dracgap, a novel drosophila gene, inhibits egfr/ras...
TRANSCRIPT

INTRODUCTION
The development of multicellular organisms depends, to alarge extent, on external stimuli controlling a variety of cellularprocesses, such as cell growth and proliferation, cell survival,and cell determination and differentiation. These stimuli, in theform of extracellular ligands, interact with and activate orrepress transmembrane receptor proteins. The Ras superfamilyof small GTPases, which includes among others the Ras andRho families, links receptor stimulation to activation ofcytosolic protein kinase cascades which, in turn, orchestratecytoplasmic and nuclear events (Bourne et al., 1990).
The mammalian Rho-family comprises at least 14 differentproteins; the best characterised are RhoA, Rac1 and Cdc42Hs(reviewed by Van Aelst and D’Souza-Schorey, 1997). Theseproteins act as signal transducers by switching betweeninactive, GDP-bound and active, GTP-bound forms. Theiractivity is regulated by the opposing effects of guaninenucleotide exchange factors (GEF), positive regulators thatenhance the exchange of bound GDP for GTP, and GTPase-activating proteins (GAP), negative regulators that stimulatethe intrinsically low GTPase activity of the Rho proteins. Theyare further regulated by guanine-nucleotide dissociationinhibitor proteins which stabilise the bound GTP. RhoGEFsand RhoGAPs, by analogy to RasGEFs and RasGAPs, arethought to be the immediate targets of activated receptors(Boguski and McCormick, 1993).
Mammalian Rho-family GTPases regulate cell proliferationand also many actin-based cellular processes such as cell
motility, axonal growth and guidance, formation of cell adhesioncomplexes, intracellular membrane trafficking and cytokinesis(reviewed by Hall, 1998). They participate in transcriptionalregulation, mainly through activation of the c-Jun NH2-terminalkinase (JNK) and p38 mitogen-activated (MAP) kinase cascades(Van Aelst and D’Souza-Schorey, 1997).
Homologues to the Rho-family GTPases and their regulatorshave been identified in D. melanogaster(DRhoA, DRhoL,DRac1, 2 and 3, DCdc42, DRhoGEF1and 2, pebble andRotundRacGAP(RnRacGAP), reviewed by Van Aelst andD’Souza-Schorey, 1997). This has facilitated the geneticanalysis of their physiological roles. In accordance with theirfunction in reorganising the actin cytoskeleton, DrosophilaRho-family proteins control growth cone guidance (reviewedby Luo et al., 1997), myogenesis (Luo et al., 1994),remodelling of synaptic terminals (Sone et al., 1997) andcytokinesis (Prokopenko et al., 1999). They also participate ingastrulation (Barrett et al., 1997; Häcker and Perrimon, 1998),embryonic segmentation (Magie et al., 1999) and cellmigration (reviewed by Montell, 1999). Epithelial planarpolarity (EPP) and embryonic dorsal closure depend both onactin cytoskeleton reorganisation and JNK activation, whichare controlled by Rho-family proteins (reviewed by Eaton,1997; Noselli and Agnès, 1999). Only one putative RacGAP,RnRacGAP, has been identified in Drosophila (Agnel et al.,1992).Its ectopic expression in cellularising embryos disruptsthe actin cytoskeleton and its overexpression in pupae slightlyalters the pattern of wing veins, wing shape and EPP (Guichardet al., 1997).
5427Development 127, 5427-5438 (2000)Printed in Great Britain © The Company of Biologists Limited 2000DEV7830
We have identified a novel Drosophilagene, DRacGAP,which behaves as a negative regulator of Rho-familyGTPases DRac1 and DCdc42. Reduced function ofDRacGAP or increased expression ofDRac1 in the wingimaginal disc cause similar effects on vein and sensoryorgan development and cell proliferation. These effectsresult from enhanced activity of the EGFR/Ras signallingpathway. We find that in the wing disc, DRac1 enhancesEGFR/Ras-dependent activation of MAP Kinase in theprospective veins. Interestingly, DRacGAP expression isnegatively regulated by the EGFR/Ras pathway in theseregions. During vein formation, local DRacGAP repression
would ensure maximal activity of Rac and, in turn, ofRas pathways in vein territories. Additionally, maximalexpression of DRacGAPat the vein/intervein boundarieswould help to refine the width of the veins. Hence, controlof DRacGAP expression by the EGFR/Ras pathway is apreviously undescribed feedback mechanism modulatingthe intensity and/or duration of its signalling duringDrosophiladevelopment.
Key words: RacGAP, Rac GTPase, Signal transduction, Apoptosis,EGFR, Drosophila melanogaster, Imaginal disc
SUMMARY
DRacGAP, a novel Drosophila gene, inhibits EGFR/Ras signalling in the
developing imaginal wing disc
Sol Sotillos and Sonsoles Campuzano*
Centro de Biología Molecular Severo Ochoa, CSIC and UAM Cantoblanco, 28049 Madrid, Spain*Author for correspondence (e-mail: [email protected])
Accepted 5 October; published on WWW 14 November 2000

5428
Initially, each of the components of the Ras superfamilywere thought to control different, independent regulatorypathways. However, nowadays there is compelling evidence ofcrosstalks among their different signalling pathways (Schwartzand Baron, 1999). For example, in mammalian cells in culture,Ras and Rho-family proteins cooperate in the control of cellproliferation. Thus, activated forms of Rho or Rac synergisewith Raf, a downstream kinase of Ras, in focus-formationassays, and dominant negative forms of Rho, Rac and Cdc42inhibit cell-transformation caused by oncogenic Ras (reviewedby Van Aelst and D’Souza-Schorey, 1997). Here we present,to our knowledge for the first time, evidence for thecooperation of Rac and Ras signalling pathways in the contextof a whole organism. We describe a novel Drosophila gene,DRacGAP, which encodes a putative GAP for Rac and Cdc42GTPases. We show that both DRacGAP and DRac1 areinvolved in the control of cell proliferation. Moreover, reducedactivity of DRacGAPor overexpression of DRac1in the wingimaginal disc cause similar defects: widening of veins,development of extra sensory organs (SOs), apoptosis andappearance of enlarged cells that differentiate multiple hairswith abnormal polarity. We show that these phenotypes resultfrom DRac1 enhancement of epidermal growth factor receptor(EGFR)/Ras signalling. This conserved pathway, whichoperates through activation of the Ras/Raf/MEK/MAP kinasecascade (Brunner et al., 1994; Díaz-Benjumea and Hafen,1994) controls multiple developmental processes (Rommel andHafen, 1998; Hackel et al., 1999) and is accurately regulated.Indeed, this pathway controls the expression of its ownnegative and positive regulators (Wasserman and Freeman,1998; Martín-Blanco et al., 1999; Perrimon and McMahon,1999). Interestingly, expression of DRacGAPis repressed byEGFR/Ras signalling in the prospective veins and accumulatesat the vein/intervein boundaries. These results suggest thatcontrol of DRacGAPexpression by the EGFR/Ras pathwayprovides a new mechanism to modulate the intensity of thispathway during Drosophiladevelopment.
MATERIALS AND METHODS
Drosophila stocksThe strains used in this study are: Df(2R)CX1, which uncovers theDRacGAP gene; Df(3R)dsx10M, which uncovers the RnRacGAP gene;Rho GTPase pathway alleles and transgenes: RhoAand UAS-DRho(Strutt et al., 1997); UAS-DRac1, UAS-DRac1N17, UAS-Dcdc42N17andUAS-Dcdc42V12(Luo et al., 1994), Cdc423 (Fehon et al, 1997), DRho-GEF2 (PZ 04291, Häcker and Perrimon, 1998), Pak4 (Hing et al,1999) and PknP (PZ 06736,Lu and Settleman, 1999); EGFR pathwayalleles and transgenes (veinlet7M (ve7M), vein1 (vn), UAS-RasV12
(Karim and Rubin, 1998), UAS-RafDN3.1(Martín-Blanco et al., 1999);UAS-rolledSem (Martín-Blanco, 1998), UAS-argos (UAS-aos andaossty; a gift from M. Freeman) and UAS-p35 (Hay et al., 1994), UAS-p21 (Karim and Rubin, 1998), UAS-CyclinE(Weigmann et al., 1997),UAS-string (Neufeld et al., 1998);Act5CFRTDraf+FRTnuc-lacZ(Struhl and Basler, 1993); y ActFRTcdc2FRTGal4, (a gift fromE. Sánchez-Herrero), neuralized-lacZ(A101-IF3), engrailed-Gal4(en-G4), Scalloped-Gal4 (Sd-G4), 69B-Gal4, MS1096-Gal4,dppdisk-Gal4 and C765-Gal4. They are described in FlyBase(http://gin.ebi.ac.ak:7081). Transgenic lines UAS-DRacGAP, UAS-DRacGAP∆EIE and UAS-DRacGAPR417Q were generated bysubcloning wild-type DRacGAPcDNA or the mutated forms (seebelow) in the pUAST vector (Brand and Perrimon, 1993). The
resulting pUAST-plasmids were used to transform w1118 embryos(Ashburner, 1989).
