
Asymptotic Techniques in Asymptotic Techniques in Enzyme KineticsEnzyme Kinetics
Presented By:– Dallas Hamann– Ryan Borek– Erik Wolf– Carissa Staples– Carrie Ruda

OutlineOutline
Compartmental AnalysisChemical ReactionsLaw of Mass ActionEnzyme ReactionsThe Equilibrium ApproximationThe Quasi-Steady-State ApproximationEnzyme Inhibition

Compartmental Compartmental DiagramsDiagrams
Compartmental Diagrams are visual models of a physical, biological, or a
biochemical system or process.

Compartmental Diagram Compartmental Diagram ElementsElements
Represents an amount of homogenous material in the process.
Labeled with a variable name.
Alcohol
Compartments

Compartmental Diagram Compartmental Diagram ElementsElements
Represents flow into or out of a compartment.
Labeled by amount of variable change per unit of time.
Inflow of Alcohol/second
Outflow of Alcohol/second
Arrows

Compartmental Diagram Compartmental Diagram Governing PrincipleGoverning Principle
Rate of change in X = inflow rate – outflow rate
X
Using this principle allows formation of a system of differential equations using a compartmental diagram. A differential equation
can be formed for each compartment.

Physiological Perspective of Physiological Perspective of Compartmental DiagramsCompartmental Diagrams
Variables represent amounts of biological substances in a physiological system.
Compartments represent these physiological systems.

Fundamental QuestionsFundamental Questions
To understand the distribution of biological substances amongst various components of a physiological system.
To understand how the distribution of biological substances change in a system.

Chemical Reactions and Law of Mass Chemical Reactions and Law of Mass ActionAction
Suppose C represents a chemical, [C] which denotes its concentration.
Now suppose two chemicals A,B react upon collisions to form a product C.
where k is the rate constant of the reaction.
CBA k

Law of Mass ActionLaw of Mass ActionA ModelA Model
= rate of accumulation of product
Which depends on:
- the energy of the collision
- geometrical shapes and sizes of the reactant molecules
dt
Cd ][

Law of Mass ActionLaw of Mass ActionA ModelA Model
dt
Cd ][ Is directly proportional to reactant concentrations
]][[][
BAkdt
Cdi.e.

Compartmental Diagram for Compartmental Diagram for Law of Mass ActionLaw of Mass Action
A B
C
]][[1 BAk
][1 Ck

Reverse ReactionsReverse Reactions
Biochemical reactions are typically bi-directional.
CBAk
k
1
1
So, we get the differential equations:
i)][]][[
][11 CkBAk
dt
Cd
ii)]][[][
][11 BAkCk
dt
Ad

EquilibriumEquilibriumA state when concentrations are no longer changing.
In equilibrium, 0
][
dt
Ad
So, 0]][[][ 11 BAkCk
Solving for
eqeqeq BAk
kC ][][][
1
1
(1)

EquilibriumEquilibriumNow, in a closed system (no other reactions are going on)
eqeqeq BCAk
kC
CAA
ACA
])[][(][
][][
][][
01
1
0
0
(constant)
(conservation of matter equation)
Substituting in (1),

EquilibriumEquilibriumSolving for
eqC][
eqeqeq
eqeqeqeq
eqeqeq
eqeqeqeq
BAk
kB
k
kC
BAk
kBC
k
kC
BCk
kBA
k
k
BCBAk
kC
][)][1(][
][][][][
][][][
)][][][(][
01
1
1
1
01
1
1
1
1
10
1
1
01
1

EquilibriumEquilibrium
eqeq
eqeq
eq
eqeq
eq
eq
eq
BK
BAC
Bkk
BAC
Bkk
BAkk
C
][
][][
][
][][
][1
][
][
0
1
1
0
1
1
01
1
Where, (equilibrium constant) has concentration units1
1
k
kKeq

ReactionsReactions
• Elementary • Enzyme

Elementary ReactionsElementary Reactions
Proceed directly from collision of reactantsFollows the Law of Mass Action directly
CBA k

EnzymesEnzymes
•An enzyme is a substance that acts as a catalyst for some chemical reaction.
•Enzymes act on other molecules (called substrates), helping convert them into products.
•Enzymes themselves are not changed by the reaction.
•Enzymes work by lowering the “free energy of activation” for the reaction.

Enzyme ReactionsEnzyme Reactions
Enzyme reactions do not follow the Law of Mass Action directly
If they did, theory would predict:
CESk
k
1
1
but it has been shown that this is not the case.
So, what do they look like?

