Differential descending control of inputs from hairy vs. glabrous skin
1
The degree of acute descending control of spinal nociception
in an area of primary hyperalgesia is dependent on the
peripheral domain of afferent input
1Drake R.A.R., 2Hulse, R.P. *1Lumb B.M and 1,3Donaldson L.F
1School of Physiology and Pharmacology, University of Bristol, 2Cancer Biology,
University of Nottingham and 3Arthritis Research UK Pain Centre and School of Life
Sciences, University of Nottingham.
Category: Neuroscience – behavioural/systems/cognitive
Key words: Nociception, Noradrenaline, Hairy skin
Total word number:
*address for correspondence:
School of Physiology and Pharmacology, The University of Bristol, Medical Sciences
Building, University Walk, Bristol BS7 8EJ.
Tel: +44 117 331 2313
Fax: +44 117 331 2288
Email: [email protected]

Differential descending control of inputs from hairy vs. glabrous skin
2
Abstract
Descending controls of spinal nociceptive processing play a critical role in the
development of inflammatory hyperalgesia. Acute peripheral nociceptor
sensitisation drives spinal sensitisation, and activates spino-supraspinal-
spinal loops leading to descending inhibitory and facilitatory controls of spinal
neuronal activity, that further modify the extent and degree of the pain state.
The afferent inputs from hairy and glabrous skin are distinct with respect to
both the profile of primary afferents classes and the degree of their peripheral
sensitisation. It is not known whether these differences in afferent input
differentially engage descending control systems to different extents or in
different ways.
Injection of Complete Freund’s adjuvant resulted in inflammation and swelling
of hairy hindpaw skin in rats, a transient thermal hyperalgesia lasting <2
hours, and long-lasting primary mechanical hyperalgesia (at least 7d). Much
longer lasting thermal hyperalgesia was apparent in glabrous skin (1h up to
>72h). In hairy skin, transient hyperalgesia was associated with sensitisation
of withdrawal reflexes to thermal activation of either A- and C-nociceptors.
The transience of the hyperalgesia was attributable to a rapidly engaged
descending inhibitory noradrenergic mechanism, which affected withdrawal
responses to both A- and C-nociceptor activation and this could be reversed
by intrathecal administration of yohimbine (alpha-2-adrenoceptor antagonist).
In glabrous skin, yohimbine had no effect on an equivalent thermal
inflammatory hyperalgesia.
We conclude that acute inflammation and peripheral nociceptor sensitisation
in hind paw hairy skin, but not glabrous skin, rapidly activates a descending

Differential descending control of inputs from hairy vs. glabrous skin
3
inhibitory noradrenergic system. This may result from differences in the
engagement of descending controls systems following sensitisation of
different primary afferent classes that innervate glabrous and hairy skin.

Differential descending control of inputs from hairy vs. glabrous skin
4
Key Points
• Acute inflammation engages various descending control systems in the
brain that alter the resulting inflammatory pain, usually by inhibiting it.
• In this study we looked at the differences in inhibition of acute (up to
3h) inflammatory pain from the smooth (glabrous) and the hairy skin in
the rat hind-paw.
• In hairy skin, inflammatory pain is rapidly inhibited by descending
systems that release noradrenaline, but not opiates, into the spinal
cord.
• In glabrous skin, neither descending noradrenergic nor opioidergic
controls affect inflammatory pain.
• These results tell us that the controls on the spinal processing of
cutaneous inflammatory pain are different depending on the skin type
affected.
Abbreviations
Periaqueductal grey (PAG), locus coeruleus (LC), rostroventral medulla
(RVM), Complete Freund’s adjuvant (CFA)

Differential descending control of inputs from hairy vs. glabrous skin
5
Introduction
Tissue damage and subsequent inflammation activates and sensitises
peripheral nociceptors. This increases afferent input to central nociceptive
circuits and leads to enhanced sensitivity to nociceptive stimuli (hyperalgesia),
and sensations of pain to innocuous stimulation (allodynia) (Meyer et al.,
2005; Latremoliere & Woolf, 2009). Peripheral nociceptive inputs drive both
the sensitisation of spinal nociceptive circuits and the activation of ascending
and descending pathways that together modulate transmission, and ultimately
perception, of nociceptive information (Mantyh & Hunt, 2004). The final
perception of the intensity and quality of pain is determined by the degree of
such modulation of information throughout the nociceptive neuraxis (Ossipov
et al., 2010).
It is well established that spinal nociceptive processing is modulated by both
inhibitory and facilitatory descending pathways that originate supraspinally
(Millan, 2002). The midbrain periaqueductal grey (PAG) forms a major
component of the descending nociceptive system and relays via brain stem
nuclei such as locus coeruleus (LC) and the rostroventral medulla (RVM)
(Mantyh, 1983; Bajic & Proudfit, 1999) to exert controls on spinal nociceptive
neurons. Descending controls from the PAG include opioidergic (Przewłocki &
Przewłocka, 2001;Vasquez & Vanegas, 2000; Kwok et al., 2013) and
monoaminergic inhibition (Pertovaara, 2006) as well as prostanergic
facilitation (Oliva et al., 2006; Leith et al., 2007; Palazzo et al., 2011).
Medullary and brain stem nuclei are major sources of spinally projecting
monoaminergic fibres (Westlund et al 1983; Yoshimura & Furue, 2006) that
are known to exert inhibitory influences, which modulate the development of

Differential descending control of inputs from hairy vs. glabrous skin
6
acute and persistent pain states (Vanegas & Schaible, 2004; Pertovaara,
2006, 2013; Yoshimura & Furue, 2006). Descending noradrenergic systems
exert these inhibitory influences largely through presynaptic inhibition,
primarily through alpha-2-adrenoceptor activation (Pertovaara, 2006;
Yoshimura & Furue, 2006). Alpha-2-agonists, in particular those with affinity
for alpha 2A receptors, are antinociceptive in both animals and humans
(Pertovaara, 2006). Descending serotonergic systems exert complex pro- and
anti-nociceptive effects at the spinal level, primarily through 5-HT1 and 5-HT3
autoreceptors (Yoshimura & Furue, 2006; Dogrul et al., 2009; Jeong et al.,
2012). Additionally there are peripheral sources of monoamines, such as
sympathetic fibres within the dorsal root ganglia, that are able to impact upon
nociception during periods of prolonged inflammation or tissue damage
(Pertovaara, 2006). Peripheral nerve stimulation evokes spinal release of
monoamines in the presence of ganglionic blocking agents (Tyce & Yaksht,
1981), however, so it is widely accepted that the major sources of spinal
noradrenaline and serotonin are projections from medullary and brain stem
nuclei (Pertovaara, 2006; Yoshimura & Furue, 2006).
Acute cutaneous inflammation leads to sensitisation of peripheral and central
nociceptive neurons resulting in primary and secondary hyperalgesia.
Descending controls of the spinal processing of input from the area of primary
hyperalgesia is initially facilitatory from, for example PAG/RVM, but are short-
lived (up to 3h), and are then overwhelmed by local spinal and supraspinal
inhibitory influences (Ren & Dubner, 1996; Guan et al., 2002; Miki et al., 2002;
Vanegas & Schaible, 2004).The dampening effect of the inhibitory systems on

Differential descending control of inputs from hairy vs. glabrous skin
7
spinal dorsal horn activity effectively reduces the extent of inflammatory pain
by limiting central sensitisation (Vanegas & Schaible, 2004).
Current understanding of the development of inflammatory hyperalgesia and
the influence of descending controls over spinal nociceptive processing is
largely derived from animal models using inflammatory insults to the glabrous
skin of the hindpaw. There is little information on the contribution of
descending controls to the development or maintenance of primary
hyperalgesia in hairy skin, which is a potential shortcoming given this is the
more prevalent skin type in mammals. Glabrous cutaneous tissue found on
the plantar surface of the paw forms a major informative surface of the body
providing critical sensory/discriminative information about the environment.
This is reflected by a greater density of primary afferents innervating glabrous
tissue and its over representation in cortical somatotopic maps (Blake et al.,
2002; Provitera et al., 2007; Boada et al., 2013). There are notable
differences in the profile of nociceptive afferent fibres that innervate glabrous
and hairy cutaneous tissue, for instance, a threefold higher proportion of fast
conducting to slowly conducting fibres innervating glabrous cutaneous tissue
(Boada et al., 2013) and a large proportion of C-polymodal nociceptors
innervating the hairy skin of the rat hind-paw (Lynn & Carpenter, 1982; Leem
et al., 1993). Critically, there are differences in the sensitisation of
unmyelinated nociceptive afferents innervating the two skin types. Following
application of conditioning stimuli or sensitising agents, a population of
polymodal C-nociceptors found in glabrous skin fail to sensitise to thermal
stimuli but a similar population of polymodal C-nociceptors found in hairy
cutaneous tissue do sensitise, with a lowering of activation threshold and/or

Differential descending control of inputs from hairy vs. glabrous skin
8
increased suprathreshold response (Campbell & Meyer, 1983; Andrew &
Greenspan, 1999; Koerber et al., 2010). We hypothesised that during
inflammation, the differences in primary afferent input from different skin types
might differentially drive descending control systems and hence differentially
affect the development of inflammatory hyperalgesia in the two skin types.

