
Journal of Asian Earth Sciences xxx (2013) xxx–xxx
Contents lists available at ScienceDirect
Journal of Asian Earth Sciences
journal homepage: www.elsevier .com/locate / jseaes
Distinct microbial communities thriving in gas hydrate-associatedsediments from the eastern Japan Sea
1367-9120/$ - see front matter � 2013 Elsevier Ltd. All rights reserved.http://dx.doi.org/10.1016/j.jseaes.2013.10.019
⇑ Corresponding authors. Tel.: +81 46 867 9717; fax: +81 46 867 9715.E-mail addresses: [email protected] (K. Yanagawa), [email protected]
s.u-tokyo.ac.jp (Y. Suzuki).1 Present address: Subsurface Geobiology Advanced Research (SUGAR) Project,
Japan Agency for Marine-Earth Science and Technology (JAMSTEC), Yokosuka, Japan.
Please cite this article in press as: Yanagawa, K., et al. Distinct microbial communities thriving in gas hydrate-associated sediments from the easterSea. Journal of Asian Earth Sciences (2013), http://dx.doi.org/10.1016/j.jseaes.2013.10.019
Katsunori Yanagawa a,⇑,1, Mariko Kouduka a, Yuri Nakamura b, Akihiro Hachikubo c, Hitoshi Tomaru d,Yohey Suzuki a,⇑a Department of Earth and Planetary Science, The University of Tokyo, 7-3-1 Hongo, Bunkyo-ku, Tokyo, Japanb Institute of Geology and Geoinformation, National Institute of Advanced Industrial Science and Technology (AIST), Tsukuba, Japanc Kitami Institute of Technology, 165 Koen-cho, Kitami, Japand Department of Earth Sciences, Graduate School of Science, Chiba University, 1-33 Yayoi-cho, Inage-ku, Chiba, Japan
a r t i c l e i n f o a b s t r a c t
Article history:Received 20 December 2012Received in revised form 4 October 2013Accepted 7 October 2013Available online xxxx
Keywords:Marine sedimentGas hydrateJapan SeaMicrobial community16S rRNA genesJS1
Marine gas hydrate and cold-seep systems, which maintain a large amount of methane in the seabed,may critically impact the geochemical and ecological characteristics of the deep-sea sedimentary envi-ronment. However, it remains unclear whether marine sediments associated with gas hydrate harbornovel microbial communities that are distinct from those from typical marine sediments. In this study,microbial community structures thriving in sediments associated with and without gas hydrate in theeastern Japan Sea were characterized by 16S rRNA gene-based phylogenetic analyses. Uncultivated bac-terial lineages of candidate division JS1 and a novel group NT-B2 were dominant in the sediments fromgas hydrate-associated sites. Whereas, microbial populations from sites not associated with gas hydratewere mainly composed of Bacteroidetes, Nitrospirales, Chlamydiales, Chlorobiales, and yet-unculturedbacterial lineages of OD1 and TM06. The good correlation between the dominance of JS1 and NT-B2and the association of gas hydrate could be attributed to the supply of more energetically favorableenergy sources in gas-rich fluids from the deep subsurface than refractory organic matter of terrigenousand diatomaceous origin.
� 2013 Elsevier Ltd. All rights reserved.
1. Introduction
Marine sediments are one of the largest methane reservoirs onEarth (Kvenvolden et al., 1993). Most of the methane under anoxicsedimentary conditions are produced and consumed microbiolog-ically by methanogenesis and anaerobic oxidation of methane(AOM), respectively (Valentine, 2011). Both microbial processesare mediated by cultivated and uncultivated lineages of Archaeaand play important ecological and biogeochemical roles in globalmarine systems (Knittel and Boetius, 2009).
To date, it remains unclear whether microbial populationsoccurring in the gas hydrate-bearing sediments are consistentlydistinct from those found in typical marine sediment on thecontinental margin. Numerous DNA-based studies on marinemicrobial life have targeted the deep subsurface in areas wheregas-rich formations are found (Inagaki et al., 2006; Kormas et al.,2003; Reed et al., 2002; Webster et al., 2006a). Uncultivable and
physiologically unknown lineages of Archaea and Bacteria arefound to be dominant in the subsurface sediments from NankaiTrough, Sea of Okhotsk, Cascadian Margin and Peru Margin (seeFry et al., 2008 and references therein). Inagaki et al. (2006) sug-gested that the novel lineages of JS1 and Deep-Sea Archaeal Group(DSAG) occur dominantly in sites associated with gas hydrate, andthat Chloroflexi-related phylotypes dominate organic-rich deepmarine sediments along the Pacific Ocean Margin (Inagaki et al.,2006). In contrast, subsurface sediment associated with methanehydrate in the Andaman Sea contains a unique microbial 16S rRNAgene phylotype composition dominated by Firmicutes (Briggset al., 2012).
The variability in microbial community might result from dif-ferences in DNA extraction, PCR primer set used, geographic isola-tion, sediment lithology, substrate availability and so on. Todistinguish microbial populations nutritionally dependent on gashydrate, a critical step is to evaluate microbial diversity in nearbysediments associated with and without gas hydrate by taking thesefactors into careful consideration. During MD179 cruise, marinesediment was collected from sites associated with and withoutgas hydrate within a semi-enclosed basin of the eastern JapanSea. The main goal of this study is to characterize microbial com-munity structures from sediments with a depth range across the
n Japan

2 K. Yanagawa et al. / Journal of Asian Earth Sciences xxx (2013) xxx–xxx
sulfate-methane transition zone (SMTZ) by 16S rRNA genephylotype analysis and to clarify the lineages of prokaryotes, themetabolic activity of which are potentially fueled by gas-rich fluidsin the eastern Japan Sea.
