
AN INVESTIGATION INTO
MATHEMATICAL MODELLING
OF INTEGRATED BIOSYSTEMS
FOR OPERATIONAL CONTROL
AND MANAGEMENT
By
Khalid Shamim
2014
A Thesis Submitted for the Degree of
Doctor of Philosophy
School of Animal & Veterinary Sciences/
School of Chemical Engineering
The University of Adelaide
Australia

i
ABSTRACT
The South Australian Research & Development Institute (SARDI) and the
Environmental Biotechnology Cooperative Research Centre (EBCRC)
undertook a project “Commercial Scale Integrated Biosystems for Organic
Waste and Wastewater Treatment for the Livestock and Food Processing
Industries”, for which this research forms a part. The Integrated Biosystems
(IBS) project laboratory was set up at the Roseworthy campus of The
University of Adelaide, South Australia. The major objective of this project
was to develop an Integrated Biosystems (IBS) on a commercial scale for
the treatment of wastewater by applying the stages of anaerobic digestion
and bioconversion stages involving algae, zooplankton and fish. The IBS
developed could be used in both rural and urban settings for efficient waste
disposal and generation of energy in the process. The overall aim of this
research was to develop a mathematical model of an IBS for operational
control and management.
The objectives of this research were to:
1. Develop a mathematical model for the anaerobic digestion system.
2. Use an existing model to simulate the aquaculture stages of the IBS
and to test its suitability for a commercial scale IBS for effective
control and management.
3. Conduct a sensitivity analysis on the parameters of the aquaculture
model.
4. Develop an automatic calibration program to validate the
aquaculture model with real time field data.
The major contribution from this research was to elucidate the key
parameters required to simulate the integrated IBS model. The data was
collected through a series of rigorous experimentation for both the anaerobic
digestion and aquaculture modules of the IBS. The data obtained was used
to parameterise a coupled anaerobic digestion-hydrodynamic ecological

ii
model. The resulting model adequately simulated key processes within the
IBS, which was further improved with a novel auto calibration algorithm.
The primary contribution of this work has been to develop an automatic
parameter calibration for the aquaculture component of the model.
Parameter calibration in aquaculture models has been time consuming as it
is basically a “trial and error” procedure. This thesis presents a significant
contribution in this area.
A literature review conducted on the models developed for the IBS and
automatic calibration revealed certain gaps in the previous research
conducted, which formed a basis for further work in this PhD study. A
significant proportion of research time was invested in set up, installation
and commissioning of the pilot plant two-stage anaerobic digestion system
and the Integrated Aquaculture (mesocosm) facility. This was done to
understand the mechanism of the IBS. Both the systems were run for a
period of 12 months for data collection. During this time, operational
glitches and troubleshooting provided the researcher an opportunity to
implement engineering skills in the IBS context and understand the
behaviour of this complex system. Batch scale experiments were conducted
in the laboratory for data collection to develop a model for the anaerobic
digestion system using microbial kinetics. The aquaculture model DYRESM
CAEDYM was used to simulate the aquaculture stages of the IBS. The
model source code was altered to run the model for the IBS set up. Real
time field data could not be obtained as the commercial scale IBS was not
constructed due to administrative hurdles which were beyond the
researcher’s control. A sensitivity analysis was conducted on selected
parameters of the model to determine the behaviour of model outputs on
controlled changes to those parameters. Finally an automatic model
calibration and validation program was written in FORTRAN 90 to
automatically validate the model with field data as opposed to the
conventional methods of manual parameter validation. Pseudo field data
was used for demonstration purposes.

iii
The primary contributions from this research have been to assess the
suitability of DYRESM CAEDYM as the modelling software for the
commercial scale IBS, run a sensitivity analysis on selected parameters of
the model and execute an automatic model calibration and validation
program.
The phytoplankton ponds had a domination of cyanobacteria growth for the
maximum part of the year, due to thermal stratification in the summer
months, which otherwise would not happen in an IBS with proper control
and management. There was negligible zooplankton and fish growth due to
diminished chlorophyte concentrations.
Results from similar IBS studies in India and France were used to compare
results with the proposed commercial IBS. The comparative IBS examples
sourced from sites in India and France show that IBS has been successfully
implemented in different parts of the world comprising tools for better
control and management of ponds. The use of mixing (agitation) and
aeration assist in mixing the ponds and the effluent uniformly which
minimises stratification in ponds and thus reduces the growth of
cyanobacteria, and in turn improves the growth of phytoplankton,
zooplankton and fish. The model DYRESM CAEDYM could incorporate
the use of mixers and aeration in the IBS ponds to overcome the problems
of algal crashes in summer.
The sensitivity analysis conducted on the parameters of the model show that
the model results for phytoplankton growth is highly sensitive to those
parameters that directly affect the growth rates e.g. maximum phytoplankton
growth rate, phytoplankton respiration coefficient and phytoplankton
temperature multiplier. The automated calibration routine incorporated a
novel methodology to calibrate and validate DYRESM CAEDYM
automatically without having to manually adjust parameters. This procedure
is a significant improvement over the conventional methodology of
validating the model by “trial and error” which was time consuming and to a
certain extent inaccurate. The simulations ran successfully to validate the
model parameters with the pseudo field data. This calibration program could

