
Metabolizm komórkowy
i sposoby uzyskiwania energii
Metabolizm – całokształt reakcji chemicznych i związanych z
nimi przemian energii zachodzący w komórkach. Podstawa
wszelakich zjawisk biologicznych.


Metabolizm komórkowy
anabolizm katabolizm
procesy biosyntezy złożonych
cząsteczek, wymagające
dostarczenia energii
procesy rozkładu dużych
cząsteczek organicznych,
dostarczające energię
Energia pozyskana z rozkładu tylko częściowo zamieniana jest w
pracę. Jest to tzw. energia swobodna (reszt rozpraszana jest w
postaci ciepła.
Energia swobodna wykorzystywana jest w syntezie, transporcie
czy ruchu cząsteczek.

Sposoby wykorzystywania
energii
aktywny transport substancji
przez błony
wzrost
rozmnażanie
ruch
odpowiedź na bodźce
utrzymanie homeostazy
Sposoby magazynowania
energii
ATP - adenozynotrifosforan
FAD - dinukleotyd flawinoadeninowy
NAD - dinukleotyd
nikotynoamidoadeninowy
NADP - fosforan dinukleotydu
nikotynamidoadeninowego
pompa protonowa
Szlaki metaboliczne

Podział bakterii ze względu na źródła energii
i węgla
Chemoorganotrofy (heterotrofy)
źródło węgla i energii: substraty organiczne np. cukry, kwasy
tłuszczowe, alkohole
Chemolitotrofy (autotrofy)
źródło energii: substraty nieorganiczne np. S2-, So, H2, Fe2+,
NH3
źródło węgla: utlenione związki węgla np. CO2, węglany
Fototrofy (autotrofy lub heterotrofy)
źródło energii: światło
źródło węgla – CO2 lub substraty organiczne

ATP – adenozynotrójfosforan
(adenosine triphosphate)
ADP – adenozynodwufosforan
(adenosine diphosphate)
NAD+ - dwunukleotyd nikotynoamidoadeninowy
(forma utleniona)
NADH – dwunukleotyd nikotynoamidoadeninowy
(forma zredukowana)

ADP - forma uniwersalnego komórkowego nośnika energii ATP w postaci
„rozładowanej”. Zawiera niewielką ilość energii do reakcji biochemicznych.
ATP- adenozynotrifosforan. Uniwersalny nośnik i przekaźnik energii w
komórkach. W typowej reakcji przekazania energii, krańcowa grupa
fosforanowa ATP przyłącza się do cząsteczki innej substancji, przenosząc z
sobą większość energii zawartej w wiązaniu chemicznym, którym była
pierwotnie połączona w cząsteczce ATP. W wyniku tej rakcji powstaje ADP, z
którego na ogół odtwarza się ATP.
NAD+ (NADH)- dinukleotyd amidoadeninowy. Koenzym wykorzystywany w
procesach metabolicznych w komórkach jako akceptor lub donor protonu.
NAD+ bierze udział w reakcjach katabolicznych, w tym w oddychaniu
komórkowym.
NADP+(NADPH) –fosforan dinukleotydu nikotynoamidoadeninowego.
Koenzym wykorzystywany w komórkach jako akceptor lub donor protonu.
Bierze udział w reakcjach anabolicznych , w tym w fotosyntezie.

Sposoby uzyskiwania energii
fotosynteza - energia światła absorbowana przez barwniki
(bakteriochlorofile i karotenoidy) zostaje przetworzona w siłę
protonomotoryczną lub energię chemiczną,
fermentacja – substrat jest metabolizowany bez udziału
zewnętrznego (egzogennego) czynnika utleniającego.W wyniku
tego procesu powstają proste związki oraz energia,
oddychanie tlenowe (respiracja) – substrat organiczny jest
przekształcany z udziałem zewnętrznego czynnika utleniającego:
tlenu, do wody i dwutlenku węgla. Energia wiązań zostaje
przekształcona w ATP,
oddychanie beztlenowe – podobnie jak w oddychaniu tlenowym
następuje utlenienie substratu organicznego lecz rolę zewnętrznych
akceptorów elektronów pełnią azotany, siarczany lub fumarany.