2. Materials and methods
2.1. Sampling sites
The study sites within a semi-enclosed and marginal basin inthe Japan Sea are mainly divided into three with respect to theoccurrence of gas hydrate: (1) those associated with actual gashydrate offshore Joestu city, (2) one site likely associated withgas hydrate off West Okushiri Ridge and (3) those not associatedwith gas hydrate on the Toyama Trough and off West Tsugaru(Fig. 1). Homogeneous deep seawater, so-called Japan Sea ProperWater, has extremely high dissolved oxygen concentrations(>210 lmol/kg), uniform salinity (�34.1‰) and low temperature(�0 �C) with a short residence time (�300 yr) (Tada, 1994). Japan
a
a
b
c
d
137º50′E37º44′N
37º21′N
-100
0m
-110
0m
-110
0m
-1000 m
-900 m
-700 m
-1200 m
-1300 m
3304
3296
330
Umitaka Spur
Joetsu Knoll Unn
Fig. 1. Site map of the research area in the eastern margin of Japan Sea during MD179 cruToyama Trough (b), a ridge 20 km southwest of West Tsugaru (c), and the West Okushi
Please cite this article in press as: Yanagawa, K., et al. Distinct microbial commuSea. Journal of Asian Earth Sciences (2013), http://dx.doi.org/10.1016/j.jseaes.2
Sea sediment is commonly of terrigenous and diatomaceous originwith a shallow SMTZ at depths of <5 m (Tomaru et al., 2011, Hachi-kubo et al., submitted for publication).
2.2. Sample collection
Sediment samples were collected during the MD179 cruiseusing the R/V Marion Dufresne in June 2010 (Fig. 1). The continuoussediment cores down to 40 m below the seafloor (mbsf) were takenusing a giant piston corer (Calypso). Microbiological analyses werecarried out from a total of seven core samples in the Japan Sea. Onthe Umitaka Spur, the Calypso core MD3304 was dropped adjacentto the gas seep site, located about 50 m off the gas seep vent(37�25.800E, 138�00.350E, 913 m below sea level [mbsl]). The twocores were taken from the surrounding sites (MD3296, locatedabout 0.5 km off methane seep vent: 37�24.810E, 138�00.800E,914 mbsl; MD3301, located about 1 km off the vent: 37�27.590N,138�04.600E, 1065 mbsl). Other cores were retrieved from non-gas seep sites in Unnamed Ridge (MD3312: 37�32.090N,
139º50′E
138º55′E
b 5km
137º50′E 138º10′E
38º10′N
38º25′N
c 5km
139º30′E
40º25′N
40º40′N
5km
138º10′E
d 5km
138º44′E
41º55′N
42º05′N
-1800 m
-1700m
-200m
-400 m
-600 m
-800 m
-1000m
-360
0m
-350
0m
-350
0m
-360
0m
-1100 m
1
3312
3326
3325
3329
amed Ridge
ise. Black dots indicate the coring sites located on the offshore Joetsu (a), the centralri Ridge (d).
nities thriving in gas hydrate-associated sediments from the eastern Japan013.10.019

K. Yanagawa et al. / Journal of Asian Earth Sciences xxx (2013) xxx–xxx 3
138�08.300E, 1026 mbsl), in the central Toyama Trough (MD3325G:38�16.520N, 137�55.040E, 1860 mbsl), in a ridge 20 km southwest ofWest Tsugaru (MD3326G: 40�31.320N, 138�37.990E, 325 mbsl), andin the possible gas seep site with strong bottom-simulating reflec-tors (BSR) located on West Okushiri Ridge (MD3329G: 42�01.290N,138�48.710E, 3448 mbsl) (Fig. 1). The sediment cores were cut into1.5-m-long sections immediately after the core retrieval. Subsam-ples for microbiological study were taken using sterilized spatulaand tip-cut syringe and stored at �80 �C for subsequent onshorelaboratory analysis.
2.3. Geochemical analysis
Porewater chemical compositions were determined in the sameway as described elsewhere (Hiruta et al., 2009). The dissolvedsulfate concentrations were measured by ion chromatography(TOA DKK, ICA-2000) with a PCI230 anion exchange column andelectrolytic conductivity detector. Dissolved gases in sedimentswere extracted according to Hachikubo et al. (submitted to SpecialIssue: Shallow Gas Hydrates). The gases were taken by a gas-tightsyringe and injected into a gas chromatograph (Shimadzu GC-14B)equipped with a flame-ionization detector for determination ofmethane concentration.
2.4. Microbial community analysis
2.4.1. DNA extractionProkaryotic DNA was extracted from 0.1 g of frozen sediment as
described previously (Kouduka et al., 2012). In brief, sedimentsample was incubated in a mixture of 0.33 N NaOH and TE bufferat 65 �C for 30 min to dissolve amorphous silica minerals. Afteralkaline incubation, aliquots were centrifuged at 5000g for 30 s.Then, the supernatant was placed into a new tube and neutralizedwith 1 M Tris–HCl (pH 6.5). After neutralization, a column packedwith polyvinylpolypyrrolidone was used for purification of sedi-ment samples containing humic substances (Holben et al., 1988).Purified DNA solution was stored at �20 �C.
2.4.2. Amplification of prokaryotic 16S rRNA, sequencing andphylogenetic analysis
Bacterial and archaeal 16S rRNA genes were amplified by PCRusing a universal primer set (Univ340F and Univ806R; Takai andHorikoshi, 2000). PCR reaction mixture was composed of each oli-gonucleotide primer, the DNA template, a LA Taq polymerase(TaKaRa Bio Inc., Shiga, Japan) and Ampdirect Plus (Shimadzu Co.,Ltd., Kyoto, Japan). Thermal cycling was performed as describedpreviously (Takai and Horikoshi, 2000). PCR products of approxi-mately 467 base pairs (bp) in size were gel-purified, cloned intovector pCR2.1-TOPO (Invitrogen, Carlsbad, CA), and then trans-formed into chemically competent Escherichia coli DH5a (TaKaRaBio Inc., Otsu, Shiga, Japan). Plasmid DNA from the clones were se-quenced using the M13 forward primer and the ABI Big-Dye reac-tion mix as described previously (Yanagawa et al., 2011). TheMothur Utility package was used for statistical analyses of the16S rRNA sequence data (Schloss et al., 2009). Partial 16S rRNAgene sequences with more than 97% similarity were assigned toidentical phylotypes. The representative sequences were alignedto the SILVA Reference Alignment using the NAST algorithm(Pruesse et al., 2007), followed by manual alignment. Phylogeneticaffiliations were assigned by maximum parsimony method withthe SILVA SSU Reference Database in ARB software (Ludwig et al.,2004).