iv
be also used to validate other outputs from the model and is a significant
contribution in this research.
The parameters which can be controlled for managing the commercial scale
IBS in an effective way would be parameters related to inflow and outflow
volumes and flow rates of effluent, retention time of the effluent, nutrient
loads, rates of mixing and aeration within the ponds and control of biomass
conversion for primary, secondary and tertiary productions. These
management strategies could also be used to operate an IBS with a variety
of different effluents to its maximum capacity and construct an IBS with
better module design.
The use of automated calibration of parameters has high applicability in the
development of mathematical models for managing the performance of
wastewater recycling technology, which is in high demand in the modern
world in order to reduce the dependence on limited water resources. The
calibration routine developed in this PhD study has demonstrated that for a
complex aquaculture model like DYRESM CAEDYM where manually
validating the parameters is a tedious task, automatic calibration routine
using GLUE methodology is an effective way to validate the model which
minimises the risks of computational errors.

v
STATEMENT OF ORIGINALITY I certify that this work contains no material which has been accepted for the
award of any other degree or diploma in any university or other tertiary
institution and, to the best of my knowledge and belief, contains no material
previously published or written by another person, except where due
reference has been made in the text. In addition, I certify that no part of this
work will, in the future, be used in a submission for any other degree or
diploma in any university or other tertiary institution without the prior
approval of the University of Adelaide and where applicable, any partner
institution responsible for the joint-award of this degree.
I give consent to this copy of my thesis, when deposited in the University
Library, being made available for loan and photocopying, subject to the
provisions of the Copyright Act 1968.
I also give permission for the digital version of my thesis to be made
available on the web, via the University’s digital research repository, the
Library catalogue and also through web search engines, unless permission
has been granted by the University to restrict access for a period of time.
Signed: ……………………………..
KHALID SHAMIM
Dated: ………………………………

vi
ACKNOWLEDGEMENTS I would like to sincerely thank my Principal Supervisor, Professor Martin S
Kumar and Co-Supervisor, Associate Professor David Lewis for their
continuous support and guidance throughout the entire project. I feel
honoured to have been supervised by these outstanding academics and thank
them for their input, encouragement and undivided attention. I would also
like to thank Environmental Biotechnology Cooperative Research Centre
(EBCRC) for awarding me a full postgraduate scholarship, and in particular
I would like to thank Dr. David Garman for his support and giving me an
opportunity to be part of the Project 6 (P6) group, for which this thesis
forms a part.
I would also like to take this opportunity to thank the many people who
assisted me in my research. Firstly, Dr. Stephen Carr for assisting me in
compiling computer programs and setting up simulations on The University
of Adelaide Server; Mr. Paul Harris for guiding me through the anaerobic
digestion process; Mrs. Sandy Wyatt and Mrs. Belinda Rodda for laboratory
support and Mr. Andrew Ward for helping me understand aquaculture
concepts.
I have also had the privilege to collaborate with several interstate and
overseas researchers that gave invaluable suggestions and scope for
exploration of novel ideas. I would like to express my gratitude to Dr. Matt
Hipsey, Dr. Daniel Botelho and Dr. Jason Antenucci from the Centre for
Water Research, University of Western Australia; Dr. Pratap
Pullammanappallil from University of Florida, USA and Mr. Dennis Trolle
from the University of Waikato, New Zealand
Finally I would like to thank my wife Afreen, my parents and my extended
family for their patience and encouragement throughout my PhD studies.

vii
LIST OF PUBLICATIONS Shamim, K., Kumar, M.S., Lewis, D.M., (2014), “Automated Parameter
Estimation and Calibration”, Hydrological Processes (In review).
Shamim, K., Kumar, M.S., Lewis, D.M., (2008), “Development of
DYRESM CAEDYM as an aquaculture model for an Integrated
Biosystems”, Environmental Biotechnology CRC Annual Conference, 8-10
December 2008, Adelaide, Australia (Poster Presentation).
Shamim, K., Kumar, M.S., Lewis, D.M., (2007), “A mathematical model
for the anaerobic digestion of raw piggery effluent”, 11th World Congress on
Anaerobic Digestion, 23-27 September 2007, Brisbane, Australia (Poster
Presentation).
Shamim, K., Kumar, M.S., Pullammanappallil, P., Ho, G (2006),
“Mathematical model development for anaerobic digestion system in an
Integrated Biosystems”, Environmental Biotechnology CRC Annual
Conference, 28-29 November 2006, Sydney, Australia (Poster Presentation).

