z2 þ b22z
22Þg
� fp21z1 þ p22z2 þ p23
1
2ðb11z
21 þ 2b12z1z2 þ b22z
22Þg�
þ q32½f 2x1x1
fp11z1 þ p12z2 þ p13
1
2ðb11z
21 þ 2b12z1z2 þ b22z
22Þg
2
þ f 2x2x2
fp21z1 þ p22z2
þ p23
1
2ðb11z
21 þ 2b12z1z2 þ b22z
22Þg
2 þ f 2x3x3
fp31z1 þ p32z2 þ p33
1
2ðb11z
21 þ 2b12z1z2 þ b22z
22Þg
2
þ 2f 2x2x3
fp21z1 þ p22z2 þ p23
1
2ðb11z
21 þ 2b12z1z2 þ b22z
22Þgfp31z1 þ p32z2 þ p33
1
2ðb11z
21 þ 2b12z1z2 þ b22z
22Þg
þ 2f 2x3x1
fp31z1 þ p32z2 þ p33
1
2ðb11z
21 þ 2b12z1z2 þ b22z
22Þgfp11z1 þ p12z2 þ p13
1
2ðb11z
21 þ 2b12z1z2 þ b22z
22Þg
þ 2f 2x1x2
fp11z1 þ p12z2 þ p13
1
2ðb11z
21 þ 2b12z1z2 þ b22z
22Þg � fp21z1 þ p22z2 þ p23
1
2ðb11z
21 þ 2b12z1z2 þ b22z
22Þg�
þ q33
�f 3x1x1
fp11z1 þ p12z2 þ p13
1
2ðb11z
21 þ 2b12z1z2 þ b22z
22Þg
2 þ f 3x2x2
fp21z1 þ p22z2 þ p23
1
2ðb11z
21 þ 2b12z1z2 þ b22z
22Þg
2
þ f 3x3x3
fp31z1 þ p32z2 þ p33
1
2ðb11z
21 þ 2b12z1z2 þ b22z
22Þg
2 þ 2f 3x2x3
fp21z1 þ p22z2
þ p23
1
2ðb11z
21 þ 2b12z1z2 þ b22z
22Þgfp31z1 þ p32z2 þ p33
1
2ðb11z
21 þ 2b12z1z2 þ b22z
22Þg þ 2f 3
x3x1fp31z1 þ p32z2
þ p33
1
2ðb11z
21 þ 2b12z1z2 þ b22z
22Þgfp11z1 þ p12z2 þ p13
1
2ðb11z
21 þ 2b12z1z2 þ b22z
22Þg þ 2f 3
x1x2fp11z1 þ p12z2
þ p13
1
2ðb11z
21 þ 2b12z1z2 þ b22z
22Þgfp21z1 þ p22z2 þ p23
1
2ðb11z
21 þ 2b12z1z2 þ b22z
22Þg�:
Model. Earth Syst. Environ. (2016) 2:106 Page 7 of 14 106
123

Comparing the coefficients of z21, z1z2 and z2
2 from both
sides, we have
and
From equations (6.10), (6.11) and (6.12), we have
1
2k1
ffiffiffiffiffik2
p0
�ffiffiffiffiffik2
pk1
ffiffiffiffiffik2
p
0 �ffiffiffiffiffik2
p 1
2k1
0
BBBB@
1
CCCCA
b11
b12
b22
0
B@
1
CA ¼X1
X2
X3
0
B@
1
CA: ð6:13Þ
The equation (6.13) gives the coefficients b11, b12 and b22
as follows:
b11 ¼k2ðX1 þ X3Þ � k1
2ðffiffiffiffiffik2
pX2 � k1X1Þ
ðk31
4þ k1k2Þ
;
b12 ¼k2
1X2
4� k1
ffiffiffik2
p
2ðX3 � X1Þ
ðk31
4þ k1k2Þ
;
b22 ¼k2ðX1 þ X3Þ þ k2
1X3
2þ k1
ffiffiffik2
p
2X2
ðk31
4þ k1k2Þ
:
The flow of the central manifold is characterized by the
reduced system as
ffiffiffiffiffik2
pb12 þ
k1
2b11
¼ q31 f 1x1x1
p211 þ f 1
x2x2p2
21 þ f 1x3x3
p231 þ 2f 1
x2x3p21p31 þ 2f 1
x3x1p31p11 þ 2f 1
x1x2p11p21
h i
þ q32 f 2x1x1
p211 þ f 2
x2x2p2
21 þ f 2x3x3
p231 þ 2f 2
x2x3p21p31 þ 2f 2
x3x1p31p11 þ 2f 2
x1x2p11p21
h i
þ q33 f 3x1x1
p211 þ f 3
x2x2p2
21 þ f 3x3x3
p231 þ 2f 3
x2x3p21p31 þ 2f 3
x3x1p31p11 þ 2f 3
x1x2p11p21
h i
¼ X1;
ð6:10Þ
�ffiffiffiffiffik2
pðb11 � b22Þ þ k1b12
¼ 2q31½f 1x1x1
p11p12 þ f 1x2x2
p21p22 þ f 1x3x3
p31p32 þ f 1x2x3
ðp21p32 þ p22p31Þ þ f 1x3x1