Overexpression experimentsDifferent UAS transgenic lines were crossed with several Gal4 linesand larvae were raised at 18°C, 25°C or 29°C. An en-G4/SM6a-TM6b/UAS-DRacGAP∆EIE stock was maintained at 18°C and used totest modifications of the DRacGAPDN phenotype in different geneticbackgrounds.
Overexpression of UAS transgenes was also carried out by the flip-out method (Struhl and Basler, 1993) in larvae of the genotype y whs-FLP/y; ActFRTcdc2FRTGal4; UAS-y+/UAS-DRacGAPDN.Expression of the Flipase(FLP) gene was induced at first, second andearly third instar larvae by 1 hour incubation at 37°C. Clones weredistinguishable as y+.
Molecular characterisation of DRacGAPA 2.3 kb cDNA containing the entire DRacGAPORF was fortuitouslyisolated from a DrosophilacDNA imaginal disc library (provided byJ. Botas) as a cloning artefact joined to a kuzbaniancDNA clone(Sotillos et al., 1997) and subcloned into pBluescript as an EcoRIfragment. The isolated DRacGAPcDNA was full length as its sizecoincides with that of DRacGAPmRNA detected by northern blot(not shown). cDNA was sequenced with the ABI Taq cyclesequencing system and analysed with the University of WisconsinGCG software package (Devereux et al., 1984). GenBank accessionnumber of DRacGAPis AJ251502.
Comparison of DRacGAPcDNA and corresponding genomicsequences, contained in bacteriophage P1DS04160 (mapped at 50C6-8, Berkeley Drosophila Genome Project), revealed that the DRacGAPgene contains 4 exons separated by three introns of 69, 63 and 513nucleotides.
Site directed mutagenesis A 0.2 kb SacII-EcoRV from DRacGAPcDNA was subcloned inpBluescript and used as template for inverted PCR-mutagenesisessentially as described by Hemsley et al. (1989). Oligonucleotidesused to generate the EIE deletion were GCCCGCGGCCTGACCGAand ATTTACGCAGTGCACAATCAGTG and those used for sitedirected mutagenesis of arg417 were CAGTTGTTCCTCGTCGGA-GCG and ATAGAGGCCAACCTCGGTCAG. The arg codon wasreplaced by the sequence shown in bold. The mutated fragments wereexcised from pBluescript and inserted in place of the wild-typefragments in subclones of the DRacGAPcDNA in pGEM1. Mutationswere confirmed by sequencing.
In situ hybridisation and immunocytochemistryIn situ hybridisation with digoxigenin-labelled probes, combined withX-gal staining, anti-β galactosidase (Promega) and phalloidin(Molecular Probes) stainings are described by Cubas et al. (1991),TUNEL labelling by Milán et al. (1997), staining with anti-doublephosphorylated-ERK (dp-ERK, Sigma), to determine the pattern ofactivated MAPK, by Martín-Blanco et al. (1999) and in situhybridisation to polythene chromosomes by Ashburner (1989).
Generation of β-galactosidase expressing clones y w hs-FLP; en-G4/SM6a-TM6b/UAS-DRacGAP∆EIE females werecrossed with Act5CFRTDraf+FRTnuc-lacZmales (Struhl and Basler,1993). Larvae were heat-shocked for 7 minutes at 37°C at 24-48 hoursafter egg laying to induce expression of the FLP gene. After 3 daysat 25°C, lacZ-expressing cells in the wing discs were identified byanti-β-galactosidase staining.
Other methodsBasic molecular biology techniques were carried out as described bySambrook et al. (1989). Statistical analyses were those described byZar (1999).
S. Sotillos and S. Campuzano

5429DRacGAP inhibits EGFR/Ras pathway
RESULTS
Molecular characterisation of the DRacGAP geneWe have isolated a D. melanogaster cDNA clone that encodesa protein of 625 amino acids whose sequence is closely relatedto RacGAPs from different organisms (Fig. 1A). This proteincontains a domain (residues 395-537) including the threeconserved amino acid blocks and the arginine finger that definethe GAP domain in GAPs for Rho-family GTPases (Ahmed etal., 1994; Scheffzek et al., 1998). It also presents two motifsusually found in these proteins (Lamarche and Hall, 1994) – aproline rich region, putative recognition motif for SH3-domaincontaining proteins, and a cysteine-rich domain, similar tothe diacylglycerol-binding domain of protein kinase C. Theputative GAP domain is most similar to those of DrosophilaRnRacGAP (Agnel et al., 1992), a mouse RacGAP(Wooltorton et al., 1999) and human n-chimaerin (Ahmedet al., 1994), which is a GAP specific for Rac GTPase(Dieckmann et al., 1991). Accordingly, and also based in thegenetic interactions described below, we have named this geneDRacGAP. The DRacGAPgene was mapped to cytologicalregion 50C4-8 by in situ hybridisation (not shown).
Expression pattern of DRacGAPDRacGAP gives rise to a unique transcript of 2.3 kb presentthroughout development (not shown). The expression pattern ofDRacGAP is highly dynamic. It is ubiquitous during the initialstages of embryogenesis and becomes restricted, after germ bandretraction, to the central and peripheral nervous system (notshown). In the early second instar wing imaginal disc, DRacGAPexpression occurs mostly in the presumptive wing region (Fig.1B). Afterwards, DRacGAP mRNA accumulation is widespreadin all imaginal discs (Fig. 1C and not shown). At late third instarDRacGAPis expressed in the eye-antenna disc in two stripes ofcells located at, and ahead of, the morphogenetic furrow and inseveral rings of cells in the presumptive antenna (Fig. 1D). Inthe wing disc, DRacGAPmRNA accumulates in the presumptiveinterveins (Fig. 1E), where it persists in pupae, being stronger atthe vein/intervein boundaries (Fig. 1G).
Analysis of DRacGAP function in the wing imaginaldiscThe pattern of expression of DRacGAPin third instar wing
discs prompted us to examine its role in vein patterning. Sincethere are no DRacGAPmutants available, we used theGal4/UAS system (Brand and Perrimon, 1993). First, weestablished transgenic flies carrying wild-type DRacGAPunder the control of the UAS element (UAS-DRacGAPlines).Overexpression of DRacGAPin the wing disc interfered withvein development. Thus, overexpression driven by the Gal4lines MS1096or C765eliminated the corrugation of the adultveins (Fig. 2A,B and not shown). Overexpression driven by en-G4 reduced the expression of a vein marker gene in the wingdisc (Fig. 2D) and resulted in thinning of adult veins (notshown). A reduction in wing size and a decrease in the numberof SOs were also found (Fig. 2B and Table 1).
To investigate whether a reduction in DRacGAPfunctionwould cause reciprocal effects, we generated two types oftransgenic flies expressing dominant negative (DN) forms ofDRacGAP under the control of UAS. In one of them, the EIEmotif (amino acids 405-407) was deleted (UAS-DRacGAP∆EIE
line). Removal of the corresponding amino acids in human n-chimaerin abolishes its RacGAP activity without impairing itsability to bind to Rac (Ahmed et al., 1994). In the other DNform, the conserved arginine residue of the arginine finger(arg417) was replaced by glutamine (UAS-DRacGAPR417Q line).The homologous arginine residues of RasGAP and RhoGAParginine fingers are essential for their ability to increase thehydrolysis of GTP bound to the corresponding GTPase(Scheffzek et al., 1998). Hence, both DN proteins shouldcompete with the wild-type protein for binding to the GTPaseand, when bound to it, should maintain it in the GTP-boundactive state, effectively mimicking a reduction in DRacGAPfunction.