Differential descending control of inputs from hairy vs. glabrous skin
9
Materials and Methods
All experiments were performed in accordance with United Kingdom Animals
(Scientific Procedures) Act (1986) and associated guidelines. Animals were
housed in standard conditions with food and water provided ad libitum.
Experiments were performed on a total of 58 male Wistar rats weighing 250-
350g. The majority of animals (n=55) received a subcutaneous (s.c.) injection
of 50 or 100µg Complete Freund’s Adjuvant (CFA) (1mg/ml; Cat No: F5881,
Sigma-Aldrich, UK) into either dorsal or ventral surface of the left hind-paw
under brief halothane (3% in O2) anaesthesia. Control animals (n=3) received
a s.c. injection of an equivalent volume of vehicle (mineral oil). CFA induces a
dose dependent increase in swelling, in both rat and mouse, up to doses of
250µg and 500µg respectively. This CFA dose/volume (50µg/50µl) gives a
mild, limited inflammation that is less than the maximal achievable with CFA,
and that does not spread to involve the sides or other surface of the hindpaw.
Greater swelling and spread of inflammation can be seen with higher doses of
CFA (Donaldson et al., 1993; Chillingworth et al., 2006). Although differences
in oedema have been previously reported with higher CFA doses (Cook &
Moore, 2006), we chose to investigate the effects of equivalent inflammatory
stimuli in this study, rather than equivalent oedematous responses.
Nociceptive behavioural testing.
To assess the effect of s.c. CFA injections at different sites on nociceptive
behaviour, some animals (n=7) underwent nociceptive testing before, three
hours, one, three and seven days after either dorsal or plantar s.c CFA
injection. Animals were habituated to the apparatus and experimenter

Differential descending control of inputs from hairy vs. glabrous skin
10
beginning 3 days prior to the start of the testing. Mechanical hyperalgesia:
Serrated laboratory forceps were adapted with strain gauges for the
measurement of the force applied across the tips (contact area 5.6mm2 per
tip, total 11.2mm2, Supp. Figure 1). The output signal was fed through a
bridge amplifier and captured for subsequent offline analysis via a CED1401
(Cambridge Electronic Design (CED), Cambridge UK) on a computer running
Spike2 software (CED, Cambridge, UK). Grams per volt were calculated
following calibration with standardised weights placed at the point of finger
grip. The gram force delivered was then divided by the total contact area to
give grams.mm-2. The tips were placed across the dorsoventral aspect of the
hind paw and ramped pressure was applied until the foot was withdrawn or
the animal displayed pain-related behaviours (vocalisation, biting etc.) at
which point the force applied was immediately stopped. As the duration and
force applied were controlled manually, and cut-off on first appearance of
pain-related behaviours, the intensity of this stimulus could be controlled to
ensure that it did not result in additional tissue damage. Use of this stimulator
did not result in any observable tissue damage in naïve or inflamed animals.
The occurrence of withdrawal/response was marked using the input from a
foot pedal through the CED1401 into a computer running Spike2 software.
Two consecutive recordings were made and mean threshold calculated for
each animal at each time point.
To test for thermal hyperalgesia a custom built Peltier heating device (contact
area 18mm2) was used to deliver a ramped thermal stimulus to the hind paw
of the rat from an initial contact temperature of 30oC. The maximum
temperature was cut off at 55oC to prevent tissue damage. Surface (contact)

Differential descending control of inputs from hairy vs. glabrous skin
11
temperature was measured with a T-type thermocouple (made in-house) and
captured for subsequent offline analysis via a CED1401 on a computer
running Spike5 software. The extent of paw oedema was assessed by
measuring the thickness across the midline dorsoventral aspect of the hind
paw using callipers before, and 1 hour, 1, 3 and 7 days after CFA injection.
Surgical preparation
Surgical preparation for electromyograph recordings was performed under
initial halothane anaesthesia (2-3% in O2) and consisted of; (i) external jugular
branch cannulation for anaesthetic maintenance (constant intravenous
infusion of alphaxalone (~40mg.kg.h; Alfaxan; Jurox Pty PLC, Australia)), (ii)
external carotid artery branch cannulation for blood pressure measurement
and (iii) tracheal cannulation for airway maintenance. Body temperature was
maintained within physiological limits (37-38oC) by means of a feedback
controlled heating blanket and rectal probe. For placement of the intrathecal
catheter (Størkson et al., 1996), a longitudinal incision was made along the
back midline, starting at the level of the iliac crest and advancing 2-3cm
rostrally. A 6cm length of 32G polyurethane catheter (OD: 0.25mm, ID:
0.13mm ) with a manufacturer-supplied internal metal stylet for increased
rigidity (Cat No: 0041; ReCathCo, USA) was back loaded into a 25G needle
(Terumo Medical Corporation) so that the end of the catheter was flush with
the needle tip. Animals were positioned with the vertebral column flexed and
the catheter-loaded needle was inserted bevel up between L5 and L6
vertebrae until a tail flick indicated penetration of dura. The catheter was then
advanced ~3cm rostrally along the intrathecal space to approximately the
level of the lumbar enlargement. The needle and stylet were then carefully

Differential descending control of inputs from hairy vs. glabrous skin
12
removed leaving the catheter in place, which was then fixed in position with
cyanoacrylate glue (Superglue). A small (~2cm) length of polyethylene tubing
(OD: 6.1mm, ID: 2.8mm) was then attached to the free end of the catheter, by
insertion of the catheter end just into the polyethylene tubing, which was then
secured with cyanoacrylate, to allow for connection to a Hamilton syringe for
drug delivery.
For the measurement of electromyographic (EMG) activity, a custom made bi-
polar electrode was made using Teflon coated stainless steel wires (0.075mm
diameter; Advent research Materials, UK) that were stripped at both ends and
one end inserted into the bicep femoris of the hind-leg using a 25-gauge
hypodermic needle. The signal across the electrodes was amplified (x1K,
Neurolog NL104 amplifier, A-B configuration; Digitimer, UK), filtered (50Hz-5
KHz, Neurolog NL125) and raw data digitised via the CED1401, and stored
for offline analysis using Spike2. Animals were left under a constant level of
anaesthesia for a minimum of 1 hour after surgical preparation before further
experiments. The level of anaesthesia was maintained by constant infusion
such that EMG activity could be detected in response to stimulation without
overt paw movement.
Preferential activation of A- and C-heat nociceptors.
A- and C-nociceptor were preferentially activated using a custom-made heat
lamp in contact with the dorsal hind paw. A constant voltage was applied to
the lamp to provide fast (7.5±1oC.s-1) or slow rates of heating (2.5±1oC.s-1),
which preferentially activate A-fibre (myelinated, capsaicin-insensitive) and C-
fibre (unmyelinated, capsaicin-sensitive) heat nociceptors respectively

Differential descending control of inputs from hairy vs. glabrous skin
13
(Yeomans & Proudfit, 1996; McMullan et al., 2004; McMullan & Lumb, 2006a;
Leith et al., 2007). Stimuli were applied with an inter-stimulus interval of at
least 8 minutes to prevent tissue damage and sensitisation. The heat ramp
apparatus was placed in contact with the foot for a minimum of 60 seconds
before the start of the heat stimulation to allow for the adaptation for low
threshold mechanoreceptors. A feedback-controlled cut-off was set at 58oC
for fast thermal ramps and 55oC for slow thermal ramps to prevent tissue
damage. In recordings in which thermal ramps reached the cut-off
temperature without the occurrence of measurable EMG activity, threshold
was recorded as cut-off + 2oC (Leith et al., 2007). For stimulation of glabrous
skin, a slow ramp (2.5±1oC.s-1) contact heat stimulus was applied immediately
behind the foot-pad using the same apparatus. As it has not yet been
determined whether this stimulus is able to preferentially activate A- or C-
nociceptors in glabrous skin no attempt was made to draw conclusions with
respects to A- or C-nociceptor activation at this site.
Subsurface heating rate measurement
To determine whether CFA-induced paw oedema affected the subcutaneous
heating rates that are required for preferential activation of A- and C-
nociceptors, subcutaneous heating rates were directly measured using a T-
type thermocouple. Heating rates were recorded before, for three hours and
on the seventh day after s.c. CFA injection. Care was taken to implant the
thermocouple as close to the dermal/epidermal border as possible as
nociceptive terminals are found in epidermis and superficial dermal layers.
Two ramps were delivered at each experimental time point.