ðp11p32
þ p12p31Þ þ f 1x1x2
ðp11p22 þ p12p21Þ þ q32½f 2x1x1
p11p12 þ f 2x2x2
p21p22 þ f 2x3x3
p31p32 þ f 2x2x3
� ðp21p32 þ p22p31Þ þ f 2x3x1
ðp11p32 þ p12p31Þ þ f 2x1x2
ðp11p22 þ p12p21Þ þ q33 f 3x1x1
p11p12
h
þ f 3x2x2
p21p22 þ f 3x3x3
p31p32 þ f 3x2x3
ðp21p32 þ p22p31Þ þ f 3x3x1
ðp11p32 þ p12p31Þ
þ f 3x1x2
ðp11p22 þ p12p21Þi¼ X2;
ð6:11Þ
�ffiffiffiffiffik2
pb12 þ
k1
2b22
¼ q31 f 1x1x1
p212 þ f 1
x2x2p2
22 þ f 1x3x3
p232 þ 2f 1
x2x3p22p32 þ 2f 1
x3x1p31p12 þ 2f 1
x1x2p12p22
h i
þ q32 f 2x1x1
p212 þ f 2
x2x2p2
22 þ f 2x3x3
p232 þ 2f 2
x2x3p22p32 þ 2f 2
x3x1p31p12 þ 2f 2
x1x2p12p22
h i
þ q33 f 3x1x1
p212 þ f 3
x2x2p2
22 þ f 3x3x3
p232 þ 2f 3
x2x3p22p32 þ 2f 3
x3x1p31p12 þ 2f 3
x1x2p12p22
h i
¼ X3:
ð6:12Þ
106 Page 8 of 14 Model. Earth Syst. Environ. (2016) 2:106
123

d
dt
z1
z2
¼ 0 �
ffiffiffiffiffik2
pffiffiffiffiffik2
p0
!z1
z2
þ F1
F2
; ð6:14Þ
where F1 ¼ q11/1 þ q12/2 þ q13/3 þ h:o:t, F2 ¼ q21/1þq22/2 þ q23/3 þ h:o:t. The stability of the bifurcating
limit cycle can be determined by the sign of the para-
metric expression
P ¼ F1111 þ F2
112 þ F1122 þ F2
222
þ F112ðF1
11 þ F122Þ � F2
12ðF211 þ F2
22Þ � F111F
211 þ F1
22F222ffiffiffiffiffi
k2
p ;
ð6:15Þ
where Fijk ¼ o3Foziozjozk
at the origin. If the value of the
above expression is negative, then the Hopf bifurcating
limit cycle is stable and we have a supercritical Hopf
bifurcation. If the value is positive, then the Hopf
bifurcating limit cycle is unstable and the bifurcation is
subcritical.
Here
F111 ¼ 2q11½f 1
x1x1p2
11 þ f 1x2x2
p221 þ f 1
x3x3p2
31 þ 2f 1x3x1
p11p31 þ 2f 1x1x2
p11p21
þ 2q12½f 2x1x1
p211 þ f 2
x2x2
� p221 þ 2f 2
x1x2p11p21 þ 2q13½f 3
x1x1p2
11 þ f 3x3x3
p231 þ 2f 3
x3x1p11p31;
F112 ¼ 2q11½f 1
x1x1p11p12 þ f 1
x2x2p21p22 þ f 1
x3x3p31p32 þ f 1
x3x1
ðp11p32 þ p12p31Þ þ f 1x1x2
ðp12p21
þ p22p11Þ þ 2q12½f 2x1x1
p11p12 þ f 2x2x2
p21p22 þ f 2x1x2
ðp12p21 þ p22p11Þ þ 2q13½f 3x1x1
p11p12 þ f 3x3x3
p31p32
þ f 3x3x1
ðp11p32 þ p12p31Þ;
F122 ¼ 2q11½f 1
x1x1p2
12 þ f 1x2x2
p222 þ f 1
x3x3p2
32 þ 2f 1x3x1
p12p32 þ 2f 1x1x2
p12p22þ 2q12½f 2
x1x1p2
12 þ f 2x2x2
p222 þ 2f 2
x1x2p12p22 þ 2q13
½f 3x1x1
p212 þ f 3
x3x3p2
32 þ f 3x3x1
p12p32Þ;F1
111 ¼ 6q11b11½f 1x1x1
p11p13 þ f 1x2x2
p21p23 þ f 1x3x3
p31p33 þ f 1x3x1
ðp11p33 þ p13p31Þ þ f 1x1x2
ðp11p23
þ p21p13Þ þ 6q12b11½f 2x1x1
p11p13 þ f 2x2x2
p21p23 þ f 2x3x3
p31p33
þ f 2x1x2
ðp11p23 þ p13p21Þ þ 6q13b11½f 3x1x1
p11p13 þ f 3x3x3
p31p33
þ f 3x3x1
ðp11p33 þ p13p31Þ;F1
