
Mathematical modeling applied to neuroscience
Amitabha BoseDepartment of Mathematical Sciences
New Jersey Institute of Technology
Hunter College High School 2013

Mathematics and the Natural World• “The Unreasonable Effectiveness of Mathematics in
the Natural Sciences” published in 1960 by Nobel Prize winning Physicist Eugene Wigner
• Gives numerous examples from physics like Newton’s gravitational laws or Einstein’s theory of relativity
• “How can it be that mathematics, being after all a product of human thought which is independent of experience, is so admirably appropriate to the objects of reality?” — Albert Einstein

Mathematics and Life Sciences
• Wigner concludes with “A much more difficult and confusing situation would arise if we could, some day, establish a theory of the phenomena of consciousness, or of biology, which would be as coherent and convincing as our present theories of the inanimate world.”
• “There is only one thing which is more unreasonable than the unreasonable effectiveness of mathematics in physics, and this is the unreasonable ineffectiveness of mathematics in biology.” — Israel Gelfand

Mathematical Physiology• Many successful applications of mathematics to “solving”
biological problems• Derivation of the Hodgkin-Huxley equations (1952)• FitzHugh-Nagumo equation (1962)• Understanding cardiac dynamics (Peskin 1970’s on, Keener 1980’s
on)• Understanding dynamics of pancreatic beta cells associated with
diabetes (Miura 1970’s, Sherman 1980’s)• Neuroscience – Rinzel, Wilson-Cowan 1970’s on, Kopell,
Ermentrout 1980’s on, Terman 1990’s on• Great book by Art Winfree “The Geometry of Biological Time
(1980)• Other important texts by Jim Murray, Keener & Snyed,
Hoppensteadt & Peskin, Edelstein-Keshet

Outline
• Basics of neuroscience, circuit theory and differential equations
• Numerous applications of math & neuroscience
Q. What is the appropriate level of detail for modeling?Q. Is there any fun mathematics to be done?
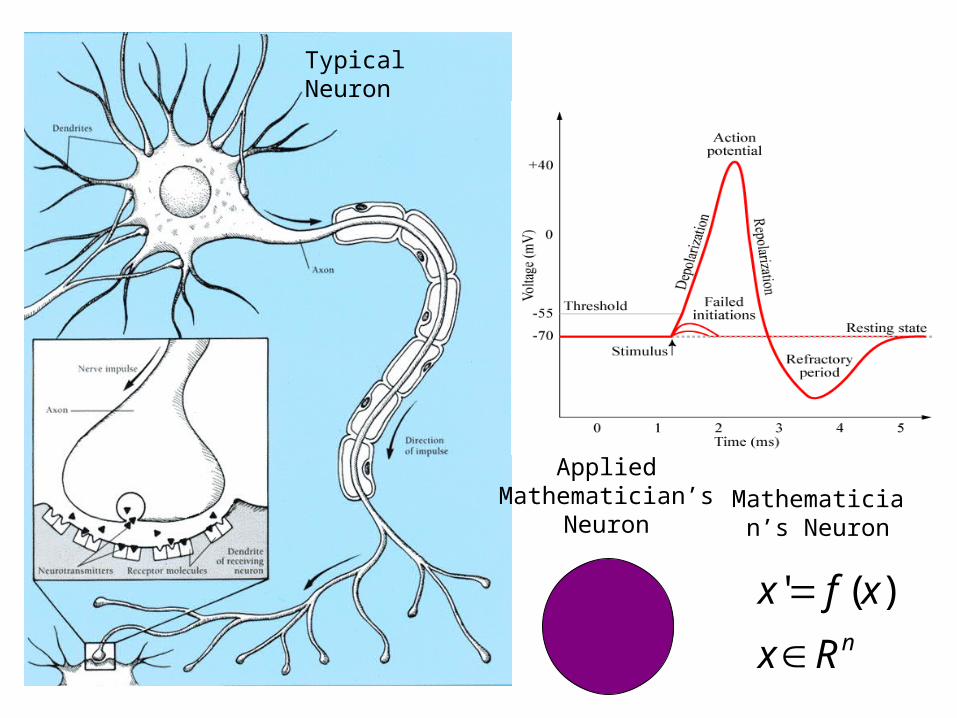
Typical Neuron
Applied Mathematician’s
NeuronMathematician’s
Neuron
nRx
xfx
)('

What is a derivative?
• A mathematical object that measures the rate of change of a quantity
• Ex: the slope of a line measures the rate of change of the “rise” to the “run”.
• Ex: think of the formula s = v·t to measure distance s. The change in distance per unit time is the velocity. As the unit of time is made smaller, this rate becomes the derivative ds/dt = v.