Differential descending control of inputs from hairy vs. glabrous skin
14
Drugs
The alpha-2-adrenoreceptor antagonist yohimbine 30µg in 10µl in vehicle
(80% saline/20%DMSO), Cat no: 1127 Tocris Biosciences, Bristol, UK) was
delivered intrathecally via the implanted catheter 2.5h after CFA (n=9) so that
the peak drug effect coincided with the third hour following CFA. The
concentration of yohimbine used was based on previous reports of effective
intrathecal alpha-2 adrenoceptor blockade (Ossipov et al., 1989; Takano &
Yaksh, 1992; Green et al., 1998). At the time of injection the tip of a 100µl
Hamilton syringe was inserted in to the PE10 tubing and 10µl of the drug was
delivered over several minutes. This was washed through with an equivalent
volume of saline. In a separate group of animals (n=6) the non-specific opioid
antagonist naloxone (3mg.kg-1 in 0.9%saline) (Sigma-Aldrich, UK, Cat no:
N7758) was delivered intraperitoneally so that the peak drug effect coincided
with the second hour following CFA.
At the end of the experiment the position of the cannula was determined by
injecting 20µl of Xylocaine (2%) through the cannula. If pinch-evoked EMG
activity was abolished the cannula was deemed to be correctly positioned, if
not the data were not included in subsequent analyses (n=1, CFA injected).
Additionally, in some experiments the location of the distal tip of the catheter
was determined by injection of pontamine Sky Blue (20µl of 5%) and visual
confirmation of the location of the catheter tip.
Statistical analyses
Data are displayed as mean + SEM unless otherwise noted in Figure legends.
Data were analysed using Graphpad Prism v4/5 using Mann Whitney test (2

Differential descending control of inputs from hairy vs. glabrous skin
15
group comparisons) or Kruskal-Wallis, one- or two-way ANOVA (3 or more
groups) where appropriate and as stated in Figure legends. *p<0.05,
**p<0.01, ***p<0.001, ****p<0.0001.

Differential descending control of inputs from hairy vs. glabrous skin
16
Results
Consequences of CFA induced inflammation of the hairy skin of the rat hind
paw.
Injection of 50µg CFA into the hind-paw dorsum resulted in paw oedema that
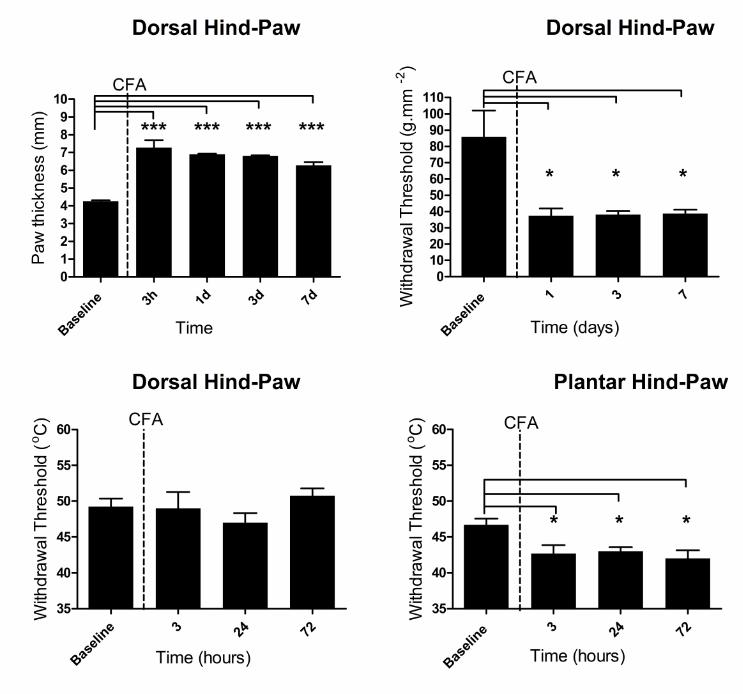
was evident after one hour and persisted for 7 days (Figure 1A; baseline:
4.2±0.1mm vs seven days: 6.3±0.2mm, p<0.001). CFA also produced a
mechanical hyperalgesia to a noxious pinch (Figure 1B; baseline 87±33g.mm-
2 vs. seven days 39±5g.mm-2, p<0.05) but did not induce a thermal
hyperalgesia in hairy skin (Figure 1C; baseline 49±1.1oC, three hours
49±2.3oC, twenty-four hours 47±1.3oC and three days 51±1.0oC). An
equivalent subcutaneous injection of CFA into the plantar surface of the hind-
paw resulted in thermal hyperalgesia that persisted for at least three days
(Figure 1D, baseline 47±0.9 oC vs. three days 42±1.2oC, p<0.05).
The ability of the thermal ramp stimuli to preferentially activate A- and C-
nociceptors is reliant on achieving subsurface cutaneous heating rates of
2.5oC.s-1 (fast) for A-nociceptors and 0.8oC.s-1 (slow) for C-nociceptors
(Yeomans & Proudfit, 1996; McMullan et al., 2004; McMullan & Lumb, 2006a;
Leith et al., 2007). As paw oedema could affect the heat transfer through the
skin, and thus subcutaneous heating rates, we first determined the effect of
acute inflammatory oedema on cutaneous heating rates. Surface heating
rates elicited by both fast and slow heating ramps were kept within limits
previously shown to elicit correct subsurface heating rates in normal skin
(Fast - 7.5 oC.s-1, Slow - 2.5 oC.s-1; McMullan et al., 2004) and the
corresponding subsurface heating rates were recorded. Subsurface heating

Differential descending control of inputs from hairy vs. glabrous skin
17
rates to both fast and slow thermal ramps were unchanged over the three
hours of acute inflammation (Supp. Figure 2, p>0.05). However, at seven
days following CFA, subsurface heating rates elicited by slow thermal ramps
were significantly faster than pre-CFA values (Suppl. Figure 2B; baseline
subsurface rate 0.6 ± 0.03oC.s-1 vs. seven day subsurface rate 1.2 ± 0.1oC.s-1;
p<0.05).
Effect of CFA induced inflammation on the spinal processing of thermal
nociceptor inputs from hairy and glabrous skin.
We sought to determine the effects of CFA induced inflammation of hairy skin
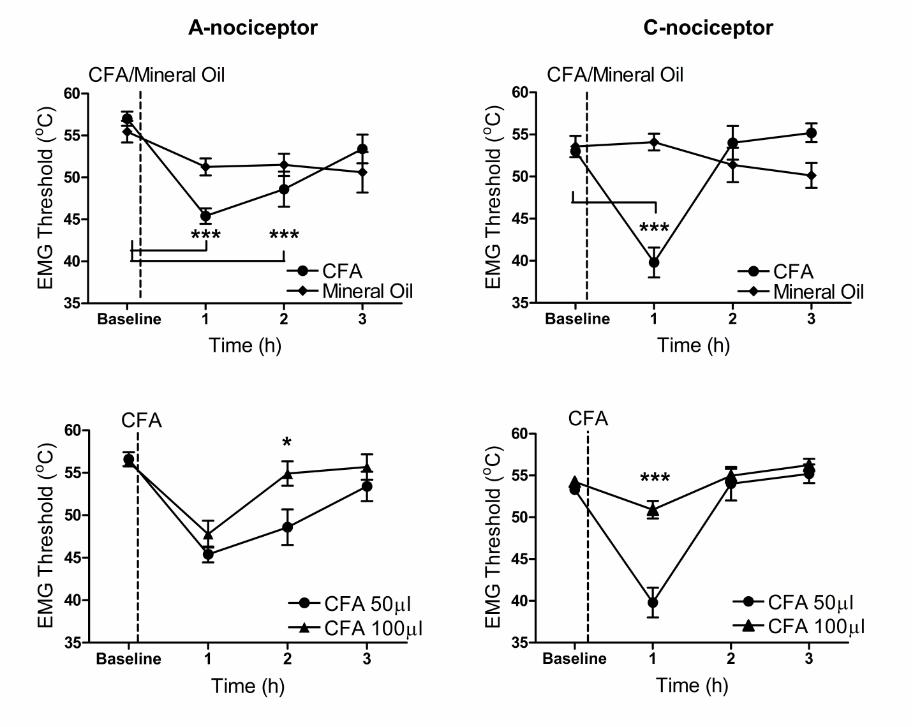
on the spinal processing of A- and C-nociceptor inputs. Withdrawal thresholds
to thermal A- and C-nociceptor stimulation were both significantly lower one
hour after CFA injection (Figure 2A; baseline A-nociceptors vs. one hour A-
nociceptors, 57 ± 0.8oC vs. 45 ± 0.9oC, p<0.001) (Figure 2B; baseline C-
nociceptors vs. one hour C-nociceptors, 53 ± 0.4oC vs. 40 ± 1.8oC, p<0.001).
After 2 hours, withdrawal threshold to C-nociceptor stimulation had returned to
slightly above baseline levels (Figure 2B; baseline vs. two hours, 53 ± 0.4oC
vs. 54 ± 2.0oC) whereas A nociceptor thresholds were still lowered (Fig 2A;
baseline vs. two hour, 57 ± 0.8oC vs. 49 ± 2.0oC, p<0.001). By three hours A
and C nociceptor thresholds had returned to baseline values (Figure 2A & B,
p>0.05). Vehicle injection had no significant effect on A- or C-nociceptor
thresholds (Fig 2A & B; p>0.05). It should be noted that in three of five
animals, at two and three hours following CFA, no EMG activity to C-
nociceptor stimulation was evoked before stimulation reached the imposed
cut-off temperature. Furthermore, in two additional animals (data not
used/shown in further analysis) the cut-off was not imposed and in these

