
Chapitre IV. Les principes de dissection génétique
1

Fonction
Gène Protéineséquence
isolement demutants
clonage du gène& analyse bioinformatique
interprétation(biochimie, biologie cellulaire)
Principaux organismes modèles eucaryotes
2
La génétique est aussi une approche expérimentale puissante pour ″disséquer‟ lesfonctions biologiques, cad identifier les gènes et les protéines impliquées dans cesfonctions. On utilise pour cela des organismes modèles qui ont la particularité de seprêter aisément à l’analyse génétique :
Les gènes identifiés chez un organisme
modèle sont souvent conservés chez des
espèces plus complexes dont les
mammifères

La levure : modèle d’étude en biologie cellulaire des eucaryotes
- voies métaboliques (enzymes)
- réplication et réparation de l’ADN- transcription de l’ADN et régulation
- maturation et traduction des ARNm- recombinaison de l’ADN
- perméabilité cellulaire (perméases, canaux, ..)- communication, réponse cellulaire (nutriments,
hormones, stress, ..)
- transport intracellulaire des protéines- cycle cellulaire (mitose, méiose)
- maturation et dégradation des protéines
- peroxisomes, mitochondries- ER, Golgi, endosomes, vacuole- cytosquelette / transport vésiculaire- développement, apoptose, ..
- prions
Fonctions à l'étude : Fonctionscellulaires
Gène Protéineséquence
isolement demutants
clonage du gène& analyse bioinformatique
interprétation(biochimie, biologie cellulaire)
3

4
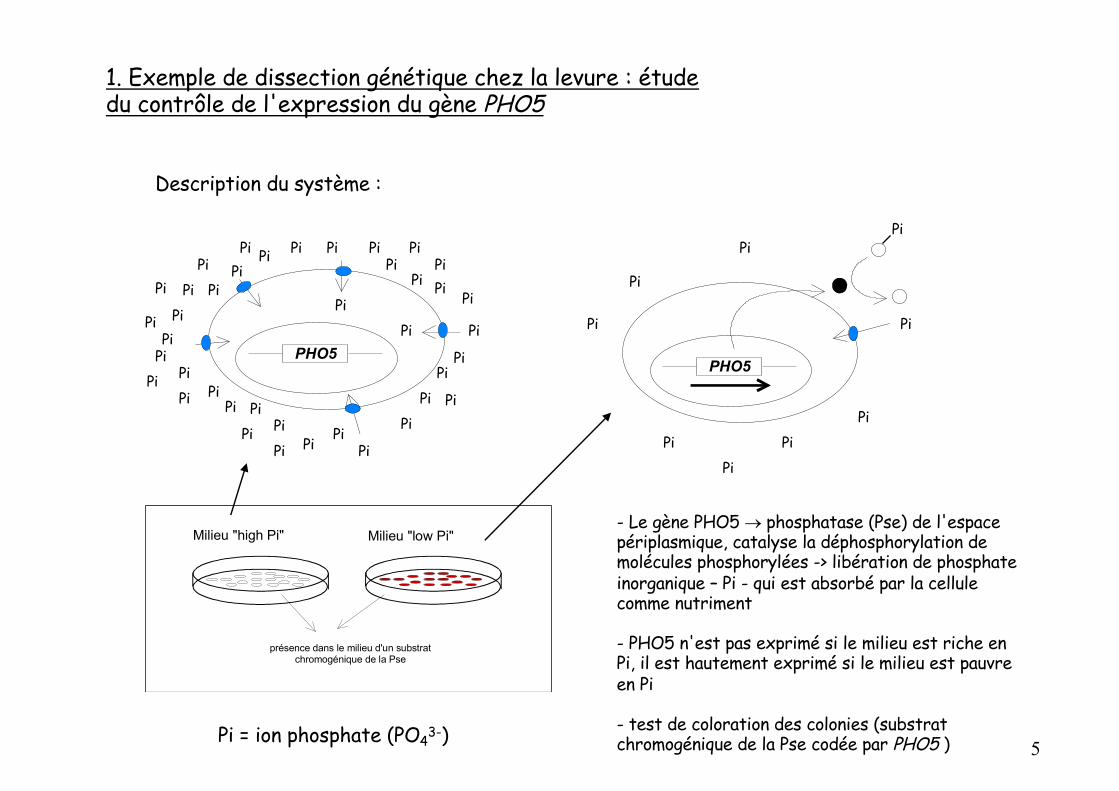
Description du système :
Milieu "high Pi" Milieu "low Pi"
présence dans le milieu d'un substrat chromogénique de la Pse
PHO5PHO5
PiPi
Pi
Pi
Pi
Pi
PiPi
PiPi
Pi
PiPi
Pi PiPiPi
Pi
Pi PiPi
Pi
PiPiPi
Pi
PiPi
PiPi
PiPiPiPi Pi
PiPi
PiPi
Pi
Pi
Pi
Pi Pi
Pi
Pi PiPi
Pi
- Le gène PHO5 ® phosphatase (Pse) de l'espace périplasmique, catalyse la déphosphorylation de molécules phosphorylées -> libération de phosphate inorganique – Pi - qui est absorbé par la cellule comme nutriment
- PHO5 n'est pas exprimé si le milieu est riche en Pi, il est hautement exprimé si le milieu est pauvre en Pi
- test de coloration des colonies (substrat chromogénique de la Pse codée par PHO5 )Pi = ion phosphate (PO43-)
5
1. Exemple de dissection génétique chez la levure : étude du contrôle de l'expression du gène PHO5