viii
Table of Contents
1 Introduction ...................................................................................... 1
1.1 Integrated Biosystems .............................................................................................. 1
1.2 IBS Project ................................................................................................................ 4
1.3 Aim of the Research .................................................................................................. 6
1.4 Background .............................................................................................................. 7
1.5 Commercial Scale Integrated Biosystems .................................................................. 9
1.5.1 Anaerobic Digestion ............................................................................................................. 10 1.5.2 Microalgal Ponds ................................................................................................................. 10 1.5.3 Zooplankton and Fish Ponds ................................................................................................ 11
1.6 Organisation of the Thesis ...................................................................................... 12
1.7 Research Program ................................................................................................... 13
2 Literature Review ............................................................................ 14
2.1 Anaerobic Digestion ............................................................................................... 14
2.1.1 Hydrolysis and Fermentation ............................................................................................... 15 2.1.2 Acetogenesis and Homoacetogenesis ................................................................................. 16 2.1.3 Methanogenesis .................................................................................................................. 16
2.2 Mathematical Models in Anaerobic Digestion ......................................................... 19
2.3 Primary and Secondary Production ......................................................................... 22
2.4 Models developed for Primary and Secondary Production ...................................... 24
2.4.1 The Hydrodynamic Model DYRESM ..................................................................................... 24 2.4.2 The Aquatic Ecological Model CAEDYM ............................................................................... 35
2.5 Techniques used in Water Quality Models Calibration ............................................ 47
2.5.1 GLUE .................................................................................................................................... 49
2.6 Summary ................................................................................................................ 52
3 Experimental Methods and Data Collection ..................................... 54
3.1 Introduction ........................................................................................................... 54
3.2 Set up of the Pilot Scale Two-‐Stage Anaerobic Digestion System ............................. 55

ix
3.2.1 Equipment: Reactors and Digesters ..................................................................................... 55 3.2.2 Analytical techniques ........................................................................................................... 57
3.3 Pilot Plant Stage 1 Experiments .............................................................................. 59
3.3.1 Acidogenesis ........................................................................................................................ 59 3.3.2 Methanogenesis .................................................................................................................. 59 3.3.3 Results ................................................................................................................................. 60
3.4 Pilot Plant Stage 2 Experiments .............................................................................. 65
3.4.1 Methods ............................................................................................................................... 65 3.4.2 Results ................................................................................................................................. 67 3.4.3 Conclusions .......................................................................................................................... 80
3.5 Set up of the Pilot Scale Integrated Aquaculture System ......................................... 82
3.6 Bioconversion of piggery effluent to algae (280 L working volume) ......................... 84
3.6.1 Objective .............................................................................................................................. 84 3.6.2 Materials and Methods ....................................................................................................... 84 3.6.3 Results ................................................................................................................................. 85 3.6.4 Key findings .......................................................................................................................... 88
3.7 Bioconversion of piggery effluent to algae (180 L working volume) ......................... 88
3.7.1 Objective .............................................................................................................................. 88 3.7.2 Materials and Methods ....................................................................................................... 89 3.7.3 Results ................................................................................................................................. 89 3.7.4 Key findings .......................................................................................................................... 92
3.8 Lab Scale Anaerobic Digestion Experiments ............................................................ 93
3.8.1 Introduction ......................................................................................................................... 93 3.8.2 Materials & Methods ........................................................................................................... 93
3.9 Conclusions ............................................................................................................ 95
4 Development of an Anaerobic Digestion Model ............................... 97
4.1 Introduction ........................................................................................................... 97
4.2 Methods ................................................................................................................. 98
4.2.1 Development of Model Equations for the Anaerobic Digestion Process ........................... 101
4.3 Results .................................................................................................................. 103
4.3.1 Methane Model ................................................................................................................. 103 4.3.2 TAN Model ......................................................................................................................... 105 4.3.3 P Model .............................................................................................................................. 107

