
I
CARACTERIZACIÓN DE LA ACTIVIDAD TRIPANOCIDA Y
CITOTÓXICA DE ACEITES ESENCIALES DE LIPPIA ALBA Y
SUS TERPENOS BIOACTIVOS SOBRE TRYPANOSOMA CRUZI
ERIKA MARCELA MORENO MORENO
UNIVERSIDAD DE SANTANDER
FACULTAD DE CIENCIAS DE LA SALUD MAESTRÍA EN INVESTIGACIÓN EN ENFERMEDADES INFECCIOSAS
BUCARAMANGA
2017

II
CARACTERIZACIÓN DE LA ACTIVIDAD TRIPANOCIDA Y
CITOTÓXICA DE ACEITES ESENCIALES DE LIPPIA ALBA Y
SUS TERPENOS BIOACTIVOS SOBRE TRYPANOSOMA CRUZI
ERIKA MARCELA MORENO MORENO
Proyecto de Grado presentado al Programa de Maestría en Investigación en
Enfermedades Infecciosas de la Facultad de Ciencias de la Salud de la Universidad
de Santander, como parte de los requisitos para la obtención del título de Magíster en
Investigación en Enfermedades Infecciosas.
Tutor: Dra. Liliana Torcoroma García Sánchez, MSc, PhD.
Co-Tutor: Sandra Milena Leal Pinto, MSc, PhD.
UNIVERSIDAD DE SANTANDER
FACULTAD DE CIENCIAS DE LA SALUD MAESTRÍA EN INVESTIGACIÓN EN ENFERMEDADES INFECCIOSAS
BUCARAMANGA
2017

III

IV
Tutor
DRA. LILIANA TORCOROMA GARCÍA SÁNCHEZ, MSc, PhD

V
No te rindas, por favor no cedas,
aunque el frío queme,
aunque el miedo muerda,
aunque el sol se ponga y se calle el viento,
aún hay fuego en tu alma,
aún hay vida en tus sueños,
porque cada día es un comienzo nuevo,
porque esta es la hora y el mejor momento.
Mario Benedetti

VI
A Miguel y Alix

VII
AGRADECIMIENTOS
Agradezco de forma muy especial a la Dra. Liliana Torcoroma García
Sánchez, por depositar su confianza en mí y haberme permitido trabajar con
su maravilloso equipo de trabajo, de quiénes aprendí valiosos
conocimientos durante estos dos años.
A todos los docentes y personal administrativo del programa de Maestría en
Investigación en Enfermedades Infecciosas de la Universidad de Santander.
A la Dra. Sandra Leal, por su apoyo y competente orientación.
A la Dra. Elena Stashenko, directora del Centro Nacional de Investigación
para la Agroindustrialización de Especies Vegetales Aromáticas y
Medicinales Tropicales (CENIVAM), quién nos facilitó los aceites
esenciales.
Al Laboratorio de Investigación e Innovación en Biotecnología
Agroambiental (LIIBAAM) de la Universidad de Santander, por la
disponibilidad de sus instalaciones.
A la Secretaría de Salud del municipio de Santander, por su gentil
colaboración en la adquisición del Benznidazol.
A la Dra. Leonor Yamile Vargas del programa de Química ambiental de la
Universidad Santo Tomás de Aquino, Bucaramanga, por su colaboración en
la purificación del médicamento de referencia.
A la Dra. Wendy Grey Nieto Pérez del laboratorio de Biotecnología de la
Fundación Cardiovascular, por su disponibilidad y orientación en la
realización de las pruebas de citometría de flujo.
A mis preciadas alumnas, Stephany, María Fernanda y Laura, por su
valiosa colaboración.
A mi gran amigo, Jorge Silva.
A mi familia.

VIII
LISTA DE FIGURAS
Figura 1 Representación esquemática de las formas extracelulares de T. cruzi.. ........ 3 Figura 2. Representación esquemática de la forma intracelular de T. cruzi ................ 4 Figura 3. Ciclo de vida de T. cruzi. .............................................................................. 5
Figura 4. Distribución global de los casos de Enfermedad de Chagas…………...…..7
Figura 5. Distribución de los principales vectores de T. cruzi en el continente
Americano.. .................................................................................................................. 9 Figura 6. Triatoma infestans ........................................................................................ 9 Figura 7. Formas de transmisión oral de T. cruzi... .................................................... 12 Figura 8. Didelphis marsupialis y Panstrongylus megistus infectados con T. cruzi . 14 Figura 9. Mecanismos causales de miocarditis en la infección con T. cruzi. ............ 15 Figura 10. Fases de la infección por T. cruzi.. ........................................................... 17 Figura 11. Estructuras del Benznidazol y Nifurtimox ............................................... 20 Figura 12. Mecanismo de acción del Benznidazol y Nifurtimox............................... 20 Figura 13. Lippia alba (Mill.) N.E. Brown. ............................................................... 25 Figura 14. Distribución geográfica de quimiotipos de Lippia alba. .......................... 27 Figura 15. Estructuras químicas de terpenos de L. alba. ........................................... 30 Figura 16. Actividad Tripanocida y Citotóxica in vitro de Aceite Esenciales de L.
alba quimiotipos Citral y Carvona. .................................................................... ……45
Figura 17. Porcentajes de muerte celular sobre formas cíclicas de T. cruzi tratadas
con 100 µg/mL de AEs de L. alba quimiotipos Citral y Carvona. ............................ 46
Figura 18. Actividad Tripanocida y Citotóxica in vitro de terpenos de L. alba ........ 48 Figura 19. Porcentajes de muerte celular sobre las formas extracelulares de T.cruzi
tratadas con 50 µg/mL de las fracciones terpenoides. ............................................... 49 Figura 20. Isobologramas de las interacciones farmacológicas entre el limoneno y
otros terpenos de AEs de L. alba. .............................................................................. 51 Figura 21. Pérdida del potencial de membrana mitocondrial en epimastigotes de
T.cruzi observados mediante microscopia de contraste de fases interferencial. ........ 52 Figura 22. Electroforesis en gel de agarosa Low Melting de epimastigotes de T. cruzi
tratados con los compuestos en estudio. .................................................................... 54 Figura 23. Fragmentación de ADN en epimastigotes de T.cruzi observados mediante
microscopia de contraste de fases interferencial.. ...................................................... 55 Figura 24. Análisis por citometría de flujo de la externalización de la fosfatidilserina
en epimastigotes de T. cruzi. ...................................................................................... 58

IX
LISTA DE TABLAS
Tabla 1. Reservorios de T. cruzi ................................................................................ 13 Tabla 2. Cuadro comparativo de acción de AEs frente a T. cruzi ............................. 24 Tabla 3. Diversidad fitoquímica de Lippia alba ........................................................ 26 Tabla 4. Aceites esenciales de L. alba quimiotipo Citral. .......................................... 34 Tabla 5. Aceites esenciales de L. alba quimiotipo Carvona. ..................................... 35 Tabla 6. Composición de los medios de cultivo utilizados para el crecimiento in vitro
de Células Vero y T. cruzi .......................................................................................... 37 Tabla 7. Matriz de interacción entre el Limoneno y el resto de terpenos .................. 40 Tabla 8. Composición química relativa y efecto antiproliferativo de los AEs extraídos
del quimiotipo Citral de L. alba sobre T. cruzi. ......................................................... 43 Tabla 9. Composición química relativa y efecto antiparasitario de los AEs extraídos
del quimiotipo Carvona de L. alba sobre T. cruzi ...................................................... 44 Tabla.10. Muerte celular de AEs de L. alba, quimiotipo Citral y Carvona a 100
µg/mL sobre las diferentes formas de T. cruzi. .......................................................... 46 Tabla 11. Efecto antiparasitario de los principales terpenos derivados de L. alba en T.
cruzi ............................................................................................................................ 47 Tabla 12. Muerte celular de terpenos de L. alba a 50 µg/mL sobre las diferentes
formas de T. cruzi. ...................................................................................................... 49 Tabla 13. Interacción farmacológica entre Limoneno y el resto de terpenos derivados
de L.alba. .................................................................................................................... 50

X
LISTA DE ABREVIATURAS
°C: Grados Celsius
CIF: Sumatoria de la CIF µg/mL: Microgramo sobre mililitro
µL: Microlitro
µM: Micromolar
ADN: Ácido Desoxirribonucleico
AE: Aceite esencial
AEs: Aceites esenciales
ANOVA: Análisis de varianza
ATP: Adenosín Trifosfato
BNZ: Benznidazol
CC50: Concentración citotóxica 50
CI50: Concentración Inhibitoria 50
cel/mL: Células por mililitro
CENIVAM: Centro Nacional de Investigaciones para la Agroindustrialización de
Especies Vegetales, Aromáticas y Medicinales Tropicales
CIF: Concentración inhibitoria fraccional
CO2: Dióxido de carbono
COX2: Ciclooxigenasa 2
DAPI: 4 ',6¬diamino¬-2, ¬fenilindol
DMSO: Dimetilsulfóxido
DMEM: Dubelcco’s Modified Eagle Medium
DO: Densidad óptica
DE: Desviación estandar
DTUs: Unidades discretas de tipificación
Em: Emisión
Ex: Excitación
GC-MS: Cromatografía de gases acoplada a espectrometría de masas
gr/mL: Gramos por mililitro
HD-MW: Hidrodestilación asistida por la radiación de un microondas
INS: Instituto Nacional de Salud
IS: Índice de selectividad
kDa: Kilo Dalton(s)
L. alba: Lippia alba
LIT: Liver Infusion Tryptose
LPS: Lipopolisacaridos
m.s.n.m: Metros sobre el nivel del mar
MC: Muerte celular
min: Minutos
mm: Milimetros
NFX: Nifurtimox
nm: Nanómetro
nM: Nanomolar
NO: Oxido nítrico
NOS: Oxido nítrico sintetasa

XI
OMS: Organización Mundial de la Salud
OxCar: Óxido de cariofileno
PBS: Phosphate Buffered Saline
PPAR: receptores activos por proliferadores de peroxisomas
PS: Fosfatidilserina
ROS: Especies reactivas de oxígeno
RPMI: Roswell Park Memorial Institute
SFBi: Suero Fetal Bovino Inactivado
T. cruzi: Trypasnosoma cruzi
TDC: Tripomastigotes derivados de células
TNF: Factor de necrosis tumoral
TryR: Tripanotión reductasa
U/mL: Unidades por mililitro

XII
TABLA DE CONTENIDO
1. INTRODUCCIÓN………………………………………………………… …1
1.1 Trypanosoma cruzi…..…………………………………………….…....…3
1.1.1 Ciclo de Vida ............................................................................................ 4
1.1.2 Epidemiología ............................................................................................ 8
1.1.3 Transmisión ............................................................................................... 8
1.1.4 Reservorios .............................................................................................. 15
1.1.5 Patogénesis de la Enfermedad de Chagas................................................ 17
1.1.6 Manifestaciones cínicas ........................................................................... 17
1.1.7 Tratamientos clásicos .............................................................................. 19
1.1.8 Tratamientos alternativos ........................................................................ 22
1.2 Lippia alba…………………………………………………..……....... …25
1.2.1 Etnofarmacología .................................................................................. ...28
1.2.2 Actividad biológica de L. alba ................................................................ 28
1.2.3 Actividad tripanocida de L. alba ............................................................. 29
1.2.4 Terpenos de L. alba ................................................................................. 29
2. OBJETIVOS ................................................................................................ …32
2.1 OBJETIVO GENERAL……………………………………………….....32
2.2 OBJETIVOS ESPECÍFICOS………………………………………….....32
3. MATERIALES Y MÉTODOS ........................................................................ 33
3.1 MATERIALES…………………………………………………..……….33
3.1.1 Aceites esenciales derivados de L. alba .................................................. 33
3.1.2 Terpenos y medicamentos ....................................................................... 36
3.1.3 Cultivos celulares .................................................................................... 37
3.2 MÉTODOS…………………………………………………………….....38
3.2.1 Citotoxicidad en Células Vero ................................................................. 38
3.2.2 Actividad antiparasitaria en Epimastigotes de T. cruzi ........................... 38
3.2.3 Actividad antiparasitaria en Tripomastigotes derivados de células ……38
3.2.4 Actividad antiparasitaria en amastigotes intracelulares de T. cruzi ......... 39
3.2.5 Evaluación del efecto combinado entre el Limoneno y el resto de
terpenos sobre T. cruzi ..................................................................................... 39
3.2.6 Cambios Morfológicos ............................................................................ 40
3.2.7 Determinación del potencial de membrana mitocondrial ........................ 40
3.2.8 Determinación de la fragmentación del ADN ......................................... 41
3.2.9 Determinación de la externalización de la fosfatidilserina ...................... 41
3.3 ANÁLISIS DE DATOS……………………………………………...… 42
4. RESULTADOS ................................................................................................ 43
4.1 COMPOSICIÓN QUÍMICA Y ACTIVIDAD TRIPANOCIDA DE AEs
DE L. alba………………………………………………………………………43
4.2 ACTIVIDAD TRIPANOCIDA DE TERPENOS DE L. alba…………. 47
4.3 INTERACCIÓN FARMACOLÓGICA DE LOS TERPENOS SOBRE T.
cruzi………………………………………………………………………….….49
4.4 MECANISMO DE MUERTE CELULAR INDUCIDO POR LOS
COMPUESTOS EN ESTUDIO………………………………………………...51

XIII
4.4.1 Cambios celulares ................................................................................... 51
4.4.2 Fragmentación del ADN .......................................................................... 54
4.4.3 Externalización de la fosfatidilserina ...................................................... 57
5. DISCUSIÓN .................................................................................................... 59
6. CONCLUSIONES ........................................................................................... 67
7. REFERENCIAS ............................................................................................... 68

XIV
RESUMEN
TÍTULO
CARACTERIZACIÓN DE LA ACTIVIDAD TRIPANOCIDA Y CITOTÓXICA
DE ACEITES ESENCIALES DE LIPPIA ALBA Y SUS TERPENOS BIOACTIVOS
SOBRE TRYPANOSOMA CRUZI
AUTORES
ERIKA MARCELA MORENO MORENO
PALABRAS CLAVE
Lippia alba, Aceites esenciales, Trypanosoma cruzi
DESCRIPCIÓN
La Enfermedad de Chagas es una infección causada por el protozoario Trypanosoma
cruzi, la cual es caracterizada por un prolongado curso silencioso y complejas
manifestaciones clínicas. Actualmente, no existe evidencia contundente de
quimioterapéuticas eficaces, lo que hace obligatoria la búsqueda de nuevos agentes
tripanocidas más efectivos. Bajo este escenario, fármacos extraídos de plantas
resultan prometedores, teniendo en cuenta su baja toxicidad y amplio espectro
farmacológico. De acuerdo con esto, en este trabajo se evaluó la actividad
tripanocida in vitro de aceites esenciales (AEs) de Lippia alba y de sus terpenos
bioactivos (óxido de cariofileno, citral, limoneno y carvona), caracterizándose el
mecanismo de muerte celular inducido contra T.cruzi. Los resultados fueron
expresados como la concentración inhibitoria o citotóxica 50 (CI50 y CC50,
respectivamente), realizando regresión sigmoidal mediante el Software estadístico
MsxlfitTM. El fenotipo de muerte fue rastreado por microscopía óptica y de
fluorescencia, citometría de flujo y electroforesis de ADN. El análisis estadístico se
realizó por ANOVA de Welch (software SPSS 15.0). En las condiciones testadas, los
AEs y terpenos de L. alba ejercieron efectos inhibitorios distintos en los tres estadios
parasitarios evaluados (p=0.000). Los AEs del quimiotipo Citral exhibieron
concentraciones inhibitorias medias menores (Epimastigotes: CI50 13,6±1,5µg/mL;
Tripomastigotes: CI50 21,9±1,4 µg/mL; Amastigotes: CI50 74,1±4,4µg/mL) que las
alcanzadas con los aceites del quimiotipo Carvona (Epimastigotes: CI50
88,2±3,7µg/mL; Tripomastigotes: CI50 44,9±2,5µg/mL; Amastigotes: CI50
>150µg/mL). El limoneno representó el terpeno de mayor selectividad
(Epimastigotes: IS 7,1; Tripomastigotes: IS 32,8; Amastigotes: IS 10,3) y menor
toxicidad sobre células Vero (Limoneno: CC50 297,1±2,4µg/mL). Además, el
limoneno evidenció una interacción sinérgica in vitro con el Benznidazol
(Epimastigotes CIF=0,44±0,13; tripomastigotes CIF=0,42±0,10; amastigotes
CIF=0,58±0,13µg/mL) y con el óxido de cariofileno (Epimastigotes: CIF 0,49;
Tripomastigotes: CIF 0,45; Amastigotes: CIF 0,71µg/mL). Los análisis celulares
sugieren que la muerte celular observada en el T. cruzi podría ser mediada por un
mecanismo tipo-apoptótico.

XV
ABSTRACT
TITLE
CHARACTERIZATION OF THE TRYPANOCIDAL AND CYTOTOXIC
ACTIVITY OF ESSENTIAL OILS FROM LIPPIA ALBA AND THEIR
BIOACTIVE TERPENES ON TRYPANOSOMA CRUZI
AUTHOR
ERIKA MARCELA MORENO MORENO
KEYWORDS
Lippia alba, Essential oils, Trypanosoma cruzi
DESCRIPTION
Chagas Disease is caused by infection of Trypanosoma cruzi parasite. In general, the
course of the disease is characterized by a prolonged silent phase and a complex
clinical presentation. Currently, there is no compelling evidence of effective
chemotherapeutics, making compulsory the research on more effective and selective
therapies.In this way, plants offer a promissory reserve of bioactive molecules, take
into account their low toxicity and width pharmacologic spectrum. In this work, the
in vitro trypanocidal activity of Lippia alba essential oils (EOs) and their bioactive
terpenes (caryophyllene oxide, citral, limonene and carvone) was evaluated,
following the death mechanism induced on T. cruzi cells. The results were expressed
as Inhibitory Concetration 50 (IC50) or Cytotoxic Concentration (CC50) by sigmoidal
regression using the statistical software MsxlfitTM. Optical and fluorescent
microscopy, flow cytometry, and DNA electrophoresis were used for
characterization of the cell death phenotype. The statistical analysis was made by
Welch’s ANOVA (software SPSS 15.0). In the conditions tested, the L. alba EOs
and their terpenes exhibed a different tripanocidal performance on the diverse
parasites stages (p=0.000). Citral chemotype oils evidenced lower IC50
(Epimastigotes: IC50 13,6±1,5µg/mL; Trypomastigotes: IC50 21,9±1,4 µg/mL, and
Amastigotes: IC50 74,1±4,4µg/mL) than Carvone chemotype EOs (Epimastigotes:
IC50 88,2±3,7µg/mL; Trypomastigotes: IC50 44,9±2,5µg/mL, and Amastigotes: IC50
>150µg/mL). The better selectivity index (SI) was achieved by the monoterpene
limonene (Epimastigotes: SI=7,1; Trypomastigotes: SI=32,8, and Amastigotes:
SI=10,3), with a CC50 value on Vero cells of 297,1±2,4µg/mL. Limonene presented
in vitro synergistic interaction with Benznidazole (with a Fractional Inhibitory
Concentration (FIC) of 0,44±0,13, 0,42±0,10 and 0,58±0,13µg/mL for
epimastigotes, trypomastigotes, and intracellular amastigotes, respectively), and with
caryophyllene oxide (Epimastigotes: FIC=0,49; Tripomastigotes: FIC=0,45;
Amastigotes: FIC=0,71). The cellular analysis suggested that these oils or their
bioactive terpenes (caryophyllen oxide/limonene) could be inducing cell death by an
apoptotic like mechanisms on T. cruzi.

Introducción
1
1. INTRODUCCIÓN
La Enfermedad de Chagas, constituye una de las más importantes enfermedades
tropicales desatendidas a nivel mundial y uno de los problemas más graves de salud
pública en América Latina. Se estima que más de 7 millones de personas están
infectadas en todo el mundo, en donde el 99% de los casos ocurren en el continente
Latinoamericano (Rassi et al., 2012; OMS, 2015). Actualmente, se estima que los
costos globales de esta enfermedad, originada por la infección del protozoo flagelado
intracelular Trypanosoma cruzi, son de aproximadamente US $ 7,19 mil millones por
año (Lee et al., 2013) con pérdidas económicas regionales de US $1,2 mil millones
anuales (Hotez et al., 2008; Hotez et al., 2013). Por su parte, la prevalencia de
Tripanosomiasis en Colombia se ha estimado entre 700.000 y 1.200.000 infectados y
más de 8.000.000 de personas en riesgo (INS, 2012; Cruz y Moreno, 2015).
A pesar del gran avance en el conocimiento de la biología del protozoario T. cruzi y
de los esfuerzos eficaces centrados en el control vectorial y transmisión por
transfusiones sanguíneas (Coura y Dias, 2009; Coura y Viñas, 2010; Coura, 2015),
grandes desafíos aún deben ser enfrentados; en particular aquellos asociados a la
cambiante epidemiología de la enfermedad, atribuida a la variedad de vectores,
reservorios y diferentes vías de transmisión y a la falta de terapias profilácticas y
esquemas de quimioterapéuticos eficaces.
Actualmente, el manejo terapéutico convencional de la Enfermedad de Chagas se
reduce al uso exclusivo de dos medicamentos, Nifurtimox (Lampit®, Bayer) y
Benznidazol (Rochagan® en Brazil y Radanil® en Argentina, Roche), introducidos
en la terapia clínica desde hace más de 40 años (Bustamante y Tarleton, 2014).
Ambos comparten deficiencias relacionadas principalmente con los largos periodos
de tratamiento, sus altos grados de toxicidad (que conllevan a la aparición de efectos
adversos severos como intolerancia digestiva, anorexia grave y compromisos
neurológicos (Rassi et al., 2012; Bern, 2015)), eficiencia tripanocida variable y
eficacia limitada en la fase crónica de la infección (Soeiro y De Castro, 2009; Urbina,
2010; Alviano et al., 2012; Morillo et al., 2015). Adicionado a esto, se ha reportado
el surgimiento de parásitos resistentes (Croft et al., 2005; Coura, 2009).
De acuerdo con lo anterior, la búsqueda de nuevos fármacos tripanocidas es un
problema en abierto, por lo que es una obligación de los países tropicales priorizar
sus esfuerzos públicos y privados, para encontrar nuevas sustancias con actividad
antiparasitaria que permitan el desarrollo de tratamientos más cortos, con mayor
efectividad en los diferentes estadios del parásito, mejores índices de selectividad y

Introducciön
2
menores efectos colaterales (Bernacchi et al., 2002). En este sentido, en las últimas
décadas se han focalizado intensas investigaciones a partir de fármacos extraídos de
plantas o sintetizados, tomando como base prototipos vegetales, los cuales han
evidenciado resultados promisorios para el tratamiento de enfermedades parasitarias
(Schmidt et al., 2012; Wink, 2012; Ramos et al., 2014).
Algunos aceites esenciales (AEs) derivados de plantas de las familias Verbenaceae,
Zingiberaceae, Piperaceae y Poaceae, entre otras (y sus componentes mayoritarios),
han sido descritos como agentes antimicrobianos de amplio espectro farmacológico
(Ramos et al., 2014), incluso con actividad significativa antihelmíntica y
antiprotozoaria (Bakkali et al., 2008; Escobar et al., 2010; Cheikh Ali et al., 2011;
Alviano et al., 2012; Borges et al., 2012; York et al., 2012; Leal et al., 2013). AEs
ricos en terpenos tales como citral (Lippia alba y Cymbopogon citratus), óxido de
cariofileno (Aframomum sceptrum, Achillea millefolium y Piper var brachypodon),
limoneno (L. origanoides y L. pedunculosa) y carvona (Origanum vulgare)
mostraron inhibir el crecimiento de formas extra e intracelulares de T. cruzi con alta
eficiencia (Santoro et al., 2007a; Santoro et al., 2007b; Cheikh Ali et al., 2011;
Santos et al., 2012; Leal et al., 2013; Kpoviessi et al., 2014).
De manera interesante, la presencia de estos terpenos ha sido identificada en AEs
extraídos de dos quimiotipos (Citral y Carvona) del arbusto aromático Lippia alba,
que crece en la región de Santander (Colombia) (Stashenko et al., 2004; García et al.,
2017). Estudios anteriores de tamizaje con estos aceites, evidenciaron acción
inhibitoria y citotóxica selectiva in vitro, contra parásitos tripanosomatídeos
(Cardoso y Soares, 2010; Escobar et al., 2010)
De acuerdo con lo anterior, en este trabajo se planteó como objetivo principal
estudiar las propiedades inhibitorias inducidas por los aceites esenciales del
quimiotipo citral y carvona de L. alba y sus terpenos bioactivos, sobre formas
epimastigotes, tripomastigotes y amastigotes de T. cruzi. Así mismo, fue realizado el
seguimiento del fenotipo de muerte celular ocasionado por el tratamiento, mediante
ensayos de microscopía óptica y fluorescencia, citometría de flujo y electroforesis de
ADN. Estos mismos estudios fueron ejecutados, utilizando como tratamiento
tripanocida los cuatro terpenos realacionados con la actividad tripanocida, que
componen estos dos tipos de aceites (Quimiotipo Carvona: carvona y limoneno; y
Quimiotipo Citral: citral y óxido de cariofileno). La CI50 fue determinada para cada
uno de los compuestos y posibles interacciones farmacológicas fueron evaluadas
mediante una matriz de combinaciones.

Introducciön
3
1.1 Trypanosoma cruzi
Trypanosoma cruzi es un protozoario hemoflagelado perteneciente al orden
Kinetoplastida y familia Trypanosomatidae (Rassi et al., 2012). Este parásito exhibe
una amplia diversidad genética, que permite su clasificación en seis unidades
discretas de tipificación (DTUs): TcI–TcVI (Zingales et al., 2009; Zingales et al.,
2012) siendo el primero, el grupo predominante en Colombia (Luna et al., 2009).
El ciclo biológico del parásito incluye tres formas celulares caracterizadas por la
posición del flagelo, kinetoplasto y núcleo (Prata, 2001): Epimastigotes,
Tripomastigotes y Amastigotes (Figura 1 y 2).
Figura 1 Representación esquemática de las formas extracelulares de T. cruzi. (A) Epimastigote (B)
Tripomastigote (Modificado de Teixeira et al., 2012).
Los epimastigotes (Figura 1A), representan la forma extracelular replicativa no
infectante del parásito, alojados en el intestino medio del vector. Poseen un aspecto
fusiforme, con una longitud aproximada de 20 μm y su kinetoplasto se sitúa en la
parte anterior al núcleo. Por otro lado, los tripomastigotes (Figura 1B) también de
20 μm de longitud, constituyen la forma extracelular infectante no replicativa,
localizada en el intestino posterior y deyecciones de los triatominos. Esta forma se
caracteriza por ser fusiforme alargada, poseer un único flagelo, una membrana
ondulante a lo largo del cuerpo y un kinetoplasto en la parte posterior al núcleo. En
Introducciön
4
los mamíferos, esta es la forma celular que difunde la infección a través de la sangre
(Shyamapada, 2014).
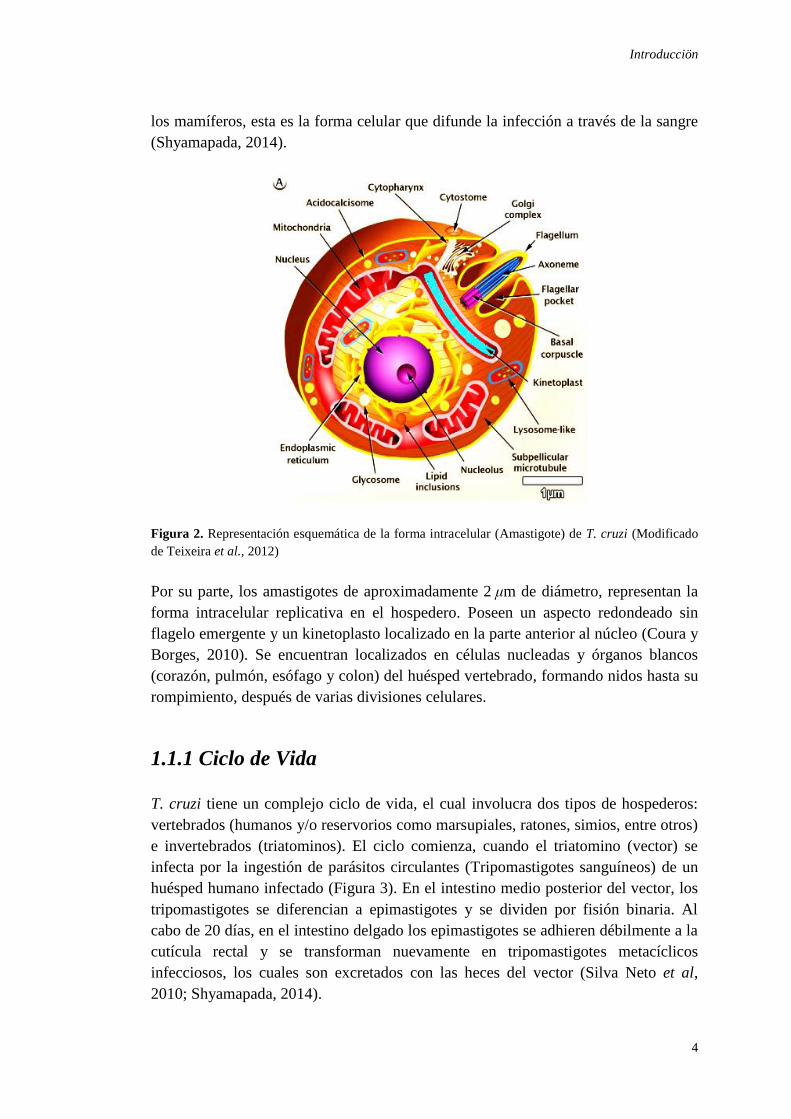
Figura 2. Representación esquemática de la forma intracelular (Amastigote) de T. cruzi (Modificado
de Teixeira et al., 2012)
Por su parte, los amastigotes de aproximadamente 2 μm de diámetro, representan la
forma intracelular replicativa en el hospedero. Poseen un aspecto redondeado sin
flagelo emergente y un kinetoplasto localizado en la parte anterior al núcleo (Coura y
Borges, 2010). Se encuentran localizados en células nucleadas y órganos blancos
(corazón, pulmón, esófago y colon) del huésped vertebrado, formando nidos hasta su
rompimiento, después de varias divisiones celulares.
1.1.1 Ciclo de Vida
T. cruzi tiene un complejo ciclo de vida, el cual involucra dos tipos de hospederos:
vertebrados (humanos y/o reservorios como marsupiales, ratones, simios, entre otros)
e invertebrados (triatominos). El ciclo comienza, cuando el triatomino (vector) se
infecta por la ingestión de parásitos circulantes (Tripomastigotes sanguíneos) de un
huésped humano infectado (Figura 3). En el intestino medio posterior del vector, los
tripomastigotes se diferencian a epimastigotes y se dividen por fisión binaria. Al
cabo de 20 días, en el intestino delgado los epimastigotes se adhieren débilmente a la
cutícula rectal y se transforman nuevamente en tripomastigotes metacíclicos
infecciosos, los cuales son excretados con las heces del vector (Silva Neto et al,
2010; Shyamapada, 2014).

Introducciön
5
Cuando el triatomino infectado ingiere sangre de huéspedes humanos susceptibles,
produce deyecciones. El prurito ocasionado por la picadura del insecto induce al
individuo al rascado; y los tripomastigotes metacíclicos encontrados en las heces del
vector, ingresan al huésped vertebrado a través de las micro laceraciones generadas
por la comezón o a través de las mucosas (Sanabria, 2016).
Figura 3. Ciclo de vida de Trypanosoma cruzi (Kessler et al., 2017).

Introducciön
6
Una vez los tripomastigotes metacíclicos ingresan al humano, invaden las células
nucleadas (macrófagos, células musculares y células epiteliales, entre otras) en el
sistema retículo endotelial localizado y tejido conectivo. En el citoplasma, se
diferencian en amastigotes esféricos y se multiplican por fisión binaria cada 12 horas
aproximadamente, durante un período de 4 a 5 días (Bern, 2015).
Cuando la célula se hincha con amastigotes, estos se transforman en tripomastigotes
sanguíneos y se liberan por la ruptura de la célula huésped. Los tripomastigotes se
propagan a través del torrente sanguíneo y la circulación linfática, e invaden tejidos
adyacentes y células musculares (cardíacas, lisas y esqueléticas) y ganglionares
principalmente, donde experimentan ciclos adicionales de multiplicación intracelular
(Mandal G et al., 2013). Finalmente, el ciclo se completa, cuando los tripomastigotes
sanguíneos son ingeridos por triatominos sanos, durante su alimentación
(Shyamapada, 2014; Bern, 2015).
1.1.2 Epidemiología
La Enfermedad de Chagas se localizó originalmente en zonas rurales del continente
Latinoamericano. No obstante, un cambio en el patrón epidemiológico ha sido
evidenciado en las últimas décadas, con mayor prevalencia, urbanización y
propagación a otros continentes. La reciente globalización, el aumento de los viajes
aéreos e inmigración, constituyen las causas principales del incremento en la
distribución de la enfermedad a Norteamérica, Europa, Australia y Japón (Gascon et
al., 2010). En los Estados Unidos, se estima que 300.000 personas están infectadas,
según los Centros para el Control y Prevención de Enfermedades (CDC, 2013).
Según la OMS, más de 7 millones de personas están infectadas con T. cruzi en todo
el mundo, en donde el 99% de las infecciones proceden de 21 países continentales de
América Latina: Argentina, Belice, Brasil, Chile, Colombia, Costa Rica, El Salvador,
Guyana Francesa, Guatemala, Guyana, Honduras, México, Nicaragua, Panamá,
Paraguay, Perú, el Estado Plurinacional de Bolivia, Surinam y Uruguay (OMS,
2015). Las áreas con mayor prevalencia se encuentran en la región del Gran Chaco,
que incluye el centro y sur de Bolivia, el norte de Argentina y el oeste de Paraguay
(Figura 4) (Bern, 2015; OMS, 2015).
En Latinoamérica, esta patología de prolongado curso silencioso y complejas
manifestaciones clínicas, constituye uno de los principales problemas de salud
pública, causando más de 7000 muertes al año, con altas tasas de morbilidad y
discapacidad de por vida. Además, más de 25 millones de personas corren el riesgo
de contraer la infección (OMS, 2015).