What is a differential equation?
• An equation composed of derivatives and functions of the dependent and independent variables.
• Ex: Damped Pendulum
• Differential equations can be “solved” by analytically or numerically integrating them for a given set of initial conditions.
2
2sin 0
d d
dt dt

Modeling a neuron as an RC circuit
• Membrane separates charge
• Ions flow through channels causing voltage changes

1,i m
m
m m
V I R gR
dVC I gV
dt
C ,
C
m c
m c
dQQ V I
dtdV
Idt
,m c iI I I
current
charge
voltage
resistance
capacitance
I
Q
V
R
C

Hodgkin-Huxley type equations,( , )
( )
( )
( )
( )
app ion i i
i i
m
i i
h
dvC I I v m h
dtdm m v m
dt v
dh h v h
dt v
( ) ( )[ ]
( ) ( )[ ]
[ ]
Na
K
p qNa NaNa Na
r sK KK K
L L L
I g m v h v v V
I g m v h v v V
I g v V

Why are action potentials important?
• Action potentials are measurable events• The timings or firing rate of action potentials can
encode information
- orientation selectivity in visual cortex- coincidence detection for sound localization- place cells in hippocampus
• Neurons can communicate with one another using action potentials via synapses or gap junctions.

Crustacean Pyloric Rhythm (CPG)
PD
LP PY
Bean, Nature Rev. Neuro. 2007
Nadim et al, 2000’s

Visual Cortex• Neurons fire at preferential orientations
Hubel and Wiesel 1962 and related work
Many mathematical models for describing this phenomenon

The brain is good at detecting edges, but not so good at other things…

Count the black dots
What kind of mathematical model can explain this?

Perceptual Bistability
Rinzel and collaborators are developing mathematical models to explain bistability

Auditory CortexCoincidence detection
• Neurons have higher firing rate when they get coincidental input from left and right ears
• Owls use this to locate prey and prey to locate owls
• Jeffress delay line model for barn owls (1948)

Model for coincidence detection(Cook et al, 2003, Grande & Spain 2004)
O1 O2
NL
P
t
t
P
O1 O2
Calculate NL firing rate as a function of phase independent of frequency of O1 and O2

Place cells• Pyramidal cells in hippocampus fire only when animal is in a
specific, known location (transient & stable)• Uses visual cues to trigger memory recall • O’Keefe (1971)

Model for place cell firing(Bose, Booth,Recce 2000)
PLACE FIELD
T P
I1 I2
InhibitionExcitation
P = Place cells
Model Predictions• Location within place field• Length of place field• Behavior in running wheel

Parkinson’s Disease• Believed to be a disorder of the Basal Ganglia• Results in tremor, uncontrollable motions,
inability to begin movement • No experimental consensus on what causes PD
or how it can be treated• Mathematicians have gotten involved with
experimentalist to figure out the underlying neural mechanisms
• Peter Tass’ group (Germany) • Terman, Rubin, Wilson (US)• Kopell, Wilson (US)

PD results when the output from Gpi/SNr becomes too synchronized
I
For severe cases, Deep Brain Stimulation has shown remarkable promise for alleviating PD symptons. We don’t know why DBS works?!

Rubin & Terman (2004) proposed that DBS targets STN
• Normal state: Irregular, no correlations in STN cells
• Parkinsonian state: Rhythmic, STN cells cluster

Modeling Sleep Rhythms
• Your brain is very active when you are asleep!• The function of sleep is still not totally
understood, but it is important for learning, for the immune system, for growth…
• REM sleep – Rapid Eye Movement sleep (dreaming)
• Mathematicians are creating models to help explain different electrical rhythms seen in different stages of sleep

Stages of sleep for humans
• we focus on the transitions between sleep/wake and REM/NREM sleep
• sleep patterns of rodents are similar, with more brief awakenings

Model (Kumar, Bose, Mallick 2012)
CRF
R-ON
GABA
R-OFF
MRF
POAH
HOM
CIRC
GABASNr
ORX
excitationinhibition
Wakefulness associated areasMRF : Mid brain reticular formationORX : Lateral hypothalamic
orexinergic neurons.
Sleep associated areasPOAH : Preoptic anterior hypothalamus CRF : Caudal reticular formation
REM sleep associated areasR-ON : LDT/ PPT cholinergic neuronsR-OFF : LC Noradrenergic neuronsGABA : GABAergic inter neurons in LC
Other inputsHom : Homeostatic sleep driveCirc : Circadian clock from SCNGABASNr : GABaergic neurons from
substantia niagra pars reticulata
Feed-forward control of one flip-flop circuit by another

Sleep/Wake Transitions
No REM activity in this trace
wakesleep wake sleepFast transitions controlled by POAH
Consistent with Szymusiak et al 1998
REM-ON
REM-OFF
POAH MRF

Transitions to REM sleep
GABA-ergic input from SNR can instigate REM-on activity consistent with the Pal & Mallick (2009) conjecture
wake

Sleep deprivation
Prolonging the wake state leads to a longer subsequent sleep episode
See Phillips & Robinson (2008) for a systematic study of effects of sleep deprivation

Loss of orexin input to sleep promoting areas disrupts sleep/wake transitions
Consistent with studies of narcolepsy (Peyron et al 2000)`
gopi = 0.6gopi = 1.0 gopi = 0.1