Differential descending control of inputs from hairy vs. glabrous skin
18
animals we found thresholds for withdrawal to C-nociceptor activation to be
greater than 60oC. In contrast thresholds of withdrawal to A-nociceptor
activation where always below the imposed temperature cut-off.
Doubling the amount of injected CFA resulted in significantly less sensitisation
of the reflex pathway over the three hour period (Figure 2C & D). Compared
to the 50µg dose, a 100µg CFA injection caused similar reduction in A-
nociceptor withdrawal threshold at one hour, but surprisingly significantly less
of a reduction in C-nociceptor threshold at this time (Figure 2D; 50µg at 1h vs.
100µg at 1h, 40 ± 1.8oC vs. 51 ± 1.0oC, p<0.001). At two hours, both C- and
A-nociceptor withdrawal threshold had return to baseline values and A-
nociceptor withdrawal thresholds were significantly higher when compared to
threshold values for the 50µg dose at the same time point (Figure 2C; 50µl at
two hour vs. 100µl at two hour, 49 ± 2.0oC vs 55 ± 1.4oC, p<0.05).
Although CFA is known to produce persistent afferent sensitisation lasting
days/weeks (Ren & Dubner, 1999), following dorsal CFA we found thermal
nociceptive behaviour to be unchanged at 3 days (Figure 1). Additionally,
seven days after CFA injection, withdrawal thresholds to A- and C-nociceptor
activation were also unchanged (C-nociceptor naïve vs. C-nociceptor seven
day CFA, 53 ± 0.6oC. vs. 55 ± 2.0oC; Mann-Whitney p>0.05, n=4; A-
nociceptor naïve vs. A-nociceptor seven day CFA, 58 ± 0.5oC vs. 58 ± 0.6oC;
Mann-Whitney p>0.05, n=4, not shown). In contrast, although the withdrawal
threshold did increase slightly towards baseline over three hours, injection of
CFA into the glabrous skin of the plantar hind-paw resulted in a significant
reduction in withdrawal thresholds to the thermal stimulus for the three hours
studied (Figure 3 - p<0.05 at all time points). It has been previously shown

Differential descending control of inputs from hairy vs. glabrous skin
19
that s.c. CFA delivered to the plantar surface of the paw leads to thermal
hyperalgesia lasting more than one week (Ren & Dubner, 1999). Differences
in sensitisation might be attributable to differences in inflammatory reaction
and/or oedema. There was significantly greater swelling in the plantar surface
of the hindpaw 3 h after injection of CFA compared to the dorsal surface
(Figure 4. 3h dorsal 5.9±0.2mm n=7; plantar 6.9±0.2mm, n=6, p<0.05). The
degree of swelling of the hindpaw was not significantly different after injection
of 100µg CFA, compared with 50µg CFA injected into the dorsal surface
(Figure 4, 3h 100µg dorsal 6±0.2mm, n=5).
Effect of spinal alpha-2-adrenoceptor receptor blockade on the processing of
thermal nociceptor inputs during hind paw inflammation.
Following inflammation of hairy skin the observation of sensitisation followed
by a reversal over minutes/hours suggested that descending inhibitory
mechanisms could be affecting spinal processing in acute inflammation.
Intrathecal delivery of the alpha-2-adrenoceptor antagonist, yohimbine (30µg
in 10µl) (Takano & Yaksh, 1992; Green et al., 1998; Hughes et al., 2013), 2
hours after CFA, when thresholds had returned (C-nociceptors) or were
returning to baseline (A-nociceptors), significantly inhibited the reversal of
both A- and C-nociceptor withdrawal thresholds (Figure 5A; 3 hour A-
nociceptor CFA only vs. 3 hour A-nociceptor CFA+yohimbine, 55 ± 1.3oC vs.
50 ± 0.9oC, p<0.05) (Figure 5B; 3 hour C-nociceptor CFA only vs. 3 hour C-
nociceptor CFA+yohimbine, 55 ± 1.1oC vs. 47 ± 2.3oC, p<0.05). In contrast,
yohimbine did not affect the withdrawal to thermal stimulation applied to

Differential descending control of inputs from hairy vs. glabrous skin
20
inflamed glabrous skin (Figure 3), which was apparent at 1 hour, and
maintained for the 3 hours studied (Figure 3; CFA only 3h threshold vs.
CFA+yohimbine 3h withdrawal threshold, 47 ± 0.7oC vs. 47 ± 0.4oC).
Intrathecal delivery of the same volume of vehicle did not affect withdrawal
thresholds in either experiment (n=3 for both experiments, data not shown).
Spinal and supraspinal opioidergic systems are known to have robust
inhibitory effects on spinal nociceptive processing during inflammation (Stanfa
& Dickenson, 1995; Przewłocki & Przewłocka, 2001). We therefore also
determined the contribution of opioidergic systems to the degree of
hyperalgesia in acute hind paw CFA inflammation. Systemic delivery of the
pan-opioid receptor antagonist naloxone (3mg.kg-1 (Stanfa & Dickenson,
1995)) had no effect on either A- or C-nociceptor withdrawal thresholds in
hairy skin at either 2 or 3 hours following CFA (Figure 5C + D). It has been
previously shown that naloxone has no effect on withdrawal latencies to
thermal stimuli at 4 hours following plantar inflammation (Hylden et al., 1991;
Tsuruoka & Willis, 1996).

Differential descending control of inputs from hairy vs. glabrous skin
21
Discussion
Two key observations arise from these data; (i) there is a differential
descending noradrenergic inhibitory control on different skin types in acute
inflammation, and (ii) there is a greater descending inhibitory control of C-
compared to A-nociceptor evoked reflexes from hairy skin.
Subcutaneous injection of CFA is a well characterised and widely used
cutaneous inflammatory model, primarily used to study inflammatory
hypersensitivity in glabrous (plantar) skin (Iadarola et al., 1988; Ren &
Dubner, 1999). The oedema generated in glabrous or hairy skin is not
equivalent for the same dose of CFA (data herein, (Cook & Moore, 2006)), but
as CFA produces a dose-dependent inflammatory response (Donaldson et al.,
1993) that should be equivalent at different sites, inflammation of hairy and
glabrous skin might be predicted to induce equivalent primary hyperalgesia.
Literature reports that CFA injection results in a robust primary mechanical
and, usually, following plantar injection, thermal behavioural hypersensitivity
to stimulation from as early as one hour and reportedly lasting for weeks
(Iadarola et al., 1988; Ren & Dubner, 1999; Terayama et al., 2002; Soignier et
al., 2011). Plantar inflammation also results in mechanical hypersensitivity
(data herein, (Ren & Dubner, 1999; Cook & Moore, 2006; Soignier et al.,
2011). In contrast, dorsal (hairy) hindpaw inflammation results in no change in
thermal reflex withdrawal response, despite thermal sensitisation of single
peripheral nociceptors in the same model (Dunham et al., 2008; Koerber et
al., 2010). CFA induced inflammation results in robust long-lasting peripheral
nociceptor thermal sensitisation, in rats, guinea pigs and primates, (Kocher et