Introducciön
7
Figura 4. Distribución global de los casos de Enfermedad de Chagas notificados a la OMS en todo el
mundo, 2010-2013. La migración mundial ha dado lugar a la difusión de la enfermedad de Chagas en
todo el mundo, inclusive en regiones que antes se pensaba no eran endémicas para la infección (OMS,
2015).
En los últimos años, una evidente disminución de la prevalencia de la enfermedad ha
sido presenciada, debido a los exitosos programas de control vectorial y de
transfusiones sanguíneas realizados en varios países latinoamericanos (Dias, 2015).
En 1985, la OMS estimó que casi 18 millones de personas padecían la infección. No
obstante, 30 años después, el último informe actualizado de la OMS, declara que solo
5.7 millones de personas están infectadas en el continente latinoamericano (OMS,
2015).
Bolivia exhibe las tasas de prevalencia e incidencia más alta; sin embargo, Argentina,
Brasil y Colombia contienen una gran mayoría de pacientes con cardiopatía
chagásica. Argentina, Paraguay y Bolivia evidencian la mayor seroprevalencia de T.
cruzi en donantes de sangre (2% - 3%). Por su parte, la transmisión congénita es
mayor en México (1788 casos en 2010) y Argentina (1457 casos en 2010) (OMS,
2015)
De acuerdo con estas cifras, el costo de la Enfermedad de Chagas fue calculado en
338,500 (95% UI: 183,800- 846,400) años de vida ajustados por discapacidad
(DALYs) (Murray et al., 2015), siendo así, la patología de mayor impacto
socioeconómico en América Latina, con un costo económico estimado de US $ 1.200
millones al año. A estos costos deben sumarse, los gastos médicos generados por el

Introducciön
8
tratamiento de personas infectadas que desarrollan patología cardiaca o digestiva
(Hotez et al., 2013; Lee et al., 2013).
En Colombia, la prevalencia de la enfermedad ha sido estimada entre 700.000 y
1.200.000 infectados y cerca de 8 millones de personas en riesgo de adquirir la
infección (INS, 2012; Cruz y Moreno, 2015). Los departamentos que presentan la
mayor tasa de infección incluyen Arauca (21,1%), Casanare (10%), Santander
(6,3%), Norte de Santander (5,2%), Boyacá (3,7%), Cundinamarca (1,9%) y Meta
(1,7%) (Rueda et al., 2014).
1.1.3 Transmisión
La enfermedad de Chagas se transmite a los seres humanos y a más de 150 especies
de animales domésticos y salvajes, principalmente a través de las heces de insectos
hematófagos de la familia Reduviidae y subfamilia Triatominae (Rassi et al., 2012).
A la fecha se han identificado alrededor de 140 especies de triatominos, de los cuales
26 han sido reportados en Colombia (Rueda et al., 2014) y solo unos pocos son
vectores competentes (Triatoma infestans, Rhodnius prolixus, Triatoma dimidiata y
Panstrongylus megistus) para la transmisión del protozoario (Gorla et al., 2010).
Aunque la transmisión vectorial sigue siendo el mecanismo más común para la
adquisición de T. cruzi, vías alternas como la transmisión congénita, transfusiones,
trasplantes de órganos e ingesta de alimentos contaminados, se han convertido
actualmente en importantes rutas de transmisión (Rassi y Marín, 2010).
1.1.3.1 Transmisión vectorial
La transmisión antropo-zoótica de T. cruzi, representa la forma más común de
infección en las zonas endémicas para la Enfermedad de Chagas (Figura 5). En
Colombia, se estima que existen alrededor de 437.960 habitantes infectados mediante
este modo de transmisión (Esteban et al., 2016). Habitualmente, la presencia de los
vectores se asocia con viviendas de bajos recursos, las cuales tienen lugares aptos
para la proliferación del triatomino (techos construidos con ramas, grietas, casas
hechas de materiales de paja, barro, adobe) (Murcia et al., 2013; Moretti, 2015;
Mantilla et al., 2011).

Introducciön
9
Figura 5. Distribución de los principales vectores de Trypanosoma cruzi en el continente Americano.
El área endémica para la transmisión vectorial se extiende desde el sur de los Estados Unidos de
América (EE.UU.) hasta el sur de Argentina y Chile (Sosa-Estani y Segura, 2015).
Triatoma infestans constituye el vector principal de T. cruzi en gran parte de América
del Sur (Figura 6) (Coura, 2015). Se presume su origen en Bolivia y de allí su
trasladado a Argentina, Chile, Paraguay, Uruguay y Brasil (Noireau, 2009). No
obstante, en el año de 1997 en Uruguay, 1999 en Chile y 2006 en Brasil, el vector
logró ser eliminado a través de la Iniciativa del Cono Sur (Coura y Dias, 2009).
Figura 6. Triatoma infestans, vector principal de T. cruzi (Teixeira et al., 2011)

Introducciön
10
Rhodnius prolixus representa uno de los vectores más importantes para T. cruzi
debido a su rápido ciclo de desarrollo, intensa dispersión, antropofilia y alta
susceptibilidad a la infección y transmisión del protozoario. Es considerado el vector
principal de Colombia, Guyana Francesa y Venezuela, desde donde se cree fue
difundido a través de las aves o pajillas de las palmeras, a los países de América
Central, con excepción de Panamá y Costa Rica (Guhl, 2007). Se ha reportado su
eliminación en Guatemala, Honduras, Nicaragua y El Salvador, con posibles focos
aislados (Coura y Dias, 2009).
En América del Sur, Triatoma dimidiata se localiza en la región centro-oeste de
Colombia, la costa del Ecuador, Norte de Perú y Venezuela. Constituye uno de los
principales vectores en Costa Rica, donde se distribuye ampliamente gracias a su
capacidad para colonizar viviendas humanas (Ramírez et al., 2005; Guhl, 2007). En
países de América Central como Nicaragua, Honduras y Guatemala, donde R.
prolixus ha sido eliminado, T. dimidiata se ha convertido en el principal objetivo.
Una de las particularidades de esta especie, es que sus ninfas se camuflan con la
tierra en el suelo de las viviendas.
Actualmente, Panstrongylus megistus representa el vector potencial más importante
de Brasil, dada su amplia distribución geográfica, alta susceptibilidad a T. cruzi y su
versatilidad como vector salvaje con fácil adaptación doméstica. El hábitat natural
de P. megistus comprende con frecuencia nidos silvestres de marsupiales, en los
ecotopos salvajes del sudeste y sur de Brasil (Coura y Dias, 2009).
1.1.3.2 Transmisión por transfusión sanguínea y trasplante de
órganos
La transmisión de la infección por T. cruzi a través de la transfusión sanguínea,
constituye el segundo mecanismo de transmisión más frecuente en América Latina.
No obstante, debido al aumento de las migraciones de pacientes con enfermedad de
Chagas a países no endémicos, nuevos escenarios globales de este mecanismo han
surgido en los últimos años (Schmunis, 2007; Coura y Viñas, 2010).
La transmisión de T. cruzi por transfusiones sanguíneas, fue sugerida inicialmente
por Dias en 1945 y los primeros casos reportados de personas que adquirieron la
infección mediante este mecanismo, fueron descritos por Freitas y colaboradores en
1952. En Estados Unidos, Suiza, Japón, España y Canadá, la transmisión a través de
vías iatrogénicas, ha sido bien documentada a partir de casos no detectados de la
enfermedad (Gascon et al., 2010).
Por su parte, el control de donantes de sangre en el cono sur, principalmente en
Brasil, Uruguay, Chile y otros países del Sur y Centro América, ha traído consigo la

Introducciön
11
evidente disminución de la infección a través de la transfusión sanguínea. No
obstante, a pesar de la depreciación de las infecciones, este mecanismo sigue siendo
un gran desafío, tanto para países endémicos que no han introducido controles en
relación con sus donantes de sangre; como en países no endémicos que reciben de
manera permanente miles de inmigrantes con enfermedad de Chagas (Coura y Viñas,
2010; Coura et al., 2014). Actualmente, se estima que aproximadamente 300.167
inmigrantes latinoamericanos en EE.UU, están infectados con T. cruzi (Minneman et
al., 2012).
Por otro lado, el trasplante de órganos en pacientes con enfermedad de Chagas y el
uso de órganos de donantes infectados, ha constituido una problemática de debate
durante muchos años en áreas endémicas. La mayor parte de la experiencia publicada
respecto al trasplante de pacientes infectados con T. cruzi, están relacionados con el
trasplante renal, donde se ha puesto de manifiesto que la reactivación de la
enfermedad, se produce principalmente durante el primer año postrasplante con una
incidencia del 15% a 35% (Riarte et al., 1999). La probabilidad de transmisión de T.
cruzi con trasplante de hígado o riñón parece ser menor que el de trasplante de
corazón. El riesgo sustancial de transmisión en receptores de trasplante cardíaco
oscila entre el 75% y 100%; y la incidencia de la reactivación de la miocardiopatía
chagásica tras el trasplante cardíaco se sitúa entre el 21 y el 45% (Kransdorf et al.,
2014). Debido a ello, no se recomeinda el uso de aloinjertos cardíacos de donantes
potenciales que sean seropositivos para la infección por T. cruzi (Kransdorf et al.,
2014).
1.1.3.3 Transmisión congénita
La transmisión congénita constituye el escenario epidemiológico más importante de
la enfermedad de Chagas crónica, teniendo en cuenta la evidente mejora en el control
vectorial. Esta forma de transmisión representa el 22% de casos nuevos de T. cruzi en
el año 2010 (OMS, 2015). De acuerdo con la OMS, el número de mujeres infectadas
en edad fértil es de aproximadamente 1,8 millones y se estima que 14.400 recién
nacidos se infectan cada año (OMS, 2006). La prevalencia de la infección en mujeres
embarazadas varía entre diferentes países y localidades rurales o urbanas, con tasas
de infección congénita de <1 a 70,5% (Oliveira et al., 2010). En Argentina, la tasa de
transmisión oscila entre 1% y 12% (Gürtler et al., 2003; De Rissio et al., 2010). Por
su parte, en Chile, la velocidad de transmisión congénita del parásito es del 8,4%
(Jercic et al., 2010). En Colombia, Cucunubá et al, (2012) determinó que la
prevalencia general de infección por T. cruzi en mujeres embarazadas de 5
departamentos fue del 2,7%, de la siguiente manera: Casanare: 4,0%; Santander:
3,3%; Boyacá: 3,2%; Arauca: 2,1% y meta 0,2%. Un año después, Manrique-Abril y
colaboradores, en dos municipios Boyacenses indicaron una prevalencia de infección
congénita de 4,0% y 2,8% para Miraflores y Moniquira, respectivamente (Manrique-

Introducciön
12
Abril et al., 2013). En éste contexto y teniendo en cuenta los datos locales, se podría
estimar que la prevalencia de infección por T. cruzi en mujeres embarazadas de
poblaciones endémicas colombianas oscila entre el 2% y 4 % (Cucunubá et al, 2017).
1.1.3.4 Transmisión oral
La transmisión oral de T. cruzi probablemente representa el mecanismo más antiguo
y frecuente entre los reservorios silvestres y domésticos del ciclo (Shikanai-Yasuda y
Carvalho, 2012; Coura, 2015). No obstante, en los seres humanos, esta forma
transmisión ocurre de manera inusual, a través de la ingestión de alimentos
contaminados o por el consumo de carne cruda (Figura 7) (Díaz y González, 2014).
Figura 7. Formas de transmisión oral de T. cruzi. Frutas o alimentos son rociadas con secreciones de
las glándulas anales odoríferas de los animales del género Didelphis (Zarigüeya) infectadas con T.
cruzi. Consumo de carne cruda de animales infectados. Vectores infectados son triturados con el
alimento durante su preparación (jugo de caña). Alimentos contaminados con deyecciones de
triatominos (Díaz y González, 2014).
En 1969, Shaw y colaboradores describieron los cuatro primeros casos agudos de la
Enfermedad de Chagas, producidos en Belém PA, Brasil, probablemente a través de
la vía oral. Desde ese tiempo, cientos de casos agudos originados por la transmisión
oral, han sido descritos en la Amazonía (Steindel et al., 2008; De Barros et al.,
2009).
A la fecha, el mayor brote de transmisión oral de T. cruzi reportado se produjo en
Caracas, Venezuela, involucrando 103 casos agudos de la enfermedad (De Noya et
al., 2010). En Colombia, esta vía de transmisión ha sido documentada recientemente
en el departamento de Santander, con evidencia de infecciones agudas de forma
severa y letal (Díaz et al., 2015).

Introducciön
13
1.1.4 Reservorios
Un total de 180 especies de mamíferos pertenecientes a ocho órdenes diferentes,
ampliamente distribuidos en todas las regiones del neotrópico fitogeográfico, han
sido identificados como posibles reservorios de Trypanosoma cruzi (Tabla 1)
(Herrera, 2010).
Tabla 1. Reservorios de T. cruzi (Modificado de Herrera, 2010).
ORDEN NOMBRE COMÚN
Carnivora
Cerdocyon thous azarae Zorro
Canis familiaris Perro doméstico
Eyra barbara Huron, Comadreja
Felis catus Gato doméstico
Nasua nasua Cuatí
Potos flavus Kinkajou
Procyon lotor Mapache
Chiroptera
Antrozous pallidus pacificus Murciélago pálido
Artibeus luturatus Murciélago frutero grande
Lonchophylla mordaz Murciélago nectifero
Mycronycteris megalotis Murciélago orejudo
Noctilio labials Murciélago blanco
Cingulata
Dasypus kapleri Armadillos, Cachicamos
Dasypus novemcinctus Armadillo, Cachicamo de nueve
bandas
Cabassous unicinctus Armadillo de cola desnuda del oriente
Didelphidomorphia
Didelphis albiventris Faro, Rabipelado
Didelphis marsupialis Zarigüeya
Gracilinanus agilis Comadrejita agil
Monodelphis domestica Zarigüeya de cola corta
Philander oposum Comadreja de cuatro ojos
Lagomorpha
Sylvilagus floridanus Conejo de monte
Perisodactyla
Bos Taurus Toro
Sus scrofa Marrano
Tayassu pecari Chácharo
Pilosa
Bradipus tridactytus Oso perezoso de tres dedos
Mirmecophaga tridactyla Oso hormiguero gigante
Tamandúa tetradactyla Oso hormiguero
Primata
Alouatta caraya Mono aullador
Ateles belzebuth Mono araña
Callitrix penicillata Mono titi de pinceles negros
Cebus apella Mono carablanca
Leontopithecus rosalia Mono león dorado
Rodentia
Akodon sp. Ratón campestre
Cavia sp. Cuy, Acure
Nectomys squamipes Rata de agua
Rattus Rata negra, Rata casera
Rhiphidomys macrurus Rata trepadora

Introducciön
14
La especie de armadillo Tatusia novencincta constituye el mamífero placentario más
infectado con T. cruzi. Por su parte, los marsupiales del género Didelphis (Figura 8)
representan los primeros hospederos de T. cruzi en América del Sur. Las zarigüeyas
(Didelphis sp.) poseen la capacidad peculiar de mantener amastigotes en los tejidos y
epimastigotes en las secreciones luminales de las glándulas anales, desde donde las
formas parasitarias contaminan el medio ambiente (Yeo et al., 2005).
Figura 8. Didelphis marsupialis y Panstrongylus megistus infectados con T. cruzi (Coura, 1966).
Los roedores también juegan un papel importante en la difusión de T. cruzi, ya que
varias especies con una amplia distribución ecológica pueden ser infectados por el
protozoario. La prevalencia del 30% de las infecciones en Rattus rattus se
correlaciona con las altas tasas de infecciones en los seres humanos (Galuppo et al.,
2009). Con respecto al orden Carnivora, perros y gatos muestran una alta prevalencia
de infección natural de T. cruzi. Ellos representan los principales animales
domésticos que participan en la transmisión peridomiciliaria y domiciliaria de esta
infección parasitaria (Teixeira et al., 2006).
La infección de T. cruzi en la mayoría de los animales salvajes y domésticos cursa
asintomática. En general en estos huéspedes, las infecciones permanecen latentes
durante años, sin la producción de una patología que ponga en peligro la vida del
animal. No obstante, mamíferos infectados con T. cruzi pueden tener manifestaciones
clínicas y patológicas similares a las descritas para la Enfermedad de Chagas en
humanos. (Teixeira et al., 2006).
Introducciön
15
1.1.5 Patogénesis de la Enfermedad de Chagas
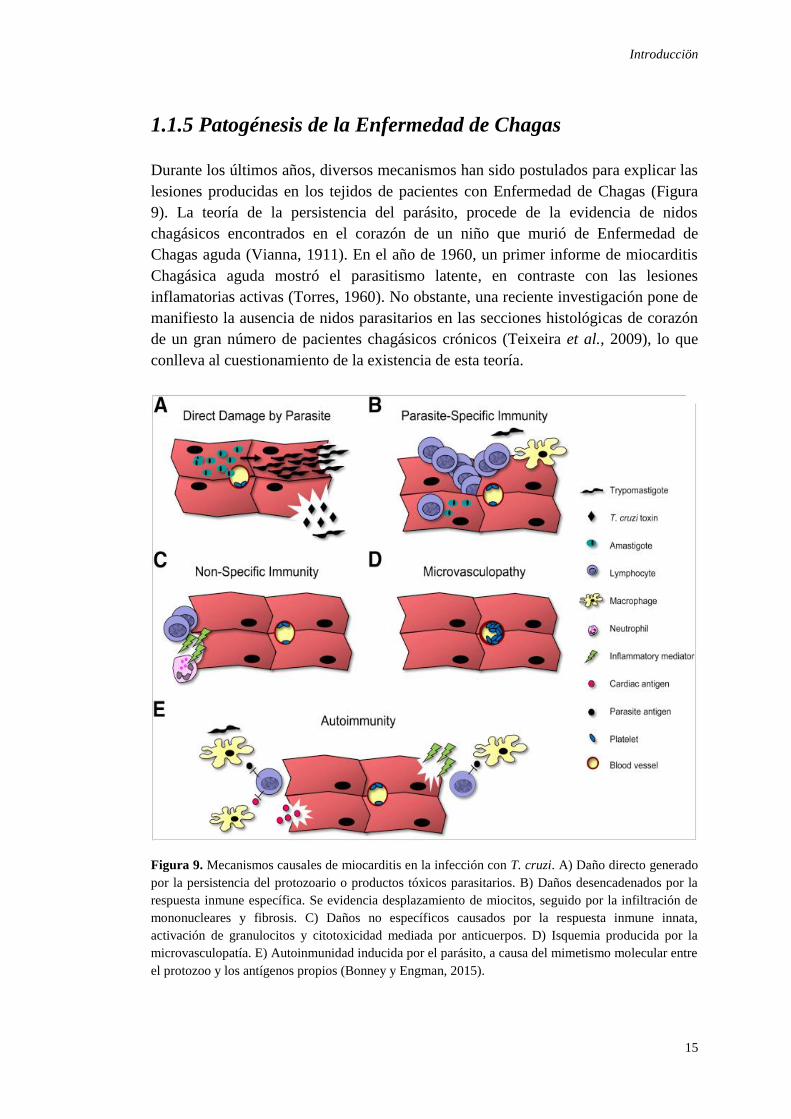
Durante los últimos años, diversos mecanismos han sido postulados para explicar las
lesiones producidas en los tejidos de pacientes con Enfermedad de Chagas (Figura
9). La teoría de la persistencia del parásito, procede de la evidencia de nidos
chagásicos encontrados en el corazón de un niño que murió de Enfermedad de
Chagas aguda (Vianna, 1911). En el año de 1960, un primer informe de miocarditis
Chagásica aguda mostró el parasitismo latente, en contraste con las lesiones
inflamatorias activas (Torres, 1960). No obstante, una reciente investigación pone de
manifiesto la ausencia de nidos parasitarios en las secciones histológicas de corazón
de un gran número de pacientes chagásicos crónicos (Teixeira et al., 2009), lo que
conlleva al cuestionamiento de la existencia de esta teoría.
Figura 9. Mecanismos causales de miocarditis en la infección con T. cruzi. A) Daño directo generado
por la persistencia del protozoario o productos tóxicos parasitarios. B) Daños desencadenados por la
respuesta inmune específica. Se evidencia desplazamiento de miocitos, seguido por la infiltración de
mononucleares y fibrosis. C) Daños no específicos causados por la respuesta inmune innata,
activación de granulocitos y citotoxicidad mediada por anticuerpos. D) Isquemia producida por la
microvasculopatía. E) Autoinmunidad inducida por el parásito, a causa del mimetismo molecular entre
el protozoo y los antígenos propios (Bonney y Engman, 2015).

Introducciön
16
La lisis celular producida por T. cruzi, desencadenada por la multiplicación de
amastigotes y liberación adyacente de tripomastigotes, constituye la causa principal
del daño en el tejido cardiaco. Sin embargo, la presencia de una robusta infiltración
leucocitaria y mediadores inflamatorios liberados por los linfocitos, macrófagos,
eosinófilos, neutrófilos y mastocitos, acrecientan daños más graves en los
cardiomiocitos (Dos Santos y Hudson, 1980; Bonney y Engman, 2015). Se ha
evidenciado que T. cruzi es capaz de perturbar el sistema bradicinina mediante la
activación de los receptores B2 en la invasión de las células endoteliales, lo que lleva
a la vasodilatación y edema intersticial posterior (Todorov et al., 2003)
Por otra parte, se ha demostrado que el daño oxidativo a los complejos respiratorios
mitocondriales en células infectadas con T. cruzi, conducen a la reducción de
generación de ATP mitocondrial y deficiencias en los mecanismos antioxidantes
celulares, como niveles reducidos de la superóxido dismutasa (Wen et al., 2004).
El hallazgo de pérdidas significativas de células neuronales en pacientes con
enfermedad de Chagas y ausencia de T. cruzi, constituye la base de la hipótesis de la
liberación de neurotoxinas generadas por el parásito oculto en alguna parte del
cuerpo humano (Köeberle, 1970). Sin embargo, a la fecha, la neurotoxina hipotética
nunca ha sido demostrada. En este contexto, la ausencia de cualquier tipo de toxina
producida por el protozoo, sugiere que las anormalidades autoinmunes dependientes
en el sistema nervioso autónomo, pueden perpetuar el ciclo de la cardiotoxicidad por
catecolaminas, miocitólisis e insuficiencia cardiaca (Dávila, 2004; Dávila et al.,
2005).
Por otro lado, la teoría de la autoinmunidad sugiere que el daño cardíaco produce una
alteración de la auto-tolerancia, lo que resulta en una reacción inmune contra las
proteínas propias. Se ha sugerido, que la autoinmunidad es desencadenada por daños
parasitarios inducidos en los cardiomiocitos (Fujinami et al., 2006; Hyland et al.,
2007) y por mimetismo molecular entre epítopos inmunológicamente similares del
protozoario y las proteínas del huésped (Cunha-Neto et al., 2006). Durante la
infección, daños mecánicos causados directamente por T. cruzi y una robusta
respuesta inmune, dan lugar a la liberación de grandes cantidades de antígenos
propios, en un entorno particularmente rico en mediadores inflamatorios (Engman y
Leon, 2002). Este medio propicio de estímulos inmunes, supera el umbral de
activación necesario para la ruptura de la auto-tolerancia, lo que resulta en la
autoinmunidad dirigida contra múltiples antígenos. Por su parte, factores
inflamatorios como el interferón-γ y el óxido nítrico, promueven la activación de
células T autorreactivas (Cardillo et al., 2015).

Introducciön
17
1.1.6 Manifestaciones clínicas
La Enfermedad de Chagas se desarrolla en tres fases definidas, las cuales presentan
una sintomatología variable y en ocasiones inespecífica (Figura 10).
Figura 10. Fases de la infección por T. cruzi. La fase aguda de la infección por T. cruzi se caracteriza
por la detección microscópica del parásito durando 8 a 12 semanas. Las personas con enfermedad
crónica sin manifestaciones clínicas, cursan con la forma indeterminada de la enfermedad. Se estima
que del 20 a 30% de las personas que inicialmente tienen la fase indeterminada, desarrollan a lo largo
de los años enfermedades cardíacas o gastrointestinales (Bern, 2015).
1.1.6.1 Fase aguda
La fase aguda se caracteriza por un cuadro clínico inespecífico que puede durar cerca
de 4 a 8 semanas, y en algunas ocasiones, se puede extender hasta 4 meses. Durante
el período de incubación de T. cruzi (72 horas), el protozoo sufre ciclos de

Introducciön
18
replicación en la célula huésped y el sistema inmune desencadena una respuesta
inflamatoria de la piel (chagoma: nódulo subcutáneo, redondeado, eritematoso duro e
indoloro) o la conjuntiva (edema bipalpebral unilateral conocido como signo de
Romaña). Estas lesiones se presentan en menos del 5% de los casos con
manifestaciones clínicas. Los síntomas característicos de la enfermedad incluyen
malestar general, fiebre no mayor de 38.5°C, la cual es de larga duración y constante;
dolor de cabeza, dolor articular y muscular, anorexia, vómitos, diarrea, somnolencia,
apatía, linfadenopatía, hepatoesplenomegalia, edema y convulsiones (Rassi et al.,
2012; Bern, 2015). Las infecciones agudas pueden ser detectadas por la presencia de
anticuerpos IgM específicos contra T. cruzi o frotis sanguíneos coloreados con
Giemsa, en donde se observa directamente el parásito.
En la Enfermedad de Chagas aguda, la tasa de mortalidad es inferior al 10% y la
principal causa de muerte es atribuida a la insuficiencia cardiaca, meningitis o
encefalitis (Rassi et al., 2012; Bern, 2015; Sanabria, 2016).
1.1.6.2 Fase indeterminada
En esta fase de la enfermedad la mayoría de los individuos inmunocompetentes se
encuentran asintomáticos y debido a la eficacia de la inmunidad adaptativa
específica, el parásito logra ser mantenido en bajos o indetectables niveles (Bonney y
Engman, 2015). Se estima que más de dos tercios de personas infectadas con T.
cruzi, permanecen en la fase intermedia durante toda su vida. Por lo general, esta fase
se detecta durante pruebas de admisión de trabajo o tamizaje en los donantes de
sangre. La esperanza de vida de estos pacientes es similar a las registradas para los
individuos no infectados de la misma región (Rassi et al., 2012; Bern, 2015).
1.1.6.3 Fase crónica
Las manifestaciones clínicas de la Enfermedad de Chagas crónica, se observa en
menos de un tercio de los individuos que cursan con la fase indeterminada de la
infección, después de aproximadamente 25 años a partir de la adquisición del
protozoario (Prata, 2001). Entre los pacientes chagásicos con manifestaciones
clínicas, el 94,5% de los casos se verán afectados por problemas de corazón; en
dónde el 38,5% de estos pacientes mueren repentinamente y el 56% desarrollan
insuficiencia cardíaca. El 5,5% de los individuos restantes, presentan síndromes
digestivos como el megaesófago o megacolon. La muerte por enfermedad de Chagas
crónica es a menudo el resultado de la insuficiencia cardíaca congestiva después de
un desarrollo gradual de la disfunción miocárdica debido al daño causado por la
inflamación crónica (Teixeira et al., 2006; Rassi et al., 2012).

Introducciön
19
1.1.6.4 Enfermedad de Chagas congénita
Las manifestaciones clínicas de la enfermedad de Chagas congénita varían desde
casos asintomáticos hasta infecciones mortales; las cuales están relacionadas con el
nivel de parasitemia al nacer (Torrico et al., 2004a; Torrico et al., 2004c). Los signos
y síntomas de la enfermedad pueden estar presentes en el nacimiento, o aparecer en
cuestión de días posteriores al parto (Carlier et al., 2011). La prevalencia de
infección congénita asintomática oscila entre un 40% y 100% (Blanco et al., 2000;
Torrico et al., 2004b). Sin un tratamiento eficaz, los recién nacidos podrían
desarrollar la fase indeterminada de la enfermedad, e inclusive algunos de ellos
presentarían la enfermedad crónica con las manifestaciones típicas cardíacas y
gastrointestinales (Schijman, 2006).
1.1.6.5 Enfermedad de Chagas oral
El periodo de incubación de esta infección, oscila entre 3 a 22 días después de
consumido el protozoario. Las manifestaciones clínicas incluyen fiebre prolongada,
náuseas, vómito, diarrea, cefalea, mialgias, dolor epigástrico, artralgia,
hepatoesplenomegalia, miocarditis, derrame pericárdico y taponamiento cardiaco que
genera finalmente la muerte. Esta patología presenta una alta tasa de mortalidad y se
considera de peor pronóstico entre los pacientes más jóvenes (Bern, 2015; Díaz et al.,
2015).
1.1.7 Tratamientos clásicos
Actualmente, el Benznidazol (BNZ) y Nifurtimox (NFX), introducidos en la terapia
clínica aproximadamente hace más de 40 años, constituyen los dos únicos fármacos
disponibles y de elección para la Enfermedad de Chagas (Bustamante y Tarleton,
2014; Bern, 2015; Benziger et al., 2017). Estas drogas se proporcionan con
frecuencia a todos los niños con infección y personas con enfermedad crónica hasta
la edad de 50 años sin enfermedad cardíaca, y están contraindicadas en el embarazo y
en pacientes con enfermedad renal o hepática avanzada (Rassi et al., 2012).
1.1.7.1 Benznidazol
El Benznidazol (N-bencil-2- (2-nitro-1H-imidazol-1-il) acetamida) es un compuesto
nitro-heterocíclico desarrollado por Roche (USA) (Figura 11A) y utilizado como
tratamiento de primera elección para la Enfermedad de Chagas, desde hace más de 4
décadas (Rassi et al., 2012; Bern, 2015).

Introducciön
20
Figura 11. Estructuras del Benznidazol (A) y Nifurtimox (B) (Castro et al., 2015)
El tratamiento con BNZ, se administra generalmente por vía oral durante 60 días, con
una dosis diaria de 5-7mg/kg para adultos y 10 mg/kg para niños (OMS, 2015). Diversas
investigaciones han evidenciado que la baja solubilidad del BNZ combinada con altas
dosis de tratamiento durante un largo período de tiempo, desencadena una serie de
reacciones adversas que incluyen erupciones cutáneas, dermatitis exfoliativa,
fotosensibilidad, supresión de la médula ósea (trombocitopenia, neutropenia y
agranulocitosis) y neuropatía periférica (Weiss et al., 2011; Rassi et al., 2012). La
biodisponibilidad del fármaco también se ve afectada por su baja solubilidad,
ocasionando una disminución de su efectividad durante la fase crónica de la infección
(Weiss et al., 2011).
El Benznidazol y Nifurtimox actúan mediante la formación de metabolitos electrofílicos
y/o radicales libres (Figura 12). El grupo nitro (tipo R-NO2) de ambos fármacos se
reduce a un grupo amino por acción de la NADPH - Citocromo P450 Reductasa,
ocasionando un anión radical nitro (R-NO2-) derivado del NFX y metabolitos
electrofílicos procedentes del BNZ. Este radical experimenta un ciclo redox con oxígeno
molecular, y por acción de la Superóxido Dismutasa (SOD) se produce O2 y H2O2. El
anión superóxido (O2 U-) y el H2O2, en presencia de Fe3+ forman el radical hidroxilo
(OH-) (reacción de Haber-Weiss), el cual se une a lípidos, proteínas y ADN parasitario,
produciendo la destrucción del protozoo (Maya et al., 2007). Por su parte, el efecto
tripanocida del BNZ no depende principalmente de la producción de radicales, si no de
la formación de metabolitos electrofílicos que se unen de manera covalente a
macromoléculas y generan un daño a las células (Maya et al., 2004; Apt et al., 2008).
Figura 12. Mecanismo de acción del Benznidazol y Nifurtimox (Maya et al., 2007).