x
4.4 Comparison of model data vs. measured data ...................................................... 108
4.5 Discussion ............................................................................................................. 112
4.6 Further Work ........................................................................................................ 113
5 Modelling Commercial Scale Integrated Biosystems ...................... 115
5.1 Introduction ......................................................................................................... 115
5.1.1 Model Description ............................................................................................................. 117
5.2 Methods ............................................................................................................... 118
5.2.1 Research Site ..................................................................................................................... 118 5.2.2 Alterations to the original source code of DYRESM CAEDYM ............................................ 119 5.2.3 Input Data .......................................................................................................................... 121
5.3 Results .................................................................................................................. 125
5.3.1 Temperature ...................................................................................................................... 125 5.3.2 Phytoplankton growth in algal pond 1 ............................................................................... 126 5.3.3 Phytoplankton growth in algal pond 2 ............................................................................... 130 5.3.4 Zooplankton growth .......................................................................................................... 134 5.3.5 Fish growth ........................................................................................................................ 136
5.4 Discussion ............................................................................................................. 139
5.4.1 Limitations of the model .................................................................................................... 142 5.4.2 Comparison with algal data obtained from Bolivar Wastewater Treatment Plant ........... 143
5.5 Typical Outputs from established IBS .................................................................... 145
5.5.1 Central Institute of Freshwater Aquaculture, India ........................................................... 145 5.5.2 IBS set up in France ............................................................................................................ 147
5.6 Conclusions .......................................................................................................... 149
6 Sensitivity Analysis ........................................................................ 152
6.1 Introduction ......................................................................................................... 152
6.2 Methods ............................................................................................................... 154
6.3 Results .................................................................................................................. 157
6.3.1 Maximum Phytoplankton Growth Rate (Pmax) ................................................................... 157 6.3.2 Phytoplankton Respiration Coefficient (kr) ........................................................................ 158 6.3.3 Phytoplankton Temperature Multiplier (vR) ...................................................................... 159 6.3.4 Minimum Phytoplankton Internal P (IPmin) ........................................................................ 160 6.3.5 Maximum Phytoplankton Internal P (IPmax) ....................................................................... 161

xi
6.3.6 Maximum Rate of Phytoplankton P Uptake (UPmax) .......................................................... 162 6.3.7 Minimum Phytoplankton Internal N (INmin) ....................................................................... 163 6.3.8 Maximum Phytoplankton Internal N (INmax) ...................................................................... 164 6.3.9 Maximum Rate of Phytoplankton N Uptake (UNmax) ......................................................... 165 6.3.10 Half Saturation Constant for Phytoplankton P Uptake (KP) ............................................. 166 6.3.11 Half Saturation Constant for Phytoplankton N Uptake (KN) ............................................ 167 6.3.12 Parameter for initial slope of P-‐I curve (Ik) ...................................................................... 168
6.4 Discussion ............................................................................................................. 169
6.5 Conclusions and Further Work .............................................................................. 171
7 Automated Parameter Estimation and Calibration ........................ 172
7.1 Introduction ......................................................................................................... 172
7.2 Methods ............................................................................................................... 173
7.2.1 Incorporating Monte Carlo and GLUE calibration in DYRESM CAEDYM ............................ 173 7.2.2 Analysis of the auto calibration program .......................................................................... 174 7.2.3 Flowchart of the GLUE program ........................................................................................ 176
7.3 Results .................................................................................................................. 177
7.3.1 GLUE Calibration for Chlorophyte growth ......................................................................... 177 7.3.2 GLUE calibration for Cyanobacteria growth ...................................................................... 181
7.4 Discussion and Conclusions ................................................................................... 184
7.5 Further Work ........................................................................................................ 186
8 Summary and Conclusions ............................................................. 187
8.1 General Discussions and Conclusions .................................................................... 187
8.2 Summary of the Research Results ......................................................................... 187
8.2.1 Laboratory Experiments and Development of Anaerobic Digestion Model ...................... 187 8.2.2 Modelling the Aquaculture Component of the IBS ............................................................ 189 8.2.3 Sensitivity Analysis ............................................................................................................. 191 8.2.4 Automated Parameter Estimation and Calibration ........................................................... 191
8.3 Summary .............................................................................................................. 192
8.4 Recommendations for further studies .................................................................. 192
9 References ..................................................................................... 195

xii
Appendix A Anaerobic Digestion Models .......................................... 219
Appendix B Data from Batch Scale Anaerobic Digestion
Experiments 229
Appendix C DYRESM CAEDYM INPUT FILES ....................................... 233
Appendix D Nutrient Data in the IBS ponds ....................................... 235
Appendix E Program Code for Auto Calibration of DYRESM
CAEDYM 250
Appendix F GLUE Calibration Numerical Outputs .............................. 260