Fotosystemy
Budowa - kompleks antenowy zbierający światło, zawierający setki
cząsteczek pigmentów absorbujących światło (karoteny, chlorofil lub
bakteriochlorofil) oraz centrum aktywne, w którym znajdują się dwie
specjalne cząsteczki chlorofilu.
Rola - zbieranie energii świetlnej i przekształcenie jej w energię
chemiczną.
LHC I i LHC II – light harvesting complex
Fotosynteza
Fotosynteza – seria złożonych reakcji przekształcających energię
świetlną w energię chemiczną. Process odbywa się w organellach
zwanych chloroplastami
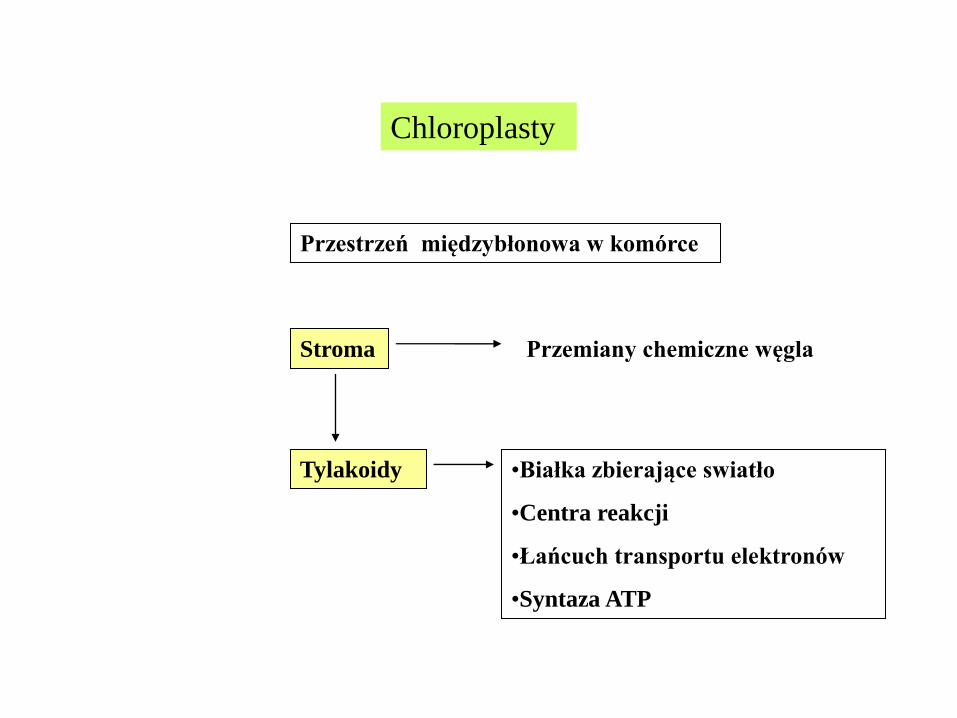
Chloroplasty
Przestrzeń międzybłonowa w komórce
Stroma Przemiany chemiczne węgla
Tylakoidy •Białka zbierające swiatło
•Centra reakcji
•Łańcuch transportu elektronów
•Syntaza ATP


Centrum reakcji i kompleksy zbierające światło, widok prostopadły do
płaszczyzny błony komórkowej (góra) i z boku (dół). Kolor niebieski – LH2,
kolor zielony – LH1, kolor żółty – centrum reakcji.

światło
światło
P680
Fotosystem II
h
Plastochinon
Kompleks
cytochromów
Plastocyjanina
e-
P700
Fotosystem I e
-
e-
e-
Ferredoksyna
NADP+
h e
-
e-
0
+400
2H+
(ze środowiska)
NADPH
H2O ½ O2 + 2H+
e-
ATP
ADP+Pi
-600
+1000
Pote
ncj
ał o
ksy
dac
yjn
o-r
edukcy
jny (
mV
)
P680*
P700*
Fotosynteza oksygenowa ( z wytworzeniem tlenu) (w algach i cyjanobakteriach fotosystemy współdziałają ze sobą)

Światło powoduje wyrzucenie z chlorofilu w fotosystemie PSII
elektronu o dużej energii, który przesuwa wzdłuż łańcucha
transportu elektronów. Powoduje to powstanie siły
protonomotorycznej po obu stronach błony. Energia ta następnie
zostaje wykorzystana do syntezy ATP (proces fotofosforylacji).
Elektrony z fotosystemu PSI redukują NADP. Utleniony chlorofil w
fotosystemie PSII pobiera elektrony pochodzące z rozkładu wody.
Towarzyszy temu wydzielenie się cząsteczki tlenu.
W reakcjach niezależnych od światła (reakcje ciemne) energia
pochodząca z fotosyntezy wykorzystana jest do syntezy związków
węgla z CO2.


