Download - Biokémia II

Szarka András
Biokémia II. Biokémiai szabályozás
A Biokémia II. – Biokémiai szabályozás című tankönyv másodéves biomérnök hallgatók
számára készült, akik a Biokémia I. tantárgy során már elsajátították a biokémiai,
molekuláris biológiai alapismereteket. Ezeket alapul véve, ezekre építkezik a tankönyv,
amely kiemelten a biokémiai szabályozásokkal foglalkozik. Az építkezés szó igen találó,
hiszen a szabályozástechnika részelemeinek ismertetését követően egyszerűbb, majd
egyre összetettebb szabályozási problémák megismerése felé haladunk. Így
foglalkozunk az enzimaktivitás különböző szabályozási lehetőségével, mint az
allosztérikus, a foszforilációval és a limitált proteolízissel történő szabályozás, majd
terítékre kerül a fehérjék élettartamának szabályozása. A következő nagyobb blokk a
génkifejeződés témakörét tárgyalja prokarióta, majd eukarióta sejtek esetében. Egyetlen
egy sejt-, szervezetszintű szabályozással foglalkozó tankönyv sem mehet el szó nélkül a
jelátviteli útvonalak mellett, így a harmadik jelentősebb terület a mi esetünkben is ezzel
a területtel foglalkozik. Az első három blokk funkcionális egységet a negyedik blokkban
nyer, amelyben a szabályozási részelemek működését három kiemelt szabályozási
probléma, az éhezés – jól tápláltság, vércukorszint szabályozás, a programozott sejthalál,
illetve a sérült fehérjeválasz révén mutatjuk be.
Kulcsszavak: biokémia, sejtbiológia, szabályozás, anyagcsereutak, enzimek, molekuláris
biológia, centrális dogma, transzkripció, jelátvitel, sejthalál, vércukorszint szabályozás
Budapesti Műszaki és Gazdaságtudományi Egyetem
Semmelweis Egyetem
Typotex Kiadó
2014

COPYRIGHT: 2014-2019, Dr. Szarka András, Budapesti Műszaki és Gazdaságtudományi Egyetem,
Semmelweis Egyetem
Creative Commons NonCommercial-NoDerivs 3.0 (CC BY-NC-ND 3.0)
A szerző nevének feltüntetése mellett nem kereskedelmi céllal szabadon másolható, terjeszthető,
megjelentethető és előadható, de nem módosítható.
Lektorálta: Barna János Ph.D.
ISBN 978-963-279-170-8
Készült a Typotex Kiadó gondozásában
Felelős vezető: Votisky Zsuzsa
Készült a TÁMOP-4.1.2/A/1-11/1-2011-0079 számú, „Konzorcium a biotechnológia és bioinformatika
aktív tanulásáért” című projekt keretében.

© Szarka András www.interkonyv.hu
Tartalomjegyzék
Előszó ....................................................................................................................................................................... 5
1. Bevezetés ........................................................................................................................................................... 6
2. Biokatalizátorok – enzimek ....................................................................................................................... 9
2.1. Néhány szóban az enzimkinetikáról ........................................................................................ 11
2.2. Az enzimes katalízis háttere ........................................................................................................ 13
3. Enzimszabályozás ....................................................................................................................................... 17
3.1. Allosztérikus enzimek .................................................................................................................... 21
3.1.1. Kooperativitás ........................................................................................................................ 23
3.1.2. A hemoglobin szerkezete és oxigénkötése ................................................................. 25
3.2. Fehérjék térszerkezetének, aktivitásának befolyásolása foszforiláció/defoszforiláció által .............................................................................................. 31
3.3. Limitált proteolízis .......................................................................................................................... 36
3.3.1. Inaktív zimogének és aktiválódásuk ............................................................................. 36
3.3.1.1. Pepszinogén .................................................................................................................. 36
3.3.1.2. Enterális proteázok zimogénjei ........................................................................... 37
3.3.1.3. Proteázinhibitorok .................................................................................................... 38
4. Fehérjefolding, fehérjelebontás – ubikvitin-proteaszóma rendszer .................................... 39
4.1. Poszttranszlációs módosulások ................................................................................................. 39
4.1.1. Fehérjefolding ......................................................................................................................... 39
4.1.1.1. Hősokk-fehérjék – molekuláris chaperonok .................................................. 40
4.2. Sérült fehérjék proteolízise ......................................................................................................... 42
4.2.1. Proteaszóma ............................................................................................................................ 42
4.2.1.1. A proteaszóma felépítése ........................................................................................ 42
4.2.2. A „halál csókja” – Ubikvitináció ....................................................................................... 43
4.2.3. Számos fehérje irányított lebontás révén szabályozott ....................................... 45
5. A génexpresszió szabályozása ............................................................................................................... 47
5.1. Génexpresszió, a gén kifejeződése ............................................................................................ 47
5.1.1. Sejtdifferenciáció ................................................................................................................... 48
5.1.1.1. Azonosságok és különbségek a sejtek között................................................. 48
5.1.2. A génexpresszió szabályozásának szintjei ................................................................. 49
5.1.3. A génszabályozó fehérjék DNS-kötő részei ................................................................ 50
5.1.4. DNS-felismerő fehérjeszerkezeti egységek ................................................................ 52
5.1.4.1. Hélix-turn-hélix részletek ....................................................................................... 53
5.1.4.2. DNS-kötő cink-ujj részletek ................................................................................... 55
5.1.4.3. β-redő .............................................................................................................................. 56
5.1.4.4. Leucin-cipzár ................................................................................................................ 56
5.1.4.5. Hélix-loop-hélix részlet ............................................................................................ 57
5.1.5. DNS-kötő fehérjék detektálása ........................................................................................ 59
5.2. A genetikai kapcsolók munka közben ..................................................................................... 60
5.2.1. A triptofán-represszor, mint egyszerű génkapcsoló .............................................. 60

www.interkonyv.hu © Szarka András
5.2.2. A lac-operon: transzkripciós aktiválás/represszálás ............................................ 64
5.3. Az eukarióta génexpresszió szabályozása ............................................................................. 66
5.3.1. Eukarióta génregulációs fehérjék .................................................................................. 67
5.3.2. Eukarióta DNS-reguláló szekvenciák ........................................................................... 68
5.3.3. Génaktivátor fehérjék és a transzkripció .................................................................... 69
5.3.3.1. A génregulációs fehérjék szinergikusan dolgoznak .................................... 72
5.3.3.2. Eukarióta represszorok ........................................................................................... 72
5.3.3.3. Inzulátor DNS-szekvenciák .................................................................................... 74
5.4. Az X-kromoszóma inaktiválása nőkben ................................................................................. 74
5.5. DNS-metiláció .................................................................................................................................... 78
5.5.1. DNS-metiláció a genomi mintázatban .......................................................................... 79
5.5.2. CG-szigetek ............................................................................................................................... 81
6. Sejtkommunikáció ...................................................................................................................................... 83
6.1. Az extracelluláris szignálmolekulák ........................................................................................ 84
6.1.1. A szignálmolekulák hatótávolsága ................................................................................ 85
6.2. Autokrin szabályozás ..................................................................................................................... 87
6.3. Gap junction ........................................................................................................................................ 87
6.4. Plazmamembrán-receptorok ...................................................................................................... 88
6.4.1. Receptor-ioncsatornák ....................................................................................................... 88
6.4.2. Hét transzmembrán szegmenssel rendelkező, G-fehérje-kapcsolt receptorok ..................................................................................................................................... 89
6.4.3. Protein-kinázok, foszfoprotein-foszfatázok .............................................................. 93
6.4.3.1. cAMP-dependens protein-kináz (protein-kináz-A) ..................................... 93
6.4.3.2. A cGMP-dependens protein-kináz (protein-kináz-G) ................................. 94
6.4.3.3. Protein kináz-C ............................................................................................................ 94
6.4.4. Transzmembrán protein kinázok: növekedési faktor-receptor ....................... 95
6.4.5. Foszfoprotein-foszfatázok ................................................................................................. 96
6.5. A cAMP mediátorrendszer ........................................................................................................... 96
6.6. Inozitol-foszfolipid jelátviteli rendszer .................................................................................. 99
6.7. A guanilát-cikláz és a cGMP ...................................................................................................... 102
6.7.1. Nitrogén-monoxid .............................................................................................................. 102
7. Összetett folyamatok szabályozása .................................................................................................. 103
7.1. Éhezés – jól tápláltság ................................................................................................................. 103
7.1.1. A glikolízis – glukoneogenezis szabályozása .......................................................... 103
7.1.2. A glikogén felépítés – lebontás szabályozása ......................................................... 107
7.2. Programozott sejthalál: Apoptózis ........................................................................................ 110
7.3. Sérült fehérjeválasz (UPR) az endoplazmás retikulumban ........................................ 115
Utószó ................................................................................................................................................................ 120
Ábraanyag ........................................................................................................................................................ 121
Felhasznált és ajánlott irodalom ............................................................................................................ 126

Előszó
A Biokémia II. – Biokémiai szabályozás című tankönyv másodéves biomérnök
hallgatók számára készült, akik a Biokémia I. tantárgy során már elsajátították a
biokémiai, molekuláris biológiai alapismereteket. Ezeket alapul véve, ezekre építkezik a
tankönyv, amely kiemelten a biokémiai szabályozásokkal foglalkozik. A
szabályozástechnika, legyen az biológiai vagy más természetű, mindig a legnagyobb
kihívást jelentő területek egyike. A tankönyvet igyekeztünk úgy felépíteni, hogy a
részelemek ismertetését követően egyszerűbb, majd egyre összetettebb problémák felé
haladjunk. Reméljük, sikerült mindezt világos gondolatmenet mentén megtennünk, és
legalább akkora örömet jelent a tankönyv olvasása, a sejtünk titkaiba való bevezetés,
mint a tankönyv kigondolása és megírása jelentett a szerző számára.
Budapest, 2013. nyár
Szarka András

www.interkonyv.hu © Szarka András
1. Bevezetés
A tankönyv elején érdemes tisztázni, hogy mivel is foglalkozik a biokémiai szabályozás-
technika.
A kérdés megválaszolásához vegyük a következő példát. Tekintsünk úgy a sejtre,
mint egy vegyipari üzemre. Nézzük meg, milyen hasonlóságok és analógiák vonhatók az
üzem és a sejt között:
üzem sejt
fal sejtmembrán
különböző specializálódott egységek sejtorganellumok, kompertimentumok
például: energiaellátó központ
raktár
gépgyártó sor
végszerelde
mitokondrium (kloroplaszt-növényekben)
vakuólum (bizonyos értelemben az ER is)
riboszómák
endoplazmás retikulum (ER)
központi irányítás,
információszolgáltatás,
elosztás
sejtmag
1.1. ábra. A sejt mint vegyipari üzem

1. Bevezetés 7
© Szarka András www.interkonyv.hu
Az üzemet nyilvánvalóan el kell látni energiával. A sejt esetében ez az energia
leggyakrabban a glükózból származik. A hulladék anyagokat is el kell távolítani a sejt
(üzem) zavartalan működéséhez. Ilyen hulladék a CO2, illetve egyéb toxikus vegyületek,
amelyek a sejt működése során keletkeznek.
Az üzem életének szigorúan szabályozott körülmények között kell folynia:
a. Szigorúan meghatározott mennyiségű „gépet”, terméket kell előállítania. Sem
a túltermelés, sem a hiány nem megengedett.
b. Az egyes gyártási lépések között sem tolerálható a köztes anyagok hiánya,
feleslege.
c. Az energiafelhasználás (anyagfelhasználás) optimalizálására törekszünk.
Ezek együttesen biztosítják az optimális gépmennyiséget.
d. Fontos az egészséges munkakörülmények biztosítása, a külső vagy belső
gyártás során keletkező mérgek, szennyezőanyagok eliminálása.
Mindezek biztosításához szigorúan össze kell hangolni a részlépéseket, munka-
folyamatokat, az egyes alapanyagok, köztitermékek, késztermékek transzport-
folyamatait.
A sejt esetében ezen folyamatokkal foglalkozik a biokémiai szabályozástechnika.
Tanulmányaink során:
1. megismerjük a sejt gépeit, az enzimeket (mi az enzimek szerepe, milyen fontos
tulajdonságokkal bírnak)
2. a biokémiai alapfolyamatokat több szinten szabályozhatjuk (a sejt működési
hatásfoka [intenzitása], valamint a sejt gépeinek száma is szabályozható)
a) az enzimaktivitás szabályozásának lehetőségei:
i. gátlások
ii. szabályozások
1. allosztérikus enzimek
2. proteolitikus hasítás
3. foszforiláció
a fehérjék születése: – transzláció
– poszttranszlációs módosulások
a fehérjék aktív állapota
a fehérjék bukása –ubikvitizáció → lebomlások, proteaszóma
b) a gépek számának változtatása – az enzimmennyiségek változtatása,
szabályozása
i. sejtdifferenciáció és génexpresszió szabályozása
ii. a génexpresszió szabályozásában résztvevő DNS-t kötő fehérjék:

8 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
1. hélix-turn-hélix
2. cink-ujj fehérje
3. leucin-cippzár
4. hélix-loop-hélix
iii. a DNS-t felismerő fehérjeszakaszok azonosítása
iv. a gének ki- és bekapcsolása
1. lac-operon
2. triptofán-represszor
3. eukarióta sejtek transzkripciós szabályozása
a. transzkripciós faktorok
b. promóterek
c. génregulációs szekvenciák
d. génregulációs fehérjék
v. sejtdifferenciálódás
vi. poszttranszkripciós szabályozások
vii. a sejtszintű információáramlás: szignál-transzdukció
1. általános kitekintés: receptorok, ligandok
2. extracelluláris jeltovábbítás: paraszimpatikus és endokrin
lehetőségek
3. sejtmag receptorok: szteroid receptorok
4. sejtfelszíni receptorok:
a. ioncsatorna-kapcsolt
b. G-fehérjéhez kapcsolt
c. enzimkapcsolt
viii. célsejt adaptáció – receptor leszabályozás
ix. összetett folyamatok
1. glikolízis, glikogén lebontás/felépítés, vércukorszint-
szabályozás
2. apoptózis, sérült fehérjeválasz (UPR)

© Szarka András www.interkonyv.hu
2. Biokatalizátorok – enzimek
Az enzimek szabályozásával kapcsolatos ismeretek megértéséhez szükségünk van
bizonyos termodikai alaptudásra.
Nézzük a következő reakciót:
papír+O2 füst+hamu+hő+CO2+H2O
A papír elég, hőenergiát juttat az atmoszférába, valamint vizet és széndioxidot,
viszont a füst és a hamu ezekkel a dolgokkal és a melegített levegővel sohasem képes
papírrá visszaalakulni. Amikor a papír elég, kémiai energiája, mint hőenergia
disszipálódik. Ezzel egy időben a papír atomjai, molekulái szétszóródnak, rendezetlenek
lesznek. A termodinamika nyelvén mondva szabadenergia-veszteség, szabadenergia-
csökkenés következik be. Ezt az energiát használjuk fel munkavégzésre vagy kémiai
reakciók végrehajtására.
Általánosan elmondhatjuk: a kémiai reakciók mindig a szabadenergia-csökkenés
irányába mennek végbe. Vagyis a reakciók spontán iránya az energetikailag
kedvezőbb irány.
Normál körülmények között a szén energetikailag legstabilabb formája a széndioxid
és a hidrogéné a víz. Az élő szervezetek mégsem válnak széndioxidköddé és a papír sem
kezd el lángolni kezeink között. Ennek oka, hogy az élő szervezet és a füzet molekulái
egy viszonylagos stabil állapotban találhatóak és ahhoz, hogy alacsonyabb
energiaszintre kerüljenek, energiát kell befektetni. Magyarul a molekuláknak aktivációs
energiára van szükségük ahhoz, hogy egy jóval stabilabb (alacsonyabb energiájú)
állapotba kerüljenek (2.1. ábra). Az égő füzet (papír) esetében az aktivációs energiát egy
meggyújtott gyufa jelenti. A vizes oldatban található molekulák számára az energiagáton
történő átjutást a szomszédos molekulákkal történő ütközés teremti meg (amely a
hőmérséklet emelkedésével egyre gyakrabban következik be).

10 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
2.1. ábra. Az enzimek, mint biokatalizátorok
Az energiagát átugrását az élő szervezetben az enzimek segítik. Ez az energiagát-
csökkentés annyira jelentős mértékű lehet, hogy az enzimek akár (több)milliószorosára
is gyorsíthatják a reakciókat. Emiatt szabályozásuk kiemelt fontossággal bír a sejt
életében, metabolizmusában. A reakció végén maguk változatlanok maradnak. Ezért
tekinthetjük őket biokatalizátoroknak. Hasonlóan bármely katalizátorhoz,
kizárólag termodinamikailag lehetséges reakciókat katalizálnak, mivel „csak”
az aktivációs energiát csökkentik, ezáltal hihetetlenül megnövelve a
reakcióba lépő molekulák számát (2.1. ábra);
tekintve, hogy általában fehérjemolekulák, reakcióra és/vagy szubsztrátra
specifikusak;
(Hasonlóan bármely katalizátorhoz) Maguk nem változnak a reakció során.
A hasonló reakciókat katalizáló enzimeket egyazon funkcionális csoportba soroljuk.
1. Oxidoreduktázok pl.: etanol + NAD+ acetaldehid + NADH + H+
2. Transzferázok pl.: glukóz + ATP glukóz-6 foszfát + ADP
3. Hidrolázok pl.: glukóz-6-foszfát + H2O glukóz + Pi
4. Liázok pl.: 2-foszfoglicerát foszfoenol-piruvát + H2O
5. Izomerázok pl.: glukóz-6-foszfát fruktóz-6 foszfát
6. Ligázok pl.: glutamát + NH3 + ATP glutamin + ADP + Pi

2. Biokatalizátorok – enzimek 11
© Szarka András www.interkonyv.hu
A csoporton belül minden enzim specifikus, csak egy adott reakciót katalizál
viszonylag szűk szubsztrát tartományon belül (gyakran csak egyetlen meghatározott
molekula [szubsztrát] átalakítására képes).
Például: A hexokináznak a D-glukóz szubsztrátja, de az L-glukóz már nem
szubsztrátja.
A trombin a fibrinben csak egy adott arginin és a szomszédos glicin között
hasít, máshol nem.
Természetesen ez a specifikusság különböző mértékű lehet, előfordul, hogy egy adott
reakciótípusra, vagy adott funkciós csoportra korlátozódik csak, de ismertek kizárólag
egyszubsztrátos enzimek is.
Az enzimek hálózatot alkotnak. Az egyik enzim terméke igen gyakran egy másik
szubsztrátja és így tovább. Ennek eredménye az anyagcsere utak hálózata, az
energiaellátás biztosítása és a különböző molekulák szintézise.
2.1. Néhány szóban az enzimkinetikáról
Az enzimes reakciókkal kapcsolatos, alapvető reakciókinetikai összefüggéseket, törvény-
szerűségeket a következő egyszerű, egyszubsztrátos enzimes reakción szeretnénk
bemutatni:
Az adott idő alatt az enzim által megkötött és átalakított szubszrátmolekulák száma
behatárolt.
Ha növeljük a szubsztrátok számát, egy maximum értékig növekszik az átalakított
szubsztrát/keletkezett termék mennyisége. Ebben a pontban az enzimünk telítődött
szubsztrátjával és a reakció sebessége (Vmax) csak attól függ, hogy milyen gyorsan képes
az enzim a szubsztrátot átalakítani. A maximális sebességet az enzim koncentrációjával
osztva megkapjuk az enzim átviteli számát, mely az angolszász irodalomban turnover
number néven ismeretes. Ennek értéke egy és tízezer között szokott lenni, általában ezer
körüli.
Az enzimműködés kvantitatív jellemzése elengedhetetlen az enzimek megismerése
során, illetve széleskörű, irányított ipari felhasználásukhoz.
Tekintsük ismételten a következő egyszubsztrátos reakciót:

12 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
A reakcióval kapcsolatban a következő egyszerűsítéseket tesszük:
Feltételezzük, hogy az enzim (E) és a termék (P) között annyira ritkán játszódik le
enzim-szubsztrát komplexet (ES) eredményező reakció, hogy ezt el is hanyagolhatjuk.
Ekkor az EP komplexet nem szükséges megjelenítenünk és a reakció előrehaladását,
azaz a sebességét a következőképp fejezhetjük ki:
V = kcat [ES]
Ebben az esetben [ES] az enzim-szubsztrát komplex koncentrációja, kcat, az átviteli
szám, ami megadja, hogy egy enzimmolekula egy másodperc alatt hány szubsztrát-
molekulát alakít át. Az ES komplex koncentrációja az enzim és szubsztrát molekulák
összekeverését követően hirtelen nő, míg el nem éri az egyensúlyi állapotot (steady
state). Ebben az állandósult állapotban koncentrációja jó közelítéssel állandónak vehető:
ES fogyás = ES képződés
k-1[ES] + kcat [ES] = k1 [E][S]
A szabad enzim koncentrációja: [E] = [E0] - [ES]
Behelyettesítve:
Vezessük be a Km Michaelis-konstanst:
Km =
Ekkor
Illetve visszaírva: V = kcat [ES]
Megkapjuk a híres Michaelis–Menten-egyenletet:
Ha növeljük a szubsztrát mennyiségét, egy maximum értékig növekszik az átalakított
szubsztrát/keletkezett termék mennyisége. Ebben a pontban az enzimünk telítődött
szubsztrátjával (tehát minden enzimmolekula szubsztrátot köt) és a reakció sebessége
Vmax. Ezt a sebességet abban a pontban érjük el, ahol:
2. Biokatalizátorok – enzimek 13
© Szarka András www.interkonyv.hu
V = Vmax = kcat [E0]
Ez alapján a Michaelis–Menten-egyenlet általános formája a következő:
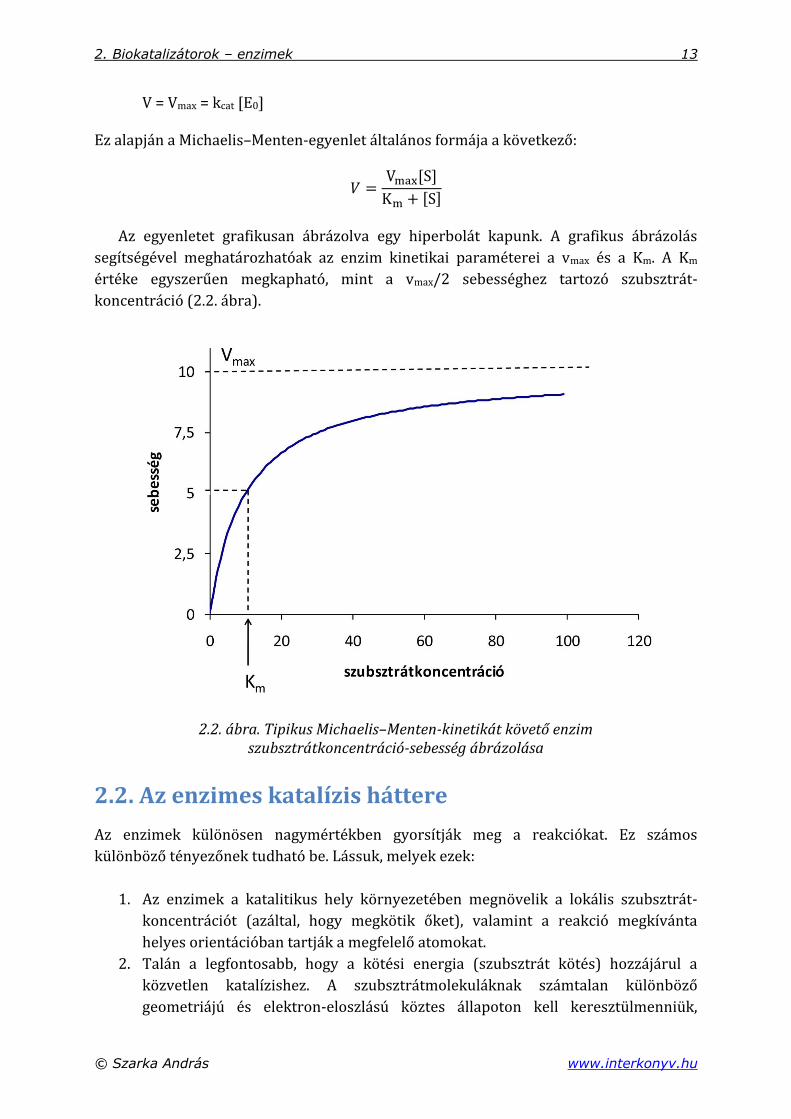
Az egyenletet grafikusan ábrázolva egy hiperbolát kapunk. A grafikus ábrázolás
segítségével meghatározhatóak az enzim kinetikai paraméterei a vmax és a Km. A Km
értéke egyszerűen megkapható, mint a vmax/2 sebességhez tartozó szubsztrát-
koncentráció (2.2. ábra).
2.2. ábra. Tipikus Michaelis–Menten-kinetikát követő enzim szubsztrátkoncentráció-sebesség ábrázolása
2.2. Az enzimes katalízis háttere
Az enzimek különösen nagymértékben gyorsítják meg a reakciókat. Ez számos
különböző tényezőnek tudható be. Lássuk, melyek ezek:
1. Az enzimek a katalitikus hely környezetében megnövelik a lokális szubsztrát-
koncentrációt (azáltal, hogy megkötik őket), valamint a reakció megkívánta
helyes orientációban tartják a megfelelő atomokat.
2. Talán a legfontosabb, hogy a kötési energia (szubsztrát kötés) hozzájárul a
közvetlen katalízishez. A szubsztrátmolekuláknak számtalan különböző
geometriájú és elektron-eloszlású köztes állapoton kell keresztülmenniük,

14 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
mielőtt a reakció végét jelentő formába jutnak. A legstabilabb átmeneti állapot
felvételéhez szükséges szabadenergia-mennyiség az aktivációs energia. Igazából
ez határozza meg a reakció sebességét. Az enzimek jóval nagyobb affinitással
bírnak az átmeneti állapotú szubsztrát, mint a stabil forma irányába. Ez a szoros
kötés nagymértékben csökkenti az aktivációs energiát, így jelentősen
meggyorsítja a reakciót.
Az enzimek tehát többféle generális stratégiát vetnek be a katalízis során. A
legfontosabbak:
a) az enzim megköti és pontosan orientálja egymáshoz a szubsztrátokat
b) a szubsztrát megkötésével az enzim átrendezi annak elektroneloszlását,
részlegesen + és - részeket eredményezve
c) az enzim megfeszíti a megkötött szubsztrátmolekulát, ezzel az átmeneti állapot
felé tolva
2.4. ábra. Az enzimek legfontosabb katalitikus stratégiái
Katalitikus antitestek
Az orientációs hatás jelentőségét segítenek megérteni az ún. katalitikus antitestek. Ezek,
az enzimhez hasonló térszerkezetük révén, stabilizálják az átmeneti állapotú
molekulákat, így meggyorsítva az alapreakciót, igaz, nem az enzimhez hasonló
mértékben (kb. 10 000- szeresére).
Természetesen az enzimek más úton-módon is hozzájárulnak a reakció minél
gyorsabb végbemeneteléhez. Pl. pontosan pozícionált atomokat tartalmaznak, amelyek
képesek a szubsztrát (átmeneti állapotú molekula) elektronszerkezetét deformálni.
A következőkben a lizozim segítségével nézzünk meg egy konkrét példát az enzimes
katalízisre.
A lizozim tulajdonképpen egy természetes antibiotikum. Megtalálható a tojás-
fehérjében, a nyálban, a könnyben, illetve egyéb szekrétumokban. A bakteriális sejtfal
poliszacharidjait bontja, egészen pontosan az 1-4-es glikozidos kötéseket az N-acetil-
muraminsav és az N-acetilglukózamin között. Ennek hatására a baktériumok nem
képesek ellenállni a turgornyomásnak és szétdurrannak.
Az enzimet még Fleming fedezte fel és nevezte el 1922-ben. Kisméretű (14602 Da),
könnyen izolálható. Így nem csoda, hogy ez volt az első enzim, amelynek röntgen-
diffrakciós szerkezete ismertté vált.

2. Biokatalizátorok – enzimek 15
© Szarka András www.interkonyv.hu
2.5. ábra. A lizozim katalitikus mechanizmusa
A lizozim által katalizált reakció, poliszacharidláncok hidrolízise, egy energetikailag
kedvező, tehát szabadenergia-csökkenéssel járó reakció. Az enzim távollétében azonban
vígan elvannak ezek a poliszacharid láncok még vizes közegben (oldatban) is, bármilyen
észrevehető bomlás, hidrolízis nélkül. Ez érthető, hisz a korábban említett energiagát
jelen esetben is érezteti hatását. A két cukormolekulát összekötő kötés kizárólag abban
az esetben hasad fel a hozzá ütköző vízmolekula hatására, ha a poliszacharidlánc adott
szerkezeti torzuláson megy keresztül, felveszi az átmeneti állapotot, amelyben a kötés
körüli atomok geometriája és elektroneloszlása megváltozik. Ez random módon csak
igen ritkán következik be, azonban jelentősen megváltozik a helyzet, amennyiben az
oldat lizozimet is tartalmaz. A lizozim aktív helye, tekintve, hogy szubsztrátja egy
polimer, egy hosszanti árok, amely egyszerre hat egymáshoz kapcsolt cukrot tud
megkötni. A lizozim sajátságos módon tartja a szubsztrátját, úgy, hogy kifordítsa a
kötésben részt vevő cukrok egyikét, kitörve a stabilis konformációjából. A felhasítandó
kötést úgy pozícionálja, hogy közel kerüljön az aktív hely két savas karakterű
aminosavához (egy glutamáthoz és egy aszpartáthoz) (2.5. ábra). Az enzim az enzim-
szubsztrát komplexben a bal oldali cukrot kifeszített, egyenes konformációban tartja,
miközben a 35-ös glutamáttal mint savval támadja a cukor-cukor kötést, protont
donálva a jobb oldali cukornak, illetve ellensúlyként az 52-es aszpartáttal támadja a C1-
szénatomon (2.5. ábra). Ennek következtében kialakul egy kovalans kötés az 52-es
aszpartát és a cukormolekula C1-szénatomja között. Mindeközben a 35-ös glutamát egy
vízmolekulát (pirossal jelölve) polarizál, hogy annak oxigénatomja megtámadhassa a C1-
szénatomot, így kiváltva az 52-es aszpartátot. A vízmolekula (pirossal jelölve) reakciója
befejezvén a hidrolízist, visszaállítja az enzim eredeti állapotát, miközben létrejön az
enzim-termék komplex (2.5. ábra).
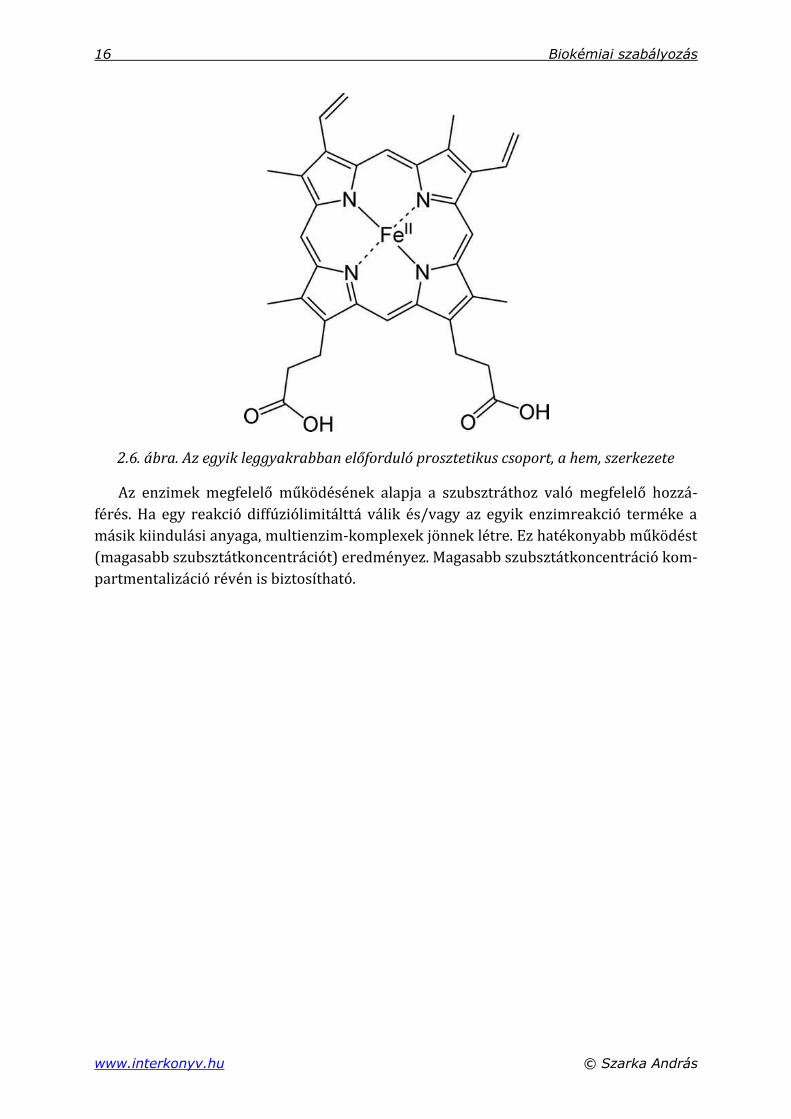
Az enzimek működésükhöz nem fehérje és nem aminosav részeket is igényelnek.
Ezek egy része kovalensen kapcsolódik az anya fehérjelánchoz. Ezeket nevezzük
prosztetikus csoportoknak. A fehérjeláncokhoz kapcsolódó nem aminosav alkotók
lehetnek fémionok vagy szerves molekulák (koenzimek, lipidek, cukrok) (2.6. ábra).
16 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
2.6. ábra. Az egyik leggyakrabban előforduló prosztetikus csoport, a hem, szerkezete
Az enzimek megfelelő működésének alapja a szubsztráthoz való megfelelő hozzá-
férés. Ha egy reakció diffúziólimitálttá válik és/vagy az egyik enzimreakció terméke a
másik kiindulási anyaga, multienzim-komplexek jönnek létre. Ez hatékonyabb működést
(magasabb szubsztátkoncentrációt) eredményez. Magasabb szubsztátkoncentráció kom-
partmentalizáció révén is biztosítható.

© Szarka András www.interkonyv.hu
3. Enzimszabályozás
A korábbiakban láttuk, hogy egy-egy enzim több milliószorosára, akár milliárdszorosára
képes az általa katalizált reakció sebességét megnövelni. Ha figyelembe vesszük, hogy
egy élő sejt az enzimek ezreit tartalmazza a kis citoszolban vagy más kis térfogatú
kompartimentumban, továbbá, hogy ezek egy összetett metabolikus hálózatot alkotnak,
melyben igen gyakran az egyik enzim terméke a másik szubsztrátja, továbbá, hogy ez a
hálózat az elágazási pontok tömkelegét tartalmazza, beláthatjuk, hogy mindezek
következtében pontos, precíz szabályozásra van szükség a problémamentes anyagcsere,
sejtműködés lebonyolításához. Az anyagcsere összetettségéről és a szabályozási feladat
által képviselt kihívásról képet kaphatunk, ha a 3.1. ábrára tekintünk, melyen néhány
unatkozó biokémikus összefoglalta a sejtben folyó biokémiai reakciókat.
3.1. A sejtben folyó biokémiai reakciók összessége
A sejt életfolyamatainak szabályozása tehát igen jelentős mértékben az azokat kata-
lizáló enzimek mennyiségének, illetve aktivitásának szabályozására vezethető vissza.