xiii
Table of Figures Figure 1.1 Schematic of material flow within an Integrated Biosystems ............... 3
Figure 1.2 Schematic of the IBS Project ................................................................. 5
Figure 1.3 Flowchart of the IBS process ............................................................... 11
Figure 2.1 Flow Chart of processes in anaerobic digestion
(Pullammanappallil, Chynoweth et al. 2001) ........................................................ 18
Figure 2.2 Schematic Flow Chart of DYRESM (Robson and Hamilton
2004) ...................................................................................................................... 25
Figure 2.3 DYRESM simulation process (Imerito 2007) ..................................... 26
Figure 2.4 Surface energy flux exchanges ............................................................ 29
Figure 2.5 Overview of CAEDYM state variables showing the water
column, benthic and sediment components (Hipsey, Romero et al. 2006) ........... 36
Figure 2.6 Schematic of phytoplankton dynamics within CAEDYM
(Hipsey, Romero et al. 2006) ................................................................................ 38
Figure 2.7 Schematic of zooplankton dynamics within CAEDYM (Hipsey,
Romero et al. 2006) ............................................................................................... 42
Figure 2.8 Schematic of fish dynamics within CAEDYM (Hipsey, Romero
et al. 2006) ............................................................................................................. 46
Figure 3.1 Schematic diagram of the pilot scale anaerobic digestion system ....... 57
Figure 3.2 Thermophilic (stainless steel) digesters in the pilot scale research
facility .................................................................................................................... 58
Figure 3.3 Mesophilic (polybag) digesters in the pilot scale research facility ...... 58
Figure 3.4 pH profile during acidogenesis in T1 and T2 treating piggery
effluent .................................................................................................................. 61
Figure 3.5 Ammonia (TAN) levels in T1 and T2 during acidification of raw
piggery effluent ..................................................................................................... 62
Figure 3.6 Soluble Phosphorus (SP) levels in T1 and T2 during the
acidification of raw piggery effluent ..................................................................... 62
Figure 3.7 Gas production during anaerobic acidification of piggery effluent
in T1 (55 0C) and T2 (38 0C) ................................................................................. 63
Figure 3.8 Gas production during anaerobic digestion of acidified piggery
effluent in polydigesters 1 and 2 operating under ambient conditions .................. 65

xiv
Figure 3.9 pH profile during acidogenesis in reactors T1 and T2 treating raw
piggery effluent ..................................................................................................... 71
Figure 3.10 VFA profile during acidogenesis in reactors T1 and T2 treating
raw piggery effluent .............................................................................................. 72
Figure 3.11 TKN profile during acidogenesis in reactors T1 and T2 treating
raw piggery effluent .............................................................................................. 72
Figure 3.12 Ammonia nitrogen profile during acidogenesis in reactors T1
and T2 treating raw piggery effluent ..................................................................... 73
Figure 3.13 TP profile during acidogenesis in reactors T1 and T2 treating
raw piggery effluent .............................................................................................. 73
Figure 3.14 SP profile during acidogenesis in reactors T1 and T2 treating
raw piggery effluent .............................................................................................. 74
Figure 3.15 Biogas production and methane composition during
acidogenesis in reactors T1 and T2 treating raw piggery effluent ........................ 74
Figure 3.16 pH profile during anaerobic digestion in polydigesters D1, D2,
D3 & D4 treating acidified piggery effluent from the acidogenic reactors ........... 77
Figure 3.17 VFA profile during anaerobic digestion in polydigesters D1,
D2, D3 & D4 treating acidified piggery effluent from the acidogenic
reactors .................................................................................................................. 78
Figure 3.18 TKN profile during digestion in polydigesters D1, D2, D3 & D4
treating acidified piggery effluent from the acidogenic reactors .......................... 78
Figure 3.19 Ammonia profile during digestion in polydigesters D1, D2, D3
& D4 treating acidified piggery effluent from the acidogenic reactors ................ 79
Figure 3.20 Soluble P profile during digestion in polydigesters D1, D2, D3
& D4 treating acidified piggery effluent from the acidogenic reactors ................ 79
Figure 3.21 Biogas production and methane composition during digestion in
polydigesters D1, D2, D3 & D4 treating acidified piggery effluent from the
acidogenic reactors ................................................................................................ 80
Figure 3.22 Schematic diagram of the pilot scale Integrated Aquaculture
System ................................................................................................................... 83
Figure 3.23 Clear Perspex tanks set up at differential heights for micro-algal
culture while blue fibre glass tanks are used for fish culture. These tanks are
part of the indoor integrated aquaculture system (mesocosm) at Roseworthy
Laboratory ............................................................................................................. 83