Widmo błony bakterii R. sphaeroides z zaznaczonymi chromoforami: Car –
karotenoidy, LH1 – kompleks zbierający światło , RC – centrum reakcji,
Bpheo –bakteriofeofityna, Bchl – bakteriochlorofil..
Kim, E.-J., et al., International Journal of Hydrogen Energy, 2003. 31: p. 531-538
Fotosynteza anoksygenowa (bez wytworzenia tlenu)
w bakteriach fotosyntetyzujących
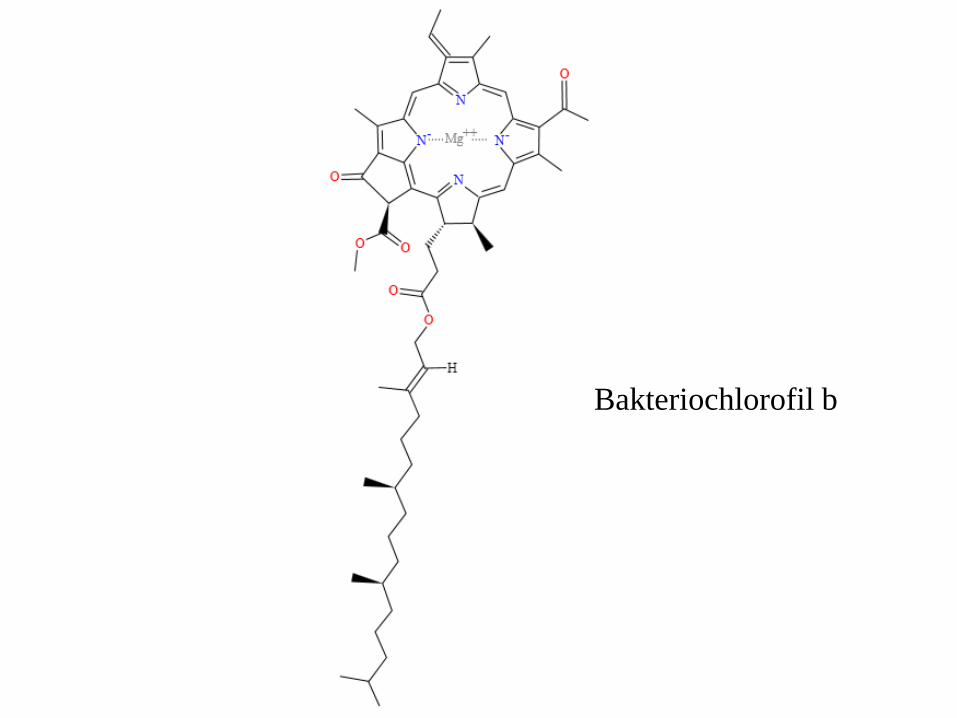
Bakteriochlorofil b
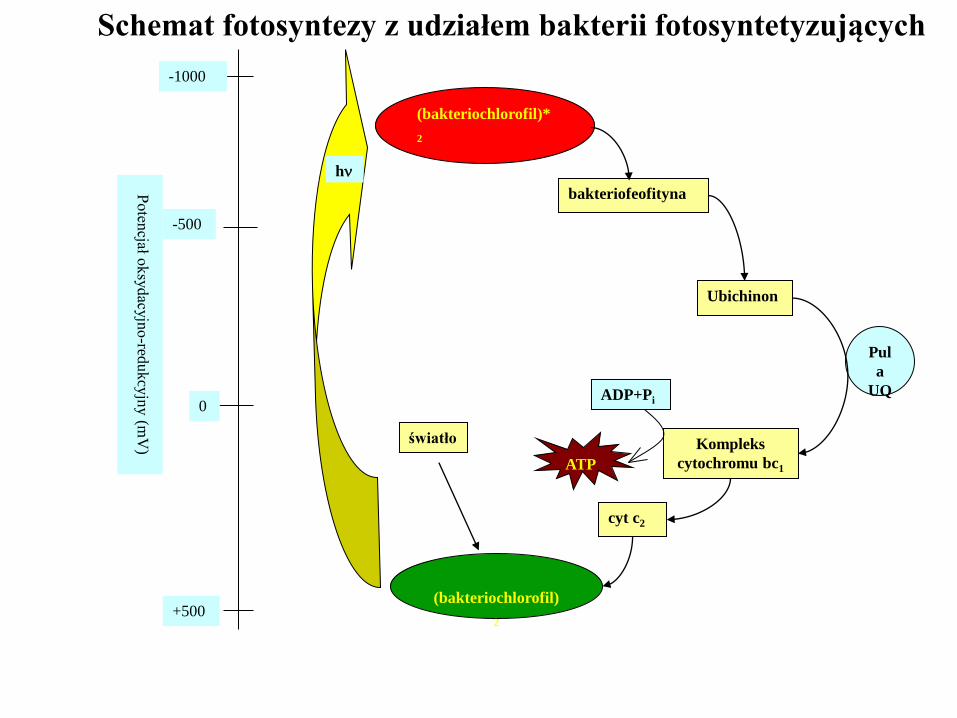
(bakteriochlorofil)
2
bakteriofeofityna
Kompleks
cytochromu bc1
Ubichinon
h
(bakteriochlorofil)*
2
Pul
a
UQ
cyt c2
-1000
-500
0
+500
Poten
cjał oksy
dacy
jno
-redukcy
jny (m
V)
ADP+Pi
ATP
światło
Schemat fotosyntezy z udziałem bakterii fotosyntetyzujących