122 ¼ 2q11½f 1x1x1
ð2p12p13b12 þ p11p13b22Þ þ f 1x2x2
ð2p22p33b12 þ p21p23b22Þ þ f 1x3x3
ð2p32p33b12 þ p31p33b22Þþ f 1
x3x1ð2p13p32b12 þ p11p33b22 þ p13p31b22 þ 2p12p33b12Þ
þ f 1x1x2
ð2p12p23 � b12 þ p13p21b22 þ p11p23b22 þ 2p22p13b12Þþ 2q12½f 2
x1x1ð2p12p13b12 þ p11p13b22Þ þ f 2
x2x2� ð2p22p23b12
þ p21p23b22Þ þ f 2x1x2
ð2p12p23b12 þ p11p23b22 þ 2p13p22b12
þ p13p21b22Þ þ 2q13½f 3x1x1
ð2p13p12b12 þ p11p13b22Þþ f 3
x3x3ð2p32p33b12 þ p31p33b22Þ þ f 3
x3x1
ð2p32p13b12 þ p31p13b22 þ 2p12p13b12 þ p33p11b22Þ;
F211 ¼ 2q21½f 1
x1x1p2
11 þ f 1x2x2
p221 þ f 1
x3x3p2
31 þ 2f 1x3x1
p11p31
þ 2f 1x1x2
p11p21 þ 2q22½f 2x1x1
p211 þ f 2
x2x2� p2
21
þ 2f 2x1x2
p11p21 þ 2q23½f 3x1x1
p211 þ f 3
x3x3p2
31
þ 2f 3x3x1
p11p31;F2
12 ¼ 2q21½f 1x1x1
p11p12 þ f 1x2x2
p21p22 þ f 1x3x3
p31p32
þ f 1x3x1
ðp11p32 þ p12p31Þ þ f 1x1x2
ðp12p21 þ p22p11Þþ 2q22½f 2
x1x1p11p12 þ f 2
x2x2p21p22 þ f 2
x1x2ðp12p21 þ p22p11Þ
þ 2q23½f 3x1x1
p11p12 þ f 3x3x3
p31p32 þ f 3x3x1
ðp11p32 þ p12p31Þ;F2
22 ¼ 2q21½f 1x1x1
p212 þ f 1
x2x2p2
22 þ f 1x3x3
p232 þ 2f 1
x3x1p12p32
þ 2f 1x1x2
p12p22 þ 2q22½f 2x1x1
p212 þ f 2
x2x2p2
22 þ 2f 2x1x2
p12p22þ 2q23½f 3
x1x1p2
12 þ f 3x3x3
p232 þ f 3
x3x1p12p32Þ;
F2112 ¼ 2q21½f 1
x1x1ð2p11p13b12 þ p12p13b11Þ þ f 1
x2x2ð2p21p23b12
þ p22p23b11Þ þ f 1x3x3
ð2p31p33b12 þ p32p33b11Þþ f 1
x3x1ð2p11p33b12 þ p13p32b11 þ p12p33b11
þ 2p13p31b12Þ þ f 1x1x2
ð2p13p21 � b12 þ p12p23b11
þ 2p11p23b12 þ p22p13b11Þ þ 2q22½f 2x1x1
ð2p11p13b12
þ p12p13b11Þ þ f 2x2x2
� ð2p21p23b12 þ p22p23b11Þþ f 2
x1x2ð2p11p23b12 þ p12p23b11 þ 2p13p21b12
þ p13p22b11Þ þ 2q23½f 3x1x1
ð2p11p13b12 þ p12p13b11Þþ f 3
x3x3ð2p31p33b12 þ p32p33b11Þ þ f 3
x3x1ð2p31p13b12
þ p33p12b11 þ 2p11p33b12 þ p32p13b11Þ;F2
222 ¼ 6q21b22½f 1x1x1
p12p13 þ f 1x2x2
p22p23 þ f 1x3x3
p32p33
þ f 1x3x1
ðp12p33 þ p13p32Þ þ f 1x1x2
ðp12p23 þ p13p22Þþ 6q22b22½f 2
x1x1p12p13 þ f 2
x2x2p22p23 þ f 2
x1x2ðp12p23
þ p13p22Þ þ 6q23b22½f 3x1x1
p12p13 þ f 3x3x3
p32p33
þ f 3x3x1
ðp12p33 þ p13p32Þ:
Global stability of the system (2.1) around E�
Theorem 6.1 The interior equilibrium E� is globally
asymptotically stable if the condition
ðiÞ a1a2b1b2rk[ ðx1� þ kÞðwþ kÞða1b1c2 þ a2b2c1Þ is
satisfied:
Proof Let
Lðx1; x2; x3Þ ¼ L1ðx1; x2; x3Þ þ L2ðx1; x2; x3Þ þ L3ðx1; x2; x3Þð6:16Þ
be a positive Lyapunov function, where
L1 ¼ s1
�x1 � x1� � x1� lnð x1
x1�Þ�; L2 ¼ s2
�x2 � x2� � x2� lnð x2
x2�Þ�;
L3 ¼ s3
�x3 � x3� � x3� lnð x3
x3�Þ�;
s1; s2 and s3 being positive real constants.