18 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
A szabályozás több szinten valósulhat meg:
1. Befolyásolható az enzimmolekulák száma – ez majdnem kizárólagosan a gén-
expresszió szabályozása révén történik.
2. A sejt specializált kompertimentumokba telepíti a folyamatokat (enzimeket), itt a
szubsztrátkoncentrációt a citoszoltól eltérően alakíthatja.
3. Célzott fehérjelebontással az enzim (fehérjék) féléletidejét befolyásolhatja.
4. A leggyorsabb és legáltalánosabb módszer a reakció sebességének befolyáso-
lására az enzimaktivitás közvetlen és reverzibilis befolyásolása speciális mole-
kulák segítségével, amelyek végső soron a fehérje térszerkezetének változásán
keresztül fejtik ki hatásukat.
A szabályozási szintek tárgyalását a legáltalánosabb 4. pontban ismertetett esettel, a
már elkészült fehérjemolekulák aktivitásának befolyásolásával kezdjük. Kulcsmondatunk
a következő lesz: A fehérjék térszerkezete és funkciója szinonim fogalom.
Ez a mondat igen jól összefoglalja a fejezet lényegét, amennyiben befolyásoljuk,
megváltoztatjuk egy fehérje térszerkezetét, akkor, az maga után vonja annak funkció- és
aktivitásváltozását is!
Az ilyen jellegű szabályozás igen gyakran feed-back gátlás formájában valósul meg.
Feed-back gátlás során az enzimet, amely a metabolikus út egy korai szakaszán
található, egy a metabolikus út későbbi szakaszának terméke gátolja. Így amikor a
termék elkezd felgyülemleni, az gátolja az első enzimet, limitálva (megakadályozva),
hogy további szubsztrátmolekulák lépjenek be a metabolikus útvonalba (3.2. ábra).

3. Enzimszabályozás 19
© Szarka András www.interkonyv.hu
3.2. ábra. Feed-back gátlás. A metabolikus út egy korai szakaszán található enzimet a metabolikus út egy későbbi szakaszának terméke gátolja
Amikor az útvonalak elágaznak vagy egymást keresztezik, akkor többszörös szabá-
lyozópontok alakulnak ki, amelyeket a következő végtermékek kontrollálnak, mindegyik
saját szintézisét befolyásolva (3.3. ábra).

20 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
3.3. ábra. Többszörösen elágazó (aminosav) metabolikus út szabályozása feed-back gátlással
A feed-back inhibíció azonnal működésbe lép és gyorsan megszűnik, amint a termék
szintje csökken. Amennyiben ez a típusú szabályozás csökkenti az enzim aktivitását,
negatív szabályozásról beszélünk (3.2. ábra).
Amennyiben a szabályozómolekula növeli az enzim aktivitását, pozitív szabályo-
zásról beszélünk. Ebben az esetben a metabolikus útvonal egyik ágának terméke egy
másik metabolikus útvonal enzimét aktiválja. Pl. az ADP feldúsulása aktiválja a cukor
lebontásában résztvevő enzimeket, hogy az ADP ATP-vé alakulását elősegítse.
Gyakran előfordul, hogy az anyagcsereút elő anyaga aktiválja a belépést katalizáló
enzimet.

3. Enzimszabályozás 21
© Szarka András www.interkonyv.hu
Egymással ellentétes folyamatok esetén egy bizonyos molekula az egyik út számára
allosztérikus aktivátorként, míg a visszafelé folyó út enzime számára allosztérikus
inhibitorként viselkedik, így biztosítva az ellentétes utak koordinálását.
3.1. Allosztérikus enzimek
A szabályozás elnevezése igen találó magában hordozza, annak lényegét:
Allos= másik
Steros=szilárd vagy háromdimenziós (térbeli)
Egy szubsztráttól különböző molekula az enzim egy regulációs helyéhez kötődik
(amely hely nem azonos az aktív hellyel), ezáltal befolyásolva annak aktivitását. A szabá-
lyozómolekula alakja gyakran teljesen eltérő a szubsztráthoz képest. Tehát az aktív
helyhez kötődik a szubsztrát, a regulátormolekula pedig a regulációs helyhez. A két
kitüntetett helynek kommunikálnia kell, hogy a regulátormolekula bekötése befolyásol-
hassa az aktív helyet.
Ez a kommunikáció a fehérje térszerkezet-változásán keresztül valósul meg. A bekö-
tődő szabályozómolekula konformáció változást, ez pedig aktivitásváltozást eredményez.
Úgy gondoljuk, hogy a fehérjék jelentős része allosztérikus:
– enzimek,
– receptorok,
– szerkezeti fehérjék,
– motorfehérjék.
A ligandok azt a konformációt stabilizálják, amelyikhez a legerősebben kötődnek, így
kellően magas ligandkoncentrációnál a fehérje ezen konformációja lesz „bekapcsolva”.
Az alaposabb megértéshez vegyük a következő példát.
Adott egy fehérje, amelynek két kötőhelye van elkülönülten. Az egyikkel a
szubsztrátját, a glukózt, a másikkal egy regulátormolekulát, X-et köt.
Ha a glukóz bekötődése megváltoztatja az X-kötőhely alakját, akkor a két kötőhely
kapcsolt.
Ha mindkét ligand ugyanazon fehérje konformációt részesíti előnyben, akkor
bármelyik bekötése növeli a fehérje másik irányába mutatott affinitását. Tehát ha a zárt
forma köti legjobban a glukózt és ez a zárt forma szintén kedvezőbb az X-kötőhely
számára, hogy X-et kössön, akkor a fehérje szorosabban köti a glukózt, ha X jelen van,
mintsem ha nincs. A szabályozás ebben az esetben pozitív szabályozás (3.4. ábra).

22 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
3.4. ábra. Pozitív allosztérikus reguláció
Következésképpen a kötőhelyek összekötöttsége negatívan befolyásolja a kötődést,
ha a két különböző molekula a különböző konformációkhoz szeret jobban kötődni.
Ebben az esetben az első ligand bekötődése csökkenti a második (másik) ligand
bekötődésének valószínűségét (3.5. ábra).
3.5. ábra. Negatív allosztérikus reguláció

3. Enzimszabályozás 23
© Szarka András www.interkonyv.hu
A kötöttség mértéke mennyiségileg oda-visszaható. Ha a glukóznak nagy hatása van
X kötődésére, akkor X-nek is nagy hatása van a glukóz bekötődésére. Mivel X-molekula
nem a katalitikus (aktív) helyhez kötődik, nem szükségszerű, hogy bármilyen kémiai
kapcsolat legyen közte és a glukóz vagy bármelyik másik ligand között, amely az aktív
helyhez kötődik. Ahogy láttuk az X-molekula egyszerűen képes bekapcsolni (+ reguláció)
vagy kikapcsolni (- reguláció) az enzimet.
Ezáltal az allosztérikus fehérjék általános kapcsolóként működnek, amelyek
megteremtik a lehetőséget, hogy egy molekula egy másik molekula sorsát meghatározza
a sejten belül.
3.1.1. Kooperativitás
Egy alegységes enzim feed-back inhibíciója során 90% aktivitásról 10%-ra szabályo-
zódik vissza, ha az inhibitor koncentráció 100-szorosára nő.
Ez nem ad lehetőséget éles szabályozásra. A legtöbb ligand által be- és kikapcsolható
enzim több egymással megegyező enzimatikus alegységből áll. Ebben az elrendeződés-
ben, ha egy ligand beköt az egyik alegységen levő kötőhelyre, kivált egy allosztérikus
változást az alegységen belül, amely később átterjedhet a szomszédos alegységekre, elő-
segítve őket, hogy ugyanezt a ligandot megkössék. Ennek eredményeképp egy koope-
ratív allosztérikus átalakulás jön létre (3.6. ábrán kék vonallal jelölve), lehetővé téve,
hogy a sejtben bekövetkező kisarányú ligand koncentráció-változás teljesen aktiválja,
vagy teljesen inaktiválja az egész apparátust (enzimet, fehérjét).
3.6. ábra. Egy- és több alegységes enzimek inhibitor koncentráció – relatív enzimaktivitás összefüggése
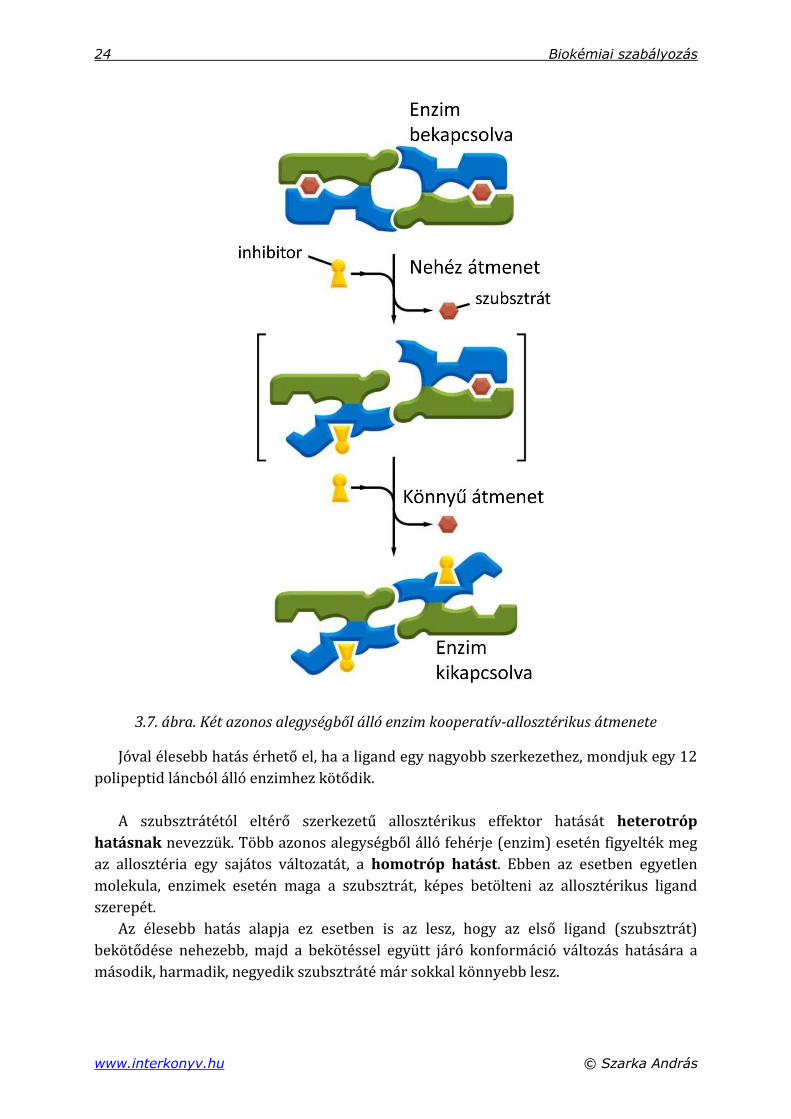
Az első ligand bekötése nehezebb, mivel ennek során egy energetikailag kedvező
kapcsolat bomlik fel az alegységek között, viszont a második ligand bekötődése jóval
könnyebb, mivel ez visszaállítja a szimmetrikus molekula monomer-monomer kapcso-
latát (és ezzel egyidejűleg teljesen inaktiválja az enzimet) (3.7. ábra).
24 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
3.7. ábra. Két azonos alegységből álló enzim kooperatív-allosztérikus átmenete
Jóval élesebb hatás érhető el, ha a ligand egy nagyobb szerkezethez, mondjuk egy 12
polipeptid láncból álló enzimhez kötődik.
A szubsztrátétól eltérő szerkezetű allosztérikus effektor hatását heterotróp
hatásnak nevezzük. Több azonos alegységből álló fehérje (enzim) esetén figyelték meg
az allosztéria egy sajátos változatát, a homotróp hatást. Ebben az esetben egyetlen
molekula, enzimek esetén maga a szubsztrát, képes betölteni az allosztérikus ligand
szerepét.
Az élesebb hatás alapja ez esetben is az lesz, hogy az első ligand (szubsztrát)
bekötődése nehezebb, majd a bekötéssel együtt járó konformáció változás hatására a
második, harmadik, negyedik szubsztráté már sokkal könnyebb lesz.

3. Enzimszabályozás 25
© Szarka András www.interkonyv.hu
A pozitív homotróp hatás érvényesülése esetén a hiperbolikus telítési görbe torzul:
szigmoid lesz. Kis szubsztrát koncentráció esetén kicsi a kötődés valószínűsége, a
szubsztrát koncentráció növekedtével (és a konformáció változással) a kötődési esély
megnő. Ennek eredménye lesz a szubsztrát koncentráció emelkedésével meredeken
emelkedő telítési görbe.
A kooperativitás egyértelmű előnye tehát a szűkebb ligand/szubsztrát koncentráció-
tartományban bekövetkező éles aktivitásváltozás. Ez pedig igen komoly regulációs
előnyt jelent.
A kooperativitás és az allosztérikus szabályozás bemutatására a legjobb példát a
hemoglobin oxigénkötése adja. Így a következőkben ezzel részletesen is foglalkozni
fogunk.
3.1.2. A hemoglobin szerkezete és oxigénkötése
A hemoglobin szerkezetét és oxigénkötésének sajátságait, szabályozását egy másik
oxigén kötésére képes molekula, a mioglobin szerkezetével, oxigénkötési sajátságaival
együtt tárgyaljuk. Tesszük ezt azért, mert a párhuzamos tárgyalási mód igen fontos
biokémiai szabályszerűségek megvilágítására ad módot. A két molekula igen hasonló
szerkezettel rendelkezik, mindkettő globin fehérjeláncból és a hozzá kovalensen
kapcsolódó hem prosztetikus csoportból áll. Igen fontos különbség azonban, hogy míg a
mioglobin monomer, addig a hemoglobin tetramer (3.8. és 3.9. ábra).
3.8. ábra. A mioglobin monomer szerkezete

26 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
3.9. ábra. A hemoglobin tetramer szerkezete
Mielőtt a két oxigénkötésre képes molekula szerkezetének mélységeibe merülnénk,
ejtsünk néhány szót az oxigéntárolás/szállítás szerepéről. Anyagcserénk oxigén nélkül
elképzelhetetlen lenne, ez könnyen belátható, ha csak a terminális oxidáció irdatlan
oxigénigényére gondolunk. A hemoglobin vitathatatlan szerepét az oxigénszállításban
aláhúzza a megfigyelés, mely szerint a hemoglobinmentes vér oxigénkoncentrációja 5 ml
oxigén/liter, mindez hemoglobinnal 250 ml oxigén/liter.
A hemoglobin, mint minden hemoprotein, vasat (Fe[II]) és az azt koordináló
protoporfirin-IX vázat tartalmaz. Ez a váz adja a hemoproteinek jellegzetes színét. A vas
koordinációs kötései közül 4 a protoporfirin gyűrű N-atomjaival jön létre, 2 további
kötés a hem síkjának két oldalán. Egy a globinlánc egy (proximális) hisztidinjével jön
létre, ebben az irányban kiemelkedik a dezoxi-hemoglobinban a porfinváz síkjából a
vasatom. A másik oldalon található az oxigén kötőhely. Itt érdemes megjegyeznünk, hogy
a szén-monoxid igen veszélyes, mivel 300-szor jobban kötődik a hemoglobinhoz, mint az
oxigén. Fontos tudnunk, hogy kizárólag a ferro(II)-hemoglobin köt oxigént, a ferri (III)
vagy más néven methemoglobin nem képes oxigént kötni.
Ha végignézzük a hemoproteinek funkcióit, megállapíthatjuk, hogy az oxigénszállítás,
kötés (Hb, Mb) Fe (II) állapotban valósul meg, azonban az elektrontranszfer-
reakciókban résztvevő hemoproteinek (pl. citokrómok) esetében fontos szerepet kap a
Fe (II) – Fe (III) átmenet, hasonlóan a redoxireakciókhoz (pl. citokróm P450 enzimek),
ahol a Fe (II) megköti az oxigént majd redukálja, miközben Fe (II) – Fe (III) átmenet
következik be.

3. Enzimszabályozás 27
© Szarka András www.interkonyv.hu
A rövid kitérőt követően térjünk vissza a két molekula térszerkezetéhez. A
hemoglobint tehát 4 polipeptidlánc alkotja, melyek mindegyikéhez egy-egy hem tartozik.
Ezekhez egy-egy oxigénmolekula kötődhet. A felnőtt hemoglobin A-t (adult) két alfa és
két béta lánc alkotja (3.9. ábra). A magzati hemoglobin F (foetalis) két alfa és két gamma
láncból áll. A láncokat ionos és apoláris kötések tartják össze. Ha egy pillantást vetünk a
mioglobin és a hemoglobin oxigéntelítési (szaturációs) görbéjére (3.10. ábra) megálla-
píthatjuk, hogy azok jelentős különbséget mutatnak, míg a mioglobin klasszikus hiper-
bolikus telítési görbével jellemezhető, addig a hemoglobin telítési görbéje szigmoid.
3.10. ábra. A hemoglobin és a mioglobin oxigéntelítési (szaturációs) görbéje
Az ábrát figyelmesebben tanulmányozva megállapíthatjuk, hogy a mioglobin
alkalmatlan az oxigénszállításra, mivel a szövetekben nem képes leadni azt. Láthatjuk,
hogy a hiperbolikus telítési görbe miatt a tüdőre jellemző oxigén parciális nyomáson
telítődik (közel 100%-os szaturáció) oxigénnel, azonban a szövetekre jellemző parciális
nyomáson még mindig igen magasan halad a görbe. A parciális nyomáskülönbségek csak
igen csekély mértékű oxigén leadását tennék lehetővé (3.10. ábrán magenta nyilakkal
jelölt). Nagyobb mennyiségű oxigén leadása csak igen csekély oxigén parciális nyomás
mellett lehetséges. Ezzel függ össze a mioglobin fiziológiás szerepe: az izmokban történő
oxigéntárolás. Oxigén tárolására kifejezetten kedvező ez a karakterisztika.
A mioglobinnal szemben a hemoglobin szigmoid telítési görbével jellemezhető, így a
tüdőben telítődik oxigénnel (közel 100%-os a szaturáció ebben az esetben is), a
szövetekben tapasztalható alacsonyabb parciális nyomáson pedig leadja azt (3.10. ábrán

28 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
kék nyíllal jelölve). A hemoglobin így ideális jelölt az oxigén szállítására. Mi állhat az
eltérő karakterisztika hátterében? Ahogy azt korábban említettük, a fehérjék szerkezete
és funkciója között szoros összefüggés van. A mioglobin monomer, így a szokásos
hiperbolikus telítési görbével jellemezhető, a hemoglobin azonban tetramer, melynek
láncai között érvényesül a kooperativitás, ami szigmoiddá teszi telítési görbéjét.
Lássuk csak, hogyan támogatja a hemoglobin szerkezete a kooperativitás megvaló-
sulását! Amennyiben a hemoglobin négy alfa vagy négy béta-láncból állna, hiperbolikus
oxigéntelítési görbéje lenne. A deoxihemoglobinban (oxigént nem kötő forma) a vasatom
0,06 nm-rel kilóg a porfiringyűrű síkjából (3.11. ábra). Az oxigénkötés megváltoztatja a
hemoglobin negyedleges szerkezetét. Az oxigén megkötésével a vasatom behúzódik a
gyűrűbe, a vele kovalensen összekötött fehérjelánc szerkezete is megváltozik, ezért
megváltoznak az alfa- és béta-láncok közötti H-híd-kötések és apoláris kölcsönhatások,
valamint felszakad a deoxihemoglobint stabilizáló 8 ionpár.
3.11. ábra. A hemoglobin oxigénkötésre elszenvedett térszerkezet-változása
A deoxihemoglobinban tehát ionpárok kötik össze a globinláncokat (Tense forma),
míg az oxihemoglobinban (Relaxed forma) ezek felszakadnak. Így már érthető, hogy
miért lesz szigmoid a hemoglobin oxigéntelítési görbéje, hisz a feszített deoxi-formát
stabilizáló ionpárok akadályozzák az oxigén megkötését. (A deoximioglobin és az
oximioglobin szerkezete hasonló.) Felmerül a kérdés, hogy mi stabilizálja a
deoxihemoglobin negyedleges szerkezetét?

3. Enzimszabályozás 29
© Szarka András www.interkonyv.hu
A válasz megadásában segít a megfigyelés, mely szerint a hemoglobinoldatban
nagyobb affinitással köti az oxigént, mint a vörösvértesten belül. Tehát a sejten belül
valami lerontja a hemoglobin oxigén iránti affinitását.
Ez a „faktor” egy glikolízis-mellékreakció során képződő, a vörösvértestben nagy
mennyiségben termelődő kis molsúlyú anyag, a 2,3-biszfoszfoglicerát (3.12. ábra).
3.12. ábra. A 2,3-biszfoszfoglicerát
Nézzük meg, pontosan milyen szerepet tölt be a 2,3-biszfoszfoglicerát a hemoglobin-
oxigénkötés szabályozásában! A 2,3-biszfoszfoglicerát ekvimorális arányban kötődik a
deoxihemoglobinhoz, 1 tetramer hemoglobin-molekula 1 molekula 2,3-biszfoszfoglice-
rátot (2,3-BPG) köt. A 2,3-BPG-nek fiziológiás viszonyok között 5 negatív töltése van
(3.12. ábra). A hemoglobin 2,3-BPG kötőhelyét a béta-láncok 4 His, illetve 2 Lys
aminosava alakítja ki. A pozitív töltésű aminosav-oldalláncok és a negatív töltésű 2,3-
BPG közötti kölcsönhatások olyan kötődést hoznak léte, amely stabilizálja a deoxi-Hb
negyedleges szerkezetét (3.13. ábra).
A 2,3-BPG az oxihemoglobinhoz nem kötődik, az oxigenálódás folyamán a láncok
elmozdulása miatt a BPG kötőhely megváltozik, szűk lesz a 2,3-BPG számára. Így az 2,3-
BPG tulajdonképpen a hemoglobin allosztérikus regulátora.
3.13. ábra. A hemoglobin és a 2,3-biszfoszfoglicerát kapcsolata

30 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
A 2,3-BPG koncentrációja nagyjából 4,5 mM és szabályozott; a szabályozás pontos
mechanizmusa jelenleg ismeretlen.
A glikolízisben résztvevő egyes enzimek szintézisének zavarai a vörösvértestekben
befolyásolják a 2,3-BPG szintet. Így hexokináz-hiányos állapotokban a 2,3-BPG szint
csökken, ezért az oxigéntelítési görbe balra tolódik, a hemoglobin oxigénaffinitása nő.
Piruvát-kináz hiányos állapotokban a görbe jobbra tolódik, mert a 2,3-BPG
koncentráció emelkedett, a hemoglobin oxigénaffinitása csökken.
A krónikus hipoxia esetén a 2,3-BPG szint megemelkedik 4,5 mM-ról 8,0 mM-ra.
Magaslati levegőn (4500 m) 2 nap alatt 7,0 mM-ra emelkedik. Tulajdonképpen ezt
használják ki a hegymászók és ezért építenek akklimatizációs táborokat. Ilyenkor, pár
nap alatt megemelkedik a vér 2,3-BPG szintje, aminek hatására a hemoglobin
oxigénkötése leromlik, így nagyobb lesz a tüdő és szövetek telítettsége közötti
különbség, jobbra tolódik a hemoglobin oxigéntelítési görbéje és jóval nagyobb
mennyiségű oxigént tud leadni a szövetekben. Tengerszintre visszatérve a 2,3-BPG
szintje viszonylag gyorsan visszaáll.
A tárolt vér 2,3-BPG tartalma folyamatosan csökken, ezért oxigén affinitása nő.
Transzfúzió esetén ez előnytelen, ezért a tárolt vérhez inozint adnak, amelynek hatására
2,3-BPG képződik a glikolízis során.
A hemoglobin F kisebb mértékben köti a 2,3-BPG-t, mint a Hb A, így a Hb F
oxigénaffinitása nagyobb, mint a Hb A-é. A Hb F Hb A-nál nagyobb oxigénaffinitása
elősegíti az oxigén bejutását az anyai keringésből a magzati vérkeringésbe.
Bohr-effektus
A hemoglobin és a mioglobin között további lényeges különbség az, hogy a hemoglobin
protonokat és széndioxidot is köt, illetve szállít. A pH csökkenésével a hemoglobin
oxigénaffinitása csökken (pH 7,6 – pO2= 40 Hgmm értéknél a visszatartott oxigén 80%,
míg pH 6,8 – pO2= 40 Hgmm értéknél a visszatartott oxigén 45%). A keletkezett
széndioxid kb. 15%-a a hemoglobinhoz kötődik. Fokozott anyagcsere (pl. izommunka)
esetén több széndioxid termelődik, ilyenkor az oxigénszükséglet is fokozódik. A
laktáttermelés megemelkedik, a vér pH értéke csökken, a hemoglobin oxigéntelítési
görbéje jobbra tolódik (3.14. ábra). A jelenséget Christian Bohr dán fiziológus (Niels
Bohr fizikus édesapja) írta le, tiszteletére Bohr-effektusnak nevezték el. A képződött
széndioxidot, illetve a protonok egy részét a hemoglobin úgy köti meg, hogy a béta-lánc
N-terminális aminocsoportjával karbamátot (OOC-HN-R) képez. További sókötések
jönnek létre, ami tovább csökkenti a hemoglobin oxigén affinitását. A tüdőalveolusokban
a hemoglobin leadja a felvett széndioxidot, illetve protonokat és oxigént vesz fel.
Mindezek eredményeképpen fokozott izommunka során a szövetekben leadható oxigén
mennyisége jelentősen megnő.

3. Enzimszabályozás 31
© Szarka András www.interkonyv.hu
3.14. ábra. A hemoglobin oxigéntelítési görbéje különböző pH-értéken (Bohr-effektus)
3.2. Fehérjék térszerkezetének, aktivitásának befolyásolása foszforiláció/defoszforiláció által
Míg az előző fejezetben ismertetett allosztérikus szabályozás során a regulátormolekula
másodrendű kötésekkel pillanatszerű asszociáció/disszociáció során kapcsolódott a
fehérjéhez és így igen gyorsan bekövetkezett a térszerkezet-/aktivitásváltozás, addig a
fehérjék foszforilációja kovalens módosításról lévén szó: 1. jóval lassabb, 2. tartósabb, 3.
enzim-katalizált (mind a foszforiláció, mind a defoszforiláció) folyamat (3.15. ábra). A
kétféle szabályozási mód (allosztérikus, foszforiláció/defoszforilációval történő)
kiegészíti egymást az eukarióta sejtekben.
A foszforiláció két módon is érintheti, módosíthatja a fehérjéket:
1. Minden PO4 csoport 2 negatív töltést hordoz. Ennek következtében jelentős szer-
kezeti változást generálhat: pl. pozitívan töltött aminosav részleteket vonzhat
magához. A foszfátcsoportot eltávolítva természetesen visszaáll az eredeti
helyzet.

32 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
2. A fehérjéhez kapcsolódott foszfátcsoport egy olyan szerkezet része is lehet,
amelyet más fehérjék kötőhelye ismer fel. Nagyobb fehérjékben gyakran for-
dulnak elő olyan kis fehérjedomének, modulok, amelyek kötőhelyül szolgálhatnak
más foszforilált fehérjék részére. Egy ilyen modul az SH2-domén, amely foszfo-
tirozint tartalmazó peptidszakaszokhoz köt, de rajta kívül még vagy 10 hasonló
fehérjerészlet ismeretes. A foszforiláció ilyeténképp fehérjeszerkezetek össze- és
szétszerelésében is fontos tényező.
A fehérjefoszforiláció mint szabályozási mechanizmus elterjedtségét mi sem mutatja
jobban, mint az a megfigyelés, mely szerint az emlőssejtek 10 000 fehérjéjének mintegy
harmada található foszforilált állapotban az adott pillanatban. A PO43--csoport az ATP-
terminális foszfátcsoportjának terhére történő felvitelét transzferázok, a protein
kinázok katalizálják (3.15 ábra).
3.15. ábra. Fehérje foszforiláció/defoszforiláció
Az ATP hidrolízis miatt a reakció egyirányú. A sejtben több száz különböző protein
kináz található, amelyek a fehérjék szerin-/treonin- (Ser/Thr kinázok) vagy tirozin (Tyr
kinázok) oldalláncainak hidroxilcsoportjára transzferálják a foszfátcsoportot (3.16.
ábra).

3. Enzimszabályozás 33
© Szarka András www.interkonyv.hu
3.16. ábra. Szerin-/treonin- és tirozin-foszforiláció
A foszfátcsoport eltávolítását a foszfoprotein-foszfatázok katalizálják. Hasonlóan a
kinázokhoz a sejt foszfoprotein-foszfatázok tömkelegét tartalmazza, amelyek specifi-
citása jelentős különbséget mutat, némelyikük a fehérjék tágabb csoportját képes foszfo-
rilálni, vagy defoszforilálni, míg mások kizárólagosan csak egy fehérjével képesek ezt
megtenni.
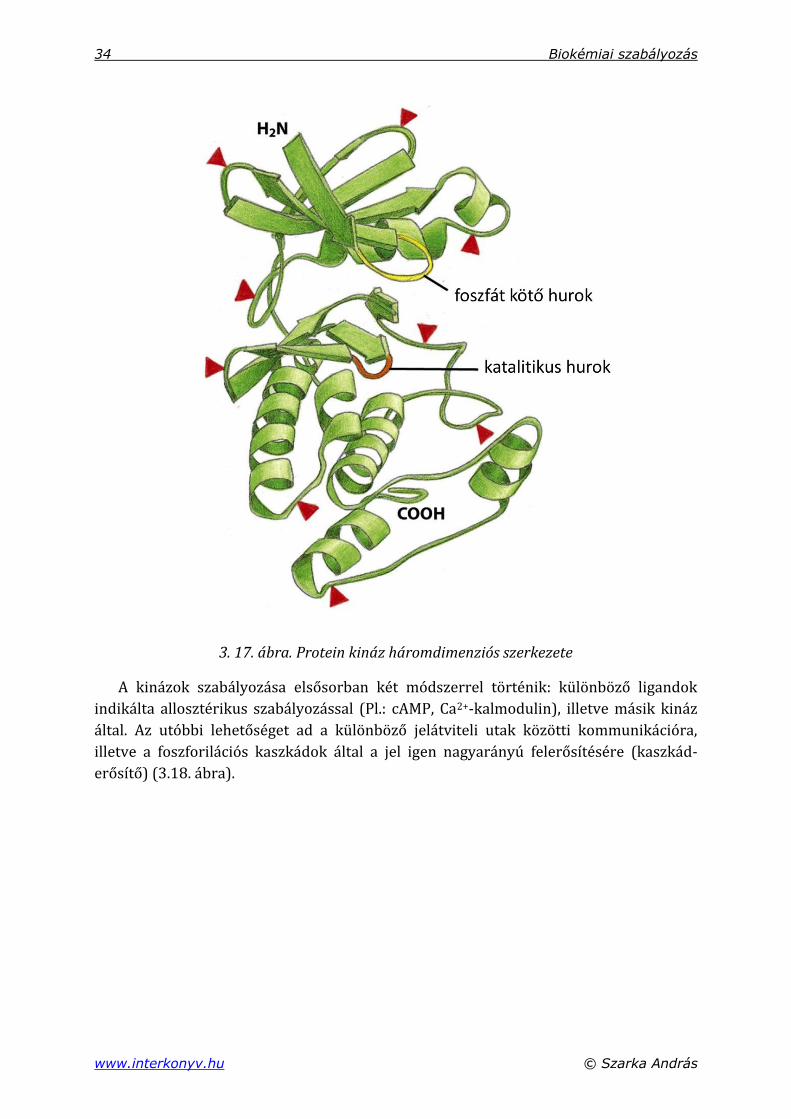
A protein kinázok egy nagy enzimcsalád tagjai, amelynek tagjai egy nagyjából 290
aminosavas-katalitikus (kináz) doménnel rendelkeznek. A fehérjelánc további részei
nagyobb változatosságot mutatnak az egyes családtagok között (3.17. ábra). Ezek
felelősek a szubsztrát felismerésért, illetve a kinázaktivitás szabályozásáért.
34 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
3. 17. ábra. Protein kináz háromdimenziós szerkezete
A kinázok szabályozása elsősorban két módszerrel történik: különböző ligandok
indikálta allosztérikus szabályozással (Pl.: cAMP, Ca2+-kalmodulin), illetve másik kináz
által. Az utóbbi lehetőséget ad a különböző jelátviteli utak közötti kommunikációra,
illetve a foszforilációs kaszkádok által a jel igen nagyarányú felerősítésére (kaszkád-
erősítő) (3.18. ábra).
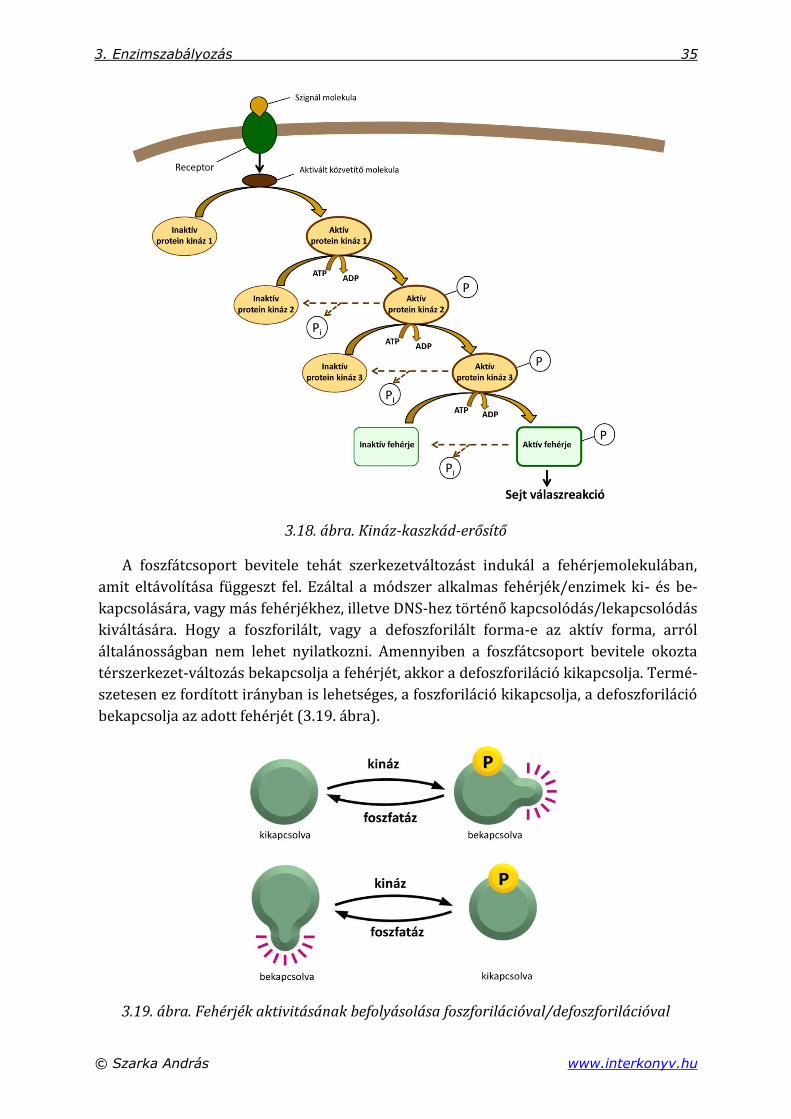
3. Enzimszabályozás 35
© Szarka András www.interkonyv.hu
3.18. ábra. Kináz-kaszkád-erősítő
A foszfátcsoport bevitele tehát szerkezetváltozást indukál a fehérjemolekulában,
amit eltávolítása függeszt fel. Ezáltal a módszer alkalmas fehérjék/enzimek ki- és be-
kapcsolására, vagy más fehérjékhez, illetve DNS-hez történő kapcsolódás/lekapcsolódás
kiváltására. Hogy a foszforilált, vagy a defoszforilált forma-e az aktív forma, arról
általánosságban nem lehet nyilatkozni. Amennyiben a foszfátcsoport bevitele okozta
térszerkezet-változás bekapcsolja a fehérjét, akkor a defoszforiláció kikapcsolja. Termé-
szetesen ez fordított irányban is lehetséges, a foszforiláció kikapcsolja, a defoszforiláció
bekapcsolja az adott fehérjét (3.19. ábra).
3.19. ábra. Fehérjék aktivitásának befolyásolása foszforilációval/defoszforilációval

36 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
3.3. Limitált proteolízis
A harmadik (már kész) fehérjék aktivitásának szabályozására alkalmas módszer neve is
igen beszédes:
Proteolízis: peptidkötések hasítása,
Limitált: csak jól definiát helyen történik.
A módszer által elérhető, hogy kizárólag a betöltött funkció helyén (erre jó példa az
emésztőenzimek aktiválása, amelyek kizárólag a gyomorba/bélbe jutva aktiválódhatnak,
nehogy megemésszék az előállító szövetet), illetve bizonyos körülmények között (erre jó
példát szolgáltatnak a véralvadási enzimek, amelyek kizárólag érfal sérüléskor aktivá-
lódhatnak) legyenek aktívak. A limitált proteolízissel aktiválódó fehérjék, enzimek elő-
alakként, zimogén formában szintetizálódnak, majd később aktiválódnak, miközben
adott helyen hasad a fehérjeláncuk. Fehérjehidrolitikus enzimek másik szabályozási
módja a proteázinhibítorokon keresztüli szabályozás. Röviden erről is szólunk.
3.3.1. Inaktív zimogének és aktiválódásuk
A limitált proteolízissel aktiválódó fehérjék két különböző stratégiát választhatnak:
1. Az aktív centrum kialakul a szintézist követően, de a fehérje egy másik része
lefedi (erre szolgáltat jó példát a pepszinogén).
2. Az aktív centrum csak a limitált proteolízist követő szerkezetváltozás kapcsán
alakul ki (erre szolgáltat jó példát a tripszinogén).
Vegyük szemügyre alaposabban mindkét esetet:
3.3.1.1. Pepszinogén
A pepszinogént a gyomor fősejtjei termelik, gasztrin stimulusra szekretálódik. Meglehe-
tősen széles szubsztrátspecificitással rendelkezik, az aromás aminosavak és dikarbon-
savak melletti peptidkötéseket bontja.
A pepszin zimogén előalakjából a pepszinogénből keletkezik. Aktív centruma már a
pepszinogénben kialakul, de az aktivitásért felelős két aszpartát ionos kötést létesít az
előalak távolabbi részén található argininekkel, lizinekkel. Ezek az ionos kötések savas
pH-n (pH<5) megszűnnek, mivel az aszpartát karboxil csoportjának disszociációja
visszaszorul. Ekkor a fehérjelánc kinyílik és a térszerkezet-változásnak köszönhetően
önemésztődik az előalak és kialakul a teljes aktivitással rendelkező pepszin (3.20. ábra).