Wings of flies expressing UAS-DRacGAP∆EIE in the wingdisc driven by en-G4 (referred to hereafter in the text asDRacGAPDN flies) displayed loss of wing tissue (wingnotching), veins of increased width, fusion of adjacent veinsand appearance of vein material connecting neighbouring veins(Fig. 2H). In addition, they had extra SOs along the vein L3and in the proximal regions of veins L4 and L5 (Fig. 2J, andTable 1). DRacGAPDN wings showed enlarged cells (indicatedby low trichome density, Fig. 2H) and alteration in the numberand polarity of the trichomes (Fig. 2I). The enlarged cells werealso visible in the imaginal disc (phalloidin staining, whichlabels cortical actin; Fig. 2K,L) indicating the early onset of
Table 1. Quantitative analysis of the genetic interactions Wing phenotype (%)
Genotype Weak Intermediate Strong Number of sensilla n T
MS1096, UAS-DRacGAP/+ n.d. 1.4±0.16 10 29en-G4/+, UAS-DRacGAPR417Q/+ 20 40 40 5 10 25en-G4/+; UAS-DRacGAP∆EIE/+ − 30 70 5 36 25en-G4/+; UAS-DRacGAP∆EIE/+ − 17 83 9±1.1*,§ 12 29en-G4/UAS-DRac n.d. 7.7±0.48‡ 12 18en-G4/UAS-DRac; UAS-DRacGAP∆EIE/+ n.d. 9.8±0.62‡ 22 18Cdc423/+, en-G4/+; UAS-DRacGAP∆EIE/+ − 85 15 5.45±0.1§ 20 29en-G4/+; UAS-DRacGAP∆EIE/Pak4 − 86 14 n.d. 28 25en-G4/+; UAS-DRACGAP∆EIE/ve7Mvn − 92 8 n.d. 12 25en-G4/+; UAS-DRACGAP∆EIE/aossty n.d. 11.9±0.7* 22 29
Wing phenotypes were classified as weak (presence of enlarged cells with polarity defects, as shown in Fig. 2G), intermediate (showing in addition widenedveins) and strong (with additional wing notching, as shown in Fig. 2H). Number of sensilla are the average number of campaniform sensilla (in the L3, L4 and L5veins and anterior crossvein)±s.e.m. Wild-type wings only show four campaniform sensilla on the L3 vein and one on the anterior crossvein. n, number of wingsexamined; T, temperature (°C); n.d., not determined. (*,‡,§) Differences between the indicated values are statistically significant (P<0.05 in all comparisons;Mann-Whitney U-Test).

5430
this defect. Overexpression of UAS-DRacGAPR417Q
produced the same effects, although they were weakerthan with UAS-DRacGAP∆EIE (Fig. 2G and Table 1).Accordingly, all genetic interactions shown below wereperformed using the UAS-DRacGAP∆EIE transgene.Overexpression of eitherUAS-DRacGAPDN transgenewith other Gal4 lines (MS1096, 69B) or in clones ofGal4-expressing cells (see Materials and methods)caused the same phenotypes (not shown).
All the defects of DRacGAPDN flies were rescued bycoexpression of wild-type DRacGAP (Fig. 3A,B), whileremaining unaltered in the presence of UAS-lacZ,indicating that they result from reduced DRacGAPactivity. Moreover, the mutant phenotype ofDRacGAPDN flies was increased in flies with only onecopy of the endogenous DRacGAPgene (compare Figs3C and 2H; 100% of the flies presented the phenotypeshowed in Fig. 3C). Still, DRacGAPDN proteins mayinterfere to some extent with the function of the relatedRnRacGAP, since heterozygosity for RnRacGAP(inDf(3R)dsx10M/+ flies) moderately increased the loss ofwing tissue of DRacGAPDN flies (compare Figs 3D and2H).
DRac1 and Dcdc42 are putative targets ofDRacGAPThe above results suggest that the small GTPase(s)whose activity is downregulated by DRacGAP shoul berequired for vein and SO development and for cellproliferation. By analogy with its closest homologues,DRacGAP should regulate RacGTPase. Indeed, wefound that overexpression of wild-type DRac1 (UAS-DRac1) driven by en-G4caused vein thickening,
S. Sotillos and S. Campuzano
Fig. 1. Molecular characterisation and expression ofDRacGAP. (A) Conceptual translation of DRacGAPcDNAand its alignment with DrosophilaRnRacGAP (GenBankaccession number gi1335764), mouse RacGAP (AF079974)and human n-chimaerin (pir508242). Sequencescorresponding to the cysteine-rich domain and the GAPdomain (boxes I, II and III) are underlined in lilac, yellow,orange and light green, respectively. A potential argininefinger (Scheffzek et al., 1998) is marked by the red line. Aputative SH3 binding motif (Lamarche and Hall, 1994) isunderlined in dark green. Identical and similar amino acidsare shown in black boxes. Percentages of identity/similarity inthe GAP domain between DRacGAP and RnRacGAP, mouseRacGAP or human n-chimaerin are 54/73%, 51/67% and28/52%, respectively. (B-E,G) Expression pattern ofDRacGAPin wing (B,C,E,G) and eye (D) imaginal discsfrom early (B) and late (C) second instar larvae, late thirdinstar larvae (D,E) and in 24-28 hours after pupariumformation pupal wing (G), visualised by in situ hybridisation.Note in B the stronger expression of DRacGAPin thepresumptive wing (W) than in the presumptive notum region(N). Asterisks in D indicate the two stripes of DRacGAPexpression in the eye disc at and ahead of the morphogeneticfurrow (determined by comparing the pattern of transcriptionof DRacGAPwith β-galactosidase staining in neuralized-lacZeye discs (not shown). (E,G) DRacGAPis very weaklyexpressed in the presumptive vein territories, visualised in Fby expression of ve/rho. L2-L5, longitudinal veins. Themagnification in B is 2× that in C, and in E and F, 4× C.

5431DRacGAP inhibits EGFR/Ras pathway
development of extra SOs and wing enlargement (Fig. 4A andTable 1). Coexpression of UAS-DRac1and UAS-DRacGAPDN
had a synergistic effect (Table 1 and Fig. 4B, note the extensivedevelopment of vein tissue; DRacGAPDN flies did not showa mutant phenotype at the temperature of the experiment),suggesting that DRacGAP reduces DRac1 activity.Consistently, the lethality of Sd-G4; UAS-DRac1larvae wasreversed by coexpression of UAS-DRacGAP(Fig. 4D) and thewing size reduction associated with a decreased activity ofDRac1 in Sd-G4; UAS-DRac1DN flies (UAS-DRac1N17Fig. 4E)was enhanced by coexpression of UAS-DRacGAP(Fig. 4F).Similar experiments could not be performed in the case of Rho
or Cdc42 GTPases since overexpression of UAS-DRhoor UAS-Dcdc42V12 was lethal even at 18°C. In order to investigatewhether DRho and DCdc42 are targets of DRacGAP, wereasoned that since GAPs are negative regulators of theGTPases, the phenotypic effects of decreased DRacGAPactivity should be reduced by a lowered activity of theregulated GTPase. According to this genetic test, DRhoA didnot seem to be a substrate for DRacGAP in the wing disc sincea decrease in DRhoAfunction (in DRhoA/+or DRho-GEF2/+backgrounds) did not appreciably modify the DRacGAPDN
mutant phenotype (not shown). On the contrary, a partial rescueof the DRacGAPDN phenotype was observed in a Cdc423/+
Fig. 2. Functional analysisof DRacGAP. (A-D) Overexpression ofDRacGAPinterferes with vein and SOdevelopment and reduces wing size.(A) Dorsal view of a wild-type wingshowing longitudinal veins L3 and L4and one of the campaniform sensilla ofL3 (arrow). (B) A similar region of aMS1096; UAS-DRacGAPwing showsloss of the dorsal corrugation of L3 andL4 veins and of campaniform sensilla(arrow). Note the reduction in thedistance between veins L3 and L4.(C-E) ve/rho expression responds tothe Rac pathway. Late third instar wingdiscs from wild-type (C), en-G4/+;UAS-DRacGAP/+ (D) and en-G4/+;UAS-DRacGAP∆EIE/+ (E) larvaeshowing that expression of ve/rhoinveins and wing margin (arrowheadsand arrow, respectively) is reduced (D)or expanded (E) in the posteriorcompartment. (G-L) Wing phenotypesof flies with reduced DRacGAPactivity. (F) Wild-type wing.(G-J) Flies overexpressingUAS-DRacGAPR417Q(G) or UAS-DRacGAP∆EIE (H-J) driven by en-G4show veins of increased width andfusion of veins (arrowhead in H), lossof wing tissue (H) and enlarged cells(arrows in G and H). (I,J) Magnifiedviews of en-G4/+; UAS-DRacGAP∆EIE/+ wings. Note in I thedevelopment of enlarged cells, eachone possessing several abnormallyoriented trichomes (arrowhead, normaltrichome orientation is indicated by thearrow) and in J the extra SOs(campaniform sensilla, some areindicated by arrows) along the L3 vein.(See also Table 1). (K,L) Anterior (K)and posterior (L) compartments of thesame en-G4/+;UAS-DRacGAP∆EIE/+wing disc stained with FITC-phalloidin. Note cells of increased sizein the posterior compartment (arrows).Flies were raised at 25°C (A, C-I, K,L)or 29°C (B, J). A, anteriorcompartment; P, posteriorcompartment. Line in F marks theanteroposterior compartment boundary.