Differential descending control of inputs from hairy vs. glabrous skin
22
al., 1987; Davis et al., 1993; Andrew & Greenspan, 1999; Djouhri & Lawson,
1999; Djouhri et al., 2001; Dunham et al., 2008; Koerber et al., 2010)
irrespective of whether hairy or glabrous skin is inflamed, which is inconsistent
with the behavioural data. We therefore determined whether the observed
lack of thermal behavioural responses in acute inflammation might be
attributable to descending inhibitory control of nociceptive processing from
inflamed hairy skin sites. The observation that plantar inflammation produces
greater swelling, but little obvious descending inhibition, whereas dorsal
inflammation produces less swelling but more profound inhibition with
increasing inflammatory severity, suggest that it is the overall degree of
inflammation, rather than just the oedema, that engages descending control
systems. This is consistent with previous findings, where much higher CFA
doses produced acute thermal analgesia i.e. less nociception than control, in
hairy but not glabrous skin (Cook & Moore, 2006). Differences in inflammatory
oedema in different sites may not therefore accurately predict nociceptive
responses, as this aspect of inflammation may more accurately reflect local
tissue compliance (Reed & Rubin, 2010) rather than nociceptive input.
Processing of noxious inputs is known to be subject to both descending
inhibitory and facilitatory controls (Millan, 2002; Vanegas & Schaible, 2004).
Primary inflammatory hyperalgesia is widely accepted to be dominated by
descending inhibitory controls, which are both rapidly activated and long-
lasting. These are hypothesised to protect against spinal hyperexcitabilty
following inflammation by limiting mechanisms of central sensitisation (Ren &
Dubner, 1996; Vanegas & Schaible, 2004). However, they are not usually
reported to completely obliterate the nociceptive sensitivity resulting from the

Differential descending control of inputs from hairy vs. glabrous skin
23
insult (Tsuruoka & Willis, 1996; Tsuruoka et al., 2003; Vanegas & Schaible,
2004). This is in contrast to our findings where thermal hyperalgesia in hairy
skin was more short lived, particularly so when inflammation was more severe
(100µg CFA dose).
Descending fibres from areas such as LC and RVM exert profound
modulatory influences on spinal nociceptive processing (Fields et al., 1991;
Tsuruoka & Willis, 1996; Miki et al., 2002; Millan, 2002; Tsuruoka et al., 2003)
through monoaminergic systems (Pertovaara, 2006; Yoshimura & Furue,
2006). Descending monoaminergic systems, particularly noradrenaline acting
through spinal alpha-2 receptors, are reported to modulate the development
of inflammatory hyperalgesia (Tsuruoka & Willis, 1996; Tsuruoka et al., 2003;
Pertovaara, 2006). Our findings confirm this for hairy cutaneous tissue, and
additionally indicate that, at acute time points, yohimbine blockade
differentially modulates thermal inflammatory sensitivity in hairy/glabrous skin.
It has been proposed that the descending noradrenergic systems form a
stimulus driven negative feedback circuit that impinges onto spinal nociceptive
processing and so regulates the onward flow of ascending nociceptive
information (Pertovaara, 2006, 2013). In hairy skin, our finding of a greater
inhibition in more severe inflammation would support this. In contrast, in
glabrous skin hypersensitivity persists (>4h) (Tsuruoka & Willis, 1996) and at
acute time points (<3h) we find little evidence of either monoaminergic or
opioidergic inhibition, consistent with previous findings (Hylden et al., 1991;
Tsuruoka & Willis, 1996).

Differential descending control of inputs from hairy vs. glabrous skin
24
Although yohimbine has high affinity for alpha-2 receptors and is commonly
used as a specific alpha-2-adrenoceptor antagonist, it has moderate affinity at
alpha-1-adrenoceptors, 5-HT1A, 5-HT1B and dopamine D2 receptors (Ki
determinations generously provided by the National Institute of Mental
Health’s Psychoactive Drug Screening Program,
(http://pdsp.med.unc.edu/pdsp.php)). As a result of the latter effects, systemic
yohimbine can decrease 5-HT and increase dopamine and noradrenaline
levels in rat CNS, probably through blockade of autoreceptors (Paalzow &
Paalzow, 1983; Millan et al., 2000). In inflammation, spinal 5-HT1A receptors
both enhance (Zhang et al., 2001; Wang et al., 2003) and inhibit (Liu et al.,
2002; Yu et al., 2011; Horiguchi et al., 2013) acute inflammatory nociception;
known descending 5-HT3 receptor-mediated facilitatory pathways do not
seem to contribute to inflammatory nociception (Asante & Dickenson, 2010).
There are no published data on contributions of spinal D2 receptors to acute
inflammatory nociception. Although this concentration of yohimbine
administered intrathecally has been widely interpreted as exerting actions
primarily at alpha-2 adrenoceptors (Ossipov et al., 1989; Takano & Yaksh,
1992; Green et al., 1998; De Felice et al., 2011; Little et al., 2012; Hughes et
al., 2013), we cannot exclude an additional contribution of modulation of
alpha-1-adreno or 5-HT receptors at the spinal level to the observed effects of
yohimbine. Actions of yohimbine on 5-HT1A receptors would be consistent
with a concurrent anti-nociceptive effect of 5-HT in this model (Liu et al., 2002;
Yu et al., 2011; Horiguchi et al., 2013).
The nocifensive flexor reflex is thought to provide an objective measure of
noxious threshold and is the most commonly used in pain studies in both

Differential descending control of inputs from hairy vs. glabrous skin
25
humans and animals (Wiesenfeld-Hallin, 1995; Sandrini et al., 2005). In
animals the nocifensive flexor reflex typically involves recording
electromyographic activity from the biceps femoris evoked by stimulation of
the hind-paw (Wiesenfeld-Hallin, 1995), alpha motor neurons serving the
biceps femoris have receptive fields on both the dorsal and plantar surface
(Cook & Woolf, 1985), thus the effects of drugs on reflexes evoked from both
paw surfaces can be directly compared and contrasted. Thus, whilst the
musculotopic organisation of reflexes may have meant that recording from
different muscular sites may have yielded slightly different results
(Schouenborg et al., 1994; Harris & Clarke, 2002), these may not have been
directly comparable for stimulation of different sites on the hindpaw. In
addition, as alpha-2-adrenoceptor agonists are analgesic (Pertovaara, 2006)
and presumably inhibit the sensory afferent arm of the withdrawal reflex, it
could be hypothesised that the result of yohimbine blockade would affect, to a
greater or lesser extent, all muscle groups involved in noxious withdrawal
reflexes. This is supported by the behavioural data.
A differential descending inhibitory control on glabrous versus hairy skin may
relate to the functional importance of the footpad/glabrous skin. The plantar
surface forms the main exploratory hindpaw surface in rodents, providing
important sensory/discriminative information about the environment (Boada et
al., 2013). Pain provides a critical protective mechanism in which nociceptive
sensitisation, including increased reflex sensitivity, serves to protect injured
areas that might regularly come into contact with physical stimuli
(Schouenborg et al., 1994) and it has been shown that load bearing parts of
the paw have special significance in generating reflex sensitisation (Clarke &

Differential descending control of inputs from hairy vs. glabrous skin
26
Harris, 2004). However, effective locomotion is essential to survival, being
central to feeding and escape behaviours, and therefore the positive impact of
nociceptive reflex hypersensitivity and its protective function must be traded
off against its negative impact on locomotion. Thus peripheral domains that
rarely come into contact with nociceptive stimuli (dorsal hind-paw) may have a
reduced capacity to produce reflex sensitisation following inflammation.
The differences in descending inhibition of spinal processing of afferent input
from different peripheral domains may also represent a difference in both
innervation and the effects of inflammation on innervating neuronal properties.
The inflammatory sensitisation of unmyelinated afferents differs in glabrous
versus hairy skin in both rat and primate (Campbell & Meyer, 1983; Kocher et
al., 1987; Andrew & Greenspan, 1999; Dunham et al., 2008), in that
peripheral thermal sensitisation occurs in C-nociceptors innervating hairy skin
(Campbell & Meyer, 1983; Davis et al., 1993; Dunham et al., 2008; Koerber et
al., 2010), but not in glabrous skin (Meyer & Campbell, 1981; Campbell &
Meyer, 1983; Andrew & Greenspan, 1999; Du et al., 2006), where C-
nociceptors may become desensitised instead (Campbell & Meyer, 1983;
Andrew & Greenspan, 1999). C-nociceptors play critical roles in driving
mechanisms of central sensitisation and of descending controls from
supraspinal sites (Woolf & Wall, 1986; Sivilotti et al., 1993; Suzuki et al., 2002;
Ikeda et al., 2003; Mantyh & Hunt, 2004; You et al., 2010). Sensitisation of C-
nociceptors in inflamed hairy skin would drive ascending, and therefore also
descending systems to a greater extent than in glabrous skin, where C-
nociceptors do not readily sensitise, leading to the differences in inhibition
observed. This is supported by the enhanced inhibition seen in a more severe