2.4.3. OTU-based community cluster analysisA comparison of the microbial community structures was per-
formed using an operational taxonomic unit (OTU)-based approach
Please cite this article in press as: Yanagawa, K., et al. Distinct microbial commuSea. Journal of Asian Earth Sciences (2013), http://dx.doi.org/10.1016/j.jseaes.2
at the 97% identity level. The distance was determined by Yue &Clayton theta structural diversity measure (Schloss et al., 2009).
2.4.4. Nucleotide sequence accession numbersThe prokaryotic 16S rRNA gene sequences determined in this
study have been deposited in the DDBJ/EMBL/GenBank databasesunder accession numbers AB770165 to AB770469.
3. Results
3.1. Microbial communities inside the Umitaka Spur (MD3296,MD3301 and MD3304)
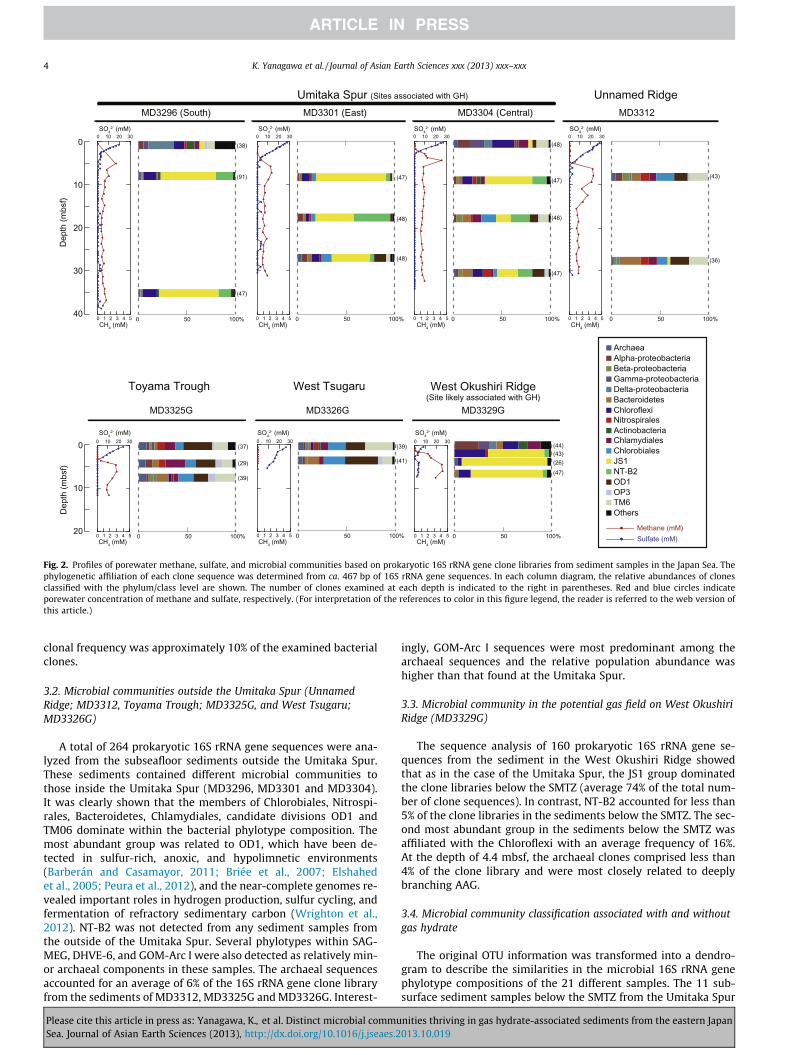
Using sediment core samples from the Umitaka Spur (MD3296,MD3301 and MD3304), within which the several gas seepageswere observed, a total of 507 prokaryotic 16S rRNA gene sequenceswere analyzed for the purpose of phylogenetic identification ofsubseafloor microbial community. The archaeal clones comprisedless than 4% of total prokaryotic clone sequences, and were closelyrelated to uncultivated lineages, the marine benthic group D(MBG-D; Vetriani et al., 1999), Deep-Sea Archaeal Group (DSAG;alternatively classified as the marine benthic group B [MBG-B]; Ta-kai and Horikoshi, 1999), the South African Gold Mine Euryarchae-otal Group (SAGMEG; Takai et al., 2001), the Deep-SeaHydrothermal Vent Euryarchaeotal Group 6 (DHVE-6; Takai andHorikoshi, 1999), and the ancient archaeal group (AAG; Takai andHorikoshi, 1999). No 16S rRNA gene sequences from lineages ofknown methanogens and methane-oxidizing archaea were de-tected in the clone library, with the exception of the GOM Arc Igroup of the order Methanosarcinales. This group is phylogeneti-cally close to the ANME-2 of anaerobic methane-oxidizing archaea(Mills et al., 2003), but there is no evidence of anaerobic oxidationof methane by the members of the GOM arc I lineage.
The surface sediments (5–52 cmbsf) harbored diverse phyloge-netic lineages of bacteria including members of the Chloroflexi,Firmicutes, Actinobacteria, Chlamydiales, Alpha-, Gamma-, andDelta-proteobacteria (Fig. 2). On the other hand, microbial commu-nity structures from the sediment samples deeper than SMTZ wasrelatively simple and remarkably uniform. The prokaryotic 16SrRNA gene clone libraries were dominated by uncultivated phylo-types closely related to the candidate division JS1, comprising anaverage of 44% of the 16S rRNA clone library from the sedimentsbelow the SMTZ (Fig. 2). The first JS1 sequence was identified fromsubseafloor sediments at 78 mbsf in the Japan Sea (Rochelle et al.,1994). Subsequently, JS1 has been found to be ubiquitous in vari-ous marine environments, such as near-surface marine sediments,deeply buried sediments, mud volcanoes, cold-seep sediments, gashydrate-bearing sediments, hydrothermal sediments, and ben-zene-degrading and acetate-utilizing sulfate-reducing sediments(Inagaki et al., 2002, 2003, 2006; Kormas et al., 2003; Li et al.,1999; Newberry et al., 2004; Phelps et al., 1998; Reed et al.,2002; Teske et al., 2002; Webster et al., 2004, 2006a, 2006b,2007). However, representatives of this group have not been culti-vated yet, and thus their metabolism remains unknown.