xv
Figure 3.24 Mean cell density in 280 L algal culture ............................................ 86
Figure 3.25 Mean TAN concentration in 280 L algal culture ............................... 87
Figure 3.26 Mean SP concentration in 280 L algal culture ................................... 87
Figure 3.27 Mean pH in 280 L algal culture ......................................................... 88
Figure 3.28 Mean cell density for 180 L algal culture .......................................... 90
Figure 3.29 Mean TAN for 180 L algal culture .................................................... 91
Figure 3.30 Mean SP for 180 L algal culture ........................................................ 91
Figure 3.31 Mean pH for 180 L algal culture ........................................................ 92
Figure 3.32 Experimental apparatus for the laboratory study of anaerobic
digestion. ............................................................................................................... 95
Figure 4.1 Decrease in COD of the raw piggery effluent ..................................... 98
Figure 4.2 Cumulative methane output ................................................................. 99
Figure 4.3 Increase in TAN of the raw piggery effluent ....................................... 99
Figure 4.4 Increase in soluble P of the raw piggery effluent .............................. 100
Figure 4.5 Scaled/Normalized Plot for Anaerobic Digestion of Piggery
Effluent ................................................................................................................ 101
Figure 4.6 Modelled methane data at 55 0C ........................................................ 104
Figure 4.7 Temperature Response for Growth and Death Rates in CH4
modelling ............................................................................................................. 105
Figure 4.8 Modelled Total Ammonia Nitrogen (TAN) data at 55 0C ................. 106
Figure 4.9 Temperature Response for Growth and Death Rates in TAN
modelling ............................................................................................................. 106
Figure 4.10 Modelled Soluble P data at 55 0C .................................................... 107
Figure 4.11 Temperature Response for Growth and Death Rates in P
modelling ............................................................................................................. 108
Figure 5.1 Schematic of the proposed commercial scale IBS ............................. 119
Figure 5.2 Meteorological data comprising short wave radiation, air
temperature, rainfall, wind speed, cloud cover and vapour pressure
(clockwise, starting from left) ............................................................................. 123
Figure 5.3 Simulated pond temperature (all ponds are the same
temperature) ........................................................................................................ 125
Figure 5.4 Simulated chlorophyte growth in algal pond 1 .................................. 126
Figure 5.5 Simulated cyanobacteria growth in algal pond 1 ............................... 127
Figure 5.6 Simulated freshwater diatoms growth in algal pond 1 ....................... 128

xvi
Figure 5.7 Nutrient Profile in Algal Pond 1 ........................................................ 129
Figure 5.8 Simulated chlorophyte growth in algal pond 2 .................................. 130
Figure 5.9 Simulated cyanobacteria growth in algal pond 2 ............................... 131
Figure 5.10 Simulated fresh water diatoms growth in algal pond 2 .................... 132
Figure 5.11 Nutrient Profile in Algal Pond 2 ...................................................... 133
Figure 5.12 Simulated zooplankton growth in zooplankton pond ...................... 134
Figure 5.13 Nutrient Profile in Zooplankton Pond ............................................. 135
Figure 5.14 Simulated fish growth in fish pond .................................................. 136
Figure 5.15 Simulated zooplankton growth in fish pond .................................... 137
Figure 5.16 Nutrient Profile in Fish Pond ........................................................... 138
Figure 5.17 Algal succession data obtained from Bolivar Wastewater
Treatment Plant ................................................................................................... 144
Figure 5.18 Algal succession data obtained from Bolivar Wastewater
Treatment Plant for the period 2000 - 2001 ........................................................ 144
Figure 5.19 Diagram of the pilot-scale IBS for recycling swine manure in
France .................................................................................................................. 149
Figure 6.1 Chlorophyll-a response to change in Pmax values .............................. 157
Figure 6.2 Chlorophyll-a response to change in kr values .................................. 158
Figure 6.3 Chlorophyll-a response to change in vR values .................................. 159
Figure 6.4 Chlorophyll-a response to changes in IPmin values ............................ 160
Figure 6.5 Chlorophyll-a response to changes in IPmax values ............................ 161
Figure 6.6 Chlorophyll-a response to changes in UPmax values .......................... 162
Figure 6.7 Chlorophyll-a response to changes in INmin values ........................... 163
Figure 6.8 Chlorophyll-a response to changes in INmax values ........................... 164
Figure 6.9 Chlorophyll-a response to changes in UNmax values ......................... 165
Figure 6.10 Chlorophyll-a response to changes in KP values ............................. 166
Figure 6.11 Chlorophyll-a response to changes in KN values ............................. 167
Figure 6.12 Chlorophyll-a response to changes in Ik values ............................... 168
Figure 7.1 FlowChart of the Monte Carlo & GLUE Calibration script .............. 176
Figure 7.2 Nash Sutcliffe Coefficients for GLUE Calibration ............................ 177
Figure 7.3 Comparison between simulated (model) and field data before
GLUE calibration for chlorophyte growth .......................................................... 179
Figure 7.4 Comparison between simulated (model) and field data after
GLUE calibration for chlorophyte growth .......................................................... 179

xvii
Figure 7.5 Nash – Sutcliffe Coefficients for GLUE calibration for
cyanobacteria growth .......................................................................................... 181
Figure 7.6 Comparison between simulated (model) and field data before
GLUE calibration for cyanobacteria growth ....................................................... 183
Figure 7.7 Comparison between simulated (model) and field data after
GLUE calibration for cyanobacteria growth ....................................................... 183