Introducciön
21
En el año 2013, Pinazo et al, analizaron 54 pacientes tratados con BNZ, con el fin de
determinar una posible asociación entre las reacciones adversas y la concentración de
BNZ en suero. Aunque los autores no encontraron diferencias estadísticamente
significativas entre las reacciones desencadenadas en los pacientes que detuvieron el
tratamiento con respecto a aquellos que lo terminaron; ellos pudieron determinar que
el BNZ administrado a 5 mg/kg/día resultó en altas concentraciones tripanocidas (3-6
μg/mL), que indican una dosis óptima terapéutica en suero (Pinazo et al., 2013).
1.1.7.2 Nifurtimox
Nifurtimox (N-(3-metil-1,1-dioxo-1,4-tiazinan-4-il)- 1-(5-nitro-2-furil) metanimina)
constituye el tratamiento principal para la enfermedad de Chagas en los Estados
Unidos (Figura 11B). Durante más de 10 años, la compañía farmacéutica Bayer ha
suministrado gratuitamente a la OMS 1 millón de tabletas al año, con asistencia
financiera para su distribución, primordialmente en Honduras y El Salvador
(Martínez-Parra et al., 2015).
El tratamiento con NFX se administra generalmente, de acuerdo a las
recomendaciones de la OMS: de 8 a10 mg /kg diarios en tres dosis divididas para
adultos y de 15 a 20 mg /kg diarios en cuatro dosis divididas para los niños, durante
60 a 90 días (OMS, 2015). Los efectos adversos predominantes del medicamento son
las molestias gastrointestinales, tales como náuseas, vómitos y dolor abdominal. No
obstante, un aproximado del 30% de los pacientes experimentan perturbaciones del
sistema nervioso central como polineuritis, confusión, ataques focales y/o
generalizados, e incluso psicosis, que son superadas cuando el tratamiento se detiene
(Weiss et al., 2011; Rassi et al., 2012).
En general, NFX ha demostrado mayor toxicidad y efectos adversos que BNZ,
incluyendo el aumento del estrés oxidativo en páncreas de rata (Mecca et al., 2007) y
corazón (Mecca et al., 2008). En este contexto, el daño cardíaco causado por el
parásito durante una infección crónica, podría aumentar con la administración de
dicho medicamento. Es por ello, que el NFX no representa el tratamiento de elección
para la Enfermedad de Chagas, en la mayoría de los países endémicos.
El mecanismo de acción del fármaco se basa en la reducción cíclica del grupo nitro,
produciéndose un nitro radical que sufre auto-oxidación, y formación del superóxido
(Figura 12) (Maya et al., 2007). La Nitrorreductasa tipo 1, constituye la enzima
principal implicada en la bioactivación de los fármacos nitroheterocíclicos. En este
contexto, el deterioro de su actividad confiere resistencia a ambos fármacos, NFX y
BNZ (Wilkinson et al., 2008). Hall et al. (2012) demostraron que la reducción de
BNZ genera un derivado de hidroxilamina que finalmente se convierte en glioxal, un
compuesto altamente citotóxico y mutagénico. No obstante, este hallazgo fue
contrarrestado dos años después, en un estudio de metabolómica realizado en T. cruzi

Introducciön
22
tratados con BNZ, en dónde no se detectaron niveles de glioxal (Trochine et al.,
2014).
A pesar de sus efectos secundarios indeseables y la existencia de cepas de parásitos
naturalmente resistentes a los dos compuestos (Filardi y Brener, 1987; Campos et al.,
2013), la principal limitación del uso del NFX y el BNZ, radica en su ineficiente
actividad tripanocida en la fase crónica de la enfermedad (Oliveira et al., 2015;
Urbina, 2015).
En este contexto, dos estudios clínicos (TRAENA (Tratamiento en pacientes adultos)
y BENEFIT (Evaluación del Benznidazol para Interrumpir la Tripanosomiasis))
realizados en una cohorte de pacientes adultos con enfermedad de Chagas crónica,
fueron ejecutados en los últimos años (Zingales et al., 2014; Morillo et al., 2015;
Oliveira et al., 2015; Urbina, 2015). Ambas investigaciones demostraron la
ineficiencia del BNZ para reducir el deterioro clínico cardiaco, morbilidad o
mortalidad de los participantes, a pesar de su efecto tripanocida (Morillo et al., 2015;
Oliveira et al., 2015; Urbina, 2015).
Por su parte, desde el año 2015 un ensayo clínico de fase II/fase III
(CHICAMOCHA-3) empezó a ejecutarse, con la finalidad de deducir la eficacia del
Nifurtimox. El estado actual de esta investigación permanece en curso
(ClinicalTrials.gov).
1.1.8 Tratamientos alternativos
Debido a que los tripanosomatideos y hongos sintetizan ergosterol de manera similar,
fármacos antifúngicos tales como el Posaconazol y Ravuconazol, que actúan
inhibiendo la biosíntesis del ergosterol, han sido estudiados en los últimos años para
evaluar su actividad antichagásica. Estos azoles han mostrado resultados promisorios
en estudios preclínicos, con potentes y selectivos hallazgos frente a amastigotes
intracelulares y modelos murinos en fase aguda de la enfermedad (Urbina, 2009;
Urbina, 2010; Urbina, 2015; Keenan et al., 2015). No obstante, la efectividad de
estas moléculas en estudio clínicos, no ha sido exitosa. CHAGAZASOL determinó la
eficacia y seguridad del Posaconazol en comparación con el BNZ en pacientes
adultos con Chagas crónico (Molina et al., 2014); y STOP CHAGAS
(ClinicalTrials.gov) evaluó la eficacia de esta misma molécula, pero en individuos
que cursaban con la fase indeterminada sin compromiso cardiaco.
Desafortunadamente, en ambos estudios no se evidenció curación parasitaria, ni
tratamiento exitoso. Respecto a la toxicidad y seguridad, ningún efecto adverso grave
fue observado (Keenan et al., 2015).

Introducciön
23
El Fexinidazol representa otro compuesto nitroheterocìclico redescubierto en la
exploración de nitroimidazoles como drogas potenciales para la tripanosomiasis
africana humana. Esta molécula ha mostrado un amplio espectro antiprotozoario, con
buenos resultados en la investigación preclínica (Bahia et al., 2012).
1.1.8.1 Aceites esenciales
Los aceites esenciales (AEs) son extractos volátiles obtenidos a través de destilación
al vapor de diferentes partes de plantas aromáticas (flores, semillas, hojas, ramas,
cortezas frutos y raíces) (Stashenko, 2009); y su principal actividad biológica radica
en actuar como mecanismos de defensa frente a la acción de diferentes
microorganismos fitopatógenos (Bakkali et al., 2008, Schelz et al., 2010).
Durante las últimas décadas, los AEs y sus componentes mayoritarios han sido
descritos como agentes antimicrobianos de amplio espectro farmacológico (Ramos et
al., 2014), con actividad significativa antihelmíntica y antiprotozoaria (Bakkali et al.,
2008; Escobar et al., 2010; Cheikh Ali et al., 2011; Alviano et al., 2012; Borges et
al., 2012; York et al., 2012). Algunos terpenos constituyentes de estas fracciones
aceitosas, tales como citral (Lippia alba y Cymbopogon citratus), óxido de
cariofileno (Aframomum sceptrum, Achillea millefolium y Piper var brachypodon),
limoneno (L. origanoides y L. pedunculosa) y carvona (Origanum vulgare) han
mostrado actividad tripanocida eficiente en formas extra e intracelulares de T. cruzi
(Santoro et al., 2007a; Santoro et al., 2007b; Santoro et al., 2007c; Cardoso y Soares,
2010; Escobar et al., 2010; Kpoviessi et al., 2014; Santos et al., 2014) (Tabla 2).
En el año 2007, Santoro et al., demostraron el efecto tripanocida de AEs derivados
de Cymbopogon citratus contra T. cruzi, obteniendose CI50 significativas en los tres
estadíos parasitarios (Epimastigotes: 126,5 µg/mL; Tripomastigotes: 15,5 µg/mL;
Amastigotes: 5,1µg/mL) después de 24 horas de tratamiento. Estos resultados fueron
atribuidos a su componente principal, el citral: una mezcla de neral y geranial
(Santoro et al., 2007a). En ese mismo año, este grupo de investigadores evidenció el
efecto antiproliferativo de AEs extraídos de la mata de clavo (Syzigium aromaticum),
la albahaca (Ocimum basilicum) y la milenrama (Achillea millefolium) sobre formas
extracelulares de T. cruzi. En esta investigación, el aceite de clavo, cuyo compuesto
principal lo constituyó el Eugenol, evidenció un valor de CI50 de 99,5 µg/mL sobre
epimastigotes y 57,5 µg/mL para Tripomastigotes (Santoro et al., 2007c).
Introducciön
24
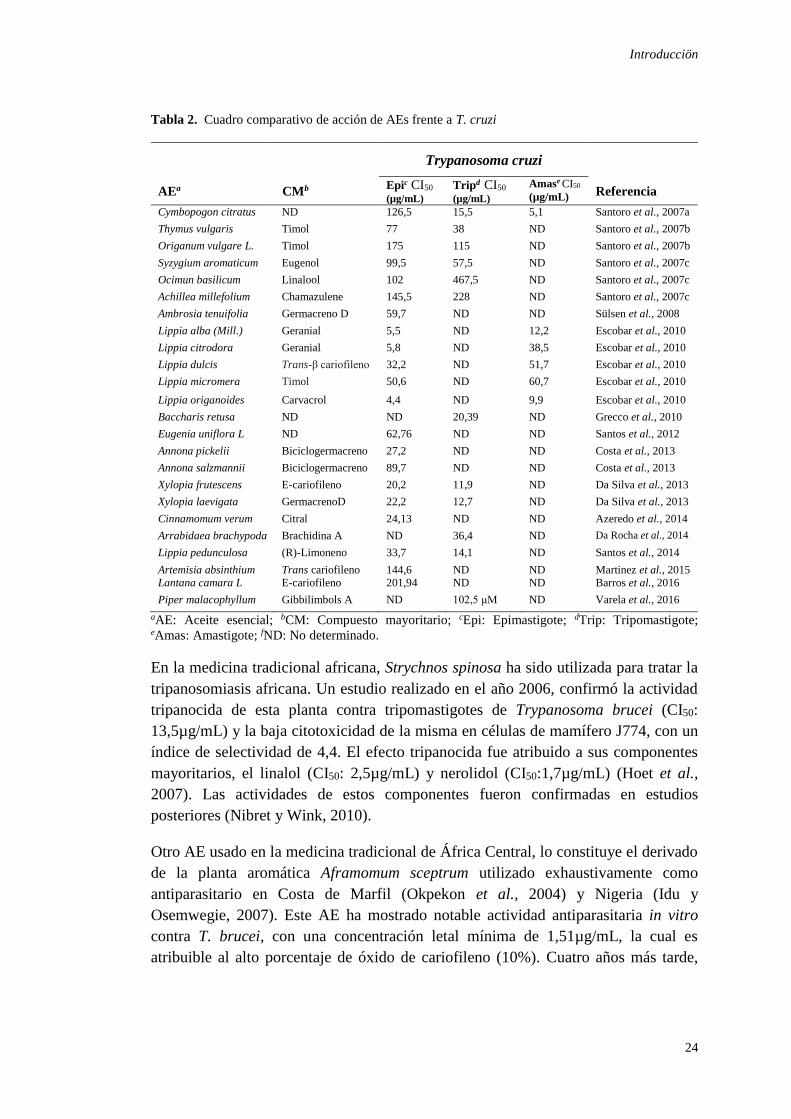
Tabla 2. Cuadro comparativo de acción de AEs frente a T. cruzi
Trypanosoma cruzi
AEa CMb Epic CI50
(µg/mL) Tripd CI50
(µg/mL)
Amase CI50
(µg/mL) Referencia
Cymbopogon citratus ND 126,5 15,5 5,1 Santoro et al., 2007a
Thymus vulgaris Timol 77 38 ND Santoro et al., 2007b
Origanum vulgare L. Timol 175 115 ND Santoro et al., 2007b
Syzygium aromaticum Eugenol 99,5 57,5 ND Santoro et al., 2007c
Ocimun basilicum Linalool 102 467,5 ND Santoro et al., 2007c
Achillea millefolium Chamazulene 145,5 228 ND Santoro et al., 2007c
Ambrosia tenuifolia Germacreno D 59,7 ND ND Sülsen et al., 2008
Lippia alba (Mill.) Geranial 5,5 ND 12,2 Escobar et al., 2010
Lippia citrodora Geranial 5,8 ND 38,5 Escobar et al., 2010
Lippia dulcis Trans-β cariofileno 32,2 ND 51,7 Escobar et al., 2010
Lippia micromera Timol 50,6 ND 60,7 Escobar et al., 2010
Lippia origanoides Carvacrol 4,4 ND 9,9 Escobar et al., 2010
Baccharis retusa ND ND 20,39 ND Grecco et al., 2010
Eugenia uniflora L ND 62,76 ND ND Santos et al., 2012
Annona pickelii Biciclogermacreno 27,2 ND ND Costa et al., 2013
Annona salzmannii Biciclogermacreno 89,7 ND ND Costa et al., 2013
Xylopia frutescens E-cariofileno 20,2 11,9 ND Da Silva et al., 2013
Xylopia laevigata GermacrenoD 22,2 12,7 ND Da Silva et al., 2013
Cinnamomum verum Citral 24,13 ND ND Azeredo et al., 2014
Arrabidaea brachypoda Brachidina A ND 36,4 ND Da Rocha et al., 2014
Lippia pedunculosa (R)-Limoneno 33,7 14,1 ND Santos et al., 2014
Artemisia absinthium Trans cariofileno 144,6 ND ND Martinez et al., 2015
Lantana camara L E-cariofileno 201,94 ND ND Barros et al., 2016
Piper malacophyllum Gibbilimbols A ND 102,5 μM ND Varela et al., 2016
aAE: Aceite esencial; bCM: Compuesto mayoritario; cEpi: Epimastigote; dTrip: Tripomastigote; eAmas: Amastigote; fND: No determinado.
En la medicina tradicional africana, Strychnos spinosa ha sido utilizada para tratar la
tripanosomiasis africana. Un estudio realizado en el año 2006, confirmó la actividad
tripanocida de esta planta contra tripomastigotes de Trypanosoma brucei (CI50:
13,5µg/mL) y la baja citotoxicidad de la misma en células de mamífero J774, con un
índice de selectividad de 4,4. El efecto tripanocida fue atribuido a sus componentes
mayoritarios, el linalol (CI50: 2,5µg/mL) y nerolidol (CI50:1,7µg/mL) (Hoet et al.,
2007). Las actividades de estos componentes fueron confirmadas en estudios
posteriores (Nibret y Wink, 2010).
Otro AE usado en la medicina tradicional de África Central, lo constituye el derivado
de la planta aromática Aframomum sceptrum utilizado exhaustivamente como
antiparasitario en Costa de Marfil (Okpekon et al., 2004) y Nigeria (Idu y
Osemwegie, 2007). Este AE ha mostrado notable actividad antiparasitaria in vitro
contra T. brucei, con una concentración letal mínima de 1,51µg/mL, la cual es
atribuible al alto porcentaje de óxido de cariofileno (10%). Cuatro años más tarde,

Introducciön
25
Cheikh Ali et al. (2011), evidenciaron una concentración letal menor (0,1µg/mL) de
este AE en T. brucei.
Por su parte, un estudio realizado en Colombia en el año 2010, puso de manifiesto
que especies de Lippia spp. (Verbenaceae) ricas en neral, geranial, geraniol y timol,
son particularmente activas contra T. cruzi. El AEs de L. alba exhibió la más alta
actividad frente a epimastigotes (CI50: 5,5 μg/mL) y amastigotes intracelulares (CI50:
12,2 μg/mL). Por su parte, los terpenos timol (CI50: 3,2 μg/mL) y S-carvona (CI50:
6,1 μg/mL), constituyeron los compuestos más activos en amastigotes intracelulares
de T. cruzi con un índice selectivo superior a 10 (Escobar et al., 2010).
1.2 Lippia alba
Lippia alba (L. alba) es un arbusto aromático perteneciente a la familia Verbenaceae
y nativo de América Latina. Se conoce popularmente como prontoalivio (Colombia),
cidreira (Brasil) y salvia morada en Argentina. Crece en regiones tropicales y
subtropicales con alturas máximas de 2300 m.s.n.m, (Vit et al., 2002; Parra et al.,
2010), llegando a alcanzar longitudes de hasta 2 metros. Sus hojas verdes se
desarrollan en forma peciolada, asociadas a flores pequeñas (3-5 mm) de tonalidades
rosas, violetas, blancas o amarillas (Guzmán et al., 2004; Parra et al., 2010) (Figura
13).
Figura 13. Lippia alba (Mill.) N.E. Brown.
La planta presenta alta plasticidad fenotípica, atribuida a su versatilidad genética,
ontogenia, ubicación geográfica e influencia de factores medioambientales (clima,
disponibilidad de nutrientes, luminosidad, condiciones de colecta y siembra) (Palacio
et al, 2007; Parra et al, 2007; Cicció y Ocampo, 2010; Teles et al, 2012; Ehlert et al,
2013); y una amplia diversidad fitoquímica (Tabla 3).

Introducciön
26
Tabla 3. Diversidad fitoquímica de Lippia alba (Modificado de Cicció y Ocampo 2010).
País Componentes Mayoritarios
Argentina Lippiona, Dihidrocarvona, Citral, 1-8-Cineol, Piperitona
(36,7%), Limoneno (34,2%).
Brasil Linalool (55%), γ-Terpineno (46,7%), β-Cariofileno
(24,3%), Geranial (12,9), Neral (9,6%), 2-undecanona
(9,0%), p- Cimeno (8,7%).
Colombia Carvona (41-57%), Limoneno (24-37%), Mircenona (39-
63%), Geranial (30,5%), Neral (23,6%).
Costa Rica
Carvona (62,4%), Limoneno (20,8%-25%), Mircenona.
Cuba Carvona (29-40%), β-Guayeno (9,8-11,5%) Piperitenona
(6,4-8,3%), Limoneno (5,8-6.5%).
Estados
Unidos
Metil-chavicol (56,5%), 1,8-Cineol (12,6%), Cariofileno
(7%).
Guatemala Mircenona (37-58,2%), Geranial (27%), 1,8-cineol
(25,4%), Neral (18,9%), Omicenona (16,3%).
Guayana
Francesa
Carvona (67,8%), Limoneno (18,1%)
Perú Carvona (63,4%), Germacreno D (5,6%), Limoneno
(5,1).
Región Caribe Neral (33,1%), Geranial (20,8%), α-Guayeno (15,4%), β-
Ocimeno (13,3).
Uruguay Alcanfor (18,2%), 1-8-Cineol (16,5%)
La variedad química de L. alba sugiere la aparición de múltiples quimiotipos
(individuos de una misma especie con composición química diferente). En este
contexto, Hennebelle et al., (2006) propusieron siete quimiotipos basados en los
componentes mayoritarios del aceite esencial.
Quimiotipo I o Citral: representado por AEs con alto contenido de Citral (Mezcla de
Geranial y Neral); Linalool y β-Cariofileno. Esta variedad ha sido reportada en
diferentes regiones de Colombia (Arauca, Bolívar y Bucaramanga). Así mismo,
análisis de espectrometría de masas han permitido determinar algunos constituyentes
adicionales como geraniol (6,3%), 6-metil-5-heptano-2-ona (6%) y limoneno (3,7%)
(Mesa et al., 2009).
Quimiotipo II o Tagetenona: caracterizado por presentar altos contenidos de
Tagetenona, un terpeno conformado por las cetonas ocimenona y mircenona.
Quimiotipo III o Carvona: constituido por cetonas cíclicas (Dihidrocarvona,
Piperitenona, Lippiona o Carvona) y Limoneno. Ha sido descrito en Colombia con
Introducciön
27
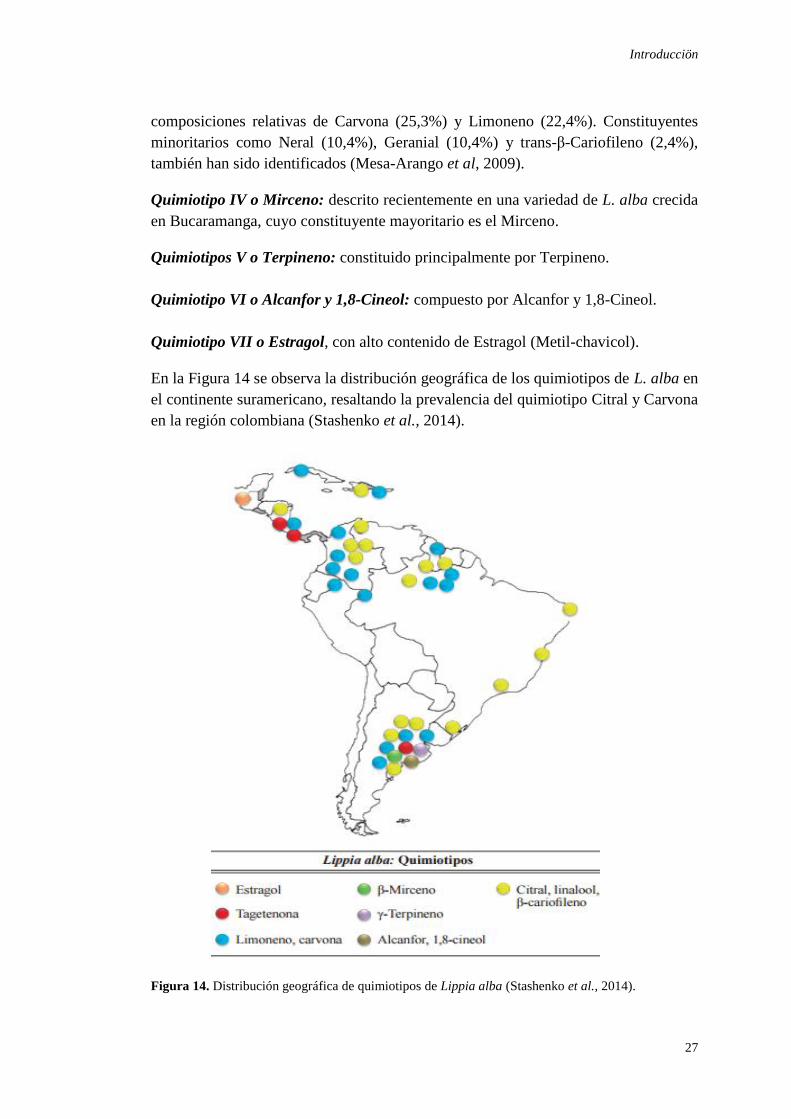
composiciones relativas de Carvona (25,3%) y Limoneno (22,4%). Constituyentes
minoritarios como Neral (10,4%), Geranial (10,4%) y trans-β-Cariofileno (2,4%),
también han sido identificados (Mesa-Arango et al, 2009).
Quimiotipo IV o Mirceno: descrito recientemente en una variedad de L. alba crecida
en Bucaramanga, cuyo constituyente mayoritario es el Mirceno.
Quimiotipos V o Terpineno: constituido principalmente por Terpineno.
Quimiotipo VI o Alcanfor y 1,8-Cineol: compuesto por Alcanfor y 1,8-Cineol.
Quimiotipo VII o Estragol, con alto contenido de Estragol (Metil-chavicol).
En la Figura 14 se observa la distribución geográfica de los quimiotipos de L. alba en
el continente suramericano, resaltando la prevalencia del quimiotipo Citral y Carvona
en la región colombiana (Stashenko et al., 2014).
Figura 14. Distribución geográfica de quimiotipos de Lippia alba (Stashenko et al., 2014).

Introducciön
28
1.2.1 Etnofarmacología
L. alba representa la séptima especie más citada en la medicina tradicional de Brasil
(Hennebelle et al., 2008). Los “curanderos” utilizan sus hojas como una infusión
para tratar problemas de hipertensión, digestivos, resfriados y curación de heridas
locales (Di Stasi et al., 2002; Oliveira et al., 2006; Pinto et al., 2006). En Boyacá,
Colombia, se usa frecuentemente como analgésico y para problemas digestivos
(diarrea, dolor de estómago) y respiratorios (Gripa y tos) (Toscano-González, 2006).
La administración por vía oral representa la vía más citada y según Hennebelle et al.,
(2008) sus usos principales están constituidos por problemas digestivos,
respiratorios, cardiovasculares y sedativos.
1.2.2 Actividad biológica de L. alba
La fitoterapia se ha convertido en una alternativa muy importante de diversas
patologías, dado los efectos secundarios que se atribuyen a las drogas sintéticas.
Durante las últimas décadas, L. alba y sus constituyentes mayoritarios han
demostrado gran potencial terapéutico, como analgésicos, antiinflamatorios,
antiespasmódicos (Haldar et al., 2012; Mamun-OrRashid et al., 2013) y
antimicrobianos de amplio espectro (Escobar et al., 2010; Fabri et al., 2011, Nuñez et
al., 2012; Gómez et al., 2013; Olivero-Verbel et al., 2014; Vera et al., 2014).
Extractos etanólicos de esta variedad, presentan amplia actividad antibacteriana
principalmente contra Staphylococcus aureus (Oliveira et al., 2006; Sena Filho et al.,
2006) y otros microorganismos (Aguiar et al., 2008).
El mayor número de estudios antimicrobianos reportados, han sido principalmente
sobre la actividad de los AEs de L. alba como agentes anti fúngicos en fitopatógenos
(Shukla et al., 2009; Anaruma et al., 2010; Tomazoni et al., 2016) y patógenos
humanos (Mesa-Arango et al., 2009; Costa et al., 2014; Geromini et al., 2015).
Sin embargo, en el último año, estudios publicados referentes a la actividad biológica
de este arbusto aromático, han sido referidos al efecto antitumoral y citotóxico sobre
células leucémicas humanas (Bai et al., 2016; García et al., 2017). Además, L. alba
también ha evidenciado tener acción anestésica y antioxidativa (Cunha et al., 2010;
Azambuja et al., 2011; Barbas et al., 2017). Estas propiedades bioactivas son
atribuidas principalmente a sus constituyentes mayoritarios tales como citral,
geranial, óxido de cariofileno, carvona, neral, linalool y limoneno (Hennebelle et al.,
2008; García et al., 2017).

Introducciön
29
1.2.3 Actividad tripanocida de L. alba
A la fecha, son escasos los estudios reportados en la literatura, específicos de la
actividad tripanocida ejercida por L. alba y/o sus compuestos mayoritarios.
En Colombia, un estudio realizado en el año 2010, puso de manifiesto que especies
de Lippia spp. (Verbenaceae) ricas en neral, geranial, geraniol y timol, presentaban
actividad contra T. cruzi. El AEs de L. alba exhibió efecto tripanocida frente a
epimastigotes (CI50: 5,5 μg/mL) y amastigotes intracelulares (CI50: 12,2 μg/mL). Por
su parte, los terpenos timol (CI50: 3,2 μg/mL) y S-carvona (CI50: 6,1 μg/mL),
constituyeron los compuestos más activos en amastigotes intracelulares de T. cruzi
con un índice selectivo superior a 10 (Escobar et al., 2010).
En ese mismo año, Cardoso y Soares (2010) evaluaron el efecto del citral
(componente mayoritario del quimiotipo Citral de L. alba) sobre el proceso de
metaciclogénesis en T. cruzi. Ellos demostraron que bajas concentraciones de citral
(20 μg/mL) no afectan la viabilidad del epimastigote, ni interfieren en el proceso de
diferenciación. No obstante, concentraciones superiores a 60 μg/mL, producían un
porcentaje de muerte significativo (100%). Además, determinaron que la
concentración calculada que inhibió el 50% de la metaciclogénesis fue de 31 μg/mL
después de 24 horas de tratamiento. Por otra parte, los tripomastigotes derivados de
células demostraron igual suceptibilidad al citral, con una CI50 de 24,5 μg/mL
(Cardoso y Soares, 2010).
Recientemente, Santos et al. (2014) determinó que el limoneno (componente
mayoritario del quimiotipo Carvona de L. alba) posee actividad tripanocida in vitro
en las formas extracelulares de T. cruzi (Epimastigote: CI50 33,7 μg/mL;
Tripomastigotes: CI50 14,1 μg/mL). El tratamiento con este terpeno, causó además
una reducción moderada en el porcentaje de macrófagos infectados y en el número
de parásitos intracelulares, en concentraciones no tóxicas para las células mamíferas
(Santos et al., 2014).
1.2.4 Terpenos de L. alba
En las últimas décadas, las especies de Lippia han sido química y
farmacológicamente investigadas. Se ha determinado que especies pertenecientes a
la familia Verbenaceae, son productores constantes de monoterpenos y
sesquiterpenos, como el limoneno, citral, óxido de cariofileno y carvona (Hennebelle
et al., 2006) (Figura 15), entre otros compuestos (alcaloides, carotenoides,
flavonoides, iridoides, ácidos fenólicos, saponinas, esteroles, azúcares, taninos),
responsables de los efectos biológicos de los aceites esenciales.

Introducciön
30
Figura 15. Estructuras químicas de terpenos encontrados en aceites esenciales de L. alba.
1.2.4.1 Citral
Citral, un monoterpeno aldehído sintetizado por varios géneros de plantas, es una
mezcla de geranial y neral, con fuertes actividades antimicrobianas demostradas
(Cardoso y Soares, 2010; Saddiq y Khayyat, 2010; Jafri et al., 2014). Constituye el
principal componente del aceite esencial de la hierba limón y tiene actividad contra
T. cruzi (Santoro et al., 2007a). Resultados tripanocidas similares han sido
encontrados en aceites derivados de una variante Colombiana (Santander) de Lippia
alba quimiotipo Citral, los cuales eran ricos en citral (mezcla de neral y geranial),
geraniol timol y óxido de cariofileno (Escobar et al., 2010). A este metabolito se le
ha reportado la capacidad de inducir estrés oxidativo y apoptosis en células
tumorales, mediada por el aumento de p53, como consecuencia del daño oxidativo a
nivel del ADN y del estrés del retículo endoplasmático (Ji et al., 2006; Kapur et al.,
2016; Thomas et al., 2016). Además se le han anexado cualidades interesantes como
agente quimioprotector (efecto anti-oxidante cinco veces superior al del α-tocoferol)
(Stashenko et al., 2014) y con actividad inhibitoria fuerte sobre la expresión
citoquinas y mediadores inflamatorios (Liao et al., 2015).
1.2.4.2 Óxido de Cariofileno
El óxido de cariofileno es un sesquiterpeno bicíclico natural, constituyente de varios
AEs y con propiedad antimicrobiana de amplio espectro (Yang et al., 2000; Polanco-
Hernández et al., 2013). La presencia de este terpeno ha sido asociada a la inhibición
del crecimiento de T. brucei con AEs derivados de Aframomum sceptrum (Cheikh‐

Introducciön
31
Ali et al., 2011). Adicionalmente, el óxido de cariofileno también ha sido descrito
como agente inductor de estrés oxidativo e inhibidor de vías de supervivencia en
células tumorales (Park et al., 2011) y con potente actividad anti-inflamatoria de la
respuesta inmune mediada por células T (Ku y Lin, 2013)
1.2.4.3 Limoneno
El limoneno, es un terpeno perteneciente al grupo limonoide, cuya estructura posee
un carbono asimétrico como esterocentro. Como antimicrobianos, múltiples trabajos
han definido sus efectos antibacterianos (Soković et al., 2010), antivirales (Gómez et
al., 2013; Astani y Schnitzler, 2014), antimicóticos (Chee et al., 2009) y
antiparasitarios (Graebin et al., 2010; Santos et al., 2014). Su presencia además, ha
sido atribuida con muerte celular apoptótica por incremento del estrés oxidativo en
células susceptibles de cáncer de prostata (Rabi y Bishayee, 2009) y a través de vías
de muerte mitocondrial dependientes de caspasa en células leucémicas (Ji et al.,
2006).
1.2.4.4 Carvona
Carvona es un terpenoide que se encuentra naturalmente en muchos aceites
esenciales, con fuerte evidencia de actividad antimicrobiana (Aggarwal et al., 2002;
Escobar et al., 2010). Ha sido descrito como un potente antioxidante, incluso
superior al α-tocoferol (Stashenko et al., 2004) y con importante efecto
quimioprotector en células cancerígenas (Zheng et al., 1992).
Finalmente, teniendo en cuenta que la evolución de la infección del T. cruzi es
condicionada por la presencia del parásito, frente al cual el huésped mamífero monta
una respuesta inmune exacerbada (con daño celular y neuronal) y fibrosis
subsecuente de los tejidos (Nagajyothi et al., 2012; Cunha¬Neto et al., 2014), la
búsqueda y desarrollo de nuevas aproximaciones farmacológicas más eficientes y
selectivas, deben apuntar a la completa eliminación del parásito, con adecuada
modulación de la respuesta inmune del huésped. De acuerdo con lo anterior, aceites
esenciales de Lippia alba y sus terpenos bioactivos resultan promisorios para el
desarrollo de terapias más integrales contra la Enfermedad de Chagas,en la cual se
pueden abordar la eliminación selectiva del parásito, quimioprotección,
inmunomodulación y baja toxicidad, a través de posibles sinergias de sus
componentes (Zheng et al., 1992; Mesa-Arango et al. 2009; Escobar et al., 2010; Ku
y Lin, 2013; Stashenko et al., 2014; Liao et al., 2015; García et al., 2017).