The second most abundant bacterial group from the sedimentsamples deeper than SMTZ was related to an uncultivated lineageof NT-B2, which accounted for an average of 16% of the clone li-brary from the sediments below the SMTZ. NT-B2 was only previ-ously found in subseafloor sediments from the Nankai TroughForearc Basin (Reed et al., 2002), the Peru Margin (Webster et al.,2006a) and the South Pacific Gyre (Durbin and Teske, 2011). Otherthan JS1 and NT-B2, we recovered sequences affiliated with theChloroflexi, which have been found as dominant populations inthe deep marine sediment (Inagaki et al., 2006). However the
nities thriving in gas hydrate-associated sediments from the eastern Japan013.10.019
ArchaeaAlpha-proteobacteriaBeta-proteobacteriaGamma-proteobacteriaDelta-proteobacteriaBacteroidetesChloroflexiNitrospiralesActinobacteriaChlamydialesChlorobialesJS1NT-B2OD1OP3TM6Others
MD3296 (South) MD3301 (East) MD3304 (Central) MD3312
MD3325G MD3326G
Dep
th (m
bsf)
0
10
30
40
20
Dep
th (m
bsf)
0
10
20
MD3329G
Umitaka Spur (Sites associated with GH) Unnamed Ridge
West Okushiri Ridge(Site likely associated with GH)
Toyama Trough West Tsugaru
0 1 2 3 4 5
0 10 20 30
0 1 2 3 4 5
0 10 20 30
0 1 2 3 4 5
0 10 20 30
0 1 2 3 4 5
0 10 20 30
0 1 2 3 4 5
0 10 20 30
0 1 2 3 4 5
0 10 20 30
0 1 2 3 4 5
0 10 20 30
Methane (mM)Sulfate (mM)
(38)
(91)
(47)
(47)
(48)
(48)
(48)
(47)
(46)
(47)
(43)
(36)
(37)
(29)
(39)
(39)
(41)
(44)(43)(26)(47)
0 100%50 0 100%50 0 100%50
0 100%50 0 100%50 0 100%50
SO42- (mM) SO4
2- (mM) SO42- (mM) SO4
2- (mM)
SO42- (mM) SO4
2- (mM) SO42- (mM)
CH4 (mM) CH4 (mM) CH4 (mM) CH4 (mM)
CH4 (mM) CH4 (mM) CH4 (mM)
0 100%50
Fig. 2. Profiles of porewater methane, sulfate, and microbial communities based on prokaryotic 16S rRNA gene clone libraries from sediment samples in the Japan Sea. Thephylogenetic affiliation of each clone sequence was determined from ca. 467 bp of 16S rRNA gene sequences. In each column diagram, the relative abundances of clonesclassified with the phylum/class level are shown. The number of clones examined at each depth is indicated to the right in parentheses. Red and blue circles indicateporewater concentration of methane and sulfate, respectively. (For interpretation of the references to color in this figure legend, the reader is referred to the web version ofthis article.)
4 K. Yanagawa et al. / Journal of Asian Earth Sciences xxx (2013) xxx–xxx
clonal frequency was approximately 10% of the examined bacterialclones.
3.2. Microbial communities outside the Umitaka Spur (UnnamedRidge; MD3312, Toyama Trough; MD3325G, and West Tsugaru;MD3326G)
A total of 264 prokaryotic 16S rRNA gene sequences were ana-lyzed from the subseafloor sediments outside the Umitaka Spur.These sediments contained different microbial communities tothose inside the Umitaka Spur (MD3296, MD3301 and MD3304).It was clearly shown that the members of Chlorobiales, Nitrospi-rales, Bacteroidetes, Chlamydiales, candidate divisions OD1 andTM06 dominate within the bacterial phylotype composition. Themost abundant group was related to OD1, which have been de-tected in sulfur-rich, anoxic, and hypolimnetic environments(Barberán and Casamayor, 2011; Briée et al., 2007; Elshahedet al., 2005; Peura et al., 2012), and the near-complete genomes re-vealed important roles in hydrogen production, sulfur cycling, andfermentation of refractory sedimentary carbon (Wrighton et al.,2012). NT-B2 was not detected from any sediment samples fromthe outside of the Umitaka Spur. Several phylotypes within SAG-MEG, DHVE-6, and GOM-Arc I were also detected as relatively min-or archaeal components in these samples. The archaeal sequencesaccounted for an average of 6% of the 16S rRNA gene clone libraryfrom the sediments of MD3312, MD3325G and MD3326G. Interest-
Please cite this article in press as: Yanagawa, K., et al. Distinct microbial commuSea. Journal of Asian Earth Sciences (2013), http://dx.doi.org/10.1016/j.jseaes.2
ingly, GOM-Arc I sequences were most predominant among thearchaeal sequences and the relative population abundance washigher than that found at the Umitaka Spur.
3.3. Microbial community in the potential gas field on West OkushiriRidge (MD3329G)
The sequence analysis of 160 prokaryotic 16S rRNA gene se-quences from the sediment in the West Okushiri Ridge showedthat as in the case of the Umitaka Spur, the JS1 group dominatedthe clone libraries below the SMTZ (average 74% of the total num-ber of clone sequences). In contrast, NT-B2 accounted for less than5% of the clone libraries in the sediments below the SMTZ. The sec-ond most abundant group in the sediments below the SMTZ wasaffiliated with the Chloroflexi with an average frequency of 16%.At the depth of 4.4 mbsf, the archaeal clones comprised less than4% of the clone library and were most closely related to deeplybranching AAG.
3.4. Microbial community classification associated with and withoutgas hydrate
The original OTU information was transformed into a dendro-gram to describe the similarities in the microbial 16S rRNA genephylotype compositions of the 21 different samples. The 11 sub-surface sediment samples below the SMTZ from the Umitaka Spur
nities thriving in gas hydrate-associated sediments from the eastern Japan013.10.019

K. Yanagawa et al. / Journal of Asian Earth Sciences xxx (2013) xxx–xxx 5
and West Okushiri Ridge formed a distinct cluster from the others(cluster I; Fig. 3). The structural feature of the 16S rRNA gene phyl-otype composition forming the cluster I was the dominance of JS1and NT-B2 as described above. The other cluster consisted of sitesnot associated with gas hydrate (i.e., Unnamed Ridge, ToyamaTrough and West Tsugaru; cluster II), which contained the mem-bers of OD1, TM06, Chlorobiales, Nitrospirales, Bacteroidetes andChlamydiales. These results indicate that the upwelling gas fluxis an important parameter controlling the subseafloor microbialcommunity structure.