5432
background (Table 1) pointing to DCdc42 as a possible targetof DRacGAP. Supporting the involvement of DRac1 andDCdc42 in vein development, reduction of their activity in fliesexpressing either UAS-DRac1N17 or UAS-Dcdc42N17 driven byen-G4caused a mild loss of vein phenotype (not shown andsee also Eaton et al., 1996).
Many of the Rho/Rac/Cdc42 effectors are serine/threoninekinases (Aspenström, 1999). Two of these kinases, p21-activatedkinase (Pak) and protein kinase N (Pkn), have been characterisedin Drosophila (Harden et al., 1996; Hing et al., 1999; Lu andSettleman, 1999). In an attempt to investigate whether any ofthem is involved in the processes affected by DRacGAP, weanalysed the phenotype of RacGAPDN flies in Pkn or Pakheterozygous backgrounds. While reduced Pkn function (PknP/+)did not appreciably modify their phenotype (not shown), asubstantial rescue of vein thickening and wing notching wasobserved in a Pak4/+ background (Fig. 4H; Table 1).
DRacGAP and DRac1 affect the EGFR/Ras signallingpathwayAs shown above, overexpression of UAS-DRacGAPDN or UAS-DRac1caused vein enlargement and the appearance of extraSOs, two structures requiring EGFR/Ras/Raf/MAPK activity(Clifford and Schüpbach, 1989; Díaz-Benjumea and García-Bellido, 1990; Díaz-Benjumea and Hafen, 1994). Since Rac andRas pathways cooperate in mammalian cells in the control ofcell proliferation (see Introduction), we investigated whether thephenotypes associated to increased Rac signalling could be dueto overactivity of the Ras pathway. This appears to be the case,since a reduction in EGFR signalling (by expression of UAS-RafDN, Martín-Blanco et al, 1999) reduced their vein, wingnotching and large cell phenotypes (compare Fig. 5B,D withFig. 4G,A, respectively). Similarly, when levels of the EGFRactivators ve/rho(Sturtevant et al., 1993) andvein(vn; Schneppet al., 1996; Simcox et al., 1996) were decreased (in ve7M vn/+flies), the wing notching associated with DRacGAPDN
expression was substantially corrected (Table 1). In contrast,activation of EGFR signalling enhanced the mutant phenotypeof DRacGAPDN flies. Thus, although flies heterozygous for
argos (aossty/+), a repressor ligand of EGFR (Freeman et al.,1992; Schweitzer et al., 1995), are phenotypically wild type, itscombination with DRacGAPDN significatively increased thenumber of campaniform sensilla of DRacGAPDN flies (Table 1).
Note that the loss of vein tissue and reduced size of en-G4;UAS-RafDN wings were partially rescued by the expression ofeither UAS-DRacGAPDN or UAS-DRac1(Fig. 5A-D). Theseresults further suggest that the efficacy of the EGFR pathwaywas enhanced by increased Rac activity. Signalling byEGFR/Ras leads to phosphorylation and consequent activationof MAPK (Gabay et al., 1997). Rac signalling also ultimatelyactivated MAPK since expression of UAS-DRacGAPDN widenedthe domains of accumulation of dp-ERK in the presumptiveveins and spread them into the interveins (Fig. 5E,F). This effectwas enhanced by coexpression of UAS-DRac1 (Fig. 5G). Theactivation of the EGFR pathway was also visualised by theincreased expression of the downstream gene ve/rho (Martín-Blanco et al., 1999 (Fig. 2E). As expected, overexpression ofDRacGAPproduced the opposite effect (Fig. 2D).
Expression of DRacGAPappears to be decreased in thedomains of EGFR activation. This is most apparent in thenotum region of second instar wing disc (Fig. 1B), where theEGFR activator veinis expressed (Simcox, 1997), and in thepresumptive veins of third instar wing disc (Fig. 1E), territoriesof maximal EGFR signalling (Gabay et al., 1997). Thisobservation suggests that the EGFR pathway may repress theexpression of DRacGAP. In agreement with this notion,expression of DRacGAPwas either decreased or enhanced incells expressing UAS-RasV12 or UAS-aos(Fig. 5I-K) in whichthe EFGR/Ras pathway is activated (Karim and Rubin, 1998)or repressed (Freeman et al., 1992; Schweitzer et al., 1995),respectively.
Overexpression of DRacGAPDN induces apoptosisThe above results suggest a cooperation between Rac and Rassignalling pathways in the activation of the MAPK. In thecontext of this cooperation, we analysed the other phenotypesof DRacGAPDN flies.
Apoptotic cells are rare in wild-type wing discs (Milán et
S. Sotillos and S. Campuzano
Fig. 3. DRacGAPDN protein interferes withDRacGAP function. Wing phenotypes of en-G4; UAS-DRacGAP∆EIE flies (A) are reversedby coexpression of UAS-DRacGAP(B) andseverely or moderately enhanced in fliesheterozygous for Df(2R)CX1(C) andDf(3R)dsx10M (D) (compare C and D with theen-G4/+; UAS- DRacGAP∆EIE/+ wing raised at25°C shown in Fig. 2H). HeterozygousDf(2R)CX1/+or Df(3R)dsx10M/+ flies arephenotypically wild type (not shown). Flieswere grown at 29°C (A,B) or 25°C (C,D).Arrowheads in C,D indicate enlarged veins.Wings are shown with anterior up and distal tothe right. All wings are shown at the samemagnification.

5433DRacGAP inhibits EGFR/Ras pathway
al., 1997). However, they were very abundant in DRacGAPDN
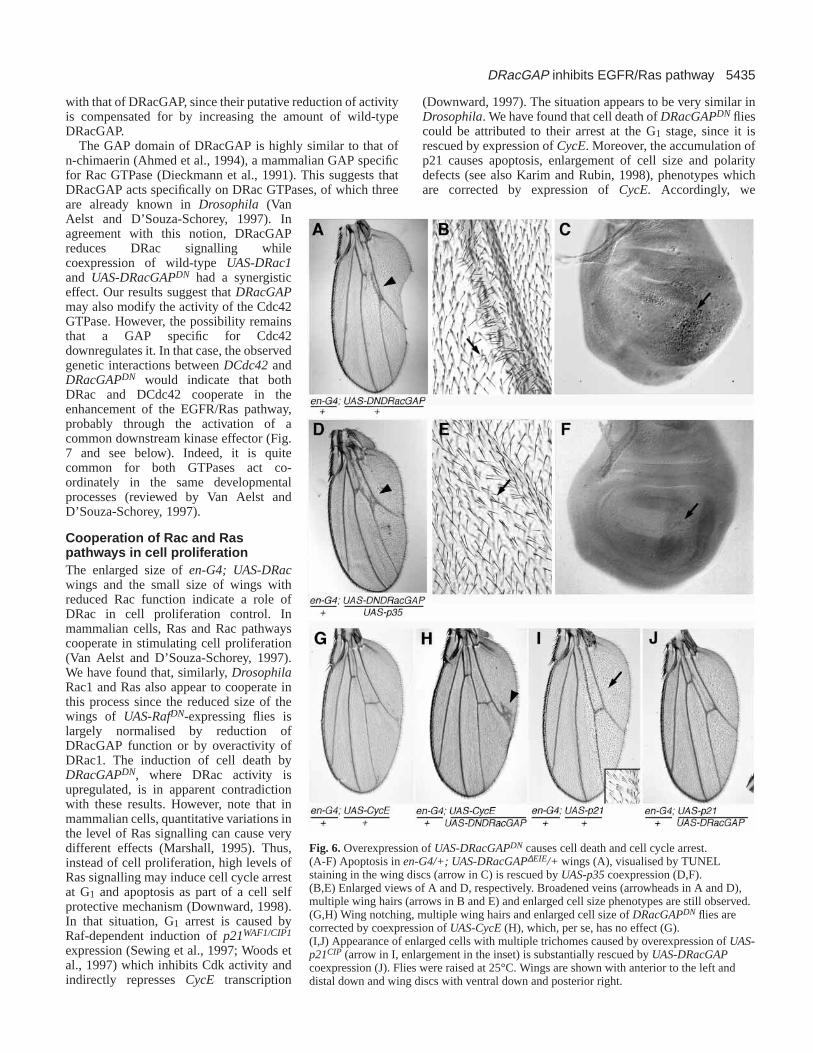
discs (Fig. 6C), indicating that the loss of tissue inDRacGAPDN wings (Figs 2H, 6A) was due to apoptosis.Moreover, this loss was rescued by coexpression of thebaculovirus caspase inhibitor p35 (Hay et al., 1994; Fig. 6D,F).The remaining DRacGAPDN phenotypes were not rescued,indicating that they are not indirect consequences of apoptosis.