Procédure suivie :
1 - Isolement de mutants (phénotype: gène PHO5 dérégulé)
2 – Classement des mutants (® combien de gènes interviennentdans la régulation de PHO5 ?)
3 – Relations entre les gènes ainsi répertoriés
4 – Clonage des gènes
® séquence, protéine, …
6
Objectif de la dissection génétique : identifier les gènes/protéines impliquées dans la régulation du gène PHO5

1.1 Mutagenèse et isolement de mutants Pse- et Psec
Milieu "high Pi" Milieu "low Pi"
levures sauvages haploïdes
Répliques par tampon de velours
mutant Pse constitutive
mutant Pse -
- Traitement chimique mutagène de cellules sauvages haploïdes des deux signes (a et a) (donc, deux cultures et deux mutagenèses séparées)
- Agent mutagène = EMS = éthyl-méthane sulfonate = agent alkylant
- Obtention d'un grand nombre de colonies
- Réplique par tampon de velours sur milieux pauvre ou riche en Pi (avec substrat chromogénique de Pho5)
- Obtention d'un grand nombre de clones de phénotype "phosphatase moins" et "phosphatase constitutive"
7

1.2. Classement des mutants
- phénotype récessif ou dominant ? ® croisement de chaque mutant avec la souche sauvage de signe sexuel contraire et analyse du 2n hétérozygote
Principe :
Mutant
Sauvage Phénotype du diploïde
Type de mutation
Pse- Pse+ Pse+ Récessive Pse- Pse+ Pse- Dominante PseC Pse+ Pse+ Récessive PseC Pse+ PseC Dominante
mutation récessive = mutation de perte de fonction(complémentée par l’allèle sauvage)
mutation dominante = mutation de gain de fonction(non complémentée par l’allèle sauvage)
Ú
ÚÚ
Résultat : 3 types de mutants sont obtenus :
- mutants Pse- récessifs- mutants Psec récessifs- mutants Psec dominants
8

- Les mutants Pse- sont-ils tous mutés dans le même gène? Ou dans plusieurs gènes, mais alors combien? ® test de complémentation entre les différents mutants récessifs Pse-
Principe :
n
n
2nx
x
xx Complémentation
(Phénotype sauvage)
Mut
ants
aff
ecté
s da
ns
des
gène
s di
ffér
ents
n
n2n
x
x
xx Pas de complémentation
(Phénotype mutant)
Mut
ants
aff
ecté
s da
ns
le m
ême
gène
9