This function is well-defined and continuous in Int(Rþ3).
It can be easily verified that the function Lðx1; x2; x3Þ is
zero at the equilibrium point E� and is positive for all other
Model. Earth Syst. Environ. (2016) 2:106 Page 9 of 14 106
123

positive values of ðx1; x2; x3Þ; and thus E� is the global
minimum of Lðx1; x2; x3Þ.Since the solutions of the system are bounded and
ultimately enter the set X ¼ fðx1; x2; x3Þ; x1 [ 0; x2 [ 0;
x3 [ 0 : x1 þ x2
e1þ x3
e2�M þ �; 8�[ 0g, we restrict our
study in X. The time derivative of L along with the
solutions of the system (2.1) gives (cf. Sarwardi et al.
(2010), Sarwardi et al. (2012))
dL
dt¼� s1
h rk
ðx1� þ kÞðx1 þ kÞ �c1x2�
ða1 þ x1 þ b1x2Þða1 þ x1� þ b1x2�Þ
� c2x3�ða2 þ x1 þ b2x3Þða2 þ x1� þ b2x3�Þ
iðx1 � x1�Þ2
� s2
h c1e1ða1 þ b1Þða1 þ x1 þ b1x2Þða1 þ x1� þ b1x2�Þ
i
� ðx2 � x2�Þ2 � s3
h c2e2ða2 þ b2Þða2 þ x1 þ b2x3Þða2 þ x1� þ b2x3�Þ
i
ðx3 � x3�Þ2
þc1
�s2b1e1x2� � s1ða1 þ x1�Þ
�ðx1 � x1�Þðx2 � x2�Þ
ða1 þ x1 þ b1x2Þða1 þ x1� þ b1x2�Þ
þc2
�s3b2e2x3� � s1ða2 þ x1�Þ
�
ða2 þ x1 þ b2x2Þða2 þ x1� þ b2x2�Þ� ðx1 � x1�Þðx3 � x3�Þ:
ð6:17Þ
Letting s1 ¼ 1, s2 ¼ a1þx1�b1e1x2�Þ and s3 ¼ a2þx1�
b2e2x3�Þ ; we have
dL
dt� �
h rk
ðx1� þ kÞðx1 þ kÞ �c1x2�
ða1 þ x1 þ b1x2Þða1 þ x1� þ b1x2�Þ
� c2x3�ða2 þ x1 þ b2x3Þða2 þ x1� þ b2x3�Þ
iðx1 � x1�Þ2
\�h rk
ðx1� þ kÞðwþ kÞ �c1
a1b1
� c2
a2b2
iðx� x�Þ2
\0; by condition (i);
ð6:18Þ
along all the trajectories in the positive octant except
ðx1�; x2�; x3�Þ. Also dLdt¼ 0 when ðx1; x2; x3Þ ¼ ðx1�; x2�; x3�Þ.
The proof follows from (6.16) and Lyapunov-Lasalle’s
invariance principle (cf. Hale (1989)).
Numerical simulations
Numerical simulations have been carried out by making
use of MATLAB-R2010a and Maple-12. The analytical
findings of the present study are summarized and repre-
sented schematically in Tables 1 and 2. These results are all
verified by means of numerical illustrations of which some
chosen ones are shown in the figures. We took a set of
admissible parameter values: r ¼ 1:7; k ¼ 200; a1 ¼ a2 ¼100; b1 ¼ b2 ¼ 0:5; c1 ¼ c2 ¼ 1:8; d1 ¼ 0:82; d2 ¼ 0:62;
e1 ¼ 0:8143; e2 ¼ 0:6250: For this set of parameter val-
ues, the system possessed an unique interior equilib-
rium pointE� ¼ ð169:1663564; 55:36073780; 62:98120968Þ:
The system parameter r is the growth rate of the prey pop-
ulation. It plays a crucial role in regulating the dynamical
behaviour of the proposed system. For this reason, we try to
determine the system’s possible outcomes by varying this
parameter within its feasible range. The interior equilibrium
E� is stable for r[ rc ¼ 1:320961640 (cf. Figs. 4, 5 for local
stability and Fig. 6 for global stability). The system (2.1)
experiences a Hopf bifurcation when the parameter r crosses
the critical value rc from left to right, i.e., when r ¼ rc, all the
species coexist oscillating. Following the steps discussed in
Subsect. 6.1, we have found the value of P ¼1:042431405[ 0; which indicates that the obtained Hopf
bifurcation is a subcritical bifurcation (cf. Fig. 7).
It is observed that, if the interference coefficient b1
(interference effect on the first predator due to the pres-
ence of the second predator) increases it stabilizes the
system for b1 ¼ 0:7; while it is unstable at b1 ¼ 0:6:
When the parameter b1 exceeds its value 20, the first
predator population disappears from the system. Simi-
larly, the interference on the second predator due to the
presence of first predator, parameterized by b2 also plays
an important role to stabilize the system. If the interfer-
ence coefficient b2 increases it stabilizes the system for
b2 ¼ 0:6; while it is unstable at b2 ¼ 0:5: It also regulates
the existence of second predator in the system. As the
parameter b2 exceeds 20.9, the second predator population
gets extinguished.