atp

nikotynazmide
Budowa NAD+ i NADH

Rodzaje fermentacji etanolowa
mlekowa
propionowa
mrówczanowa
masłowa
metanowa
mieszana
Fermentacja

Fermentacja alkoholowa
C6H12O6 2C2H5OH + 2CO2 + 40 kcal
ATP + H2O ADP + Pi – 7.3 kcal
7.3 kcal 1ATP
14.6 kcal 2ATP
40 -14.6 25.4 kcal (ciepło)
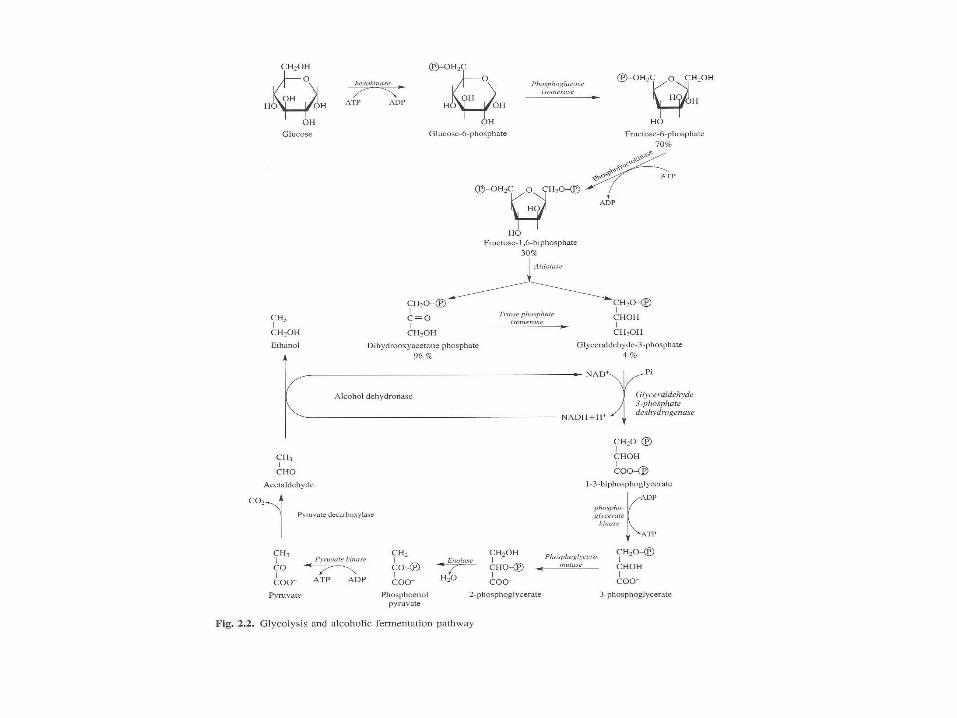
Gli
kol
iza
duz
a