Viability of the DRacGAPDN cells was monitored by clonalanalysis. Lineage clones (labelled with lacZ) were induced inDRacGAPDN first instar larvae and werevisualized 72 hours later. Clones weresignificantly more frequent in the Acompartment of the presumptive wing (51 Aclones and 14 P clones were found in 28 discs).This difference in clone frequency indicates alower viability of DRacGAPDN-expressingcells, which should lead to their gradual lossfrom the disc epithelium by ‘cell competition’(Simpson, 1979) and/or the apoptosis of theDRacGAPDN cells.
DRacGAPDN caused cell cycle arrestDRacGAPDN wings displayed enlarged cells(Figs 2I, 6B) and showed abnormal EPPmanifested by the appearance of severalrandomly orientated trichomes per cell (Fig.2I). Enlarged cells may result from cell cyclearrest, since blocking cell cycle progressiondoes not impair cell growth (reviewed byConlon and Raff, 1999). To analyse whetherDRacGAPDN cells were arrested at the G1 or G2phases of the cell cycle, we attempted tonormalise their size by expressing eitherCyclinE (CycE) or string, which driveprogression from G1 to S and G2 to M,respectively (Edgar and O´Farrell, 1990;Knoblich et al., 1994). Reversion to normal cellsize was found only by expressing UAS-CycE(Fig. 6H), suggesting arrest at G1. Cell deathand the development of multiple hairs percell were similarly rescued, whereas the veinphenotype was not modified (Fig. 6H).
As indicated above, the efficacy of the Raspathway seems to be enhanced in DRacGAPDN
flies. Overactivation of the Ras pathway mayalso contribute to the G1 arrest of DRacGAPDN
cells since we have found that although at 18°Coverexpression of UAS-rolledSem(the activatedform of MAPK) or UAS-DRacGAPDN did notenlarge cells or alter EPP, such modificationswere observed upon coexpression of both genes(not shown). Although Ras is an inducer of cellproliferation, in mammalian cells, prolonged,high-intensity Ras/Raf signalling arrests cellsin G1 (Sewing et al., 1997; Woods et al., 1997).This is mediated by Raf-dependent activationof the cyclin-dependent kinase (cdk) inhibitorp21WAF1/CIP1, which, indirectly, represses CycEtranscription (Downward, 1997). Indeed,expression of UAS-p21WAF1/CIP1in the imaginalwing disc caused mutant phenotypes somewhat
similar to those of DRacGAPDN, namely, the appearance ofenlarged cells with altered hair polarity (Karim and Rubin,1998 and Fig. 6I) and apoptosis (not shown). As expected,these phenotypes were partially reversed by UAS-CycEoverexpression (not shown), confirming that p21 arrests cellsin G1 (de Nooij and Hariharan, 1995). Remarkably, expressionof wild-type UAS-DRacGAPsignificantly rescued the mutantphenotype caused by expression of UAS-p21WAF1/CIP1 (Fig.6J), suggesting a reduction in p21 activity associated with
Fig. 4. DRacGAP reduces DRac1 pathway activity. (A,B) Reduced activity ofDRacGAP increases DRac1 signalling. (A) Overexpression of UAS-DRac1driven byen-G4at 18°C (A) causes wing phenotypes similar to those of en-G4/+; UAS-DRacGAP∆EIE/+ flies (G). (B) Mutant phenotype is enhanced by coexpression of bothtransgenes (see also Table 1). Note the extensive vein thickening (arrowheads).(C-F) In contrast, overexpression of DRacGAPreduces DRac1activity. Lethality ofSd-G4/+; UAS-DRac1/+flies is rescued by coexpression of UAS-DRacGAP(D) andthe reduced size of Sd-G4/+; UAS-DRac1N17/+ wings (E, compare with the wild-typewing shown in C) is enhanced by coexpression of UAS-DRacGAP (F). Sd-G4/+; UAS-DRacGAP/+flies raised at 18°C are phenotipically wild type. DRacGAPDN
phenotypes (G) are partially rescued when one dose of Pakis eliminated (H, see alsoTable 1). Flies were grown at 18°C (A-F) and 25°C (G-H). All wings are shown at thesame magnification.

5434
decreased Rac activity. Since transcription of UAS-p21isdriven by the heterologous UAS promoter, this suggests thatDRacGAP may be involved in posttranslational modificationof p21. Interestingly, involvement of Rho in p21 stability hasbeen described in cultured mammalian cells (Olson et al.,1998).
DISCUSSION
The activity of the signalling pathways that operate duringdevelopment must be tightly regulated to achieve differentthresholds required for the different cellular responses. Aninitial weak activation of a pathway could lead to transcriptionof activators of the pathway and/or repression of its inhibitors,to gain further amplification of the signal. During Drosophilavein development both positive (ve/rho) and negative (argos,sproutyand kekkon) regulators of the EGFR/Ras pathway aretranscribed in response to the pathway (Perrimon andMcMahon, 1999). In this report we show that signalling
downstream of Rho-family GTPases cooperate with theEGFR/Ras pathway in activation of the MAPK. Furthermore,expression of DRacGAP, a negative regulator of Rho-familyGTPases, is suppressed by EGFR/Ras signalling in theprospective vein regions of the wing disc. This transcriptionalrepression contributes to upregulation of the EGFR/Raspathway in these regions thus providing an additional controllevel of the pathway.
Function of DRacGAP in the wing discThe functional analysis of DRacGAP, performed byoverexpressing wild-type and dominant negative forms,indicates that it negatively regulates cell proliferation and veinand SO development. It cannot be ruled out completely thatDN forms of DRacGAP may also compete for targets withother RacGAPs. In fact, reducing the dosage of the relatedRnRacGAP(and of other uncharacterised genes included in thechromosomal deficiency used) slightly enhanced thephenotype of DRacGAPDN flies. In any case, the functions ofthose other uncharacterised RacGAPs appear to be redundant
S. Sotillos and S. Campuzano
Fig. 5. Cross-regulatory interactions between Racand EGFR/Ras pathways. (A-D) DRac1canpartially compensate for reduced Raf signalling.(A,C) Flies overexpressing UAS-RafDN in the Pcompartment cells at 25°C (A) show loss ofposterior compartment veins (L4 and L5) andreduced size. These phenotypes are less extreme at18°C (C). They are partially rescued bycoexpression of UAS-DRacGAP∆EIE (B) or UAS-DRac1(D). (Arrows in B and D indicate therescued veins). (E-G) MAPK activation, visualisedby staining with anti-dp-ERK, is enhanced byincreased Rac signalling. The domains ofaccumulation of dp-ERK in the veins in wild-typelate third instar wing discs (E, arrowheads) areexpanded and extend into the intervein regions(arrow) when DRacGAP activity is reduced in en-G4/+; UAS-DRacGAP∆EIE/+ flies (F). Expansionof dp-ERK is also found in the anteriorcompartment vein L3 most likely due to the lateexpression of enin anterior compartment cells(Blair, 1992). Note the dramatic expansion of dp-ERK in the wing pouch when DRac activity isfurther upregulated (in en-G4/UAS-DRac1; UAS-DRacGAP∆EIE/+, (G), compare with the wing ofthe same genotype shown in Fig. 4B).(H-K) DRacGAPexpression is regulated by theEGFR pathway. Expression of DRacGAP(arrow)in the P compartment in early third instar wingdiscs (H) is repressed in en-G4/+; UAS-RasV12/+discs (I) whereas it is enhanced in en-G4/+; UAS-argos/+ discs (K). Similar enhancement ofDRacGAPexpression is caused by overexpressionof argosin dpp-G4/ UAS-aosdiscs (J). (Note thatthe disc shown in J is slightly tilted in relation tothose shown in H,I,K). Larvae were raised at 18°C(C,D,H,I) and 25°C (A,B,E-G,J,K). Wing discs areshown with ventral up and posterior right.
5435DRacGAP inhibits EGFR/Ras pathway
with that of DRacGAP, since their putative reduction of activityis compensated for by increasing the amount of wild-typeDRacGAP.
The GAP domain of DRacGAP is highly similar to that ofn-chimaerin (Ahmed et al., 1994), a mammalian GAP specificfor Rac GTPase (Dieckmann et al., 1991). This suggests thatDRacGAP acts specifically on DRac GTPases, of which threeare already known in Drosophila(VanAelst and D’Souza-Schorey, 1997). Inagreement with this notion, DRacGAPreduces DRac signalling whilecoexpression of wild-type UAS-DRac1and UAS-DRacGAPDN had a synergisticeffect. Our results suggest that DRacGAPmay also modify the activity of the Cdc42GTPase. However, the possibility remainsthat a GAP specific for Cdc42downregulates it. In that case, the observedgenetic interactions between DCdc42andDRacGAPDN would indicate that bothDRac and DCdc42 cooperate in theenhancement of the EGFR/Ras pathway,probably through the activation of acommon downstream kinase effector (Fig.7 and see below). Indeed, it is quitecommon for both GTPases act co-ordinately in the same developmentalprocesses (reviewed by Van Aelst andD’Souza-Schorey, 1997).