3. Enzimszabályozás 37
© Szarka András www.interkonyv.hu
3.20. ábra. A pepszinogén aktiválódása limitált proteolízissel
3.3.1.2. Enterális proteázok zimogénjei
Az enterális proteázok, előalak formájában, a hasnyálmirigyben termelődnek, ahonnan
szekretin és kolecisztokinin stimulusra a vékonybélbe választódnak ki. Ezt követően a
vékonybélben termelődő enteropeptidáz a tripszinogént limitált proteolízissel
tripszinné alakítja, majd ez végzi a többi enzim aktiválását (kimotiszinogén, proelasztáz,
prokarboxipeptidáz-A) (3.21. ábra). Amennyiben az enzimek aktiválása hiányt szenved,
emésztési gondokkal, amennyiben viszont túlaktiválás (illetve túl korai aktiválás)
következik be, nekrotizáló hasnyálmirigy gyulladással (pancreatatis) kell szembenézni.

38 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
3.21. ábra. Az enterolis proteázok zimogénjükből történő aktíválódása limitált proteolízissel
3.3.1.3. Proteázinhibitorok
A két legismertebb proteázinhibítor a pancreas tripszininhibitor és az α1–proteáz
inhibitor.
A Pancreas tripszininhibítor tulajdonképpen egy 6000 Da-os műszubsztrát, amelyet
a tripszin nagyon lassan bont a Lys15 – Ala16 aminosavak között több hónapos
féléletidővel.
Az α1–proteázinhibítor egy akut fázisfehérje, feltehetően a Ser-proteázokkal behatoló
baktériumokkal szembeni védekezés fontos eleme. Elsősorban elasztázinhibitor, de
gyakorlatilag bármely szerinproteázt gátolja. Az inhibítor COO--csoportja és a Ser OH-ja
között lassan hidrolizáló észterkötés alakul ki.
A Met358 nélkülözhetetlen az enzimkötődésben. Dohányosokban ez a metionin
oxidálódik metionin-szulfoxiddá és ezáltal inaktíválódik. Az elégtelen inhibitorfunkció
veleszületett is lehet. Ennek következtében az elasztázok kontrollálatlanul működnek,
melynek legszembetűnőbb következménye a tüdőtágulás (emphysema).

© Szarka András www.interkonyv.hu
4. Fehérjefolding, fehérjelebontás – ubikvitin-proteaszóma rendszer
Ugyan a fehérjék végleges, funkcionális térszerkezetének felvétele, a szintézisüket köve-
tő feltekeredésük, foldingjuk szorosan nem tartozik a biokémiai szabályozástechnika
témakörébe, röviden mégis foglalkozunk vele. Tesszük ezt legalább két ok miatt. Ahogy
korábban láttuk, a fehérjék térszerkezete, annak megváltozása direkt kapcsolatban van
funkciójuk megváltozásával. A hibásan feltekeredett fehérjék, az ismételten sikertelen
feltekeredésüket követően egy központi lebontó apparátus, az ubikvitin-proteaszóma
rendszer ügyfelei lesznek. Szintén ennek a fehérjelebontó rendszernek ügyfelei lesznek
azok a fehérjék, amelyekre már nincs szükség, így a fehérjék élettartam-szabályozásán
keresztül a tématerület végső soron kapcsolódik a sejtben zajló szabályozástechnikai fo-
lyamatokhoz. Végül, de nem utolsó sorban, rendkívül érdekes, ezért sem lenne érdemes
megfosztani magunkat tárgyalásától.
4.1. Poszttranszlációs módosulások
A transzláció során szintetizált fehérjeláncok jelentős része további reakciók tárgya lesz,
hogy elnyerje végső, biológiailag aktív formáját. Ezeket a transzlációt követő, fehérje-
láncot érintő reakciókat összefoglalóan poszttranszlációs módosulásoknak nevezzük.
Poszttranszlációs módosulásokon mind a prokarióta, mind az eukarióta sejtek fehérjéi
keresztülmennek.
4.1.1. Fehérjefolding
A fehérje foldingja során elnyeri aktív háromdimenziós konformációját. Lezajlanak a
poszttranszlációs módosulások, illetve a fehérjelánc hidrofób részeit befelé fordítja,
kvázi beburkolja külső hidrofil felszínével, elzárja a vizes közegtől. Így a vizes közegben
energetikailag kedvezőbb szerkezetet vesz fel.
A foldingnak viszonylag gyorsan kell lezajlania. Ezt támogatja a kialakult aminosav
sorrend is a fehérjékben.
Számos esetben, ahogy az N-terminális megszintetizálódik a riboszómán, elkezdődik a
lánc feltekeredése. A több doménnel rendelkező fehérjék néhány másodperccel a riboszó-
mát elhagyva már egy viszonylag kompakt szerkezetet vesznek fel, amely többé-kevésbé
tartalmazza a másodlagos szerkezetet (α-hélix, β-redő). A legtöbb fehérje esetében ez a
flexibilis és viszonylagosan nyitott szerkezet az „olvadt gombóc” állapot a kiindulópont a

40 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
további foldinghoz, amely során kialakul a másodlagos kötésekkel stabilizált végleges
negyedleges szerkezet.
Egy átalagos fehérje szintézise néhány percen keresztül folyik, számos fehérje
esetében a folding jelentős része lezajlik, mire a C-terminális szabaddá válik a riboszó-
máról (4.1. ábra).
4.1. ábra. Fehérjék transzlációjával párhuzamosan zajló folding
Az „olvadt gombóc” állapotból több lehetséges út vezethet a végleges szerkezetig.
Ezen útvonalak intermedierei aggregálódhatnak, ezáltal lekerülhetnek az útvonalról,
végső soron mint sikertelen folding-próbálkozások megemésztődhetnek.
Ezeket a részlegesen, vagy nem megfelelő módon feltekert fehérjéket a hősokk-
fehérjék tudják megóvni az aggregációtól, illetve segítségükre vannak a helyes térszer-
kezethez vezető út megtalálásában is.
4.1.1.1. Hősokk-fehérjék – molekuláris chaperonok
Radioaktív aminosavakkal követett fehérjeszintézis és foldingkísérletek során kimutat-
ták, hogy a fehérjék 20%-a hsp70, 10%-a pedig hsp60 ügyfél. Mindkét fehérje saját se-
gédfehérjék kis csoportjával dolgozik együtt, hogy más fehérjék feltekeredését segítsék.
A hsp70 már a szintézis során, mielőtt az még elhagyná a riboszómát, hozzá-
kapcsolódik a fehérjék nagyjából hét aminosav hosszú hidrofób aminosav részleteihez
(4.2. ábra).

4. Fehérjefolding, fehérjelebontás – ubikvitin-proteaszóma rendszer 41
© Szarka András www.interkonyv.hu
4.2. ábra. A hsp70-család munka közben
Ezzel szemben a hsp60-szerű fehérjék egy nagy hordóalakú szerkezetet alkotnak,
amelyek a teljes fehérjelánc szintézisét követően lépnek csak működésbe (4.3. ábra).
Gyakorlatilag egy „izolációs kamrába” kerül a fehérje, amely megakadályozza aggregá-
cióját és kedvező feltételeket teremt a folding újbóli megkísérléséhez. Működésük során
a hősokk-fehérjék ATP-t használnak fel, hogy helyes konformációba tekerjék a fehérje-
láncokat.
Ugyan most csak két hősokk fehérjéről szóltunk, azt meg kell említenünk, hogy a
különböző sejtszervecskékben számtalan ilyen fehérje található és vesz részt a fehérjék
megfelelő térszerkezetének kialakításában.
4.3. ábra. A hsp60 család szerkezeti felépítése és működése
Jogosan merül fel bennünk a kérdés, hogyan ismerik fel, választják ki a hősokk
fehérjék a helytelen térszerkezettel rendelkező fehérjéket?
A tökéletlen folding miatt a részlegesen, vagy teljesen feltekeredni képtelen fehérjék
esetében hidrofób felület marad, ehhez a hősokk fehérje köt. Egyfelől megakadályozza
az aggregációját, másfelől segíti a refoldingot.
A fehérjék előtt foldingjukat illetően tehát 4 különböző út állhat:1. kapásból kialakul
a megfelelő térszerkezet; 2. hősokk fehérjék segítségével, refoldingot követően alakul ki
a helyes térszerkezet; 3. nem alakul ki a megfelelő térszerkezet és a sejt lebontja; 4.
aggregálódik (amennyiben ennek mértéke bizonyos határt átlép, az a sejt pusztulását
okozhatja) (4.4. ábra).

42 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
aggregáció
OK Refolding Lebontás
OK
4.4. ábra. Fehérjeminőségi kontroll
Az első két lehetséges útvonalról már röviden szóltunk, ezt követően vizsgáljuk meg
a sérült, nem megfelelő módon feltekert vagy már szükségtelenné vált fehérjék
lebontását.
4.2. Sérült fehérjék proteolízise
A folyamat a fehérjék felületén rendellenesen elhelyezkedő hidrofób rész(ek) felisme-
résével kezdődik, majd a komplex proteolítikus rendszerhez, a proteaszómához történő
szállítással folytatódik, ahol legvégül rövid peptidekre történő bontásukkal fejeződik be.
4.2.1. Proteaszóma
A sejtek meglehetősen gyorsan igyekeznek megszabadulni a transzlációs, illetve folding
hulladékfehérjéktől. Az újonnan szintetizált fehérjék mintegy 1/3-át lebontásra ítéli a
fehérjeminőségi kontroll.
A fehérjék számára a végállomást egy ATP-dependens proteázkomplex, a protea-
szóma jelenti, amely a sejt fehérjéinek mintegy 1 %-át alkotja. A proteaszóma számos
kópiában megtalálható a citoszolban és a nukleuszban is. Ezen kívül endoplazmás reti-
kulumba irányított fehérjéket is bont. Amennyiben az endoplazmás retikulum fehér-
jéinek foldingja, illetve összeszerelése sikertelen, érzékeli egy endoplazmás retikulum
lumenében található minőségellenőrző rendszer. A minőségi kontrollon megbukott
fehérjék retrotranszlokációra kerülnek és a citoszolban található proteaszómákon
kerülnek lebontásra.
4.2.1.1. A proteaszóma felépítése
Minden proteaszóma két fő szerkezeti elemből, egy üreges csőből és a hozzá kapcsolódó
sapkából áll. Az üreges cső a 20S magi proteaszóma. Több fehérje alegységből épül fel,
amelyek 4 heptamerikus gyűrűt alkotnak (4.5. ábra). Alegységeinek egy része különböző
proteáz, amelyek aktív helye a cső belseje felé néz. A cső mindkét vége egy nagy
fehérjekomplex-szel a 19S sapkával asszociál. A 19S sapka hozzávetőlegesen 20 külön-
böző polipeptidből áll. Ebből 6 ATP hidrolízisére képes, a cső végénél ezek az ATP-ázok
kitekerik a fehérjéket és a cső belsejébe juttatják, ahol lezajlik proteolízisük. A protea-
szóma kritikus sajátsága és egyben komplexitásának oka, hogy egy sima proteázzal
ellentétben az első peptidkötés után is megkötve tartja a fehérjét és csak akkor engedi el,
ha már apró darabokra hasogatta. A 19S sapka, mint egy szabályozott kapu működik,
eldönti, hogy beengedje-e a proteolítikus bendőbe a fehérjét. Szintén ez a szerkezeti
elem felel a célfehérjék proteaszómához kötéséért.

4. Fehérjefolding, fehérjelebontás – ubikvitin-proteaszóma rendszer 43
© Szarka András www.interkonyv.hu
4.5. ábra. A proteaszóma felépítése
4.2.2. A „halál csókja” – Ubikvitináció
Kevés kivételtől eltekintve a proteaszóma olyan fehérjéket köt, majd hasít, amelyek egy
speciális jellel rendelkeznek. A „halál csókja” egy 76 aminosavból álló polipeptid, az
ubikvitin (általában több kópiában), kovalensen kötődik. A sejtben mind szabad formá-
ban, mind fehérjéhez kötötten előfordul. A fehérjéhez kapcsolt ubikvitinláncnak többféle
jelentése is lehet, attól függően, hogy hány ubikvitinmolekula és milyen módon kap-
csolódik össze (4.6. ábra).
4.6. ábra. A fehérjéhez kapcsolt ubikvitinláncok különböző szignaling jelentése

44 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
A lebontásra ítélt fehérjéket egy kifinomult ubikvitinkonjugáló rendszer jelöli meg.
Az ubikvitint egy ATP-függő ubikvitin aktiváló enzim (E1) teszi alkalmassá a fehérjékhez
történő kötődésre, konjugálódásra. A konjugálódásra előkészített ubikvitin az E1-ről
szinte azonnal az E2 ubikvitin konjugáló enzimre kerül. Az E2 az E3 kiegészítő fehérjével
együtt látja el feladatát. Az E2-E3 komplexet ubikvitin-ligáznak is nevezik. Az E3 a cél-
fehérjén belül specifikus degradációs szignálokhoz kötődik, segítve az E2-t, hogy
poliubikvitin-láncot kössön a szubsztrátfehérje egy lizin oldalláncához. Az ubikvitin C-
terminálisa az őt megelőző ubikvitin egy lizinéhez kapcsolódik, így létrehozva a
poliubikvitin-láncot. Ezt a poliubikvitin-láncot ismeri majd fel a proteaszóma receptora
(4.7. ábra).
4.7. ábra. A fehérjék ubikvitinációja
Emlős sejtekben nagyjából 30 strukturálisan hasonló, de különböző E2, illetve több
száz különböző E3 található, amelyek mind specifikus E2-enzimekhez kapcsolódnak. A
sejtekben tehát több hasonlóan szervezett, de különböző, specifikus ubikvitin-proteo-
szóma útvonal található. Ezeknek közös a kiindulópontja (E1-enzim) és a végállomása
(proteaszóma).
A különböző ubikvitin-ligázok különböző degradációs szignálokat ismernek fel.
Degradációs szignál lehet:
– denaturált,
– téves térszerkezetű fehérje,
– oxidált aminosav(ak),
– módosult (károsodott) aminosav(ak).

4. Fehérjefolding, fehérjelebontás – ubikvitin-proteaszóma rendszer 45
© Szarka András www.interkonyv.hu
Normálisan ezek a szignálok a fehérje belsejében helyezkednek el, így nem
jelentenek „veszélyt” a megfelelően feltekert fehérjék számára. Felmerül a kérdés, hogy
mi a helyzet a készülő/félkész fehérjékkel, amelyek így még a felszínükön degradációs
szignálokat tartalmaznak? Úgy tűnik, hogy ezek igen gyakran az ubikvitin-proteaszóma
rendszerbe kerülnek és lebontódnak.
4.2.3. Számos fehérje irányított lebontás révén szabályozott
Az ubikvitin-proteaszóma rendszer lehetőséget ad a rövid féléletidejű fehérjék élet-
idejének szabályozására is. Egyes fehérjék mindig rövid életűek, míg mások életideje
függ a sejt állapotától. Bizonyos metabolikus állapotban stabilak, míg egy másikban
rövid életidejűek. Ez utóbbiakra jó példát szolgáltatnak a mitotikus ciklinek, amelyek
stabilak egészen a mitózis végéig.
Felmerül a kérdés, hogyan kontrollált a fehérjék szabályozott lebontása?
Egyfelől lehetőség van az ubikvitin-ligáz szabályozására bármelyik korábban
ismertetett módon: 1. foszforiláció által, 2. allosztétikus ligand által, 3. fehérjemolekula
által (4.8. ábra).
4.8. ábra. Az ubikvitin-ligáz szabályozása
Másfelől a folyamat szabályozható a degradációs szignálon keresztül. Intracelluláris
vagy környezeti hatásra degradációs szignál keletkezhet a molekulán belül 1. foszfori-
lációval, 2. de/maszkoló fehérje, fehérjealegység disszociációja révén, 3. proteolízissel
(4.9. ábra).

46 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
4.9. ábra. Degradációs szignálok kialakulása

© Szarka András www.interkonyv.hu
5. A génexpresszió szabályozása
Az előző fejezetek során megvizsgáltuk, miképpen lehet a már elkészült fehérje
aktivitását, illetve annak élettartamát befolyásolni. Tekintve, hogy a fehérjeszintézis
meglehetősen nagy energiaigénnyel bír, mindenképpen célszerű az adott fehérje iránti
igénynek megfelelően, már korábban szabályozni a gének átírását. Így mind az RNS
szintézise (transzkripció), mind az azt követő érési folyamatok, a fehérje szintézise,
illetve az azt követő érési folyamatok elkerülhetőek, megspórolhatóak (5.1. ábra).
Természetesen ez a szabályozási mód jóval hosszabb átfutási idővel rendelkezik, ebben
az esetben órák, napok alatt lehetséges az egyes folyamatok aktivitásába beavatkozni.
5.1. ábra. A molekuláris biológia centrális dogmája
A molekuláris biológia centrális dogmája (5.1. ábra) értelmében (néhány RNS-vírus
kivételével) minden fehérjeszerkezetre vonatkozó információ, illetve azok kifejező-
désének körülményei a sejt DNS-állománya által meghatározott. Természetesen néhány
fontos pontosítást, körülményt figyelembe kell vennünk:
Az RNS a nukleuszból történő kilépés előtt érik, illetve a splicing merőben megvál-
toztathatja az információ jelentését.
5.1. Génexpresszió, a gén kifejeződése
Egy bakteriális DNS-állomány általában néhány millió bázispárból (bp) áll. A mi emberi
genomunk mintegy 3*109 bp-t számlál. Bár igen jelentős tudományos siker volt a humán
genom szekvenciájának megismerése, a nukleotid sorrend önmagában értelmetlen
adathalmaz. A nukleotid szekvenciák ismerete nem tesz minket képessé arra, hogy
pusztán rájuk alapozva létrehozzunk egy élőlényt, ahogy angol szavak tömkelege sem
tesz képessé minket arra, hogy pusztán rájuk alapozva rekonstruáljuk Shakespeare
bármely művét. Hiszen teljes joggal tehetjük fel a következő kérdések bármelyikét:
Mikor, milyen körülmények között készül az adott gén terméke és ha elkészült mit tesz?
A következő fejezetben szeretnénk tisztázni a génexpresszió szabályozásának legfon-
tosabb szabályait, illetve mechanizmusát. Mindazokat a szabályokat, amelyek alapján a
gének egy alcsoportja szelektíven kifejeződik az egyes sejtekben.

48 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
5.1.1. Sejtdifferenciáció
A különböző sejttípusok (pl. emlős neuron, hámsejt…) mind szerkezetileg, mind funkcio-
nálisan annyira különböznek, hogy nehéz elképzelni, hogy ugyanazt a genomot tartal-
mazzák. Ezért, valamint amiatt, hogy a sejtdifferenciáció gyakran irreverzibilis, a
biológusok kezdetben azt hitték, hogy a differenciálódás során a gének szelektíven
elveszhetnek a sejtből. Ma már tudjuk, hogy a sejtdifferenciáció a génexpresszió
megváltozását jelenti, nem pedig a nukleotidszekvencia megváltozását. A háttérben
tehát nem a különböző DNS, hanem a különböző RNS, illetve fehérjeállomány áll.
Ez utóbbi kijelentést kísérleti bizonyítékkal is alátámasztották.1958-ban, John
Gurdon és munkatársai, az Oxfordi Egyetem kutatója, afrikai karmosbékán (Xenopus
laevis) végzett kísérleteik során egy teljesen differenciálódott békasejtmagot egy
sejtmagtalanított békapetesejtbe ültettek. A beültetett sejtmaggal a petéből egy teljesen
életképes, normális ebihal keletkezett. Tekintve, hogy az ebihal a differenciálódott sejtek
legkülönbözőbb együttesét tartalmazza, bizonyítottá vált, hogy a differenciálódott (és
később beültetett) sejt nem vesztette el egyetlen fontos géndarabját sem (5.2. ábra).
5.2. ábra. A sejtdifferenciáció a génexpresszió megváltozása, nem a nukleotidszekvenciáé.
Hasonló eredményeket értek el sárgarépa esetében, mikor sejttenyészetbe vitték az
izolált sejtet, majd abból egy sejtet kiválasztottak. A későbbiek során belőle teljes,
életképes növény növekedett. Emlősök esetében is számos esetben sikerült a fenti tételt
bizonyítani: birka (Dolly), kecske, sertés, egér, illetve madarak (liba) esetében is.
A fenti kijelentés alól néhány kivétel azért akad, így például DNS-átrendeződés
történik az egyed fejlődése során, pl. az immunrendszer változatosságának kialakulása
során.
A sejtdifferenciáció (génexpresszió) mibenléte talán jobban megérthető, ha szem-
ügyre vesszük a sejtek között fennálló különbségeket, azonosságokat.
5.1.1.1. Azonosságok és különbségek a sejtek között
1. Számos olyan folyamat van, ami közös az egyes sejtekben. Természetesen ezek közös
fehérjeállományt igényelnek, jelentenek. Ilyenek például: strukturális fehérjék kro-
moszómacsomagoló fehérjék, RNS-, DNS-polimerázok, DNS-reparáló enzimek, ribo-
szómális fehérjék, citoszkeletális fehérjék, központi metabolizmus fehérjéi.

5. A génexpresszió szabályozása 49
© Szarka András www.interkonyv.hu
2. Ugyanakkor rendkívül nagy különbségeket is tapasztalhatunk, hiszen sok fehérje
csak az adott sejttípusban található meg, erre jó példaa csak vörösvértestekben kife-
jeződő hemoglobin.
3. Adott időben egy átlag humán sejt 10 000-20 000 gént fejez ki a megközelítőleg
25 000-ből. Ha megnézzük a különböző gének expressziós mintázatát, rendkívül
nagy variációt láthatunk a különböző sejttípusok között. A különbségek lehetnek
annyira élesek, mint a hemoglobin esetében, de a legtöbbször csak finom eltérések
fedezhetők fel.
4. Habár nagy az mRNS-beli különbség a különböző sejttípusok között, ezt jóval
meghaladja az eltérés, amely a fehérjék esetében tapasztalható. Elsősorban az
alternatív splicing, a poszttranszlációs módosulások tovább növelik a különbségeket.
Itt érdemes megjegyeznünk, hogy a legtöbb sejt képes külsődleges jelek hatására
megváltoztatni a génexpressziós mintázatát.
Például a májsejtek glukokortikoid hormonok hatására számos fehérje kifejeződését
fokozzák. A glukokortikoid hormonok éhezés, intenzív fizikai munka hatására kerülnek
elválasztásra és a májsejtek számára jelként szolgálnak, hogy fokozza a glukóz
aminosavakból, illetve más kis molsúlyú anyagokból történő előállítását. Ennek
eredményeképpen például megnövekszik a tirozin-aminotranszferáz szintje, amely a
tirozinból történő glukózelőállítás fontos kulcsenzime. Amennyiben a hormon szintje
lecsökken, vele együtt ezen fehérjék kifejeződése, szintje is csökken.
Más típusú sejtek máshogy reagálnak a glukokortikoid stimulusra. Példának okáért a
zsírsejtekben glukokortikoid hatására lecsökken a tirozin-aminotranszferáz szintje, míg
más sejtek sehogy sem reagálnak.
A sejtdifferenciáció egyik általános sajátsága, hogy a különböző sejttípusok máshogy
reagálnak az extracelluláris szignálokra. Ezzel, megadva az alapot az extracelluláris
szignálok kiváltotta szabályozásra. A génexpressziós mintázatnak azonban számos olyan
jellemzője van, amely nem változik meg és ezáltal az egyes sejttípusok számára állandó,
megkülönböztető karaktert biztosít.
5.1.2. A génexpresszió szabályozásának szintjei
A génexpresszió a DNS-től induló, RNS-en keresztül menő, fehérjékig tartó útvonala
során számos ponton szabályozható. Röviden tekintsük át milyen szabályozási szintek-
ről beszélhetünk (5.3. ábra).
1. Transzkripciós kontroll: Mikor és milyen gyakran íródik át mRNS-re az adott
gén?
2. RNS-processziós kontroll: Az adott mRNS hogyan érik?
3. RNS-transzport kontroll: Melyik érett, komplett RNS exportálódik a sejtmagból
és a citoszólban hol lokalizálódik?
4. Transzlációs kontroll: A citoszólban melyik mRNS fog átíródni fehérjére?

50 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
5. RNS-degradációs kontroll: Szelektíven destabilizálódó mRNS, amelyről nem
készül fehérjelánc.
6. Fehérjeaktivációs kontroll: Szelektíven aktiválódó, deaktiválódó, degradálódó,
kompartmentalizálódó fehérjék.
5.3. ábra. A génkifejeződés szabályozásának szintjei
A legtöbb gén esetében a transzkripciós kontroll kiemelkedően fontos. Ez könnyen
belátható, ha belegondolunk, ez az egyetlen lehetőség, hogy a sejt ne gyártson felesleges
köztitermékeket (5.3. ábra). Mindenképpen a szabályozás egy gazdaságos módjának
nevezhetjük, ellenben korántsem a leggyorsabbnak, hiszen a további öt ponton is át kell
jutnia a génnek, majd a fehérjének. A 3. fejezetben ismertetett fehérjeaktivációs kontroll
ettől jóval gyorsabb beavatkozásra is lehetőséget ad.
A következő fejezetben megtárgyaljuk azon szabályozó DNS- és fehérjeelemeket,
amelyek segítségével a transzkripció befolyásolható.
5.1.3. A génszabályozó fehérjék DNS-kötő részei
Hogy dönti el a sejt, hogy több ezer génje közül melyiket írja át?
Az átírni kívánt génszakasz közelében található egy DNS-regulációs szakasz. Ezek
között vannak egészen egyszerűek, mint egy külső szignál által bekapcsolt kapcsoló és
egész bonyolultak, mint egy kis mikroprocesszor, amelyek többféle szignált vesznek,
kombinálnak és be- vagy kikapcsolják a szomszédos gént.
Mindkét fajta kapcsoló alkalmatosság két fő szerkezeti egységből épül fel. Egy rövid
irányító DNS-szakaszból és egy (vagy több) génregulációs fehérjéből, melyek az
előbbiekhez kötődnek.
A λ-represszor volt az elsőként leírásra került ilyen DNS-kötő, transzkripciós
szabályozófehérje. Letompítja a virális géneket, a vírus alszik, de szaporodik.

5. A génexpresszió szabályozása 51
© Szarka András www.interkonyv.hu
A fehérjével interakcióra képes DNS-részletek azonosítása 4 lépésben történhet meg:
1. fehérjeizolálás;
2. specifikus kötődés létrehozása (bizonyítása);
3. a fehérjével kötődő DNS szekvenálása;
4. DNS-lenyomatolás.
Természetesen adódik a kérdés, hogy mi módon ismerik fel a fehérjék a jellegzetes
DNS-szekvenciákat? Mi hordozza számukra a kötődéshez szükséges információt?
Tekintve, hogy a DNS-ben tárolt információt a bázissorrend jelenti, az első elképzelés
szerint a fehérjék a kettősszálú DNS-ben a bázisok közötti H-hidakhoz férkőznének és ily
módon ismernék fel a jellegzetes részleteket. Ekkor azonban fel kellene, hogy nyíljék a
kettősszálú DNS-lánc.
Jelenleg már ismeretes, hogy a DNS-t felismerő, azokhoz specifikusan kötő
fehérjék a DNS külső felszínét ismerik fel. Így nem szükséges, hogy felnyíljék a DNS-
kettősszál. Gondoljunk csak bele: a DNS-ben található mindegyik bázispár éle a DNS-
kettőshélix felületén van. Így a felületen a bázisoknak megfelelően különböző funkciós
csoportok vannak: H-kötés donorok, H-kötés akceptorok, illetve hidrofób csoportok (5.4.
ábra). Ezek a csoportok pedig a bázissorrendnek megfelelően szabályosan váltakoznak.
Így tulajdonképpen a DNS felülete is kellő információval szolgál (ha nem is annyira
specifikusan, mint a bázisok maguk) a DNS-kötő fehérjék számára a kötődés pontos
helyéről (5.4. ábra).
5.4. ábra. A DNS-kötő fehérjék kapcsolódási helye a DNS-molekula külső felszínén
A DNS-ben a párokat alkotó bázisok és a hozzájuk tartozó cukoregységek között
létrejövő glikozidos kötések nem pontosan egymással szemben helyezkednek el. Az
egyik oldalon 180o-nál nagyobb, a másik oldalon ennél kisebb szöget zárnak be

52 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
egymással. A kettőshélix külső részét képező dezoxiribóz-foszfát váz csavarulatai között
elhelyezkedő, szintén helikális árok ennek megfelelően torzul. Azon az oldalon, ahol a
bezárt szög nagyobb, található a nagyobb helikális árok (nagyárok), ahol pedig kisebb,
ott a kisárok (5.5. ábra).
5.5. ábra. A nagyárok és a kisárok értelmezése a glikozidos kötések egymással bezárt szögei alapján
A fehérjék számára csak a nagyárokban áll rendelkezésre elegendő információ, így a
DNS-kötő fehérjék minden esetben a DNS nagyárkába fekszenek be (5.4. ábra).
Ezeken kívül a hélixben foglalt DNS-szekvenciától függően torzul a hélix és ezek a
torzulások is információval szolgálnak a fehérjéknek a megfelelő szakasz felismeréséhez.
A DNS-ben szabályosan 36o a szögelfordulás. Ez azt jelenti, hogy 10 nukleotidpárra jut
egy 1 hélixfordulat. Az egymást követő sok AAAA azonban ezt torzítja és egy hajlítást
visz a DNS-be.
A DNS szerkezete flexibilis, sokszor a kötődő fehérjéhez illeszkedik, így még erő-
sebbé téve a kötődést. Az energianyereség (ami a deformálódásból, kötődésből szár-
mazik) nukleotidszekvencia-függő.
A „genetikai kapcsolók” fontos részei a 20 bp-nál rövidebb DNS-szekvenciák, ame-
lyeket a kötőhelyek felismernek. Több ezer ilyen DNS-szekvenciát azonosítottak, ame-
lyeket különböző génregulációs fehérjék, vagy génregulációs fehérjeszettek ismernek fel.
5.1.4. DNS-felismerő fehérjeszerkezeti egységek
A biológiában, a molekuláris felismerés két molekula felületeinek tökéletes illesz-
kedésén múlik (ilyen például az antigén-antitest illeszkedés, kapcsolat is). Egy további
igen kiváló példa a génregulációs fehérjék és a DNS között kialakuló kapcsolat. A
génregulációs fehérje felismeri a specifikus DNS-szakaszt, mivel a fehérje felülete

5. A génexpresszió szabályozása 53
© Szarka András www.interkonyv.hu
nagymértékben komplementer az adott régióban a kettős hélix speciális felületi
sajátságaival. A fehérje nagyszámú kapcsolódást alakít ki a DNS-sel hidrofób, ionos, H-
híd kötések révén. Ezek egyenként gyengék, de a kettő vagy több kapcsolódás erős
és specifikus kötődést eredményez (5.4. ábra).
A kapcsolódások mindegyike egyedi, mindegyik fehérje tartalmaz egy speciális DNS
kötő szakaszt – ezek rendszerint α-hélixet vagy β-redőt jelentenek.
A következőkben tekintsük át milyen DNS-t felismerő fehérjerészleteket, fehérje-
családokat ismerünk.
5.1.4.1. Hélix-turn-hélix részletek
Az elsőként leírt DNS-kötő fehérjecsalád. Eredetileg baktériumokban írták le, de
eukarióta sejtekben is előfordul. A családot több száz fehérje alkotja. Szerkezetüket
tekintve két α-hélixből és az őket összekötő rövid hajlatból állnak. A rövid hajlat a két
hélixet fix szögben tartja. A C-terminális a DNS-felismerő hélix, a nagyárokba ez köt be
(5.6. ábra). Ennek aminosav egységei minden fehérjében különböznek. A másik hélix
szerkezeti feladatokat lát el, irányban tartja, pozícionálja a felismerő hélixet.
5.6. ábra. A hélix-turn-hélix fehérjerészlet felépítése és kapcsolódása a DNS-hez
A hélix-turn-hélix szerkezeten kívül eső részek nagy szerkezeti különbséget
mutatnak a fehérjék között. Ezek révén mindegyik fehérje rá jellemző módon képes a
hélix-turn-hélix részletét a DNS-hez tartani. Az egyéb részek is képesek a DNS-sel
kapcsolatba lépni, mintegy finomra hangolva a DNS és a fehérje közötti kapcsolatot.
Igen gyakran dimer formában kötnek a DNS-hez. A dimerként való kötődés közös
tulajdonsága számos szekvencia specifikus DNS-kötő fehérjének. Ennek megfelelően a
DNS két oldala is hasonló, szimmetrikus szekvenciájú (5.7. ábra).
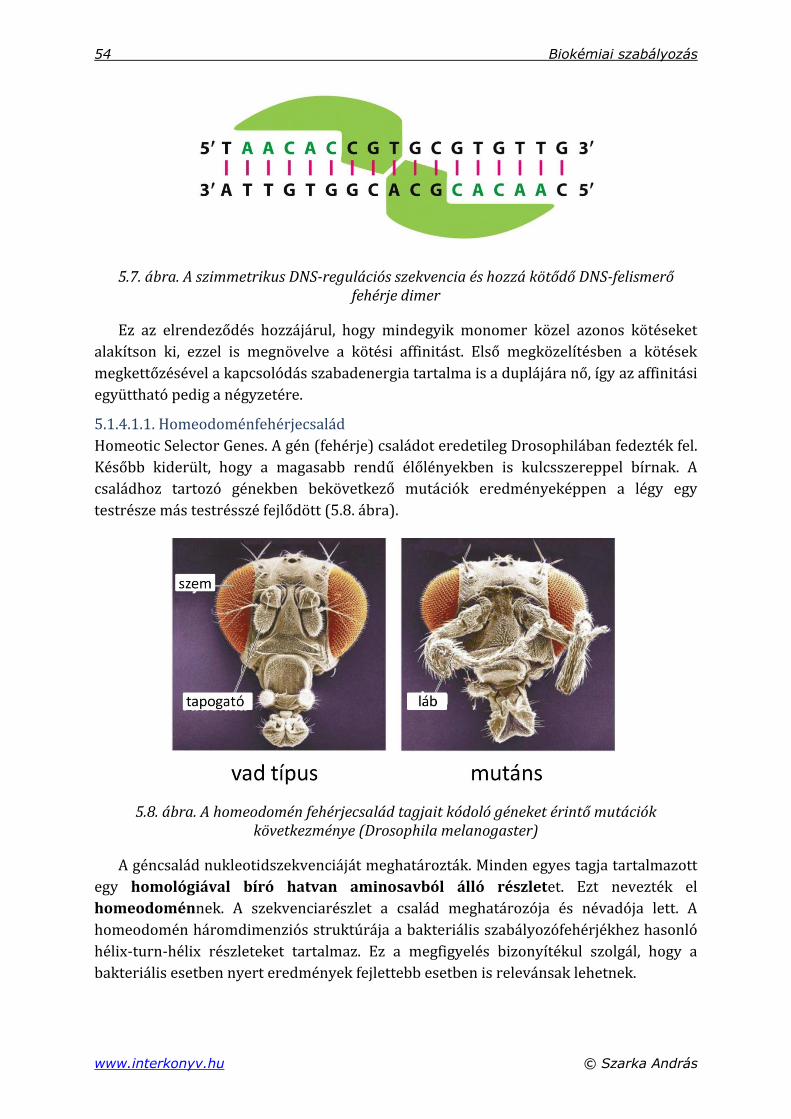
54 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
5.7. ábra. A szimmetrikus DNS-regulációs szekvencia és hozzá kötődő DNS-felismerő fehérje dimer
Ez az elrendeződés hozzájárul, hogy mindegyik monomer közel azonos kötéseket
alakítson ki, ezzel is megnövelve a kötési affinitást. Első megközelítésben a kötések
megkettőzésével a kapcsolódás szabadenergia tartalma is a duplájára nő, így az affinitási
együttható pedig a négyzetére.
5.1.4.1.1. Homeodoménfehérjecsalád
Homeotic Selector Genes. A gén (fehérje) családot eredetileg Drosophilában fedezték fel.
Később kiderült, hogy a magasabb rendű élőlényekben is kulcsszereppel bírnak. A
családhoz tartozó génekben bekövetkező mutációk eredményeképpen a légy egy
testrésze más testrésszé fejlődött (5.8. ábra).
5.8. ábra. A homeodomén fehérjecsalád tagjait kódoló géneket érintő mutációk következménye (Drosophila melanogaster)
A géncsalád nukleotidszekvenciáját meghatározták. Minden egyes tagja tartalmazott
egy homológiával bíró hatvan aminosavból álló részletet. Ezt nevezték el
homeodoménnek. A szekvenciarészlet a család meghatározója és névadója lett. A
homeodomén háromdimenziós struktúrája a bakteriális szabályozófehérjékhez hasonló
hélix-turn-hélix részleteket tartalmaz. Ez a megfigyelés bizonyítékul szolgál, hogy a
bakteriális esetben nyert eredmények fejlettebb esetben is relevánsak lehetnek.