Differential descending control of inputs from hairy vs. glabrous skin
27
inflammation where nociceptors would be sensitised more rapidly and afferent
barrage would be greater (Figure 3 A & B).
Previous findings from our laboratory indicate that descending inhibitory
controls, arising from the PAG preferentially target C-nociceptive inputs to
spinal nociceptive networks (McMullan & Lumb, 2006b). Here, during acute
inflammation of hairy skin, we observe a fast acting and potent inhibition of C-
nociceptor evoked withdrawals; at two and three hours following CFA the
withdrawal thresholds to C-nociceptor stimulation were greater than baseline
values and in 3/5 animals we observed no evoked EMG activity before the
cut-off temperature was reached. Additionally, in the two cases where the cut-
off was removed, thresholds for EMG activity were greater than 60oC (data
not shown). In contrast, at two hours following CFA, thresholds for A-
nociceptor evoked withdrawals were still significantly lower than baseline
values and, at three hours following CFA, all 5 animals responded to the
thermal stimuli before the cut-off temperature was reached. It is likely that the
effect of yohimbine on C- versus A-nociceptor evoked withdrawals failed to
reach significance (data not shown) as a result of the inherent
underestimation of the full effect of yohimbine on C-nociceptor withdrawal
thresholds due to the imposed temperature cut-off (55oC). Descending
inhibitory systems have been shown to preferentially target thermal spinal
nociceptive processing in both normal and inflamed rats (Kauppila et al.,
1998; Howorth et al., 2009), consistent with our observations. Additionally, as
noxious mechanical stimuli are thought to be conveyed by A-fibre nociceptors
(Ziegler et al., 1999; Magerl et al., 2001) an alternative interpretation of these

Differential descending control of inputs from hairy vs. glabrous skin
28
findings is that there as a preferential inhibition of spinal nociceptive
processing of C- versus A-nociceptor inputs.
There may be a possible evolutionary advantage of greater inhibitory control
on C- compared to A-nociceptor-evoked reflexes which could relate to the
different functions of these afferents. In humans, rapidly conducting A-
nociceptors convey sharp, well localised painful sensations (Magerl et al.,
2001), information which is encoded with a high fidelity by spinal neurons (rat)
(McMullan & Lumb, 2006a). In contrast, slowly conducting C-nociceptors
convey dull and diffuse pain (Ziegler et al., 1999; Magerl et al., 2001) encoded
with low fidelity by spinal neurons in rats (McMullan & Lumb, 2006a). Thus A-
nociceptors are hypothesised to convey more detailed information relating to
the protective function of pain, i.e. localisation and magnitude assessment. A
greater acute inhibition of C-nociceptor inputs could serve to filter extraneous
C-nociceptive information and limit saturation in the reflex pathway (Tsuruoka
et al., 2012) while concurrently preserving A-nociceptive information which is
essential to the protective function of reflex withdrawals. Thus differential
inhibitory control of inputs from body areas with different functions, and from
different nociceptor types would be hypothesised to limit central sensitisation
and protect against spinal hyperexcitabilty following inflammation, whilst
maintaining adequate locomotor and protective nociceptive function to enable
healing/survival.

Differential descending control of inputs from hairy vs. glabrous skin
29
Competing interests:
The authors have no competing interests to declare.
Author’s contributions:
Experiments were performed in laboratories in the School of Physiology and
Pharmacology, University of Bristol. RARD, BML & LFD contributed to
experimental design, and data analysis and interpretation, RPH designed and
built the mechanical stimulator, RARD contributed data collection, RARD,
BML & LFD wrote the manuscript, and all authors approved final manuscript
submission.
Funding
This work was funded by the Medical Research Council (DTP studentship
supporting RARD, Project grant G0801381 (BML)) and a BBSRC/MRC/BPS
Strategic Skills award (LFD & BML).
Acknowledgements:
We would like to thank Prof. Bruce Matthews for assistance with the design
and build of the mechanical pincher, and additionally Dr. Louise Hickey and
Mr Sam Hughes for their helpful discussions and input on technical issues.
Supporting information
The online supporting information gives further detail on the mechanical
stimulation device, and the validation of the heat ramp stimulations in inflamed
skin.

Differential descending control of inputs from hairy vs. glabrous skin
30
References:
Andrew D & Greenspan J (1999). Mechanical and heat sensitization of cutaneous nociceptors after peripheral inflammation in the rat. J Neurophysiol 82, 2649–2656.
Asante CO & Dickenson AH (2010). Descending serotonergic facilitation mediated by spinal 5-HT3 receptors engages spinal rapamycin-sensitive pathways in the rat. Neurosci Lett 484, 108–112.
Bajic D & Proudfit HK (1999). Projections of neurons in the periaqueductal gray to pontine and medullary catecholamine cell groups involved in the modulation of nociception. J Comp Neurol 405, 359–379.
Blake DT, Byl NN & Merzenich MM (2002). Representation of the hand in the cerebral cortex. Behav Brain Res 135, 179–184.
Boada MD, Houle TT, Eisenach JC & Ririe DG (2013). Differing neurophysiologic mechanosensory input from glabrous and hairy skin in juvenile rats differing neurophysiologic mechanosensory input from glabrous and hairy skin in juvenile rats. J Neurophysiol 104, 3568–3575.
Campbell JN & Meyer RA (1983). Sensitization of unmyelinated nociceptive afferents in monkey varies with skin type. J Neurophysiol 49, 98–110.
Chillingworth NL, Morham SG, Donaldson LF (2006). Sex differences in inflammation and inflammatory pain in cyclooxygenase-deficient mice. Am J Physiol Regul Integr Comp Physiol 291, 327–334.
Clarke RW & Harris J (2004). The organization of motor responses to noxious stimuli. Brain Res Brain Res Rev 46, 163–172.
Cook AJ & Woolf CJ (1985). Cutaneous receptive field and morphological properties of hamstring flexor α-motorneurones in the rat. J Physiol 364, 249–263.
Cook CD & Moore KI (2006). Effects of sex, hindpaw injection site and stimulus modality on nociceptive sensitivity in arthritic rats. Physiol Behav 87, 552–562.

Differential descending control of inputs from hairy vs. glabrous skin
31
Davis KD, Meyer RA & Campbell JN (1993). Chemosensitivity and sensitization of nociceptive afferents that innervate the hairy skin of monkey. J Neurophysiol 69, 1071–1081.
Djouhri L, Dawbarn D, Robertson A, Newton R & Lawson SN (2001). Time course and nerve growth factor dependence of inflammation-induced alterations in electrophysiological membrane properties in nociceptive primary afferent neurons. J Neurosci 21, 8722–8733.
Djouhri L & Lawson SN (1999). Changes in somatic action potential shape in guinea-pig nociceptive primary afferent neurones during inflammation in vivo. J Physiol 520, 565–576.
Dogrul A, Ossipov MH & Porreca F (2009). Differential mediation of descending pain facilitation and inhibition by spinal 5HT-3 and 5HT-7 receptors. Brain Res 1280, 52–59.
Donaldson LF, Seckl JR & McQueen DS (1993). A discrete adjuvant-induced monoarthritis in the rat: effects of adjuvant dose. J Neurosci Methods 49, 5–10.
Du J, Zhou S & Carlton SM (2006). Kainate-induced excitation and sensitization of nociceptors in normal and inflamed rat glabrous skin. Neuroscience 137, 999–1013.
Dunham JP, Kelly S & Donaldson LF (2008). Inflammation reduces mechanical thresholds in a population of transient receptor potential channel A1-expressing nociceptors in the rat. Eur J Neurosci 27, 3151–3160.
Fields HL, Heinricher MM & Mason P (1991). Neurotransmitters in nociceptive modulatory circuits. Annu Rev Neurosci 14, 219–245.
Green GM, Lyons L & Dickenson AH (1998). Alpha2-adrenoceptor antagonists enhance responses of dorsal horn neurones to formalin induced inflammation. Eur J Pharmacol 347, 201–204.
Guan Y, Terayama R, Dubner R & Ren K (2002). Plasticity in excitatory amino acid receptor-mediated descending pain modulation after inflammation. J Pharmacol Exp Ther 300, 513–520.
Harris J & Clarke RW (2002). Organisation of sensitisation of hind limb withdrawal reflexes from acute noxious stimuli in the rabbit. J Physiol 546, 251–265.
Horiguchi N, Ago Y, Hasebe S, Higashino K, Asada K, Kita Y, Takuma K & Matsuda T (2013). Isolation rearing reduces mechanical allodynia in a mouse model of chronic inflammatory pain. Pharmacol Biochem Behav 113, 46–52.
Howorth PW, Thornton SR, O’Brien V, Smith WD, Nikiforova N, Teschemacher AG & Pickering AE (2009). Retrograde viral vector-mediated inhibition of pontospinal noradrenergic neurons causes hyperalgesia in rats. J Neurosci 29, 12855–12864.
Hughes SW, Hickey L, Hulse RP, Lumb BM & Pickering AE (2013). Endogenous analgesic action of the pontospinal noradrenergic system spatially restricts and