Glikoliza gora

Glikolza dól
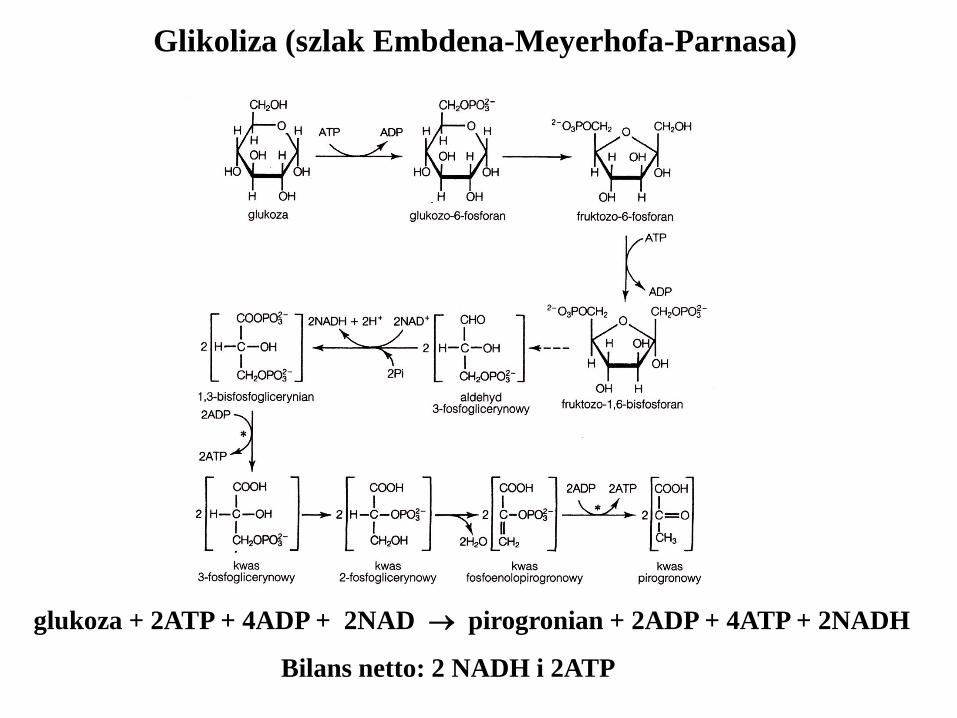
Glikoliza (szlak Embdena-Meyerhofa-Parnasa)
glukoza + 2ATP + 4ADP + 2NAD pirogronian + 2ADP + 4ATP + 2NADH
Bilans netto: 2 NADH i 2ATP