5. A génexpresszió szabályozása 55
© Szarka András www.interkonyv.hu
A Drosophila genom (proteom) több mint 60 homeodomén fehérjét tartalmaz. A
gyümölcslegyeken kívül számos organizmusban írták le a család tagjainak jelenlétét, így
például élesztőben, különböző növényekben és az emberben is.
A bakteriális fehérjében a hélix-turn-hélix részlet különböző szerkezeti környe-
zetben található. Ezzel szemben a homeodomén család tagjaiban hasonlóak a különböző
hélix-turn-hélix részeket körülvevő szerkezeti elemek (ezek alkotják a homeodomén
fennmaradó részét). Így arra következtethetünk, hogy a hélix-turn-hélix részlet mindig
hasonlóan helyezkedik a DNS-hez. Egy drosophila és egy élesztő homeodomén család-
tagot összehasonlítva megállapították, hogy, bár csak mindössze 17 aminosav egyezett a
60-ból, a konformációjuk és a DNS-felismerés módja nagyon hasonlított egymásra.
5.1.4.2. DNS-kötő cink-ujj részletek
Míg a hélix-turn-hélix fehérjék kizárólag aminosav részekből épülnek fel, addig a cink-ujj
fehérjék egy vagy több strukturális szerepet betöltő cinket is tartalmaznak. Bár az
elnevezés azonos minden Zn-et tartalmazó DNS-kötő fehérje esetében, a név eredete az
elsőként felfedezett ujj alakú fehérjétől származik.
A később napvilágra került szerkezeti tanulmányok kimutatták, hogy több szerkezeti
csoportba lehet a család tagjait osztani. Jelen esetben ezek közül a két legfontosabbat
említjük meg:
1. Az elsőt az eukarióta rRNS expresszióját aktiváló fehérjében írták le. Szerkezete
rendkívül egyszerű, egy α-hélix és egy β-redő, amelyet egy Zn tart össze (5.9.
ábra). Ezek az egységek igen gyakran klasztereket alkotnak, így folyamatosan
kitöltve a nagyárkot (egyik DNS kötő -hélix a másik után) (5.9. ábra). Így egy
erős és specifikus fehérje-DNS kapcsolat alakulhat ki a bázisegységek folyamatos
ismétlődése révén. A szerveződés előnye, hogy az ismétlődések számának
változtatásával változtatható a kötődés erőssége, specifikussága is.
2. A Zn-ujjak másik családja az intracelluláris receptorokban található (lásd 6.
fejezet). Szerkezetük különbözik az előző csoportba tartozó fehérjékétől: két α-
hélix pakolódik össze Zn segítségével (hasonló a hélix-turn-hélix szerkezethez). A
hélix-turn-hélix szerkezethez hasonlóan dimereket képez, amely lehetővé teszi,
hogy az egyik α-hélix a nagyárokba feküdjön.

56 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
5.9. ábra. A cink-ujj fehérjerészletek első csoportjának szerkezeti felépítése, illetve a klaszterképzés
Habár különböző szerkezetűek az imént tárgyalt fehérjék, közös vonásuk, hogy Zn-t
használnak szerkezeti elemként, illetve egy α-hélix révén ismerik fel a nagyárokban
található információt (szekvenciát).
5.1.4.3. β-redő
Az eddig ismertetésre került DNS-kötő fehérjékben minden esetben az α-hélix szolgált
felismerő részletként. A génregulációs fehérjék egy csoportjában β-redő részletek fek-
szenek a DNS nagyárkába. A kétszálú β-redő szerkezetről az aminosav oldalláncok a DNS
irányába lógnak. A felismerés az adott aminosavtól függ, amelyek a β-redő adott részét
alkotják.
5.1.4.4. Leucin-cipzár
Sok génregulációs fehérje homodimer (feltehetően az erős specifikus kötés miatt). Gyakran a dimerizálódásért felelős rész más, mint a DNS-kötésért felelős rész. A leucin-cipzár ötvözi a két funkciót. A cipzár elnevezés a szerkezetéből fakad, mivel a két α-hélix, hidrofób kötései révén egy összecsavarodást hajt végre gyakran a leucinrészleteknél (5.10. ábra).

5. A génexpresszió szabályozása 57
© Szarka András www.interkonyv.hu
5.10. ábra. A leucin-cipzár fehérjerészlet szerkezete és kötődése a DNS-hez
Kevéssel a dimerizációs szakaszon túl egy y-t formálva szétválnak, hogy lehetővé
tegyék az oldalláncaiknak a nagyárokba kötődést. A dimerek úgy ülnek a DNS-en, mint a
ruhacsipesz a ruhaszárító kötélen (5.10. ábra). A lehetőségek sorát bővíti, hogy több
DNS-kötő fehérje, így a leucin-cipzár is heterodimereket is képes formálni. Így igen
változatos erősségű és specificitású DNS-felismerő fehérje (komplexek) jöhetnek létre.
Természetesen a heterodimerek variálódásának, promiszkuitásának, határa van, így el-
kerülhető a regulációs káosz. Hogy képződik-e két leucin-cipzár rész között hetero-
dimer, az dönti el, hogy a két α-hélix hidrofób felülete mennyire passzol egymáshoz. Ez
végső soron az aminosav sorrendtől függ. Így egy adott leucin-cipzár fehérje csak
viszonylag szűk leucin-cipzár partnerállományból válogathat az adott sejten belül.
A heterodimerizáció jó példa a kombinatorikus kontrollra, amikor nem egy külön-
álló, önálló fehérje, hanem különböző fehérjék kombinációja felügyeli a sejt folyamatait.
5.1.4.5. Hélix-loop-hélix részlet
Egy rövidebb és egy hosszabb α-hélix szakaszból és az őket összekötő hurokból áll. A
hurok flexibilitása lehetővé teszi, hogy az egyik hélix visszakanyarodjon és a másikkal
átellenben csomagolódjon. Az 5. 11. ábrán jól látható, hogy ez a duplahélix-szerkezet a
DNS-hez kapcsolódik, egy másik hélix-loop-hélix szerkezethez hasonlóan, mint a leucin-
cipzár.

58 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
5.11. ábra. A hélix-loop-hélix fehérjerészlet szerkezete és kapcsolódása a DNS-hez
A leucin-cipzárhoz hasonlóan képezhet homo- és heterodimereket. Sok hélix-loop-
hélix fehérje elveszti a DNS-hez kötő α-hélixét, ezek a csonka fehérjék teljes hosszúságú
hélix-loop-hélix fehérjékkel heterodimereket alkotnak, azonban a kötődés a DNS-hez
ebben az esetben lazább, mint a teljes hosszúságú fehérjék esetében. Ez tulajdonképpen
egy szabályozási lehetőség, mivel a csonka fehérje azáltal, hogy egy teljes fehérjéhez
kapcsolódik, megakadályozza azt, hogy egy másik teljes hosszúságú fehérjével
homodimerizálódjon és aktív legyen (5.12. ábra).
5.12. ábra. Hélix-loop-hélix fehérjerészlet aktivitásának szabályozása heterodimerizációval
Nyílván több olvasóban is felmerült a kérdés, hogy vajon melyik fehérjerészlet
melyik DNS-szekvenciát ismeri fel? A tárgyalt DNS-felismerő részletek szerkezeti
keretet biztosítanak, hogy az adott aminosavak adott DNS-szakaszokat ismerjenek fel.
De vajon van-e pontos fehérje-bázis megfeleltetés? Példának okáért a G-C párost
milyen aminosav ismeri fel? A válasz, hogy nincs, bár adott bázis-aminosavpárosítások
gyakoribbak, mint mások. 20 aminosavból kombinálódnak, több lehetőség is van, hogy

5. A génexpresszió szabályozása 59
© Szarka András www.interkonyv.hu
az adott szekvenciának megfelelő felületet kialakítsák. Talán a közeljövőben lehet majd
adott DNS-szekvenciát felismerő fehérjét tervezni. A probléma megoldása hihetetlen
távlatokat nyitna a molekuláris biotechnológia és a molekuláris biológiai terápia terén.
5.1.5. DNS-kötő fehérjék detektálása
A biokémiai szabályozások részletesebb megismeréséhez elengedhetetlen a DNS-kötő
fehérjék izolálása, szerkezetük megismerése. A továbbiakban néhány gyakran
használatos gyakorlati módszert ismertetünk a DNS-kötő fehérjék tisztítására.
1. Gél-mobility shift assay
Az első lépés a kérdéses DNS-szakasz klónozásos vagy kémiai úton történő
előállítása, majd radioaktív jelölése. Az így elkészített DNS-preparátumhoz adjuk
a sejtlizátumot, majd megfuttatjuk poliakrilamid gélen. Amennyiben a DNS-szál
fehérjét kötött, lassabban vándorol (5.13 ábra).
5.13. ábra. A gél mobility shift assay (electrophoretic mobility shift assay, EMSA) működési elve
2. DNS-affinitás kromatográfia
Ha a génregulációs fehérje által felismert DNS-szakasz ismert, egy porózus
mátrixhoz kötik a kémiai úton előállított kettősszálú DNS-t. Ez megköti,
gyakorlatilag „kiválogatja” a kötésre képes fehérjét. Ha a teljes genom

60 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
szekvenciája ismert, a fehérjerészlet aminosav szekvenciájából kiindulva
megkereshető a regulációs fehérje génje. Az RNS meghatározható, így klónozható.
3. A felismert DNS-szekvencia meghatározása
A génregulációs fehérjék egy része izolálható a megváltozott mutáns egyedekből,
például a homeodomén fehérjék génjeit is így sikerült fejlődési rendellenességgel
rendelkező Drosophilákból izolálni. Ezeket túltermeltetik, majd izolálják a
fehérjét. Rövid DNS-szakaszokkal párosítják (csak amelyiket szorosan köti); sok
ilyen kör után meghatározzák a DNS-szakasz szekvenciáját, a konszenzus-
szekvenciát. Adatbázisokkal vetik össze és megnézik, hogy melyik génben lehet
az adott szekvencia. A módszer hátránya, hogy nem teljesen megbízható; sok
szervezet termel hasonló génregulációs fehérjéket, amelyek közel azonos DNS-
szekvenciákat ismernek fel.
4. Kromatin-immunoprecipitációs technika
A DNS-hez kötött fehérjéket formaldehid segítségével kovalensen keresztkötik (a
DNS-sel), majd lizálják a sejteket. Az izolált DNS-t kis fragmentumokra szeletelik.
Ezt követően az adott génregulációs fehérje ellen termeltetett antitesttel
kihalásszák a DNS-fehérje komplexeket. Így azokat a DNS-részleteket
határozhatjuk meg, amelyekhez a sejtben a regulációs fehérje kötődik.
5.2. A genetikai kapcsolók munka közben
A genetikai kapcsolók szerkezetének megismerését követően itt az ideje, hogy végre
megismerjük működésüket. A megismerés legjobb eszköze, ha végignézünk egy egy-
szerűbb és egy valamivel komplikáltabb prokarióta génexpressziós szabályozást, majd a
bakteriális analógia segítségével áttérünk a jóval összetettebb eukarióta szabályozásra.
5.2.1. A triptofán-represszor, mint egyszerű génkapcsoló
Ha pusztán mennyiségi szempontból vizsgáljuk meg az Escherichia Coli genomját, illetve
proteomját, megállapíthatjuk, hogy azt 4,6 * 106 bp és 4 300 fehérje alkotja. Egy adott
időpontban azonban mindössze csak egy részük íródik át, készül el. A baktérium éle-
tében kiemelkedő szereppel bír a tápanyaghoz való hozzáférés (rendkívül sok hason-
lóságot mutatva jó néhány embertársunkkal). Számos gén kifejeződése a rendelkezésre
álló táplálékoktól függ. Erre igen jó példa a triptofán szintéziséhez szükséges 5 gén. A
bioszintetikus útvonal enzimeit kódoló gének egy operonba rendeződtek. Az operon
struktúrgének (fehérjéket kódoló gének) egy kisebb csoportjából áll, amelyek igen
gyakran egy bioszintetikus, metabolikus útvonal egymást követő lépéseinek kata-
líziséért felelős fehérjéket kódolnak (egyik gén a másik után), illetve az operon részét
képezi a gének szabályozásáért felelős DNS-szakasz (regulátor szakasz és promóter).
Mindezen gének egy promóterről íródnak át, egyetlen hosszú mRNS-t eredményezve
(5.14. ábra).

5. A génexpresszió szabályozása 61
© Szarka András www.interkonyv.hu
5.14. ábra. A triptofán operon elemei
Amennyiben a triptofán a baktérium számára készen rendelkezésére áll (mondjuk,
ha egy emlős belében található, amelyik megeszi a fehérjét tartalmazó táplálékot), a sejt
kikapcsolja a triptofán szintéziséhez szükséges enzimek génjeit.
A transzkripció iniciációja kiemelt fontosságú, hiszen a sejt ekkor dönti el, hogy
melyik fehérjét és milyen arányban (túlsúllyal) fejezze ki. A bakteriális RNS-polimeráz
egy több alegységes komplex. A komplex egyik tagja, a különálló, de a komplexhez
asszociáló alegység, a σ-faktor (5.15. ábra). A σ-faktornak kiemelt szerepe van abban,
hogy az RNS-polimeráz felismerje a DNS-en a transzkripció megkezdéséért felelős
szignált (azaz a promóter szekvenciát).
Az RNS-polimeráz csak lazán kötődik a DNS-hez, gyorsan végigszalad (csúszik) a
DNS-molekulán, majd ledisszociál róla. Ha azonban a polimeráz a promóter régióra
csúszik, amely tehát egy speciális DNS-szekvencia, amely kijelöli az RNS-polimeráz
számára, hogy itt kell az RNS-szintézist elkezdeni, akkor szorosan hozzáköt. A polimeráz
a σ-faktor segítségével ismeri fel a promótert. A faktor, az előző fejezetben részletezett
módon specifikus kötéseket alakít ki a DNS külső felszínén a bázisokkal. Ezt követően az
RNS-polimeráz felnyitja a DNS-kettőshélixet (5.15. ábra). Egy rövidke szakasz
kinyitásával megkezdődik az RNS szintézise. A kitekeredést a polimeráz és a DNS-lánc
komformációváltozása stabilizálja, a folyamat (a replikációval ellentétben) ATP-t nem
igényel.
Az első tíz nukleotid összekapcsolódása után az RNS-polimerázról leválik a σ-faktor
(5.15. ábra). A polomeráz konformációja megváltozik, ennek következtében a szintézis
sebessége megváltozik, beindul a gyors RNS-szintézis. Ez egészen a második szignálig, a
terminátor szekvenciáig tart, itt az RNS-polimeráz elengedi mind a DNS-t, mind a frissen
szintetizált RNS-t (5.15. ábra).

62 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
5.15. ábra. A transzkripció lépései (1.promóter felismerése a -faktor által, 2.a DNS-szál felnyitása, 3. RNS-szál szintézisének kezdete(iniciáció), 4. az első tíz nukleotid szintézise,5.
a -faktor leválása a komplexről, RNS-szintézis felgyorsul (elongáció), 6. a szintézis a terminátor régióig folyik, majd a kész RNS-szál leválik a komplexről, 7. a -faktor ismét
elfoglalja helyét az RNS-polimeráz komplexben)
A triptofán szintéziséért felelős gének promóterében található egy operátornak
nevezett szabályozó. Ez tulajdonképpen egy rövidke (jól definiált nukleotidsorrenddel
rendelkező), szabályozó DNS-szakasz, melyet egy génszabályozó fehérje, a triptofán-
represszor ismer fel. A triptofán-represszor, egy hélix-turn-hélix családba tartozó
fehérje (lásd előző fejezet). A promóter és az operátor szakaszok úgy helyezkednek el,
hogy ha a triptofán-represszor beköt az operátor szakaszhoz, az megakadályozza (az
átlapoló elhelyezkedés révén) az RNS-polimeráz bekötődését a promóterhez, így a
triptofánt szintetizáló enzimek nem tudnak átíródni (5.16. ábra).
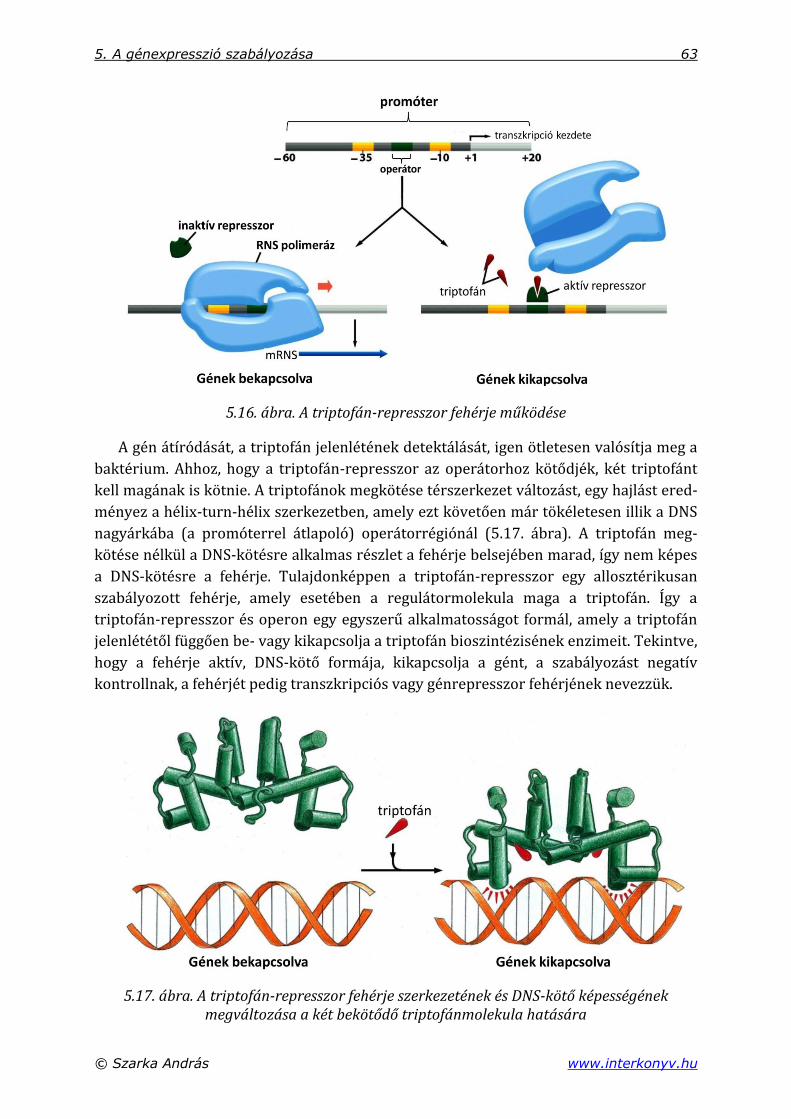
5. A génexpresszió szabályozása 63
© Szarka András www.interkonyv.hu
5.16. ábra. A triptofán-represszor fehérje működése
A gén átíródását, a triptofán jelenlétének detektálását, igen ötletesen valósítja meg a
baktérium. Ahhoz, hogy a triptofán-represszor az operátorhoz kötődjék, két triptofánt
kell magának is kötnie. A triptofánok megkötése térszerkezet változást, egy hajlást ered-
ményez a hélix-turn-hélix szerkezetben, amely ezt követően már tökéletesen illik a DNS
nagyárkába (a promóterrel átlapoló) operátorrégiónál (5.17. ábra). A triptofán meg-
kötése nélkül a DNS-kötésre alkalmas részlet a fehérje belsejében marad, így nem képes
a DNS-kötésre a fehérje. Tulajdonképpen a triptofán-represszor egy allosztérikusan
szabályozott fehérje, amely esetében a regulátormolekula maga a triptofán. Így a
triptofán-represszor és operon egy egyszerű alkalmatosságot formál, amely a triptofán
jelenlététől függően be- vagy kikapcsolja a triptofán bioszintézisének enzimeit. Tekintve,
hogy a fehérje aktív, DNS-kötő formája, kikapcsolja a gént, a szabályozást negatív
kontrollnak, a fehérjét pedig transzkripciós vagy génrepresszor fehérjének nevezzük.
5.17. ábra. A triptofán-represszor fehérje szerkezetének és DNS-kötő képességének megváltozása a két bekötődő triptofánmolekula hatására

64 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
Néhány bakteriális promóter szekvencia csak alacsony hatásfokkal működik, mivel
az RNS-polimeráz csak alacsony hatásfokkal ismeri fel és/vagy az RNS-
polimeráznak nehézségei adódnak a DNS-kettős hélix felnyitásával és a
transzkripció megindításával.
Mindkét esetben, az alacsony hatásfokkal működő promóterek esetében is, elindul-
hat a transzkripció, amennyiben egy génregulációs fehérje kapcsolódik a promóter
közelébe a DNS-lánchoz és kapcsolatba lép az RNS-polimerázzal. Így a transzkripció
megindításának valószínűsége jelentős mértékben megnő. Tekintve, hogy ezen fehérjék
aktív DNS-kötő formája bekapcsolja a gént, a folyamatot pozitív kontrollnak, a fehérjéket
pedig transzkripciós vagy génaktivátor fehérjéknek nevezik.
Az esetek egy részében a génaktivátor fehérjék oly módon segítenek az RNS-poli-
meráz promóterhez kötődésében, hogy egy újabb kapcsolódó felületet biztosítanak a
polimeráznak. Más esetekben a polimeráz transzkripciós állapotba alakulását segítik elő
azáltal, hogy stabilizálják a DNS-kötő forma és a transzkripciós forma közti átmeneti
állapotot.
Ahogy a negatív kontroll (a transzkripciós represszor) esetében láttuk, úgy a pozitív
szabályozás esetében is a transzkripciós aktivátorfehérje képes egyszerű genetikai
kapcsolóként működni.
A pozitív kontrollra jó példát szolgáltat a bakteriális katabolit aktivátor fehérje
(catabolit activator protein: CAP). A CAP képessé teszi az E. colit arra, hogy alternatív
szénforrásokat használjon fel, ha az egyébként preferált szénforrása, a glukóz, nem áll
rendelkezésre (azáltal, hogy különböző géneket kapcsol be).
A glukózszint csökkenése elindítja az intracelluláris szignálmolekula, a cAMP
szintjének emelkedését, amely képes a CAP-vel történő összekapcsolódásra. A kapcsolat
létrejöttét követő térszerkezet-változásnak köszönhetően (allosztérikus reguláció) a
CAP alkalmassá válik, hogy a célpromóterek mellett specifikus DNS-szakaszokhoz
kössön, így bekapcsolva a megfelelő géneket. A kérdéses gén be- vagy kikapcsolása tehát
attól függ, hogy a cAMP szintje miként alakul a sejtben.
Sok esetben a represszorok és az aktivátorok felépítése hasonló. Pl.: a triptofán-
represszor és a CAP-aktivátor is hélix-turn-hélixrészletet tartalmaz és egy kis kofaktort
igényel a DNS-hez kötődéshez. Igazság szerint néhány bakteriális fehérje (CAP,
bakteriofág λ-represszor) mind aktivátorként, mind represszorként képes viselkedni
attól függően, hogy az általuk felismert DNS-szekvencia hogyan helyezkedik el a
promóterhez képest. Ha a fehérje kötőhely átlapol a promóterrel, a polimeráz nem tud
kötődni és a fehérje mint represszor hat. Ha azonban a fehérje a promóterszekvencia
mellett kötődik, mint egy jelzőbója segíti az RNS-polimeráz bekötődését, a transzkripció
megindulását. Ebben az esetben mint aktívátor viselkedik.
5.2.2. A lac-operon: transzkripciós aktiválás/represszálás
A pozitív és a negatív szabályozás kombinációjára jó példát szolgáltat a CAP- és lac-
represszor által megvalósított szabályozás, mely révén a baktérium alkalmazkodni tud,
képessé válik glukóz hiányában más alternatív szén- és energiaforrások (pl. laktóz)

5. A génexpresszió szabályozása 65
© Szarka András www.interkonyv.hu
felhasználására. A laktóz sejtbe juttatásáért, illetve lebontásáért felelős géneket és
szabályozó részleteket a lac-operon tartalmazza. Ahogy korábban láttuk, a glukózszint
csökkenésével megemelkedő cAMP-szint emelkedésével és CAP-fehérjével való
kapcsolat révén a CAP képessé válik a DNS-hez való kötődésre és elősegíti az alternatív
szén/energiaforrások (pl.: laktóz) felhasználását célzó fehérjék átírását. Az azonban
komoly veszteséggel járna, ha a CAP laktóz távollétében is kifejezné a fehérjéket, ezért a
lac-represszor ezeket, laktóz távollétében, kikapcsolja. Így tulajdonképpen két szignál
integrálása történik meg és a lac-operon magas szinten csak akkor expresszált ha:
1. nincs jelen glukóz, 2. és jelen van laktóz.
Minden más esetben a gének ki vannak kapcsolva.
Nézzük meg, hogyan történik mindez a gyakorlatban. Laktóz adagolására a kevés
jelenlévő laktóz-permeázon keresztül kis mennyiségű laktóz képes a sejtbe jutni és ott a
kevés -galaktozidáz segítségével a laktóz egy izomerévé, allolaktózzá alakul. Az allo-
laktóz fog tulajdonképpen induktorként viselkedni, mivel hozzáköt a lac-represszorhoz,
amely allolaktózt nem kötő formája képes a DNS-hez kötni, a lac-promóterrel átlapoló
DNS-szekvenciához. Az allolaktóz bekötődése térszerkezet-változást eredményez a lac-
represszor fehérjében, amelynek hatására az már nem képes a DNS-hez kötődni, így
felszabadul a gátlás alól a lac-operon (5.18. ábra).
Tekintve, hogy a lac-operon promótere egy gyenge promóter, a megfelelő mértékű
génexpresszióhoz szükséges a glukóz távollétében megemelkedő cAMP-szint hatására
bekötődő CAP DNS-hez kötése. A kikapcsolás természetesen éppen fordított módon
történik meg: a glukóz adagolása csökkenti a sejtben a cAMP-szintjét, mivel így a cAMP
nem tud a CAP-hoz kötődni, leválik a DNS-ről, ennek eredménye a gén kikapcsolása lesz
(5.18. ábra).

66 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
5.18. ábra. A lac-operon kettős szabályozása a lac-represszor fehérje, illetve a CAP–cAMP-komplex által
5.3. Az eukarióta génexpresszió szabályozása
A bakteriális génexpresszió szabályozása, ahogy láttuk, viszonylag egyszerű, de korláto-
zásokat tartalmaz. A promóter szomszédságában egész egyszerűen nincs elegendő hely,
hogy elég legyen a nagyobb számú DNS-regulációs szekvencia számára. Éppen ezen
korlátok miatt a szignálok tucatjainak kezelésére alkalmatlan.
Vajon, mi módon képesek az eukarióta sejtek ezen korlátokon felülemelkedni?
Az eukarióta transzkripció szabályozása három fő részletben is különbözik a bak-
teriálistól:
1. Olyan génregulációs fehérjéket alkalmaznak, amelyek képesek akár a több ezer
bázispárnyira lévő promóter befolyásolására, így tehát egy promótert
gyakorlatilag számtalan DNS-szakasz regulálhat a teljes DNS-állományon belül.
2. Az eukarióta RNS-polimeráz-II (amely az összes fehérjét kódoló gén transzkrip-
ciójáért felelős) önmaga nem képes elindítani a transzkripciót. Fehérjék egy
csoportjának közreműködését igényli, melyeket általános transzkripciós fakto-
roknak nevezünk. Ezen fehérjék transzkripciós iniciációs komplexszé történő

5. A génexpresszió szabályozása 67
© Szarka András www.interkonyv.hu
összeszerelése a transzkripció megindulásának (iniciáció) előfeltétele. Ezek
teljesen általános (konzervált) fehérjék minden egyes sejtben azonosak, amely-
ben RNS-polimeráz-II van, nem specifikusak, mint a génregulációs fehérjék. Ezen
fehérjék összeszerelésének befolyásolása egy jó szabályozási lehetőség, sok euka-
rióta génregulációs fehérje a különböző regulációs szignálok eredményeképpen
ezen a ponton fejti ki hatását.
3. Az eukarióta DNS kromatinba csomagolódása miatt a bakteriális szabályozási
lehetőségek nem alkalmazhatók ebben az esetben.
5.3.1. Eukarióta génregulációs fehérjék
Az eukarióta sejtek is, a prokariótákhoz hasonlóan, génregulációs fehérjéket alkal-
maznak, melyek közt aktivátorok és represszorok egyaránt megtalálhatók.
A DNS-szakaszt, melyhez a regulációs fehérjék kötnek, enhancereknek nevezzük. Az
elnevezés alapja az a megfigyelés volt, mely szerint ezek jelentős mértékben megnövelik
a transzkripciót.
1979-ben került leírásra az a megfigyelés, mely szerint az aktivátorfehérje a
promótertől akár több ezer bp távolságra is köthet, illetve attól függetlenül képes
génexpressziót befolyásoló hatását kifejteni, hogy a génhez képest upstream vagy
downstream kötődik. Természetesen adódik a kérdés, hogyan képesek így a promóterrel
kommunikálni? A leggyakrabban alkalmazott módszer, hogy a DNS az enhancer és a
promóter között egy hurkot formál, így az enhancerhez kapcsolódott aktívátorfehérjék
és a promóterhez kapcsolódott fehérjék (RNS-polimeráz, általános transzkripciós
faktorok) egymással kapcsolatot teremthessenek.
A DNS mint egy kötél viselkedik, segítve a két fehérjecsoportosulás találkozását
(5.19. ábra). Baktériumokban is ismeretes ez a módszer, azonban sokkal ritkábban és
rövidebb DNS-szakaszokkal valósul meg.
5.19. ábra. A DNS hurokképzése az enhancer- és a promóterrégiók között lehetővé teszi egymástól távoli szekvenciák egymásra hatását

68 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
5.3.2. Eukarióta DNS-reguláló szekvenciák
Tekintve, hogy az eukarióta génregulációs fehérjék a promótertől igen távol a DNS-hez
kötődve befolyásolni képesek a transzkripciót, a transzkripciót befolyásoló génregu-
lációs régió igen nagy lehet (5.20. ábra). A kiterjedt génregulációs szakaszok miatt benne
foglaltatik a promóter (a hozzá kapcsolódó fehérjék: RNS-polimeráz, általános transz-
kripciós faktor), illetve a regulációs szekvenciák (a hozzá kapcsolódó fehérjék: regulá-
ciós fehérjék) (5.20. ábra).
A magasabb szintű eukarióta sejtekben akár 50 000 bp távolságra is lehet a
regulációs szekvencia, illetve fehérje a promótertől. A köztük lévő flexibilis DNS-szakasz
segíti a promóter és enhancer összehangolását (5.19. és 5.20. ábra). Szintén fontos
megemlítenünk, hogy az eukarióta DNS nukleoszómákba és kromatinba pakolódik,
ezáltal is lecsökkentve a távolságot a két DNS-szakasz (regulációs és promóter) között
(5.21. ábra).
A következőkben tekintsük át röviden a két nagyobb DNS-hez kötődő fehérjecsoport
sajátságait:
Általános transzkripciós faktorok
Nagy mennyiségben előforduló,
Kis változatosságot mutató fehérjék,
Ahol RNS-polimeráz II van.
Génregulációs fehérjék
Több ezer fajta, a 25 000 humán gén 5-10%-a
ilyet kódol, egyik génregulációs szakaszról a
másikra változnak, kis mennyiségben
(általában kevesebb, mint 0,01%-a a sejt teljes
fehérjeállományának).
A megfelelő DNS-szakaszt ismerik fel a
korábban tárgyalt fehérjerészletek
segítségével, illetve más DNS-felismerő
fehérjék felépítésében vesznek részt.
A génregulációs fehérjék egy organizmus egyes génjeit kapcsolják ki vagy be. Ahogy
korábban említésre került, a különböző sejttípusok a génregulációs fehérjék egyedi
összeállítását tartalmazzák. Így génexpressziós mintázatuk is egyedi, eltérő sajátosságot
adva az egyes sejttípusoknak.
Az eukarióta sejt minden egyes génje különbözőképpen regulálódik. Ez meg-
lehetősen nagy szabályozási diverzitást jelent. Éppen ezért igen nehéz általános érvényű
szabályokat felállítani a szabályozásukról, azonban a főbb irányvonalak megállapít-
hatóak.

5. A génexpresszió szabályozása 69
© Szarka András www.interkonyv.hu
5.20. ábra. Az eukarióta gén szabályozási régiójának elemei
5.3.3. Génaktivátor fehérjék és a transzkripció
A legtöbb génregulációs fehérje moduláris felépítéssel rendelkezik. Két fő domén
különböztethető meg: a DNS-felismerő részlet (ezeket már korábban tárgyaltuk), illetve
az aktivációs domén. Ez utóbbi felelős a transzkripció gyorsításáért. A moduláris
szerkezet leírása hibridfehérjék segítségével történt, melyekben az egyik fehérje DNS-
felismerő részét fuzionáltatták a másik fehérje aktivációs részével.
Lássuk, hogyan működhetnek a génaktivátor fehérjék! Tulajdonképpen két fő hatásmódozatot különböztethetünk meg:
1. Vonzzák, pozícionálják, módosítják az általános transzkripciós faktorokat és az
RNS-polimeráz II-t a promóteren. Mindezek hatására elindulhat a transzkripció
(5.20. ábra);
2. Ezen direkt hatáson kívül megváltoztathatják a kromatinszerkezetet a promóter
körül (5.21. ábra).
1. A direkt hatás két módon érvényesülhet: Az RNS-polimeráz lépésenként állhat
össze aktív holoenzimmé, in vitro leírt sorrendben, de ismeretesek olyan sejtek
is, amelyekben már összeállt formában kerül a promóterre az RNS-polimeráz
holokomplex. Az RNS-polimeráz holoenzim az általános transzkripciós faktoro-
kon és az RNS-polimerázon kívül egy 20 fehérjéből álló, mediátornak is nevezett
komplexet is tartalmaz.