Differential descending control of inputs from hairy vs. glabrous skin
32
temporally delays the progression of neuropathic pain following tibial nerve injury. Pain 154, 1680–1690.
Hylden J, Thomas D, Iadarola M, Nahin R & Dubner R (1991). Spinal opioid analgesic effects are enhanced in a model of unilateral inflammation/hyperalgesia: possible involvement of noradrenergic mechanisms. Eur J Pharmacol 194, 135–143.
Iadarola M, Brady L, Draisci G & Dubner R (1988). Enhancement of dynorphin gene expression in spinal cord following experimental inflammation: stimulus specificity, behavioral parameters and opioid receptor binding. Pain 35, 313–326.
Ikeda H, Heinke B, Ruscheweyh R & Sandkühler J (2003). Synaptic plasticity in spinal lamina I projection neurons that mediate hyperalgesia. Science 299, 1237–1240.
Jeong H-J, Mitchell V a & Vaughan CW (2012). Role of 5-HT1 receptor subtypes in the modulation of pain and synaptic transmission in rat spinal superficial dorsal horn. Br J Pharmacol 165, 1956–1965.
Kauppila T, Kontinen VK & Pertovaara A (1998). Influence of spinalization on spinal withdrawal reflex responses varies depending on the submodality of the test stimulus and the experimental pathophysiological condition in the rat. Brain Res 797, 234–242.
Kocher L, Anton F, Reeh P & Handwerker H (1987). The effect of carrageenan-induced inflammation on the sensitivity of unmyelinated skin nociceptors in the rat. Pain 29, 363–373.
Koerber HR, McIlwrath SL, Lawson JJ, Malin SA, Anderson CE, Jankowski MP & Davis BM (2010). Cutaneous C-polymodal fibers lacking TRPV1 are sensitized to heat following inflammation, but fail to drive heat hyperalgesia in the absence of TPV1 containing C-heat fibers. Mol Pain 6, 58.
Latremoliere A & Woolf CJ (2009). Central sensitization: a generator of pain hypersensitivity by central neural plasticity. J Pain 10, 895–926.
Leem JW, Willis WD & Chung JM (1993). Cutaneous sensory receptors in the rat foot. J Neurophysiol 69, 1684–1699.
Leith JL, Wilson AW, Donaldson LF & Lumb BM (2007). Cyclooxygenase-1-derived prostaglandins in the periaqueductal gray differentially control C- versus A-fiber-evoked spinal nociception. J Neurosci 27, 11296–11305.
Liu Z-Y, Zhuang D-B, Lunderberg T & Yu L-C (2002). Involvement of 5-hydroxytryptamine(1A) receptors in the descending anti-nociceptive pathway from periaqueductal gray to the spinal dorsal horn in intact rats, rats with nerve injury and rats with inflammation. Neuroscience 112, 399–407.
Lynn B & Carpenter S (1982). Primary afferent units from the hairy skin of the rat hind limb. Brain Res 238, 29–43.

Differential descending control of inputs from hairy vs. glabrous skin
33
Magerl W, Fuchs PN, Meyer RA & Treede RD (2001). Roles of capsaicin-insensitive nociceptors in cutaneous pain and secondary hyperalgesia. Brain 124, 1754–1764.
Mantyh PW (1983). Connections of midbrain periaqueductal gray in the monkey. II. Descending efferent projections. J Neurophysiol 49, 582–594.
Mantyh PW & Hunt SP (2004). Setting the tone: superficial dorsal horn projection neurons regulate pain sensitivity. Trends Neurosci 27, 582–584.
McMullan S & Lumb BM (2006a). Spinal dorsal horn neuronal responses to myelinated versus unmyelinated heat nociceptors and their modulation by activation of the periaqueductal grey in the rat. J Physiol 576, 547–556.
McMullan S & Lumb BM (2006b). Midbrain control of spinal nociception discriminates between responses evoked by myelinated and unmyelinated heat nociceptors in the rat. Pain 124, 59–68.
McMullan S & Lumb BM (2006c). Spinal dorsal horn neuronal responses to myelinated versus unmyelinated heat nociceptors and their modulation by activation of the periaqueductal grey in the rat. J Physiol 576, 547–556.
McMullan S, Simpson DA & Lumb BM (2004). A reliable method for the preferential activation of C- or A-fibre heat nociceptors. J Neurosci Methods 138, 133–139.
Meyer RA & Campbell JN (1981). Myelinated nociceptive afferents account for the hyperalgesia that follows a burn to the hand. Science 213, 1527–1529.
Meyer RA, Ringkamp M, Campbell JN & Raja SN (2005). Neural mechanisms of hyperalgesia after tissue injury. Johns Hopkins APL Tech Dig 26, 56–66.
Miki K, Zhou QQ, Guo W, Guan Y, Terayama R, Dubner R & Ren K (2002). Changes in gene expression and neuronal phenotype in brain stem pain modulatory circuitry after inflammation. J Neurophysiol 87, 750–760.
Millan MJ (2002). Descending control of pain. Prog Neurobiol 66, 355–474.
Millan MJ, Newman-Tancredi A, Audinot V, Cussac D, Lejeune F, Nicolas J, Coge F, Galizzi J, Boutin J, Rivet J, A D & Gobert A (2000). Agonist and antagonist actions of yohimbine as compared to fluparoxan at alpha(2)-adrenergic receptors (AR)s, serotonin (5-HT)(1A), 5-HT(1B), 5-HT(1D) and dopamine D(2) and D(3) receptors. Significance for the modulation of frontocortical monoaminergic transmission and depressive states. Synapse 35, 79–95.
Oliva P, Berrino L, de Novellis V, Palazzo E, Marabese I, Siniscalco D, Scafuro M, Mariani L, Rossi F & Maione S (2006). Role of periaqueductal grey prostaglandin receptors in formalin-induced hyperalgesia. Eur J Pharmacol 530, 40–47.
Ossipov MH, Dussor GO & Porreca F (2010). Central modulation of pain. J Clin Invest 120, 3779–3787.

Differential descending control of inputs from hairy vs. glabrous skin
34
Ossipov MH, Suarez LJ & Spaulding TC (1989). Antinociceptive interactions between alpha , -adrenergic and opiate agonists at the spinal level in rodents. Anesth Analg 68, 194–200.
Paalzow G & Paalzow L (1983). Yohimbine both increases and decreases nociceptive thresholds in rats: evaluation of the dose-response relationship. Naunyn Schmiedebergs Arch Pharmacol 322, 193–197.
Palazzo E, Guida F, Gatta L, Luongo L, Boccella S, Bellini G, Marabese I, de Novellis V, Rossi F & Maione S (2011). EP1 receptor within the ventrolateral periaqueductal grey controls thermonociception and rostral ventromedial medulla cell activity in healthy and neuropathic rat. Mol Pain 7, 82.
Pertovaara A (2006). Noradrenergic pain modulation. Prog Neurobiol 80, 53–83.
Pertovaara A (2013). The noradrenergic pain regulation system: A potential target for pain therapy. Eur J Pharmacol 716, 2–7.
Provitera V, Nolano M, Pagano A, Caporaso G, Stancanelli A & Santoro L (2007). Myelinated nerve endings in human skin. Muscle Nerve 35, 767–775.
Przewłocki R & Przewłocka B (2001). Opioids in chronic pain. Eur J Pharmacol 429, 79–91.
Reed RK & Rubin K (2010). Transcapillary exchange: role and importance of the interstitial fluid pressure and the extracellular matrix. Cardiovasc Res 87, 211–217.
Ren K & Dubner R (1996). Enhanced descending modulation of nociception in rats with persistent hindpaw inflammation. J Neurophysiol 76, 3025–3037.
Ren K & Dubner R (1999). Inflammatory models of pain and hyperalgesia. ILAR J 40, 111–118.
Sandrini G, Serrao M, Rossi P, Romaniello A, Cruccu G & Willer JC (2005). The lower limb flexion reflex in humans. Prog Neurobiol 77, 353–395.
Schouenborg J, Weng H & Holmberg H (1994). Modular organization of spinal nociceptive reflexes�: A new hypothesis. Physiology 9, 261–265.
Sivilotti LG, Thompson SWN & Woolf CJ (1993). Rate of rise of the cumulative depolarization evoked by repetitive stimulation of small-caliber afferents is a predictor of action potential windup in rat spinal neurons in vitro. J Neurophysiol 69, 1621–1631.
Soignier RD, Taylor BK, Baiamonte BA, Lee FA, Paul D & Gould HJ (2011). Measurement of CFA-induced hyperalgesia and morphine-induced analgesia in rats: dorsal vs plantar mechanical stimulation of the hindpaw. Pain Med 12, 451–458.
Stanfa L & Dickenson A (1995). Spinal opioid systems in inflammation. Inflamm Res 44, 231–241.