xviii
List of Tables
Table 2.1 Description of the seven phytoplankton groups configurable
within CAEDYM .................................................................................................. 37
Table 3.1 Effluent characteristics in T1 and T2 .................................................... 60
Table 3.2 Effluent characteristics in the polydigesters operating at ambient
temperature ............................................................................................................ 64
Table 3.3 Performance parameters of acidogenic reactor T1 (38 0C) during
period 1 (Days 1-65) ............................................................................................. 67
Table 3.4 Performance parameters of acidogenic reactor T2 (55 0C) during
period 1 (Days 1-65) ............................................................................................. 67
Table 3.5 Performance parameters of acidogenic reactor T1 (38 0C) during
period 2 (Day 65-112) ........................................................................................... 69
Table 3.6 Performance parameters of acidogenic reactor T2 (55 0C) during
period 2 (Day 65-112) ........................................................................................... 70
Table 3.7 Performance parameters of Polydigesters D1 and D3 (ambient
temperature) during Period 1 (Days 1-65) ............................................................ 75
Table 3.8 Performance parameters of Polydigesters D2 and D4 (ambient
temperature) during Period 1 (Days 1-65) ............................................................ 75
Table 3.9 Performance parameters of Polydigesters D1 and D3 (ambient
temperature) during Period 2 (Days 65-112) ....................................................... 76
Table 3.10 Performance parameters of polydigesters D2 and D4 (ambient
temperature) during period 2 (Days 65-112) ......................................................... 76
Table 3.11 Comparison of data between experiments 1 and 2 .............................. 92
Table 3.12 Characteristics of the raw piggery effluent from the Roseworthy
piggery ................................................................................................................... 93
Table 3.13 Characteristics of the raw piggery effluent data .................................. 94
Table 3.14 Data from experiments to be used in IBS model development ........... 96
Table 4.1 Fitted r2 values for CH4 ....................................................................... 103
Table 4.2 Refitted Growth and Death Rates for CH4 with r2 values ................... 103
Table 4.3 Fitted r2 values for TAN ...................................................................... 105
Table 4.4 Refitted Growth and Death Rates for TAN with r2 values .................. 105
Table 4.5 Fitted r2 values for Soluble P .............................................................. 107

xix
Table 4.6 Refitted Growth and Death Rates for Soluble P with r2 values .......... 107
Table 4.7 Comparison of modelled CH4 and measured CH4 data ....................... 109
Table 4.8 Comparison of modelled TAN and measured TAN data .................... 110
Table 4.9 Comparison of modelled P and measured P data ................................ 111
Table 5.1 Nutrient Concentrations in different stages of the IBS ....................... 141
Table 5.2 Nutrient and Plankton outputs from Central Institute of
Freshwater Aquaculture system .......................................................................... 146
Table 6.1 Parameters used in sensitivity analysis (Schladow and Hamilton
1997) .................................................................................................................... 155
Table 6.2 Parameters used in the sensitivity analysis study ................................ 156
Table 6.3 Comparison of percentage variation in input parameter and output
Chl-a .................................................................................................................... 170
Table 7.1 GLUE Calibration results for 10 random simulations ........................ 178
Table 7.2 Numerical Values of the Calibrated Parameters ................................. 180
Table 7.3 GLUE Calibration Results for 10 random simulations ....................... 182
Table 7.4 Numerical Values of the Calibrated Parameters ................................. 184

1
1 Introduction
1.1 Integrated Biosystems
Integrated Biosystems (IBS) connect different food production activities
with other operations such as waste treatment and fuel generation
(Warburton, Ramage et al. 2002). An Integrated Biosystem is a continuous
closed loop or open system comprising of production and consumption
where outputs from one operation become inputs to another. This enables
the reuse of resources and minimises environmental impact.
“Integrare” is a latin verb which means to make whole and to complete by
adding parts or to combine parts into a whole. The concept of IBS is not
new. There has been evidence found on an ancient Egyptian painting of
about 2000B.C. that seems to present an IBS for pond aquaculture where
nutrients in the pond water were used to cultivate flowers, vegetables and
fruits. Other early civilisations, such as those in China and Mexico have also
developed integrated farming systems that are unique to their regions. IBS is
widely practised in China today for the production of food, fuel and
aquaculture species (Zhang 1990).
Examples of different kinds of IBS are described below (Warburton,
Ramage et al. 2002):
1. Simple systems: e.g. livestock manure used directly as a fertiliser in
agriculture.
2. Intermediate systems: e.g. organic waste → compost → agriculture.
3. Closed systems: e.g. livestock manure → fodder crop → feed
→livestock.
4. Fuel generation: e.g. organic waste → biogas.
5. Nutrient stripping and bioconversion: e.g. wastewater effluent from
sewage treatment or livestock is stored into lagoons and used to grow
floating aquatic plants (e.g. duckweed). Duckweed consumes the
nutrients from the effluent and in turn reduces the high nutrient load to a