70 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
Számos aktivátor fehérje kölcsönhat a holoenzim-komplexszel, energetikailag
(még) kedvezőbbé téve a promóterhez kötődést. Tulajdonképpen az egész
folyamat úgy fogható fel, mintha az aktivátorok a holoenzimet vonzanák,
pozícionálnák a promóterhez (5.20. ábra).
Ezt az elképzelést természetesen kísérleti úton is bizonyították.
Egy szekvencia specifikus (DNS-felismerő) részletet fúzionáltattak közvetlenül a
mediátor egyik komponenséhez (teljes transzkripciós RNS-polimeráz-komplex). Ez a
hibridfehérje, amelyből az aktivációs domén hiányzott, jelentős mértékben elősegítette a
transzkripció megkezdését, ha a DNS-szekvencia, amelyhez kötődött, a promóter-
szekvencia szomszédságában volt. Habár a holoenzim-komplex promóterhez irányítása
látványos, egyszerű módja az aktiválás folyamatának szemléltetésére, valószínűleg az
aktivátorok működése ennél jóval bonyolultabb. Ilyen lehet egyes esetekben az általános
transzkripciós faktorok lépésenkénti összeszerelése a promóteren. Más esetekben a
megkötődést követő átrendeződésük szükséges. A legtöbb holoenzim-komplexből
hiányoznak az általános transzkripciós faktorok (elsősorban a TFIID és a TFIIA),
amelyeknek a promóteren kell külön beépülniük.
Ezen lépések bármelyike egy-egy lassú (sebesség meghatározó) lépés lehet a
transzkripció megkezdése során. Az aktivátorok hozzájárulhatnak a teljes forma
kialakításához, lehetőséget adva a szabályozásra.
Számos aktivációs fehérjéről kimutatták, hogy kapcsolatba lép általános
transzkripciós faktorokkal és számos direkt módon is segíti a promóteren történő
összeszerelésüket.
2. A lokális kromatinszerkezet megváltoztatása
A regulációs és a promóter részek kromatinszerkezetének megváltoztatása révén is
kitűnően befolyásolható a génexpresszió.
A leggyakoribb két lokális kromatinszerkezet-változás: a nukleoszóma-remodellezés
és a kovalens hisztonmódosítás (5.21. és 5.22. ábra). Számos génszabályozó fehérje hasz-
nálja mindkét módszert, az enzimek két csoportjához, a hiszton-acetiltranszferázhoz
(hiszton-acetilázt) és az ATP-dependens kromatin-remodellező komplexhez kötve és
ezáltal létrehozva aktív, funkcióképes formájukat (5.21. és 5.22. ábra).
Általánosságban elmondhatjuk, hogy a kromatinszerkezetben bekövetkező változás
nagyobb hozzáférhetőséget biztosít az alapjául szolgáló DNS-nek. Ez a jobb hozzáfér-
hetőség hozzájárul, hogy az általános transzkripciós faktorok és az RNS-polimeráz
holoenzim összeszerelődjön a promóteren és ahhoz, hogy további génregulációs fehér-
jék kapcsolódjanak a regulációs szakaszhoz.
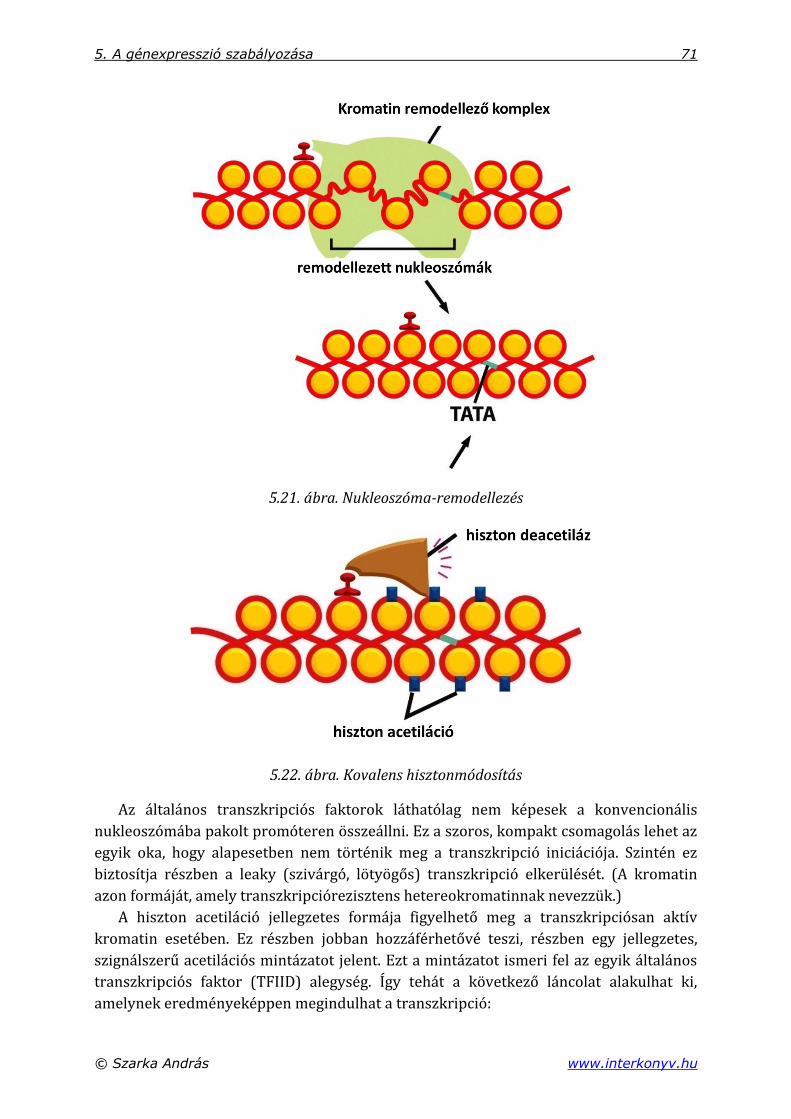
5. A génexpresszió szabályozása 71
© Szarka András www.interkonyv.hu
5.21. ábra. Nukleoszóma-remodellezés
5.22. ábra. Kovalens hisztonmódosítás
Az általános transzkripciós faktorok láthatólag nem képesek a konvencionális
nukleoszómába pakolt promóteren összeállni. Ez a szoros, kompakt csomagolás lehet az
egyik oka, hogy alapesetben nem történik meg a transzkripció iniciációja. Szintén ez
biztosítja részben a leaky (szivárgó, lötyögős) transzkripció elkerülését. (A kromatin
azon formáját, amely transzkripciórezisztens hetereokromatinnak nevezzük.)
A hiszton acetiláció jellegzetes formája figyelhető meg a transzkripciósan aktív
kromatin esetében. Ez részben jobban hozzáférhetővé teszi, részben egy jellegzetes,
szignálszerű acetilációs mintázatot jelent. Ezt a mintázatot ismeri fel az egyik általános
transzkripciós faktor (TFIID) alegység. Így tehát a következő láncolat alakulhat ki,
amelynek eredményeképpen megindulhat a transzkripció:

72 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
Génregulációs fehérjék → aktiválják a hiszton-acetilázokat → közvetetten segítik az
általános transzkripciós faktorok összeszerelését a promóteren → transzkripció.
5.3.3.1. A génregulációs fehérjék szinergikusan dolgoznak
A génregulációs fehérjék több különböző lépést képesek a transzkripció inicializációja
végett befolyásolni. Amennyiben egyszerre több faktor hatása érvényesül a reakció-
sebesség megnövelése érdekében, a reakciósebesség növekménye nem csupán az egyes
hatások összege, hanem ennél jelentősebb. Példának tekinthetjük a következő esetet,
amikor A-faktor hatására 1-szeres energiagát-csökkentés következik be, melynek ered-
ménye 100-szoros reakciósebesség-növekedés. Amennyiben B-faktorral, melynek
ugyanolyan mértékű hatása van külön, mint az A-faktornak külön, egyszerre hatnak,
annak 2-szeres energiagát-csökkentés lesz az eredménye, amely 10 000-szeres reakció-
sebesség-növekedést fog eredményezni. Ez a szinergia a transzkripció megkezdését
illetően is érvényes, különös tekintettel arra, hogy a génaktivátor fehérjék számos lépést
befolyásolnak.
5.3.3.2. Eukarióta represszorok
Az eukarióta génregulációs fehérjék legtöbbje, több (néha egész sok) alegységből álló
komplexet alkotnak a DNS-en. Ezen alegységek különböző funkciókkal rendelkeznek. A
komplexek igen gyakran csak a megfelelő DNS-szekvencia jelenlétében állnak össze.
Néhány alaposan tanulmányozott esetben például két egymás iránt kis affinitást mutató
génregulációs fehérje kooperál, hogy hozzákössön egy DNS-szekvenciához. Önállóan
egyik fehérje sem rendelkezik elegendő affinitással az effektív DNS-hez kötődéshez. A
DNS-kötődést követően egy olyan felületet alkot a dimer, amelyet egy harmadik fehérje
ismer fel, amelyik egy aktivációs domént hordoz. Az így létrejött komplex már
elindíthatja a transzkripciót.
Mindezek alapján a következő általános megállapítást tehetjük: azok a fehérje-
fehérje kapcsolatok, amelyek az oldatban túl gyengének bizonyulnak, hogy a fehérje
összeállását eredményezzék, elegendőnek bizonyulnak a DNS felületén. A DNS ez
esetben mint egy kristálygóc vagy összeszerelési mag szerepel.
Egy adott génregulációs fehérje több regulációs komplexben is részt vehet. A fehérje
egyik esetben egy aktivátor része lehet, míg más esetben egy represszorban szerepelhet
(5.23. ábra). Így nem igazán lehet a különálló génregulációs fehérjéket aktivátornak vagy
represszornak nevezni. Ehelyett mint regulációs egységek szerepelnek, melyek végső
sorsa a különálló egységekből szerveződő komplexekben dől el. Ez a sors (össze-
szerelődés) a regulációs DNS-szekvenciától, illetve a sejtben éppen jelenlévő regulációs
fehérjéktől függ.
A DNS-hez nem kötő, de DNS-kötő regulációs fehérjékben részt vevő génregulációs
fehérjéket koaktivátoroknak, vagy korepresszoroknak nevezzük. Ugyanazon koaktivá-
torok vagy korepresszorok különböző DNS-kötő fehérjékre épülhetnek (5.23. ábra). A
koaktivátorok, korepresszorok különböző feladatokat láthatnak el: kromatin remodel-
lező komplexekkel, hiszton-módosító enzimekkel, RNS-polimeráz holoenzimekkel, álta-
lános transzkripciós faktorokkal kapcsolódhatnak.

5. A génexpresszió szabályozása 73
© Szarka András www.interkonyv.hu
5.23. ábra. A szabályozó fehérjeelemek összeszerelődése különböző funkciójú regulátorkomplexekké a DNS felületén
Néhány esetben a DNS-szekvencia, melyhez a génregulátor fehérjék kötődnek,
megváltoztatja a konformációjukat, ezáltal befolyásolva a következő transzkripciós
aktivitását.
Például, ha egyfajta DNS-szekvenciához kötődik egy szteroid humán receptor, akkor
az egy korepresszorral kapcsolódik és kikapcsolja a transzkripciót. Ha egy másik, kis-
mértékben különböző DNS-szekvenciához, akkor más konformációt felvéve egy koakti-
vátorhoz kapcsolódik és bekapcsolja a transzkripciót. Általában egy génregulációs
komplex szerveződését rövidke DNS-szakaszok vezérlik (5.23. ábra), azonban van arra
példa, hogy egy jóval kiterjedtebb DNS-fehérje komplex szerveződjék, melyet enhanszo-
szómának nevezünk. Az enhanszoszómák ismertetőjegyei az építőfehérjék (5.24. ábra),
amelyek egy jól definiált szögben hajlítják a DNS-t és ezáltal elősegítik a többi enhanszo-
szómafehérje beépülését. Tekintve, hogy az enhanszoszóma felépítéséhez számos fehér-
je szükségeltetik, ez biztosítja, hogy csak akkor fejeződik ki a gén, ha ezen fehérjék mind-
egyike jelen van a sejtben. Ez a felállás egyben magas fokú kombinációs kontrollra is
lehetőséget ad.
5.24. ábra. Az enhanszoszóma felépítése

74 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
5.3.3.3. Inzulátor DNS-szekvenciák
Bizonyára még mindenki emlékszik rá, hogy az eukarióta génkifejeződés szabályozása
során a regulációs fehérjék a promórhez képest mind upstream, mind downstream
beköthetnek az enhancer szekvenciához. Így egyértelműen adódik a kérdés, hogy mi
biztosítja azt, hogy ha egy génregulációs fehérje beköt a helyére, az a szomszédos gének
átíródását nem befolyásolja?
A legismertebb megoldás, amellyel az eukarióta sejt a probléma megoldása érde-
kében előállt, az inzulátor (határoló) elemek (5.25. ábra). Az inzulátor elemek DNS-
szakaszok, melyek specializálódott fehérjéket kötnek és két speciális tulajdonsággal
rendelkeznek: 1. pufferelik a géneket a heterokromatin represszáló hatásától; 2. képesek
az enhancerek blokkolására. Ahhoz, hogy erre képesek legyenek, a promóter és az
enhancer rész között kell lokalizálódniuk. Mindazonáltal az inzulátorok pontos műkö-
dési mechanizmusa jelenleg ismeretlen és könnyen elképzelhető, hogy a különböző
inzulátorok eltérő mechanizmusok révén fejtik ki hatásukat.
5.25. ábra. Az inzulátorszekvenciák elhelyezkedése és funkciója
5.4. Az X-kromoszóma inaktiválása nőkben
Az előző fejezetben láttuk, hogy a kromatinszerkezet befolyásolhatja a gén-
expressziót. Emlőssejtek esetében akár egy teljes kromoszóma minden egyes génjének
átírására is kiterjedhet.
A férfiak és a nők (többek között) a nemi kromoszómáikban térnek el egymástól. A
férfiak XY, a nők XX nemi kromoszómákkal rendelkeznek. A nőkben tehát kétszer annyi
kópia található az X-kromoszóma génjeiből, mint a férfiakban. Ez igen jelentős
különbséget eredményez, különösen akkor, ha figyelembe vesszük, hogy az X-
kromoszóma jóval nagyobb, több mint 1000 gént tartalmaz, mint a kevesebb mint 100
gént tartalmazó Y.
A férfiak és a nők között tehát jelentős különbség alakult ki az X-génen található
gének mennyiségében. Az emlősökben egy kompenzációs mechanizmus alakult ki, amely
révén a férfiakban és a nőkben az X-kromoszóma géntermékei között levő különbség
kiegyenlíthető.
Azok az esetek, ahol a mennyiségi kompenzációban mutáció következik be letálisak,
jól szemléltetve, hogy milyen fontos fenntartani a megfelelő nemi/autoszomális gén-
arányt, gén-termékarányt.
Lássuk csak, mi módon kerül megvalósításra a kompenzáció! A nők szomatikus
(testi) sejtjeiben az egyik X-kromoszóma transzkripciósan inaktiválódik. A transzkrip-
ciós lecsendesülésre a női embrió fejlődésének korai szakaszában kerül sor, amikor még
csak néhány ezer sejtből áll. Ekkor az X-kromoszómák egyike minden egyes testi sejtben

5. A génexpresszió szabályozása 75
© Szarka András www.interkonyv.hu
jelentős mértékben kondenzálódik – heterokromatin struktúrát vesz fel. A konden-
zálódott X-kromoszóma fénymikroszkóppal is jól látható az interfázisban lévő sejtekben.
Ezt a mikroszkóppal is jól látható, kondenzálódott kromoszómát Barr-testnek nevezték
el. Az X-inaktiválódásnak köszönhetően két X-kromoszóma létezhet egyszerre egyazon
sejtmagban ugyanazoknak (a szignáloknak kitéve) a transzkripciós regulátor fehér-
jéknek kitéve, mégis teljesen különböző mértékű génexpressziót mutatva.
Az, hogy az apai vagy az anyai X-kromoszóma inaktiválódik, teljesen random dől el.
Ha már egyszer az egyik inaktiválódott, akkor az, változatlan marad a sejt további
osztódása során is benne és utódaiban is, hűen megőrizve azt, DNS replikációkon és
mitozisokon keresztül.
Tekintve, hogy az X-inaktiválódás random megy végbe, amikor már néhány ezer
sejtből áll az embrió, minden egyes nő különböző sejtek mozaikja, melyek egyikében az
apai, másikében az anyai X-kromoszóma csendes, inaktiválódott. Ezen sejtek kisebb
klasztereket alkotnak a felnőttben, mivel a testvérsejtek egymás közelében maradnak
sejtosztódást követően a későbbi fejlődés során (5.26. ábra).

76 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
5.26. ábra. Az X-kromoszóma inaktiválódása (kondenzálódása) a női embrionális testi sejtekben
Erre kitűnő szemmel látható példát is találhatunk, hiszen a macskák esetében ez
okozza a vörös-fekete teknőspáncélszerű bundamintázatot a nőstény macskák esetében.
Az egyik X-kromoszómán található a gén, amely a vörös szőrért felelős. A másik X-
kromoszómán ugyanazon gén másik allélja, amelyik a fekete szőrzetért felelős. A ran-
dom X-inaktiváció miatt alakul ki az érdekes foltos szőrzetmintázat. A hím macskák vagy
teljesen vörösek, vagy teljesen feketék lesznek attól függően, hogy az anyjuktól melyik
színt kódoló X-kromoszómát örökölték. Habár az X-kromoszóma inaktiváció ezer és ezer
sejtosztódáson keresztül megtartott, nem mindig állandó. Nevezetesen visszafordítható
az ivarsejtek kialakulásakor, vagyis minden egyes haploid petesejt egy aktív X-
kromoszómát tartalmaz és képes az X-kromoszómához kötött termékek kifejezésére.
Hogyan következik be a transzkripciós inaktiválódás egy egész kromoszómán?
Az X-kromoszóma inaktiválódása az X-kromoszóma közepéről, egy pontról kiindulva
indul és terjed tova. Ezt a helyet X-inaktivációs központnak (XIC) nevezik. Ezt a
központot (106 bp-nyi DNS-szakasz) egy nagy szabályozó elemnek tekinthetjük, amely a

5. A génexpresszió szabályozása 77
© Szarka András www.interkonyv.hu
heterokromatin kiindulási magja és segíti annak tovaterjedését mindkét irányban a
teljes kromoszóma mentén (5.27. ábra). Az XIC-on belül kódolva található egy szokatlan
RNS-molekula: az X inaktivációs specifikus transzkript (XIST), amely kizárólag az
inaktivált X-kromoszómáról expresszálódik. Ezen RNS expressziója elengedhetetlen az
X-inaktivációhoz. Egy igen érdekes RNS-ről van szó, amely nem íródik át fehérjévé, a
sejtmagban marad és bebiztosítja az inaktív X-kromoszómát. Az XIST-RNS tovaterjedése
a kromoszómán jól korrelál a gének elcsendesülésével, jól mutatja, hogy az XIST-RNS
részt vesz a heterokromatin formálásában, valamint terjedésében (5.27. ábra).
5.27. ábra. Az X-kromoszóma inaktiválódásának mechanizmusa
X-kromoszóma heterokromatin számos sajátsággal rendelkezik, amelyek mindegyike
hozzájárul a kromoszóma transzkripciós csendességéhez. Ezek a XIST, a hiszton-2A, az
alacsony szinten acetilált H3-, H4-hisztonok és végül a specifikus helyen metilált H3-
hiszton és DNS.
Az X-kromoszómával kapcsolatos tudásunk még meglehetősen hiányos.
Még megválaszolásra vár:
1. Hogyan dől el, hogy melyik (az apai, vagy az anyai) X-kromoszóma inaktiválódik?
2. Milyen mechanizmus védi a másik X-kromoszómát az inaktiválódástól?
3. Hogyan koordinálja az XIST-RNS a heterokromatin szerveződését?
4. Hogyan marad inaktív a kromoszóma a sejtosztódások tömkelegén keresztül?

78 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
5.5. DNS-metiláció
A regulációs fehérjéken kívül maga a DNS is fontos szabályozóelemként szerepelhet,
kovalensen módosítható. Gerincesekben a citozin metilálásának van fontos szerepe a
gének (in)aktiválásában. A citozin metilálása nem befolyásolja a bázispárok kialakulását.
A gerincesek DNS-ében a metiláció kizárólag a citozin nukleotidokat érinti, ott is a CG-
báziskettősben találhatókat, melyek ugyanazzal a szekvenciával párosodnak természete-
sen ellentétes orientációval: a CG-vel szemben lévő DNS-szálon GC-báziskettős található
(5.28. ábra).
Egy viszonylag egyszerű módszer biztosítja, hogy a meglévő DNS-metilációs mintázat
öröklődjék tovább a DNS-leányszálakba. A metiltranszferáz enzimek azokon a CG-bázis-
párokon dolgoznak, amelyek már metilált CG-bázispárokkal vannak párban. A szülői
szálakon található metilációs mintázat mintegy mintául szolgál a leányszálak metilációs
mintázatához, ezáltal az direkt módon továbböröklődik (5.28. ábra). A DNS-metilációs
mintázat ilyeténképp történő stabil továbböröklődéséért a DNS-metiltranszferázok
felelősek.
5.28. ábra. A DNS-metiláció mechanizmusa, a fenntartó metilázok működése
Fontos megjegyeznünk, hogy a DNS-metilációs mintázat a gerincesek fejlődése alatt
dinamikus változást mutat. Kevéssel a megtermékenyítést követően egy demetilációs
hullám vonul végig a teljes genomon, amely során a metilcsoportok jelentős többsége
eltűnik a DNS-ről. Ez a demetilálódás bekövetkezhet egyrészről a metiltranszferázok
szupressziója miatt, replikációról replikációra történő passzív demetilációval, vagy egy
specifikus demetiláló enzim által. A fejlődés egy későbbi szakaszában, amikor a magzat a
méh falába ágyazódik be, egy új metilációs mintázat alakul ki. Az új metilációs mintázat
kialakulásáért a de novo DNS-metiltranszferázok felelősek, amelyek a metilálatlan CG-
párosokat veszik kezelésbe. Ha már egyszer kialakult a metilációs mintázat, annak
fenntartásáért a fenntartó DNS-metiltranszferázok felelősek. Mind a fenntartó, mind a de
novo metiltranszferázokban bekövetkezett mutáció korai embrionális halált ered-
ményezett egerekben, ami egyértelműen arra utal, hogy a megfelelő metilációs mintázat
sorsdöntő a normális fejlődésben.

5. A génexpresszió szabályozása 79
© Szarka András www.interkonyv.hu
Gerincesekben a DNS-metiláció elsősorban transzkripciósan inaktív régiókban
található, úgy, mint az inaktív X-kromoszóma, vagy az adott szövetben inaktivált gének
esetében.
Ez a megfigyelés arra enged következtetni, hogy a gének inaktiválásában lehet
szerepe. A gerincesek sejtjeiben számos fehérje ismeri fel a metilált DNS-t és köt hozzá.
Ezen DNS-kötő fehérjék ezt követően kapcsolatba kerülnek kromatin remodellező
komplexekkel, illetve hiszton deacetiláló enzimekkel, amelyek kondenzálják a kromatint,
transzkripciósan inaktívvá téve azt. Igen ám, de a DNS-metilálás önmagában nem
elegendő, hogy transzkripciósan inaktív legyen a DNS.
Ezt egy ötletes kísérlettel bizonyították is, mely során teljes mértékben metilált,
illetve teljes mértékben demetilált izomspecifikus aktint tartalmazó plazmidot hoztak
létre. Amikor ezeket izomsejt tenyészetbe vitték, a teljes mértékben metilált plazmid
ugyanolyan magas mértékben kifejeződött, mint a nem metilált változat.
Ezen kívül, ha bekapcsolnak egy csendes gént, a metiláció csak később, jó néhány
transzkripciót követően kerül le róla. Valamint az X-kromoszóma inaktivációja még jóval
a detektálható metilációja előtt bekövetkezik. Mindezek arra utalnak, ha egyszer kikap-
csoltak egy gént, a metiláció csak megerősíti annak represszióját. A primer mecha-
nizmus nem a metiláció, az csak egy további biztosíték. Két különböző típusú gerinces
sejtben egy adott gén expressziója 106-szorosa lehet a másikénak. Baktériumban egy
expresszált és egy nem expresszált gén transzkripciója között mindössze 1000-szeres
különbség lehet. Gerincesekben a DNS metiláció is hozzájárulhat (a kromatinban
bekövetkező változásán keresztül) ehhez a nagy különbséghez. A több ezer gén
(amelyek normális körülmények között ki vannak kapcsolva) lötyögős transzkripciója is
szerepet játszhat, hogy a DNS-metiltranszferáz mutáns egérembriók életképtelenek.
A baktériumokban lötyögős a transzkripciós szabályozása, vagyis a nem expresszált
gének is transzkripcióra kerülnek bizonyos százalékban. Ez a kontroll gerincesekben,
emlősökben sokkal szigorúbb, amelyhez a DNS metilációja is hozzájárulhat. Nagyobb
különbség van az expresszált és a nem expresszált transzkript szintjében.
Gerincesekben a transzkripciós csend szintén fontos szerepet játszik az ugráló
elemek (transzpozonok) proliferációjának visszaszorításában. Amíg a kódoló rész csak
néhány százaléka a teljes genomnak, addig az ugráló elemek majdnem a felét alkot-
hatják. Ezek elkészíthetik saját maguk kópiáját és ezeket bárhová beilleszthetik a geno-
mon belül, képesek lévén géneket és regulációs szakaszokat megszakítani. A metiláció
révén visszaszorítható ezen elemek transzkripciója, így elősegítvén a genom integri-
tásának megőrzését.
5.5.1. DNS-metiláció a genomi mintázatban
Az emlőssejtek, diploidok tartalmaznak egy génszettet az apától és egyet az anyától.
Néhány esetben a gén expressziója attól függ, hogy az az anyától vagy az apától
származik (5.29.ábra). A jelenséget genomi lenyomatnak nevezik.
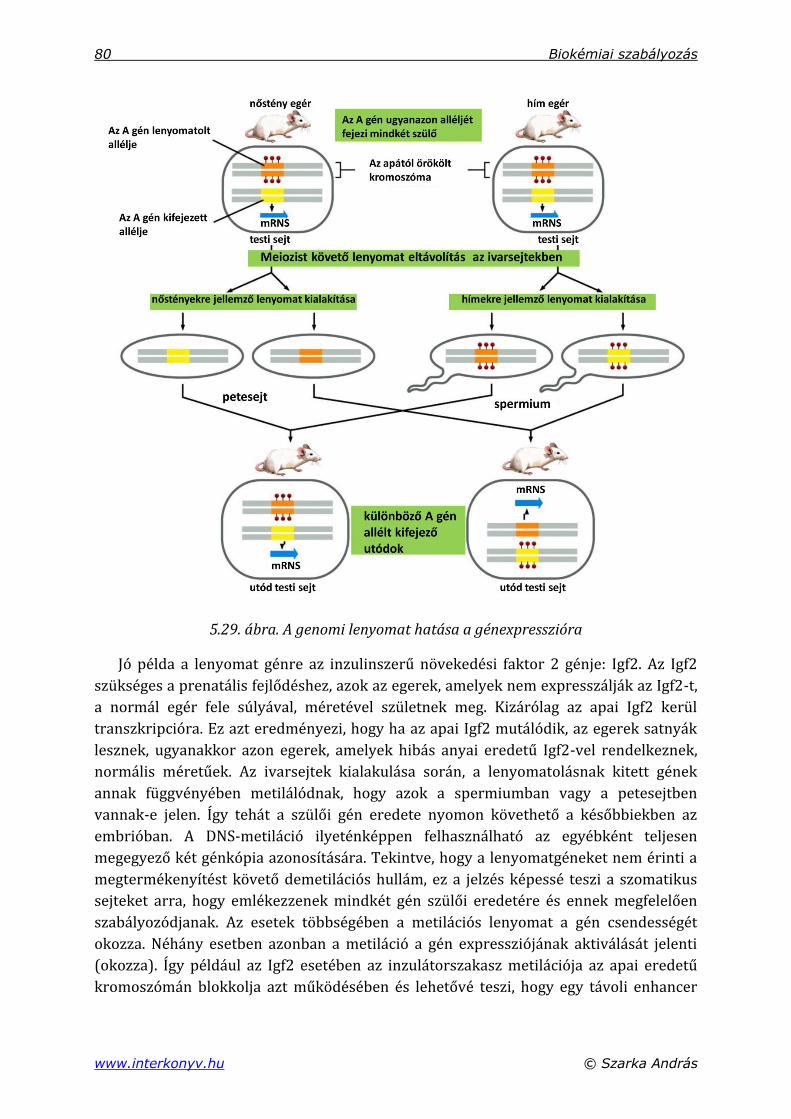
80 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
5.29. ábra. A genomi lenyomat hatása a génexpresszióra
Jó példa a lenyomat génre az inzulinszerű növekedési faktor 2 génje: Igf2. Az Igf2
szükséges a prenatális fejlődéshez, azok az egerek, amelyek nem expresszálják az Igf2-t,
a normál egér fele súlyával, méretével születnek meg. Kizárólag az apai Igf2 kerül
transzkripcióra. Ez azt eredményezi, hogy ha az apai Igf2 mutálódik, az egerek satnyák
lesznek, ugyanakkor azon egerek, amelyek hibás anyai eredetű Igf2-vel rendelkeznek,
normális méretűek. Az ivarsejtek kialakulása során, a lenyomatolásnak kitett gének
annak függvényében metilálódnak, hogy azok a spermiumban vagy a petesejtben
vannak-e jelen. Így tehát a szülői gén eredete nyomon követhető a későbbiekben az
embrióban. A DNS-metiláció ilyeténképpen felhasználható az egyébként teljesen
megegyező két génkópia azonosítására. Tekintve, hogy a lenyomatgéneket nem érinti a
megtermékenyítést követő demetilációs hullám, ez a jelzés képessé teszi a szomatikus
sejteket arra, hogy emlékezzenek mindkét gén szülői eredetére és ennek megfelelően
szabályozódjanak. Az esetek többségében a metilációs lenyomat a gén csendességét
okozza. Néhány esetben azonban a metiláció a gén expressziójának aktiválását jelenti
(okozza). Így például az Igf2 esetében az inzulátorszakasz metilációja az apai eredetű
kromoszómán blokkolja azt működésében és lehetővé teszi, hogy egy távoli enhancer

5. A génexpresszió szabályozása 81
© Szarka András www.interkonyv.hu
elindítsa az Igf2 transzkripcióját. Az anyai eredetű kromoszómán az inzulátor nem
metilálódik és így az Igf2 nem íródik át (5.30. ábra).
5.30. ábra. Az Igf2-gén metilációs lenyomata
A lenyomat jelensége az epigenetikus változat egy példája, ami a fenotípus
örökölhető változata, ami nem a DNS nukleotid szekvenciájában megírt, kódolt változat
eredménye. Hogy miért létezik, teljességgel rejtély. Gerincesekben ez a méhlepényes
emlősök sajátja és az összes lenyomat gén a fötális fejlődésben játszik szerepet. A DNS-
nek tehát nemcsak a szekvenciája örökíthető.
5.5.2. CG-szigetek
A DNS-reparáló enzimek miatt a metilált citozin-részletek az evolúció folyamán szépen
lassan kivesznek a genomból. A nem metilált citozin esetleges dezaminálása uracilt
eredményez, ami normálisan nem fordul elő a DNS-ben és így a DNS-reparáló uracil
DNS-glikoziláz felismeri és kivágja, majd citozinnal pótolja. Igen ám, de a metilált citozin
esetleges dezaminációja timint eredményez, ami így ugyebár nem felismerhető, nem
megkülönböztethető az egyébként nem mutáns normális timintől. Habár erre is létezik
egy javító mechanizmus, hogy eltávolítsa ezen timineket, mégis sok ilyen dezaminált
mutáció nem kerül felismerésre, így ezek a metilált citozin-részletek szépen lassan
timinné alakulnak az evolúció alatt. Az evolúció alatt négy CG-párosból több mint három
elveszett ilyetén módon, a gerincesekben tátongó CG-űrt hagyva maga után. A
megmaradt CG-nukleotidpárosok meglehetősen egyenlőtlenül oszlanak el a genomban.

82 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
A genom 1000, 2000 bp-os részleteiben 10-20-szor gyakrabban találhatók, mint
átlagosan. Ezeket a részeket CG-szigeteknek nevezik. Ezen részletek egy-két fontos
kivételtől eltekintve nem metilált CG-részleteket tartalmaznak minden sejttípusban.
Gyakran helyezkednek el a house-keeping gének promóterei körül. A house-keeping
gének, azok a gének, amelyek kiemelt fontossággal bírnak (átírásuk elengedhetetlen) a
sejt életképességét tekintve, ezért majd minden sejt expresszálja őket (pl. rRNS-ek,
citoszkeletális fehérjék, glikolízis enzimeinek génjei stb). Továbbá néhány
szövetspecifikus gén (amelyek csak az adott sejttípusban expresszálódnak, de azokban
szinte állandó módon) környékén is fellelhetők.
A CG-szigetek eloszlását megérthetjük, hogyha figyelembe vesszük, hogy a CG
metilációval a DNS inaktiválása a cél. Gerincesekben a metil-citozin – timin mutáció csak
akkor örökíthető tovább, ha az még az ivari vonalon bekövetkezik. A legtöbb gerinces
ivarsejtben található gén metilált, tehát inaktív. A hosszú evolúciós idő alatt ezek a
metilált CG-részletek spontán dezaminálódtak és elvesztek. Azonban az aktív állapotban
tartott gének (és így nyilvánvalóan a fontos életfontosságú housekeeping gének) aktívak
maradnak és így nem is metilálódnak. Tehát ha mutáció történik, ez javítódik is egyből.
Így a modern időkre ezek mint CG-szigetek megőrződtek. Továbbá bármely olyan CG-
mutáció, amely egy gén funkcióját vagy regulációját elrontja, kiszelektálódik, így néhány
CG-sziget egyszerűen csak a normálisnál nagyobb sűrűségű kritikus CG-szekvencia szám
eredménye. Az emlősgenom nagyjából 20 000 CG-szigetet tartalmaz. Ezek a szigetek
nagyrészt a transzkripciós egység 5’ végét, tehát egyben a gén végét is jelölik. Ily módon
a CG-szigetek az emlősgenomban felhasználhatók a DNS-szekvencián belül a gének
azonosítására.