4. Discussion
In this study, we revealed the distinct microbial 16S rRNA genephylotype compositions in marine sediments from seven sitesassociated with and without gas hydrate in the eastern JapanSea. One of the key features is the dominance of uncultivated bac-terial lineages in the sediments associated with gas hydrate. TheOTU-based community cluster analysis also clarified that microbial16S rRNA gene phylotype compositions were mainly divided intotwo clusters associated with and without the occurrence of gas hy-drate. These results suggest that gas hydrates might have an im-pact on development and/or maintenance of microbialcommunity structures in marine subsurface sediments.
The candidate division JS1 was characterized to be dominant inthe Umitaka Spur, known to harbor gas hydrate. The JS1 was foundto be a relatively major component in many subsurface marinesediments (e.g., Blazejak and Schippers, 2010; Fry et al., 2008),which may require specific metabolic functions to adapt to the glo-bal sedimentary habitat. Especially, 16S rRNA gene sequences ofJS1-related bacteria were frequently recovered from cold seeps[e.g., Japan Trench (Inagaki et al., 2002), Gulf of Mexico (Orcuttet al., 2010)], and methane hydrate-bearing sediments [e.g., PeruMargin (Inagaki et al., 2006), Nankai Trough (Reed et al., 2002)].Quantitative PCR-based study also showed the dominance of JS1in cold seep sediments (Zhang et al., 2012). These findings indicatethat JS1 has a preference of habitat enriched in methane.
The NT-B2 phylotypes co-occurred with JS1 in the sedimentsassociated with gas hydrate, whereas no clonal signals of NT-B2phylotypes were detected in those not associated with gas hydrate.This indicates that NT-B2 and JS1 might favor similar physical andchemical conditions for their growth. Although NT-B2 and JS1 areboth present in methane hydrate-bearing subsurface sedimentsfrom the Nankai Forearc Basin (Reed et al., 2002), NT-B2 is not al-ways present with JS1 in subseafloor sediments (Webster et al.,2006a).
MD3296 Um
MD3296 UmMD3296 Um
MD3301 Um
MD3301 Um
MD3301 Um
MD3304 Um
MD3304 Um
MD3304 Um
MD3304 Um
MD3312 Unn
MD3312 Unn
MD3325G ToMD3325G To
MD3325G To
MD3326G W
MD3326G W
MD3329G W
MD3329G W
MD3329G W
MD3329G W
Fig. 3. OTU-based cluster analysis of 21 subseafloor microbial communities in the Japacommunity structure similarity coefficient. The distance was determined from Yue & Cl
Please cite this article in press as: Yanagawa, K., et al. Distinct microbial commuSea. Journal of Asian Earth Sciences (2013), http://dx.doi.org/10.1016/j.jseaes.2
Sediment samples from the Unnamed Ridge, Toyama Troughand West Tsugaru formed cluster II represented by the predomi-nance of OD1 and TM6 in addition to the phylum-level lineageswith cultivated representatives. The OD1 sequences have beenmainly detected from anoxic and sulfur-rich environments (Briéeet al., 2007). Recent metagenomic analyses have shown that OD1might be involved in sulfur cycling (Elshahed et al., 2005; Wrigh-ton et al., 2012). However the predominant incidence of the OD1clones in marine sedimentary habitat has been rarely reported(Durbin and Teske, 2011; Jorgensen et al., 2012). The detection ofOD1 16S rRNA phylotypes may be affected by the DNA extractionmethod and the PCR condition of the experiments: e.g., the primerset used in this study has a much higher coverage to known se-quences of OD1 (Peura et al., 2012). Therefore, the OD1 phylotypesmay have been previously underestimated in marine sediments.
Downward diffusion of sulfate from the seafloor, upward diffu-sion of methane, microbial consumption of sulfate and methane,and methanogenesis appear to shape SMTZ in marine sediments(Reeburgh, 2007). Despite the fact that microbial communitiesare very sensitive to redox conditions in the water column (Wrigh-ton et al., 2012), changes in dominant microbial populations corre-sponding to porewater geochemical properties were not evidentfor the sites not associated with gas hydrate. One explanationcould be that most of microbial populations at the sites not associ-ated with gas hydrate mediate the respiratory and fermentativedegradation of refractory organic matter of terrigenous and diato-maceous origins (Lloyd et al., 2013; Orcutt et al., 2013; Rinke et al.,2013). As the deposition of organic matter is considered to be notso different among all investigated areas in the Japan Sea, it is pos-sible that the same sediment microbial communities that are notassociated with gas hydrates could similarly thrive in sedimentsassociated with gas hydrate. However, as distinct microbial 16SrRNA gene phylotypes were present in sediments associated withgas hydrate, it is possible that these microbial communities alsorely on energy sources derived from upwelling gas-rich fluids.These gas-rich fluids are likely produced in the deep subsurface un-der high-temperature conditions associated with thermal degrada-tion of organic matters (Hachikubo et al., submitted to J. AsianEarth Sci.). Hence, they would be enriched not only with methanebut also alkanes, alkenes, alcohols, ketones, carboxylic acids andaromatic compounds (Seewald, 2003). These substrates might beutilized by the distinct microbial communities found in the sedi-ments associated with gas hydrate.
Gas hydrate is actively being formed at or near the seafloor inthe eastern Japan Sea. The near-seafloor gas hydrate is associatedwith a vertical seismic anomaly, called a gas chimney. Gas
itaka Spur 0.5 mbsf
itaka Spur 8 mbsfitaka Spur 35.2 mbsf
itaka Spur 8.7 mbsf
itaka Spur 17.8 mbsf
itaka Spur 27 mbsf
itaka Spur 0 mbsf
itaka Spur 8.8 mbsf
itaka Spur 17.6 mbsf
itaka Spur 30.8 mbsf
amed Ridge 8.2 mbsf
amed Ridge 27.5 mbsf
yama Trough 0 mbsfyama Trough 4.5 mbsf
yama Trough 7.6 mbsf
est Tsugaru 0 mbsf
est Tsugaru 4.1 mbsf
est Okushiri Ridge 0 mbsf
est Okushiri Ridge 0.5 mbsf
est Okushiri Ridge 4.4 mbsf
est Okushiri Ridge 6.3 mbsf
Cluster II(Sites not associated with gas hydrate)
Cluster I(Sites associted with gas hydrate)
n Sea. The dendrogram represents the similarity of the samples based on the HYC
ayton theta values. The distance from the tip of the dendrogram to the root is 0.5.