Analogously, if the parameter d1; denoting the death rate
of the first predator increases then the size of the first
predator decreases as well as the second predator popula-
tion increases. If d1 decreases, the first predator population
increases and second predator population decreases. If the
death rate d1 is gradually increased to a certain level the
first predator population goes to extinction. A similar result
is observed for the case of the second predator’s death rate.
The above observations ensure that the model under con-
sideration is consistent with biological observations
(Figures are not reported here).
Concluding remarks
The problem described by the system (2.1) is well posed.
The x1; x2 and x3 axes are invariant under the flow of the
system. To our knowledge this is the first attempt to study
an ecological system with quasi-linear/bilinear growth of
the prey population. Generally, researchers only studied
biological model systems with logistic/linear growth of
prey population. One of the important results is that the
prey population becomes unbounded in absence of its
admissible predator in the long run. But in the presence of
predator the prey population can be bounded under
106 Page 10 of 14 Model. Earth Syst. Environ. (2016) 2:106
123

suitable combinations of system parameters. As a conse-
quence the total environmental population is bounded
above (cf. Subsect. 3.3). Therefore, any solution starting in
the interior of the first octant never leaves it. This
mathematical fact is consistent with the biological inter-
pretation of the system. Due to the inclusion of quasi-lin-
ear/bilinear growth of the prey population, the axial
equilibrium point is driven away by the system, a fact
Table 2 The set of system parameter (including the critical parameter rc) values and their corresponding Figures with description
No. Fixed parameters r Figures Description
1 r ¼ 1:37[ rc ¼ 1:320961640;
k ¼ 200; a1 ¼ 100; a2 ¼ 100;
b1 ¼ 0:5; b2 ¼ 0:5; c1 ¼ 1:8;
c2 ¼ 1:8; d1 ¼ 0:82; d2 ¼ 0:62;
e1 ¼ 0:8143; e2 ¼ 0:6250:
1.37 Fig. 1: (a)-(b) 2D view of Hopf
bifurcation
2 . . .. . .. . .. . .. . .. . .. . . 1.320961640 Fig. 2 Limit cycle
3 . . .. . .. . .. . .. . .. . .. . . r 2 ½0:8; 2:0 Fig. 3 Hopf bifurcation
(growth rate r vs.
population volumes)
4 . . .. . .. . .. . .. . .. . .. . . 1.4700000000 Fig. 4 2D view of local stability
5 . . .. . .. . .. . .. . .. . .. . . 1.4700000000 Fig. 5 3D view of local stability
6 . . .. . .. . .. . .. . .. . .. . . 1.4700000000 Fig. 6 Global stability
7
r ¼ 1:37; k ¼ 200;
a1 ¼ 100; a2 ¼ 100;
b1 ¼ 0:5; b2 ¼ 0:5;
c1 ¼ 1:8; c2 ¼ 1:8;
d1 ¼ 0:82; d2 ¼ 0:62;
e1 ¼ 0:8143; e2 ¼ 0:6250
rc ¼ 1:320961640;P ¼ 1:0424314050
Fig. 7 Subcritical Hopf bifurcation
Table 1 Schematic representation of our analytical findings
Equilibria Feasibility conditions/parametric restrictions Stability conditions/parametric restrictions Nature
E0 No conditions No conditions Unstable
E1 c1e1 [ d1, x11[ b1d1
c1e1�d1e1x11
þ x21[ kð1�b1e1Þ�a1
b1
LAS
E2 c2e2 [ d2, x12[ b2d2
c2e2�d2e2x12
þ x32[ kð1�b2e2Þ�a2
b2
LAS
E� x1� [ max a1d1
c1e1�d1; a2d2
c2e2�d2
n ok\min fa1 þ b1x2�; a2 þ b2x3�g LAS
E� .....................