Michaelis – Menten Model Michaelis – Menten Model (1913)(1913)
Idea Chemical Reaction Scheme Compartmental Diagram System of Differential Equations

IdeaIdea
2-step process
Step 1: Enzyme E first converts the substrate S into complex C
Step 2: Complex then breaks down into a product P, releasing the enzyme E in the process

Chemical Reaction Chemical Reaction SchemeScheme
EPCES k
k
k
2
1
1
is the dissociation constant2kNote:

Compartmental DiagramCompartmental Diagram
ck 1
sek1
ck2

Differential EquationsDifferential EquationsFrom the compartmental diagram, we get the following differential equations:
sekckdt
ds11 sekckk
dt
de121 )(
ckksekdt
dc)( 211
ckdt
dp2

So What Can We Do With All So What Can We Do With All Of This?Of This?

The Equilibrium The Equilibrium ApproximationApproximation
Michaelis – Menten (1913)
Dr. Maud MentenLeonor Michaelis
Picture Not Available

PurposePurpose
To estimate the reaction velocity of an enzyme reaction.
dt
dpV i.e. To estimate
EPCES k
k
k
2
1
1
for

AssumptionAssumptionSubstrate is in “instantaneous equilibrium” with the complex.
0dt
dsi.e.
Consider : 011 sekck
sekck 11
or

sekck 11
note: cee o
SolvingSolving for cfor c
1
1
k
kks
seck
k
1
1 letting
secks
So, by substitution )( cesck os

sekck 11 SolvingSolving for cfor c(Continued)(Continued)
os secksc
os seskc )(
sk
sec
s
o
scseck os
thus,

Derive an expression for VDerive an expression for V
Recall from previous differential equations:
ckdt
dpV 2
Substituting in the new value for c:
sk
sekV
s 02

ObservationsObservations1) The maximum reaction velocity occurs when C is biggest ( ).
maxV0ec
i.e. When all enzyme is complexed with substrate.
022max ekckdt
dpV
will be limited by the amount of enzyme present and the dissociation constant so,
is called “rate limiting” for this reaction.
02max ekV 2k
EPC k 2

Observations(cont.)Observations(cont.)
2) For large substrate concentrations, we will rewrite V as:
sk
sVV
s max
1
max
skV
s
s
s 1
max
s
kV
s
maxmax
1V
V
i.e. the reaction rate saturates

Observations(cont.)Observations(cont.)
3) If , sks
s
s
k
kVV
2maxthen
2maxV

Is that the only approximation Is that the only approximation technique for V?technique for V?
Clearly, that would not suffice!
Hold on to your seats for another exciting method,
because it gets even better.

Quasi-Steady-State Quasi-Steady-State ApproximationApproximation

IdeaIdea Step 1: Use nondimensionalization to redefine
rates. Step 2: Apply the Briggs –Haldane Assumption to
those rates.
J.B.S. Haldane George Edward Briggs
Picture Not Available

AssumptionAssumptionThe rates of formation and the rate of breakdown of the complex are equal.
0dt
dci.e.

RatesRates We know these differential equations:
sekckdt
ds11
sekckkdt
de121 )(
ckksekdt
dc)( 211
ckdt
dp2
cee 0

Variables and ParametersVariables and Parameters
Independent variables: s, c
Parameters: k1, k-1, k2
By nondimensionalization, we make new variables:
os
s:
oe
cx : tek o1:
oss oxec
oekt
1
:

Step 1: ReplacementStep 1: ReplacementWe start by rewriting our differential rate equations from using
our independent variables to using the new variables.
dt
d
d
ds
dt
ds
dt
dsoo
oekdt
d1:
We know: By substitution:
d
deks
dt
dsoo 1
1) Note:dt
d
d
d
dt
d

)1( xexeecee oooo
Now substitute:
Solve for: )1(1
1 xxks
k
d
d
o
d
deksxeskxek
sekckdt
ds
ooooo 111
11
)1(
into:
Let1
1
ks
k
o
so )(
xd
d
d
deks
dt
dsoo 1
oss oxec
Previous Statement:

ckksekdt
dc)( 121
oekdt
d1:
We know: By substitution:
ddx
kedt
dco 12
2)
dt
d
d
dxe
dt
dxe
dt
dcoo
dt
d
d
dx
dt
dx
Note:

)1( xee o :
into:
We don’t want to divide by zero (in case eo is small) so we will divide the right by so to minimize parameters.
d
dxkekkxexesk
ckksekdt
dc
oooo 12
121
211
)()1(
)(
)()1(1
12
k
kkxxs
d
dxe oo
)()1(1
12
oo
o
sk
kkxx
d
dx
s
e
Previous Statement:
ddx
kedt
dco 12
Now substitute:
oss oxec

)()1(1
12
oo
o
sk
kkxx
d
dx
s
e
Let
o
o
s
e and
1
12
ks
kk
o
so
)(
)1(
xd
dx
xxd
dx
d
dx
s
e
o
o
And now our parameters are: ,
Previous Statement:

Step 2: Apply assumptionStep 2: Apply assumptionNow we will apply the assumption of Briggs-Haldane:
0dt
dxeo
We assumed:
Because of these assumptions we can conclude:
xec o
0dt
dcWhich is comparable to : 0
ddx
then 012
ddx
keo 0
d
dx

Now applying those to our rates:
)(0
xd
dx
Solve for x:
x
Substitute x into
)()(xd
d
)(
d
d
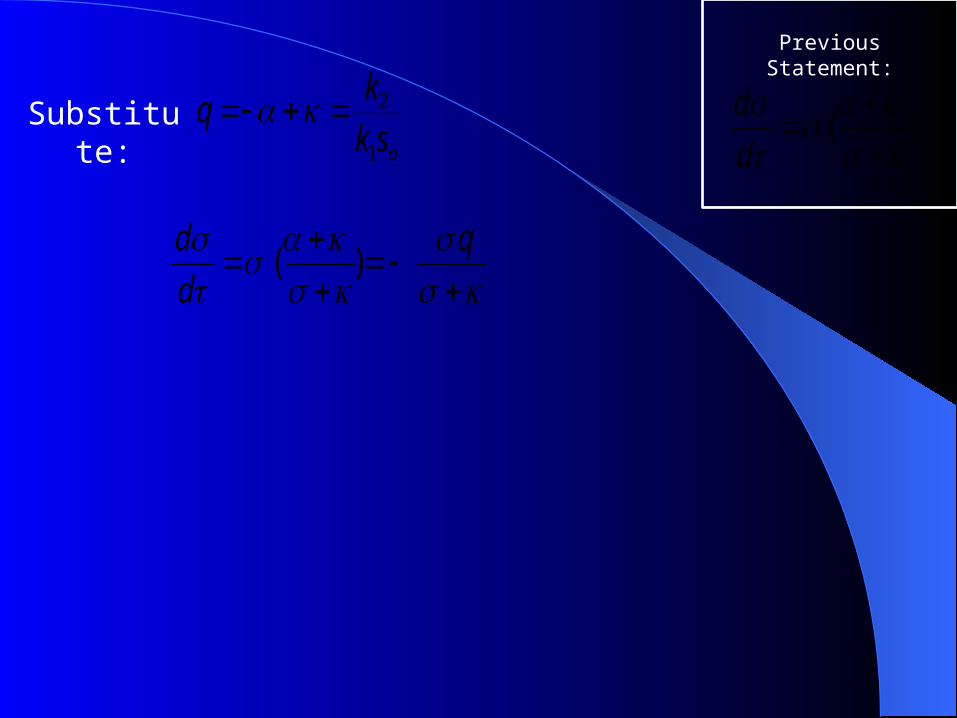
Substitute:
q
d
d)(
osk
kq
1
2
Previous Statement:
)(
d
d