Introducciön
32
2. OBJETIVOS
2.1 OBJETIVO GENERAL
Determinar la actividad tripanocida y citotóxica in vitro de aceites esenciales
obtenidos de Lippia alba (Quimiotipos Citral y Carvona) y de sus terpenos
(citral, limoneno, carvona y óxido de cariofileno), sobre formas
epimastigotes, tripomastigotes y amastigotes de Trypanosoma cruzi.
2.2 OBJETIVOS ESPECÍFICOS
Determinar la Concentración Inhibitoria media (CI50) y la Concentración
Citotóxica 50 (CC50) de los aceites esenciales de L. alba (Quimiotipos Citral
y Carvona) y de sus terpenos sobre los tres estadíos parasitarios de T. cruzi.
Establecer posibles interacciones farmacológicas entre los terpenos y/o
medicamento de referencia (Benznidazol) en la inhibición del crecimiento de
T. cruzi.
Rastrear mediante técnicas generales de biología celular, el mecanismo de
muerte celular inducido por los aceites esenciales de L. alba y sus terpenos
bioactivos sobre T. cruzi.
Objetivos

Materiales y Métodos
33
3. MATERIALES Y MÉTODOS
3.1 MATERIALES
3.1.1 Aceites esenciales derivados de L. alba
Para este trabajo, especímenes de los quimiotipos Citral y Carvona de Lippia
alba (Miller) N. E. Brown fueron plantados en el Centro Nacional de Investigaciones
para la Agroindustrialización de Especies Vegetales, Aromáticas y Medicinales
Tropicales (CENIVAM), ubicado en Bucaramanga, Colombia, a una altitud de 960 m
sobre el nivel del mar. En total fueron producidos 76 tipos diferentes de aceites, los
cuales presentaron variaciones en su quimiotipo de origen (53 Carvona y 23 Citral),
estación de plantío y colecta (lluvia y sequía), parte de la planta (tallo, hojas (verdes,
maduras y mixtas) e inflorescencias) y tiempo de extracción del aceite (30 – 90
minutos) (Tabla 4 y 5). Teniendo en cuenta, las precipitaciones medias (descritas por
el IDEAM), como época de sequía fueron seleccionados los meses de enero a marzo
y como estación seca fueron definidos los meses de abril a noviembre. Las
condiciones ambientales promedio de temperatura, humedad relativa y precipitación
para el período seco fueron: 26.3°C, 68.9% y 1.05 mm/día y para el período lluvioso,
24.5°C, 81.3% y 4.13 mm/día, respectivamente. Los AEs se extrajeron mediante
hidrodestilación asistida por microondas (HD-MW), y su caracterización química fue
llevada a cabo a través de cromatografía de gases acoplada a espectrometría de masas
(GC-MS) utilizando ionización por impacto de electrones en dos columnas de
diferente polaridad (Stashenko et al., 2004; García et al., 2017). Estos procedimentos
fueron realizados por personal adscrito al CENIVAM, bajo la dirección de la Dra.
Elena Stashenko.

Materiales y Métodos
34
Tabla 4. Aceites esenciales de L. alba quimiotipo Citral, producidos bajo diversos parámetros
ambientales y de extracción.
Est
aci
ón
del
Añ
o
Est
ad
o
Ma
teri
al
AEa
Tiempo
Extracción
(min)
Parte
planta
Composición química por GC-MS
Neral Geraniol Geranial Cariob OxCarc S
equ
ía
Fre
sco
A4 60 HJd 24.4 21.5 39.8 1.6 -
A5 90 HJ 26.1 17.2 41.4 2.3 -
A7 30 HMe 29.4 - 46.2 2.7 -
A8 30 HM 23.1 - 29.7 1.7 3.8
A27 45 Infl.f 19.6 32.3 30.6 1.9 -
A28 45 Infl. 19.3 31.5 31.3 2.3 -
Llu
via
Fre
sco
A1 30 HJ 25.5 - 36.1 4.1 1.0
A2 30 HJ 25.1 - 36.2 4.6 0.9
A9 60 HM 21.1 - 26.5 3 6.6
A25 45 HTg 22.8 5.3 27.5 4.6 2.8
A26 45 HT 36 6 45.1 3.2 1.4
Sec
o
A13 30 HJ 30.6 - 54.5 2.9 -
A14 30 HJ 31.9 - 57.3 2.6 -
A15 60 HJ 28.4 - 54 5.4 2.2
A16 60 HJ 30.8 - 52.6 3.8 3.3
A17 90 HJ 31.7 - 53.5 4.1 3
A18 90 HJ 29.3 - 54.5 3.5 3.1
A19 30 HM
33.6 - 53.8 2.7 1.9
A20 30 HM 32.1 - 54 4 2.4
A21 60 HM
24.5 - 35.1 0.8 3.9
A22 60 HM
28.8 - 39.7 1.3 4.5
A23 90 HM 28 - 37.8 6.8 2.9
A24 90 HM 24 - 34.3 2 5.7
aAE: Aceite Esencial; bCario: Cariofileno; cOxCar: Óxido de cariofileno; dHJ: Hojas jóvenes; eHM:
Hojas maduras; fInfl: Inflorescencias; gHT: Todas las hojas.
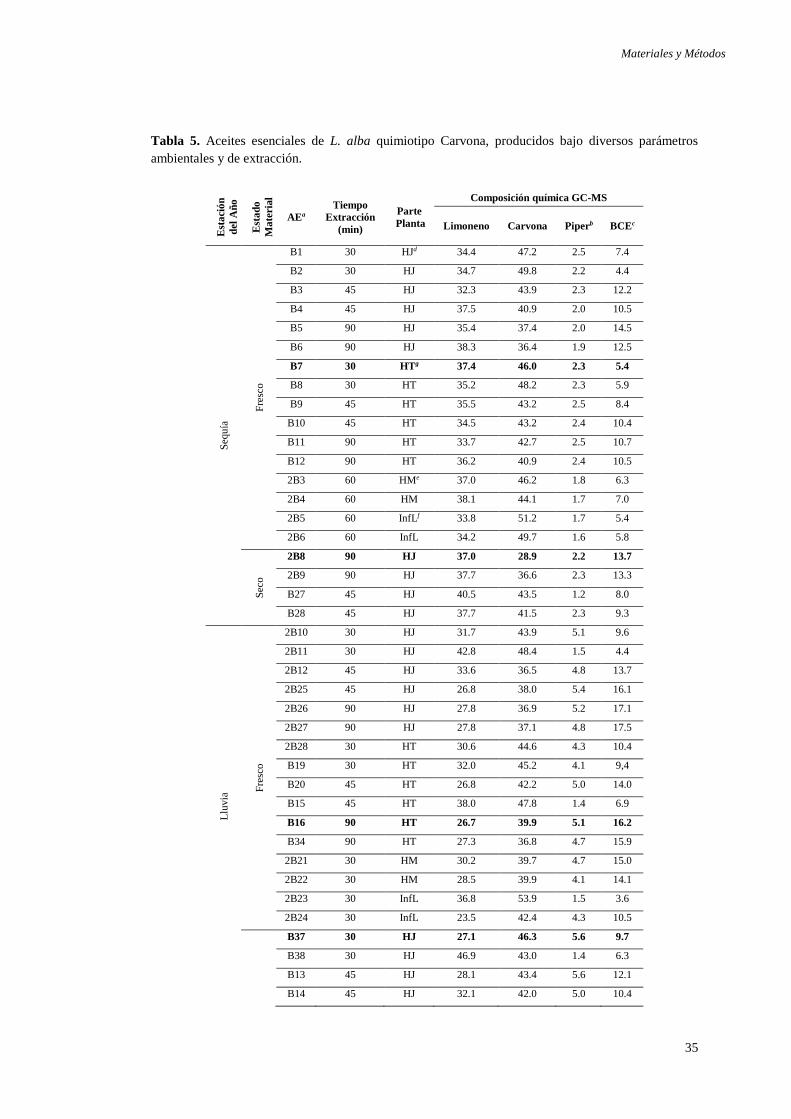
Materiales y Métodos
35
Tabla 5. Aceites esenciales de L. alba quimiotipo Carvona, producidos bajo diversos parámetros
ambientales y de extracción.
Est
aci
ón
del
Añ
o
Est
ad
o
Ma
teri
al
AEa
Tiempo
Extracción
(min)
Parte
Planta
Composición química GC-MS
Limoneno Carvona Piperb BCEc S
equ
ía
Fre
sco
B1 30 HJd 34.4 47.2 2.5 7.4
B2 30 HJ 34.7 49.8 2.2 4.4
B3 45 HJ 32.3 43.9 2.3 12.2
B4 45 HJ 37.5 40.9 2.0 10.5
B5 90 HJ 35.4 37.4 2.0 14.5
B6 90 HJ 38.3 36.4 1.9 12.5
B7 30 HTg 37.4 46.0 2.3 5.4
B8 30 HT 35.2 48.2 2.3 5.9
B9 45 HT 35.5 43.2 2.5 8.4
B10 45 HT 34.5 43.2 2.4 10.4
B11 90 HT 33.7 42.7 2.5 10.7
B12 90 HT 36.2 40.9 2.4 10.5
2B3 60 HMe 37.0 46.2 1.8 6.3
2B4 60 HM 38.1 44.1 1.7 7.0
2B5 60 InfLf 33.8 51.2 1.7 5.4
2B6 60 InfL 34.2 49.7 1.6 5.8
Sec
o
2B8 90 HJ 37.0 28.9 2.2 13.7
2B9 90 HJ 37.7 36.6 2.3 13.3
B27 45 HJ 40.5 43.5 1.2 8.0
B28 45 HJ 37.7 41.5 2.3 9.3
Llu
via
Fre
sco
2B10 30 HJ 31.7 43.9 5.1 9.6
2B11 30 HJ 42.8 48.4 1.5 4.4
2B12 45 HJ 33.6 36.5 4.8 13.7
2B25 45 HJ 26.8 38.0 5.4 16.1
2B26 90 HJ 27.8 36.9 5.2 17.1
2B27 90 HJ 27.8 37.1 4.8 17.5
2B28 30 HT 30.6 44.6 4.3 10.4
B19 30 HT 32.0 45.2 4.1 9,4
B20 45 HT 26.8 42.2 5.0 14.0
B15 45 HT 38.0 47.8 1.4 6.9
B16 90 HT 26.7 39.9 5.1 16.2
B34 90 HT 27.3 36.8 4.7 15.9
2B21 30 HM 30.2 39.7 4.7 15.0
2B22 30 HM 28.5 39.9 4.1 14.1
2B23 30 InfL 36.8 53.9 1.5 3.6
2B24 30 InfL 23.5 42.4 4.3 10.5
B37 30 HJ 27.1 46.3 5.6 9.7
B38 30 HJ 46.9 43.0 1.4 6.3
B13 45 HJ 28.1 43.4 5.6 12.1
B14 45 HJ 32.1 42.0 5.0 10.4

Materiales y Métodos
36
Tabla 5. Continuación
Est
aci
ón
del
Añ
o
Est
ad
o
Ma
teri
al
AEa
Tiempo
Extracción
(min)
Parte
Planta
Composición química GC-MS
Limoneno Carvona Piperb BCEc
aAE: Aceite Esencial; bPiper: Piperitenona; cBCE: Biciclosesquifelandreno; dHJ: Hojas jóvenes; eHM:
Hojas maduras; fInfl: Inflorescencias; gHT: Todas las hojas.
Como puede ser visto en las Tablas 4 y 5, aceites producidos bajo diversas
condiciones presentaron diferencias significativas en los porcentajes de sus terpenos
mayoritarios. De acuerdo con esto y con la finalidad de determinar el efecto de estas
variaciones en la actividad tripanocida, los 76 aceites fueron subclasificados en 12
grupos. Para este estudio, fue seleccionado un representante de cada uno de estos
subgrupos, siendo 6 óleos del quimiotipo Citral (A13, A20, A23, A24, A25 Y A28) y
6 del quimiotipo Carvona (B7, B16, B37, 2B8, 2B18 y 2B19). Todos estos aceites
fueron extraídos de hojas de L. alba. AEs extraidos de las flores fueron descartados
debido a su alta citotoxicidad sobre células normales del huésped (Vero) (datos no
mostrados).
3.1.2 Terpenos y medicamentos
Los terpenos puros S (+) carvona, D (+) limoneno, (-) (-) óxido de cariofileno y citral
(terpernos derivados de Lippia alba) fueron obtenidos comercialmente de Sigma
Aldrich. El Benznidazol utilizado como medicamento de referencia, fue adquirido de
la Secretaría de Salud del departamento de Santander, Colombia, y gentilmente
purificado por la Doctora Leonor Yamile Vargas, del programa Química ambiental
de la Universidad Santo Tomás de Aquino (Bucaramanga).
Las soluciones stock de cada uno de los aceites y terpenos fueron preparadas en
dimetilsulfóxido (DMSO, Sigma) y guardadas a 4°C, protegidas de la luz. Las
2B13 90 HJ 29.6 37.9 4.9 16.2
2B14 90 HJ 26.6 35.5 5.0 18.3
Seco 2B15 30 HT 27,6 43.0 4.8 12.2
2B16 30 HT 29.4 43.2 4.2 12.9
2B17 45 HT 26.5 41.4 5.1 13.9
2B18 45 HT 23.4 42.1 5.2 16.6
2B19 90 HT 36.4 53.3 1.6 3.2
2B20 90 HT 34.0 44.4 4.7 7.5
B25 30 HJ 38.6 41.3 2.2 9.0
B26 30 HJ 0 0 0 0
2B1 30 HT 26.4 40.2 5.6 9
2B2 30 HT 41.9 47.4 1.6 3.0
2B7 90 HT 36.1 43.1 2.3 9.5

Introducciön
37
soluciones de trabajo fueron diluidas en medio de cultivo a las concentraciones
deseadas, justo antes de cada ensayo (concentración final de DMSO <0.1%).
3.1.3 Cultivos celulares
Células epiteliales derivadas de Riñón de Mono Verde Africano (VERO, ATCC CCL-
81), fueron cultivadas en medio DMEM (Tabla 6) (Life Technology, CA, USA)
suplementado con Suero Fetal Bovino Inactivo al 10% (SFBi) (Life Technology, CA,
USA), 1000 U/mL de penicilina y 100 μg/mL de estreptomicina e incubadas a 37°C,
90% de humedad y 5% de CO2.
Tabla 6. Composición de los medios de cultivo utilizados para el crecimiento in vitro de Células Vero
y T. cruzi
Medio LIT Medio DMEM
Sales Inorgánicas KCl
NaCl
Na2HPO4-H2O
CaCl2 (Anhid.)
Fe(NO3)3-9H2O
MgSO4 (Anhid.)
KCl
NaHCO3
NaCl
NaH2PO4-H2O
Azúcares y Ácidos Orgánicos D-Glucosa
Triptosa
D-Glucosa
Piruvato de Sodio
Aminoácidos
Glicina
L-Arginine
L-Cistina 2HCl
L-Glutamina
L-Histidina -H2O
L-Isoleucina
L-Leucina
L-Lisina
L-Metionina
L-Fenilalanina
L-Serina
L-Treonina
L-Triptófano
L-Tirosina
L-Valina
Vitaminas
Pantotenato D-Calcio
Ácido Fólico
Niacinamida
Piridoxina
Riboflavina
Tiamina
I-Inositol
Aditivos Extracto de hígado
Hemina
Suero Fetal Bovino Inactivado (SFBi) 10%
Penicilina 1000 U/mL
Estreptomicina 100 µg/mL

Materiales y Métodos
38
Epimastigotes de T. cruzi cepa SYLVIO-X10, representativa del TcI (donados por el Dr.
Marcos López-Casillas, del Grupo de Investigación Biomédica Traslacional de la
Fundación Cardiovascular de Colombia) fueron mantenidos en medio Liver Infution
Triptosa (LIT) suplementado con 10% de SFBi a 28°C. Los Tripomastigotes derivados
de células fueron obtenidos mediante la infección de la monocapa de células Vero con
epimastigotes en fase estacionaria (12 días) y crecidos en las mismas condiciones
descritas anteriormente para el mantenimiento de las células de mamífero (Tabla 6).
3.2 MÉTODOS
3.2.1 Citotoxicidad en Células Vero
Células Vero (3x105 cel/mL) fueron incubadas en placas fondo plano de 96 pozos a
37°C, 5% CO2 y 95% de humedad por 24 horas hasta la formación de la monocapa.
Posteriormente, las células fueron expuestas durante 70 horas a los diferentes
compuestos, evaluando 4 concentraciones por triplicado (300, 100, 33.3 y 11.1
µg/mL). La viabilidad celular se determinó empleando el reactivo de proliferación
celular WST-1 (Roche, Mannheim, Germany). Para ello, las células tratadas fueron
expuestas durante 2 horas más con la sonda WST-1 y posteriormente la densidad
óptica fue determinada por espectrofotometría a una longitud de onda de 450 nm. El
porcentaje de citotoxicidad fue calculado mediante la siguiente ecuación: ((DO grupo
control – DO grupo tratado) / DO grupo control) x 100.
3.2.2 Actividad antiparasitaria en Epimastigotes de T. cruzi
Epimastigotes de T.cruzi en fase de crecimiento exponencial fueron cultivados en
placas de 96 pozos a una concentración de 5x105 epimastigotes/mL en medio LIT
con 10 % SFBi. Los parásitos fueron expuestos durante 72 horas a 28ºC con los AEs
y compuestos; concentraciones de 100, 33.3, 11.1, 3.7 µg/mL fueron probadas por
triplicado. Parásitos sin tratamiento fueron evaluados como controles negativos y el
benznidazol fue utilizado como control positivo. La inhibición del crecimiento de los
epimastigotes fue determinada en el microscopio de luz, mediante conteo directo de
parásitos vivos, utilizando el colorante vital azul de trypan (Gibco). Los resultados
fueron expresados como la concentración que inhibe el 50% de los parásitos (CI50).
3.2.3 Actividad antiparasitaria en Tripomastigotes derivados
de células (TDC)
Cultivo en monocapa de células Vero fueron infectados con epimastigotes de T. cruzi
en fase estacionaria e incubadas a 37ºC, 5% CO2 y 95% de humedad hasta observar

Materiales y Métodos
39
liberación de tripomastigotes derivados de células al medio de cultivo.
Posteriormente, estos tripomastigotes derivados de células (5x105 cel/mL) fueron
expuestos durante 24 horas a 37ºC, 5% CO2 y 95% de humedad con los AEs y
componentes mayoritarios. Cuatro concentraciones (100, 33.3, 11.1, 3.7 µg/mL)
fueron evaluadas por triplicado. Se evaluaron controles negativos sin tratamiento y el
benznidazol fue utilizado como control positivo.
La inhibición del crecimiento de tripomastigotes después del tratamiento, fue
determinada en el microscopio de luz, mediante conteo directo de parásitos vivos y el
resultado fue expresado como la concentración que inhibe el 50% de los parásitos
(CI50).
3.2.4 Actividad antiparasitaria en amastigotes intracelulares
de T. cruzi
Células Vero (3x105 cel/mL) fueron sembradas en placas de 16 pozos durante 24
horas e incubadas a 37ºC, 5% CO2 y 95% de humedad. Posteriormente, la monocapa
fue infectada con tripomastigotes derivados de células en un radio 1:3 (célula:
parásito) durante 24 horas para la obtención de amastigotes. Las células infectadas
fueron expuestas durante 120 horas a los compuestos estudiados (en dos dosis, una
en el tiempo 0 y la segunda a las 48 horas) bajo las mismas condiciones de
incubación. Se evaluaron controles sin tratamiento y benznidazol como medicamento
de referencia. Las placas fueron fijadas con metanol y coloreadas con Giemsa para la
evaluación de la inhibición del crecimiento. Se determinó el porcentaje de células
infectadas y no infectadas en un total de 300 células por microscopía de luz.
3.2.5 Evaluación del efecto combinado entre el Limoneno y el
resto de terpenos sobre T. cruzi
Una matriz de interacción farmacológica entre el limoneno (Fracción terpenoide de
mayor selectividad) y el resto de terpenos, fue realizada sobre las tres formas
parasitarias de T. cruzi usando el método de isobolograma de radio fijo, descrito por
Fivelman et al. (2004) con algunas modificaciones.
Para la matriz de interacción fueron usados como base los valores de
CI50 determinados para los terpenos y el Limoneno de manera individual, y utilizadas
dentro de las combinaciones de la forma como se describe en la Tabla 7. Una mezcla
adicional con Benznidazol también fue ejecutada.

Materiales y Métodos
40
Tabla 7. Matriz de interacción entre el Limoneno y el resto de terpenos usando valores de CI50
Combinación Compuesto X Compuesto Y
1 0.0 8x CI50
2 ½ CI50 4x CI50
3 CI50 2x CI50
4 2x CI50 CI50
5 4x CI50 ½ CI50
6 8x CI50 0.0
x: número de veces.
La evaluación de la susceptibilidad se realizó siguiendo la metodología descrita
anteriormente para determinar la actividad antiparasitaria en cada una de las formas
tripanocíticas. La concentración inhibitoria fraccional (CIF) fue calculada mediante la
fórmula: CIF del compuesto X= CI50 del compuesto X en combinación /CI50 del
compuesto X solo, y la sumatoria de la CIF (CIF) se determinó de la siguiente manera:
CIF= CIF del compuesto X + CIF del compuesto Y. De esta forma, se realizaron
isobologramas para determinar la naturaleza de las interacciones entre los compuestos
y/o medicamento de referencia: Interacción sinérgica: CIF <1; interacción antagónica:
CIF >1 e indiferente o aditiva con CIF =1 (Chou y Talalay, 1984; Azeredo y
Soares, 2013).
3.2.6 Cambios Morfológicos
Los cambios morfológicos en formas epimastigotas tratadas con los aceites, terpenos
bioactivos y benznidazol (forma redondeada, disminución del tamaño celular,
condensación de la cromatina nuclear, vacuolización del citoplasma, fragmentación del
núcleo y formación de cuerpos apoptóticos) fueron evaluados por microscopía de luz
mediante microscopia de contraste de fases interferencial.
Para la evaluación de la integridad del DNA, epimastigotes de T. cruzi previamente
tratados (1x106 parásitos/mL) fueron fijados con paraformaldehido al 4% durante 10
minutos y coloreados con 4 ',6diamino-2, fenilindol (DAPI, 1 µg/mL, Sigma Aldrich).
Células no tratadas se utilizaron como control negativo. La morfología de los núcleos
celulares fue observada usando un microscopio de fluorescencia (Nikon Eclipse Ni) a
una longitud de onda de excitación de 358 nm y emisión de 461 nm.
3.2.7 Determinación del potencial de membrana mitocondrial
Para la determinación del efecto sobre el potencial de membrana, los parásitos vivos
fueron coloreados con MitoTracker Red CMXRos (λEm 579nm y λEx599nm), un
derivado de Xrosamina. Para esto, epimastigotes en fase log de crecimiento (1x106
parásitos/mL) fueron incubados con 100 nM Mitotracker Red CMXRos a 28°C por 5
min en DMEM, lavadas con medio fresco e incubadas finalmente sin colorante por 20
minutos (García et al., 2007).

Materiales y Métodos
41
3.2.8 Determinación de la fragmentación del ADN
Un ensayo de TUNEL (Molecular Probes, Invitrogen) fue realizado con la finalidad
de determinar la fragmentación del ADN usando desoxinucleotidil transferasa
terminal marcada con fluoresceína dUTP. Para ello, epimastigotes tratados con la
CI50 X2 de cada compuesto durante 48 horas (1x106 parásitos/mL), fueron lavados
dos veces con PBS, fijados con 4% de paraformaraldehído e incubados con 0,25% de
triton por 20 minutos a temperatura ambiente. Los parásitos se incubaron durante 60
minutos a 37°C con el cóctel de reacción TUNEL (Buffer TDT, nucleótidos con
EdUTP, Desoxinucleotidil transferasa terminal (TdT)) y se adiciono suero albumina
bovino al 3%. Una incubación posterior fue ejecutada con el coctel de reacción
Click-IT (Alexa Fluor® 488) durante 30 minutos, y finalmente se adiciono Hoechst
por 15 minutos protegidos de la luz. Los parásitos fueron analizados mediante
microscopía de fluorescencia (Nikon Eclipse Ni) a una longitud de onda de
excitación de 358 nm y de emisión a 461 nm. Se empleó como control positivo un
cultivo viejo de epimastigotes, de 15 días de incubación. La cuantificación de la
técnica, se realizó a través del conteo de parásitos con fragmentación del ADN
(Emisión fluorescencia verde) en 300 células observadas. Los resultados se
expresaron como porcentaje de epimastigotes TUNEL positivo.
La fragmentación de ADN también fue evaluada por la presencia de un patrón
oligonucleosomal, evidenciado en una electroforesis de agarosa. Brevemente, 3 uL
de ADN parasitario extraído mediante QIAamp® DNA Mini Kit (Qiagen) fue
mezclado con 8 uL de Buffer de carga (Sigma Aldrich) y servido en un gel de
agarosa Low Melting 1,8% a 50 voltios durante 3 horas.
3.2.9 Determinación de la externalización de la
fosfatidilserina
La caracterización de la muerte celular fue determinada mediante el kit de apoptosis
PE Annexin V/Dead with SYTOX® Green (Molecular Probes, Invitrogen) de
acuerdo con las indicaciones del fabricante. La externalización de la fosfatidilserina
se determinó empleando Anexina V recombinante conjugada con la ficobiliproteína
naranja fluorescente R-PE y el fenotipo de las células necróticas utilizando el
colorante de ácidos nucleicos SYTOX. Brevemente, epimastigotes tratados con la
CI50 X2 durante 48 horas (1x106 parásitos/mL) fueron lavados y resuspendidos en el
Buffer Anexina-Binding 1X. Se adicionaron 5 μL de R-PE annexin V y 1 μL de 1
μM SYTOX® Green Stain e incubaron durante 15 minutos a 37ºC, 5% CO2 y 95%
de humedad. Como control positivo de apoptosis se utilizó un cultivo viejo de
epimastigotes, de 15 días de incubación. Parásitos sin ningún tratamiento

Materiales y Métodos
42
representaron el control negativo. Se realizó la lectura en el citómetro de flujo
(FACSCanto II con filtros para RPE (Ex: 488nm / Em: 575 nm), SYTOX (Ex: 503
nm /Em: 524 nm)), facilitado por la Fundación Cardiovascular de Colombia.
3.3 ANÁLISIS DE DATOS
Se utilizó el Software estadístico MsxlfitTM (ID Business Solution, Guildford, UK)
para calcular mediante análisis de regresión sigmoidal, la concentración inhibitoria o
citotóxica 50 (CI50 y CC50, respectivamente) de cada compuesto. Los experimentos se
realizaron por triplicado en tres momentos independientes. El análisis estadístico fue
realizado mediante el software SPSS 15.0, utilizando un ANOVA de Welch para
evaluar la existencia de diferencias entre los valores medios de CI50 en cada estadio
parasitario, así como para analizar los datos de citotoxicidad. Se utilizó la prueba de
Tukey para hacer comparaciones múltiples, estableciendo un nivel de confianza del
95% en todos los casos.

Resultados
43
4. RESULTADOS
4.1 COMPOSICIÓN QUÍMICA Y ACTIVIDAD
TRIPANOCIDA DE AEs DE L. alba
En esta investigación 12 aceites esenciales (AEs) extraídos de los quimiotipos citral y
carvona de Lippia alba, obtenidos bajo condiciones estandarizadas y controladas de
siembra, colecta y extracción, fueron caracterizados y testados en su potencial
tripanocida contra Trypanosoma cruzi.
En términos generales, todos los AEs estudiados ejercieron efectos inhibitorios
distintos en los tres estadios parasitarios evaluados (Epimastigotes: F=1320.080;
p=0.000; Tripomastigotes: F=628.786; p=0.000; Amastigotes: F= 853.422; p=0.000).
En las tablas 8 y 9 se presenta la composición química relativa de los AEs
(quimiotipos Citral y Carvona) y su actividad biológica sobre T. cruzi y Células
Vero.
Tabla 8. Composición química relativa y efecto antiproliferativo de los AEs extraídos del quimiotipo
Citral de L. alba sobre T. cruzi.
AEa
Composición química por GC-MS Epi d
CI50 g ±
DE j
(µg/mL)
Tripe
CI50 ±
DE
(µg/mL)
Amas f
CI50 ±
DE
(µg/mL)
Vero
CC50h±
DE
(µg/mL) Neral
(%)
Geraniol
(%)
Geranial
(%)
Cariob
(%)
OxCarc
(%)
ISi IS IS
A13 30,6 - 54,5 2,9 - 17,3±1,7 7,0 21,4±1,6 5,7 87,8±5,4 1,4 121,3±10,1
A20 32,1 - 54 4 2,4 8,5±1,2 7,8 13,9±0,9 4,7 >33,3 NDk 65,9±5,9
A23 28 - 37,8 6,8 2,9 8,1±1,3 6,2 17,4±1,3 3,0 >33,3 ND 50,5±6,2
A24 24 - 34,3 2 5,7 15,8±1,6 5,7 28,7±1,8 3,1 69,0±3,0 1,3 90,4±7,1
A25 22,8 5,3 27,5 4,6 2,8 17,9±0,7 5,3 19,4±0,9 4,9 >33,3 ND 94,8±9,2
A28 19,3 31,5 31,3 2,3 - 13,7±2,6 7,1 30,7±1,9 3,1 65,5±4,8 1,5 96,6±11,0
BNZ - - - - - 16,9±0,9 8,2 1,2±0,1 116,3 6,2 ± 0,9 22,4 138,7±2,3
aAE: Aceite esencial; bCario: Cariofileno; cOxCar: Óxido de Cariofileno; dEpi: Epimastigote; eTrip:
Tripomastigote; fAmas: Amastigote; gCI50: Concentración Inhibitoria 50; hCC50: Concentración
Citotóxica 50; iIS: Índice de selectividad definido como CC50/IC50; jDE: Desviación Estándar; kND:
No determinado.

Resultados
44
Tabla 9. Composición química relativa y efecto antiparasitario de los AEs extraídos del quimiotipo
Carvona de L. alba sobre T. cruzi
AEa
Composición química por GC-MS Epi d
CI50 g ±
DEj
(µg/mL)
Tripe
CI50 ±
DE
(µg/mL)
Amas f
CI50 ±
DE
(µg/mL)
Vero
CC50h± DE
(µg/mL) Limoneno
(%)
Carvona
(%) Piperb
(%) BCEc
(%)
ISi IS IS
B7 37,4 46,0 2,3 5,4 80,9±2,4 2,5 36,8±2,1 5,5 > 150 NDk 203,0±7,5
B16 26,7 39,9 5,1 16,2 97,4±3,2 2,2 57,4±3,0 3,8 > 150 ND 215,6±9,6
B37 27,1 46,3 5,6 9,7 91,9±3,9 2,1 43,3±1,8 4,5 > 150 ND 195,8±18,2
2B8 37,0 28,9 2,2 13,7 96,1±4,4 1,9 47,2±3,8 4,0 > 150 ND 186,3±11,7
2B18 23,4 42,1 5,2 16,6 85,6±4,8 1,9 33,7±3,1 4,9 > 150 ND 165,1±10,2
2B19 36,4 53,3 1,6 3,2 77,5±3,5 2,9 51,0±1,4 4,4 > 150 ND 226,1±8,5
BNZ - - - - 16,9±0,9 8,2 1,2±0,1 116,3 6,2±0,9 22,4 138,7±2,3
aAE: Aceites esencial; bPiper: Piperitenona; cBCE: Biciclosesquifelandreno; dEpi: Epimastigote; eTrip:
Tripomastigote; fAmas: Amastigote; gCI50: Concentración Inhibitoria 50; hCC50: Concentración
Citotoxica 50; iIS: Indice de selectividad definido como CC50/IC50; jDE: Desviación Estándar; kND:
No determinado.
Como se observa en las Tablas 8 y 9, y en la Figura 16, el desempeño tripanocida de
los AEs extraídos de plantas del quimiotipo Citral exhibieron mayor actividad
(Media Epimastigotes: CI50 13,6±1,5 µg/mL; Tripomastigotes: CI50 21,9±1,4 µg/mL;
Amastigotes: CI50 74,1±4,4 µg/mL), que las obtenidas con el quimiotipo Carvona
(Media Epimastigotes: CI50 88,2±3,7 µg/mL; Tripomastigotes: CI50 44,9±2,5 µg/mL;
Amastigotes: CI50 >150 µg/mL). En este sentido, los mejores valores de CI50 sobre
las formas parasitarias extracelulares fueron alcanzados por los aceites A20
(Epimastigotes: CI50 8,5 ± 1,2 µg/mL; Tripomastigotes: CI50 13,9 ± 0,9 µg/mL) y
A23 (Epimastigotes: CI50 8,1 ± 1,3 µg/mL; Tripomastigotes: CI50 17,4 ± 1,3 µg/mL),
del quimiotipo citral. Al comparar estos dos aceites, no se observaron diferencias
significativas en su actividad tripanocida contra Epimastigotes (p= 1.000), así como
contra Tripomastigotes (p= 0,161). Para estos AEs, no fue posible determinar las
concentraciones inhibitorias 50 en amastigotes, debido a la toxicidad ejercida sobre
las células Vero (A20: CC50= 65,9 ± 5,9 µg/mL; A23: CC50= 50,5 ± 6,2 µg/mL); estos
aceites no mostraron inhibición del parásito en las concentraciones evaluadas (33,3,
11,1 y 3,7 µg/mL). Por otro lado, el aceite A13, presentó actividad tripanocida
importante en las tres formas parasitarias (Epimastigotes: CI50 17,3 ± 1,7 µg/mL,
IS=7,0; Tripomastigotes: CI50 21,4 ± 1,6 µg/mL, IS=5,7; Amastigotes: CI50 87,8 ±
5,4 µg/mL, IS=1,4) y demostró ser el aceite del quimiotipo Citral menos tóxico para
las células mamíferas (CC50 121,3 ± 10,1 µg/mL) (Tabla 8, Figura 16).
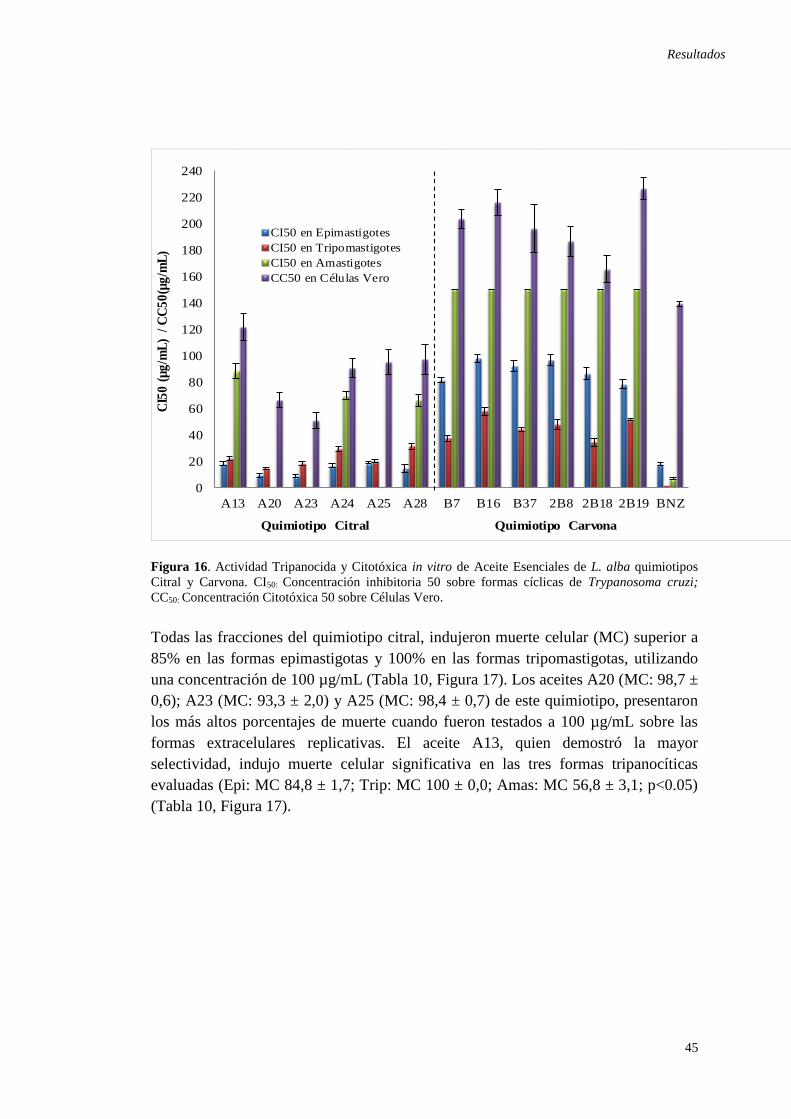
Resultados
45
0
20
40
60
80
100
120
140
160
180
200
220
240
A13 A20 A23 A24 A25 A28 B7 B16 B37 2B8 2B18 2B19 BNZ
CI5
0 (
µg
/mL
) /
CC
50
(µg
/mL
)
Quimiotipo Citral Quimiotipo Carvona
CI50 en Epimastigotes
CI50 en Tripomastigotes
CI50 en Amastigotes
CC50 en Células Vero
Figura 16. Actividad Tripanocida y Citotóxica in vitro de Aceite Esenciales de L. alba quimiotipos
Citral y Carvona. CI50: Concentración inhibitoria 50 sobre formas cíclicas de Trypanosoma cruzi;
CC50: Concentración Citotóxica 50 sobre Células Vero.
Todas las fracciones del quimiotipo citral, indujeron muerte celular (MC) superior a
85% en las formas epimastigotas y 100% en las formas tripomastigotas, utilizando
una concentración de 100 µg/mL (Tabla 10, Figura 17). Los aceites A20 (MC: 98,7 ±
0,6); A23 (MC: 93,3 ± 2,0) y A25 (MC: 98,4 ± 0,7) de este quimiotipo, presentaron
los más altos porcentajes de muerte cuando fueron testados a 100 µg/mL sobre las
formas extracelulares replicativas. El aceite A13, quien demostró la mayor
selectividad, indujo muerte celular significativa en las tres formas tripanocíticas
evaluadas (Epi: MC 84,8 ± 1,7; Trip: MC 100 ± 0,0; Amas: MC 56,8 ± 3,1; p<0.05)
(Tabla 10, Figura 17).