1 2 3 4 5 6 7 8 wt 9 + + + - - + + + + 10 + - + + + + + + + 11 - + - + + + - - + 12 + + + + + - + + + 13 + + + - - + + + + 14 + + + - - + + + + 15 + - + + + + + + + 16 + + + - - + + + + 17 - + - + + + - - + 18 - + - + + + - - + wt + + + + + + + + +
Ex. : les mutations récessives Pse- : croisement deux à deux de 8 mutants (1-8, signe a) avec 10 autres mutants (9-18, signe a) et analyse du phénotype Pse des 2n :
® 4 classes de complémentation :
I : 1, 3, 7, 8, 11, 17, 18 II : 2, 10, 15
III : 4, 5, 9, 13, 14, 16IV : 6, 12
Même type d'analyse pour les mutations récessives PseC ® 2 classes de complémentation I, II
Conclusions :
classe I = gène PHO5 (il sera établi que PHO5 est le gène de la Pse)classe II = gène PHO4 (facteur positif de l'expression de PHO5)classe III = gène PHO2 (facteur positif de l'expression de PHO5)classe IV = gène PHO81 (facteur positif de l'expression de PHO5)
classe I = gène PHO80 (facteur négatif de l'expression de PHO5)classe II = gène PHO85 (facteur négatif de l'expression de PHO5) 10
Phénotype Pse des diploïdes

PHO5PHO5 PHO5PHO5
Pi
+
¯Pi
PHO5PHO5
Pi ¯Pi Pi ¯Pi
Pse+ Pse- Pse-Pse+
Un facteur positif active l’expression de PHO5 quand le milieu est pauvre en Pi (il n’agit pas quand le milieu est riche en Pi). Si sa fonction est perdue (mutant pho2, pho4, pho81), PHO5 n’est plus exprimé sur ce milieu, et ce défaut peut-être complémenté par l’apport du gène sauvage codant pour ce facteur positif (le phénotype mutant est récessif)
+
11

PHO5PHO5 PHO5PHO5
Pi
_
¯Pi
PHO5PHO5
_
Pi ¯Pi Pi ¯Pi
Pse+ Psec Psec
Pse+
Un facteur négatif empêche l’expression de PHO5 quand le milieu est riche en Pi (il n’agit plus quand le milieu est pauvre en Pi). Si sa fonction est perdue (mutant pho80, pho85), l’expression de PHO5 est constitutive, et ce défaut peut-être complémenté par l’apport du gène sauvage codant pour ce facteur négatif (phénotype récessif)
12

- quid des mutants PseC dominants ? Il est probable que ces mutations de gain de fonctionaltèrent l’un ou l’autre gène PHO identifiés grâce aux mutations récessives (ex. p. 14). Pourtester cette hypothèse, on réalise un test de liaison génétique entre chaque mutant dominantconstitutif et un représentant de chaque classe de mutants pho récessifs (croisement,isolement d'un 2n, analyse de tétrades) : si la mutation PseC est située dans un de ces gènesPHO, une forte liaison génétique apparaitra.
ex :
Résultat de l'analyse :
- certaines mutations PseC dominantes sont très étroitement liées au (et donc affectent très vraisemblablement le) gène PHO4, et les autres au gène PHO81 (ces mutations sont désignées Pho4C
et Pho81C ).- aucune des mutations PseC isolées n’est liée à PHO2 ou à PHO5 (!)
Pse -Pse -DP
2n
pho4(Pse - )
mutantPse c
Pse cPse c
Grosse majorité detétrades = DP
PHO4*
*Pse-
PseC
Ex :Si la majorité des méiosesaboutit à un DP, alors onpeut conclure que la mu-tations Psec est généti-quement liée à PHO4 (uni-quement phénotypes pa-rentaux)
13