Cooperation of Rac and Raspathways in cell proliferationThe enlarged size of en-G4; UAS-DRacwings and the small size of wings withreduced Rac function indicate a role ofDRac in cell proliferation control. Inmammalian cells, Ras and Rac pathwayscooperate in stimulating cell proliferation(Van Aelst and D’Souza-Schorey, 1997).We have found that, similarly, DrosophilaRac1 and Ras also appear to cooperate inthis process since the reduced size of thewings of UAS-RafDN-expressing flies islargely normalised by reduction ofDRacGAP function or by overactivity ofDRac1. The induction of cell death byDRacGAPDN, where DRac activity isupregulated, is in apparent contradictionwith these results. However, note that inmammalian cells, quantitative variations inthe level of Ras signalling can cause verydifferent effects (Marshall, 1995). Thus,instead of cell proliferation, high levels ofRas signalling may induce cell cycle arrestat G1 and apoptosis as part of a cell selfprotective mechanism (Downward, 1998).In that situation, G1 arrest is caused byRaf-dependent induction of p21WAF1/CIP1
expression (Sewing et al., 1997; Woods etal., 1997) which inhibits Cdk activity andindirectly represses CycEtranscription
(Downward, 1997). The situation appears to be very similar inDrosophila. We have found that cell death of DRacGAPDN fliescould be attributed to their arrest at the G1 stage, since it isrescued by expression of CycE. Moreover, the accumulation ofp21 causes apoptosis, enlargement of cell size and polaritydefects (see also Karim and Rubin, 1998), phenotypes whichare corrected by expression of CycE. Accordingly, we
Fig. 6. Overexpression of UAS-DRacGAPDN causes cell death and cell cycle arrest.(A-F) Apoptosis in en-G4/+; UAS-DRacGAP∆EIE/+ wings (A), visualised by TUNELstaining in the wing discs (arrow in C) is rescued by UAS-p35coexpression (D,F).(B,E) Enlarged views of A and D, respectively. Broadened veins (arrowheads in A and D),multiple wing hairs (arrows in B and E) and enlarged cell size phenotypes are still observed.(G,H) Wing notching, multiple wing hairs and enlarged cell size of DRacGAPDN flies arecorrected by coexpression of UAS-CycE (H), which, per se, has no effect (G).(I,J) Appearance of enlarged cells with multiple trichomes caused by overexpression of UAS-p21CIP (arrow in I, enlargement in the inset) is substantially rescued by UAS-DRacGAPcoexpression (J). Flies were raised at 25°C. Wings are shown with anterior to the left anddistal down and wing discs with ventral down and posterior right.

5436
hypothesise that induction of cell cycle arrest and apoptosis byoveractivity of the Rac pathway could be a consequence ofincreased Ras signalling. Rac would potentiate the reduced Rafsignalling occurring in RafDN flies, allowing wing disc cells toproliferate, but in the presence of wild-type Raf, overactivityof Rac would enhance Raf signal to such a high level as toinduce p21expression and ultimately, apoptosis, as previouslyshown to occur after expression of constitutively active Ras(Karim and Rubin, 1998). This interpretation is supportedby the partial rescue of the wing notching phenotype ofDRacGAPDN flies in a ve vnheterozygous background and inRafDN flies where EGFR/Ras signalling is reduced.
Note that cell size and polarity phenotypes of DRacGAPDN
are similarly induced by p21 and reversed by CycEexpression,indicating they can be attributed to cell cycle arrest. Indeed, itis known that cell cycle arrest may cause cell enlargement(Neufeld et al., 1998; Weigmann et al., 1997; reviewed byConlon and Raff, 1999). Moreover, Drosophila mutationsaffecting cell cycle machinery are very frequently associatedwith defects in the development and polarity of the wing hairs(Weigmann et al., 1997).
Cross regulatory interactions between Rac and Raspathways The effects on vein and SO development caused by increasedDRac1 activity are very similar to those associated withoveractivity of the EGFR/Ras pathway (Baker and Rubin,1992; Brunner et al., 1994; Karim and Rubin, 1998; Sturtevantet al., 1993; J. Culí, E. Martín-Blanco and J. Modolell, personal
communication). We show that both pathways cooperate toimplement these cell fates. Thus, the appearance of thickenedveins and extra SOs in DRacGAPDN flies was enhanced orsuppressed by increased or decreased activity of the EGFR/Raspathway, respectively. Conversely, overactivity of DRacrescued the loss of veins of RafDN flies. These geneticinteractions are also manifested in the effects of DRac on theexpression of a downstream target of the EGFR/Ras pathway(rho/ve). rho/ve expression is increased in flies with enhancedDRac signalling, and reduced in flies overexpressingDRacGAP, indicating that Rac signalling activates the Raspathway. The rescue of the loss-of-vein and reducedproliferation phenotypes of RafDN flies by overactivity ofDRac1 suggests that DRac1 functions downstream of Raf. Thepossibility of DRac1 acting in parallel with Raf to activatetargets of the MAPK is unlikely since MEK activity isincreased by enhancing Rac signaling, as visualised by theexpanded distribution of phosphorylated-MAPK, the substrateof MEK. How can Rac signaling impinge on Raf-inducedMEK activation? Any of the proteins which interact with Rafor MEK (Sternberg and Alberola-Ila, 1998) may be the targetfor posttranslational modification by any of the kinases actingas effectors of Rac (reviewed by Aspenström, 1999). Ourresults suggest that Rac activation of Ras pathway may bemediated by PAK, a serine/threonine kinase which binds to andis activated by Rac and Cdc42 (Aspenström, 1999) (Fig. 7).Mammalian PAK1 phosphorylates MEK1 on a site crucial forits in vivo binding to Raf, stabilising the binding of Raf toMEK1 and thereby enhancing Raf-mediated MEK1 activation(Frost et al., 1997). Moreover, mammalian Pak3 is able topositively regulate Raf1 activity through phosphorylation(King et al., 1998). Accordingly, rescue of the mutantphenotype of RafDN by overexpression of Rac might beaccomplished by overactivated PAK enhancing the activity ofthe residual Raf present in RafDN flies.
Note that overexpression of DRaconly caused thickening ofthe veins but did not induce development of ectopic veins,indicating that Rac activity is required to enhance the intensityof the Ras pathway in previously determined vein regions. Thisposes the question of how DRac activity is controlled. Sinceexpression of DRac1is generalised (our unpublished data),control of its activity should rely on the opposing effects ofRacGEFs and RacGAPs. Activation of GEFs appears todepend on their subcellular localisation (Van Aelst andD’Souza-Schorey, 1997). Similarly, since most of themammalian Rac-GAP proteins are ubiquitously distributed andcontain putative protein interaction domains, it is assumed thatthey are regulated by protein-protein interactions, which mayaffect their activity or their subcelullar localisation. However,expression of the human n- and β-chimaerins RacGAPs istissue specific (Van Aelst and D’Souza-Schorey, 1997) anddownregulation of humanβ-chimaerin in brain cells may be afactor in the development of malignant brain gliomas (Yuan etal., 1995). Hence, restricted expression of RacGAPscould bean alternative mechanism for Rac control. Patterned expressionis found in the case of DRacGAP; its expression in late thirdinstar larvae and early pupae is almost excluded from the veinregions and it accumulates at the intervein cells adjacent to theveins. The functional relevance of this distribution is stressedby the observation that expression of DRacGAP in veinterritories reduced Ras signalling while reduction of DRacGAP
S. Sotillos and S. Campuzano
EGFR
Ras
Raf
MAPK
ve
PAK
DRac1
DCdc42
DRacGAP
EGFR
Ras
Raf
MAPK
ve
PAK
DRac1
DCdc42
DRacGAP
DRacGAP
INTERVEINVEIN
MEK MEK
DRacGAP
Fig. 7.A model for Ras pathway self-stimulation mediated byincreased Rac activity. Schematic representation of EGFR/Raspathway in vein and intervein territories in the imaginal wing disc atlate third instar. Activity of the Ras pathway is higher in veins than inthe interveins, thus inhibiting DRacGAPtranscription in veinterritories. Reduced DRacGAP allows maximal activity in veins ofDRac1 GTPase and possibly also of the related DCdc42 GTPase.Alternatively, DCdc42 GTPase activity may be modulatedindependently of DRacGAP. Both GTPases bind to and activatePAK. This kinase would phosphorylate MEK, facilitating its bindingto Raf and thus increasing Ras signaling.

5437DRacGAP inhibits EGFR/Ras pathway
activity in the intervein territory expanded the domains of Raspathway activity.