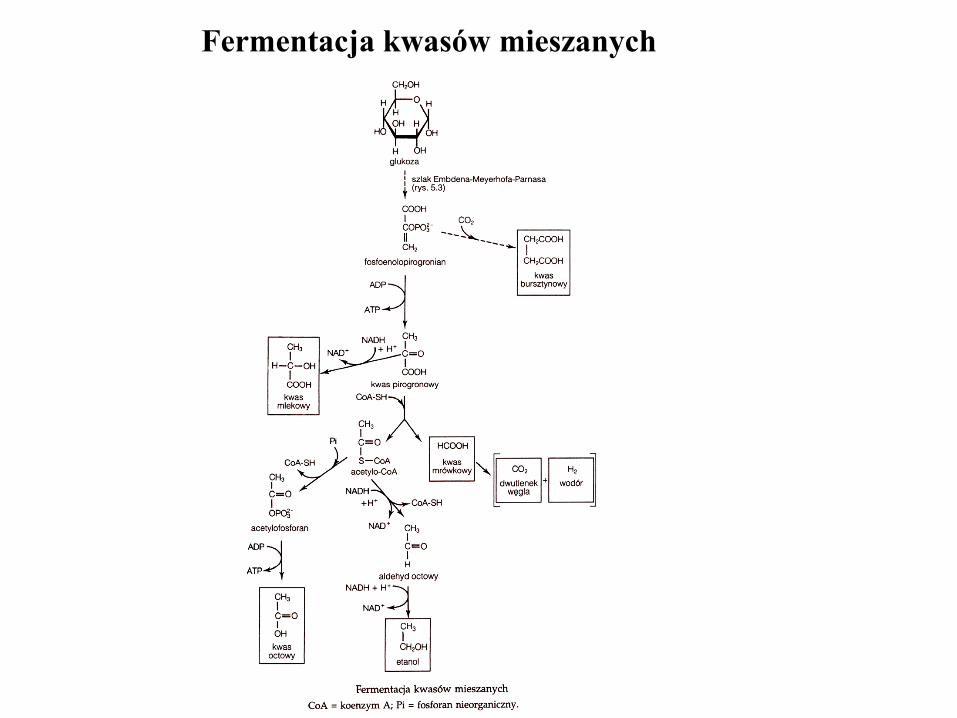
Fermentacja kwasów mieszanych

Oddychanie tlenowe
W obecności tlenu pirogronian uzyskany w procesie glikolizy jest
przekształcany w acetylo-CoA i CO2. Następnie grupa acetylowa
w acetylo-CoA ulega przekształceniu do CO2 i H2O w cyklu kwasu
cytrynowego (cykl kwasów trójkarboksylowych). Proces ten dostarcza
energii oraz prekursorów koniecznych do biosyntezy.
W wyniku jednego cyklu uwalniają się dwie cząsteczki CO2,
powstaje jedna cząsteczka ATP, trzy cząsteczki NADH lub NADPH
i jedna FADH (dinukleotyd flawino-adeninowy- akceptor i donor
wodoru w komórkowych reakcjach metabolicznych, także w procesie
fotosyntezy)
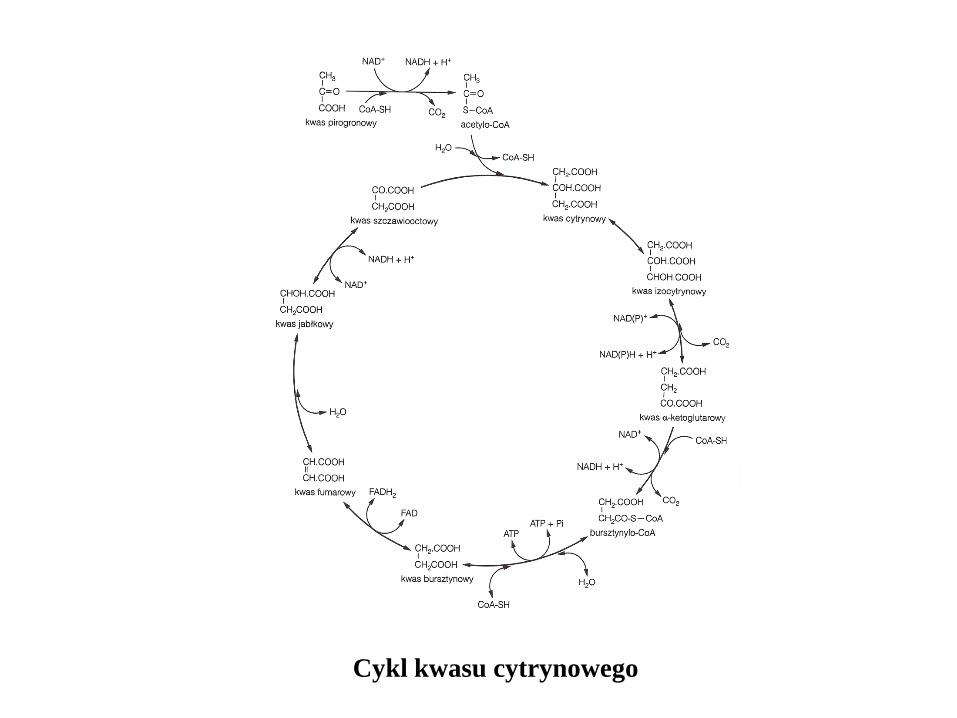
Cykl kwasu cytrynowego

Łańcuch transportu elektronów
(łańcuch oddechowy)
Powstający NADH (potencjał redoks –320 mV) przekazuje elektrony
do tlenu (potencjał redoks +820 mV) w kilku etapach, wykorzystując
nośniki elektronów o coraz wyższym potencjale redoks. Umożliwia to
stopniowe uwalnianie energii zmagazynowanej w elektronach NADH.
Ten łańcuch wyspecjalizowanych cząsteczek nazwano łańcuchem
transportu elektronów lub łańcuchem oddechowym. W bakteriach
zlokalizowany jest on w błonie cytoplazmatycznej.
Uwolniona energia jest wykorzystywana przez komórkę do
pompowania protonów poprzez błonę cytoplazmatyczną na zewnątrz.
Tendencja protonów do powrotu na wewnętrzną stronę błony
nazywana jest siłą protonomotoryczną (PMF). Siła ta może być
wykorzystana do syntezy ATP (fosforylacja oksydatywna).