nities thriving in gas hydrate-associated sediments from the eastern Japan013.10.019

6 K. Yanagawa et al. / Journal of Asian Earth Sciences xxx (2013) xxx–xxx
chimneys are formed where the upward migration of gas-richfluids is extensive. A gas chimney in the Umitaka Spur is estimatedto have a diameter of 1–2 km (Matsumoto et al., 2005), and thelarge area around the gas chimney could be influenced by thegas-rich fluids. The distinct microbial communities were alsofound in the marine sediments from the West Okushiri Ridge,which suggests upward migration of gas-rich fluid at our studysite. This area is known for shallow SMTZ, which is likely causedby anaerobic oxidation of methane coupled to sulfate reduction.At the Northern Okushiri Ridge, the previous study has confirmedmethane seep-associated chemosynthetic biological communities(Naganuma et al., 1999). Furthermore, gas hydrate was found at�90 mbsf during ODP Leg127 (Kvenvolden, 1995). Hence, it is ex-pected that gas hydrate is present nearby the investigated site.
JS1- and NT-B2-related bacteria may prefer habitats enriched incompounds derived from gas-rich fluid and are widely distributedin the Pacific and the eastern Japan Sea. It remains unclear whatkinds of substances preferentially serve as energetically favorableelectron donors. To firmly establish that these lineages of bacteriaare diagnostic to the occurrence of gas hydrate, the correlation be-tween gas hydrate and microbial community should be clarified inthe sediments from different geographical, oceanographic and geo-logic settings in the world, coupled with quantitative analyses oforganic and inorganic compounds associated with gas hydrate.
5. Conclusions
Here we described microbial community structures at the sitesassociated with and without gas hydrate in the eastern Japan Sea.Prokaryotic 16S rRNA gene-based phylogenetic analyses showedthat the majority of microbial communities found in marine sedi-ments associated with and without gas hydrate are clearly differ-ent. The physiologically unknown lineages of bacteria JS1 andNT-B2 that flourished in gas-rich sediments at depth might bedependent on chemical compounds enriched in gas-rich fluids astheir preferential energy sources. These notions will be furtherevaluated by our future study.
Acknowledgements
We thank the shipboard science party of the MD179 cruise andthe crews and operation teams of the R/V Marion Dufresne for theirsupport in sample collection. This work was supported in part bythe MH21 Research Consortium in Japan and the Grant-in-Aid forExploratory Research (KAKENHI: 23651015).
References
Barberán, A., Casamayor, E.O., 2011. Euxinic freshwater hypolimnia promotebacterial endemicity in continental areas. Microbial Ecology 61, 465–472.
Blazejak, A., Schippers, A., 2010. High abundance of JS-1- and Chloroflexi-relatedBacteria in deeply buried marine sediments revealed by quantitative, real-timePCR. FEMS Microbiology Ecology 72, 198–207.
Briée, C., Moreira, D., López-García, P., 2007. Archaeal and bacterial communitycomposition of sediment and plankton from a suboxic freshwater pond.Research in Microbiology 158, 213–227.
Briggs, B.R., Inagaki, F., Morono, Y., Futagami, T., Huguet, C., Rosell-Mele, A.,Lorenson, T.D., Colwell, F.S., 2012. Bacterial dominance in subseafloor sedimentscharacterized by methane hydrates. FEMS Microbiology Ecology 81, 88–98.
Durbin, A.M., Teske, A., 2011. Microbial diversity and stratification of South Pacificabyssal marine sediments. Environmental Microbiology 13, 3219–3234.
Elshahed, M.S., Najar, F.Z., Aycock, M., Qu, C., Roe, B.A., Krumholz, L.R., 2005.Metagenomic analysis of the microbial community at Zodletone Spring(Oklahoma): insights into the genome of a member of the novel CandidateDivision OD1. Applied and Environmental Microbiology 71, 7598–7602.
Fry, J.C., Parkes, R.J., Cragg, B.A., Weightman, A.J., Webster, G., 2008. Prokaryoticbiodiversity and activity in the deep subseafloor biosphere. FEMS MicrobiologyEcology 66, 181–196.
Hachikubo, A., Yanagawa, K., Tomaru, H., Lu, H., Matsumoto, R., Molecular andisotopic analysis of volatiles in gas hydrates and in pore water from Joetsu
Please cite this article in press as: Yanagawa, K., et al. Distinct microbial commuSea. Journal of Asian Earth Sciences (2013), http://dx.doi.org/10.1016/j.jseaes.2
Basin, eastern margin of Japan Sea. Journal of Asian Earth Sciences, Submitted.(Special Issue: Shallow Gas Hydrates).
Hiruta, A., Snyder, G.T., Tomaru, H., Matsumoto, R., 2009. Geochemical constraintsfor the formation and dissociation of gas hydrate in an area of high methaneflux, eastern margin of the Japan Sea. Earth and Planetary Science Letters 279,326–339.
Holben, W.E., Jansson, J.K., Chelm, B.K., Tiedje, J.M., 1988. DNA probe method for thedetection of specific microorganisms in the soil bacterial community. Appliedand Environmental Microbiology 54, 703–711.
Inagaki, F., Sakihama, Y., Inoue, A., Kato, C., Horikoshi, K., 2002. Molecularphylogenetic analyses of reverse-transcribed bacterial rRNA obtained fromdeep-sea cold seep sediments. Environmental Microbiology 4, 277–286.
Inagaki, F., Suzuki, M., Takai, K., Oida, H., Sakamoto, T., Aoki, K., Nealson, K.H.,Horikoshi, K., 2003. Microbial communities associated with geological horizonsin coastal subseafloor sediments from the Sea of Okhotsk. Applied andEnvironmental Microbiology 69, 7224–7235.
Inagaki, F., Nunoura, T., Nakagawa, S., Teske, A., Lever, M., Lauer, A., Suzuki, M.,Takai, K., Delwiche, M., Colwell, F.S., Nealson, K.H., Horikoshi, K., D’Hondt, S.,Jørgensen, B.B., 2006. Biogeographical distribution and diversity of microbes inmethane hydrate-bearing deep marine sediments, on the Pacific Ocean Margin.Proceedings of the National Academy of Sciences of the United States ofAmerica 103, 2815–2820.