Bringing it all togetherBringing it all together
0dt
dp
dt
dc
dt
dsWe know:
Assume 0dt
dc by the Quasi-Steady-State Assumption.
sodt
ds
dt
dp
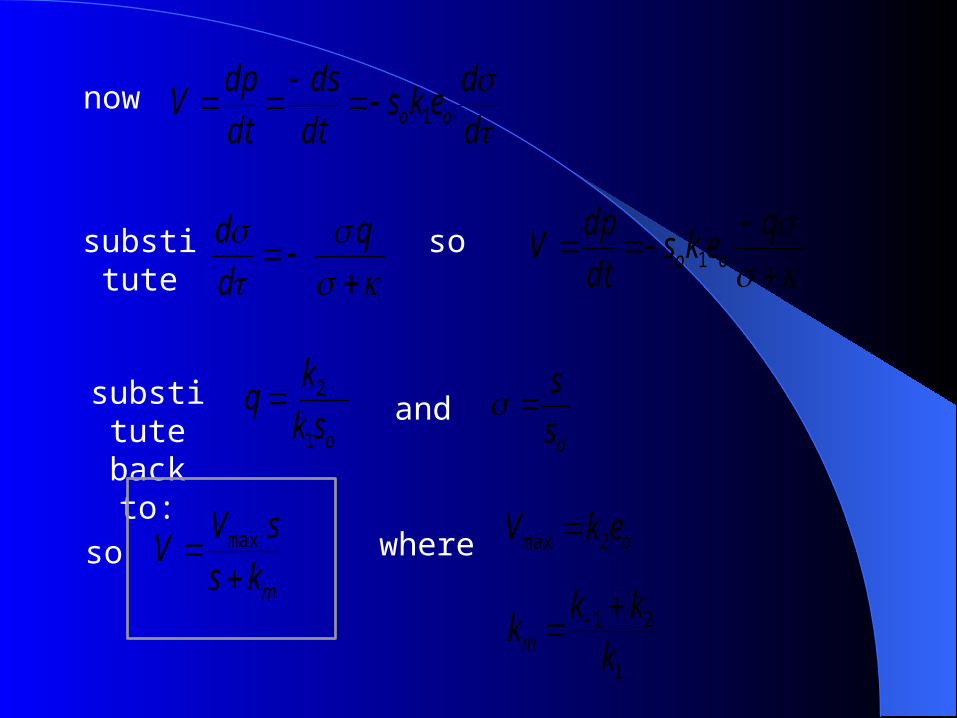
nowd
deks
dt
ds
dt
dpV oo 1
substitute
q
d
d so
q
eksdt
dpV oo 1
substituteback to: osk
kq
1
2 andos
s
somks
sVV
max where oekV 2max
1
21
k
kkkm

Equilibrium vs. Equilibrium vs. Quasi-Steady-StateQuasi-Steady-State
These two reaction schemes are similar, but not the same.
Differences include: the Equilibrium Approximation is simple to apply but has less scope, while the Quasi-
Steady-State uses nondimensionalization and applies to a greater scope.
In other reaction schemes the estimates are not as similar.

Enzyme InhibitionEnzyme Inhibition
Enzyme inhibitors are substances that inhibit the catalytic action of an enzyme.
Slow down or decrease enzyme activity to zero. Examples: nerve gas and cyanide are irreversible
inhibitors or catalytic poisons (these examples reverse the activity of life supporting enzymes).
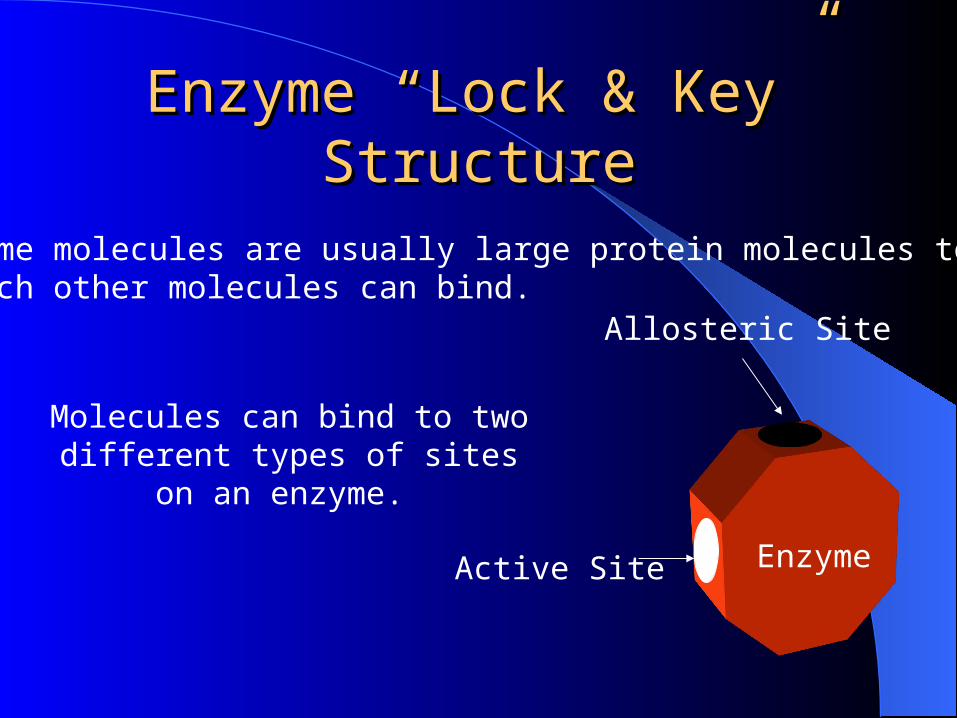
Enzyme “Lock & Key” Enzyme “Lock & Key” StructureStructure
Enzyme
Molecules can bind to two different types of sites on an enzyme.
Active Site
Allosteric Site
Enzyme molecules are usually large protein molecules to which other molecules can bind.