• Gap-junctions are physical connections between neurons that allow current to flow between them
• Can an action potential in cell 1 evoke an action potential from cell 2?• Dynamics of gap-junctionally connected neurons have been subject of
prior investigation (Sherman-Rinzel 93, Chow-Kopell 98, Lewis et al. 2000, 01, 03, Medvedev et al. 2000, 08, 10, and many more)
• Lewis and Rinzel (2000) asked the question of whether periodic activity could be sustained in a network of neurons connected by gap-junctions. For specific network architectures (like cycles) they provide estimates on frequency based on rates of spontaneous activity.
• Gansert, Nadim and Golowasch (2007) asked how the size and shape of a neuron affects the ability of these networks to sustain activity.
Neuroscience to graph theory
Cell 1 Cell 2
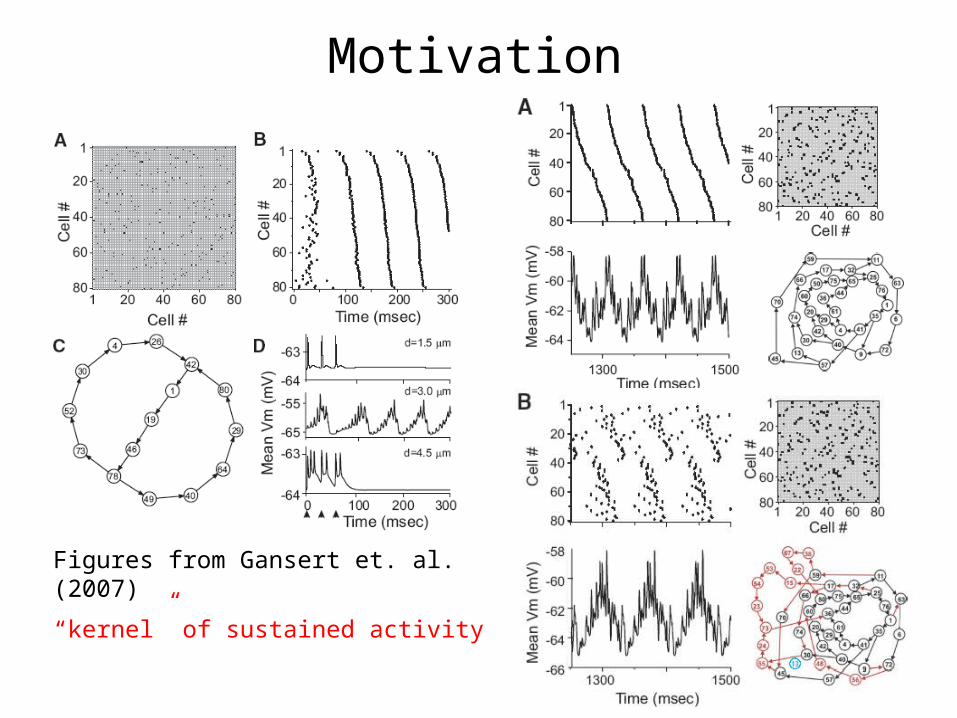
Motivation
Figures from Gansert et. al. (2007)
“kernel” of sustained activity

Some interesting questions in the context of generating rhythmic activity
● Are there specific architectures that promote sustained rhythmic activity? (Cycles for example)
● How do rules of nodal interaction affect the global dynamics ?
● How important are intrinsic dynamics of individual node in the sustainment of activity?
● In what way are dynamics related to graph structure?

Graph Properties
• G(n,p)={all graphs with n nodes and a probability p of an edge between any two nodes}
• The graph property Q consists of a subset of G(n,p) that share a common feature.
Ex. Q1 = {all graphs with a triangle}
Q2 = {all graphs that are connected}
G1 G2 G3 G4
G1,G 4 Q2 G2,G4 Q1 G3 Q1, Q2

Thresholds for 3 important monotone properties (Erdos, Renyi 1960)
• A property Q is monotone if whenever G Q and G H, then H Q.
• A function p*(n) is said to be a threshold function for a monotone property Q if
p(n)/p*(n) 0 implies that almost no G has Q, and
p(n)/p*(n) implies that almost every G has Q
• Appearance of the first edge at p ~ O(1/n2)• Appearance of k-cycles at p ~ O(1/n)• Disappearance of last isolated node at p ~ O((log n)/n)

Relating Dynamics and Graph structure
• In Singh et al (2011, SIADS), we show how random graph structure is related to periodic activity for both spiking and bursting neurons.
• Very non-intuitive results arise involving the giant component of the random graph
• We are currently investigating several theoretical questions in this area.
• Network and graph theory has seen a lot of interdisciplinary work in the area of physics.

Conclusion
• Mathematics turns out to be a good language to understand neuroscience.
• Mathematical modeling in close conjunction with experimental work is beginning to make inroads into the understanding of biological systems. There is still a lot of work to be done.
• Science can be advance by considering interdisciplinary approaches.