PHO5PHO5 PHO5PHO5
Pi
+
¯Pi
PHO5PHO5
Pi ¯Pi Pi ¯Pi
Pse+ Psec Psec
Pse+
Un facteur positif active l’expression de PHO5 quand le milieu est pauvre en Pi. Ce facteur est inhibé quand le milieu est riche en Pi. Si une mutation rend ce facteur insensible à cette inhibition, l’expression de PHO5 est constitutive. Et ce défaut n’est pas complémenté par l’apport du gène sauvage codant pour ce facteur positif (phénotype dominant).
Conclusion : intervention de deux facteurs négatifs (Pho80, Pho85) et de trois facteurs positifs (Pho2, Pho4, Pho81), et deux des facteurs positifs (Pho4 et Pho81) sont inhibés quand le milieu est riche en Pi ® cascade de régulation ?
+ + + ++
14

1.3. Etablissement des relations d'épistaticité entre les gènes PHO
® pour ordonner les éléments de régulation Pho
Comment ? analyse du phénotype de doubles mutants Pse- Psec : quel phénotype l'emporte ?
Ex. : Phénotype du bouble mutant pho4 pho80. Comment isoler ce double mutant ? Croisement entre les deux mutants et analyse des quatre ascospores issues de la méiose: si une tétrade DNP est obtenue, cad comprenant 2 spores Pse+, les deux autres spores sont obligatoirement doubles mutantes pho4 pho80
Pse +Pse +DNP
2n
pho4(Pse - )
pho80(Pse c )
ce s deux sporessont obligatoirement
doubles mutantes
pho4 PHO80+ x PHO4+ pho80
pho4 PHO80+
PHO4+ pho80
PHO4 PHO80PHO4 PHO80pho4 pho80pho4 pho80
Si dans une méiose, on trouvedeux cellules Pse +, on peutdéduire que les deux autressont des doubles mutantspho4 pho80 (DNP)
15

Résultat de l’analyse des tests d'épistaticité : phénotype des doubles mutants
Mutation Pse-
Mutation PseC
Phénotype du double mutant
pho2 pho80 Pse- pho2 pho85 Pse- pho2 pho4C Pse-
pho2 est épistatique sur les quatre autres mutations
pho2 pho81c Pse-
pho4 pho80 Pse- pho4 pho85 Pse-
pho4 est épistatique sur les trois autres mutations
pho4 pho81c Pse-
pho81 pho80 PseC pho81 pho85 PseC pho81 pho4C PseC
pho81 est hypostatique par rapport aux trois autres mutations
16

PHO5
Pho4
Pho80, Pho85
Pho81
P
PHO5
Pho80, Pho85
Pho81
P
PHO5
Pho80, Pho85
Pho81
Modèle de régulation du gène PHO5 (Oshima, 1982)
Pho2
Pho4
Pho2
Pho4
Pho2
® modèle de régulation du gène PHO5 (Oshima, 1982)
17

4. Clonage des gènes PHO : clonage par complémentation
Milieu "high Pi"Milieu "low Pi"
mutant Pse c complémenté
mutant Pse - complémenté
Mutant Pse - (pho2, pho4, pho81)
+ mut ation ur a3
Mutant Pse c (pho80, pho85)+ mutation ura3
Transformation des cellules par les plasmides (URA3+)
contenant des segments d'ADN génomique (banque génomique)
extraction de l'ADNet transformation de
E. coli : sélection des clones AmpR
extraction des plasmides:
- re-transformation des mutants- séquençage
banque génomique :
Plasmide navette E. coli – levure = plasmide pUC19 1 dans lequel on a inséré :
- le gène URA3 de sélection dans la levure
- une origine de réplication propre à la levure
L’ensemble des fragments de la banque couvre l’entièreté du génome
Sélection des clones URA3+
(milieu sans uracile)
pho2-
PHO2
pho80 -
PHO80
mutant pho2complémenté :
mutant pho80complémenté :
Ex. pour cloner le gène PHO2
Ex. pour cloner le gène PHO80
levures de génotype pho2 ura3
levures de génotype pho80 ura3
( ura3 = mutation provoquant une auxotrophie pour l’uracile )
1 plasmide contenant ori, AmpR,
mcs, cf. TP
différents fragments d'ADN
génomique
18