Jorgensen, S.L., Hannisdal, B., Lanzén, A., Baumberger, T., Flesland, K., Fonseca, R.,Øvreås, L., Steen, I.H., Thorseth, I.H., Pedersen, R.B., Schleper, C., 2012.Correlating microbial community profiles with geochemical data in highlystratified sediments from the Arctic Mid-Ocean Ridge. Proceedings of theNational Academy of Sciences of the United States of America 109, E2846–E2855.
Knittel, K., Boetius, A., 2009. Anaerobic oxidation of methane: progress with anunknown process. Annual Review of Microbiology 63, 311–334.
Kormas, K.A., Smith, D.C., Edgcomb, V., Teske, A., 2003. Molecular analysis of deepsubsurface microbial communities in Nankai Trough sediments (ODP Leg 190,Site 1176). FEMS Microbiology Ecology 45, 115–125.
Kouduka, M., Suko, T., Morono, Y., Inagaki, F., Ito, K., Suzuki, Y., 2012. A new DNAextraction method by controlled alkaline treatments from consolidatedsubsurface sediments. FEMS Microbiology Letters 326, 47–54.
Kvenvolden, K.A., 1995. A review of the geochemistry of methane in natural gashydrate. Organic Geochemistry 23, 997–1008.
Kvenvolden, K.A., Ginsburg, G.D., Soloviev, V.A., 1993. Worldwide distribution ofsubaquatic gas hydrates. Geo-Marine Letters 13, 32–40.
Li, L.N., Kato, C., Horikoshi, K., 1999. Bacterial diversity in deep-sea sediments fromdifferent depths. Biodiversity and Conservation 8, 659–677.
Lloyd, K.G., Schreiber, L., Petersen, D.G., Kjeldsen, K.U., Lever, M.A., Steen, A.D.,Stepanauskas, R., Richter, M., Kleindienst, S., Lenk, S., Schramm, A., Jorgensen,B.B., 2013. Predominant archaea in marine sediments degrade detrital proteins.Nature 496, 215–218.
Ludwig, W., Strunk, O., Westram, R., Richter, L., Meier, H., Yadhukumar, Buchner, A.,Lai, T., Steppi, S., Jobb, G., Fôrster, W., Brettske, I., Gerber, S., Ginhart, A.W., Gross,O., Grumann, S., Hermann, S., Jost, R., Kônig, A., Liss, T., Lüßmann, R., May, M.,Nonhoff, B., Reichel, B., Strehlow, R., Stamatakis, A., Stuckmann, N., Vilbig, A.,Lenke, M., Ludwig, T., Bode, A., Schleifer, K.H., 2004. ARB: a softwareenvironment for sequence data. Nucleic Acids Research 32, 1363–1371.
Matsumoto, R., Okuda, Y., Aoyama, C., Hiruta, A., Ishida, Y., Sunamura, M.,Numanami, H., Tomaru, H., Snyder, G.T., Komatsubara, J., Takeuchi, R.,Hiromatsu, M., 2005. Methane plumes over a marine gas hydrate system inthe eastern margin of Japan Sea: a possible mechanism for the transportation ofsubsurface methane to shallow waters. In: Proceedings of the 5th InternationalConference on Gas Hydrate, Trondheim, Norway, pp. 749–754.
Mills, H.J., Hodges, C., Wilson, K., MacDonald, I.R., Sobecky, P.A., 2003. Microbialdiversity in sediments associated with surface-breaching gas hydrate moundsin the Gulf of Mexico. FEMS Microbiology Ecology 46, 39–52.
Naganuma, T., Julius Meisel, C., Wada, H., Kato, Y., Takeuchi, A., Fujikura, K., Naka, J.,Fujioka, K., 1999. Sea-floor fissures, biological communities and sediment fattyacids of the Northern Okushiri Ridge, Japan Sea: implications for possiblemethane seepage. Island Arc 8, 232–244.
Newberry, C.J., Webster, G., Cragg, B.A., Parkes, R.J., Weightman, A.J., Fry, J.C., 2004.Diversity of prokaryotes and methanogenesis in deep subsurface sedimentsfrom the Nankai Trough, Ocean Drilling Program Leg 190. EnvironmentalMicrobiology 6, 274–287.
Orcutt, B.N., Joye, S.B., Kleindienst, S., Knittel, K., Ramette, A., Reitz, A., Samarkin, V.,Treude, T., Boetius, A., 2010. Impact of natural oil and higher hydrocarbons onmicrobial diversity, distribution, and activity in Gulf of Mexico cold-seepsediments. Deep-Sea Research Part II, Topical Studies in Oceanography 57,2008–2021.
Orcutt, B.N., LaRowe, D.E., Biddle, J.F., Colwell, F.S., Glazer, B.T., Reese, B.K.,Kirkpatrick, J.B., Lapham, L.L., Mills, H.J., Sylvan, J.B., Wankel, S.D., Wheat, C.G.,2013. Microbial activity in the marine deep biosphere: progress and prospects.Frontiers in Microbiology 4, 189.
Peura, S., Eiler, A., Bertilsson, S., Nykänen, H., Tiirola, M., Jones, R.I., 2012. Distinctand diverse anaerobic bacterial communities in boreal lakes dominated bycandidate division OD1. The ISME Journal 6, 1640–1652.
Phelps, C.D., Kerkhof, L.J., Young, L.Y., 1998. Molecular characterization of a sulfate-reducing consortium which mineralizes benzene. FEMS Microbiology Ecology27, 269–279.
Pruesse, E., Quast, C., Knittel, K., Fuchs, B.M., Ludwig, W.G., Peplies, J., Glöckner, F.O.,2007. SILVA: a comprehensive online resource for quality checked and aligned
nities thriving in gas hydrate-associated sediments from the eastern Japan013.10.019

K. Yanagawa et al. / Journal of Asian Earth Sciences xxx (2013) xxx–xxx 7
ribosomal RNA sequence data compatible with ARB. Nucleic Acids Research 35,7188–7196.
Reeburgh, W.S., 2007. Oceanic methane biogeochemistry. Chemical Reviews 107,486–513.