© Szarka András www.interkonyv.hu
6. Sejtkommunikáció
Az eukarióta sejtekkel megjelent a bonyolult több kompartimentumos felépítés is, ezzel
együtt pedig, az ezek közti, majd később a többsejtű élőlényekkel a sejtek közötti
kommunikáció igénye. Egyes vélemények szerint ezzel indokolható, hogy mintegy 2,5
milliárd évvel a prokarióták után jelentek csak meg az első többsejtű élőlények.
Szomszédos vagy távoli sejtek között megvalósuló kommunikáció igen jelentős
mértékben különböző szignálmolekulákon múlik, a kommunikáció egyéb igen fontos
elemei a sejt saját fehérjéi, melyek a szignálokra reagálnak.
Amint az extracelluláris szignál eléri a sejtet, a kommunikáció (a jelátvitel) a
következő szinteken folyik (6.1. ábra):
– sejtfelszíni receptorfehérjék: kötik az extracelluláris szignálokat, ligandjaikat és
elindítják a sejtbeli folyamatokat;
– intracelluláris szignálfehérjék: a szignál szortírozása a sejtben (pl.: kinázok,
foszfatázok, GTP-kötő fehérjék);
– célfehérjék: aktiválás, sejt viselkedésének megváltozása;
– génregulációs fehérjék, ioncsatornák, metabolikus utak egységei, citoszkeleton
részletei.
6.1. ábra. A celluláris jelátvitel szintjei

84 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
6.1. Az extracelluláris szignálmolekulák
Az állati sejtek szignálmolekulák százai segítségével kommunikál. Milyen anyagok tölt-
hetnek be ilyen feladatot? Az extracelluláris szignálmolekulák meglehetősen heterogén
csoportot alkotnak. Ismertek köztük: fehérjék, peptidek, aminosavak, nukleotidok,
szteroidok, retinoidok, zsírsavak, oldott gázok (pl.: NO, CO).
Ezek jelentős része a szignált kibocsátó sejtből exocitózissal az extracelluláris térbe
ürül. Azonban erre más lehetőségek is rendelkezésre állnak, mint például: diffúzió a
sejtmembránon keresztül; a szignálmolekula a szignált kibocsátó sejthez kapcsolódva a
felszínén marad kötve, de az extracelluláris térben található, arra néz. A szignál az őt
kibocsátó sejtből tehát exocitózissal, diffúzióval és sejtfelszíni kötődés révén kerülhet ki,
mutathatja be a célsejtnek.
A szignál természetétől függetlenül a célsejt egy fehérjemolekula, a receptor segít-
ségével képes a jeleket fogni és a további feldolgozást, eseményeket elindítani. A recep-
tor rendkívül specifikusan köti ligandját (a szignálmolekulát). Az extracelluláris szignál-
molekulák általában rendkívül alacsony koncentrációban (≤ 10-8 M) vannak jelen,
viszont az őket felismerő receptor rendkívül nagy affinitással köti őket (K ≥ 108 l/mol).
A jeleket fogó receptorokat elhelyezkedésüknek (illetve az általuk megkötött
szignálnak) megfelelően két nagy csoportra osztjuk.
Sejtfelszíni receptorok: az esetek nagy részében ezen transzmembrán fehérjék kötik
a szignálmolekulát és egy intracelluláris szignálkaszkádot indítanak el, ami megváltoz-
tatja a sejt viselkedését (6.2. ábra).
6.2. ábra. Sejtfelszíni receptorok
Magreceptorok: intracelluláris receptorok – a szignálnak a sejtbe kell jutnia, hogy
aktiválhassa őket. Ez esetben a kisméretű, hidrofób szignál átjut a membránon (6.3.
ábra).

6. Sejtkommunikáció 85
© Szarka András www.interkonyv.hu
6.3. ábra. Intracelluláris magreceptorok
6.1.1. A szignálmolekulák hatótávolsága
Az egyes szignálmolekulák kommunikációs távolsága igen eltérő lehet.
1. Egyes szignálmolekulák a szignált kibocsátó sejthez tapadva maradnak, így csak a
velük kapcsolatba lépő sejtekre hatnak (6.4. A ábra). Ilyen kapcsolatfüggő
szignalizáció valósul meg a fejlődés és az immunválasz során. Ez egy
meglehetősen ritka kommunikációs forma. A legtöbb esetben a szignálok
szekretálódnak.
2. Parakrin
A szekretált molekulák ebben az esetben, mint lokális mediátorok csak a köz-
vetlen környezetben található sejtekre fejtik ki hatásukat (6.4. B ábra). Az, hogy a
parakrin szignál csak a megfelelő célsejthez jusson el azt igényli, hogy ne
diffundáljon messze. A gyakorlatban ez úgy valósulhat meg, hogy a szomszédos
sejtek gyorsan felveszik, vagy az extracelluláris mátrix lehorgonyozza őket.
3. Idegsejtek, szinapszis
Egy nagyméretű többsejtes élőlénynek a rövidtávú kommunikáció nem elegendő
sejtjei irányításához. A periféria irányítására az idegsejtek a legmegfelelőbbek
(hosszú axonnyúlványok). Ha aktiválódnak (külső szignál, vagy másik idegsejt
által) egy elektromos impulzus fut rajtuk végig, majd ha eléri a végéüket, akkor
kémiai szignálokat, neurotranszmittereket bocsátanak ki. Ezek az anyagok egy
specifikus sejtkapcsolatba, a kémiai szinapszisba kerülnek, mely biztosítja, hogy
azok csak a posztszinaptikus célsejtet érjék (6.4.C ábra).
4. A másik nagy kiterjedésű irányítási lehetőség az endokrin szabályozás. A
szervezetet, mint egy egészet szabályozza. Az endokrinsejtek szignálmolekulái-
kat, a hormonokat a véráramba juttatják, amely az egész testbe eljuttatja azokat
(6.4.D ábra). Az endokrin szállító közeg a vér, a folyamat tehát diffúziólimitált
(viszonylag lassú). A szignálhormon felhígul a vérben, interstíciumban. Alacsony
koncentrációban is hatnia kell (<10-8 M)

86 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
6.4. ábra. A szignálmolekulák hatótávolsága
Ezzel szemben az idegi (szinaptikus) jeltovábbítás, az idegsejtek elektromos
impulzusa, gyorsabb, precízebb. Gyors (> 100 m/s) elektromos impulzusterjedés.
A szinapszisokra jellemző 100 nm távolságot msec-os nagyságrend alatt teszi
meg. A neurotranszmitter alig hígul, magas lokális koncentráció jellemzi (pl.: az
acetilkolin koncentráció 5x10-4 Ma neuromuszkuláris résben). Ennek következté-
ben viszonylag alacsony a neurotranszmitter receptor affinitása. Így a neuro-
transzmitter viszonylag könnyen (gyorsan) disszociálhat a receptorról, gyorsan
megszüntetve a kiváltott választ. Az idegsejtből történt kibocsátását követően
nem sokkal eltávolítható (enzimes degradáció) a szinapszisból. Visszatranszpor-
tálódik az idegsejtbe vagy a szomszédos glia sejtekbe. Mind időben, mind térben
precízebb, mint az endokrin szabályozás.
A kiváltott válasz reakcióideje attól is függ, hogy mi az a válasz (6.5. ábra):
1. már meglévő fehérjében bekövetkező változás, ez igen gyors, s, msec-os nagyság-
rendű;
2. génexpresszió szinten bekövetkező válasz, ez lassabb, több órás nagyságrendű.

6. Sejtkommunikáció 87
© Szarka András www.interkonyv.hu
6.5. ábra. Az extracelluláris jelre adott válasz természete, reakcióideje
6.2. Autokrin szabályozás
Az eddig tárgyalt esetekben egy sejt más sejteket befolyásolt, más volt a szignált
kibocsátó és a célsejt. Sejtek, amelyek ugyanazon típusú sejteknek küldenek szignálokat,
képesek saját maguknak is. Ezt a jelenséget autokrin szignalizációnak nevezzük. A sejt
szignálmolekulákat szekretál, amelyek saját receptorához képesek bekötődni. Erre jó
példa, ha a fejlődés során egy adott sejt elkötelezte magát egy bizonyos differenciálódás
útján, akkor az elkezdhet autokrin szignálokat szekretálni saját maga számára, az adott
fejlődési irányt megerősítendő. A folyamat azonos sejttípusú szomszédos sejtek által
még hatékonyabb, alkalmas arra, hogy azonos sejttípusú sejtek azonos fejlődési döntést
hozzanak. Így az autokrin szignalizáció feltehetően egy lehetséges mechanizmus a
molekuláris „csordaszellem” vagy „közösségi hatás” megvalósítására, amelyet a fejlődés
korai szakaszában tapasztalnak, amely során az azonos sejttípusú sejtek csoportja képes
a differenciálódást kiváltó szignálra reagálni, az egyedül lévő, izolált viszont nem.
6.3. Gap junction
Specializált sejt-sejt kapcsolatok a szomszédos sejtek között. Tulajdonképpen a szom-
szédos sejtek citoplazmái között létesült vízzel kitöltött csatornák. A csatornák megte-
remtik a lehetőséget, hogy kisméretű hírvivő molekulák (intracelluláris mediátorok),

88 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
mint a Ca2+ és a ciklikus AMP (cAMP), kicserélődjenek a két sejt között, azonban a
nagyobb méretű makromolekulák, mint a fehérjék, nukleinsavak számára határt képez-
zenek. Így a sejtek a réskapcsolatok révén direkt módon kommunikálhatnak egymással,
anélkül, hogy a köztük levő plazmamembrán megakadályozná őket ebben. Néhány
véglegesen differenciálódott sejt kivételével, mint a vázizom sejtek és a vérsejtek,
minden állati sejt kommunikál szomszédos sejtjeivel, ilyen réskapcsolaton keresztül.
A következőkben tekintsük át a receptorok különböző fajtáit egy-egy jellemző példa
bemutatásával.
6.4. Plazmamembrán-receptorok
Integráns membránfehérjék (általában glikoproteinek), egy vagy több transzmembrán
szakasszal, monomer vagy polimer szerkezettel. A plazmamembrán receptorokat a
receptor-fehérje szerkezete, illetve a jelátviteli mechanizmus alapján három különböző
csoportra osztjuk. A következőkben ezen három csoportot tárgyaljuk meg kicsit
részletesebben.
6.4.1. Receptor-ioncsatornák
A receptort alkotó fehérje alegységek több transzmembrán szakasszal rendelkeznek, a
membránban egy pórus, vagy csatorna köré rendeződnek. A ligand megkötésekor
térszerkezetük megváltozik és átjárhatóvá válnak. Ekkor a csatornákon keresztül
kationok, vagy anionok jutnak át, az extracelluláris jelek hatását ezek fogják közvetíteni
az intracelluláris szereplők felé. Általában neurotranszmitterek receptorai, neurotransz-
mitterek hatásait közvetítik. Ideg és ideg, ideg és célszerv (sejtjei) közötti kommunikáció
valósul meg a segítségükkel.
A receptor aktiválásának következményei attól függnek, hogy milyen ion számára
nyit kaput. Depolarizációt, azaz stimulációt, vagy ellenkezőleg: gátló hatást eredményez-
hetnek.
Például, ha
Na+ csatornát nyit → depolarizáció, stimuláció,
Cl- csatornát nyit → depolarizáció gátlása lesz az eredmény.
Mindkét folyamat rendkívül gyors, msec nagyságrendű, ellentétben például a G-
fehérje kapcsolt receptorokkal, ahol a reakcióidő néhány másodperces.
A receptor-ioncsatornák elsőként megtisztított és legjobban tanulmányozott példá-
nya az acetilkolin-nikotin receptora. Ezért ennek működésén keresztül mutatjuk be a
receptorcsaládot.
Két altípusa ismeretes: az izom, amely egy pentamer (α2-,β-,γ-,δ-alegységekkel),
illetve a neuronális altípus, amely kétféle alegységből áll (α és β). Tekintsük át az izom-
altípus működését (6.6. ábra). Az alegységek egy alapesetben nem átjárható csatorna

6. Sejtkommunikáció 89
© Szarka András www.interkonyv.hu
köré rendeződnek (6.6. A ábra). Mindegyik alegység négy transzmembránrégióval
rendelkezik (6.6. B ábra). 2 db acetilkolin-kötőhely található receptoronként, egy az α-
és γ- és egy az α- és δ-alegységek között (6.6. C ábra). Az agonisták és az antagonisták is
ide kötődnek.
Az acetilkolin bekötése megváltoztatja a kötőhely szerkezetét, ami az alegységek
elmozdulását okozza. A csatorna citoplazmatikus oldalán található „kapu” kinyílik. Az így
megnyílt átjáró egy 0,65 nm átmérőjű, kationokra permeábilis csatorna (6.6. ábra). A
nikotinreceptor nem szelektív kationcsatorna, a méretük alapján jutnak át a különböző
kationok az elektrokémiai gradiens irányába: Na+> K+> Ca2+.
A neuronális csatorna Ca2+-ionokra áteresztőbb, permeabilitása nagyobb.
6.6. ábra. Az acetilkolin-nikotin receptorának izom-altípusa, az alegységek (a), az alegységek transzmembránrégiói(b), illetve az acetilkolin-kötőhelyek felülnézetből(c)
6.4.2. Hét transzmembrán szegmenssel rendelkező, G-fehérje-kapcsolt receptorok
Számos hormon, neurotranszmitter ilyen receptorokon keresztül befolyásolja a sejt
működését, ezeken kívül ide tartoznak a látás és szaglás folyamatában részt vevő
receptorok is.
Adódhat a kérdés, hogy mely fehérjéket nevezzük G-fehérjéknek. A fehérjék egy
jelentős része GTP kötésére képes. A GTP-kötő fehérjék között találunk transzlációs
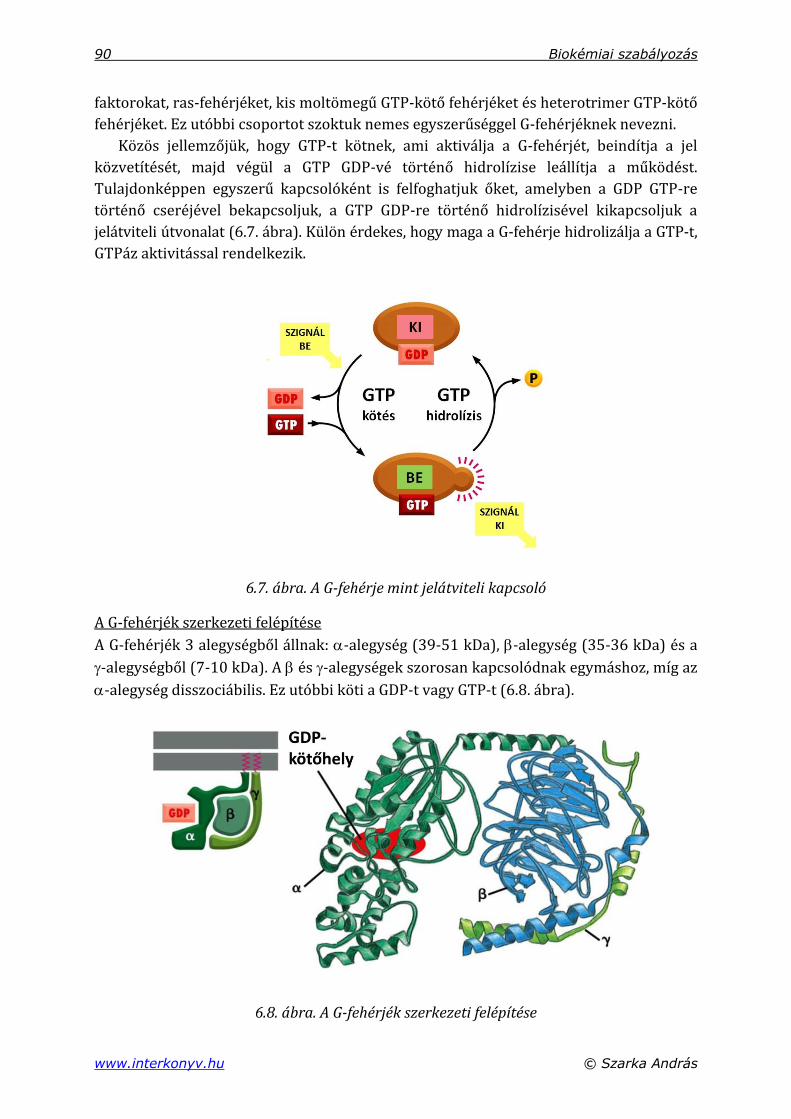
90 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
faktorokat, ras-fehérjéket, kis moltömegű GTP-kötő fehérjéket és heterotrimer GTP-kötő
fehérjéket. Ez utóbbi csoportot szoktuk nemes egyszerűséggel G-fehérjéknek nevezni.
Közös jellemzőjük, hogy GTP-t kötnek, ami aktiválja a G-fehérjét, beindítja a jel
közvetítését, majd végül a GTP GDP-vé történő hidrolízise leállítja a működést.
Tulajdonképpen egyszerű kapcsolóként is felfoghatjuk őket, amelyben a GDP GTP-re
történő cseréjével bekapcsoljuk, a GTP GDP-re történő hidrolízisével kikapcsoljuk a
jelátviteli útvonalat (6.7. ábra). Külön érdekes, hogy maga a G-fehérje hidrolizálja a GTP-t,
GTPáz aktivitással rendelkezik.
6.7. ábra. A G-fehérje mint jelátviteli kapcsoló
A G-fehérjék szerkezeti felépítése
A G-fehérjék 3 alegységből állnak: -alegység (39-51 kDa), -alegység (35-36 kDa) és a
-alegységből (7-10 kDa). A és -alegységek szorosan kapcsolódnak egymáshoz, míg az
-alegység disszociábilis. Ez utóbbi köti a GDP-t vagy GTP-t (6.8. ábra).
6.8. ábra. A G-fehérjék szerkezeti felépítése

6. Sejtkommunikáció 91
© Szarka András www.interkonyv.hu
Működési ciklusuk
Az extracelluláris agonista kötődik a hét transzmembrán szegmenssel rendelkező
receptorához, amelyhez intracellulárisan G-fehérje asszociál. A receptor-ligand
kapcsolat kialakulásakor megváltozik a komplex térszerkezete, amely ily módon kiterjed
az intracelluláris régióra is. Ennek következtében a G-fehérje -alegységéhez kötődött
GDP disszociálódik és GTP-re cserélődik. A már GTP-t kötő -alegység disszociál a -
alegységről és kölcsönhatásba lép az effektorával (ez például lehet az adenilát-cikláz, a
foszfolipáz C, sőt különböző ioncsatornák is). Az effektor működését módosítja, melynek
hatására a különböző intracelluláris mediátorok (pl cAMP, Ca2+) szintje megváltozik (6.9.
ábra). Az -alegység enzimaktivitása révén a GTP-t GDT-vé hidrolizálja, aminek hatására
az -alegység disszociál az effektorról és ismét a -alegységhez kötődik, kialakul az
inaktív GDP- komplex. Természetesen egy következő ciklus során újfent aktiválódhat
és az egész folyamat kezdődhet az elejéről.
6.9. ábra. A G-fehérjék működés közben
Csoportosításuk
A G-fehérjék csoportba osztása az -alegység szekvencia hasonlósága alapján történik.
Jelenleg 20 különböző -alegységet ismerünk, amelyek legalább 9 gén és alternatív
splicing eredményei. Ezeket 4 nagyobb családba soroljuk:
1. Gs-család: A legjobban ismert, számos hormon és neurotranszmitter, illetve a
szaglás receptorához kapcsolódik. Stimulálja (a család nevét is innen kapta) az
adenilát ciklázt, működése megnöveli a cAMP-szintet.
2. Gq-család: Szerkezete eltér a Gs-családétól, csaknem minden szövettípusban
megtalálható. A foszfolipáz C enzimet aktíválja.
3. Gi-család: Közös tulajdonságuk, hogy pertussis-toxin érzékenyek. Egyik alcso-
portjuk az adenilát cikláz gátlását (a név eredete: inhibeáló) okozza. Másik alcso-
portjuk ioncsatornákkal létesít kapcsolatot: Go. Ebbe a csoportba tartoznak a
látás folyamatában részt vevő Gt-fehérjék, amelyek a retinapálcikákban találha-
tók. Szintén fontos az a Gi-fehérje, amely a szívizomsejtekben közvetlenül
aktiválja a K+-csatornákat és hiperpolarizációt hoz létre.
4. G12-család: Szinte minden szövetben előfordulnak a család tagjai, funkciójuk
azonban többnyire még ismeretlen.

92 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
Az -alegységek több doménből épülnek fel. Az egyes doménokhoz különböző
funkció társítható:
˗ specifikus receptor felismerése
˗ guanin nukleotidkötés
˗ GTP hidrolízis
˗ -alegységek kötése
˗ effektor kapcsolat.
-alegységek
Szerepük kevésbé tisztázott, mint az -alegységé. Az -alegységet a membránhoz
rögzítik, egyes esetekben közvetlenül az effektorral is kapcsolatba lépnek.
Patobiokémiai vonatkozások
Egyes bakteriális toxinok az -alegységet adott helyen ADP-ribozilálhatják. A folyamat
során a NAD-ról ADP-ribóz egység kerül a toxin hatására az -alegységre, miközben a
nikotinsav rész felszabadul. A toxin hatása attól függ, hogy az -alegység mely
doménjére kerül az ADP-ribóz részlet, mely domén funkciója esik ki ezáltal.
A Vibrio cholerae toxinja a Gs-alegységét ADP ribozilálja a GTPáz-funkcióért felelős
doménen. Így a módosítás hatására kiesik a GTPáz-aktivitás, aminek következtében a G-
fehérje nem tud kikapcsolni. A folyton aktív G-fehérje pedig megállás nélkül aktív
állapotban tartja effektorát, az adenilát-ciklázt. A kontrollálatlan adenilát-cikláz aktivitás
következtében pedig jelentős mértékben megnő a cAMP-szintje és vele együtt minden
olyan folyamat kontroll nélkül felerősödik, amelyet a cAMP-szint regulál. Tekintve, hogy
a cholera-toxin a gyomor, bélcsatorna sejtjeivel kerül kapcsolatba, az adenilát-cikláz
állandó aktivitása is ezeket a sejteket érinti és alakít ki súlyos elektrolit- és
folyadékfelszívódási zavarokat.
A Bordatella pertussis toxinja a pertussis-toxin a Gi--alegységeket C-terminálisukhoz
közel, a receptor felismerésért, kötésért felelős részen ADP-ribozilálja. Így ADP-
riboziláció esetén a G-fehérje és a receptor nem asszociálnak. A toxin jelenlétében így
kiesnek az ezen fehérjék által közvetítette folyamatok.
G-fehérjék és jelerősítés
A G-fehérjék effektorai tehát igen gyakran enzimek, amelyek intracelluláris mediátorok
képződését katalizálják. Ez rendkívül jó lehetőséget ad a jel felerősítésére, hisz az
effektor enzim aktiválása több katalitikus ciklusban is fokozza az intracelluláris
mediátorok képződését. Ezen kívül fontos szem előtt tartanunk azt is, hogy egy receptor
többféle G-fehérjével is kölcsönhatásba léphet, amely szintén hozzájárulhat a jel
felerősítéséhez.

6. Sejtkommunikáció 93
© Szarka András www.interkonyv.hu
6.4.3. Protein-kinázok, foszfoprotein-foszfatázok
Az extracelluláris jelre cellulárisan igen gyakran adott válasz a reverzibilis fehérje-
foszforiláció. Milyen stimulus válthat ki reverzibilis fehérjefoszforilációt?
1. kiválthatja valamilyen másodlagos hírvivőmolekula, mint például a már
említett cAMP;
2. elképzelhető, hogy a receptor-ligand komplex közvetlenül (másodlagos
hírvivőmolekula képződése nélkül) fehérjefoszforilációval aktiválja a jelpályáját.
Ez utóbbi esetre jó példa az egyes növekedési faktor receptorok és a hozzájuk
kapcsolódó jelpályák.
Az eltérő extracelluláris jelek által aktivált jelpálya rendszerek között szoros
kölcsönhatás (crosstalk, párbeszéd) van, mely igen gyakran foszforilációs lépéseken
keresztül valósul meg.
A fehérjék foszforilációját végző kinázok és a defoszforilációt végző foszfoprotein-
foszfatázok részletesebb ismertetését korábban a 3.2. fejezetben már megtettük, jelen
esetben ennek ismétlésétől eltekintünk. A következőkben szeretnénk néhány, jelátviteli,
szabályozási szempontból fontos kinázt és a közreműködésükkel létrejövő jelpályát
bemutatni.
6.4.3.1. cAMP-dependens protein-kináz (protein-kináz-A)
A legrégebben és a legalaposabban ismert, tanulmányozott fehérje-kináz. Szerkezetét
tekintve inaktív formában tetramer. Két egyforma regulációs és két egyforma katalitikus
részből, alegységből áll (6.10. ábra). A regulációs rész a katalitikus alegységgel asszociált
formában gátolja annak működését.
6.10. ábra. A cAMP-dependens protein-kináz felépítése
A regulációs alegységen, alegységenként két-két cAMP-kötőhely található (regulációs
alegység dimerenként 2*2). A cAMP-szint növekedésével bekötnek a regulációs alegység
cAMP-kötőhelyére, melynek térszerkezete ennek hatására megváltozik (allosztérikus
reguláció) és a regulációs alegység dimer szabadon engedi a katalitikus alegység
monomereket, melyek így aktív állapotba kerülnek (6.11. ábra). Természetesen a cAMP-
szint csökkenésével azok ledisszociálnak a regulátor-alegységről, majd begyűjtenek két
katalitikus alegységet, így inaktív állapotba hozva őket (6.11. ábra).

94 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
6.11. ábra. A cAMP-dependens protein-kináz regulációja
Kétféle regulátor-alegység izotípust írtak le ezidáig: R’ és R”. Az R” targettáló
szereppel is rendelkezik, ugyanis képes az inaktív tetramert szubsztrátjának közelében
lehorgonyozni, így annak foszforilációja meggyorsítható a kináz aktiválódásakor.
6.4.3.2. A cGMP-dependens protein-kináz (protein-kináz-G)
Szerkezete nagymértékben hasonló a cAMP-dependens protein-kinázéhoz. Ez a fehérje
azonban két azonos polipeptid láncból áll, a regulátor és a katalitikus rész nem alkot
külön fehérjét, egyazon polipeptid lánc két doménjeként helyezkednek el. Ebben az
esetben a cGMP-regulátor doménhez történő kötése nem okozza az alegység disszo-
ciációját, hanem a szerkezet megváltozásához vezet, aminek a hatására a katalitikus rész
felszabadul a gátlás alól.
6.4.3.3. Protein kináz-C
Tulajdonképpen ebben az esetben nem egy adott enzimről van szó, hanem egy egymásra
nagymértékben hasonlító enzimcsaládról, amelynek jelenleg 12 tagja ismert. A család
klasszikus tagjai Ca2+/foszfolipid-dependens protein kinázok (, ’, ” és protein
kináz). A család többi atípusos tagja Ca2+-independens. Az izoenzimek szöveti ex-
pressziója igen eltérő. Egyetlen több doménből álló fehérjeláncból állnak. Az N-termi-
nálison helyezkedik el a regulációs domén, a C-terminálison pedig a katalitikus domén
(6.12. ábra). A regulátor doménhez kötődő allosztérikus ligandok együttes hatására
szabadul fel a katalitikus domén a gátlás alól. Az aktiváláshoz Ca2+-ra és foszfatidil-
szerinre van szükség. Fontos megjegyeznünk, hogy az enzim Ca2+ iránti affinitását nagy-

6. Sejtkommunikáció 95
© Szarka András www.interkonyv.hu
mértékben növeli a diacil-glicerol. Mind a protein kináz A, mind a protein kináz C széles
szubsztrát specificitású protein kinázok. Specificitásdeterminánsukban bázikus amino-
savak jelenléte szerepel a foszforilálandó szerin/treonin-csoportok közelében.
6.12. ábra. A protein kináz C felépítése és aktiválódása
6.4.4. Transzmembrán protein kinázok: növekedési faktor-receptor
Tulajdonképpen a protein kinázokkal megszakítottuk az eddig (remélhetően) logikus
fejezet felépítést. Hisz megtárgyaltuk a plazmamambrán receptorok két nagy családját, a
plazmamembrán ioncsatornákat és a 7 transzmembrán szegmenssel rendelkező G-
fehérje asszociált receptorokat. Ezen pontok után következett volna a harmadik nagy
csoport tárgyalása, amely nem más, mint a transzmembrán protein kinázoké. A tárgyalás
menetének megszakítása indokolt volt és remélhetően logikus is. Gondoljunk csak bele a
korábban tárgyalt ioncsatorna-receptorok jelentős része például a Ca2+-ionok (mint
másodlagos hírvivő) szintjének emelkedésén keresztül, vagy a hét transzmembrán
szegmenssel rendelkező receptorok a cAMP (mint másodlagos hírvivő) szintjének
változtatásán keresztül fejtik ki a hatásukat. Ez a hatás pedig mindkét esetben protein
kinázok aktivitásának befolyásolása (Protein kináz C a Ca2+ esetében és cAMP-függő
protein kináz a cAMP esetében). Ezért logikusnak tartottuk a receptorokat követő
folyamatokat rögtön a receptorok tárgyalása után ismertetni. Ez a tárgyalási mód
egyben megteremtette a lehetőséget, hogy ily módon eljussunk a receptorok harmadik
nagy csoportjához a transzmembrán protein-tirozin-kinázokhoz.
Esetükben egyetlen transzmembrán domén található. A fehérje extracelluláris
részén, az N-terminálison helyezkedik el az extracelluláris jel (pl.: növekedési hormon)
megkötéséért felelős receptor domén (6.13. ábra). Az intracelluláris részen, a fehérje C-
terminálisán pedig a kináz domén található (6.13. ábra). Szintén ezen a részen

96 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
találhatóak azok a tirozin oldalláncok, amelyeket az enzimfehérje saját magán fosz-
forilálni képes (autofoszforiláció). Az extracellulárisan kötődő növekedési faktor hatá-
sára intracellulárisan foszforilálódó tirozin oldalláncok indítják el a jelpálya működését.
6.13. ábra. Transzmembrán tirozin-kináz (receptor)
Léteznek nem receptor protein-tirozin-kinázok is. Ezen fehérjék nem a plazma-
membrán integráns alkotói, a citoplazmában, illetve a sejtmembránhoz kihorgonyozva
találhatóak. Az extracelluláris jelre (pl.: növekedési faktor, citokinek) a jelet felfogó
receptor intracelluláris részéhez képesek kötni, majd aktiválódva foszforilációs hatá-
sukat kifejtve elindítani (tovább adni) a jelátvitelt.
6.4.5. Foszfoprotein-foszfatázok
A jel lecsengése a fehérjék defoszforilációja révén történik meg, melyért a foszfoprotein-
foszfatázok felelősek. Ahogy korábban említettük, csoportosításuk a kinázokéhoz
hasonló, két nagy csoportba a szerin-/treonin- és a tirozin-oldalláncokról hasító
foszfatázokról beszélhetünk. Ismert néhány enzim, amely mind szerin-/treonin-, mind
tirozin-oldalláncokról képes a foszfátcsoport eltávolítására.
A fehérjék foszforiláltsági állapota mindig (legalább) kétkomponensű folyamat
eredménye, az illetékes protein kinázok és a hidrolízist végző foszfoprotein-foszfatázok
pillanatnyi aktivitásától függ.
6.5. A cAMP mediátorrendszer
A cAMP-mediátorrendszer működése egyértelműen a hét transzmembrán régióval
rendelkező receptorok működéséhez köthető (6.14. ábra). Ezek közül is két csoport vesz
részt a mediátorrendszer működésében:

6. Sejtkommunikáció 97
© Szarka András www.interkonyv.hu
1. Azok a receptorok, amelyek heterotrimer Gs-fehérjékkel állnak kapcsolatban.
Ezen ligand-receptor komplexek aktiválják az adenilát-ciklázt és az intracelluláris
cAMP-szint emelkedését idézik elő.
2. Azon receptorok, amelyek Gi-fehérjékkel állnak kapcsolatban. Ezen ligand-
receptor komplexek gátolják az adenilát-ciklázt és az intracelluláris cAMP-szint
csökkenését idézik elő.
Az adenilát-cikláz enzim az ATP és helyzetű foszfátját pirofoszfátként hasítja le a
cAMP képződésekor (6.14. ábra). A folyamatot a pirofoszfát anorganikus foszfátra
történő hidrolízise teszi irreverzibilissé. Az intracelluláris cAMP szint a cAMP-t előállító,
adenilát-cikláz és a cAMP-t hasító ciklikus nukleotid-foszfodiészteráz aktivitásának függ-
vénye. Az adenilát-cikláz átmeneti aktiválódását követően az aktiváló extracelluláris jel
csökkenésekor a nyugalmi cAMP-szintet a ciklikus nukleotid-foszfodiészterázok állítják
vissza.
A ciklikus nukleotid-foszfodiészterázoknak több típusa ismert. Nem minden sejt-
típusban fordul elő az enzim mindegyik típusa. A legáltalánosabban elterjedt az alacsony
Km értékű (Km ~ 10-6 M), illetve a magas Km értékű (Km ~ 10-4) enzimtípus.
A cAMP mediátorrendszer működését más jelpályák is befolyásolják (igen gyakran a
ciklikus nukleotid foszfodiészteráz-aktivitások befolyásolásán keresztül).
6.14. ábra. A cAMP-mediátorrendszer működés közben

98 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
A cAMP mediátorrendszer tulajdonképpen az eukarióta sejtek legegyszerűbb ismert
jelpályája, mivel minden hatásáért a cAMP-dependens protein kináz (protein kináz A) a
felelős. A cAMP felszabadítja a protein kináz A katalitikus alegységét a regulációs
alegység okozta gátlás alól. Az aktivált katalitikus alegység különböző fehérjéket
foszforilál, ezáltal aktíválja vagy inaktiválja azokat (6.11. ábra). Számos szubsztrát
fehérjéje ismert, köztük megtalálható metabolikus utak eznimei, de akár ioncsatorna
fehérjerészletek is. A mediátorrendszer így képes például a Ca2+ jelpálya működését is
befolyásolni.
A különböző extracelluláris jelek, különböző sejtekben ugyanazon jelpályákat hasz-
nálják, az extracelluláris jelre adott válasz mégis sejtspecifikus, mivel különbözőképpen
differenciálódott sejtekben a protein kináz A különböző, a sejt differenciáltságának
megfelelő fehérjéket képes foszforilálni. Tehát a sejt differencilódási módja szabja meg,
hogy milyen fehérjéket képes foszforilálni, ezáltal aktivitását befolyásolni a cAMP-
dependens protein kináz.
A cAMP-dependens protein kináz (PKA) a génszintű regulációban
A CRE, vagy cisz regulációs elem, egy rövidke DNS-szakasz (enhancer), amelyhez egy
cAMP-dependens protein kináz által foszforilált CREB nevű fehérje képes kötni. A CRE
egy rövid palindróm DNS-szekvencia, amely a cAMP által szabályozott gének
promóterén helyezkedik el. A CREB-dimer foszforilációja szükséges, hogy a CBP-nek
nevezett koaktivátor fehérje a CREB-hez kötődjék. A CBP segítségével alakul ki a
komplex fehérjeszerkezet, amely segít az RNS-polimeráz II bekötődésében, az iniciációs
komplex kialakulásában. A CBP hisztonacetiláló aktivitással is rendelkezik, ezáltal is
tovább segítve a transzkripció megindulását (6.15. ábra).
6.15. ábra. A cAMP-dependens protein kináz (PKA) szerepe a génszintű szabályozásban

6. Sejtkommunikáció 99
© Szarka András www.interkonyv.hu
6.6. Inozitol-foszfolipid jelátviteli rendszer
A receptorok egy jelentős részének aktiválása a sejtekben foszfatidilinozitol-származé-
kok hidrolízisét fokozza. Az intracelluláris mediátorok a foszfatidilinozitolokra speci-
fikus foszfolipáz-C hatására keletkeznek. Az enzim legfontosabb szubsztrátja a plazma-
membrán-lipid kettős rétegében elhelyezkedő foszfatidilinozitol-4,5-biszfoszfát (PIP2).
A foszfolipáz C hatására végbemenő hidrolízis eredménye a diacil-glicerol és az inozitol-
1,4,5-trizfoszfát. Ez a két vegyület a jelpálya mediátora. A sejten belül a jelátvitelt két
különböző irányba viszik tovább.
A foszfolipáz C kétféle receptor hatására aktiválódhat: 1. Hét transzmembrán szeg-
menssel rendelkező receptorok aktiválhatják, G-fehérjéken keresztül, illetve 2. tirozin
kináz aktivitású receptorok aktiválhatják. A két receptorcsalád eltérő foszfolipáz C izo-
enzimeket aktivál.
Foszfolipáz C
A foszfolipáz C aktivitásához Ca2+-t igényel. Háromféle izoformája van a , ,foszfolipáz
C, amelyek közül a és a jelátviteli szerepet tölt be, a szerepe egyelőre ismeretlen.
Izoformái közül a foszfolipáz-1 a legtöbb szövetben megtalálható, G-fehérjék
hatására aktiválódik. A Gq a-alegysége kölcsönhatásba lép az enzimmel és megnöveli
annak vmax értékét. A és a izoforma aktivitását a Gq a nem befolyásolja. Azon
hormonok, neurotranszmitterek és más szignálok, amelyek receptora a membránban Gq-
fehérjéhez kapcsolódik, foszfolipáz C aktiválást okoznak.
A foszfolipáz C -t a tirozin kináz receptorok aktiválják. A receptorral való kölcsön-
hatás során az enzim foszforilálódik. Az aktiválódással egyidejűleg az enzim a citoplaz-
mából a membránhoz, a szubsztrátjához (a plazmamembrán egyik alkotója: a foszfati-
dilinozitol-4,5-biszfoszfát) közel lokalizálódik.
Ahogy láttuk, az eltérő receptorok eltérő izoenzimeket aktiválnak, a végeredmény
azonban mindig ugyanaz: IP3 és diacil glicerol keletkezése foszfatidilinozitol-4,5-
biszfoszfátból (6.16. ábra).