ðiÞ r[ d1 þ d2; ðiiÞx11[
a2d2
c2e2 � d2
; ðiiiÞ x12[
a1d1
c1e1 � d1
Persistence
E� ..................... Stated in the Proposition 3.3 Boundedness
E� ..................... P[ 0 (cf. equation (6.15)) SHB
E� x1� [ max a1d1
c1e1�d1; a2d2
c2e2�d2
n oa1a2b1b2rk[ ðx1� þ kÞðwþ kÞ � ða1b1c2 þ a2b2c1Þ GAS
LAS locally asymptotically stable, GAS globally asymptotically stable, HB Hopf bifurcation, SHB subcritical Hopf bifurcation
Model. Earth Syst. Environ. (2016) 2:106 Page 11 of 14 106
123
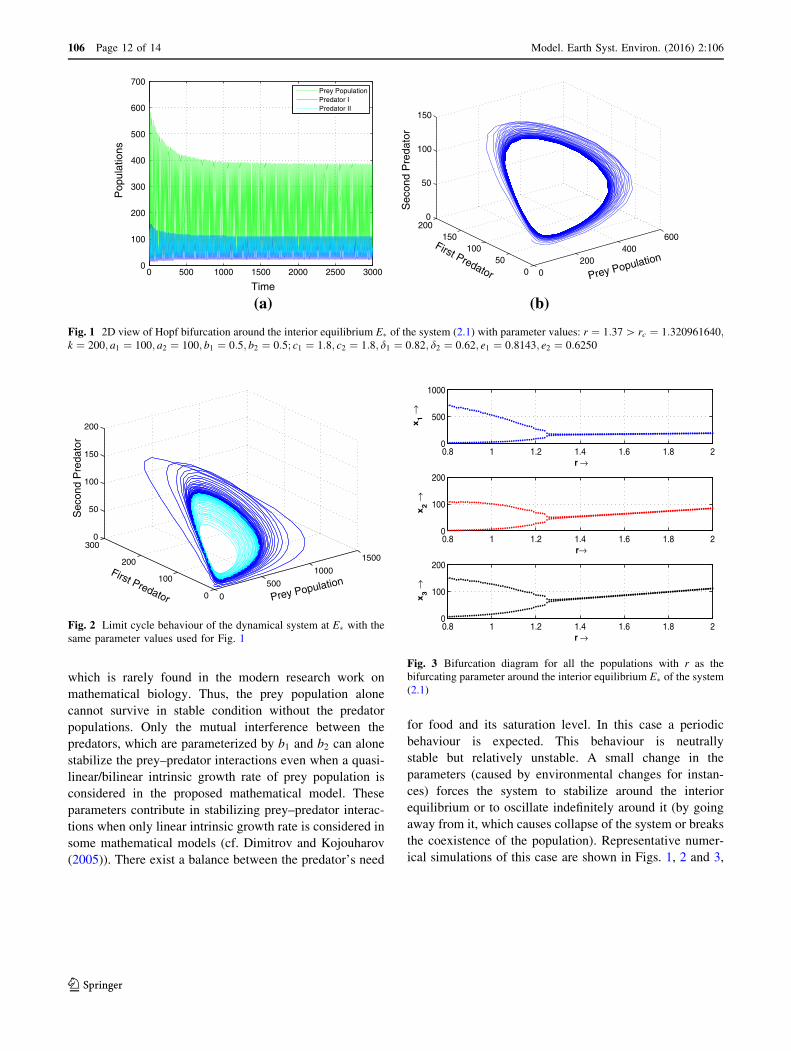
which is rarely found in the modern research work on
mathematical biology. Thus, the prey population alone
cannot survive in stable condition without the predator
populations. Only the mutual interference between the
predators, which are parameterized by b1 and b2 can alone
stabilize the prey–predator interactions even when a quasi-
linear/bilinear intrinsic growth rate of prey population is
considered in the proposed mathematical model. These
parameters contribute in stabilizing prey–predator interac-
tions when only linear intrinsic growth rate is considered in
some mathematical models (cf. Dimitrov and Kojouharov
(2005)). There exist a balance between the predator’s need
for food and its saturation level. In this case a periodic
behaviour is expected. This behaviour is neutrally
stable but relatively unstable. A small change in the
parameters (caused by environmental changes for instan-
ces) forces the system to stabilize around the interior
equilibrium or to oscillate indefinitely around it (by going
away from it, which causes collapse of the system or breaks
the coexistence of the population). Representative numer-
ical simulations of this case are shown in Figs. 1, 2 and 3,
(a)
0 500 1000 1500 2000 2500 30000
100
200
300
400
500
600
700
Time
Pop
ulat
ions
Prey PopulationPredator IPredator II
(b)
0200
400600
050
100150
2000
50
100
150
Prey PopulationFirst Predator
Sec
ond
Pre
dato
r
Fig. 1 2D view of Hopf bifurcation around the interior equilibrium E� of the system (2.1) with parameter values: r ¼ 1:37[ rc ¼ 1:320961640;k ¼ 200; a1 ¼ 100; a2 ¼ 100; b1 ¼ 0:5; b2 ¼ 0:5; c1 ¼ 1:8; c2 ¼ 1:8; d1 ¼ 0:82; d2 ¼ 0:62; e1 ¼ 0:8143; e2 ¼ 0:6250
0
500
1000
1500
0
100
200
3000
50
100