Interestingly, in the Drosophilawing disc, DRacGAPtranscription is repressed by the EGFR/Ras pathway (Fig. 7).Activity of this pathway is finely tuned by it controlling theexpression of its own inhibitors and activators (Wasserman andFreeman, 1998; Martín-Blanco et al., 1999; Perrimon andMcMahon, 1999). Our results indicate that Ras signaling canself-stimulate through activation of the Rac pathway byrepression of DRacGAP(Fig. 7). During vein formation, theEGFR/Ras pathway, once it has attained a certain threshold,should repress expression of DRacGAPin the prospective veinregions thus ensuring maximal activity of Rac and, in turn, ofRas pathways in these territories of the imaginal wing disc,which should trigger vein differentiation. In contrast, maximalexpression of DRacGAPat the vein/intervein boundariesshould locally decrease Rac and Ras signalling, and incollaboration with Notch and Dpp pathways (for a review seede Celis, 1998) help to refine the final width of the veins.Hence, this regulatory loop is another feedback mechanismmodulating the activity of the EGFR/Ras pathway duringDrosophiladevelopment.
We are most grateful to J. Modolell, J. Culí and M. J. García fortheir constant help and advice in the course of this work; A. Baonza,J. F. de Celis, E. Martín-Blanco and colleagues in our laboratory forconstructive criticisms on the manuscript; E. Caminero for experttechnical assistance, J. Pinilla for statistical analysis and A. Baonza,J. Botas, R. Fehon, M. Freeman, L. Luo, E. Martín-Blanco, M.Mlodzick, G. Rubin, E. Sánchez-Herrero, L. Zipursky and theBloomington Stock Center for materials and stocks. A postdoctoralfellowship from the Comunidad Autónoma de Madrid to S. S. isacknowledged. This work was supported by grants from ComunidadAutónoma de Madrid (07B/0033/1997), Dirección General deInvestigación Científica y Técnica (PB93-0181 and PB98-0682 to J.Modolell) and an institutional grant from Fundación Ramón Arecesto the Centro de Biología Molecular Severo Ochoa.
REFERENCES
Agnel, M., Röder, L., Vola, C. and Griffin-Shea, R.(1992). A Drosophilarotund transcript expressed during spermatogenesis and imaginal discmorphogenesis encodes a protein which is similar to human Rac GTPase-activating (racGAP) proteins. Mol. Cell Biol.12, 5111-5122.
Ahmed, S., Lee, J., Wen, L.P., Zhao, Z., Ho, J., Best, A., Kozma, R. andLim, L. (1994). Breakpoint cluster region gene product-related domain ofn-chimaerin. J. Biol. Chem.269,17642-17648.
Ashburner, M. (1989). Drosophila: A Laboratory Handbook. Cold SpringHarbor: Cold Spring Harbor Laboratory Press.
Aspenström, P.(1999). Effectors for Rho GTPases. Curr. Opin. Cell Biol.11,95-102.
Baker, N. E. and Rubin, G. M.(1992). Ellipsemutations in the Drosophilahomologue of the EGF receptor affect pattern formation, cell division, andcell death in eye imaginal discs. Dev. Biol.150,381-396.
Barrett, K., Leptin, M. and Settleman, J. (1997). The Rho GTPase and aputative RhoGEF mediate a signaling pathway for the cell shape changes inDrosophilagastrulation. Cell 91, 905-915.
Blair, S. S. (1992) engrailed expression in the anterior lineagecompartment of the developing wing blade of Drosophila. Development115, 21-33.
Boguski, M. S. and McCormick, F.(1993). Proteins regulating Ras and itsrelatives. Nature366,643-654.
Bourne, H. R., Sanders, D. A. and McCormick, F.(1990). The GTPasesuperfamily: a conserved switch for diverse cell functions. Nature348,125-132.
Brand, A. H. and Perrimon, N. (1993). Targeted gene expression as a means
of altering cell fates and generating dominant phenotypes. Development118,401-415.
Brunner, D., Oellers, N., Szabad, J., Biggs III, W. H., Zipursky, S. L. andHafen, E. (1994). A gain-of-function mutation in DrosophilaMAP kinaseactivates multiple receptor tyrosine kinase signalling pathways. Cell 76,875-888.
Clifford, R. J. and Schüpbach, T. (1989). Coordinately and differentiallymutable activities of torpedo, the Drosophila melanogasterhomolog of thevertebrate EGF receptor gene. Genetics123,771-787.
Conlon, I. and Raff, M. (1999). Size control in animal development. Cell 96,235-244.
Cubas, P., de Celis, J. F., Campuzano, S. and Modolell, J.(1991). Proneuralclusters of achaete-scuteexpression and the generation of sensory organs inthe Drosophila imaginal wing disc. Genes Dev.5, 996-1008.
de Celis, J. F.(1998). Positioning and differentiation of veins in the Drosophilawing. Int. J. Dev. Biol.42, 335-343.
de Nooij, J. C. and Hariharan, I. K. (1995). Uncoupling cell fatedetermination from patterned cell division in the Drosophilaeye. Science270,983-985.
Devereux, J., Haeverli, P. and Smithies, O.(1984). A comprehensive set ofsequence analysis programs for the VAX. Nuc. Acids Res.12, 387-395.
Díaz-Benjumea, F. J. and García-Bellido, A.(1990). Behaviour of cellsmutant for an EGF receptor homologue of Drosophila in genetic mosaics.Proc. R. Soc. Lond.242,36-44.
Díaz-Benjumea, F. J. and Hafen, E.(1994). The sevenless signalling cassettemediates DrosophilaEGF receptor function during epidermal development.Development120,569-578.
Dieckmann, D., Brill, S., Garrett, M. D., Totti, N., Hsuan, J., Monfries, C.,Hall, C., Lim, L. and Hall, A. (1991). Bcrencodes a GTPase activatingprotein for p21Rac. Nature351,400-402.
Downward, J. (1997). Routine role for Ras. Curr. Biol. 7, R258-R260.Downward, J. (1998). Ras signalling and apoptosis. Curr. Opin. Genet. Dev.
8, 49-54.Eaton, S.(1997). Planar polarization of Drosophilaand vertebrate epithelia.
Curr. Opin. Cell Biol.9, 860-866.Eaton, S., Wepf, R. and Simons, K.(1996). Roles of Rac1 and Cdc42 in
planar polarization and hair outgrowth in the wing of Drosophila. J. CellBiol. 135,1277-1289.
Edgar, B. A. and O´Farrell, P. H. (1990). The three postblastoderm cellcycles of Drosophilaembryogenesis are regulated in G2 by string. Cell62,469-480.
Fehon, R. G., Oren, T., LaJeunesse, D. R., Melby, T. E. and McCartney,B. M. (1997). Isolation of mutations in the Drosophilahomologues of thehuman Neurofibromatosis 2and yeast CDC42genes using a simple andefficient reverse-genetic method. Genetics146, 245-252.
Freeman, M., Klämbt, C., Goodman, C. S. and Rubin, G. M.(1992). Theargosgene encodes a diffusible factor that regulates cell fate decisions inthe Drosophilaeye. Cell 69, 963-975.
Frost, J. A., Steen, H., Shapiro, P., Lewis, T., Ahn, N., Shaw, P. E. andCobb, M. H. (1997). Cross-cascade activation of ERKs and ternary complexfactors by Rho family proteins. EMBO J.16, 6426-6438.
Gabay, L., Seger, R. and Shilo, B. Z.(1997). In situ activation pattern ofDrosophilaEGF receptor pathway during development. Science277,1103-1106.
Guichard, A., E. Bergeret and R. Griffin-Shea.(1997). Overexpression ofRnRacGAP in Drosophila melanogaster deregulates cytoskeletalorganisation in cellularising embryos and induces discrete imaginalphenotypes. Mech. Dev.61, 49-62.
Hackel, P. O., Zwick, E., Prenzel, N. and Ullrich, A.(1999). Epidermalgrowth factor receptors: critical mediators of multiple receptor pathways.Curr. Opin. Cell Biol.11, 184-189.
Häcker, U. and Perrimon, N.(1998). DRhoGEF2encodes a member of theDbl family of oncogenes and controls cell shape changes during gastrulationin Drosophila. Genes Dev.12, 274-284.
Hall, A. (1998). Rho GTPases and the actin cytoskeleton. Science279,509-514.
Harden, N., Lee, J., Loh, H. Y., Ong, Y. M., Tan, I., Leung, T., Manser, E.and Lim, L. (1996). A Drosophilahomolog of the Rac- and Cdc42-activated serine/threonine kinase PAK is a potential focal adhesion and focalcomplex protein that colocalizes with dynamic actin structures. Mol. Cell.Biol. 16, 1896-1908.
Hay, B. A., Wolff, T. and Rubin, G. M. (1994). Expression of baculovirusP35 prevents cell death in Drosophila. Development120,2121-2129.