Resultados
46
Tabla 10. Muerte celular de AEs de L. alba, quimiotipo Citral y Carvona a 100 µg/mL sobre las
diferentes formas de T. cruzi.
aAE: Aceite esencial; bEpi: Epimastigote; cTrip: Tripomastigote; dAmas: Amastigote; eDE: Desviación
Estándar; fND: No determinado.
Figura 17. Porcentajes de muerte celular sobre formas cíclicas de Trypanosoma cruzi tratadas con
100 µg/mL de los 12 aceites esenciales de L. alba quimiotipos Citral y Carvona.
Por su parte, los AEs del quimiotipo Carvona, presentaron valores de CI50 entre 77,5
µg/mL y 97,4 µg/mL sobre las formas epimastigotes y entre 33,7 µg/mL y 57,4
µg/mL frente a tripomastigotes, con un valor medio de porcentaje de muerte de
56,04% ± 2,25 y 81,06% ± 3,08 respectivamente, a concentraciones de 100 µg/mL
0
10
20
30
40
50
60
70
80
90
100
A13 A20 A23 A24 A25 A28 B7 B16 B37 2B8 2B18 2B19
% M
uer
te C
elu
lar
Quimiotipo Citral Quimiotipo Carvona
Epimastigotes
Tripomastigotes
Amastigotes
Muerte celular (%) ± DEe - 100 µg/mL
AE a Epi b Trip c Amas d
Qu
imio
tip
o
Cit
ral
A13 84,8 ± 1,7 100 ± 0,0 56,8 ± 3,1
A20 98,7 ± 0,6 100 ± 0,0 NDf
A23 93,3 ± 2,0 100 ± 0,0 ND
A24 87,6 ± 2,5 100 ± 0,0 74,6± 3,7
A25 98,4 ± 0,7 100 ± 0,0 ND
A28 85,3 ± 3,5 100 ± 0,0 70,4± 2,4
Qu
imio
tip
o
Ca
rvo
na
B7 60,1 ± 1,6 84,5± 1,7 17,0 ± 3,3
B16 50,9 ± 2,2 83,6 ± 6,9 39,0 ± 4,0
B37 53,5 ± 2,4 76,1 ± 1,5 20,0 ± 5,5
2B8 51,3 ± 2,1 78,5 ± 3,1 6,6 ± 4,0
2B18 57,5 ± 2,6 86,8 ± 3,4 19,1 ± 6,2
2B19 62,9 ± 2,6 77,0 ± 2,0 20,2 ± 3,1

Resultados
47
(Tabla 10, Figura 16 y 17). Dentro de los aceites de este quimiotipo, los de mejor
desempeño fueron B7 (Epimastigotes: MC 60,10%, CI50 80,9 ± 2,4 µg/mL, IS=2,5;
Tripomastigotes: MC 84,46%, CI50 36,8 ± 2,1 µg/mL, IS=5,5) y 2B18
(Epimastigotes: MC 57,53%, CI50 85,6 ± 4,8 µg/mL, IS=1,2; Tripomastigotes: MC
86,76%, CI50 33,7 ± 3,1 µg/mL, IS=4,9) (Tabla 9 y 10, Figura 16 y 17). Vale la pena
mencionar que, ninguno de estos aceites presentó actividad antiparasitaria
significativa sobre las formas intracelulares replicativas del parásito (Media CI50
>150 µg/mL; Media MC 20,35% ± 4,37). Así mismo los aceites de este quimiotipo
mostraron ser menos citotóxicos para las células hospederas (Vero), que sus
homólogos del quimiotipo Citral, con un valor medio de CC50 de 198,6 ± 10,9 µg/mL
y 86,6 ± 8,3 µg/mL, respectivamente (Tabla 8 y 9, Figura 16).
4.2 ACTIVIDAD TRIPANOCIDA DE TERPENOS
DE L. alba
Teniendo en cuenta la composición química de los aceites estudiados (Tablas 8 y 9),
y lo descrito en la literatura (García et al., 2017), fueron seleccionados cuatro
terpenos que han sido relacionados con la actividad biológica, para su estudio como
agentes tripanocidas individuales. De esta manera, para el quimiotipo Citral fueron
escogidos los terpenos citral y (-) óxido de cariofileno; y para el quimiotipo Carvona,
los monoterpenos D (+) Limoneno y S (+) Carvona. Para estos ensayos, sobre las
formas extracelulares e intracelulares de T. cruzi, fueron utilizadas las versiones
comerciales (de Sigma Aldrich), de estos compuestos. En la tabla 11 se presentan las
concentraciones inhibitorias medias (CI50) obtenidas en los tres estadios parasitarios
y los valores de CC50 determinados sobre las células Vero.
Tabla 11. Efecto antiparasitario de los principales terpenos derivados de L. alba en Trypanosoma
cruzi
Terpenos
Epia
CI50d± DE
(µg/mL)
Tripb
CI50 ± DE
(µg/mL)
Amasc
CI50 ± DE
(µg/mL)
Vero
CC50e±DE
(µg/mL) IS f IS IS
OxCarg 29,8 ± 1,7 4,3 21,6 ± 0,3 5,9 47,4 ± 1,0 2,7 128,0 ± 4,2
Limoneno 41,8 ± 2,5 7,1 9,0 ± 0,8 32,8 28,7 ± 0,7 10,3 297,1 ± 2,4
Citral 37,2 ± 0,7 2,4 20,8 ± 1,2 4,3 49,0 ± 2,3 1,8 90,2 ± 3,9
Carvona 176,9 ± 7,9 1,4 123,9 ± 7,8 1,9 > 100 NDi 240,1 ± 4,1
BNZh 16,9 ± 0,9 8,2 1,2 ± 0,1 116,3 6,2 ± 0,9 22,4 138,7 ± 2,3
aEpi: Epimastigote; bTrip: Tripomastigote; cAmas: Amastigote; dIC50: Concentración Inhibitoria 50; eCC50: Concentración citotóxica 50; fIS: Índice de selectividad definido como Vero CC50/IC50; gOxCar: Oxido de Cariofileno; hBNZ: Benznidazol; iND: No determinado.
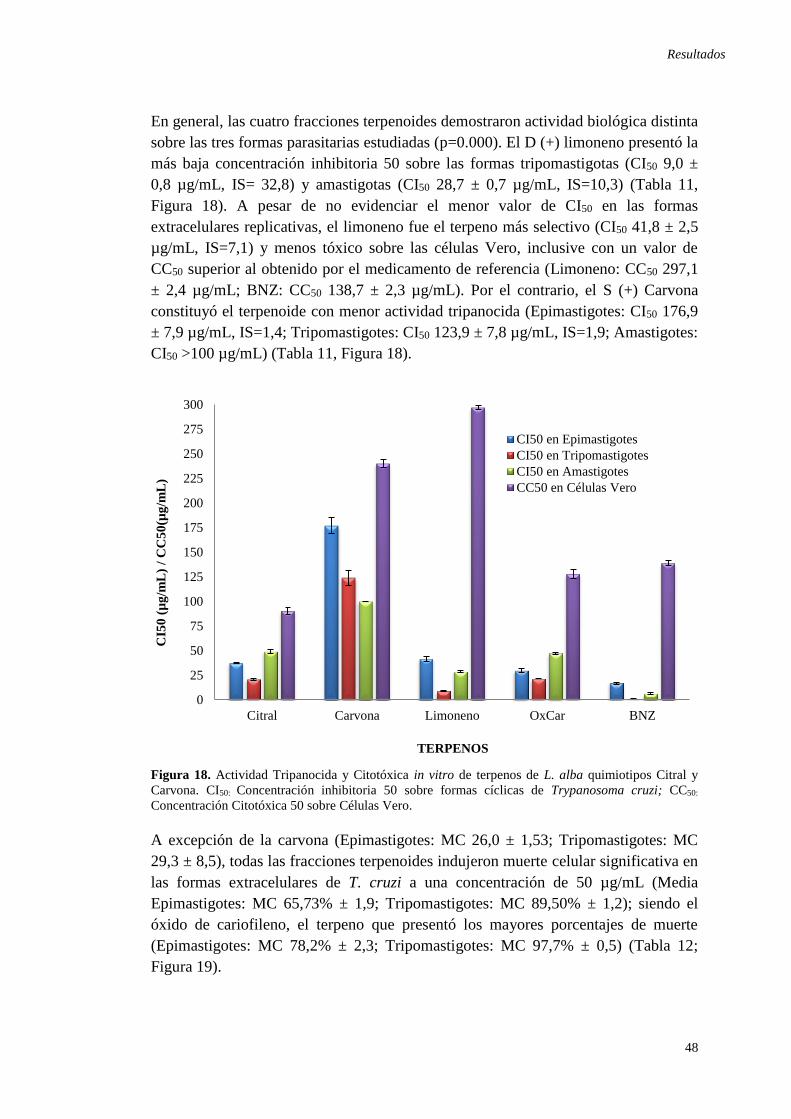
Resultados
48
En general, las cuatro fracciones terpenoides demostraron actividad biológica distinta
sobre las tres formas parasitarias estudiadas (p=0.000). El D (+) limoneno presentó la
más baja concentración inhibitoria 50 sobre las formas tripomastigotas (CI50 9,0 ±
0,8 µg/mL, IS= 32,8) y amastigotas (CI50 28,7 ± 0,7 µg/mL, IS=10,3) (Tabla 11,
Figura 18). A pesar de no evidenciar el menor valor de CI50 en las formas
extracelulares replicativas, el limoneno fue el terpeno más selectivo (CI50 41,8 ± 2,5
µg/mL, IS=7,1) y menos tóxico sobre las células Vero, inclusive con un valor de
CC50 superior al obtenido por el medicamento de referencia (Limoneno: CC50 297,1
± 2,4 µg/mL; BNZ: CC50 138,7 ± 2,3 µg/mL). Por el contrario, el S (+) Carvona
constituyó el terpenoide con menor actividad tripanocida (Epimastigotes: CI50 176,9
± 7,9 µg/mL, IS=1,4; Tripomastigotes: CI50 123,9 ± 7,8 µg/mL, IS=1,9; Amastigotes:
CI50 >100 µg/mL) (Tabla 11, Figura 18).
Figura 18. Actividad Tripanocida y Citotóxica in vitro de terpenos de L. alba quimiotipos Citral y
Carvona. CI50: Concentración inhibitoria 50 sobre formas cíclicas de Trypanosoma cruzi; CC50:
Concentración Citotóxica 50 sobre Células Vero.
A excepción de la carvona (Epimastigotes: MC 26,0 ± 1,53; Tripomastigotes: MC
29,3 ± 8,5), todas las fracciones terpenoides indujeron muerte celular significativa en
las formas extracelulares de T. cruzi a una concentración de 50 µg/mL (Media
Epimastigotes: MC 65,73% ± 1,9; Tripomastigotes: MC 89,50% ± 1,2); siendo el
óxido de cariofileno, el terpeno que presentó los mayores porcentajes de muerte
(Epimastigotes: MC 78,2% ± 2,3; Tripomastigotes: MC 97,7% ± 0,5) (Tabla 12;
Figura 19).
0
25
50
75
100
125
150
175
200
225
250
275
300
Citral Carvona Limoneno OxCar BNZ
CI5
0 (
µg
/mL
) /
CC
50
(µg
/mL
)
TERPENOS
CI50 en Epimastigotes
CI50 en Tripomastigotes
CI50 en Amastigotes
CC50 en Células Vero
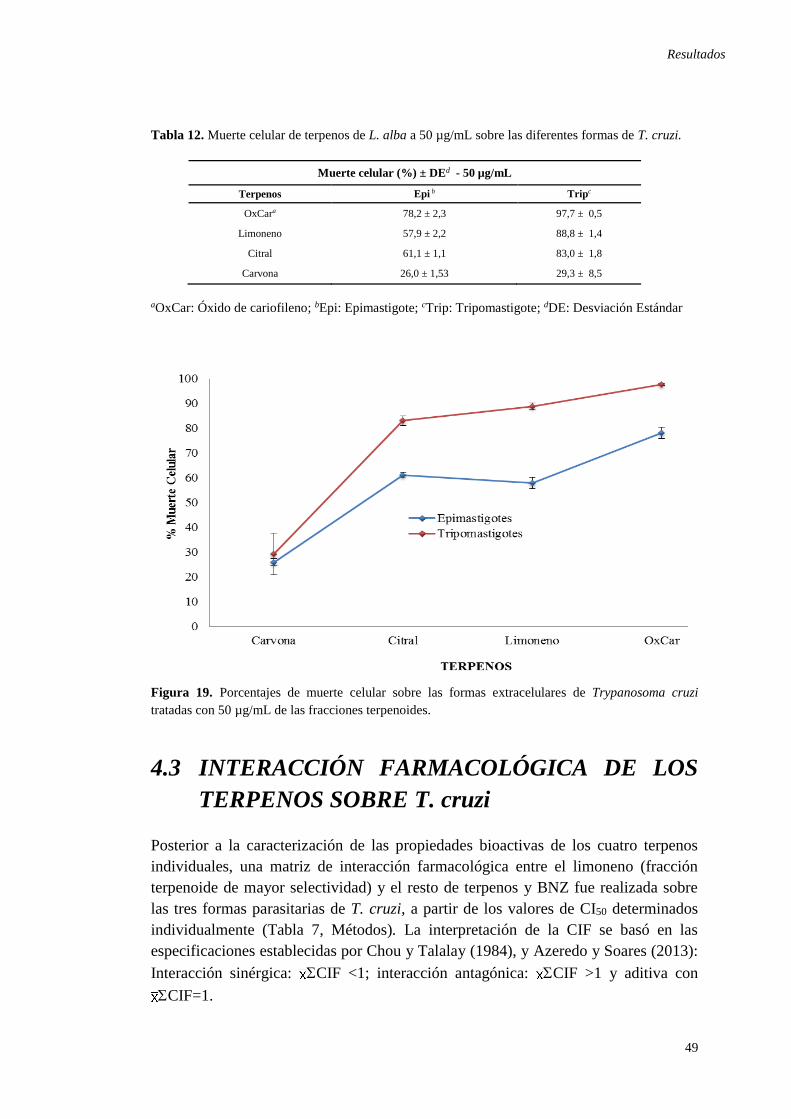
Resultados
49
Tabla 12. Muerte celular de terpenos de L. alba a 50 µg/mL sobre las diferentes formas de T. cruzi.
Muerte celular (%) ± DEd - 50 µg/mL
Terpenos Epi b Tripc
OxCara 78,2 ± 2,3 97,7 ± 0,5
Limoneno 57,9 ± 2,2 88,8 ± 1,4
Citral 61,1 ± 1,1 83,0 ± 1,8
Carvona 26,0 ± 1,53 29,3 ± 8,5
aOxCar: Óxido de cariofileno; bEpi: Epimastigote; cTrip: Tripomastigote; dDE: Desviación Estándar
Figura 19. Porcentajes de muerte celular sobre las formas extracelulares de Trypanosoma cruzi
tratadas con 50 µg/mL de las fracciones terpenoides.
4.3 INTERACCIÓN FARMACOLÓGICA DE LOS
TERPENOS SOBRE T. cruzi
Posterior a la caracterización de las propiedades bioactivas de los cuatro terpenos
individuales, una matriz de interacción farmacológica entre el limoneno (fracción
terpenoide de mayor selectividad) y el resto de terpenos y BNZ fue realizada sobre
las tres formas parasitarias de T. cruzi, a partir de los valores de CI50 determinados
individualmente (Tabla 7, Métodos). La interpretación de la CIF se basó en las
especificaciones establecidas por Chou y Talalay (1984), y Azeredo y Soares (2013):
Interacción sinérgica: CIF <1; interacción antagónica: CIF >1 y aditiva con
CIF=1.

Resultados
50
A excepción de la combinación limoneno + carvona (Epimastigotes: CIF = 1,10;
Tripomastigotes: CIF = 1,04), todas las interacciones evaluadas presentaron un
efecto sinérgico, para las formas epimastigotes, tripomastigotes y amastigotes de T.
cruzi (Tabla 13). La combinación limoneno + BNZ, mostró el mayor efecto sinérgico
frente a los tres estadios parasitarios: Epimastigotes: CIF = 0,44; Tripomastigotes:
CIF = 0,42; Amastigotes: CIF = 0,58 (Tabla 13, Figura 20D), siendo la mezcla de
4 veces la CI50 de limoneno y la mitad de la CI50 de Benznidazol (limoneno
4xCI50=BNZ ½CI50) la de mejor efecto sobre la actividad tripanocida del
medicamento de referencia, causando una reducción de su CI50 sobre Amastigotes,
Epimastigotes y Tripomastigotes de 14, 16 y 17 veces, respectivamente. No obstante,
para esta asociación, una actividad sinérgica también fue observada sobre las células
Vero (CIF = 0,54). La mezcla Limoneno + óxido de cariofileno, representó la
segunda combinación de mayor eficacia en las formas parasitarias evaluadas
(Epimastigotes: CIF = 0,49; Tripomastigotes: CIF = 0,45; Amastigotes: CIF =
0,71) y además, evidenció un efecto antagonista sobre las células Vero (CIF = 1,22)
(Tabla 13, Figura 20A).
Tabla 13. Interacción farmacológica entre Limoneno y el resto de terpenos derivados de L.alba.
aEpi: Epimastigote; bTrip: Tripomastigote; cAmas: Amastigote; dOxCar: Oxido de Cariofileno; eBNZ:
Benznidazol; fCIF: Concentración Inhibitoria Fraccional; gDE: Desviación estándar; hND: No
determinado
Los datos de la interacción farmacológica también se expresaron gráficamente como
isobologramas, trazando las concentraciones inhibitorias fraccionales medias de cada
combinación (Figura 20).
L Limoneno+Terpenos
CIFf µg/mL
± DEg
Interacción
farmacológica
Epia OxCard 0,49 ± 0,13 Sinérgico
Carvona 1,10 ± 0,08 Antagonista
Citral 0,74 ± 0,13 Sinérgico
BNZe 0,44 ± 0,13 Sinérgico
Tripb OxCar 0,45 ± 0,09 Sinérgico
Carvona 1,04 ± 0,04 Antagonista
Citral 0,60 ± 0,14 Sinérgico
BNZ 0,42 ± 0,10 Sinérgico
Amasc OxCar 0,71 ± 0,23 Sinérgico
Carvona NDi ND
Citral 0,82 ± 0,15 Sinérgico
BNZ 0,58 ± 0,13 Sinérgico
Vero OxCar 1,22 ± 0,16 Antagonista
Carvona 0,98 ± 0,07 Aditivo
Citral 1,04 ± 0,07 Antagonista
BNZ 0,54 ± 0,18 Sinérgico

Resultados
51
Figura 20. Isobologramas de las interacciones farmacológicas entre el limoneno y otros terpenos de
AEs de L. alba: A) con óxido de cariofileno; B) con carvona; C) con citral; y D) con el medicamento
de referencia Benznidazol (BNZ). Los análisis fueron realizados sobre las tres formas parasitarias de
T. cruzi y células Vero. Las líneas de puntos corresponden a un efecto aditivo. Los puntos debajo,
sobre y por encima de la línea indican un efecto sinérgico, aditivo y antagónico, respectivamente.
4.4 MECANISMO DE MUERTE CELULAR INDUCIDO
POR LOS COMPUESTOS EN ESTUDIO
La evaluación del fenotipo de muerte celular desencadenado por el tratamiento en
epimastigotes de T. cruzi con el AE (A23), terpenos (óxido de cariofileno, limoneno o
citral) e interacción de mayor sinergia (limoneno+BNZ) fue realizada mediante el
análisis de cambios morfológicos, pérdida del potencial de membrana mitocondrial,
fragmentación nuclear y externalización de la fosfatidilserina.
4.4.1 Cambios celulares
Para la evaluación de los cambios morfológicos inducidos por el tratamiento con los
diversos compuestos (AEs, terpenos y BNZ), fueron realizados análisis por microscopía
óptica y de fluorescencia, mediante contraste de fase y coloraciones nucleares (DAPI) y
0
0,2
0,4
0,6
0,8
1
1,2
0 0,2 0,4 0,6 0,8 1 1,2
CIF
Óxid
o d
e C
ario
file
no
CIF Limoneno
Tripomastigotes
Epimastigotes
Células Vero
Amastigotes
A
0
0,2
0,4
0,6
0,8
1
1,2
0 0,2 0,4 0,6 0,8 1 1,2
CIF
Ca
rvo
na
CIF Limoneno
Tripomastigotes
Epimastigotes
Células Vero
B
0
0,2
0,4
0,6
0,8
1
1,2
0 0,2 0,4 0,6 0,8 1 1,2
CIF
Cit
ra
l
CIF Limoneno
Tripomastigotes
Epimastigotes
Células Vero
Amastigotes
C
0
0,2
0,4
0,6
0,8
1
1,2
0 0,2 0,4 0,6 0,8 1 1,2
CIF
BN
Z
CIF Limoneno
Tripomastigotes
Epimastigotes
Células Vero
Amastigotes
D

Resultados
52
de potencial Redox de membrana mitocondrial (Mitotracker Red CMXRos (Pendergrass
et al., 2004)). Como puede ser observado en la Figura 21, todos los tratamientos
estudiados indujeron alteraciones significativas en la estructura de la membrana,
provocando una conformación esférica del epimastigote. Adicionalmente, fue
evidenciada pérdida del volumen citoplasmático (Figura 21 DIC), reducción del
potencial de membrana mitocondrial (Figura 21 MitoTracker) y la adquisición de un
patrón moteado de la cromatina, con zonas de condensación en el núcleo (Figura 21
DAPI). Estas alteraciones sugieren el desencadenamiento de un posible mecanismo de
muerte celular tipo apoptótico. Por otra parte, el tratamiento de los parásitos con óxido
de cariofileno provocó también pérdida del flagelo. Vale la pena mencionar, que las
células hospederas tratadas bajo las mismas condiciones, no presentaron alteraciones
morfológicas apreciables (datos no mostrados). De manera distinta, el tratamiento con el
BNZ arrojó características morfológicas sugestivas de necrosis, con aumento del
volumen citoplasmático, pérdida de la continuidad de la membrana y conservación del
potencial energético mitocondrial.
DIC DAPI MitoTracker
Figura 21. Pérdida del potencial de membrana mitocondrial en epimastigotes de T.cruzi observados
mediante microscopía de contraste de fases interferencial (DIC). El ADN parasitario fue visualizado
con la sonda DAPI y la pérdida del potencial de membrana se demostró a través de la disminución de
la intensidad de fluorescencia del colorante MitoTrackerRed.
O
xC
AR
A23 E
PI.
SIN
TR
AT
AR
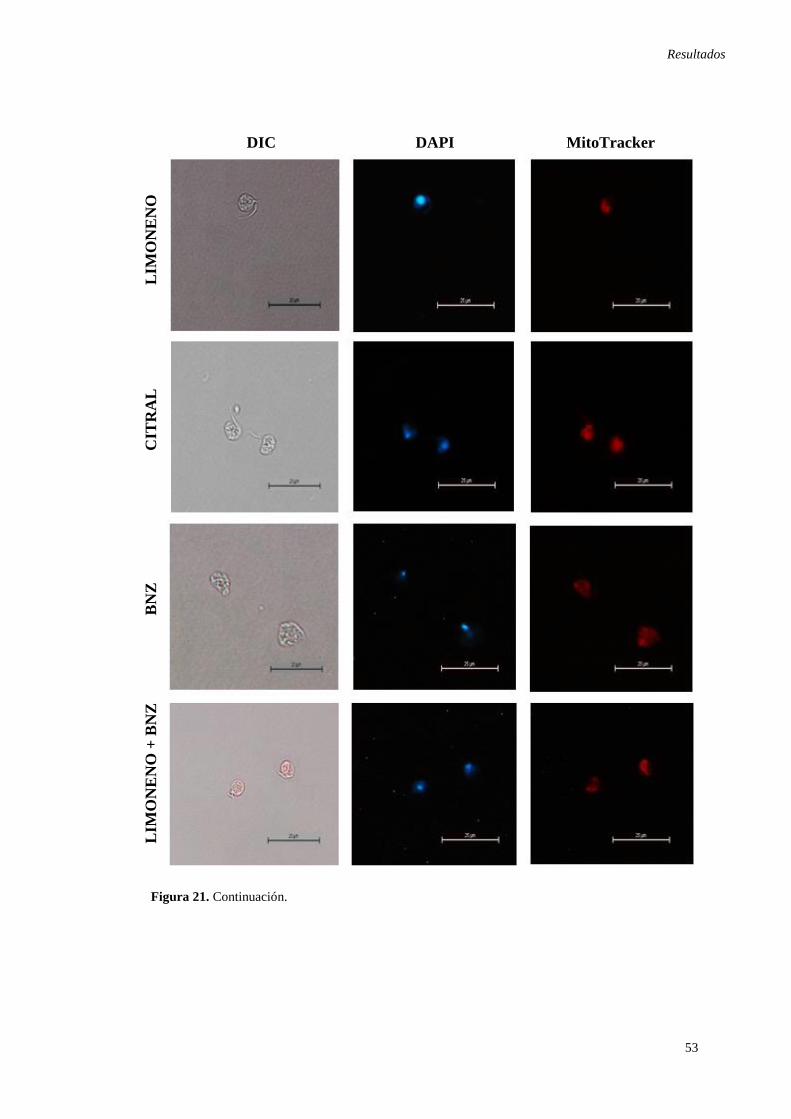
Resultados
53
DIC DAPI MitoTracker
Figura 21. Continuación.
LIM
ON
EN
O +
BN
Z
B
NZ
C
ITR
AL
LIM
ON
EN
O

Resultados
54
4.4.2 Fragmentación del ADN
Para determinar un posible efecto sobre la activación de endonucleasas, un análisis de
fragmentación del ADN por electroforesis en gel de agarosa fue realizado sobre
epimastigotes de T.cruzi, tratados con dos veces la CI50 (CI50X2) de cada compuesto,
durante 48 horas. Los tratamientos estudiados indujeron degradación del ADN, con
desaparición de las bandas, pero sin la obtención de un patrón de escalera visible. Este
patrón oligonucleosomal fue obtenido únicamente con el control utilizado proveniente de
un cultivo de más de dos semanas. Los fragmentos se observaron en la región
comprendida entre las bandas con pesos moleculares de 200 pb y 1000 pb. Un fragmento
adicional se evidenció inferior a la banda de 100 pb (Figura 22, flechas). Por el contrario,
en epimastigotes no tratados se evidenció la preservación de una banda de ADN fuerte,
cercana al sitio de siembra (en la electroforesis).
Figura 22. Electroforesis en gel de agarosa Low Melting al 1.8% del ADN de epimastigotes de T.
cruzi tratados con los compuestos en estudio. Las flechan indican la presencia de fragmentación
oligonucleosomal. aEpi: Epimastigote; bOxcar: Óxido de Cariofileno; cMP: Marcador de Peso
Molecular GeneRuler 1Kb DNA Ladder (Invitrogen); dCP: Control positivo: Cultivo de parásito de 15
días; eBNZ: Benznidazol.
Para confirmar esta posible degradación del ADN documentada en el gel de
electroforesis, fue realizado un ensayo de TUNEL por microscopía de fluorescencia.
Epimastigotes tratados por 48 horas con IC502X, fueron incubados con desoxinucleotidil
transferasa terminal marcada con fluoresceína dUTP y la sonda fluorescente HOECHST.
La fragmentación se evidenció por la emisión de fluorescencia verde en el núcleo y en el
kinetoplasto. Simultáneamente se demostró presencia de ADN con fluorescencia azul en

Resultados
55
ensayos de TUNEL negativos. Los epimastigotes tratados con los terpenos limoneno,
óxido de cariofileno y la combinación limoneno + BNZ, presentaron TUNEL positivo
con porcentajes de 94%, 99% y 98%, respectivamente. No se observó fragmentación
significativa en los cultivos de epimastigotes sin tratamiento (Figura 23).
DIC HOECHST TUNEL
Figura 23. Fragmentación de ADN en epimastigotes de T.cruzi observados mediante microscopia de
contraste de fases interferencial (DIC). El ADN parasitario fue visualizado con la sonda fluorescente
azul HOECHST y los extremos libres de ADN se observaron de color verde, indicando epimastigotes
positivos para el ensayo de TUNEL. Las fotografías son representativas de 10 campos observados.
L
IMO
NE
NO
OxC
AR
A23
EP
I. S
IN T
RA
TA
R
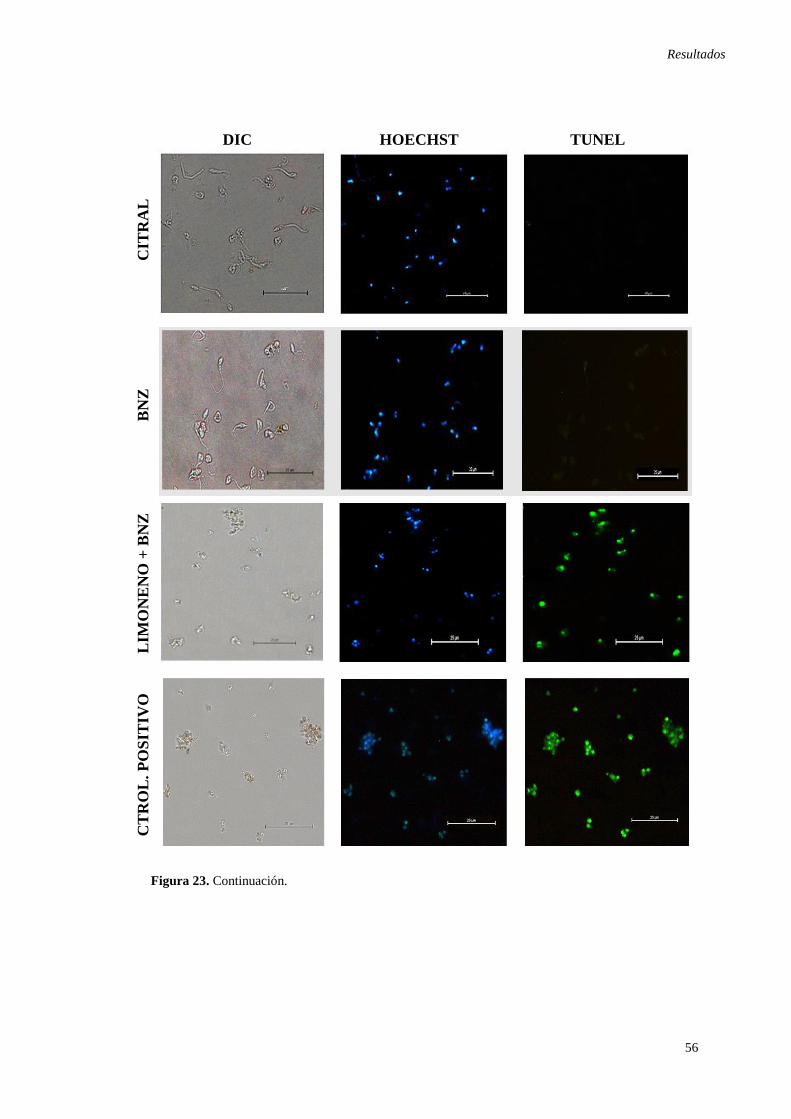
Resultados
56
DIC HOECHST TUNEL
Figura 23. Continuación.
CT
RO
L.
PO
SIT
IVO
L
IMO
NE
NO
+ B
NZ
B
NZ
CIT
RA
L

Resultados
57
4.4.3 Externalización de la fosfatidilserina
Un análisis de citometría de flujo se empleó en epimastigotes de T. cruzi para
determinar el mecanismo de muerte celular inducido por los compuestos en estudio.
Los parásitos fueron co-incubados con las sondas Anexina V y SYTOX Green, para
evaluar la expresión de la fosfatidilserina y la discontinuidad de la membrana,
respectivamente. Epimastigotes control sin tratamiento mostraron 99.7% de
viabilidad (Figura 24A), mientras que los cultivos incubados por 48 horas con dos
veces la CI50 (CI50X2) de cada compuesto, ratificaron muerte celular con porcentajes
significativos de inhibición (óxido de cariofileno: 95,7%; limoneno: 89,2%; citral:
78,4% y limoneno + BNZ: 95,9%, p=0.000). Ningún tratamiento evidenció valores
estadísticamente importantes de SYTOX-Negativo + Anexina V-positiva (p= >0.05),
excepto el control de apoptosis (Cultivo de parásito de 15 días) (13,6%) (Figura
24F). Por el contrario, todos los cultivos tratados mostraron altos porcentajes de
SYTOX-positivo + Anexina V-positiva, en comparación con los epimastigotes sin
tratamiento; resultados que probablemente corresponden a una apoptosis
tardía/Necrosis secundaria (Figura 24).