100 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
6.16. ábra. A foszfolipáz-C aktiválása és működése
Az IP3 hatására megemelkedik a citoszol szabad Ca2+-koncentrációja. Az IP3
vízoldható molekula lévén eljut az intracelluláris Ca2+ raktárakhoz, ahonnan aztán Ca2+-t
szabadít fel. Ilyen intracelluláris Ca2+ raktár az endoplazmás retikulum, illetve
izomsejtekben a szarkoplazmás retikulum (6.17. ábra). Ezekben a Ca2+ raktárakban, a
Ca2+ különböző fehérjékhez kötve található. A módszer előnye, hogy nem csapódik ki a
magas (mM-os) koncentráció ellenére sem. A harántcsíkolt, a szív és a simaizmok
legfontosabb Ca2+-kötő fehérjéje a kalszekvesztrin (63 kDa), a legtöbb sejtben
megtalálható Ca2+-kötő, -raktározó fehérje pedig kalretikulin (46 kDa).
Ezen fehérjék nagy kapacitással (~ 50 Ca2+ kötőhely/molekula) és kis affinitással (Kd
~ 1 mM) kötik a Ca2+-t. Így valósulhat meg a Ca2+ raktárakból történő felszabadulása. A
felszabadulás 2 különböző ioncsatornán keresztül valósulhat meg:
1. Az endoplazmás retikulum membránjában található IP3-receptoron keresztül. A
folyamat a következőképpen alakul: az extracelluláris szignál dokkol a plazma-
membránban található receptorán (7 transzmembrán szegmenssel rendelkező
Gq-fehérjével asszociált, vagy tirozin kináz aktivitással rendelkező receptor), ezt
követően megtörténik a foszfolipáz C intracelluláris aktivációja, amely a plazma-
membránban található szubsztrátjának hasítása révén IP3 képződését eredmé-
nyezi. Az IP3 vízoldható molekula lévén eljut az endoplazmás retikulum memb-
ránban található receptorához, ahol dokkol. A receptor egyben egy Ca2+ csatorna
is. A receptor szerkezetét tekintve 4 alegységből áll, az IP3 dokkolása megváltoz-
tatja a tetramer térszerkezetét. A konformáció változás előidézi a Ca2+ mobili-
zációját.
2. Rianodin-receptor. Ez a receptor is az endoplazmás retikulum, illetve szív-,
harántcsíkolt- és simaizmok esetében a szarkoplazmás retikulum membránjában

6. Sejtkommunikáció 101
© Szarka András www.interkonyv.hu
található. Mind szerkezetét, mind funkcióját tekintve hasonlít az IP3-receptorhoz.
Nevét onnan kapta, hogy egy rianodin nevű növényi alkaloida is aktíválja. Haránt-
csíkolt izomban a szarkoplazmás retikulum rianodin receptorai a depolarizáció
hatására aktiválódnak, majd rajtuk keresztül Ca2+ kiáramlás történik, ami aztán
előidézi az izomkontrakciót. A szívizomban a feszültségfüggő plazmamembrán
depolarizáció következtében a feszültségfüggő Ca2+ csatornákon keresztül az
extracelluláris térből a sejtbe lépő Ca2+ váltja ki az intracelluláris Ca2+ raktárból
történő Ca2+ mobilizálást.
Az agonisták hatására bekövetkező intracelluláris Ca2+ felszabadulást az IP3 váltja ki
az IP3-receptorcsatornát aktiválva (6.17. ábra).
6.17. ábra. Az IP3-jelátviteli útvonal
Az IP3 két különböző úton metabolizálódik, vagy foszfatázok direkt módon inozitollá
alakítják, vagy első lépésben egy kináz IP4-gyé alakítja és ezt követően az előző esethez
hasonlatosan foszfatázok inozitollá alakítják. Összefoglalásul elmondhatjuk, hogy az IP3
közvetítette folyamatok jelentős része a Ca2+-szint változásán keresztül regulálódik.
A foszfatidilinozitol-4,5-biszfoszfát foszfolipáz C által történő hasításának (IP3
melletti) másik terméke a diacil-glicerol. A diacil-glicerol a sejtmembránban marad és
aktiválja a membrán közelében, de a citoplazmában található protein kináz C-t, ami
ennek hatására membránhoz kötődik. Az aktiválás átmeneti, mivela diacil-glicerol
metabolizmus gyors és a membránban foszfatidsavvá alakul, hogy aztán újra felhasz-
nálódhasson foszfatidil-inozitolok szintézisére.

102 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
6.7. A guanilát-cikláz és a cGMP
A cGMP és szerepe kevésbé ismert, mint a rokon vegyület cAMP. Feltehetően ezen
molekula is intracelluláris mediátor szerepét tölti be. A cGMP GTP-ből keletkezik
guanilát-cikláz hatására. Két különböző guanilát-cikláz enzim ismert jelenleg.
1. Az úgy nevezett partikuláris enzim a plazmamembránban található és egyúttal
receptorként is funkcionál. Jelenleg ismert kevés extracelluláris szignáljainak
egyike a pitvari nátriumürítő faktor, mely vazodilatációt, diurézist, nátriurézist és
az aldoszteron szintézis gátlását okozza.
2. A másik ismert enzim a szolubilis guanilát-cikláz, a citoplazmában található. Az
enzim két, egy 77 és egy 70 kDa-os, alegységből álló heterodimer. Fontos
megjegyeznünk, hogy hem prosztetikus csoportot tartalmaz. A NO és minden
olyan anyag, amelyből NO keletkezik, a szolubilis guanilát ciklázt aktiválja. A NO a
hem csoporttal lép reakcióba és nitrozohem képződik, mely hatására az enzim
aktiválódik, ennek hatására pedig megemelkedik a cGMP szintje.
6.7.1. Nitrogén-monoxid
A teljes kép alkotásához feltétlenül szót kell ejtenünk erről a gáz halmazállapotú
szignálmolekuláról. A NO a sejtekben in vivo keletkezik argininből nitrogén-oxid-szintáz
(NOS) hatására. Az enzim konstitutív és indukálható formában fordul elő. A NO a
keletkezés helyéről diffúzióval jut el a környező sejtekhez. Hatásának erőssége és
tartama attól függ, hogy milyen messze diffundál. Féléletideje általában 10-30 sec között
van, mivel a sejtekben és a keringésben gyorsan átalakul.
Az előzőekben ismertetett jelátviteli útvonalak működését egy példán keresztül
érthetjük meg jobban. A példa legyen az érfal simaizom relaxációja NO hatására. Az
endotél sejtek érlumen felöli felszínén különböző extracelluláris szignálok, mint például
acetilkolin, bradikinin, trombin, vagy adenin-nukleotidok dokkolhatnak receptorukon,
amelynek hatására megnő az intracelluláris Ca2+-koncentráció. Ez az emelkedett Ca2+-
koncentráció aktiválja a nitrogén-oxid-szintázt, aminek hatására az argininből NO-t állít
elő. A keletkezett NO átdiffundál a szomszédos simaizom sejtekbe és ott a szolubilis
guanilát-cikláz hem prosztetikus csoportját nitrozo-hemmé alakítja, aminek hatására az
aktiválódik és megemelkedik az intracelluláris cGMP szinte. A megemelkedő cGMP
aktiválja a cGMP-dependens protein kinázt, amely hatására csökken az intracelluláris
Ca2+-szint és ez utóbbi a simaizom sejtek relaxációját (vazodilatációt) eredményezi. Ezen
hatásmechanizmus mentén működik a szívkoszorús erek szűkületében szenvedők által
szedett érfal tágító nitroglicerin tabletta is.

© Szarka András www.interkonyv.hu
7. Összetett folyamatok szabályozása
A könyv vége felé járva az író megenged magának egy ugyancsak nagyképű hasonlatot.
Ahogy a nagy Jókai megjutalmazza a kitartó olvasót és a kevésbé izgalmas, cselekmény-
dús, de a történet megértése szempontjából nem mellékes, sőt megértéséhez szükséges
leíró részeket követően kibontakozik a valódi eseménysodor, úgy a biokémiai szabá-
lyozási folyamatokban szerepet játszó részegységek kevésbé izgalmas leírását követően
itt az ideje a komplex, rendkívül izgalmas szabályozási útvonalak ismertetésének, a
szabályozási eseménysodor főáramába kerülni.
7.1. Éhezés – jól tápláltság
Arra már korábban is utaltunk, hogy nemcsak a prokarióta élőlények számára fontos a
táplálék megszerzése, a kiegyensúlyozott anyagcsere. Vegyük példának saját magunkat
(rendkívül praktikus biológiai, biokémiai alanyok vagyunk, hisz mindig saját magunk
rendelkezésére állhatunk).
Mi történik, ha felkelést követően elfelejtünk reggelizni, rohanunk az egyetemre és
ott csak később az első órát követően jut időnk a Ch büfé nagyszerű választékából
valamilyen táplálékot magunkhoz venni?
A kérdés megválaszolásához vissza kell nyúlni alap biokémiai ismereteinkhez. A
felkelést követően éhezni fogunk, megvonjuk szervezetünktől az alapvető szén- és ener-
giaforrást, a glükózt. Ekkor tehát a feladat a következő: a glükózt felhasználó folyamatok
(legfőképp a glikolízis) lassítása (gátlása), a glükózt felépítő, eredményező folyamatok
gyorsítása (ezek közül elsősorban a glukoneogenezist és a glikogén mobilizációját érde-
mes megemlíteni). Első körben tehát a glikolízis és a glukoneogenezis, majd a glikogén
felépítés és lebontás szabályozásával érdemes foglalkoznunk a probléma megértéséhez.
7.1.1. A glikolízis – glukoneogenezis szabályozása
Ahogy ezt az előbbiek során megállapítottuk, a glükózt lebontó glikolízis és a glükózt
felépítő glukoneogenezis egymással szorosan összefüggő folyamatok. Tekintve, hogy a
két reakcióút egymás inverze, a szoros kapcsolat megjelenik a közös intermedierek és az
inverz reakciókat katalizáló enzimek révén. Ha belegondolunk, hogy egy glukózmolekula
glikolízis során történő lebontása mindössze 2 ATP molekulát eredményez és egy glukóz
molekula glukoneogenezis során történő felépítése 6 ATP molekulát igényel, be-
láthatjuk, hogy a szervezetünk, (máj)sejtjeink rendkívül logikusan működnek, amennyi-
ben elkerülik a párhuzamos glukóz-lebontó és - felépítő folyamatok működtetését. Így
nem meglepő, hogy az egyik folyamat gátlása egyszersmind a másik folyamat stimu-

104 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
lációját eredményezi. Ha csökken a glikolízis sebessége, akkor megnő a glukoneo-
genezisé és vica versa. Mindkét folyamat szabályozása az irreverzibilis lépéseket
katalizáló enzimek aktivitásának befolyásolásán keresztül történik. A glikolízis során
szabályozott enzimek: a hexokináz, a foszfofruktokináz I és a piruvát-kináz. A glukoneo-
genezisben pedig a piruvát-karboxiláz és a fruktóz-1,6-biszfoszfatáz. A folyamatok
szabályozása során három tényező kap fontos szerepet. Ezek közül csak egy lesz az
általunk kiválasztott éhezés – jól tápláltság, de a teljes kép érdekében a másik kettővel is
foglalkozunk.
1. Milyen az adott sejt energiaállapota? Mit is jelent ez pontosan? Tulajdonképpen
ez esetben a sejt ATP-, AMP- és citrát-szintje bír fontos szabályozó szereppel.
Logikus, hogy amennyiben a sejt energetikailag jól töltött, tehát az ATP-szint
magas, akkor ne bontson le több cukormolekulát újabb ATP molekula nyerése
érdekében. Az ATP, így nem meglepő módon allosztérikus gátlószere a glikolízis
legfontosabb szabályozópontját jelentő foszfofruktokináz I enzimnek (7.1. ábra).
Tekintve, hogy a sejtben az ATP szint jelentősen meghaladja az AMP-ét
(ATP>>ADP>>AMP) viszonylag kis relatív ATP szint csökkenés viszonylag nagy
relatív AMP szint emelkedést eredményez. Az AMP viszont allosztérikus
aktivátora a foszfrofruktokináz I enzimnek, ezáltal az AMP szint emelkedése
stimulálja a glikolízist, ezzel egyidejűleg gátolja a glukoneogenezist, mivel az AMP
allosztérikus inhibitora az inverz reakciót katalizáló fruktóz-1,6-biszfoszfatáznak
(7.1. ábra). Az AMP szint emelkedése (ATP szint csökkenése) így biztosítja a
reakciók glikolízis irányába történő eltolását.
A citrát szintén allosztérikusan gátolja a foszfofruktokináz I enzimet (7.1. ábra).
Ezáltal a zsírsavak, vagy ketontestek oxidációja során keletkező citrát glukózt
képes „megmenteni” az igen erős glükóz függésben élő agysejtek számára.
Ugyanakkor a zsírsavak oxidációja során keletkező acetil-KoA allosztérikus
aktivátora a glukoneogenezis első lépését katalizáló piruvát-karboxiláznak, így
serkenti a glukoneogenezist. A piruvát-kináz egyidejű gátlása révén viszont
visszaszorítja a glikolízist (7.1. ábra).

7. Összetett folyamatok szabályozása 105
© Szarka András www.interkonyv.hu
7.1. ábra. A foszfofruktokináz allosztérikus regulátorai
2. Az intracelluláris pH igen fontos szabályozó szereppel bír, ugyanis a H+-kon-
centráció emelkedése a foszfofruktokináz I gátlását okozza. Az anaerob glikolízis
így a fokozott laktát (H+) felszaporodás révén csökkenti a glikolízis sebességét.
3. Elértünk igazi célunkig, a glikolízis, glukoneogenezis hormonális szabályozásáig.
A szabályozás a biokémiai szabályozások egyik legszebb példája, ugyanis elemei
között szinte minden eddig ismertetésre került részlet megtalálható és mesteri
módon áll össze egy egésszé. A glikolízis és a glukoneogenezis hormonális sza-
bályozásában a glukagon és az inzulin játszik szerepet. Elérkezett az idő, hogy
visszatérjünk főhősünkhöz, a reggelijét nélkülöző egyetemistához. Tekintsük
végig, hogy mi történik a szervezetében! A vércukorszint csökkenés hatására a
hasnyálmirigy -sejtjeiben glukagon választódik el és kerül a vérkeringésbe. Az
extracelluláris szignálmolekula tehát ez esetben a glukagon lesz, amely hét
transzmembrán szegmenssel rendelkező receptorai megtalálhatók a májsejtek
plazmamembránjában. Az éhezés (vércukorszint csökkenés) hatására elválasz-
tásra került glukagon dokkol receptorain, amelyekhez intracellulárisan G-fehér-
jék asszociálnak. A ligandkötés hatására a Gs-alegység ledisszociál a alegysé-
gekről és effektorát, az adenilát ciklázt stimulálja. A stimuláció révén megnövek-
szik az intracelluláris mediátor, a cAMP szintje. A megemelkedett cAMP szint
hatására a cAMP-dependens protein kináz allosztérikusan aktiválódik és
foszforilálni fogja a májsejtekben kifejezésre került szubsztrátjait.

106 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
Az első – egyetemistánk szempontjából – említésre érdemes szubsztrátja a
foszfofruktokináz II enzim lesz. A foszfofruktokináz II egy érdekes kettős
funkciójú enzim, amely mind kinázként, mind foszfatázként képes működni. Az,
hogy kinázként, vagy foszfatázként működik, egyedül foszforiláltsági állapotán
múlik (7.2. ábra). Amennyiben az enzim foszforilált állapotban van, foszfatázként
működik és szubsztrátjából a fruktóz-2,6-biszfoszfátból fruktóz-6-foszfátot állít
elő (7.2. ábra). Amennyiben defoszforilált állapotban van, kinázként működik és
szubsztrátjából a fruktóz-6-foszfátból fruktóz-2,6-biszfoszfátot állít elő (7.2.
ábra). A foszfátcsoport okozta térszerkezet váltás tehát egyféle kapcsolóként
működik a foszfatáz és a kinázaktivitás között.
7.2. ábra. A glikolízis/glukogeogenezis hormonális szabályozása: a foszforuktokináz II foszforilációval/defoszforilációval történő szabályozása, a cAMP szint hatása a glikolízis,
glukoneogenezis szabályozására
Lássuk csak, miként kapcsolódik mindez egyetemistánkhoz. Az előzőekben ismer-
tetett mechanizmus révén éhezésben megnő az intracelluláris cAMP szint, ami aktiválja
a cAMP-dependens protein kinázt, amely foszforilálni fogja a foszfofruktokináz II
enzimet, amely ennek hatására foszfatázként fog működni és a fruktóz-2,6-biszfoszfátot
fruktóz-6-foszfáttá és szervetlen foszfáttá hasítja (7.2. ábra). Tehát jelentősen lecsökken
a sejt fruktóz-2,6-biszfoszfát szintje, ami viszont a glikolízis legfontosabb szabályozó-
pontjának, a foszfofruktokináz I enzimnek legfontosabb allosztérikus aktivátora (és az
inverz lépést katalizáló fruktóz-1,6-biszfoszfatáz allosztérikus inhibitora) (7.2. ábra). A
folyamat eredményeként tehát lényegesen lelassul a glikolízis sebessége és meggyorsul

7. Összetett folyamatok szabályozása 107
© Szarka András www.interkonyv.hu
a glukoneogenezisé. A folyamatot kiegészíti, hogy a cAMP dependens protein kináz a
glikolízis utolsó lépését katalizáló enzimet, a piruvát-kinázt is foszforilálja, ami ennek
hatására inaktíválódik.
Mindezen folyamatok hatására főhősünk májsejtjeiben lelassul a glikolízis és
megindul a glükóz de novo szintézise, a glukoneogenezis révén.
7.1.2. A glikogén felépítés – lebontás szabályozása
A glikolízis fékezése és a glukoneogenezis beindítása, stimulálása csak egy módja a
vércukorszint állandó szinten tartásának. Elsősorban a máj- és izomsejtek képesek a
glükózt polimerként, glikogén formájában raktározni. A glikogénben történő raktározás
nagy előnye, hogy igény esetén rendkívül gyorsan glükózként mobilizálható a benne
tárolt glükóz. Hátránya, hogy meglehetősen nagy a helyigénye, jóval nagyobb, mint
amikor a glükózból zsírsavak képződnek és a tárolás lipid formájában történik, viszont
ez utóbbi esetben a mobilizálás során képződő termékek nem glukoplasztikus anyagok,
általuk a vércukorszint nem tartható állandó szinten.
Lássuk csak, hogy egyetemistánk szervezetében miként épül fel a glikogén, miként
bomlik le és hogyan szabályozódik e két folyamat. Jól táplált állapotban, amikor bővében
vagyunk a glükóznak, a sejtbe lépő glükóz glükóz-6-foszfáttá alakul, amelyet a foszfo-
glukomutáz enzim glükóz-1-foszfáttá alakít. A keletkezett glükóz-1-foszfát UTP-vel
(uridil-trifoszfát) aktíválódik az UDP-glükóz-pirofoszforiláz enzim által katalizált reak-
cióban. Az UDP-glükóz képződése mellett pirofoszfát szabadul fel, amely szervetlen
foszfátra hasad, így a reakció során két nagyenergiájú kötés használódik fel, erősen exer-
gonikussá téve azt. A következő lépésben a glikogén-szintáz által katalizált reakcióban
az UDP-glukózról a glikogén láncvégi glukózához kapcsolódik az újabb glükóz molekula,
eggyel megnövelve a glikogénláncot alkotó glükóz molekulák számát. A glikogén-szintáz
egy legalább négy glükózból álló láncot képes csak tovább hosszabbítani. Az első glükóz
molekula egy glikogenin nevű fehérjetirozin oldalláncához kapcsolódik. A glikogén több
ezer kDa molekulatömegű hatalmas polimer számos elágazással. Az elágazásoknak igen
fontos szerepe van, mert általuk egyszerre több glükózszálon tud elindulni a polimer
lebontása, a glükóz molekulák visszanyerése, szükség esetén jelentősen meggyorsítva a
cukorhoz jutás sebességét. A glikogén-szintáz mellett így igen fontos szerepe van a
szintézis során az elágazásokat kialakító glikozil-4-6-transzferáz enzimnek. Amennyiben
egy legalább 11 glükóz egységből álló lánc elkészült, az enzim egy nagyjából 7 glükóz
egységből álló láncrészt áthelyez és 1-6 kötésekkel a glükóz lánchoz kapcsolja.
Az ellentétes folyamatot, a glikogén lebontását, a glikogén-foszforiláz katalizálja. A
reakció szervetlen foszfátot igényel, ugyanis a lehasadó glükóz molekula foszforilálódik
és glükóz-1-foszfátként szabadul fel. Az elágazásokat külön enzim az oligo-6-4-glukán-
transzferáz enzim bontja. A képződött glukóz-1-foszfátból glukóz-6-foszfát lesz, amely
vagy a glikolízisben alakul tovább (izomsejtek esetében), vagy a glükóz-6-foszfatáz által
katalizált reakcióban glükózzá alakul (májsejtek esetében) és a keringésbe kerülve segít
állandó szinten tartani a vércukorszintet.

108 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
Ennyi felvezetés után megint csak forduljunk kedvenc éhező egyetemistánk irányába
és nézzük meg, mi történik szervezetében a glikogénnel. A glikogénszintézis és-lebontás
központi enzimei a glikogén-szintáz és a glikogén-foszforiláz. Mindkét enzim aktivitása
foszforiláció/defoszforiláció révén szabályozott. Míg a glikogén-szintáz foszforilált
formában inaktív (defoszforilált formában aktív), addig a glikogén-foszforiláz foszforilált
formában aktív (és defoszforilált formában inaktív). Tehát a foszforiláció (igen ötletes és
logikus módon) ellentétesen befolyásolja a glikogén felépítésért és lebontásért felelős
kulcsenzimeket, biztosítva, hogy egyszerre glikogén felépítés és lebontás ne folyjon a
szervezetünkben (igen hasonlóan a glükóz lebontó és felépítő folyamatokhoz).
Az izom és máj glikogén funkciója eltérő, ennek megfelelően felépítésének és
lebontásának szabályozása is eltérő. Ahogy korábban említettük, az izomglikogén az
izomsejt energiaraktára, amely fokozott izomműködés esetén a megnövekedett energia-
szükségletet hivatott fedezni. Az 1930-as évek második felében Carl és Gerty Cori
felfedezték, hogy a vázizom sejtekben található glikogén-foszforiláz két egymásba
átalakuló formában létezik, a katalitikusan aktív glikogén-foszforiláz a és a kevésbé
aktív glikogén-foszforiláz b formában. Earl Sutherland ezt követő kísérletei
kimutatták, hogy a nyugalomban levő izomban a foszforiláz b dominál, azonban erőteljes
izomműködés során az epinefrin nevű hormon kiváltja a foszforiláz b specifikus szerin
oldalláncának foszforilációját, mely hatására az átalakul az aktív foszforiláz a formába. A
foszforiláz b foszforilálásáért (aktiválásáért) felelős enzim, a foszforiláz b kináz maga is
foszforiláció révén aktiválódik. A foszforiláz b kinázt aktiváló enzim pedig nem lesz más,
mint a már annyit emlegetett cAMP-dependens protein kináz. A cAMP-dependens
protein kináz – ahogy korábban azt már megtanultuk – pedig az emelkedő cAMP szint
hatására aktíválódik. A cAMP szint emelkedését izomsejtekben az epinefrin, máj-
sejtekben pedig a glukagon hét transzmembrán szegmenssel rendelkező receptorához
való kötődése váltja ki, amely aztán egy Gs fehérjén keresztül aktiválja az adenilát-
ciklázt (7.3. ábra).
Az izom- és májglikogén-foszforilázok izoenzimek, melyek különböző gének
termékei és egymástól eltérő módon regulálódnak. Az izomglikogén-foszforiláz kovalens
módosításon (foszforiláción) alapuló szabályozásán kívül még két allosztérikus
szabályozási mechanizmus is befolyásolja aktivitását. A Ca2+, amely az izomkontrakció
kiváltója, képes a foszforiláz b kinázhoz kötődni és aktiválni azt, ezáltal az foszforilálja és
az inaktív foszforiláz b formából az aktív foszforiláz a formába alakítja (7.3. ábra). A Ca2+
a foszforiláz b kináz alegységéhez kapcsolódik, amely nem más, mint a kalmodulin. A
heves izommunka során az ATP lebontásból nagy mennyiségben képződő AMP képes a
foszforilázhoz kötődni, allosztérikus regulátor kötőhelyén keresztül és igen gyorsan
(allosztérikus reguláció) átalakítani azt a jóval aktívabb formájába (7.3. ábra). Megfelelő
ATP szint esetén az ATP képes bekötődni a foszforiláz fehérjére, megakadályozva az
AMP kötődését.
Amikor az izomsejt visszatér nyugalmi helyzetébe, a foszforiláz a foszfatáz, vagy más
néven a foszfoprotein-foszfatáz 1 eltávolítja a foszfatáz a foszfátcsoportját visszaalakítva
a kevésbé aktív foszforiláz b-vé.

7. Összetett folyamatok szabályozása 109
© Szarka András www.interkonyv.hu
7.3. ábra. A glikogénlebontás (felépítés) szabályozása izom- és májsejtekben
Az izom izoformához hasonlatosan a májglikogén-foszforiláz is kettős hormonális és
allosztérikus szabályozás alatt áll. A defoszforilált forma az inaktív ez esetben is. Éhező
egyetemista főhősünk esetében tapasztalható hasonló alacsony vércukorszint esetén a
glukagon (az epinefrinhez hasonlatosan) megemeli a cAMP szintet és az előbb
ismertetett foszforilációs kaszkádon keresztül aktiválja a glikogén-foszforilázt (7.3.
ábra). Amikor a (a Ch büfé áldásos szolgáltatását kihasználva egyetemistánk) vércukor-
értéke újra normalizálódik, a cukor bejut a májsejtekbe és ott a foszforiláz az
allosztérikus inhibíciós helyéhez kapcsolódik, aminek hatására az térszerkezetet vált és
a szerin oldalláncon foszforilált foszforiláz a a foszfatáz szubsztrátja lesz, amely
eredményeként az eltávolítja a foszfátcsoportot és ezáltal inaktiválja azt. A glikogén-
foszforiláz allosztérikus glükóz kötőhelye révén egyféle glükóz szenzormolekulaként
működik, a vércukorszintnek megfelelően szabályozza aktivitását.

110 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
A glikogén-szintáz regulációja ezzel pont ellentétes. Az aktív enzim a defoszforilált
állapotú glikogén-szintáz a. A cAMP és kalmodulin-dependens protein kinázok hatására
foszforilálódik és a kevésbé aktív glikogén-szintáz b-vé alakul. Aktív állapotba (a
glikogén-foszforilázt is defoszforiláló, ezáltal inaktíváló) foszfoprotein-foszfatáz 1 által
katalizált defoszforáláció által kerül. A foszfoprotein-foszfatáz aktvitását a glikogén-
foszforiláz a gátolja. A foszforiláz a foszforiláz b-vé történő átalakulással a gátló hatás is
kiesik, így a glukózszint növekedésével nem csak a glikogénlebontás csökken, de megnő
a szintézis jelentősége is. A glikogén-szintáz b defoszforilációját a vércukorszint
emelkedéssel megemelkedő inzulinszint serkenti. Ez a szabályozás tehát biztosítja, hogy
egyszerre ne történjen teles intenzitású glikogénszintézis és lebontás.
Most már mindannyian belegondolhatunk, hogy mi is történik éhező, majd jóllakott,
fizikai aktivitást végző, majd pihenő egyetemista főhősünk izom- és májsejtjeiben.
7.2. Programozott sejthalál: Apoptózis
Talán első alkalommal a fejezet címére pillantva mindenkit rossz érzés kerít hatalmába,
de megígérjük, ez nem lesz tartós. Kezdjük is a jobb kedvre derítést a programozott
sejthalál nélkülözhetetlen, élethez szükséges funkcióinak áttekintésével:
1. A többsejtű élőlényekre mint egy jól szervezett közösségre gondolhatunk. Ebben
a jól szervezett közösségben a sejtek száma szigorúan szabályozott. Amennyi-
ben egy sejtre már nincs többé szükség, az öngyilkosságot követ el, aktiválva egy
intracelluláris halálprogramot. Éppen ezért nevezik ezt a folyamatot progra-
mozott sejthalálnak, apoptózisnak, amely egy görög szó és annyit tesz, hogy
„lehull”, utalva a falevelek lehullására, amely azért is találó, mert a leveleket tartó
száracska sejtjei is apoptózissal halnak el minden ősszel a falevelek lehullásakor.
Tehát a celluláris turnover, amely szükséges a folytonos megújuláshoz, az
osztódás/sejthalál egyensúlyának megtartásához nélkülözhetetlen folyamat. Erre
jó példát szolgáltathat a májsejtek számának szabályozása: amennyiben (külö-
nösen, ha induktív hatású) gyógyszert kezdünk el szedni, a májsejtek száma
megnő, alkalmazkodva a fokozott biotranszformációs igényhez, azonban ha a
gyógyszer szedését befejezzük, a májsejtek számának vissza kell térnie a kiindu-
lás körüli szintre. Ez utóbbi folyamatot programozott sejthalál (apoptózis) révén
oldja meg szervezetünk.
2. Igen fontos a nem megfelelően differenciálódott, vagy feleslegben lévő sejtek
eltávolításához is. A szervezetében meglepően nagy méretet is ölthet az apotózis
révén eltávolításra kerülő sejtek száma. Egy egészséges felnőtt szervezetében
sejtek milliárdjai pusztulnak el a csontvelőben és a bélrendszerben minden egyes
órában.
3. Nélkülözhetetlen szerepet tölt be az egyedfejlődés során is. Tulajdonképpen a
szervezetünk, mint egy „szobrász”, az apoptózist egyféle molekuláris „vésőként”
használja. Erre igen jó példa az egérmancs kialakulása, ahol az eredetileg

7. Összetett folyamatok szabályozása 111
© Szarka András www.interkonyv.hu
úszóhártya-szerű képződményeket, amelyek összekötik az egérmancs ujjacskáit,
apoptózissal „faragja” le a szervezet. Szintén az apoptotikus „véső” áll az ebihal-
béka átalakulás, az ebihalra jellemző képletek eltűnésének hátterében. A fejlődő
szervezet idegrendszerében az idegsejtek több mint a fele nem sokkal megszü-
letésüket követően elhalálozik.
4. Számos esetben a sejtek funkciójukat csak apoptózisukat követően tudják
betölteni. Ilyen funkcióhoz kötött sejtelhalás történik például a szemlencse sejtek
esetében a látás folyamatában, vagy a bőr külső felszínét alkotó sejtmag és
organellum vesztett sejtek esetében, amelyek a mechanikai védelemért felelősek.
5. Enyhébb sejtkárosító tényezők is kiválthatják. Ez esetben a későbbi, súlyosabb
következmények elkerülése végett kerül sor az apotózisra, például vírus-
fertőzött vagy malignusan transzformált sejtek esetében.
A sejthalál két fő formája, a szép és a csúnya halál: apoptózis vs nekrózis
Az apoptózis szó jelentése, bár utal az elmúlásra, azt könnyedén, kíméletesen teszi, hisz
a falevelek lehullása az élet természetes és nem egyértelműen szomorú velejárója.
Apoptózis során egy program szerint, különböző stációk során szépen lebontja, majd
összecsomagolja magát a halálra elszánt sejt, anélkül, hogy a többi, körülötte levő
sejtnek bármiféle baja esnék. Nézzük végig, milyen stációkon megy keresztül apoptózisa
során a sejt.
A sejt elkezd zsugorodni, ehhez nagymértékben hozzájárul a citoszkeleton kollapszu-
sa, a citoszkeletális fehérjék lebontása. A felszíne felhólyagosodik (blebbing), a sejtmag
kisebbedik, a kromatinállománya kondenzálódik, gyakorlatilag a nukleázok felszeletelik
a DNS állományt. A sejtmaghártya redőződik, majd feltöredezik. A nukleáris lamin is
hidrolízisre kerül. Nagyon fontos megjegyeznünk, hogy mindezen folyamatok közben a
sejtmembrán végig sértetlen és az organellum membránok is épen maradnak. Végül a
sejt sejtmembránnal határolt apoptotikus testekre esik szét. ezeket az apoptotikus
testeket makrofágok és a szomszédos sejtek veszik fel, kebelezik be (7.4. ábra).
A sejtmembrán végig ép marad, makromolekulák nem jutnak ki az interstíciális
térbe, így gyulladásos reakció sem indul be, a környező sejteket semmilyen inzultus sem
éri az apoptotizáló sejt részéről. A környező sejtek épen maradnak, az apoptózis
tipikusan egy sejtre terjed ki, egy sejtet, különálló sejteket érintő folyamat (ez termé-
szetesen nem zárja ki, hogy tömeges legyen, de a sejtek egyenként esnek át rajta).
Az apoptózis tehát egy aktív, energiaigényes (a program végrehajtása, a sejtszervezet
„szétszerelése” energiát igényel), gyorsan (néhány óra alatt), csendesen (gyulladás-
mentesen) lezajló folyamat. Nevezhetjük szép halálnak.
Ezzel szemben a nekrózis a csúnya halál. Ezt már előrevetíti nevének eredete is,
hisz a görög nekrosz szóból ered, ami halott, megöl, pusztulás, megölés jelentéssel bír. A
nekrózis mindig patológiás folyamat (ellentétben a számos fiziológiás példát
felvonultató apoptózissal szemben). Általában erőteljesebb behatások váltják ki az
igen erőteljes, tömeges sejtpusztulást. Tipikusan nekrotikus sejthalállal halnak el
sejtjeink, ha mondjuk egy jó nagyot rásózunk a kézfejünkre kalapáccsal, vagy ha tömény

112 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
lúgba mártjuk kezünket, más testrészünket, de nekrotikus sejthalált halnak szív- vagy
agysejtjeink is egy esetleges isémia kapcsán.
Az előző példával ellentétben a nekrózist a sejt, a sejtmag, az organellumok
duzzadása, a lizoszómák felnyílása és az egész sejt kontrollálatlan szétesése jellemzi.
Ennek következtében a sejt tartalma kiáramlik az interstíciális térbe és ott gyulladásos
reakciókat vált ki, amely a többi környező sejtet is károsítja (7.4. ábra).
7.4. ábra. A nekrótikus és az apoptotikus sejthalál közötti morfológiai és folyamatbeli különbségek
Már igen korán megfigyelték, hogy a különböző sejttípusok igen sztereotip
morfológiai változásokon (melyek hátterében a korábban ismertetett folyamatok állnak)
esnek át apoptózisuk során. Mindez arra utalt, hogy konzervatív biokémiai változások és
apparátus áll a háttérben.