Składniki łańcucha transportu elektronów
Flawoproteiny np. ferredoksyny - białka żelazowo-siarczkowe
przenoszące tylko jeden elektron, silne reduktory o bardzo niskim
potencjale redoks,
chinony - zwiazki aromatyczne ulegające odwracalnej redukcji,
mogą pobierać jeden lub dwa elektrony, a z każdym przenoszonym
elektronem pobierają ze środowiska jeden proton, potencjał redoks
około +30 mV,
cytochromy - białka zawierające jedną lub więcej grup hemowych,
pobierają elektrony i przekazują je dalej poprzez naprzemienną
redukcję i utlenianie jonu żelaza, potencjał redoks cytochromu c wynosi
+230 mV.
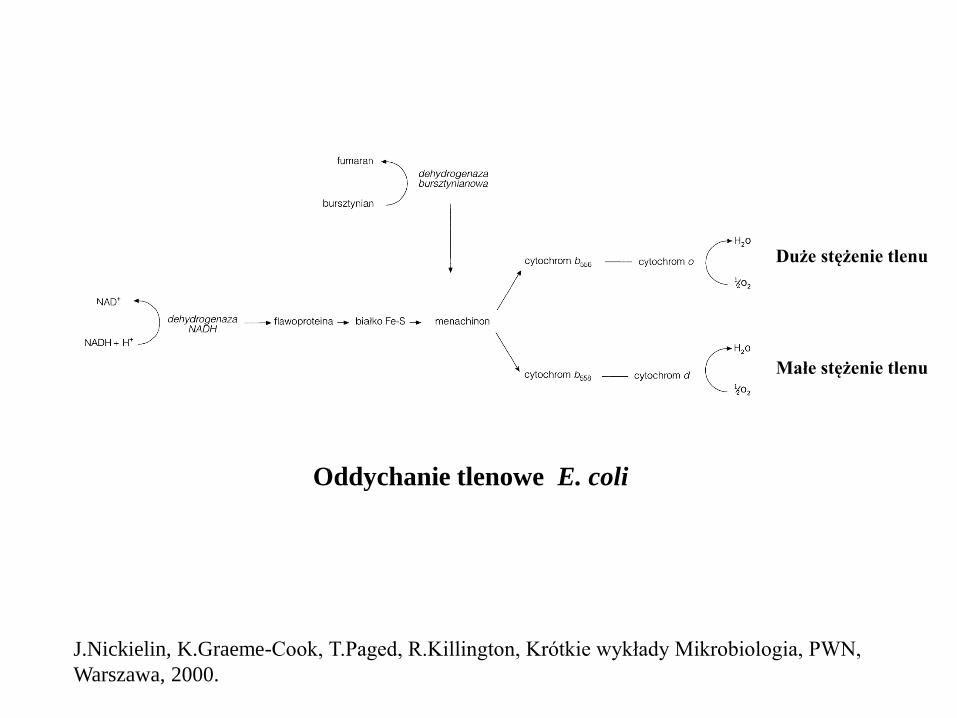
Oddychanie tlenowe E. coli
Małe stężenie tlenu
Duże stężenie tlenu
J.Nickielin, K.Graeme-Cook, T.Paged, R.Killington, Krótkie wykłady Mikrobiologia, PWN,
Warszawa, 2000.

Beztlenowe oddychanie E. Coli z wykorzystaniem azotanu
J.Nickielin, K.Graeme-Cook, T.Paged, R.Killington, Krótkie wykłady Mikrobiologia, PWN,
Warszawa, 2000.

Wydajność tworzenia ATP z glukozy
Droga kataboliczna mole ATP/mol glukozy
Oddychanie tlenowe 34
glikoliza 2
fermentacja 2-3

Wydajność tworzenia ATP z glukozy
Droga kataboliczna mole ATP/mol glukozy
Oddychanie tlenowe 34
glikoliza 2
fermentacja 2-3