Hemsley, A., Arnheim, N., Toney, M., Cortopassi, G. and Galas, D.(1989).

5438
A simple method for site-directed mutagenesis using the polymerase chainreaction. Nucl. Acid Res.17, 6545-6551.
Hing, H., Xiao, J., Harden, N., Lim, L. and Zipursky, S. L. (1999). Pakfunctions downstream of Dock to regulate photoreceptor axon guidance inDrosophila. Cell 97, 853-863.
Karim, F. D. and Rubin, G. M. (1998). Ectopic expression of activated Ras1 induces hyperplastic growth and increased cell death in Drosophilaimaginal tissues. Development125,1-9.
King, A. J., Sun, H., Diaz, B., Barnard, D., Miao, W., Bagrodia, S. andMarshall, M. S. (1998). The protein kinase Pak3 positively regulates Raf-1 activity through phosphorylation of serine 338. Nature396,180-183.
Knoblich, J. A., Sauer, K., Jones, L., Richardson, H., Saint, R. and Lehner,C. F. (1994). Cyclin E controls S phase progression and its down-regulationduring Drosophila embryogenesis is required for the arrest of cellproliferation. Cell 77, 107-120.
Lamarche, N. and Hall, A. (1994). Gaps for Rho-related GTPases. TrendsGenet.10, 436-440.
Lu, Y. and Settleman, J.(1999). The DrosophilaPkn kinase is a Rho/Raceffector target required for dorsal closure during embryogenesis. Genes Dev.13, 1168-1180.
Luo, L., Jan, L. Y. and Jan, Y. N. (1997). Rho family small GTP-bindingproteins in growth cone signalling. Curr. Opin. Neurobiol.7, 81-86.
Luo, L., Liao, Y. J., Jan, L. Y. and Jan, Y. N.(1994). Distinct morphogeneticfunctions of similar small GTPases: DrosophilaDrac1 is involved in axonaloutgrowth and myoblast fusion. Genes Dev.8, 1787-1802.
Magie, C. R., Meyer, M. R., Gorsuch, M. S. and Parkhurst, S. M.(1999).Mutations in the Rho1 small GTPase disrupt morphogenesis andsegmentation during early Drosophiladevelopment. Development126,5353-5364.
Marshall, C. J. (1995). Specificity of receptor tyrosine kinase signaling:Transient versus sustained extracellular signal-regulated kinase activation.Cell 80, 179-185.
Martín-Blanco, E. (1998). Regulatory control of signal transduction duringmorphogenesis in Drosophila. Int. J. Dev. Biol.42, 363-368.
Martín-Blanco, E., Roch, F., E., N., Baonza, A., Duffy, J. B. and Perrimon,N. (1999). A temporal switch in DER signaling controls the specificationand differentiation of veins and interveins in the Drosophila wing.Development126,5739-5747.
Milán, M., Campuzano, S. and García-Bellido, A.(1997). Developmentalparameters of cell death in the wing disc of Drosophila. Proc. Natl. Acad.Sci. USA94, 5691-5696.
Montell, D. J. (1999). The genetics of cell migration inDrosophilamelanogasterand Caenorhabditis elegansdevelopment. Development126,3035-3046.
Neufeld, T. P., de la Cruz, A. F. A., Johnston, L. A. and Edgar, B. A.(1998).Coordination of growth and cell division in the Drosophilawing. Cell 93,1183-1193.
Noselli, S. and Agnès, F.(1999). Roles of the JNK signaling pathway inDrosophilamorphogenesis. Curr. Opin. Genet. Dev.9, 466-472.
Olson, M. F., Paterson, H. F. and Marshall, C. J.(1998). Signals from Rasand Rho GTPases interact to regulate expression of p21Waf1/Cip1. Nature394,295-299.
Perrimon, N. and McMahon, A. P.(1999). Negative feedback mechanismsand their roles during pattern formation. Cell 97, 13-16.
Prokopenko, S. N., Brumby, A., O´Keefe, L., Prior, L., He, Y., Saint, R.and Bellen, H. J.(1999). A putative exchange factor for Rho1 GTPase isrequired for the initiation of cytokinesis in Drosophila. Genes Dev.13,2301-2314.
Rommel, C. and Hafen, E.(1998). Ras – a versatile cellular switch. Curr.Opin. Genet. Dev.8, 412-418.
Sambrook, J., Fritsch, E. F. and Maniatis, T.(1989). Molecular Cloning.Cold Spring Harbor: Cold Spring Harbor Laboratory Press.
Scheffzek, K., Ahmadian, M. R. and Wittinghofer, A. (1998). GTPase-activating proteins: helping hands to complement an active site. TrendsBiochem. Sci.23, 257-262.
Schnepp, B., Grumbling, G., Donaldson, T. and Simcox, A.(1996). Vein isa novel component in the Drosophila epidermal growth factor receptorpathway with similarity to the neuregulins. Genes Dev.10, 2302-2313.
Schwartz, M. A. and Baron, V. (1999). Interactions between mitogenicstimuli, or, a thousand and one connections. Curr. Opin. Cell Biol.11,197-202.
Schweitzer, R., Howes, R., Smith, R., Shilo, B. Z. and Freeman, M.(1995).Inhibition of DrosophilaEGF receptor activation by the secreted proteinArgos. Nature376,699-702.
Sewing, A., Wiseman, B., Lloyd, A. C. and Land, H.(1997). High-intensityRaf signal causes cell cycle arrest mediated by p21Cip1. Mol. Cell. Biol.17,5588-5597.
Simcox, A. (1997). Differential requirement for EGF-like ligands inDrosophilawing development. Mech. Dev.62, 41-50.
Simcox, A. A., Grumbling, G., Schnepp, B., Bennington-Mathias, C.,Hersperger, E. and Shearn, A. (1996). Molecular, phenotypic, andexpression analysis of vein, a gene required for growth of the Drosophilawing disc. Dev. Biol.177,475-489.
Simpson, P.(1979). Parameters of cell competition in the compartments of thewing disc of Drosophila. Dev. Biol.69, 182-193.
Sone, M., Hoshino, M., Suzuki, E., Kuroda, S., Kaibuchi, K., Nakagoshi,H., Saigo, K., Nabeshima, Y. and Hama, C.(1997). Still life, a protein insynaptic terminals of Drosophilahomologous to GDP-GTP exchangers.Science275,543-547.
Sotillos, S., Roch, F. and Campuzano, S.(1997). The metalloprotease-disintegrin Kuzbanian participates in Notch activation during growthand patterning of the Drosophilaimaginal discs. Development124,4769-4779.
Sternberg, P. W. and Alberola-Ila, J.(1998). Conspiracy theory: RAS andRAF do not act alone. Cell 95, 447-450.
Struhl, G. and Basler, K. (1993). Organizing activity of wingless protein inDrosophila. Cell 72, 527-540.
Strutt, D. I., Weber, U. and Mlodzik, M. (1997). The role of RhoA in tissuepolarity and Frizzled signaling. Nature387, 292-295.
Sturtevant, M. A., Roark, M. and Bier, E. (1993). The Drosophila rhomboidgene mediates the localized formation of wing veins and interactsgenetically with components of the EGF-R signaling pathway. Genes Dev.7, 961-973.
Van Aelst, L. and D’Souza-Schorey, C.(1997). Rho GTPases and signalingnetworks. Genes Dev.11, 2295-2322.
Wasserman, J. D. and Freeman, M.(1998). An autoregulatory cascade ofEGF receptor signaling patterns the Drosophilaegg. Cell95, 355-364.
Weigmann, K., Cohen, S. M. and Lehner, C. F.(1997). Cell cycleprogression, growth and patterning in imaginal discs despite inhibition ofcell division after inactivation of DrosophilaCdc2 kinase. Development124,3555-3563.
Woods, D., Parry, D., Cherwinski, H., Bosch, E., Lees, R. and McMahon,M. (1997). Raf-induced proliferation or cell cycle arrest is determined bythe level of Raf activity with arrest mediated by p21Cip1. Mol. Cell. Biol.17, 5598-5611.
Wooltorton, E. J., Haliotis, T. and Mueller, C. R.(1999). Identification andcharacterization of a transcript for a novel Rac GTPase-activating proteinin terminally differentiating 3T3-L1 adipocytes. DNA Cell Biol.18, 265-273.
Yuan, S., MIller, D. W., Barnett, G. H., Hahn, J. F. and Williams, B. R.(1995). Identification and characterization of human beta 2-chimaerin:association with malignant transformation in astrocytoma. Cancer Res.55,3456-3461.
Zar, J. H. (1999). Biostatistical Analysis.New Jersey: Prentice Hall Intl. Inc.
S. Sotillos and S. Campuzano