Resultados
58
Figura 24. Análisis por citometría de flujo de la externalización de la fosfatidilserina en epimastigotes
de T. cruzi. A) Parásitos sin tratamiento. Cultivos incubados con: B) óxido de cariofileno; C)
limoneno; D) citral; E) limoneno + BNZ y F) control positivo de apoptosis (Cultivo de parásito de 15
días). El porcentaje de células positivas para cada perfil, se indica en los diferentes cuadrantes. Las
gráficas son representativas de dos experimentos independientes.

Discusión
59
5. DISCUSIÓN
Reconocida por la Organización Mundial de la Salud (OMS) como un grave
problema de salud pública en América Latina, la Enfermedad de Chagas constituye
una de las más importantes infecciones tropicales desatendidas y la principal causa
de miocardiopatía infecciosa a nivel mundial (OMS, 2015; Cucunubá et al., 2016).
Actualmente, esta parasitosis afecta alrededor de 7 millones de personas en el mundo
con un aproximado de 7000 muertes anuales. Esta enfermedad de prolongado curso
silencioso, representa una importante carga económica calculada en 338,500 (95%
UI: 183,800- 846,400) años de vida ajustados por discapacidad (DALYs) (Murray et
al., 2015) y un costo anual de 7,19 mil millones de dólares (Hotez et al., 2013; Lee et
al., 2013). En Colombia, el costo económico del tratamiento estimado por paciente-
año es de US $ 1,028, en función de la gravedad y el nivel de cuidado implementado,
con un costo de vida útil promedio de US $ 11,619 por paciente (Castillo et al.,
2008).
A pesar de su impacto socio-económico, la enfermedad de Chagas continúa siendo
endémica en muchas regiones del Centro y Sur de América (OMS, 2015), afectando
inclusive otras zonas del mundo como Europa, América del Norte, Japón y Australia,
debido a la migración de poblaciones humanas (Gascon et al., 2010; Jackson et al.,
2014).; sin que existe evidencia contundente de una medida profiláctica o de un
tratamiento adecuado para la eliminación de la infección o para la cura de la fase
crónica de la enfermedad (Bernacchi et al., 2002; Morens et al., 2004).
En la actualidad el manejo terapéutico de la Tripanosomiasis Americana continúa
siendo un desafío. Los fármacos convencionales, esencialmente tripanocidas
Nifurtimox (Lampit®, Bayer) y Benznidazol (Rochagan® en Brazil y Radanil® en
Argentina, Roche) (Bustamante y Tarleton, 2014), han fallado en alcanzar la cura
parasitológica y la detención de la progresión de la enfermedad, principalmente en
las fases tardías de esta patología. Como consecuencia, el interés por la “terapéutica
de multi-componentes” ha venido en aumento (Kong et al., 2008) y en este contexto,
fármacos extraídos de plantas o sintetizados a partir de prototipos vegetales
constituyen una excelente opción, ya que ofrecen una importante plataforma para
identificar y desarrollar drogas sinérgicas contra enfermedades de génesis compleja
como el Chagas (Dancey y Chen, 2006).
Vale la pena mencionar, que los protozoarios contienen diferentes vías metabólicas
esenciales, ausentes en los seres humanos, que representan blancos farmacológicos
ideales. Es el caso de la vía del tripanotión (responsable del metabolismo tiol), quien
desempeña actividades protectoras contra el estrés oxidativo, mediante la renovación
del NADPH dependiente de especies de tiol. Estudios anteriores han demostrado que

Discusión
60
la inhibición de esta vía, bloquea la capacidad del parásito para resistir el ataque
oxidativo (de ROS y NOS) montado por las defensas del macrófago (Hossain et al.,
2016). Teniendo en cuenta esta sensibilidad del protozoo al estrés oxidativo, los
derivados nitroimidazólicos, así como nitrofuranos, fundamentan su actividad
tripanocida por inducción de especies reactivas de oxígeno (ROS). Sin embargo,
estos fármacos presentan limitaciones asociadas a su alta toxicidad y efectos
adversos severos, además de su potencial mutagénico. Ambos tienen efectos sobre
las formas tripomastigotes en la fase aguda de la enfermedad, pero no se ha
evidenciado un beneficio contundente de estos medicamentos en la fase crónica
(Morillo et al., 2015).
La acción del nifurtimox depende de la generación de radicales de oxígeno y
peróxido, y el efecto del benznidazol obedece a la unión covalente de metabolitos
electrofílicos a macromoléculas intracelulares, como el glutatión (en menor medida)
y tripanotión y tripanotión reductasa (principalmente). La inhibición del benznidazol
a estos dos últimos, provoca la disminución de los niveles de tiol, que son esenciales
para el mantenimiento del potencial de óxido reducción del parásito. Así mismo, se
han descrito recientemente efectos adicionales del benznidazol, como el daño al
ADN genómico del protozoo, que conlleva al desenrollamiento extenso de la
heterocromatina y sobreexpresión de proteínas de reparación del ADN. Estos
mecanismos podrían contribuir al desarrollo de resistencia a la droga, por parte del
parásito, así como al efecto mutagénico observado sobre las células del huésped
(Rajao et al., 2014).
En este contexto, la revisión de nuevas moléculas se ha centrado principalmente en la
utilización de agentes que alteran el potencial redox celular y se aprovechan de las
deficiencias del parásito, en sus mecanismos de defensa contra el estrés oxidativo
(Wilkinson et al., 2002; Paulino et al., 2005). Un efecto tripanocida similar, ha sido
descrito para mezclas de aceites esenciales (AEs) ricas en terpenos (Santoro et al.,
2007a y b; Cardoso y Soares, 2010; Escobar et al., 2010; Kpoviessi et al., 2014). Los
AEs son extractos volátiles obtenidos a través de destilación al vapor de las plantas
(Post-White y Nicholson, 2007) y sus valores medicinales han aumentado
considerablemente en las últimas décadas. Como antimicrobianos, múltiples trabajos
han definido sus efectos antibacterianos (Murbach Teles Andrade et al., 2014),
antivirales (Gilling et al., 2014), antimicóticos (Venturi et al., 2015) y
antiparasitarios (Kpoviessi et al., 2014). En el caso de T. cruzi, resultados
promisorios fueron obtenidos sobre cepas tratadas con AEs derivados de
Cymbopogon citratus, los cuales exhibieron valores de CI50 de 15,5 µg/mL y 5,1
µg/mL, contra tripomastigotes y amastigotes, respectivamente (Santoro et al.,
2007a). Estos efectos fueron atribuidos al monoterpeno oxigenado citral, uno de sus
componentes mayoritarios (Santoro et al., 2007b). Resultados similares fueron
encontrados en aceites derivados de una variante Colombiana (Santander) de Lippia

Discusión
61
alba quimiotipo Citral, los cuales eran ricos en citral (mezcla de neral y geranial),
geraniol, timol y óxido de cariofileno (Escobar et al., 2010).
En el presente trabajo, se evidenciaron valores medios de CI50 de 13,6 µg/mL y 21,9
µg/mL para mezclas complejas derivadas de L. alba ricas en citral y óxido de
cariofileno (quimiotipo Citral) y concentraciones inhibitorias medias de 88,2 µg/mL
y 44,9 µg/mL para el quimiotipo Carvona de esta misma planta (mezcla rica en
carvona y limoneno) en epimastigotes y tripomastigotes, respectivamente. De manera
individual, el citral mostró inhibir eficazmente las formas epimastigotas de T. cruzi,
con una CI50 de 37,2 µg/mL. En los tripomastigotes derivados de células se demostró
igual susceptibilidad a este terpeno (CI50 20,8 µg/mL). Estos resultados concuerdan
con los obtenidos por Cardoso y Soares (2010), quienes determinaron actividad
tripanocida in vitro del citral en las formas extracelulares no replicativas, con una
CI50 de 24,5 μg/mL.
El mecanismo antiproliferativo de este terpenoide se ha estudiado ampliamente en
otros modelos celulares, principalmente en células cancerígenas. El citral ha sido
identificado como el mayor componente de la supresión de Ciclooxigenasa 2
(COX2) y como un activador de PPAR (Receptores Activados por Proliferadores de
Peroxisomas) α y γ. La supresión del promotor de COX2 dependiente de PPAR γ fue
observada al tratamiento con este terpenoide, en macrófagos humanos estimulados
por lipopolisacaridos (LPS), de manera dosis dependiente. Evidencia creciente indica
que la expresión de COX2 es regulada de manera distinta en diferentes tipos
celulares y juega un rol importante en la génesis tumoral, la inflamación, el
desarrollo y la homeostasis circulatoria; por lo que se sugiere al citral como un
fármaco promisorio para enfermedades inflamatorias y el cáncer (Katsukawa et al.,
2010). Otras investigaciones, definieron propiedades anti-oxidantes fuertes (5 veces
superiores al α-tocoferol) (Rabi y Bishayee, 2009; Stashenko et al., 2014) y
antigenotóxicas potentes para AEs de L. alba, quimiotipo citral y carvona, y sus
principales constituyentes (citral, carvona y limoneno), siendo el citral el compuesto
más activo (López et al., 2011). Así mismo, se evidencia la capacidad de este terpeno
para modular el daño del ADN inducido por doxorrubicina (fármaco cardiotóxico) y
la inhibición de la expresión de COX2 y factor de necrosis tumoral (TNF) alfa en
macrófagos activados por LPS (de Paula et al., 2014). Por otro lado, citral bloquea la
expresión génica de la enzima Óxido Nítrico Sintasa (NOS) inducida por LPS
(iNOS) de forma concentración dependiente. Esta inhibición estaría suprimiendo de
manera potencial la actividad transcripcional y la translocación del factor nuclear
kappa de las células B activadas (NF-ƙB). Estos resultados sugieren que citral es un
agente anti-inflamatorio, y sus efectos pueden deberse a la inhibición de la
producción de óxido nítrico (NO), a través de la supresión de la activación de NF-ƙB.
(Lee et al., 2008). Corroborando esta actividad anti-inflamatoria, Liao et al., (2015),

Discusión
62
determinó el efecto inmunomodulador del citral al inhibir mediadores pro-
inflamatorios en macrófagos estimulados con LPS (NLRP2, TNF-α, IL-6 e IL-1β).
De forma semejante, en el trabajo de Ku y Lin, (2013), fue demostrada una potente
actividad anti-inflamatoria para el óxido de cariofileno, el cual incrementó
marcadamente el coeficiente de mediadores de la respuesta inmune Th2/Th1. En
nuestro estudio, este sesquiterpeno fue encontrado como componente de los aceites
de L. alba de mayor actividad tripanocida, tales como A23 y A20 (quimiotipo Citral).
En estos AEs, fueron utilizados tiempos de extracción superiores a los 60 minutos, lo
cual garantizó el incremento de este tipo de metabolitos de mayor peso molecular.
Diversas investigaciones han asociado la presencia del óxido de cariofileno a la
actividad tripanocida. De esta manera, Cheikh Ali et al., (2011), describieron la
capacidad de inhibir el crecimiento de T. brucei por AEs derivados de Aframomum
sceptrum (concentraciones letales mínimas de 0,1 µg/mL), relacionando esta
actividad a la presencia de este compuesto. Coherente con estos resultados, en
nuestro estudio, la actividad individual del óxido de cariofileno, demostró un efecto
tripanocida significativo (Epimastigotes: CI50 29,8 µg/mL; Tripomastigotes: CI50 21,6
µg/mL; Amastigotes: CI50 47,4 µg/mL) contra T. cruzi.
En modelos celulares de cáncer de próstata y cáncer de mama, se ha logrado
identificar que el óxido β-cariofileno, aislado a partir de aceites esenciales de plantas
medicinales como la guayaba (Psidium guajava), el orégano (Origanum vulgare L.),
la canela (Cinnamomum spp.), el Clavo de olor (Caryophyllata eugenia) y la
pimienta negra (Piper nigrum L.) inhiben varias vías de señalización intracelular que
lideran la muerte celular programada. Algunas de estas vías son PI3K, AKT, mTOR,
S6K1 y proteína activada por mitógeno quinasa (MAPK), las cuales desempeñan un
papel importante en la proliferación celular, supervivencia, angiogénesis y metástasis
de las células tumorales. Adicionalmente la génesis del fenotipo apoptótico por parte
del óxido de cariofileno, también ha sido vinculada al incremento de estrés oxidativo
por ROS mitocondriales. De acuerdo con lo anterior, podría especularse que el efecto
tripanocida observado para este terpeno, estaría relacionado con mecanismos
similares a los utilizados por BNZ y NFX, los cuales actúan aprovechando las
escasas defensas del parásito contra el estrés oxidativo (Paulino et al., 2005).
Adicionalmente, estos hallazgos sugieren que este sesquiterpeno podría interferir con
múltiples cascadas de señalización implicadas en el origen de patologías tumorales e
inflamatorias (Park et al., 2011), como la Enfermedad de Chagas
Otro de los terpenos bioactivos de L. alba abordados en este estudio, fue el limoneno.
Sobre este monoterpeno se han realizado múltiples investigaciones que describen su
baja toxicidad (al ser ampliamente usado como aditivo de alimentos) (Sun, 2007), su
alta biodisponibilidad (Miller et al., 2013) y sus efectos antiproliferativos selectivos
sobre diversas líneas tumorales (Manuele et al., 2009; Rabi y Bishayee, 2009; Xiu-

Discusión
63
Zhen, 2009; Kapoor, 2013). En este sentido, estudios in vivo e in vitro han ratificado
sus propiedades anticancerígenas, pricipalmente sobre tumores sólidos de próstata y
mama (Rabi y Bishayee, 2009; Vigushin et al., 1998; Miller et al., 2013). En cáncer
de próstata, fue determinada la capacidad de inducir muerte celular apoptótica por
estrés oxidativo, de manera selectiva sobre el tejido tumoral (Rabi y Bishayee, 2009).
Esta selectividad fue asociada a los niveles incrementados de estrés celular
(comparados con las normales), inducidos por la alta tasa metabólica (proliferativa) y
por la deficiencia en los mecanismos de defensa antioxidante de la célula tumoral
(Szatrowski and Nathan, 1991; Trachootham et al., 2006). En estas células,
cantidades mayores de ROS provenientes de la mitocondria, son necesarias para la
proliferación resultante de la estimulación de factores de crecimiento (Trachootham
et al., 2006). Debido a este incremento en el nivel basal de ROS, la célula tumoral es
más susceptible a sufrir muerte celular por el tratamiento con agentes pro-oxidantes.
De esta manera, muchos quimioterapéuticos lideran la apoptosis por un incremento
en el estrés oxidativo e inhibición de sistemas antioxidantes, que presionan a las
células del tumor más allá de su punto de quiebre, en términos de lipo-peroxidación,
daño del ADN y oxidación de proteínas. Es el caso de inhibidores de histonas
deacetilasas (Adachi et al., 2004), inhibidores de proteasoma (Perez-Galan et al.,
2006) y agentes redox ciclantes (Lecane et al., 2005).
Teniendo en cuenta la acción antiproliferativa y pro-oxidante del limoneno, en
modelos celulares altamente susceptibles al estrés oxidativo y con pobres defensas
antioxidantes, y la similitud de esta característica con T. cruzi, en este estudio fue
indagada la actividad individual de este monoterpeno, a pesar de su pobre desempeño
en la mezcla de AEs del quimiotipo Carvona (rico en limoneno) (Media
Epimastigotes: CI50 88,2±3,7 µg/mL; Tripomastigotes: CI50 44,9±2,5 µg/mL;
Amastigotes: CI50 >150 µg/mL). Los ensayos realizados, demostraron que el
limoneno presentó el mejor efecto tripanocida (de compuestos individuales
probados) sobre las formas presentes en el hospedero mamífero (Tripomastigotes:
CI50 9,0 µg/mL; Amastigotes: CI50 28,7 µg/mL). Estos resultados fueron similares a
los obtenidos por Santos et al. (2014), quienes evidenciaron una CI50 de 14,1 µg/mL
para este terpenoide, en las formas tripomastigotas de T. cruzi. Además estos autores
determinaron una reducción moderada en el porcentaje de macrófagos infectados y
en el número de parásitos intracelulares, en concentraciones no tóxicas para las
células mamíferas (Santos et al., 2014)
De acuerdo con esto, se concluyó que el efecto tripanocida del limoneno, fue
únicamente evidente fuera de su contexto del aceite esencial del quimiotipo Carvona
de L. alba. Esta falta de actividad antipoliferativa puede ser explicada por las altas
concentraciones medias del monoterpoeno carvona en el aceite (en media de 41.9%),
el cual ha sido descrito como un fuerte antioxidante (Stashenko et al., 2004). Con el
fin de dilucidar un posible antagonismo en su actividad tripanocida entre estos dos

Discusión
64
compuestos (limoneno y carvona), en este trabajo fueron realizados ensayos de
interacción farmacológica. Estos ensayos confirmaron que la presencia de carvona en
la mezcla disminuyó el desempeño del limoneno sobre las formas tripomastigotas
(CIF: 1,04 µg/mL) y epimastigotas (CIF: 1,10 µg/mL) de T. cruzi.
Basados en estos resultados promisorios del limoneno, indagaciones posteriores de
posibles interacciones farmacológicas sinergísticas de este terpeno fueron realizados
usándolo en combinación con los otros dos terpenos bioactivos de este estudio (citral
y óxido de cariofileno), así como con el medicamento de referencia (BNZ). Los
resultados encontrados indicaron un efecto sinérgico en todas las combinaciones
evaluadas, con valores de CIF menores a 0,8 µg/mL, sobre las tres formas
parasitarias. La mejor sinergia fue alcanzada con la mezcla limoneno + BNZ,
causando una reducción de la CI50 del fármaco sobre amastigotes, epimastigotes y
tripomastigotes de 14, 16 y 17 veces, respectivamente. Estas interacciones positivas
podrían ser el resultado de la acción de estos compuestos sobre mecanismos diversos
y aditivos que lideran la muerte celular en líneas susceptibles, tales como: a)
disrupción de la polimerización de microtúbulos e inducción del estrés del retículo
endoplasmático (citral) (Chaimovitsh et al., 2012); b) inhibición de vías de
supervivencia como PIP3/AKT (limoneno, óxido de cariofileno y citral) (Park et al.,
2011; Jia et al., 2013; Xia et al., 2013); c) inducción de estrés oxidativo (limoneno,
citral y oxido de cariofileno) (Rabi y Bishayee, 2009; Park et al., 2011; Kapur et
al.,2016; Thomas et al.,2016); d) e inducción de apoptosis a través de caspasas, entre
otros (limoneno, carvona) (Yu et al., 2008).
De acuerdo con lo anterior, las interacciones farmacológicas encontradas entre el
limoneno y los terpenos citral y óxido de cariofileno, así como con el BNZ, resultan
interesantes si se tiene en cuenta que el uso de múltiples fármacos con diferentes
modos de acción, contra una patología como la Enfermedad de Chagas, potenciaría
su tratamiento de una manera más eficaz. Diversos resultados favorables por el
sinergismo farmacológico incluyen el aumento de la eficacia del efecto terapéutico,
disminución de la dosis (para evitar toxicidad) y retraso en el desarrollo de
resistencias (Chou, 2006). En este contexto, en las últimas décadas, las interacciones
farmacológicas han sido ampliamente exploradas y se han convertido en la elección
principal para el tratamiento de enfermedades complejas como el cáncer y
enfermedades infecciosas, incluída la Tripanosomiasis Americana (Azeredo et al.,
2013; de Figueiredo Diniz et al., 2013; Polanco-Hernández et al., 2013).
Vale la pena mencionar, que en este estudio fue rastreado el fenotipo de muerte
inducido por el tratamiento con los aceites esenciales y sus terpenos bioactivos. En
este sentido, las observaciones de microscopía óptica y de fluorescencia, sobre cepas
de T. cruzi tratadas durante 48 horas con los terpenos en estudio, sugirieron la
inducción de un posible mecanismo de muerte celular programada (redondeamiento

Discusión
65
celular, pérdida del volumen citoplasmático, ausencia del flagelo, disminución del
potencial de membrana y fragmentación del núcleo). En estudios anteriores con
tripanosomatídeos tratados con inhibidores de esteroles (Menna-Barreto et al., 2009)
o con inhibidores de enzimas mitocondriales (García et al., 2007), estas
características fueron asociadas a posibles mecanismos de muerte celular programada
tipo apoptosis o autofagia. No obstante, la existencia de una muerte celular
programada en parásitos protozoarios en los últimos años, ha sido objeto de grandes
polémicas respecto a sus orígenes evolutivos. En Plasmodium y Leishmania diversas
investigaciones han evidenciado características típicas de muerte celular por
apoptosis, tales como condensación nuclear, encogimiento celular, fragmentación del
ADN y acumulación de citocromo c oxidasa (Arambage et al., 2009; Ameisen et al.,
1995; Duszenko et al., 2006; Das et al., 2008; Ali et al., 2010). A la fecha, en
microorganismos unicelulares han sido confirmadas las funciones de sólo dos
factores apoptóticos homólogos de las nucleasas de mamíferos (factor de inducción
de apoptosis y Endonucleasa G) (Akematsu y Endoh, 2010; Gannavaram et al.,
2008). Sin embargo, se recomienda cierta precaución al interpretar estos hallazgos,
ya que muchas de estas nucleasas también tienen funciones no apoptóticas.
Con la finalidad de esclarecer estos hallazgos generales, en este trabajo fueron
realizados ensayos con técnicas de citometría de flujo y coloraciones específicas para
el ADN y el potencial de membrana mitocondrial. De esta forma, para la detección
de la fragmentación nuclear, epimastigotes tratados por 48 horas con los terpenos
citral, óxido de cariofileno, limoneno y la combinación limoneno-BNZ fueron
coloreados con DAPI (Jiménez et al., 2008). En todos los tratamientos fue obtenido
un patrón moteado característico de degradación del ADN. Esta fragmentación fue
confirmada por la obtención de positividad en la prueba de TUNEL. Sin embargo,
debido a que el ensayo de TUNEL no consigue discriminar entre muerte celular
apoptótica y necrótica (Charriaut-Marlangue y Ben-Ari, 1995; Grasl-Kraupp et al.,
1995), un análisis posterior de externalización de fosfatidilserina fue realizado por
citometría de flujo. Para esto, se utilizó la sonda Anexina V y SYTOX Green
(discontinuidad de la membrana) sobre epimastigotes de T. cruzi, tratados con los
compuestos en estudio. Ningún tratamiento mostró valores estadísticamente
importantes de SYTOX-Negativo + Anexina V-positiva (p= >0.05). Por el contrario,
todos los cultivos exhibieron altos porcentajes de SYTOX-positivo + Anexina V-
positiva; resultados que sugieren un posible desencadenamiento de una apoptosis
temprana (antes de 48 horas) que evoluciona rápidamente a apoptosis tardía/necrosis
secundaria, asociada a la fragmentación del ADN. De manera compatible con estos
resultados, las coloraciones específicas para la evaluación de la actividad
mitocondrial, indicaron que las células de T. cruzi tratadas con los compuestos
estudiados presentaban disminución en el potencial de membrana de la organela.

Discusión
66
Finalmente, nuestros resultados permiten concluir que los aceites esenciales
derivados de L. alba quimiotipo Citral, cultivados bajo condiciones estandarizadas y
controladas de siembra, colecta y extracción; presentan una actividad tripanocida
significativa sobre las tres formas cíclicas del T. cruzi. Ensayos posteriores realizados
con las fracciones terpenoides, confirmaron la asociación de esta actividad con la
presencia de los terpenos citral y óxido de cariofileno. En el caso de los aceites del
quimiotipo Carvona, este estudio permitió establecer una relación de antagonismo
entre la carvona y el monoterpeno limoneno, con respecto a su desempeño
tripanocida, teniendo en cuenta que pruebas de citotoxicidad con el limoneno, de
manera indiviual, otorgaron las mejores cualidades antiproliferativas contra T. cruzi.
De igual forma, el limoneno fue el terpeno de mayor selectividad en este estudio
(CC50 297,1 ± 2,4 µg/mL). Adicionalmente, en los ensayos de interaccion
sinergística, el limoneno mostró mejorar la eficiencia tripanocida del citral y óxido
de cariofileno, así como del compuesto de referencia, sobre las tres formas
parasitarias. En el rastreamiento de muerte celular, se evidenció la inducción de un
posible fenotipo tipo-apóptótico. En este contexto, se podría concluir que la
combinación limoneno + óxido de cariofileno, presentó características de desempeño
promisorias (actividad tripanocida significativa sobre los tres estadíos de T.cruzi y
selectividad, mediada por la induccion de un posible mecanismo de muerte no
inflamatorio sobre el parásito), las cuales deben ser confirmadas en posteriores
estudios pre-clínicos. Como ventajas previas a estos estudios, vale la pena mencionar
la seguridad de la baja toxicidad adjudicada al limoneno, la cual ha sido constatada
mediante estudios in vivo (Vigushin et al., 1998), y el uso seguro de este terpeno
como aditivo alimenticio, durante muchas décadas (Van Der Logt et al., 2004). Así
mismo, el limoneno y los otros terpenos bioactivos como agentes tripanocidas,
dilucidados en este trabajo, también ofrecen un potencial uso como agentes
inmunomoduladores en el tratamiento de fases tardías de la Enfermedad de Chagas.
De esta forma, estas terapias podrían ser aplicadas como adyuvantes a los
tratamientos convencionales, mejorando sus niveles de tolerancia y su eficacia
parasitológica. En este sentido, los aceites esenciales de L. alba representarían una
fuente renovable para la extracción de este tipo de compuestos y su explotación
comercial futura.

Conclusiones
67
6. CONCLUSIONES
Los aceites esenciales del quimiotipo Citral exhibieron mayor actividad tripanocida in
vitro, en comparación con el quimiotipo Carvona. La actividad antiparasitaria de estos
AEs fue correlacionada con la presencia de altas concentraciones de citral y óxido de
cariofileno.
El mejor desempeño antiproliferativo sobre T. cruzi fue alcanzado por los AEs A23 y
A20, con valores de CI50 respectivos, sobre epimastigotas de 8,1 y 8,5 µg/mL; y sobre
tripomastigotas de 17,4 y 13,9; y con IS>3.
Sobre amastigotes intracelulares la mejor actividad antiparasitaria fue observada con el
AE A28 (CI50 65,5±4,8).
Entre los terpenos evaluados, el limoneno demostró tener la mayor actividad
antiproliferativa y selectiva in vitro contra las tres formas parasitarias de T. cruzi
(epimastigotes: CI50 41,8 ± 2,5 µg/mL, IS=7,1; tripomastigotes: CI50 9,0 ± 0,8 µg/mL,
IS= 32,8; amastigotes CI50 28,7 ± 0,7 µg/mL, IS=10,3), con porcentrajes de muerte de
88,8 % y 57,9% en tripomastigotes y amastigotes respectivamente, a una concentración
de 50 µg/mL. Adicionalmente, este monoterpeno mostró muy baja toxicidad sobre
células de mamífero Vero (CC50 297,1 ± 2,4 µg/mL).
La combinación del limoneno con óxido de cariofileno, citral o BNZ, fue sinergística
para la actividad antiparasitaria. Por el contrario, hubo antagonismo del efecto
tripanocida en la mezcla limoneno y carvona.
El mejor desempeño tripanocida fue obtenido por la combinación limoneno+óxido de
cariofileno (epimastigotes: CIF 0,49; tripomastigotes: CIF 0,45; amastigotes: CIF
0,71). En esta interacción, la presencia del limoneno disminuyó 8 veces los efectos
tóxicos del óxido de cariofileno, sobre células normales e incremento 13 y 16 veces su
actividad sobre las formas amastigotas y tripomastigotas del parásito. Adicionalmente,
en la mezcla limoneno+BNZ, se potencializó la actividad tripanocida de ambos
compuestos sobre las tres formas de T. cruzi (epimastigotes: CIF 0,44; tripomastigotes:
CIF 0,42; amastigotes: CIF 0,58), aunque con incremento también de su citotoxicidad
sobre células de mamífero (CIF = 0,54).
El mecanismo de muerte celular inducido por el tratamiento del parásito con los terpenos
limoneno, óxido de cariofileno y la combinación limoneno+BNZ, fue aparentemente tipo
apoptótico con redondemamiento de las células, pérdida del potencial de membrana
mitocondrial, fragmentación del ADN y externalización de la fosfatidilserina.

Referencias
68
7. REFERENCIAS
Adachi, M., Zhang, Y., Zhao, X., Minami, T., Kawamura, R., Hinoda, Y., & Imai, K. (2004).
Synergistic effect of histone deacetylase inhibitors FK228 and m-carboxycinnamic acid bis-
hydroxamide with proteasome inhibitors PSI and PS-341 against gastrointestinal adenocarcinoma
cells. Clinical cancer research, 10(11), 3853-3862.
Aggarwal, K. K., Khanuja, S. P. S., Ahmad, A., Santha Kumar, T. R., Gupta, V. K., & Kumar, S.
(2002). Antimicrobial activity profiles of the two enantiomers of limonene and carvone isolated from
the oils of Mentha spicata and Anethum sowa. Flavour and Fragrance Journal, 17(1), 59-63.
Aguiar, J. S., Costa, M. C., Nascimento, S. C., & Sena, K. X. (2008). Antimicrobial activity of Lippia
alba (Mill.) NE Brown (Verbenaceae). Revista Brasileira de Farmacognosia, 18(3), 436-440.
Akematsu, T., & Endoh, H. (2010). Role of apoptosis-inducing factor (AIF) in programmed nuclear
death during conjugation in Tetrahymena thermophila. BMC cell biology, 11(1), 13.
Ali, M., Al-Olayan, E. M., Lewis, S., Matthews, H., & Hurd, H. (2010). Naturally occurring triggers
that induce apoptosis-like programmed cell death in Plasmodium berghei ookinetes. PLoS One, 5(9),
e12634.
Alviano, D. S., Barreto, A. L. S., de Almeida Dias, F., de Almeida Rodrigues, I., dos Santos Rosa, M.
D. S., Alviano, C. S., & de Araújo Soares, R. M. (2012). Conventional therapy and promising plant-
derived compounds against trypanosomatid parasites. Frontiers in microbiology, 283(3), 60-69.
Ameisen, J. C., Idziorek, T., Billaut-Mulot, O., Loyens, M., Tissier, J. P., Potentier, A., & Ouaissi, A.
(1995). Apoptosis in a unicellular eukaryote (Trypanosoma cruzi): implications for the evolutionary
origin and role of programmed cell death in the control of cell proliferation, differentiation and
survival. Cell death and differentiation, 2(4), 285-300.
Anaruma, N. D., Schmidt, F. L., Duarte, M. C. T., Figueira, G. M., Delarmelina, C., Benato, E. A., &
Sartoratto, A. (2010). Control of Colletotrichum gloeosporioides (Penz.) Sacc. in yellow passion fruit
using Cymbopogon citratus essential oil. Brazilian Journal of Microbiology, 41(1), 66-73.
Apt, W., Heitmann, I., Jercic, L., Isabel, M., Jofré, L., Noemí, I., & Zulantay, I. (2008). Guías clínicas
de la enfermedad de Chagas: Parte I. Introducción y epidemiología. Revista chilena de
infectología, 25(3), 189-193.
Arambage, S. C., Grant, K. M., Pardo, I., Ranford-Cartwright, L., & Hurd, H. (2009). Malaria
ookinetes exhibit multiple markers for apoptosis-like programmed cell death in vitro. Parasites &
Vectors, 2(1), 32.
Astani, A., & Schnitzler, P. (2014). Antiviral activity of monoterpenes beta-pinene and limonene
against herpes simplex virus in vitro. Iranian journal of microbiology, 6(3), 149.
Azambuja, C. R., Mattiazzi, J., Riffel, A. P. K., Finamor, I. A., de Oliveira Garcia, L., Heldwein, C.
G., & Llesuy, S. F. (2011). Effect of the essential oil of Lippia alba on oxidative stress parameters in
silver catfish (Rhamdia quelen) subjected to transport. Aquaculture, 319(1), 156-161.
Azeredo, C. M., & Soares, M. J. (2013). Combination of the essential oil constituents citral, eugenol
and thymol enhance their inhibitory effect on Crithidia fasciculata and Trypanosoma cruzi
growth. Revista Brasileira de Farmacognosia, 23(5), 762-768.