Reed, D.W., Fujita, Y., Delwiche, M.E., Blackwelder, D.B., Sheridan, P.P., Uchida, T.,Colwell, F.S., 2002. Microbial communities from methane hydrate-bearing deepmarine sediments in a forearc basin. Applied and Environmental Microbiology68, 3759–3770.
Rinke, C., Schwientek, P., Sczyrba, A., Ivanova, N.N., Anderson, I.J., Cheng, J.-F.,Darling, A., Malfatti, S., Swan, B.K., Gies, E.A., Dodsworth, J.A., Hedlund, B.P.,Tsiamis, G., Sievert, S.M., Liu, W.-T., Eisen, J.A., Hallam, S.J., Kyrpides, N.C.,Stepanauskas, R., Rubin, E.M., Hugenholtz, P., Woyke, T., 2013. Insights into thephylogeny and coding potential of microbial dark matter. Nature 499, 431–437.
Rochelle, P.A., Cragg, B.A., Fry, J.C., John Parkes, R., Weightman, A.J., 1994. Effect ofsample handling on estimation of bacterial diversity in marine sediments by16S rRNA gene sequence analysis. FEMS Microbiology Ecology 15, 215–225.
Schloss, P.D., Westcott, S.L., Ryabin, T., Hall, J.R., Hartmann, M., Hollister, E.B.,Lesniewski, R.A., Oakley, B.B., Parks, D.H., Robinson, C.J., Sahl, J.W., Stres, B.,Thallinger, G.G., Van Horn, D.J., Weber, C.F., 2009. Introducing mothur: open-source, platform-independent, community-supported software for describingand comparing microbial communities. Applied and EnvironmentalMicrobiology 75, 7537–7541.
Seewald, J.S., 2003. Organic–inorganic interactions in petroleum-producingsedimentary basins. Nature 426, 327–333.
Tada, R., 1994. Paleoceanographic evolution of the Japan Sea. Palaeogeography,Palaeoclimatology, Palaeoecology 108, 487–508.
Takai, K., Horikoshi, K., 1999. Genetic diversity of archaea in deep-sea hydrothermalvent environments. Genetics 152, 1285–1297.
Takai, K., Horikoshi, K., 2000. Rapid detection and quantification of members of thearchaeal community by quantitative PCR using fluorogenic probes. Applied andEnvironmental Microbiology 66, 5066–5072.
Takai, K., Moser, D.P., DeFlaun, M., Onstott, T.C., Fredrickson, J.K., 2001. Archaealdiversity in waters from deep South African gold mines. Applied andEnvironmental Microbiology 67, 5750–5760.
Teske, A., Hinrichs, K.U., Edgcomb, V., Gomez, A.D., Kysela, D., Sylva, S.P., Sogin, M.L.,Jannasch, H.W., 2002. Microbial diversity of hydrothermal sediments in theGuaymas Basin: evidence for anaerobic methanotrophic communities. Appliedand Environmental Microbiology 68, 1994–2007.
Please cite this article in press as: Yanagawa, K., et al. Distinct microbial commuSea. Journal of Asian Earth Sciences (2013), http://dx.doi.org/10.1016/j.jseaes.2
Tomaru, H., Muramatsu, Y., Anzai, H., Snyder, G., Matsumoto, R., 2011.Geochemistry of pore waters in methane rich sediments, eastern margin ofthe Japan Sea. In: Proceedings of the 7th International Conference on GasHydrates, Edinburgh, Scotland, UK.
Valentine, D.L., 2011. Emerging topics in marine methane biogeochemistry. AnnualReview of Marine Science 3, 147–171.
Vetriani, C., Jannasch, H.W., MacGregor, B.J., Stahl, D.A., Reysenbach, A.L., 1999.Population structure and phylogenetic characterization of marine benthicarchaea in deep-sea sediments. Applied and Environmental Microbiology 65,4375–4384.
Webster, G., Parkes, R.J., Fry, J.C., Weightman, A.J., 2004. Widespread occurrence of anovel division of bacteria identified by 16S rRNA gene sequences originallyfound in deep marine sediments. Applied and Environmental Microbiology 70,5708–5713.
Webster, G., John Parkes, R., Cragg, B.A., Newberry, C.J., Weightman, A.J., Fry, J.C.,2006a. Prokaryotic community composition and biogeochemical processes indeep subseafloor sediments from the Peru Margin. FEMS Microbiology Ecology58, 65–85.
Webster, G., Watt, L.C., Rinna, J., Fry, J.C., Evershed, R.P., Parkes, R.J., Weightman, A.J.,2006b. A comparison of stable-isotope probing of DNA and phospholipid fattyacids to study prokaryotic functional diversity in sulfate-reducing marinesediment enrichment slurries. Environmental Microbiology 8, 1575–1589.
Webster, G., Yarram, L., Freese, E., Köster, J., Sass, H., Parkes, R.J., Weightman, A.J.,2007. Distribution of candidate division JS1 and other Bacteria in tidalsediments of the German Wadden Sea using targeted 16S rRNA gene PCR-DGGE. FEMS Microbiology Ecology 62, 78–89.
Wrighton, K.C., Thomas, B.C., Sharon, I., Miller, C.S., Castelle, C.J., VerBerkmoes, N.C.,Wilkins, M.J., Hettich, R.L., Lipton, M.S., Williams, K.H., Long, P.E., Banfield, J.F.,2012. Fermentation, hydrogen, and sulfur metabolism in multiple uncultivatedbacterial phyla. Science 337, 1661–1665.
Yanagawa, K., Sunamura, M., Lever, M.A., Morono, Y., Hiruta, A., Ishizaki, O.,Matsumoto, R., Urabe, T., Inagaki, F., 2011. Niche separation of methanotrophicarchaea (ANME-1 and -2) in methane-seep sediments of the eastern Japan Seaoffshore Joetsu. Geomicrobiology Journal 28, 118–129.
Zhang, Y., Su, X., Chen, F., Jiao, L., Jiang, H., Dong, H., Ding, G., 2012. Abundance anddiversity of candidate division JS1- and Chloroflexi-related bacteria in cold seepsediments of the northern South China Sea. Frontiers of Earth Science. http://dx.doi.org/10.1007/s11707-012-0324-0.
nities thriving in gas hydrate-associated sediments from the eastern Japan013.10.019