Differential descending control of inputs from hairy vs. glabrous skin
35
Størkson R V, Kjørsvik A, Tjølsen A & Hole K (1996). Lumbar catheterization of the spinal subarachnoid space in the rat. J Neurosci Methods 65, 167–172.
Suzuki R, Morcuende S, Webber M, Hunt SP & Dickenson AH (2002). Superficial NK1-expressing neurons control spinal excitability through activation of descending pathways. Nat Neurosci 5, 1319–1326.
Takano Y & Yaksh TL (1992). Characterization of the pharmacology of intrathecally administered alpha-2 agonists and antagonists in rats. J Pharmacol Exp Ther 261, 764–772.
Terayama R, Dubner R & Ren K (2002). The roles of NMDA receptor activation and nucleus reticularis gigantocellularis in the time-dependent changes in descending inhibition after inflammation. Pain 97, 171–181.
Tsuruoka M, Matsutani K & Inoue T (2003). Coeruleospinal inhibition of nociceptive processing in the dorsal horn during unilateral hindpaw inflammation in the rat. Pain 104, 353–361.
Tsuruoka M, Tamaki J, Maeda M, Hayashi B & Inoue T (2012). Biological implications of coeruleospinal inhibition of nociceptive processing in the spinal cord. Front Integr Neurosci 6, 87.
Tsuruoka M & Willis WD (1996). Descending modulation from the region of the locus coeruleus on nociceptive sensitivity in a rat model of inflammatory hyperalgesia. Brain Res 743, 86–92.
Tyce GM & Yaksht TL (1981). Monoamine release from cat spinal cord by somatic stimuli: an intrinsic modulatory system. J Physiol 314, 513–529.
Vanegas H & Schaible HG (2004). Descending control of persistent pain: inhibitory or facilitatory? Brain Res Rev 46, 295–309.
Wang W, Wu S-X, Wang Y-Y, Liu X-Y & Li Y-Q (2003). 5-Hydroxytryptamine1A receptor is involved in the bee venom induced inflammatory pain. Pain 106, 135–142.
Westlund KN, Bowker RM, Ziegler MG CJ (1983). Noradrenergic projections to the spinal cord of the rat. Brain Res 263, 15–31.
Wiesenfeld-Hallin Z (1995). Neuropeptides and spinal cord reflexes. Prog Brain Res 104, 271–280.
Woolf CJ & Wall PD (1986). Relative effectiveness of C primary afferent fibers of different origins in evoking a prolonged facilitation of the flexor reflex in the rat. J Neurosci 6, 1433–1442.
Yeomans DC & Proudfit HK (1996b). Nociceptive responses to high and low rates of noxious cutaneous heating are mediated by different nociceptors in the rat: Electrophysiological evidence. Pain 68, 141–150.

Differential descending control of inputs from hairy vs. glabrous skin
36
Yeomans DC & Proudfit HK (1996a). Nociceptive responses to high and low rates of noxious cutaneous heating are mediated by different nociceptors in the rat: electrophysiological evidence. Pain 68, 141–150.
Yoshimura M & Furue H (2006). Mechanisms for the anti-nociceptive actions of the descending noradrenergic and serotonergic systems in the spinal cord. J Pharmacol Sci 101, 107–117.
You H-J, Lei J, Sui M-Y, Huang L, Tan Y-X, Tjølsen A & Arendt-Nielsen L (2010). Endogenous descending modulation: spatiotemporal effect of dynamic imbalance between descending facilitation and inhibition of nociception. J Physiol 588, 4177–4188.
Yu Z, Aihui L, Jiajia X, Lixing L, Ke R, Brian B, Ming T & Zhang R-X (2011). Involvement of spinal serotonin receptors in electroacupuncture anti-hyperalgesia in an inflammatory pain rat model. Neurochem Res 36, 1785–1792.
Zhang Y, Yang Z, Gao X & Wu G (2001). The role of 5-hydroxytryptamine1A and 5-hydroxytryptamine1B receptors in modulating spinal nociceptive transmission in normal and carrageenan-injected rats. Pain 92, 201–211.
Ziegler EA, Magerl W, Meyer RA & Treede RD (1999). Secondary hyperalgesia to punctate mechanical stimuli. Central sensitization to A-fibre nociceptor input. Brain 122 (Pt 1), 2245–2257.

Differential descending control of inputs from hairy vs. glabrous skin
37
Figure legends
Figure 1: A single subcutaneous injection of CFA into the hind-paw dorsum
leads to a primary mechanical but not thermal hyperalgesia.
A. Delivery of CFA (50µl) into the hind-paw dorsum produced significant oedema
(one-way RM ANOVA; p<0.0001, F(4,3)=33.54, Bonferroni’s post-test *p<0.001
compared to baseline, n=4).
B. CFA-induced inflammation was associated with a primary mechanical
hyperalgesia (one-way RM ANOVA; p<0.001, F(3,3)=7.13, Bonferroni’s post-test
*p<0.05 compared to baseline, n=4).
C. CFA-induced inflammation did not however result in thermal hyperalgesia (n=4).
D. In contrast, a 50µl s.c. injection of CFA into the plantar surface of the hind-paw
produced a persistent primary thermal hyperalgesia (one-way RM ANOVA; p<0.05,
F(3,2)=11.88, Bonferroni’s post-test, *p<0.05 compared to baseline, n=3).
Figure 2: A single subcutaneous injection of CFA into the hind-paw dorsum
produces a transient sensitisation of withdrawal reflexes to A- and C-
nociceptor activation.
A. CFA-induced inflammation of the dorsal hind-paw resulted in a transient reduction
in the withdrawal threshold to A-nociceptor stimulation that resolved fully by 3 hours
after CFA injection (one-way RM ANOVA, p<0.0001, F(3,4)=21.22, Bonferroni’s post-
test ***p<0.001 compared to baseline, n=5).
B. Responses to C-nociceptor activation showed that inflammation resulted in an
even more transient response, resolving after 2 hours (one-way RM ANOVA,
P<0.0001, F(3,4)=20.39, Bonferroni’s post-test *p<0.001 compared to baseline, n=5).
An equivalent injection of vehicle (mineral oil) had no significant effect on withdrawal
thresholds to A- or C-nociceptor activation (Mineral oil group, A&B, Friedman test,
p>0.05, n=3)

Differential descending control of inputs from hairy vs. glabrous skin
38
C. A larger dose of CFA (100µg/100µl) resulted in a more rapid resolution of
sensitisation to A-nociceptor activation, causing a more rapid return to baseline (two-
way RM ANOVA; main dose effect: p=0.08, F(3,1)=3.98, Bonferroni’s post-test
*p<0.05 between groups, n=5 for both groups).
D. A larger dose of CFA (100µg) resulted in a more rapid resolution of sensitisation
to C-nociceptor activation so that withdrawal thresholds were significantly higher
compared to a 50µg dose at the one hour time point (two-way RM ANOVA; Main
dose effect: p=0.0022, F(3,1)=19.53, Bonferroni’s post-test *** p<0.001 between
groups, n=5 for both groups).
Figure 3: Sensitisation resulting from acute inflammation of the plantar
hindpaw is unaffected by intrathecal yohimbine.
Acute CFA-induced inflammation of the plantar hind-paw resulted in a reduction in
thermal withdrawal threshold that persisted for the 3 hours tested (one-way RM
ANOVA; p<0.0001, F(3,4)=19.30, Bonferroni’s post-test ***p<0.001, **p<0.01
compared to baseline). This was unaffected by intrathecal yohimbine at 3h following
CFA (n=5 CFA, n=4 CFA+yoh).
Figure 4: Paw swelling is different following CFA injection into dorsal or
plantar hindpaw.
Injection of 50µg CFA resulted in greater swelling in the plantar compared to the
dorsal surface of the hindpaw. Injection of a larger CFA concentration (100µg) in the
dorsal surface did not increase the swelling above that caused by 50µg. (one way
ANOVA, p<0.0001 F(12, 70)=59.13, Bonferroni post-hoc tests *p<0.05 plantar c.f.
both other groups, data shown as mean±SD).

Differential descending control of inputs from hairy vs. glabrous skin
39
Figure 5: Blockade of spinal alpha 2 adrenoceptors reveals a rapid inhibitory
control on withdrawal reflexes to A- and C-nociceptor activation during acute
inflammation.
A. The loss of acute sensitisation to A-nociceptor stimulation 3 hours after CFA
injection was partially reversed by the intrathecal injection of the noradrenergic alpha
2 receptor antagonist yohimbine (Mann-Whitney U, *p<0.05, n=7 CFA (3h), n=5 yoh).
B. The loss of acute sensitisation to C-nociceptor stimulation 3 hours after CFA
injection was reversed by yohimbine (Mann-Whitney U, *p<0.05, n=8 CFA (3h), n=5
yoh).
C. Systemic administration of the pan-opioid antagonist, naloxone, had no effect on
either A-nociceptor evoked withdrawals (CFA n=5, CFA+nal n=6) or on D. C-
nociceptor evoked withdrawals (CFA n=5, CFA+nal n=6).