Referencias
69
Bahia, M. T., de Andrade, I. M., Martins, T. A. F., do Nascimento, Á. F. D. S., de Figueiredo Diniz,
L., Caldas, I. S., & Ribeiro, I. (2012). Fexinidazole: a potential new drug candidate for Chagas
disease. PLoS Negl Trop Dis, 6(11), e1870.
Bai, J., Zheng, Y., Wang, G., & Liu, P. (2015). Protective effect of D-limonene against oxidative
stress-induced cell damage in human lens epithelial cells via the p38 pathway. Oxidative medicine and
cellular longevity, 2016.
Bakkali, F., Averbeck, S., Averbeck, D., & Idaomar, M. (2008). Biological effects of essential oils–a
review. Food and chemical toxicology, 46(2), 446-475.
Baldissera, M. D., de Freitas Souza, C., Mourão, R. H. V., da Silva, L. V. F., & Monteiro, S. G.
(2015). Trypanocidal action of Lippia alba and Lippia origanoides essential oils against Trypanosoma
evansi in vitro and in vivo used mice as experimental model. Journal of Parasitic Diseases, 1-7.
Barbas, L. A. L., Maltez, L. C., Stringhetta, G. R., de Oliveira Garcia, L., Monserrat, J. M., da Silva,
D. T., Heinzmann, B. M., & Sampaio, L. A. (2017). Properties of two plant extractives as anaesthetics
and antioxidants for juvenile tambaqui Colossoma macropomum. Aquaculture, 469, 79-87.
Barros, L. M., Duarte, A. E., Morais-Braga, M. F. B., Waczuk, E. P., Vega, C., Leite, N. F., &
Kamdem, J. P. (2016). Chemical characterization and trypanocidal, leishmanicidal and cytotoxicity
potential of Lantana camara L. (Verbenaceae) essential oil. Molecules, 21(2), 209.
Benziger, C. P., do Carmo, G. A. L., & Ribeiro, A. L. P. (2017). Chagas Cardiomyopathy: Clinical
Presentation and Management in the Americas. Cardiology Clinics, 35(1), 31-47.
Bern, C. (2015). Chagas’ disease. New England Journal of Medicine, 373(5), 456-466.
Bernacchi, A. S., Franke, D. C. B., & Castro, J. A. (2002). Trypanocidal action of 2, 4-dichloro-6-
phenylphenoxyethyl diethylamine hydrobromide (Lilly 18947) on Trypanosoma cruzi. Acta
Pharmacologica Sinica, 23(5), 399-404.
Blanco, S. B., Segura, E. L., Cura, E. N., Chuit, R., Tulián, L., Flores, I., ... & Gürtler, R. E. (2000).
Congenital transmission of Trypanosoma cruzi: an operational outline for detecting and treating
infected infants in north western Argentina. Tropical Medicine & International Health, 5(4), 293-301.
Bonney, K. M., & Engman, D. M. (2015). Autoimmune pathogenesis of Chagas heart disease: looking
back, looking ahead. The American journal of pathology, 185(6), 1537-1547.
Borges, A. R., de Albuquerque Aires, J. R., Higino, T. M. M., de Medeiros, M. D. G. F., Citó, A. M.
D. G. L., Lopes, J. A. D., & de Figueiredo, R. C. B. Q. (2012). Trypanocidal and cytotoxic activities
of essential oils from medicinal plants of Northeast of Brazil. Experimental parásitology, 132(2), 123-
128.
Bustamante, J. M., & Tarleton, R. L. (2014). Potential new clinical therapies for Chagas
disease. Expert review of clinical pharmacology, 7(3), 317-325.
Campos, M. C. O., Castro-Pinto, D. B., Ribeiro, G. A., Berredo-Pinho, M. M., Gomes, L. H. F., da
Silva Bellieny, M. S., & Leon, L. L. (2013). P-glycoprotein efflux pump plays an important role in
Trypanosoma cruzi drug resistance. Parásitology research, 112(6), 2341-2351.
Cardillo, F., de Pinho, R. T., Antas, P. R. Z., & Mengel, J. (2015). Immunity and immune modulation
in Trypanosoma cruzi infection. Pathogens and Disease, 73(9).
Cardoso, J., & Soares, M. J. (2010). In vitro effects of citral on Trypanosoma cruzi
metacyclogenesis. Memorias do Instituto Oswaldo Cruz, 105(8), 1026-1032.

Referencias
70
Carlier, Y., Torrico, F., Sosa-Estani, S., Russomando, G., Luquetti, A., Freilij, H., & Vinas, P. A.
(2011). Congenital Chagas disease: recommendations for diagnosis, treatment and control of
newborns, siblings and pregnant women. PLoS Negl Trop Dis, 5(10), e1250.
Castillo-Riquelme, M., Guhl, F., Turriago, B., Pinto, N., Rosas, F., Martínez, M. F., & Campbell-
Lendrum, D. (2008). The costs of preventing and treating chagas disease in Colombia. PLoS Negl
Trop Dis, 2(11), e336.
Castro, A., Montalto de Mecca, M., Deaz Gomez, M. I., & Castro, D. (2015). Chagas disease: The
contribution of the Center for Toxicological Research. Acta Bioquimica Clinica Latinoamericana,
49(1), 73-82.
Centers for Disease Control and Prevention. (2013)American trypanosomiasis: epidemiology and risk
factors. Atlanta (Georgia): CDC.
Chaimovitsh, D., Altshuler, O., Belausov, E., Abu‐ Abied, M., Rubin, B., Sadot, E., & Dudai, N.
(2012). The relative effect of citral on mitotic microtubules in wheat roots and BY2 cells. Plant
biology, 14(2), 354-364.
Charriaut-Marlangue, C., & Ben-Ari, Y. (1995). A cautionary note on the use of the TUNEL stain to
determine apoptosis. Neuroreport, 7(1), 61-64.
Chee, H. Y., Kim, H., & Lee, M. H. (2009). In vitro antifungal activity of limonene against
Trichophyton rubrum. Mycobiology, 37(3), 243-246.
Cheikh Ali, Z., Adiko, M., Bouttier, S., Bories, C., Okpekon, T., Poupon, E., & Champy, P. (2011).
Composition, and antimicrobial and remarkable antiprotozoal activities of the essential oil of rhizomes
of Aframomum sceptrum K. Schum. (Zingiberaceae). Chemistry & biodiversity, 8(4), 658-667.
Chou, T. C. (2006). Theoretical basis, experimental design, and computerized simulation of synergism
and antagonism in drug combination studies. Pharmacological reviews, 58(3), 621-681.
Chou, T. C., & Talalay, P. (1984). Quantitative analysis of dose-effect relationships: the combined
effects of multiple drugs or enzyme inhibitors. Advances in enzyme regulation, 22, 27-55.
Cicció, J., & Ocampo, R. (2010). Distribución biogeográfica de Lippia alba (Mill.) NE Br. ex Britton
& Wilson y quimiotipos en América y el Caribe. Normalización de Productos Naturales Obtenidos de
Especies de la Flora Aromática Latinoamericana EdiPUCRS, Porto Alegre p, 107-130.
ClinicalTrials.gov. A Study of the Use of Oral Posaconazole (POS) in the Treatment of Asymptomatic
Chronic Chagas Disease (P05267) (STOP CHAGAS). https://
clinicaltrials.gov/ct2/show/NCT01377480?term=nct01377480& rank=1 (Accessed Feb 13, 2017).
ClinicalTrials.gov. CHICAMOCHA 3 - Equivalence of Usual Interventions for Trypanosomiasis
(EQUITY) (CHICAMOCHA-3). https://clinicaltrials.gov/ct2/show/NCT02369978?term=chicamocha-
3&rank=1 Accessed Feb 21, 2017)
Costa, D. C. M., Vermelho, A. B., Almeida, C. A., Dias, E. P. D. S., Cedrola, S. M. L., Arrigoni-
Blank, M. D. F., & Alviano, D. S. (2014). Inhibitory effect of linalool-rich essential oil from Lippia
alba on the peptidase and keratinase activities of dermatophytes. Journal of enzyme inhibition and
medicinal chemistry, 29(1), 12-17.
Costa, E. V., Dutra, L. M., Salvador, M. J., Ribeiro, L. H. G., Gadelha, F. R., & de Carvalho, J. E.
(2013). Chemical composition of the essential oils of Annona pickelii and Annona salzmannii
(Annonaceae), and their antitumour and trypanocidal activities. Natural product research, 27(11),
997-1001.
Coura, J. R. (1966). Contribução ao estudo da doença de Chagas no estado da Guanabara. Revista
Brasileira De Malariologia E Doencas Tropicais, 18, 9-98.

Referencias
71
Coura, J. R. (2009). Present situation and new strategies for Chagas disease chemotherapy: a
proposal. Memórias do Instituto Oswaldo Cruz, 104(4), 549-554.
Coura, J. R. (2015). The main sceneries of Chagas disease transmission. The vectors, blood and oral
transmissions-A comprehensive review. Memórias do Instituto Oswaldo Cruz, 110(3), 277-282.
Coura, J. R., & Borges-Pereira, J. (2010). Chagas disease: 100 years after its discovery. A systemic
review. Acta tropica, 115(1), 5-13.
Coura, J. R., & Dias, J. C. P. (2009). Epidemiology, control and surveillance of Chagas disease: 100
years after its discovery. Memórias do Instituto Oswaldo Cruz, 104, 31-40.
Coura, J. R., & Viñas, P. A. (2010). Chagas disease: a new worldwide
challenge. Nature, 465(n7301_supp), S6-S7.
Coura, J. R., Viñas, P. A., & Junqueira, A. C. (2014). Ecoepidemiology, short history and control of
Chagas disease in the endemic countries and the new challenge for non-endemic countries. Memórias
do Instituto Oswaldo Cruz, 109(7), 856-862.
Croft, S. L., Barrett, M. P., & Urbina, J. A. (2005). Chemotherapy of trypanosomiases and
leishmaniasis. Trends in parásitology, 21(11), 508-512.
Cruz Bermúdez, H. F., & Moreno Collazos, J. E. (2015). Seroprevalencia de tamizaje de Chagas y
factores asociados a coinfecciòn en un banco de sangre de Colombia durante 2006-2011. Revista
Médica de Risaralda, 21(1), 26-30.
Cucunubá, Z. M., Okuwoga, O., Basáñez, M. G., & Nouvellet, P. (2016). Increased mortality
attributed to Chagas disease: a systematic review and meta-analysis. Parasites & vectors, 9(42), 1-13.
Cucunubá Z, Valencia C, Florez C, Leon C, Castellanos Y, Cardenas A. Pilot program for surveillance
of congenital Chagas disease in Colombia 2010-2011. Int J Infect Dis. 2012;16:e343
Cucunubá, Z. M., Valencia-Hernández, C. A., Puerta, C. J., Sosa-Estani, S., Torrico, F., Cortés, J. A &
Muller, E. Á. (2017). First Colombian consensus on congenital Chagas and clinical approach for
women of child-bearing age diagnosed with Chagas. Infectio, 21(4).
Cunha, M. A., de Barros, F. M. C., de Oliveira Garcia, L., de Lima Veeck, A. P., Heinzmann, B. M.,
Loro, V. L., & Baldisserotto, B. (2010). Essential oil of Lippia alba: a new anesthetic for silver
catfish, Rhamdia quelen. Aquaculture, 306(1), 403-406.
Cunha-Neto, E., Bilate, A. M., Hyland, K. V., Fonseca, S. G., Kalil, J., Engman, D. M., & Kalil, J.
(2006). Induction of cardiac autoimmunity in Chagas heart disease: a case for molecular
mimicry. Autoimmunity, 39(1), 41-54.
Cunha¬Neto, E., & Chevillard, C. (2014). Chagas disease cardiomyopathy: immunopathology and
genetics. Mediators of inflammation, 2014(1), 1¬11.
Da Rocha, C. Q., Queiroz, E. F., Meira, C. S., Moreira, D. R. M., Soares, M. B. P., Marcourt, L., &
Wolfender, J. L. (2014). Dimeric flavonoids from Arrabidaea brachypoda and assessment of their
anti-Trypanosoma cruzi activity. Journal of natural products, 77(6), 1345-1350.
Da Silva, T. B., Menezes, L. R., Sampaio, M. F., Meira, C. S., Guimarães, E. T., Soares, M. B., &
Costa, E. V. (2013). Chemical composition and anti-Trypanosoma cruzi activity of essential oils
obtained from leaves of Xylopia frutescens and X. laevigata (Annonaceae). Natural product
communications, 8(3), 403-406.

Referencias
72
Dancey, J. E., & Chen, H. X. (2006). Strategies for optimizing combinations of molecularly targeted
anticancer agents. Nature reviews Drug discovery, 5(8), 649-659.
Das, M., Mukherjee, S. B., & Shaha, C. (2001). Hydrogen peroxide induces apoptosis-like death in
Leishmania donovani promastigotes. Journal of Cell Science, 114(13), 2461-2469.
Das, R., Roy, A., Dutta, N., & Majumder, H. K. (2008). Reactive oxygen species and imbalance of
calcium homeostasis contributes to curcumin induced programmed cell death in Leishmania
donovani. Apoptosis, 13(7), 867-882.
Davila, D. F. (2004). Sympathetic nervous system activation in chagasic patients with congestive heart
failure. Journal of the American College of Cardiology, 43(9), 1723-1724.
Dávila, D. F., Núñez, T. J., Odreman, R., & de Dávila, C. A. M. (2005). Mechanisms of
neurohormonal activation in chronic congestive heart failure: pathophysiology and therapeutic
implications. International journal of cardiology, 101(3), 343-346.
De Barros Moreira Beltrão, H., de Paula Cerroni, M., de Freitas, D. R. C., das Neves Pinto, A. Y., da
Costa Valente, V., Valente, S. A., & Sobel, J. (2009). Investigation of two outbreaks of suspected oral
transmission of acute Chagas disease in the Amazon region, Para State, Brazil, in 2007. Tropical
doctor, 39(4), 231-232.
De Figueiredo Diniz, L., Urbina, J. A., de Andrade, I. M., Mazzeti, A. L., Martins, T. A. F., Caldas, I.
S., & Bahia, M. T. (2013). Benznidazole and posaconazole in experimental Chagas disease: positive
interaction in concomitant and sequential treatments. PLoS Negl Trop Dis, 7(8), e2367.
De Noya, B. A., Díaz-Bello, Z., Colmenares, C., Ruiz-Guevara, R., Mauriello, L., Zavala-Jaspe, R.,
& Rivas, L. (2010). Large urban outbreak of orally acquired acute Chagas disease at a school in
Caracas, Venezuela. Journal of Infectious Diseases, 201(9), 1308-1315.
De Paula Porto, M., Da Silva, G. N., Luperini, B. C. O., Bachiega, T. F., de Castro Marcondes, J. P.,
Sforcin, J. M., & Salvadori, D. M. F. (2014). Citral and eugenol modulate DNA damage and pro-
inflammatory mediator genes in murine peritoneal macrophages. Molecular biology reports, 41(11),
7043-7051.
De Rissio, A. M., Riarte, A. R., García, M. M., Esteva, M. I., Quaglino, M., & Ruiz, A. M. (2010).
Congenital Trypanosoma cruzi infection. Efficacy of its monitoring in an urban reference health
center in a non-endemic area of Argentina. The American journal of tropical medicine and
hygiene, 82(5), 838-845.
Di Stasi, L. C., Oliveira, G. P., Carvalhaes, M. A., Queiroz-Junior, M., Tien, O. S., Kakinami, S. H., &
Reis, M. S. (2002). Medicinal plants popularly used in the Brazilian Tropical Atlantic
Forest. Fitoterapia, 73(1), 69-91.
Dias, J. C. P. (2015). Evolution of Chagas disease screening programs and control programs: historical
perspective. Global Heart, 10(3), 193-202.
Díaz, M. L., & González, C. I. (2014). Enfermedad de Chagas agudo: transmisión oral de
Trypanosoma cruzi como una vía de transmisión re-emergente. Revista de la Universidad Industrial
de Santander. Salud, 46(2), 177-188.
Díaz, M. L., Leal, S., Mantilla, J. C., Molina-Berríos, A., López-Muñoz, R., Solari, A., & Rugeles, C.
I. G. (2015). Acute chagas outbreaks: molecular and biological features of Trypanosoma cruzi
isolates, and clinical aspects of acute cases in Santander, Colombia. Parasites & vectors, 8(1), 608.
Docampo, R. (1990). Sensitivity of parasites to free radical damage by antiparasitic drugs. Chemico-
biological interactions, 73(1), 1-27.

Referencias
73
Dos Santos, R. R., & Hudson, L. (1980). Trypanosoma cruzi: immunological consequences of parasite
modification of host cells. Clinical and experimental immunology, 40(1), 36.
Duszenko, M., Figarella, K., Macleod, E. T., & Welburn, S. C. (2006). Death of a trypanosome: a
selfish altruism. Trends in parásitology, 22(11), 536-542.
Ehlert, P. A. D., Ming, L. C., Marques, M. O. M., Fenandes, D. M., Rocha, W. A., Luz, J. M. Q., &
Silva, R. F. (2013). Influência do horário de colheita sobre o rendimento e composição do óleo
essencial de erva-cidreira brasileira [Lippia alba (Mill.) NE Br.]. Revista Brasileira de Plantas
Medicinais, 72-77.
Engman, D. M., & Leon, J. S. (2002). Pathogenesis of Chagas heart disease: role of
autoimmunity. Acta tropica, 81(2), 123-132.
Escobar, P., Milena Leal, S., Herrera, L. V., Martinez, J. R., & Stashenko, E. (2010). Chemical
composition and antiprotozoal activities of Colombian Lippia spp essential oils and their major
components. Memórias do Instituto Oswaldo Cruz, 105(2), 184-190.
Esteban, L., Montes, J. M., & Angulo, V. M. (2016). Diversidad de Triatominae (Hemiptera:
Reduviidae) en Santander, Colombia: implicaciones epidemiológicas. Biomédica, 37(1).
Fabri, R. L., Nogueira, M. S., Moreira, J. D. R., Bouzada, M. L. M., & Scio, E. (2011). Identification
of antioxidant and antimicrobial compounds of Lippia species by bioautography. Journal of medicinal
food, 14(7-8), 840-846.
Filardi, L. S., & Brener, Z. (1987). Susceptibility and natural resistance of Trypanosoma cruzi strains
to drugs used clinically in Chagas disease. Transactions of the Royal Society of Tropical Medicine and
Hygiene, 81(5), 755-759.
Fivelman, Q. L., Adagu, I. S., & Warhurst, D. C. (2004). Modified fixed-ratio isobologram method for
studying in vitro interactions between atovaquone and proguanil or dihydroartemisinin against drug-
resistant strains of Plasmodium falciparum. Antimicrobial agents and chemotherapy, 48(11), 4097-
4102.
Fujinami, R. S., Von Herrath, M. G., Christen, U., & Whitton, J. L. (2006). Molecular mimicry,
bystander activation, or viral persistence: infections and autoimmune disease. Clinical microbiology
reviews, 19(1), 80-94.
Galuppo, S., Bacigalupo, A., García, A., Ortiz, S., Coronado, X., Cattan, P. E., & Solari, A. (2009).
Predominance of Trypanosoma cruzi genotypes in two reservoirs infected by sylvatic Triatoma
infestans of an endemic area of Chile. Acta tropica, 111(1), 90-93.
Gannavaram, S., Vedvyas, C., & Debrabant, A. (2008). Conservation of the pro-apoptotic nuclease
activity of endonuclease G in unicellular trypanosomatid parasites. Journal of cell science, 121(1), 99-
109.
García, L. T., Leal, A. F., Moreno, É. M., Stashenko, E. E., & Arteaga, H. J. (2017). Differential anti-
proliferative effect on K562 leukemia cells of Lippia alba (Verbenaceae) essential oils produced under
diverse growing, collection and extraction conditions. Industrial Crops and Products, 96, 140-148.
García, L. T., Leite, N. R., Alfonzo, J. D., & Thiemann, O. H. (2007). Effects of Trypanosoma brucei
tryptophanyl-tRNA synthetases silencing by RNA interference. Memórias do Instituto Oswaldo
Cruz, 102(6), 757-762.
Gascon, J., Bern, C., & Pinazo, M. J. (2010). Chagas disease in Spain, the United States and other
non-endemic countries. Acta tropica, 115(1), 22-27.

Referencias
74
Geromini, K. V. N., Roratto, F. B., Ferreira, F. G., Camilotti, J., Vidigal, T. M. A., Valle, J. S., &
Linde, G. A. (2015). Fungicidal effect of Lippia alba essential oil on a white-rot fungus. Maderas.
Ciencia y tecnología, 17(1), 29-38.
Gilling, D. H., Kitajima, M., Torrey, J. R., & Bright, K. R. (2014). Antiviral efficacy and mechanisms
of action of oregano essential oil and its primary component carvacrol against murine
norovirus. Journal of applied microbiology, 116(5), 1149-1163.
Gómez, L. A., Stashenko, E., & Ocazionez, R. E. (2013). Comparative study on in vitro activities of
citral, limonene and essential oils from Lippia citriodora and L. alba on yellow fever virus. Natural
product communications, 8(2), 249-252.
Gorla, D., & Noireau, F. (2010). Geographic distribution of Triatominae vectors in
America. American trypanosomiasis (Chagas disease). One hundred years of research. 1st edition.
Burlington (VA): Elsevier Inc, 209-31.
Graebin, CS, Madeira, MDF, Yokoyama-Yasunaka, JK, Miguel, DC, Uliana, SR, Benítez, D., y
Eifler-Lima, VL (2010). Síntesis y actividad in vitro de los derivados de limoneno contra Leishmania
y Trypanosoma. Revista Europea de la química medicinal, 45(4), 1524-1528.
Grasl-Kraupp, B., Ruttkay-Nedecky, B., Koudelka, H., Bukowska, K., Bursch, W., & Schulte-
Hermann, R. (1995). In situ detection of fragmented DNA (TUNEL assay) fails to discriminate among
apoptosis, necrosis, and autolytic cell death: a cautionary note. Hepatology, 21(5), 1465-1468.
Grecco, S. S., Reimão, J. Q., Tempone, A. G., Sartorelli, P., Romoff, P., Ferreira, M. J., & Lago, J. H.
(2010). Isolation of an antileishmanial and antitrypanosomal flavanone from the leaves of Baccharis
retusa DC (Asteraceae). Parásitology research, 106(5), 1245-1248.
Guhl, F. (2007). Chagas disease in Andean countries. Memórias do Instituto Oswaldo Cruz, 102, 29-
38.
Gurtler, R. E., Segura, E. L., & Cohen, J. E. (2003). Congenital transmission of Trypanosoma cruzi
infection in Argentina. Emerging infectious diseases, 9(1), 29-32.
Guzmán, S., Cardozo, R., & García, V. (2004). Desarrollo agrotecnológico de Lippia alba (Miller) NE
Br ex Britton y Wilson. Guillermo de Ockhman, 7, 201-215.
Haldar, S., Kar, B., Dolai, N., Kumar, R. S., Behera, B., & Haldar, P. K. (2012). In vivo anti–
nociceptive and anti–inflammatory activities of Lippia alba. Asian Pacific Journal of Tropical
Disease, 2, S667-S670.
Hall, B. S., & Wilkinson, S. R. (2012). Activation of benznidazole by trypanosomal type I
nitroreductases results in glyoxal formation. Antimicrobial agents and chemotherapy, 56(1), 115-123.
Hennebelle, T., Sahpaz, S., Dermont, C., Joseph, H., & Bailleul, F. (2006). The essential oil of Lippia
alba: analysis of samples from French overseas departments and review of previous works. Chemistry
& biodiversity, 3(10), 1116-1125.
Hennebelle, T., Sahpaz, S., Joseph, H., & Bailleul, F. (2008). Ethnopharmacology of Lippia
alba. Journal of ethnopharmacology, 116(2), 211-222.
Herrera, L. (2010). Una revisión sobre reservorios de Trypanosoma (Schizotrypanum) cruzi (Chagas,
1909), agente etiológico de la Enfermedad de Chagas. Bol Mal Salud Amb, 50(1), 3-15.
Hoet, S., Pieters, L., Muccioli, G. G., Habib-Jiwan, J. L., Opperdoes, F. R., & Quetin-Leclercq, J.
(2007). Antitrypanosomal activity of triterpenoids and sterols from the leaves of Strychnos spinosa
and related compounds. Journal of natural products, 70(8), 1360-1363.

Referencias
75
Hossain, M. U., Oany, A. R., Ahmad, S. A. I., Hasan, M. A., Khan, M. A., & Siddikey, M. A. A.
(2016). Identification of potential inhibitor and enzyme-inhibitor complex on trypanothione reductase
to control Chagas disease. Computational Biology and Chemistry, 65, 29-36.
Hotez, P. J., Bottazzi, M. E., Franco-Paredes, C., Ault, S. K., & Periago, M. R. (2008). The neglected
tropical diseases of Latin America and the Caribbean: a review of disease burden and distribution and
a roadmap for control and elimination. PLoS Negl Trop Dis, 2(9), e300.
Hotez, P. J., Dumonteil, E., Heffernan, M. J., & Bottazzi, M. E. (2013). Innovation for the ‘bottom
100 million’: eliminating neglected tropical diseases in the Americas. In Hot Topics in Infection and
Immunity in Children IX (pp. 1-12). Springer New York.
Hyland, K. V., Leon, J. S., Daniels, M. D., Giafis, N., Woods, L. M., Bahk, T. J., & Engman, D. M.
(2007). Modulation of autoimmunity by treatment of an infectious disease. Infection and
immunity, 75(7), 3641-3650.
Idu, M., & Osemwegie, O. O. (2007). Some medicinal flora of okomu forest reserve in Southern
Nigeria. Research Journal of Medicinal Plant, 1(1), 29-31.
Instituto Nacional de Salud. (2012). Subdirección de vigilancia y control en salud pública. Protocolo
de vigilancia y control de Chagas.
Jackson, Y., Pinto, A., & Pett, S. (2014). Chagas disease in Australia and New Zealand: risks and
needs for public health interventions. Tropical Medicine & International Health, 19(2), 212-218.
Jafri, H., Husain, F. M., & Ahmad, I. (2014). Antibacterial and antibiofilm activity of some essential
oils and compounds against clinical strains of Staphylococcus aureus. Journal of Biomedical and
Therapeutic Sciences, 1(1), 65-71.
Jercic, M. I., Mercado, R., & Villarroel, R. (2010). Congenital Trypanosoma cruzi infection in
neonates and infants from two regions of Chile where Chagas' disease is endemic. Journal of clinical
microbiology, 48(10), 3824-3826.
Ji, J., Zhang, L., Wu, Y. Y., Zhu, X. Y., Lv, S. Q., & Sun, X. Z. (2006). Induction of apoptosis by d-
limonene is mediated by a caspase-dependent mitochondrial death pathway in human leukemia
cells. Leukemia & lymphoma, 47(12), 2617-2624.
Jia, S. S., Xi, G. P., Zhang, M., Chen, Y. B., Lei, B., Dong, X. S., & Yang, Y. M. (2013). Induction of
apoptosis by D-limonene is mediated by inactivation of Akt in LS174T human colon cancer
cells. Oncology reports, 29(1), 349-354.
Jimenez, V., Paredes, R., Sosa, M. A., & Galanti, N. (2008). Natural programmed cell death in T.
cruzi epimastigotes maintained in axenic cultures. Journal of cellular biochemistry, 105(3), 688-698.
Kapoor, S. (2013). D-Limonene: An emerging antineoplastic agent. Human and Experimental
Toxicology, 32(11), 1228.
Kapur, A., Felder, M., Fass, L., Kaur, J., Czarnecki, A., Rathi, K., & Patankar, M. (2016). The
monoterpene, citral, increases intracellular oxygen radicals and inhibits cancer cell proliferation by
inducing apoptosis and endoplasmic reticulum stress. Clinical cancer research, 615(22) 19106-4404.
Katsukawa, M., Nakata, R., Takizawa, Y., Hori, K., Takahashi, S., & Inoue, H. (2010). Citral, a
component of lemongrass oil, activates PPARα and γ and suppresses COX-2 expression. Biochimica
et Biophysica Acta (BBA)-Molecular and Cell Biology of Lipids, 1801(11), 1214-1220.
Keenan, M., & Chaplin, J. H. (2015). Chapter Four-A New Era for Chagas Disease Drug
Discovery?. Progress in medicinal chemistry, 54, 185-230.
Kessler, R. L., Contreras, V. T., Marliére, N. P., Guarneri, A. A., Villamizar Silva, L. H., Mazzarotto,
G. A. C. A., Batista, M., Soccol, V. T., Krieger, M. A., & Probst, C. M. (2017). Recently

Referencias
76
differentiated epimastigotes from Trypanosoma cruzi are infective to the mammalian host. Molecular
Microbiology.
Köberle, F. (1970). The causation and importance of nervous lesions in American
trypanosomiasis. Bulletin of the World Health Organization, 42(5), 739.
Kong, D. X., Li, X. J., & Zhang, H. Y. (2008). Convergent evolution of
medicines. ChemMedChem, 3(8), 1169-1171.
Kpoviessi, S., Bero, J., Agbani, P., Gbaguidi, F., Kpadonou-Kpoviessi, B., Sinsin, B., & Quetin-
Leclercq, J. (2014). Chemical composition, cytotoxicity and in vitro antitrypanosomal and
antiplasmodial activity of the essential oils of four Cymbopogon species from Benin. Journal of
ethnopharmacology, 151(1), 652-659.
Kransdorf, E. P., Zakowski, P. C., & Kobashigawa, J. A. (2014). Chagas disease in solid organ and
heart transplantation. Current opinion in infectious diseases, 27(5), 418-424.
Ku, C. M., & Lin, J. Y. (2013). Anti-inflammatory effects of 27 selected terpenoid compounds tested
through modulating Th1/Th2 cytokine secretion profiles using murine primary splenocytes. Food
chemistry, 141(2), 1104-1113.
Leal, S. M., Pino, N., Stashenko, E. E., Martínez, J. R., & Escobar, P. (2013). Antiprotozoal activity of
essential oils derived from Piper spp. grown in Colombia. Journal of Essential Oil Research, 25(6),
512-519.
Lecane, P. S., Karaman, M. W., Sirisawad, M., Naumovski, L., Miller, R. A., Hacia, J. G., & Magda,
D. (2005). Motexafin gadolinium and zinc induce oxidative stress responses and apoptosis in B-cell
lymphoma lines. Cancer research, 65(24), 11676-11688.
Lee, B. Y., Bacon, K. M., Bottazzi, M. E., & Hotez, P. J. (2013). Global economic burden of Chagas
disease: a computational simulation model. The Lancet infectious diseases, 13(4), 342-348.
Lee, H. J., Jeong, H. S., Kim, D. J., Noh, Y. H., Yuk, D. Y., & Hong, J. T. (2008). Inhibitory effect of
citral on NO production by suppression of iNOS expression and NF-κB activation in RAW264. 7
cells. Archives of pharmacal research, 31(3), 342-349.
Liao, P. C., Yang, T. S., Chou, J. C., Chen, J., Lee, S. C., Kuo, Y. H., Ho, L. C & Chao, L. K. P.
(2015). Anti-inflammatory activity of neral and geranial isolated from fruits of Litsea cubeba
Lour. Journal of Functional Foods, 19, 248-258.
López, M. A., Stashenko, E. E., & Fuentes, J. L. (2011). Chemical composition and antigenotoxic
properties of Lippia alba essential oils. Genetics and molecular biology, 34(3), 479-488.
Luna-Marín, K. P., Jaramillo-Londoño, C. L., Hernández-Torres, J., Gutiérrez-Marín, R., Vallejo, G.
A., & Angulo-Silva, V. M. (2009). ITS–RFLP-and RAPD-based genetic variability of Trypanosoma
cruzi I, human and vector strains in Santander (Colombia). Parásitology research, 105(2), 519.
Mamun-Or-Rashid, A. N. M., Sem, M. K., Jamal, M. A. H. M., & Nasrin, S. (2013). A comprehensive
ethno-pharmacological review on Lippia alba M. International Journal of Biomedical Materials
Research, 1(1), 14-20.
Mandal, G., Orta, J. F., Sharma, M., & Mukhopadhyay, R. (2013). Trypanosomatid aquaporins: roles
in physiology and drug response. Diseases, 2(1), 3-23.
Manrique-Abril, F., Ospina, J. M., Herrera, G., Florez, A. C., Pavia, P. X., Montilla, M & Bula, C. P.
(2013). Diagnóstico de enfermedad de Chagas en mujeres embarazadas y recién nacidos de Moniquirá
y Miraflores, Boyacá, Colombia. Infectio, 17(1), 28-34.