7. Összetett folyamatok szabályozása 113
© Szarka András www.interkonyv.hu
A morfológiai és sejtbiológiai történésekben főszerepet játszik egy proteáz család,
amelyek aktív helyükben cisztein tartalmaznak és szubsztrát fehérjéiket specifikus
helyen előforduló aszpartát oldalláncaik mellett hasítják. Elnevezésük is erre a két
dologra utal: kaszpázok (cysteinyl aspartate specific protease: caspase). A kaszpázok,
előalakként prokaszpáz formában termelődnek és tulajdonképpen minden sejtben
megtalálhatóak, magukban hordozva a sejt öngyilkos gépezetét. Aktiválódásuk limitált
proteolízissel történik (lásd 3.3 fejezet), amely folyamat során egy kis és egy nagy
alegységre hasadnak, létrehozva a két aktív centrummal rendelkező a2b2 hetero-
tetramert. Tekintve, hogy a kaszpázok több lépcsőben egymás hasítására is képesek, ez a
jel kaszkád-szerű erősítésével jár együtt (7.5. ábra). Emlős sejtekben ezidáig 14
különböző kaszpázt írtak le, melyek közül emberben 11 fordul elő (kaszpáz 1-10 és 14).
A kaszpázokat funkciójuk alapján két nagy csoportba sorolhatjuk:
1. Citokin aktivátor gyulladásos kaszpázok. Ebbe a csoportba tartozik a kaszpáz
1, 4, 5, 11.
2. Sejthalál-apoptotikus kaszpázok. Ebbe a csoportba tartozik a kaszpáz 2, 3, 6, 7,
8, 9, 10, 12.
A sejthalál-apoptotikuskaszpázokat N-terminális prodoménje és a programozott
sejthalál folyamatában betöltött szerepe alapján szintén két alcsoportra oszthatjuk:
1. Effektorkaszpázok: Ezen kaszpázok hasítják a celluláris célmolekulák jelentős
részét, rövid N-terminális doménnel rendelkeznek. Ide tartozik a kaszpáz 3, 6, 7.
2. Iniciátorkaszpázok. Ezen kaszpázok aktiválják az effektor kaszpázokat, hosszú
N-terminális doménnel rendelkeznek. Ez az úgy nevezett homofil interakciós
domén, amely révén adaptorfehérjék hasonló doménjével képes kapcsolatba
lépni. Ezen fehérje-fehérje kapcsolatok révén jön létre az apoptoszóma. Az
iniciátor kaszpázok ezekben a komplexekben aktiválódnak konformáció
változásuk következtében.
Az effektor kaszpázokat, nemcsak az iniciátor kaszpázok, hanem más proteázok is
képesek aktiválni, mint például a katepszinek, kalpainok, granzimok. Az aktivált kasz-
pázok jelenlegi ismereteink szerint ezt követően mintegy 300 ismert magi, citoszolban
található fehérjét hasítanak. Az apoptotikus morfológiai változások hátterében ezen
proteolítikus események állnak. Így például a kaszpáz aktiválta DNáz (CAD) felelős a
DNS fragmentációért. A sejtalak megváltozásának hátterében pedig az aktin, spektrin,
citokeratinek, a vimentin, fodrin hasítása áll. A nukleáris laminok hidrolízise pedig a
sejtmag zsugorodásához vezet.
Természetesen a kaszpáz aktivitás szigorú kontroll alatt áll, a kaszpázokat különböző
fehérjék, mechanizmusok tartják féken. Ilyen apoptózis-inhibitorfehérjék az IAP
(inhibitor of apoptozis protein) család tagjai: XIAP, c-IAP1, c-IAP2, survivin, livin és a
többiek. A család tagjai kaszpáz 3, 7, 9 gátló hatással bírnak. Minden egyes családtag

114 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
rendelkezik egy BIR doménnel, amely révén kapcsolódnak a kaszpázokhoz és gátolják
azokat. A család néhány tagja rendelkezik egy RING-finger motívummal, egy cink ujj
doménnel, amely révén ubikvitin-konjugáló enzimet vonzanak és az ubikvitinálja a
kaszpázokat.
A sejthalál folyamatáról önálló könyvek születnek, természetesen jelen fejezetben
csak a legfontosabb mozzanatokra, a fő irányvonalakra térhetünk ki. A következőkben
szeretnénk felvázolni a programozott sejthalál két főbb útvonalát, az extrinszik,
(külsődleges) és az intracelluláris, intrinszik útvonal legfőbb történéseit.
A korábbiaknak megfelelően a prokaszpáz aktiváció folyamatában fontos szerepet
kapnak az adaptor fehérjék, amelyek segítenek összegyűjteni és aktív konformációba
hozni az iniciátor prokaszpázokat.
A sejten kívüli prokaszpáz aktivációra igen kiváló példa a killer limfociták által
iniciált sejthalál. A killer limfociták képesek azáltal sejthalált előidézni, hogy egy Fas
ligand nevű fehérjét prezentálnak, amely képes a célsejtek felszínén található
halálreceptor fehérjéhez, a Fas-hoz kötődni (7.5. ábra). Az ily módon klaszterekbe
tömörült Fas fehérjék intracellulárisan adaptor fehérjéket kötnek, amelyek kötik a
prokaszpáz 8 molekulákat. Ezek térszerkezet-változásuk következtében hasítják,
aktiválják egymást. Az ily módon aktiválódott kaszpáz 8 molekulák ezt követően hasítják
az effektor kaszpázaikat és apoptózist indukálnak (7.5. ábra). Néhány esetben a
stresszelt, vagy sérült sejtek mind a Fas ligand, mind a Fas fehérje előállítására képesek,
ily módon elindítva az intracelluláris kaszpáz kaszkádot.
7.5. ábra. A halálligandon – halálreceptoron– keresztül bonyolódó külsődleges (extrinszik), illetve a mitokondrium külső membrán sérülésén, a citokrom c felszabadulásán keresztül
zajló, belsődleges (intrinszik) apoptotikus útvonalak

7. Összetett folyamatok szabályozása 115
© Szarka András www.interkonyv.hu
A stresszelt és károsodott sejtek esetében legjobban ismert intracelluláris apoptózis
útvonal a mitokondriumból kiinduló programozott sejthalál folyamata. A sérült,
stresszelt fehérjék mitokondriumának külső membránrepedését követően az
elektronátvitelben szerepet játszó fehérje, a citokróm c kikerül a citoplazmába, ahol
az Apaf-1 adaptorfehérjéhez köt és aktiválja azt; az ily módon létrejött Apaf-1
klaszterhez köt a prokaszpáz 9. A prokaszpáz aktivációja indítja el a további effektor
kaszpáz aktivációt. Az apoptózis legtöbb formája esetén találkozunk ezen mitokondriális
prokaszpáz aktivációval, amely nagymértékben hozzájárul a kaszpáz kaszkád
elindításához, gyorsításához és felerősítéséhez.
7.3. Sérült fehérjeválasz (UPR) az endoplazmás retikulumban
A transzláció során szintetizált fehérjeláncok jelentős része további reakciók tárgya lesz,
hogy elnyerje végső, biológiailag aktív formáját. Ezeket a transzlációt követő,
fehérjeláncot érintő reakciókat összefoglalóan poszttranszlációs módosulásoknak
nevezzük. Poszttranszlációs módosulásokon mind a prokarióta, mind az eukarióta sejtek
fehérjéi keresztülmennek.
Csak felsorolásképp néhány folyamatot ragadjunk ki: egyes fehérjék szerin, treonin
és tirozin részletei foszforilálódnak (kofaktorként ATP-t igényelve); több véralvadási
faktor aszpartát és glutamát részletei további karboxil-csoportokkal egészülnek ki
(kofaktorként K-vitamint igényelve); izomfehérjék és a citokróm c lizin részletei
metilálódhatnak (kofaktorként N-adenozil-metionint igényelve). A kollagének fő
szerkezetalkotója a hidroxi-prolin, aszkorbinsavat igénylő folyamat során jut hozzá
hidroxil csoportjához. A glikoproteinekre is poszttranszlációs módosulásuk során kerül
fel szénhidrát oldalláncuk. Több bakteriális és eukarióta fehérje igényel aktivitásához
kovalensen kötött prosztetikus csoportot. Ezen csoportok is a fehérjelánc riboszómáról
történő távozásakor épülnek be. Példaként említhetjük a kovalensen kötött biotint az
acetil-CoA karboxiláz esetében, valamint a hem csoportot a citokróm c esetében.
Az exportra kerülő fehérjékben gyakran alakulnak ki kovalens diszulfid kötések,
láncközi és láncok közötti cisztein részletek, között natív konformációjuk kialakulásakor.
Az így kialakult diszulfid kötések segítenek a natív fehérjeszerkezet megőrzésében az
intracelluláristól oly különböző viszonyok között.
A fehérjék foldingjában résztvevő sejtszervecskékben olyan adaptációs mechaniz-
musok, programok működnek, amelyek lehetővé teszik a nem megfelelő módon feltekert
fehérjék detektálását, illetve a sérült, nem megfelelő térszerkezettel rendelkező fehérjék
korrekcióját. Az endoplazmás retikulum lumenében zajlik a sejtfelszíni, illetve a szek-
récióra szánt fehérjék poszt-transzlációs módosítása, foldingja. Az endoplazmás retiku-
lum fehérje minőségi kontrollgépezete a folding útvonalakkal szorosan együttműködve,
igen szelektíven azon őrködik, hogy az egész kismértékű elváltozásokat, a folding
apparátus enyhe hatásfokzavarait is érzékelje. Az érzékelés eredményeként a naszcens
fehérjéket mint tévesen feltekert fehérjéket minősíti, ennek következtében akkumulálja,

116 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
illetve lebontja. Így nem meglepő, hogy az endoplazmás retikulum olyan fehérjéket is
tartalmaz, amelyek elsődleges feladata, hogy a tévesen feltekert, sérült térszerkezetű
fehérjéket detektálja és olyan génexpressziós változásokat indítson el, amely komoly
hatással van az endoplazmás retikulum foldingkapacitására. Az endoplazmás retikulum
stresszre adott válasz jó modellként szolgát egyrészt arra, hogy megértsük, mi módon
érzékeli a stresszt az organellum, másrészt arra is, hogy egy adott organellum
homeosztázisának felborulása miként hat ki szerteágazó módon a sejt különböző
funkcióira, mint a génexpresszió, metabolizmus, jelátvitel és az apoptózis.
Az endoplazmás retikulumban felgyűlt sérült szerkezetű fehérjék egy koordinált
adaptációs programot indítanak el a sejtben, ez a sérült fehérjeválasz (az unfolded
protein response: UPR). A sérült fehérjeválasz során a sejt csökkenteni igyekszik a
stresszt azáltal, hogy a fehérjefolding és degradáció folyamatait erősíti (ezekben a
folyamatokban részt vevő fehérjék kifejezését, szintézisét fokozza), valamint igyekszik
csökkenteni a fehérjefolding apparátusra nehezedő nyomást oly módon, hogy az
általános fehérjeszintézist gátolja.
Lássuk csak, mely folyamatok befolyásolják a fehérjeszintézist! Ezek között akad
fiziológiás és patológiás folyamat egyaránt.
7.6. ábra. Az endoplazmás retikulum stresszt és a következményes sérült fehérjeválaszt kiváltó különböző fiziológiás, patológiás, illetve kísérleti tényezők
Az endoplazmás retikulumban folyó fehérjefolding bármilyen zavara kiváltja a sérült
fehérjeválaszt (7.6. ábra). Ezek között számos fiziológiás, normális történés is akad, ilyen
például a professzionális szekréciós sejtek, mint az inzulintermelő béta és a plazma-
sejtek kifejlődése, differenciációja. De kiváltó ok lehet a megváltozó helyzet, mint a
glükózhiány, vagy a homociszteinémia és az isémia. Kiválthatja a szekréciós, vagy
transzmembrán fehérjék génjében bekövetkező mutáció (amelyek praktikusan az endo-
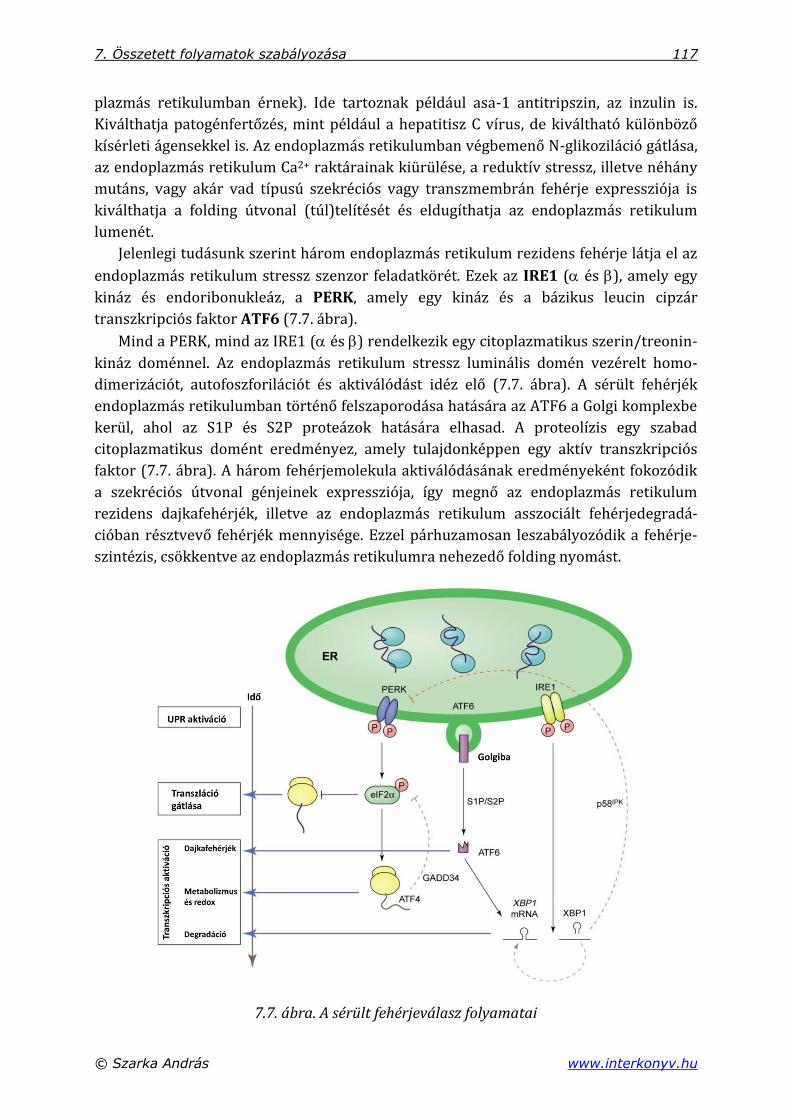
7. Összetett folyamatok szabályozása 117
© Szarka András www.interkonyv.hu
plazmás retikulumban érnek). Ide tartoznak például asa-1 antitripszin, az inzulin is.
Kiválthatja patogénfertőzés, mint például a hepatitisz C vírus, de kiváltható különböző
kísérleti ágensekkel is. Az endoplazmás retikulumban végbemenő N-glikoziláció gátlása,
az endoplazmás retikulum Ca2+ raktárainak kiürülése, a reduktív stressz, illetve néhány
mutáns, vagy akár vad típusú szekréciós vagy transzmembrán fehérje expressziója is
kiválthatja a folding útvonal (túl)telítését és eldugíthatja az endoplazmás retikulum
lumenét.
Jelenlegi tudásunk szerint három endoplazmás retikulum rezidens fehérje látja el az
endoplazmás retikulum stressz szenzor feladatkörét. Ezek az IRE1 ( és ), amely egy
kináz és endoribonukleáz, a PERK, amely egy kináz és a bázikus leucin cipzár
transzkripciós faktor ATF6 (7.7. ábra).
Mind a PERK, mind az IRE1 ( és ) rendelkezik egy citoplazmatikus szerin/treonin-
kináz doménnel. Az endoplazmás retikulum stressz luminális domén vezérelt homo-
dimerizációt, autofoszforilációt és aktiválódást idéz elő (7.7. ábra). A sérült fehérjék
endoplazmás retikulumban történő felszaporodása hatására az ATF6 a Golgi komplexbe
kerül, ahol az S1P és S2P proteázok hatására elhasad. A proteolízis egy szabad
citoplazmatikus domént eredményez, amely tulajdonképpen egy aktív transzkripciós
faktor (7.7. ábra). A három fehérjemolekula aktiválódásának eredményeként fokozódik
a szekréciós útvonal génjeinek expressziója, így megnő az endoplazmás retikulum
rezidens dajkafehérjék, illetve az endoplazmás retikulum asszociált fehérjedegradá-
cióban résztvevő fehérjék mennyisége. Ezzel párhuzamosan leszabályozódik a fehérje-
szintézis, csökkentve az endoplazmás retikulumra nehezedő folding nyomást.
7.7. ábra. A sérült fehérjeválasz folyamatai

118 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
Tekintsük végig, hogy az egyes stressz szenzor molekulák mely történésekért és mi
módon felelősek. Az ATF6 által szabályozott gének átírása akkor növekszik meg, amikor
az ATF6 citoplazmatikus doménje a sejtmagba kerül (7.7. ábra). Az IRE1 génexpressziót
fokozó hatása oly módon érvényesül, hogy az aktív fehérje endoribonukleáz doménje, az
X-box binding protein (XBP1) gén, mRNS-ből kihasít egy 26 nukleotid hosszú részletet.
Ez a hasítás egy transzlációs frameshiftet eredményez, amelynek hatására létrejön az
XBP1 transzkripciós faktor (7.7. ábra). Ez utóbbi transzkripciós faktor a sejtmagba
kerülve fokozza a degradációban részt vevő fehérjék kifejeződését. A PERK aktivációja
az eIF2 transzlációs iniciációs faktor α-alegységének foszforilációját eredményezi,
amely gátolja a funkcióképes 80S riboszóma összeszerelését. A folyamat végeredménye
így a fehérjeszintézis általános gátlása lesz.
A nem stresszelt sejtekben mindhárom fehérje a BiP dajkafehérjével asszociáltan
található meg. Az IRE1 és az ATF6 kötőhelye már ismert, a PERK BiP kötőhelye még
ismeretlen, de az IRE1-hez igen hasonló részlet található ezen molekulában is. A BiP-hez
történő asszociáció (normális esetben) bent tartja az ATF6 molekulát az endoplazmás
retikulum lumenében, így nem hasadhat és viselkedhet transzkripciós faktorként
(magyarul inaktív). Az IRE1 és a PERK pedig a dimerizálódásért felelős doménjével
kapcsolódik a BiP-hez, tehát ezen fehérjék is dimerizálódásra képtelenek, azaz
aktiválódásra képtelenek lesznek, amennyiben BiP-hez kötődnek. A BiP azon régiója,
amellyel az IRE1-hez és a PERK-hez kötődik, pedig nem más, mint amellyel a sérült
fehérjék hidrofób régiójához kötődik.
Az UPR aktivációjának folyamata ezek szerint a következőképpen képzelhető el: a
sejt normális – nem stresszelt – állapotában a BiP (az endoplazmás retikulumban
legnagyobb mennyiségben előforduló dajkafehérje) az ATF6-hoz, az IRE1-hez és a
PERK-hez kötődik. Amennyiben az endoplazmás retikulumban felszaporodnak a sérült,
helytelen térszerkezetű fehérjék, akkor ezen fehérjék hidrofób részeihez kötődik és
szabadon hagyja a három szenzormolekulát. A szabaddá vált szenzormolekulák közül az
ATF6 a Golgi apparátusba kerül, ahol a proteázok által vágódik, aktiválódik. Az IRE1 és a
PERK szabaddá válnak és dimerizálódnak, majd autofoszforilálódnak és így ők is
aktiválódnak.
Egyelőre megválaszolásra váró kérdés, hogy a három szenzormolekula által vezérelt
folyamatok kizárólag egymással párhuzamosan, vagy egymástól függetlenül is
végbemennek-e? A kísérletes stressz mindig igen jelentős mennyiségű sérült fehérjét
eredményez, így igen nehezen vizsgálható a független aktiválódás kérdésköre.
Sérült fehérjeválasz és apoptózis
A sejt az UPR összehangolt folyamatai révén igyekszik megbirkózni az endoplazmás
retikulum stresszel, azonban egy bizonyos szint felett erre képtelen lesz. Ekkor beindul a
programozott sejthalál, az endoplazmás retikulum stressz kiváltotta apoptózis folya-
mata. Az endoplazmás retikulum specifikus kaszpáz 12 képes in vitro a citokróm c-től
és az Apaf-1-től függetlenül aktiválni a kaszpáz 9-et. A kaszpáz 12 az IRE1 és az
adaptorfehérje TRAF2-vel kialakított kapcsolat révén az endoplazmás retikulum
membránjánál található. Az endoplazmás retikulum stressz hatására a TRAF2 disszo-

7. Összetett folyamatok szabályozása 119
© Szarka András www.interkonyv.hu
ciál a kaszpáz 12-ről, amely folyamatot a kaszpáz dimerizációja és aktiválódása követ
(7.8. ábra). A kaszpáz 12 UPR mediálta apoptózisban betöltött szerepét megerősítette,
hogy a kaszpáz 12 dupla hiánymutáns sejtek valamelyest rezisztensek voltak az endo-
plazmás retikulum stressz kiváltotta apoptózisra. A teljes képhez azonban hozzátartozik,
hogy a sejtek nem mutattak teljes mértékű endoplazmás retikulum stressz kiváltotta
apoptózis rezisztenciát, amely alapján azt valószínűsíthetjük, hogy, kompenzálva a
kaszpáz 12 hiányát, más jelátviteli útvonalak is hozzájárulhatnak az apoptózis kivál-
tásához.
7.8. ábra. Az endoplazmás retikulum kiváltotta UPR, illetve az UPR elbukását követő endoplazmás retikulum eredetű apoptózis folyamata
Az endoplazmás retikulum, a mitokondrium és a citoplazma Ca2+ koncentrá-
ciójának befolyásolása is feltehetően egy apoptózis-kiváltó jel lehet (7.8. ábra). A
Ca2+szint befolyásoló szerepét aláhúzza az a megfigyelés is, mely szerint a kaszpáz 12
aktivációja szintén bekövetkezhet a Ca2+-dependens proteáz kalpain általi hasítása
révén is. Az endoplazmás retikulumból történő Ca2+ kiáramlást követően igen gyorsan
bekövetkezik a Ca2+ mitokondriális felvétele, melyben különösen a mitokondrium azon
részei játszanak szerepet, amelyek szorosan és stabilan asszociálnak az endoplazmás
retikulum membránnal. A Ca2+ mitokondriális felvétele a mitokondriális membrán-
potenciál kollapszusához és így a mitokondriális citokróm c felszabadulásához, az
intrinszik apoptotikus (erősítő) útvonal beindulásához vezethet.

www.interkonyv.hu © Szarka András
Utószó
A Biokémia II. – Biokémiai szabályozás című tankönyvet igyekeztünk a legújabb
kísérletes bizonyítékok felhasználásával, de érdekes, a mindennapi életből vett példákra
alapozva elkészíteni. Célunk az volt, hogy az olvasó felejtse el, hogy egy tankönyvet tart a
kezében, és észrevétlenül merüljön el a sejtben, kísérje nyomon az izgalmas biokémiai
szabályozási folyamatokat. Reméljük, hogy izgalmas kalandokban volt része és
észrevétlenül a tudás birtokába is került. Ha így van elértük célunkat.
A tankönyv a leggondosabb írás és többszöri korrektúrát, lektorálást követően is
tartalmazhat hibákat. Ezúton arra szeretném kérni az olvasót, amennyiben hibát észlel,
vagy bármilyen javaslata, észrevétele van a tankönyvvel kapcsolatban, tegye azt meg a
következő e-mail címen: [email protected].
Köszönettel,
Szarka András
Budapest, 2013. nyár

© Szarka András www.interkonyv.hu
Ábraanyag
A tankönyv ábraanyagát két részre oszthatjuk. Az első rész saját készítésű ábrákból áll. A
második részbe tartozó ábrák, az interneten fellelhető ábraanyag direkt, vagy azok
átalakított változatának felhasználásai. Ez utóbbi csoportba tartozó ábrák listája az
eredeti linkkel együtt a következőkben található. A linkeket első ízben 2013. július,
augusztus hónapban töltöttük le, majd ellőrzés céljából 2014. január 21-én.
Internetes forrású ábrajegyzék
2.4. ábra. Az enzimek legfontosabb katalitikus stratégiái
http://www.ncbi.nlm.nih.gov/books/NBK26911/figure/A483/?report=objectonly
2.5. ábra. A lizozim katalitikus mechanizmusa
http://www.bio.miami.edu/tom/courses/protected/ECB/CH05/5_28.jpg
3.1. A sejtben folyó biokémiai reakciók összessége
http://biochemicalpathwayswallchart.blogspot.hu/2010/10/roche-biochemical-
pathways-wall-chart.html
3.2. ábra. Feed-back gátlás. A metabolikus út egy korai szakaszán található enzimet a
metabolikus út egy későbbi szakaszának terméke gátolja
http://www.ncbi.nlm.nih.gov/books/NBK26911/figure/A490/?report=objectonly
3.3. ábra. Többszörösen elágazó (aminosav) metabolikus út szabályozása feed-back
gátlással
http://www.ncbi.nlm.nih.gov/books/NBK26911/figure/A491/?report=objectonly
3.4. ábra. Pozitív allosztérikus reguláció
http://www.ncbi.nlm.nih.gov/books/NBK26911/figure/A494/?report=objectonly
3.5. ábra. Negatív allosztérikus reguláció
http://www.ncbi.nlm.nih.gov/books/NBK26911/figure/A495/?report=objectonly
3.6. ábra. Egy- és több alegységes enzimek inhibitor koncentráció – relatív
enzimaktivitás összefüggése
http://www.ncbi.nlm.nih.gov/books/NBK26911/figure/A497/?report=objectonly

122 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
3.7. ábra. Két azonos alegységből álló enzim kooperatív-allosztérikus átmenete
http://www.ncbi.nlm.nih.gov/books/NBK26911/figure/A498/?report=objectonly
3.8. ábra.A mioglobin monomer szerkezete
http://www.fizyka.umk.pl/~wiesiek/HEMOHIS.JPG
3.9. ábra.A hemoglobin tetramer szerkezete
http://www.baileybio.com/plogger/?level=picture&id=490
3.15. ábra. Fehérje foszforiláció/defoszforiláció
http://www.ncbi.nlm.nih.gov/books/NBK26911/figure/A504/?report=objectonly
3. 17. ábra. Proteinkináz háromdimenziós szerkezete
http://www.ncbi.nlm.nih.gov/books/NBK26911/figure/A505/?report=objectonly
3.19. ábra. Fehérjék aktivitásának befolyásolása foszforilációval/defoszforilációva
http://www.ncbi.nlm.nih.gov/books/NBK26911/figure/A504/?report=objectonly
4.1. ábra. Fehérjék transzlációjával párhuzamosan zajló folding
http://www.ncbi.nlm.nih.gov/books/NBK26829/figure/A1101/?report=objectonly
4.2. ábra.A hsp70-család munka közben
http://www.ncbi.nlm.nih.gov/books/NBK26829/figure/A1104/?report=objectonly
4.3. ábra. A hsp60 család szerkezeti felépítése és működése
http://www.ncbi.nlm.nih.gov/books/NBK26829/figure/A1105/?report=objectonly
4.5. ábra. A proteaszóma felépítése
http://www.nature.com/nrm/journal/v6/n1/box/nrm1552_BX3.html
4.7. ábra. A fehérjék ubikvitinációja
http://www.ncbi.nlm.nih.gov/books/NBK26829/figure/A1110/?report=objectonly
4.8. ábra. Az ubikvitin-ligáz szabályozása
http://www.ncbi.nlm.nih.gov/books/NBK26829/figure/A1113/?report=objectonly
4.9. ábra. Degradációs szignálok kialakulása
http://www.ncbi.nlm.nih.gov/books/NBK26829/figure/A1113/?report=objectonly
5.2. ábra. A sejtdifferenciáció a génexpresszió megváltozása, nem a
nukleotidszekvenciáé.
http://www.ncbi.nlm.nih.gov/books/NBK26885/figure/A1210/?report=objectonly
5.4. ábra. A DNS-kötő fehérjék kapcsolódási helye a DNS-molekula külső felszínén
http://www.ncbi.nlm.nih.gov/books/NBK26806/figure/A1235/?report=objectonly

Ábraanyag 123
© Szarka András www.interkonyv.hu
5.7. ábra. A szimmetrikus DNS-regulációs szekvencia és hozzá kötődő DNS-felismerő
fehérje dimer
http://www.ncbi.nlm.nih.gov/books/NBK26806/figure/A1239/?report=objectonly
5.10. ábra. A leucin-cipzár fehérjerészlet szerkezete és kötődése a DNS-hez
http://www.ncbi.nlm.nih.gov/books/NBK26806/figure/A1249/?report=objectonly
5.11. ábra. A hélix-loop-hélix fehérjerészlet szerkezete és kapcsolódása a DNS-hez
http://www.ncbi.nlm.nih.gov/books/NBK26806/figure/A1255/?report=objectonly
5.12. ábra. Hélix-loop-hélix fehérjerészlet aktivitásának szabályozása
heterodimerizációval
http://www.ncbi.nlm.nih.gov/books/NBK26806/figure/A1256/?report=objectonly
5.13. ábra.A gél mobility shift assay (electrophoretic mobility shift assay, EMSA)
működési elve
http://www.ncbi.nlm.nih.gov/books/NBK26806/figure/A1261/?report=objectonly
5.14. ábra. A triptofán operon elemei
http://www.ncbi.nlm.nih.gov/books/NBK26872/figure/A1271/?report=objectonly
5.15. ábra. A transzkripció lépései
http://www.ncbi.nlm.nih.gov/books/NBK26887/figure/A988/?report=objectonly
5.16. ábra. A triptofán-represszor fehérje működése
http://www.ncbi.nlm.nih.gov/books/NBK26872/figure/A1272/?report=objectonly
5.17. ábra. A triptofán-represszor fehérje szerkezetének és DNS-kötő képességének
megváltozása a két bekötődő triptofánmolekula hatására
http://www.ncbi.nlm.nih.gov/books/NBK26872/figure/A1273/?report=objectonly
5.18. ábra. A lac-operon kettős szabályozása a lac-represszor fehérje, illetve a CAP–
cAMP-komplex által
http://www.ncbi.nlm.nih.gov/books/NBK26872/figure/A1278/?report=objectonly
5.20. ábra. Az eukarióta gén szabályozási régiójának elemei
http://www.ncbi.nlm.nih.gov/books/NBK26872/figure/A1287/?report=objectonly
5.21. ábra. Nukleoszóma-remodellezés
http://www.ncbi.nlm.nih.gov/books/NBK26872/figure/A1293/?report=objectonly
5.22. ábra. Kovalens hisztonmódosítás
http://www.ncbi.nlm.nih.gov/books/NBK26872/figure/A1294/?report=objectonly

124 Biokémiai szabályozás
www.interkonyv.hu © Szarka András
5.23. ábra. A szabályozó fehérjeelemek összeszerelődése különböző funkciójú
regulátorkomplexekké a DNS felületén
http://www.ncbi.nlm.nih.gov/books/NBK26872/figure/A1301/?report=objectonly
5.24. ábra. Az enhanszoszóma felépítése
http://www.ncbi.nlm.nih.gov/books/NBK26872/figure/A1302/?report=objectonly
5.25. ábra. Az inzulátorszekvenciák elhelyezkedése és funkciója
http://www.ncbi.nlm.nih.gov/books/NBK26872/figure/A1316/?report=objectonly
5.26. ábra. Az X-kromoszóma inaktiválódása (kondenzálódása) a női embrionális testi
sejtekben
http://www.ncbi.nlm.nih.gov/books/NBK26854/figure/A1348/?report=objectonly
5.27. ábra. Az X-kromoszóma inaktiválódásának mechanizmusa
http://www.ncbi.nlm.nih.gov/books/NBK26854/figure/A1349/?report=objectonly
5.28. ábra. A DNS-metiláció mechanizmusa, a fenntartó metilázok működése
http://www.ncbi.nlm.nih.gov/books/NBK26854/figure/A1353/?report=objectonly
5.29. ábra. A genomi lenyomat hatása a génexpresszióra
http://www.ncbi.nlm.nih.gov/books/NBK26854/figure/A1357/?report=objectonly
5.30. ábra. Az Igf2-gén metilációs lenyomata
http://www.ncbi.nlm.nih.gov/books/NBK26854/figure/A1358/?report=objectonly
6.1. ábra. A celluláris jelátvitel szintjei
http://www.ncbi.nlm.nih.gov/books/NBK21059/figure/A2741/?report=objectonly
6.2. ábra. Sejtfelszíni receptorok
http://www.ncbi.nlm.nih.gov/books/NBK26813/figure/A2746/?report=objectonly
6.3. ábra. Intracelluláris magreceptorok
http://www.ncbi.nlm.nih.gov/books/NBK26813/figure/A2746/?report=objectonly
6.4. ábra. A szignálmolekulák hatótávolsága
http://www.ncbi.nlm.nih.gov/books/NBK26813/figure/A2748/?report=objectonly
6.5. ábra. Az extracelluláris jelre adott válasz természete, reakcióideje
http://www.ebookfiles.org/molecular-biology-of-the-cell-5th-edition-pdf-free-full-
download/
6.7. ábra. A G-fehérje mint jelátviteli kapcsoló
http://www.ebookfiles.org/molecular-biology-of-the-cell-5th-edition-pdf-free-full-
download/

Ábraanyag 125
© Szarka András www.interkonyv.hu
6.8. ábra. A G-fehérjék szerkezeti felépítése
http://www.ncbi.nlm.nih.gov/books/NBK26912/figure/A2797/?report=objectonly
6.9. ábra. A G-fehérjék működés közben
http://www.ncbi.nlm.nih.gov/books/NBK26813/figure/A2767/?report=objectonly
6.14. ábra. A cAMP-mediátorrendszer működés közben
http://www.ncbi.nlm.nih.gov/books/NBK26912/figure/A2806/?report=objectonly
6.15. ábra. A cAMP-dependens protein kináz (PKA) szerepe a génszintű szabályozásban
http://www.ncbi.nlm.nih.gov/books/NBK26912/figure/A2806/?report=objectonly
6.16. ábra. A foszfolipáz-C aktiválása és működése
http://www.ncbi.nlm.nih.gov/books/NBK26912/figure/A2811/?report=objectonly
6.17. ábra. Az IP3-jelátviteli útvonal
http://www.ncbi.nlm.nih.gov/books/NBK26912/figure/A2812/?report=objectonly
7.4. ábra. A nekrotikus és az apoptotikus sejthalál közötti morfológiai és folyamatbeli
különbségek
http://www.springerimages.com/Images/RSS/1-10.1007_s10103-009-0723-y-0
7.5. ábra. A halálligandon – halálreceptoron– keresztül bonyolódó külsődleges
(extrinszik), illetve a mitokondrium külső membrán sérülésén, a citokrom c
felszabadulásán keresztül zajló, belsődleges (intrinszik) apoptotikus útvonalak
http://www.nature.com/nrd/journal/v4/n5/fig_tab/nrd1717_F2.html
7.6. ábra. Az endoplazmás retikulum stresszt és a következményes sérült fehérjeválaszt
kiváltó különböző fiziológiás, patológiás, illetve kísérleti tényezők
http://www.sciencedirect.com/science/article/pii/S0962892403002782
7.7. ábra. A sérült fehérjeválasz folyamatai
http://www.sciencedirect.com/science/article/pii/S0962892403002782
7.8. ábra. Az endoplazmás retikulum kiváltotta UPR, illetve az UPR elbukását követő
endoplazmás retikulum eredetű apoptózis folyamata
http://www.sciencedirect.com/science/article/pii/S0962892403002782

www.interkonyv.hu © Szarka András
Felhasznált és ajánlott irodalom
Andersen MH, Becker JC, Straten PT (2005) Regulators of apoptosis: suitable targets for
immune therapy of cancer Nature Reviews Drug Discovery4:399–409.
Ciechanover A (2005) Proteolysis: from the lysosome to ubiquitin and the
proteasomeNature Reviews Molecular Cell Biology6: 79–87.
Rutkowski DT, Kaufman RJ. (2004) A trip to the ER: coping with stress. Trends Cell Biol.
14:20–28.
Jeremy M. Berg, John L. Tymoczko, and Lubert Stryer:Biochemistry 5th editionNew York,
W. H. Freeman, 2002. ISBN 0716730510
Robert K. Murray, Victor W. Rodwell, David Bender, Kathleen M. Botham, P. Anthony
Weil, Peter J.: Kennelly Harper's Illustrated Biochemistry, 28th edition, McGraw-Hill
Medical 2009. ISBN 0071625917
David L. Nelson and Michael M. Cox: Lehninger Principles of Biochemistry (5th Edition)
W. H. Freeman; 5th edition, 2008. ISBN 071677108X
Bruce Alberts, Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, Peter Walter:
Molecular Biology of the Cell (5th edition), Garland Science, 2008. ISBN 9780815341055
Ádám Veronika (szerk.): Orvosi Biokémia, Medicina Könyvkiadó 2006. ISBN
963242767X
Machovich Raymund, Mandl József (szerk.): Orvosi patobiokémia, Medicina, 2007.