pho80 -
PHO80
- extraction et purification de l’ADN - transformation de bactéries E. coli et sélection de colonies AmpR
E. coli
- culture de ces bactéries transformées et purification à partir de ces bactéries du plasmide
- séquençage des deux extrémités du fragment d’ADN génomique inséré dans le plasmide
- comparaison des séquences obtenues avec la séquence génomique complète de la levure 1
1 http://www.yeastgenome.org/
Pour récupérer le plasmide porteur du gène PHO80 :
milieu + ampicilline
19
« miniprep »

S1
S2
Test de complémentation de la
mutation pho80
oui
non
non
oui
nonoui
Le fragment cloné est issu du chr. 15 et contient 8 ORFs : laquelle correspond au gène PHO80 ?
ORF de PHO80 ® séquence d’aa de la protéine Pho80
Conclusion :
Chaq
ue f
ragm
ent
est
insé
ré d
ans
un
plas
mid
e et
réi
ntro
duit
dan
s la
levu
re
20

21
http://www.yeastgenome.org/
22
Régulationde PHO5
Gène Protéineséquence
isolement demutants
clonage du gène& analyse bioinformatique
interprétation(biochimie, biologie cellulaire)
Les protéines Pho impliquées dans la régulation transcriptionnelle de PHO5 étant identifiées, la suite de l’étude visera à comprendre leur mode d’action moléculaire (par des analyses biochimiques, de biologie cellulaire, ..)

® Conclusions de l'analyse :
- Pho2 et Pho4 = activateurs de transcription se liant à des séquences spécifiques en amont du gène PHO5
- Pho85 = kinase de type CDK (cyclin-dependent kinase) activée par la sous-unité Pho80 (protéine de type cycline)
- Le complexe Pho85-Pho80 phosphoryle Pho4 : la protéine Pho4 est alors inactive (car séquestrée dans le cytoplasme)
- Pho81 : se lie au complexe Pho80-Pho85 et inhibe Pho85 (qui ne peut plus se lier à Pho4) dans les conditions de carence en Pi. Pho81 reste lié au complexe mais sans l'inhiber si le Pi est abondant ; Pho4 n'est alors plus phosphorylé, il migre dans le noyau est active la transcription (conjointement avec Pho2) de PHO5
- Inhibition de Pho81 par le Pi ? Une molécule hautement phosphorylée (inositol-7-phosphate, IP7) se lie à Pho81 et change sa conformation > inhibition de Pho81, Pho85 reste alors actif
23
IP7
_

Forward genetics
Fonction è Mutants è Gène è Séquence
Avant les travaux de génomique fonctionnelle :
Reverse genetics
Fonction ç Mutants (knockout) ç Gène ç Séquence
Suite aux travaux de génomique fonctionnelle (ex. isolement des ≈ 6000 mutants de la levure) :
On recherche parmi tous les mutants possibles de levure (cf. chap II, 3.6) ceux qui présentent le phénotype recherché (perte de fonction). Quand un mutant est ainsi trouvé, on sait immédiatement de quel gène il s'agit.
On isole des mutants présentant un phénotype particulier, et on tente d'identifier le gène déficient dans ce mutant
24

25
x ~50
Plaque 96 puits > 1 mutant de levure par puit : collection d’ environ 4800 mutants (viables)
Chaque clone correspond à un mutant où un gène particulier (ici, YKR039w) a été éliminé du génome
Test de coloration sur milieu riche ou pauvre en Pi -> identification de tous les gènes impliqués dans
le contrôle de l’expression du gène PHO5