Referencias
77
Mantilla, J. C., Suárez, E. U., & Barraza, M. F. (2011). Enfermedad de Chagas: correlación clínico-
patológica. Serie de casos del Hospital Universitario de Santander-Departamento de Patología,
Universidad Industrial de Santander. Revista Colombiana de Cardiología, 18(5), 249-261.
Manuele, M. G., Barreiro Arcos, M. L., Davicino, R., Ferraro, G., Cremaschi, G., & Anesini, C.
(2009). Limonene exerts antiproliferative effects and increases nitric oxide levels on a lymphoma cell
line by dual mechanism of the ERK pathway: relationship with oxidative stress. Cancer
investigation, 28(2), 135-145.
Martínez-Díaz, R. A., Ibáñez-Escribano, A., Burillo, J., Heras, L. de las, del Prado, G., Agulló-Ortuño,
M. T., González-Coloma, A. (2015). Trypanocidal, trichomonacidal and cytotoxic components of
cultivated Artemisia absinthiumLinnaeus (Asteraceae) essential oil. Memórias Do Instituto Oswaldo
Cruz, 110(5), 693–699.
Martínez-Parra, A. G., Pinilla-Alfonso, M. Y., & Estrada-Montoya, J. H. (2015). Enfermedad de
Chagas: de las tendencias epidemiológicas rurales a su urbanización en países desarrollados. Revisión
de la literatura. Acta Odontológica Colombiana, 5(2), 85.
Maya, J. D., Cassels, B. K., Iturriaga-Vásquez, P., Ferreira, J., Faúndez, M., Galanti, N., & Morello,
A. (2007). Mode of action of natural and synthetic drugs against Trypanosoma cruzi and their
interaction with the mammalian host. Comparative Biochemistry and Physiology Part A: Molecular &
Integrative Physiology, 146(4), 601-620.
Maya, J. D., Rodriguez, A., Pino, L., Pabon, A., Ferreira, J., Pavani, M., & Morello, A. (2004). Effects
of buthionine sulfoximine nifurtimox and benznidazole upon trypanothione and metallothionein
proteins in Trypanosoma cruzi. Biological research, 37(1), 61-69.
Mecca, M. M. D., Bartel, L. C., Castro, C. R. D., & Castro, J. A. (2008). Benznidazole
biotransformation in rat heart microsomal fraction without observable ultrastructural alterations:
comparison to Nifurtimox-induced cardiac effects. Memorias do Instituto Oswaldo Cruz, 103(6), 549-
553.
Mecca, M. M., Fanelli, S. L., Bartel, L. C., de Castro, C. R., Díaz, E. G., & Castro, J. A. (2007).
Nifurtimox nitroreductase activity in different cellular fractions from male rat pancreas. Biochemical
and ultrastructural alterations. Life sciences, 81(2), 144-152.
Menna-Barreto, R. F., Salomão, K., Dantas, A. P., Santa-Rita, R. M., Soares, M. J., Barbosa, H. S., &
de Castro, S. L. (2009). Different cell death pathways induced by drugs in Trypanosoma cruzi: an
ultrastructural study. Micron, 40(2), 157-168
Mesa-Arango, A. C., Montiel-Ramos, J., Zapata, B., Durán, C., Betancur-Galvis, L., & Stashenko, E.
(2009). Citral and carvone chemotypes from the essential oils of Colombian Lippia alba (Mill.) NE
Brown: composition, cytotoxicity and antifungal activity. Memórias do Instituto Oswaldo
Cruz, 104(6), 878-884.
Miller, J. A., Lang, J. E., Ley, M., Nagle, R., Hsu, C. H., Thompson, P. A., & Chow, H. S. (2013).
Human breast tissue disposition and bioactivity of limonene in women with early-stage breast
cancer. Cancer Prevention Research, 6(6), 577-584.
Minneman, R. M., Hennink, M. M., Nicholls, A., Salek, S. S., Palomeque, F. S., Khawja, A., Albor,
L.C., Pennock, C.C., & Leon, J. S. (2012). Barriers to testing and treatment for Chagas disease among
Latino immigrants in Georgia. Journal of parásitology research, 2012.
Molina, I., Gómez i Prat, J., Salvador, F., Treviño, B., Sulleiro, E., Serre, N., & Blanco-Grau, A.
(2014). Randomized trial of posaconazole and benznidazole for chronic Chagas' disease. New England
Journal of Medicine, 370(20), 1899-1908.
Morens, D. M., Folkers, G. K., & Fauci, A. S. (2004). The challenge of emerging and re-emerging
infectious diseases. Nature, 430(6996), 242-249.

Referencias
78
Moretti, N. S., da Silva Augusto, L., Clemente, T. M., Antunes, R. P. P., Yoshida, N., Torrecilhas, A.
C., & Schenkman, S. (2015). Characterization of Trypanosoma cruzi sirtuins as possible drug targets
for Chagas disease. Antimicrobial agents and chemotherapy, 59(8), 4669-4679.
Morillo, C. A., Marin-Neto, J. A., Avezum, A., Sosa-Estani, S., Rassi Jr, A., Rosas, F., & Guhl, F
(2015). Randomized trial of benznidazole for chronic Chagas’ cardiomyopathy. New England Journal
of Medicine, 373(14), 1295-1306.
Murbach Teles Andrade, B. F., Nunes Barbosa, L., da Silva Probst, I., & Fernandes Júnior, A. (2014).
Antimicrobial activity of essential oils. Journal of Essential Oil Research, 26(1), 34-40.
Murcia, L., Carrilero, B., Munoz-Davila, M. J., Thomas, M. C., López, M. C., & Segovia, M. (2013).
Risk factors and primary prevention of congenital Chagas disease in a nonendemic country. Clinical
Infectious Diseases, 56(4), 496-502.
Murray, C. J., Barber, R. M., Foreman, K. J., Ozgoren, A. A., Abd-Allah, F., Abera, S. F., & Abu-
Rmeileh, N. M. (2015). Global, regional, and national disability-adjusted life years (DALYs) for 306
diseases and injuries and healthy life expectancy (HALE) for 188 countries, 1990–2013: quantifying
the epidemiological transition. The Lancet, 386(10009), 2145-2191.
Nagajyothi, F., Machado, F. S., Burleigh, B. A., Jelicks, L. A., Scherer, P. E., Mukherjee, S., Lisanti,
M. P., Weiss, L. M., Garg, N. J., & Tanowitz, H. B. (2012). Mechanisms of Trypanosoma cruzi
persistence in Chagas disease. Cellular microbiology, 14(5), 634¬643.
Nibret, E., & Wink, M. (2010). Volatile components of four Ethiopian Artemisia species extracts and
their in vitro antitrypanosomal and cytotoxic activities. Phytomedicine, 17(5), 369-374.
Noireau, F. (2009). Wild Triatoma infestans, a potential threat that needs to be monitored. Memórias
do Instituto Oswaldo Cruz, 104, 60-64.
Nuñez, M. B., Torres, C. A., Aguado, M. I., Bela, A. J., Dudik, H. N., & Bregni, C. (2012).
Polyphenols and antimicrobial activity in extracts of Lippia alba (Mill.). International Journal of
Medicinal and Aromatic Plants, 2(3), 361-368.
Okpekon, T., Yolou, S., Gleye, C., Roblot, F., Loiseau, P., Bories, C., & Hocquemiller, R. (2004).
Antiparasitic activities of medicinal plants used in Ivory Coast. Journal of ethnopharmacology, 90(1),
91-97.
Oliveira, D. R., Leitao, G. G., Santos, S. S., Bizzo, H. R., Lopes, D., Alviano, C. S., & Leitao, S. G.
(2006). Ethnopharmacological study of two Lippia species from Oriximiná, Brazil. Journal of
Ethnopharmacology, 108(1), 103-108.
Oliveira, G. B. F., Avezum, Á., & Mattos, A. J. C. (2015). Perspectives in Chagas disease treatment.
Global Heart, 10, 189-192.
Oliveira, I., Torrico, F., Muñoz, J., & Gascon, J. (2010). Congenital transmission of Chagas disease: a
clinical approach. Expert review of anti-infective therapy, 8(8), 945-956.
Olivero-Verbel, J., Barreto-Maya, A., Bertel-Sevilla, A., & Stashenko, E. E. (2014). Composition,
anti-quorum sensing and antimicrobial activity of essential oils from Lippia alba. Brazilian Journal of
Microbiology, 45(3), 759-767.
Organización Mundial de la Salud. (2006). Estimación cuantitativa de la enfermedad de Chagas en las
Americas.
Organización Mundial de la Salud. (2015). Investing to Overcome the Global Impact of Neglected
Tropical Diseases: Third WHO Report on Neglected Tropical Diseases
Organización Mundial de la Salud. (2015). Chagas disease in Latin America: an epidemiological
update based on 2010 estimates.

Referencias
79
Palacio, K. A., & Rodríguez, N. (2008). Plasticidad fenotípica en Lippia alba (Verbenaceae) en
respuesta a la disponibilidad hídrica en dos ambientes lumínicos. Acta Biológica Colombiana, 13(1),
187.
Park, K. R., Nam, D., Yun, H. M., Lee, S. G., Jang, H. J., Sethi, G., & Ahn, K. S. (2011). β-
Caryophyllene oxide inhibits growth and induces apoptosis through the suppression of
PI3K/AKT/mTOR/S6K1 pathways and ROS-mediated MAPKs activation. Cancer letters, 312(2),
178-188.
Parra-Garcés, M. I., Caroprese-Araque, J. F., Arrieta-Prieto, D., & Stashenko, E. (2010). Morphology,
anatomy, ontogeny and chemical composition of inflorescences volatile secondary metabolites of
Lippia alba (Verbenaceae) at three stages of development. Revista de biologia tropical, 58(4), 1533-
1548.
Paulino, M., Iribarne, F., Dubin, M., Aguilera-Morales, S., Tapia, O., & Stoppani, A. O. M. (2005).
The chemotherapy of Chagas' disease: an overview. Mini reviews in medicinal chemistry, 5(5), 499-
519.
Pendergrass, W., Wolf, N., & Poot, M. (2004). Efficacy of MitoTracker Green™ and CMXrosamine
to measure changes in mitochondrial membrane potentials in living cells and tissues. Cytometry Part
A, 61(2), 162-169.
Pérez-Galán, P., Roué, G., Villamor, N., Montserrat, E., Campo, E., & Colomer, D. (2006). The
proteasome inhibitor bortezomib induces apoptosis in mantle-cell lymphoma through generation of
ROS and Noxa activation independent of p53 status. Blood, 107(1), 257-264.
Pinazo, M. J., Guerrero, L., Posada, E., Rodríguez, E., Soy, D., & Gascon, J. (2013). Benznidazole-
related adverse drug reactions and their relationship to serum drug concentrations in patients with
chronic chagas disease. Antimicrobial agents and chemotherapy, 57(1), 390-395.
Pinto, E. D. P. P., Amorozo, M. C. D. M., & Furlan, A. (2006). Conhecimento popular sobre plantas
medicinais em comunidades rurais de mata atlântica–Itacaré, BA, Brasil. Acta Botanica
Brasilica, 20(4), 751-762.
Polanco-Hernández, G., Escalante-Erosa, F., García-Sosa, K., Rosado, M. E., Guzmán-Marín, E.,
Acosta-Viana, K. Y., & Peña-Rodríguez, L. M. (2013). Synergistic effect of lupenone and
caryophyllene oxide against Trypanosoma cruzi. Evidence-Based Complementary and Alternative
Medicine, 2013.
Post-White, J., & Nichols, W. (2007). Randomized trial testing QueaseEase TM essential oil for
motion sickness. Int J Essent Oil Ther, 1(4), 158-166.
Prata, A. (2001). Clinical and epidemiological aspects of Chagas disease. The Lancet infectious
diseases, 1(2), 92-100.
Rabi, T., & Bishayee, A. (2009). Terpenoids and breast cancer chemoprevention. Breast cancer
research and treatment, 115(2), 223-239..
Rajao, M. A., Furtado, C., Alves, C. L., Passos‐ Silva, D. G., Moura, M. B., Schamber‐ Reis, B. L., &
Mendes, I. C. (2014). Unveiling Benznidazole's mechanism of action through overexpression of DNA
repair proteins in Trypanosoma cruzi. Environmental and molecular mutagenesis, 55(4), 309-321.
Ramírez, C. J., Jaramillo, C. A., del Pilar Delgado, M., Pinto, N. A., Aguilera, G., & Guhl, F. (2005).
Genetic structure of sylvatic, peridomestic and domestic populations of Triatoma dimidiata
(Hemiptera: Reduviidae) from an endemic zone of Boyaca, Colombia. Acta tropica, 93(1), 23-29.
Ramos, E. H., Moraes, M. M., Nerys, L. L. D. A., Nascimento, S. C., Militão, G. C., de Figueiredo, R.
C., Da Camara, C. A. G., & Silva, T. G. (2014). Chemical composition, leishmanicidal and cytotoxic

Referencias
80
activities of the essential oils from Mangifera indica L. var. Rosa and Espada. BioMed research
international, 2014.
Rassi, A., & de Rezende, J. M. (2012). American trypanosomiasis (Chagas disease). Infectious disease
clinics of North America, 26(2), 275-291.
Rassi, A., & Marin-Neto, J. A. (2010). Chagas disease. The Lancet, 375(9723), 1388-1402.
Riarte, A., Luna, C., Sabatiello, R., Sinagra, A., Schiavelli, R., De Rissio, A., & Lauricella, M. (1999).
Chagas' disease in patients with kidney transplants: 7 years of experience, 1989–1996. Clinical
infectious diseases, 29(3), 561-567.
Rueda, K., Trujillo, J. E., Carranza, J. C., & Vallejo, G. A. (2014). Transmisión oral de Trypanosoma
cruzi: una nueva situación epidemiológica de la enfermedad de Chagas en Colombia y otros países
suramericanos. Biomédica, 34(4), 631-41.
Saddiq, A. A., & Khayyat, S. A. (2010). Chemical and antimicrobial studies of monoterpene:
Citral. Pesticide Biochemistry and Physiology, 98(1), 89-93.
Sanabria Calvo, M. (2016). Tripanosomiasis Americana o Enfermedad de Chagas. Revista Médica de
Costa Rica y Centroamérica, 72(616), 539-544.
Santoro, G. F., Cardoso, M. G., Guimarães, L. G. L., Freire, J. M., & Soares, M. J. (2007a). Anti-
proliferative effect of the essential oil of Cymbopogon citratus (DC) Stapf (lemongrass) on
intracellular amastigotes, bloodstream trypomastigotes and culture epimastigotes of Trypanosoma
cruzi (Protozoa: Kinetoplastida). Parásitology, 134(11), 1649-1656.
Santoro, G. F., Cardoso, M. G., Guimarães, L. G. L., Salgado, A. P. S., Menna-Barreto, R. F., &
Soares, M. J. (2007b). Effect of oregano (Origanum vulgare L.) and thyme (Thymus vulgaris L.)
essential oils on Trypanosoma cruzi (Protozoa: Kinetoplastida) growth and
ultrastructure. Parásitology research, 100(4), 783-790.
Santoro, G. F., Cardoso, M. G., Guimarães, L. G. L., Mendonça, L. Z., & Soares, M. J. (2007c).
Trypanosoma cruzi: activity of essential oils from Achillea millefolium L., Syzygium aromaticum L.
and Ocimum basilicum L. on epimastigotes and trypomastigotes. Experimental parásitology, 116(3),
283-290.
Santos, K. K., Matias, E. F., Tintino, S. R., Souza, C. E., Braga, M. F., Guedes, G. M., Rolón, M.,
Vega, C., de Arias, A. R., Costa, J. G., & Menezes, I. R. (2012). Anti-Trypanosoma cruzi and
cytotoxic activities of Eugenia uniflora L. Experimental Parásitology, 131(1), 130-132.
Santos, N. N., Menezes, L. R., Dos Santos, J. A., Meira, C. S., Guimarhes, E. T., Soares, M. B., &
Costa, E. V. (2014). A new source of (R)-limonene and rotundifolone from leaves of Lippia
pedunculosa (verbenaceae) and their trypanocidal properties. Natural product communications, 9(6),
737-739.
Santos, NN, Menezes, LR, Dos Santos, JA, Meira, CS, Guimarhes, ET, Soares, MB, y Costa, EV
(2014). Una nueva fuente de limoneno (R) y rotundifolone partir de hojas de Lippia pedunculosa
(Verbenaceae) y sus propiedades tripanocidas. Comunicaciones de productos Natural , 9 (6), 737-739.
Schelz, Z., Hohmann, J., & Molnar, J. (2010). Recent advances in research of antimicrobial effects of
essential oils and plant derived compounds on bacteria. Ethnomedicine: A Source of Complementary
Therapeutics, 6, 179-201.
Schijman, A. G. (2006). Congenital chagas disease. Perspectives in Medical Virology, 13, 223-258.
Schmidt, T. J., A Khalid, S., J Romanha, A., MA Alves, T., W Biavatti, M., Brun, R., Da Costa, F. B.,
de Castro, S.L., Ferreira, V. F., de Lacerda, M. V., Lago, J. H., León, L. L., Lopes, N. P., das Neves
Amorin, R. C., Niehues, M., Ogungbe, I. V., Pohlit, A. M., Scotti, M. T., Setzer, W. N., de N.C.

Referencias
81
Soerio, M., Steindel, M., Tempone, A. G. (2012). The potential of secondary metabolites from plants
as drugs or leads against protozoan neglected diseases-part II. Current medicinal chemistry, 19(14),
2176-2228.
Schmunis, G. A. (2007). The globalization of Chagas disease. ISBT Science Series, 2(1), 6-11.
Sena Filho, J. G., Melo, J. G., Saraiva, A. M., Gonçalves, A. M., Psiottano, M. N. C., & Xavier, H. S.
(2006). Antimicrobial activity and phytochemical profile from the roots of Lippia alba (Mill.) NE
Brown. Revista Brasileira de Farmacognosia, 16(4), 506-509.
Shikanai-Yasuda, M. A., & Carvalho, N. B. (2012). Oral transmission of Chagas disease. Clinical
Infectious Diseases, cir956.
Shukla, R., Kumar, A., Singh, P., & Dubey, N. K. (2009). Efficacy of Lippia alba (Mill.) NE Brown
essential oil and its monoterpene aldehyde constituents against fungi isolated from some edible
legume seeds and aflatoxin B 1 production. International Journal of Food Microbiology, 135(2), 165-
170.
Shyamapada, M. (2014). Epidemiological Aspects of Chagas Disease-a Review. Journal of Ancient
Diseases & Preventive Remedies.
Silva-Neto, M. A., Fampa, P., Caiaffa, C. D., Carneiro, A. B., & Atella, G. C. (2010). Cell signaling
during Trypanosoma cruzi development in triatominae. Open Parásitology Journal, 4, 188-194.
Soeiro, M. N. C., & de Castro, S. L. (2009). Trypanosoma cruzi targets for new chemotherapeutic
approaches. Expert opinion on therapeutic targets, 13(1), 105-121.
Soković, M., Glamočlija, J., Marin, P. D., Brkić, D., & van Griensven, L. J. (2010). Antibacterial
effects of the essential oils of commonly consumed medicinal herbs using an in vitro
model. Molecules, 15(11), 7532-7546.
Sosa-Estani, S., & Segura, E. L. (2015). Integrated control of Chagas disease for its elimination as
public health problem-A Review. Memórias do Instituto Oswaldo Cruz, 110(3), 289-298.
Stashenko, E. E. (2009). Aceites esenciales. Universidad Industrial de Santander.
Stashenko, E. E., Jaramillo, B. E., & Martınez, J. R. (2004). Comparison of different extraction
methods for the analysis of volatile secondary metabolites of Lippia alba (Mill.) NE Brown, grown in
Colombia, and evaluation of its in vitro antioxidant activity. Journal of Chromatography A, 1025(1),
93-103.
Stashenko, E. E., Martínez, J. R., Durán, D. C., Córdoba, Y., & Caballero, D. (2014). Estudio
comparativo de la composición química y la actividad antioxidante de los aceites esenciales de
algunas plantas del género Lippia (Verbenaceae) cultivadas en Colombia. Revista de la Academia
Colombiana de Ciencias Exactas, Físicas y Naturales, 38, 89-105.
Steindel, M., Pacheco, L. K., Scholl, D., Soares, M., de Moraes, M. H., Eger, I., & de Carvalho-Pinto,
C. J. (2008). Characterization of Trypanosoma cruzi isolated from humans, vectors, and animal
reservoirs following an outbreak of acute human Chagas disease in Santa Catarina State,
Brazil. Diagnostic microbiology and infectious disease, 60(1), 25-32.
Sülsen, V. P., Frank, F. M., Cazorla, S. I., Anesini, C. A., Malchiodi, E. L., Freixa, B., & Martino, V.
S. (2008). Trypanocidal and leishmanicidal activities of sesquiterpene lactones from Ambrosia
tenuifolia Sprengel (Asteraceae). Antimicrobial Agents and Chemotherapy, 52(7), 2415-2419.
Sun, J. (2007). D-Limonene: safety and clinical applications. Alternative Medicine Review, 12(3), 259.
Szatrowski, T. P., & Nathan, C. F. (1991). Production of large amounts of hydrogen peroxide by
human tumor cells. Cancer research, 51(3), 794-798.

Referencias
82
Teixeira, A. R., Hecht, M. M., Guimaro, M. C., Sousa, A. O., & Nitz, N. (2011). Pathogenesis of
chagas' disease: parasite persistence and autoimmunity. Clinical microbiology reviews, 24(3), 592-
630.
Teixeira, A. R., Nascimento, R. J., & Sturm, N. R. (2006). Evolution and pathology in Chagas disease:
a review. Memórias do Instituto Oswaldo Cruz, 101(5), 463-491.
Teixeira, D. E., Benchimol, M., Crepaldi, P. H., & de Souza, W. (2012). Interactive multimedia to
teach the life cycle of Trypanosoma cruzi, the causative agent of Chagas disease. PLoS Negl Trop
Dis, 6(8), e1749.
Teles, S., Pereira, J. A., Santos, C. H., Menezes, R. V., Malheiro, R., Lucchese, A. M., & Silva, F.
(2012). Geographical origin and drying methodology may affect the essential oil of Lippia alba (Mill)
NE Brown. Industrial Crops and Products, 37(1), 247-252.
Thomas, M. L., Coyle, K. M., Cruickshank, B., Giacomantonio, M., Wallace, M., Giacomantonio, C.,
& Marcato, P. (2016). Abstract B23: Citral reduces ALDH1A3 activity in breast cancer: Potential
applications in targeting breast cancer stem cells. Molecular Cancer Research, 14(2 Supplement),
B23.
Todorov, A. G., Andrade, D., Pesquero, J. B., Araujo, R. D. C., Bader, M., Stewart, J., & Neto, H. C.
F. (2003). Trypanosoma cruzi induces edematogenic responses in mice and invades cardiomyocytes
and endothelial cells in vitro by activating distinct kinin receptor (B1/B2) subtypes. The FASEB
Journal, 17(1), 73-75.
Tomazoni, E. Z., Pansera, M. R., Pauletti, G. F., Moura, S., Ribeiro, R. T., & Schwambach, J. (2016).
In vitro antifungal activity of four chemotypes of Lippia alba (Verbenaceae) essential oils against
Alternaria solani (Pleosporeaceae) isolates. Anais da Academia Brasileira de Ciências, (AHEAD), 0-
0.
Torres, C. M. (1960). Myocytolysis and fibrosis of the myocardium in Chagas' disease. Memorias do
Instituto Oswaldo Cruz, 58, 161.
Torrico, F., Alonso-Vega, C., Suarez, E., Rodriguez, P., Torrico, M. C., Dramaix, M., Truyens, C., &
Carlier, Y. (2004b). Maternal Trypanosoma cruzi infection, pregnancy outcome, morbidity, and
mortality of congenitally infected and non-infected newborns in Bolivia. The American journal of
tropical medicine and hygiene, 70(2), 201-209.
Torrico, F., Castro, M., Solano, M., Rodriguez, P., Torrico, M. C., Truyens, C., & Carlier, Y. (2004a).
Effects of maternal infection with Trypanosoma cruzi in pregnancy development and in the newborn
infant. Revista da Sociedade Brasileira de Medicina Tropical, 38, 73-76.
Torrico, M. C., Solano, M., Guzmán, J. M., Parrado, R., Suarez, E., Alonzo-Vega, C., & Torrico, F.
(2004c). Estimation of the parasitemia in Trypanosoma cruzi human infection: high parasitemias are
associated with severe and fatal congenital Chagas disease. Revista da Sociedade Brasileira de
Medicina Tropical, 38, 58-61.
Toscano Gonzalez, J. (2006). Uso tradicional de plantas medicinales en la vereda San Isidro,
municipio de San José de Pare-Boyacá: un estudio preliminar usando técnicas cuantitativas. Acta biol.
colomb, 11(2), 137-1146.
Trachootham, D., Zhou, Y., Zhang, H., Demizu, Y., Chen, Z., Pelicano, H., Chiao, P.J., Achanta, G.,
Arlinghaus, R.B., Liu, J., y Huang, P. (2006). Selective killing of oncogenically transformed cells
through a ROS-mediated mechanism by β-phenylethyl isothiocyanate. Cancer cell, 10(3), 241-252.
Trochine, A., Creek, D. J., Faral-Tello, P., Barrett, M. P., & Robello, C. (2014). Benznidazole
biotransformation and multiple targets in Trypanosoma cruzi revealed by metabolomics. PLoS Negl
Trop Dis, 8(5), e2844.

Referencias
83
Urbina, J. A. (2009). Ergosterol biosynthesis and drug development for Chagas disease. Memorias do
Instituto Oswaldo Cruz, 104, 311-318.
Urbina, J. A. (2010). Specific chemotherapy of Chagas disease: relevance, current limitations and new
approaches. Acta tropica, 115(1), 55-68.
Urbina, J. A. (2015). Recent clinical trials for the etiological treatment of chronic Chagas disease:
advances, challenges and perspectives. Journal of Eukaryotic Microbiology, 62(1), 149-156.
Van Der Logt, E. M., Roelofs, H. M., Van Lieshout, E. M., Nagengast, F. M., & Peters, W. H. (2004).
Effects of dietary anticarcinogens and nonsteroidal anti-inflammatory drugs on rat gastrointestinal
UDP-glucuronosyltransferases. Anticancer research, 24(2B), 843-850. Varela, M. T., Dias, R. Z., Martins, L. F., Ferreira, D. D., Tempone, A. G., Ueno, A. K., & Fernandes,
J. P. S. (2016). Gibbilimbol analogues as antiparasitic agents—Synthesis and biological activity
against Trypanosoma cruzi and Leishmania (L.) infantum. Bioorganic & medicinal chemistry
letters, 26(4), 1180-1183.
Venturi, C. R., Danielli, L. J., Klein, F., Apel, M. A., Montanha, J. A., Bordignon, S. A., Roehe, P.
M., Fuentefria, A. M., & Henriques, A. T. (2015). Chemical analysis and in vitro antiviral and
antifungal activities of essential oils from Glechon spathulata and Glechon marifolia. Pharmaceutical
biology, 53(5), 682-688.
Vera, S. S., Zambrano, D. F., Méndez-Sanchez, S. C., Rodríguez-Sanabria, F., Stashenko, E. E., &
Luna, J. E. D. (2014). Essential oils with insecticidal activity against larvae of Aedes aegypti (Diptera:
Culicidae). Parásitology research, 113(7), 2647-2654.
Vianna, G. O. (1911). On pathologic anatomy in Chagas disease. Mem. Inst. Oswaldo Cruz, 3, 276-
294.
Vigushin, D. M., Poon, G. K., Boddy, A., English, J., Halbert, G. W., Pagonis, C., & Coombes, R. C.
(1998). Phase I and pharmacokinetic study of D-limonene in patients with advanced cancer. Cancer
chemotherapy and pharmacology, 42(2), 111-117.
Viotti, R., Vigliano, C., Álvarez, M. G., Lococo, B., Petti, M., Bertocchi, G., & Laucella, S. (2011).
Impact of aetiological treatment on conventional and multiplex serology in chronic Chagas
disease. PLoS Negl Trop Dis, 5(9), e1314.
Viotti, R., Vigliano, C., Armenti, H., & Segura, E. (1994). Treatment of chronic Chagas' disease with
benznidazole: clinical and serologic evolution of patients with long-term follow-up. American heart
journal, 127(1), 151-162.
Vit, P., Silva, B., & Meléndez, P. (2002). Lippia alba NE Br. Ficha botánica de interés apícola en
Venezuela, No. 2 Cidrón. Rev Fac Farm Universidad de Los Andes, Mérida, Venezuela, 43.
Weiss, B., & Aksoy, S. (2011). Microbiome influences on insect host vector competence. Trends in
parásitology, 27(11), 514-522.
Wen, J. J., Vyatkina, G., & Garg, N. (2004). Oxidative damage during chagasic cardiomyopathy
development: role of mitochondrial oxidant release and inefficient antioxidant defense. Free Radical
Biology and Medicine, 37(11), 1821-1833.
Wilkinson, S. R., Obado, S. O., Mauricio, I. L., & Kelly, J. M. (2002). Trypanosoma cruzi expresses a
plant-like ascorbate-dependent hemoperoxidase localized to the endoplasmic reticulum. Proceedings
of the National Academy of Sciences, 99(21), 13453-13458.

Referencias
84
Wilkinson, S. R., Taylor, M. C., Horn, D., Kelly, J. M., & Cheeseman, I. (2008). A mechanism for
cross-resistance to nifurtimox and benznidazole in trypanosomes. Proceedings of the National
Academy of Sciences, 105(13), 5022-5027.
Wink, M. (2012). Medicinal plants: a source of anti-parasitic secondary
metabolites. Molecules, 17(11), 12771-12791.
Xia, H., Liang, W., Song, Q., Chen, X., Chen, X., & Hong, J. (2013). The in vitro study of apoptosis
in NB4 cell induced by citral. Cytotechnology, 65(1), 49-57.
Xiu-zhen, W. L. Z. (2009). Growth inhibition effects of D-limonene on Human Gastric Carcinoma
MGC803 cells [J]. Life Science Instruments, 1, 012.
Yang, D., Michel, L., Chaumont, J. P., & Millet-Clerc, J. (2000). Use of caryophyllene oxide as an
antifungal agent in an in vitro experimental model of onychomycosis. Mycopathologia, 148(2), 79-82.
Yeo, M., Acosta, N., Llewellyn, M., Sánchez, H., Adamson, S., Miles, G. A., & de Arias, A. R.
(2005). Origins of Chagas disease: Didelphis species are natural hosts of Trypanosoma cruzi I and
armadillos hosts of Trypanosoma cruzi II, including hybrids. International journal for
parásitology, 35(2), 225-233.
York, T., Van Vuuren, S. F., & De Wet, H. (2012). An antimicrobial evaluation of plants used for the
treatment of respiratory infections in rural Maputaland, KwaZulu-Natal, South Africa. Journal of
Ethnopharmacology, 144(1), 118-127.
Yu, Z., Wang, W., Xu, L., Dong, J., & Jing, Y. (2008). d-Limonene and d-carvone induce apoptosis in
HL-60 cells through activation of caspase-8. Asian J Trad Med, 3, 134-43.
Zheng, G. Q., Kenney, P. M., & Lam, L. K. (1992). Anethofuran, carvone, and limonene: potential
cancer chemoprotective agents from dill weed oil and caraway oil. Planta medica, 58(04), 338-341.
Zingales, B., Andrade, S. G., Briones, M. R. D. S., Campbell, D. A., Chiari, E., Fernandes, O., &
Miles, M. A. (2009). A new consensus for Trypanosoma cruzi intraspecific nomenclature: second
revision meeting recommends TcI to TcVI. Memorias do Instituto Oswaldo Cruz, 104(7), 1051-1054.
Zingales, B., Miles, M. A., Campbell, D. A., Tibayrenc, M., Macedo, A. M., Teixeira, M. M., &
Andrade, S. G. (2012). The revised Trypanosoma cruzi subspecific nomenclature: rationale,
epidemiological relevance and research applications. Infection, Genetics and Evolution, 12(2), 240-
253.
Zingales, B., Miles, M. A., Moraes, C. B., Luquetti, A., Guhl, F., Schijman, A. G., & Ribeiro, I.
(2014). Drug discovery for Chagas disease should consider Trypanosoma cruzi strain
diversity. Memórias do Instituto Oswaldo Cruz, 109(6), 828-833.

85