Enzyme “Lock & Key” Enzyme “Lock & Key” StructureStructure
Active Sites are sites on an enzyme where
substrate can bind to form complex.
However, when an inhibitor is bound to the
active site it is called Competitive Inhibition. Enzyme
Active Site
Allosteric Site
Competitive Inhibition
Inhibitor

Enzyme “LockEnzyme “Lock & Key” & Key” StructureStructure
Allosteric Sites are secondary sites on an
enzyme that regulate the catalytic activity of an
enzyme.
When an inhibitor is bound to the allosteric site
it is called Allosteric Inhibition.
EnzymeActive Site
Allosteric Site
Inhibitor
Allosteric Inhibition

Competitive InhibitionCompetitive Inhibition
Enzyme
Active Site
Allosteric Site
Inhibitor
Reaction Scheme
S + E C1 E + Pk1
k-1
k2
E + I C2
k3
k-3
In competitive inhibition the inhibitor “competes” with the substrate to bind to the Active Site. If the inhibitor wins there is less product formed.

Allosteric InhibitionAllosteric Inhibition
Enzyme
Active Site
Allosteric Site
InhibitorIn allosteric inhibition the inhibitor binds to the Allosteric Site. The binding of the inhibitor to the Allosteric Site changes the action of the enzyme. However, the substrate may also bind to the Enzyme at the Active Site. This leaves a very complicated reaction scheme.
Substrate

Allosteric Inhibition Reaction Allosteric Inhibition Reaction SchemeScheme
Let:S ~ SubstrateE ~ EnzymeI ~ InhibitorC1 ~ ES complexC2 ~ EI complexC3 ~ EIS complexP ~ Product
E + S C1 E + Pk1
k-1
k2
E + I C2
k3
k-3
C2 + S C3
k1
k-1
C1 + Ik3
k-3
C3

Allosteric Inhibition Reaction Allosteric Inhibition Reaction SchemeScheme
For Allosteric Inhibition we should use “Complex Free Reaction Notation.”
In Complex Free Reaction Notation one substance is implied.

Allosteric Inhibition Reaction Allosteric Inhibition Reaction SchemeScheme
E + S C1 E + Pk1
k-1
k2
E ES E + Pk1s
k-1
k2
Former Notation:
Complex Free Notation:
Let:S ~ SubstrateE ~ EnzymeI ~ InhibitorC1 ~ ES complexC2 ~ EI complexC3 ~ EIS complexP ~ Product

Allosteric Inhibition Reaction Allosteric Inhibition Reaction SchemeScheme
E ES E + Pk1s
k-1
k2
EI EISk1s
k-1
k3i k-3 k3i k-3
Complex Free Notation:

Allosteric Inhibition Reaction Allosteric Inhibition Reaction SchemeScheme
Complex Free Compartmental DiagramLet:X = ESY = EIZ = EIS E X
ZY
Pk1se
k-1x
k2x
k1sy
k-1z
k3ie k-3y k3ix k-3z

Allosteric Inhibition Reaction Allosteric Inhibition Reaction Differential EquationsDifferential Equations
zkzksykixkdt
dz3113
. . .

ConclusionConclusion
As you can see, asymptotic techniques in enzyme kinetics can get quite complex. However, these techniques give us vital information about the model without having to solve the differential equation directly. We will continue our study into next semester.

BibliographyBibliography Mathematical Physiology by Keener and Sneyd,
Springer-Verlag 1998 Mathematics Applied to Deterministic Problems in the
Natural Sciences by Lin and Segal, SIAM 1988 Mathematical Models in Biology by Leah Edelstein-
Keshet, McGraw-Hill 1988 A Course in Mathematical Modeling by Mooney and
Swift, MAA 1999 Sites of pictures:
http://www.cdnmedhall.org/Inductees/menten_98.htm
http://www.marxists.org/archive/haldane

Special Thanks to:Special Thanks to:
Dr. Jean FoleyDr. Steve DeckelmanAll of you for coming!