150
200
Prey PopulationFirst Predator
Sec
ond
Pre
dato
r
Fig. 2 Limit cycle behaviour of the dynamical system at E� with the
same parameter values used for Fig. 1
0.8 1 1.2 1.4 1.6 1.8 20
500
1000
r →
x1
→
0.8 1 1.2 1.4 1.6 1.8 20
100
200
r→
x2
→
0.8 1 1.2 1.4 1.6 1.8 20
100
200
r →
x3
→
Fig. 3 Bifurcation diagram for all the populations with r as the
bifurcating parameter around the interior equilibrium E� of the system
(2.1)
106 Page 12 of 14 Model. Earth Syst. Environ. (2016) 2:106
123
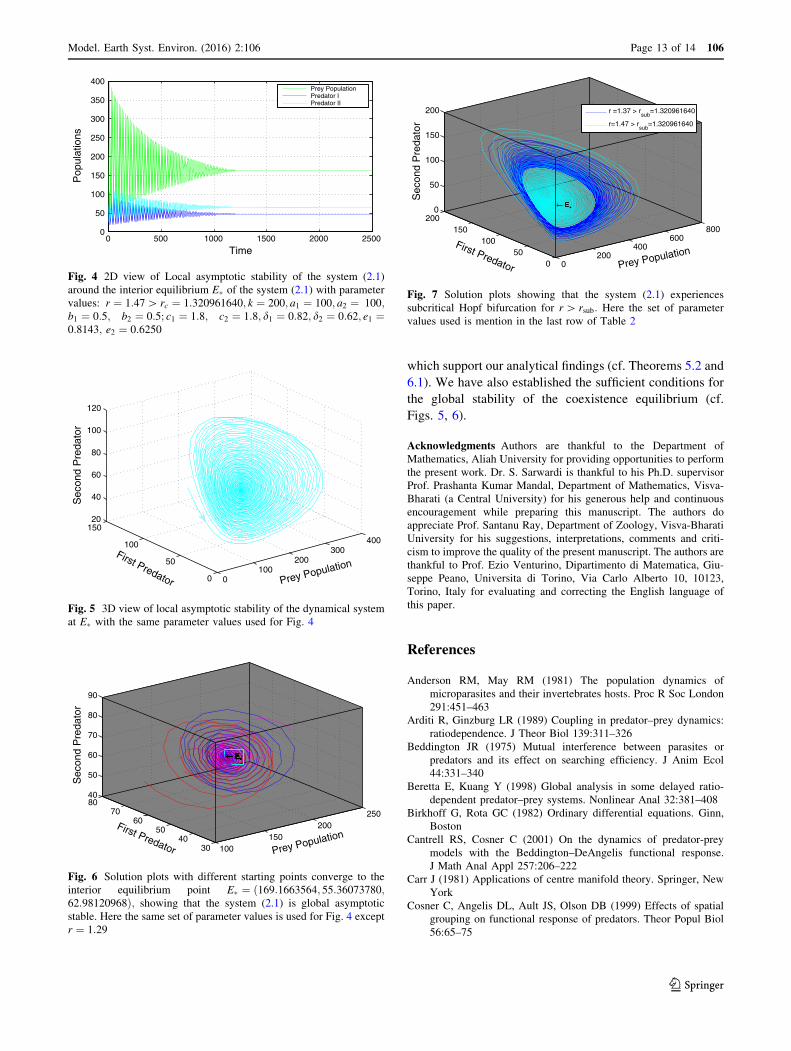
which support our analytical findings (cf. Theorems 5.2 and
6.1). We have also established the sufficient conditions for
the global stability of the coexistence equilibrium (cf.
Figs. 5, 6).
Acknowledgments Authors are thankful to the Department of
Mathematics, Aliah University for providing opportunities to perform
the present work. Dr. S. Sarwardi is thankful to his Ph.D. supervisor
Prof. Prashanta Kumar Mandal, Department of Mathematics, Visva-
Bharati (a Central University) for his generous help and continuous
encouragement while preparing this manuscript. The authors do
appreciate Prof. Santanu Ray, Department of Zoology, Visva-Bharati
University for his suggestions, interpretations, comments and criti-
cism to improve the quality of the present manuscript. The authors are
thankful to Prof. Ezio Venturino, Dipartimento di Matematica, Giu-
seppe Peano, Universita di Torino, Via Carlo Alberto 10, 10123,
Torino, Italy for evaluating and correcting the English language of
this paper.
References
Anderson RM, May RM (1981) The population dynamics of
microparasites and their invertebrates hosts. Proc R Soc London
291:451–463
Arditi R, Ginzburg LR (1989) Coupling in predator–prey dynamics:
ratiodependence. J Theor Biol 139:311–326
Beddington JR (1975) Mutual interference between parasites or
predators and its effect on searching efficiency. J Anim Ecol
44:331–340
Beretta E, Kuang Y (1998) Global analysis in some delayed ratio-
dependent predator–prey systems. Nonlinear Anal 32:381–408
Birkhoff G, Rota GC (1982) Ordinary differential equations. Ginn,
Boston
Cantrell RS, Cosner C (2001) On the dynamics of predator-prey
models with the Beddington–DeAngelis functional response.
J Math Anal Appl 257:206–222
Carr J (1981) Applications of centre manifold theory. Springer, New
York
Cosner C, Angelis DL, Ault JS, Olson DB (1999) Effects of spatial
grouping on functional response of predators. Theor Popul Biol
56:65–75
0 500 1000 1500 2000 25000
50
100
150
200
250
300
350
400
Time
Pop
ulat
ions
Prey PopulationPredator IPredator II
Fig. 4 2D view of Local asymptotic stability of the system (2.1)
around the interior equilibrium E� of the system (2.1) with parameter
values: r ¼ 1:47[ rc ¼ 1:320961640; k ¼ 200; a1 ¼ 100; a2 ¼ 100;b1 ¼ 0:5; b2 ¼ 0:5; c1 ¼ 1:8; c2 ¼ 1:8; d1 ¼ 0:82; d2 ¼ 0:62; e1 ¼0:8143; e2 ¼ 0:6250
0100
200300
400
0
50
100
15020
40
60
80
100
120
Prey PopulationFirst Predator
Sec
ond
Pre
dato
r
Fig. 5 3D view of local asymptotic stability of the dynamical system
at E� with the same parameter values used for Fig. 4
100150
200250
3040
5060
708040
50
60
70
80
90
Prey Population
← E*
← E*
← E*
← E*
← E*
← E*
← E*
← E*
First Predator
Sec
ond
Pre
dato
r
Fig. 6 Solution plots with different starting points converge to the
interior equilibrium point E� ¼ ð169:1663564; 55:36073780;62:98120968Þ; showing that the system (2.1) is global asymptotic
stable. Here the same set of parameter values is used for Fig. 4 except
r ¼ 1:29
0200
400600
800
050
100150
2000
50
100
150
200
Prey Population
← E*
← E*
First Predator
Sec
ond
Pre
dato
r
r =1.37 > rsub
=1.320961640
r=1.47 > rsub
=1.320961640
Fig. 7 Solution plots showing that the system (2.1) experiences
subcritical Hopf bifurcation for r[ rsub: Here the set of parameter
values used is mention in the last row of Table 2
Model. Earth Syst. Environ. (2016) 2:106 Page 13 of 14 106
123

Cui J, Takeuchi Y (2006) Permanence, extinction and periodic
solution of predator–prey system with Beddington–DeAngelis
functional response. J Math Anal Appl 317:464–474
Curds CR, Cockburn A (1968) Studies on the growth and feeding of
Tetrahymena pyriformis in axenic and monoxenic culture. J Gen
Microbiol 54:343–358
DeAngelis RA, Goldstein RA, Neill R (1975) Ecology 56:881–892
Dimitrov DT, Kojouharov HV (2005) Complete mathematical
analysis of predator–prey models with linear prey growth and
Beddington–DeAngelis functional response. Appl Math Comp
162:523–538
Freedman HI (1990) A model of predator–prey dynamics modified by
the action of parasite. Math Biosci 99:143–155
Gard TC, Hallam TG (1979) Persistence in Food web-1, Lotka-
Volterra food chains. Bull Math Biol 41:877–891
Hadeler KP, Freedman HI (1989) Predator–prey populations with
parasitic infection. J Math Biol 27:609–631
Hale JK (1989) Ordinary differential equations. Krieger Publisher
Company, Malabar
Haque M, Venturino E (2006) Increase of the prey may decrease the
healthy predator population in presence of a disease in the
predator. Hermis 7:39–60
Haque M, Venturino E (2006) The role of transmissible diseases in
the Holling–Tanner predator–prey model. Theor Popul Biol
70:273–288
Hassell MP, Varley GC (1969) New inductive population model for
insect parasites and its bearing on biological control. Nature
223:1133–1137
Hethcote HW, Wang W, Ma Z (2004) A predator–prey model with
infected prey. Theor Popul Biol 66:259–268
Huo HF, Li WT, Nieto JJ (2007) Periodic solutions of delayed
predator–prey model with the Beddington–DeAngelis functional
response. Chaos Solitons Fractals 33:505–512
Hwang TW (2003) Global analysis of the predator–prey system with
Beddington–DeAngelis functional response. J Math Anal Appl
281:395–401
Kar TK, Gorai A, Jana S (2012) Dynamics of pest and its predator
model with disease in the pest and optimal use of pesticide.
J Theor Biol 310:187–198
Ma WB, Takeuchi Y (1998) Stability analysis on predator–prey
system with distributed delays. J Comput Appl Math 88:79–94
Salt GW (1974) Predator and prey densities as controls of the rate of
capture by the predator Didinium nasutum. Ecology 55:434–439
Sarwardi S, Haque M, Venturino E (2010) Global stability and
persistence in LG-Holling type-II diseased predators ecosystems.
J Biol Phys 37:91–106
Sarwardi S, Mandal PK, Ray S (2012) Analysis of a competitive
prey–predator system with a prey refuge. Biosystems
110:133–148
Sarwardi S, Mandal PK, Ray S (2013) Dynamical behaviour of a two-
predator model with prey refuge. J Biol Phys 39:701–722
Venturino E (1995) Epidemics in predator–prey models: disease in
prey, in mathematical population dynamics. Analysis of hetero-
geneity 1. In: Arino O, Axelrod D, Kimmel M, Langlais M (eds),
pp 381–393
Wiggins S (2003) Introduction to applied nonlinear dynamical
systems and Chaos, 2nd edn. Springer, New York
Xiao Y, Chen L (2001) Modeling and analysis of a predator–prey
model with disease in prey. Math Biosci 171:59–82
106 Page 14 of 14 Model. Earth Syst. Environ. (2016) 2:106
123