2
level where the water can be used for crop irrigation. The duckweed is
also harvested and used as feed for livestock and fish.
6. Water reuse: e.g. recycling dams allow the same water to be used for
growing several crops.
7. Industrial by-products: e.g. fermentation of grain (to produce beer,
spirits, biofuels) produces organic residues, heat and carbon dioxide.
The organic residues can be used in aquaculture to increase the
production of cultured fish, the carbon dioxide can be used for aerated
drink production, and both heat and carbon dioxide can be used as a
catalyst to improve growing conditions in hydroponic greenhouses.
8. Settlements: e.g. integration of waste treatment systems with housing
(septic tanks).
The IBS concept consists of three basic principles:
1. Use all the available wastes and organic materials instead of discarding
them.
2. Obtain at least one or more valuable products from the wastes.
3. Develop a closed loop continuous system using organisms through
biological processes for nutrient and wastewater recycling so that the
resources are completely utilised and there is no waste disposal.
The nutrient and material flows within an IBS are shown in Figure 1.1
summarising the IBS concept discussed previously. The input which is
essentially wastewater effluent enters the IBS process where it is subject to
a first biological process (e.g. microbial activity). As a result of this, a
product and by product are formed. The by product is used as an input to the
subsequent biological process. After the second biological process, a
product and by product are formed. The by product can be recycled back
into the first biological process thus completing a continuous cycle or it can
flow on to further biological processes before completing the loop. At each
stage the product formed can be put to effective use (e.g. energy generation
etc).

3
Figure 1.1 Schematic of material flow within an Integrated Biosystems
The advantage of using IBS is that it allows the resources to be converted,
recycled and re-used and offers many opportunities for increased efficiency,
enhanced profit and to develop novel solutions for effective waste
management.
With the advent of the 21st century, global concerns have been raised related
to increase in population; and diminishing resources like fuel, water and
minerals. These have direct impact on sustainable development and
maintaining quality of life. The IBS approach can reduce the dependence on
fossil fuels. Biogas obtained can be used as an alternative fuel for generating
energy (e.g. electricity). Recycling wastewater is an effective method to
BIOLOGICAL
ACTIVITY
BIOLOGICAL
ACTIVITY
INPUT
PRODUCT
PRODUCT
INPUT
BY PRODUCT
BY PRODUCT
INPUT

4
reuse dwindling water resources and utilise it in aquaculture, agriculture and
horticulture for the benefit of everyone.
The IBS should be flexible enough to be used by both an ordinary farmer
for simple agricultural systems or by a large scale processing industry for
complex systems (e.g. abattoir, winery waste treatment) (Warburton 2001).
1.2 IBS Project
The pilot scale IBS project was set up at the Roseworthy campus of the
University of Adelaide, South Australia. The IBS consists of the modules of
anaerobic digestion and the integrated aquaculture system comprising of the
bioconversion stages of microalgae, zooplankton and fish. Raw piggery
wastewater was used as the effluent feedstock for the two-stage anaerobic
digestion system where the raw piggery effluent was first fed to a first stage
thermophilic (acidogenic) anaerobic digester followed by a second stage
mesophilic (methanogenic) anaerobic digester. The digested piggery
effluent rich in nutrients was fed to the integrated aquaculture system. The
nutrients released in the digested effluent were utilised by microalgae to
grow and multiply. The microalgae served as food source for the
zooplankton which in turn served as food source for fish. The valuable end
products from the IBS were biogas (which could be used for energy),
aquaculture fish production and recycled water (which could be used for
agriculture, horticulture etc.). A flow diagram of the IBS project is shown in
Figure 1.2 to gain a better understanding into the IBS process.

5
Figure 1.2 Schematic of the IBS Project
LIVESTOCK
(ORGANIC
WASTE)
RECYCLED
WATER
THERMOPHILIC
ANAEROBIC DIGESTION
MESOPHILIC ANAEROBIC
DIGESTION
MICROALGAE
ZOOPLANKTON FISH
BIOGAS

6
1.3 Aim of the Research
The aim of this research was to develop a mathematical model for
Integrated Biosystems (IBS) which could be used as a tool for operational
management and process control and to also investigate the suitability of
aquaculture model DYRESM CAEDYM for modelling an IBS. Previous
modelling studies on IBS have been restricted to modelling the individual
IBS units rather than the whole system.
The major contribution from this research was to elucidate the key
parameters required to simulate the integrated IBS model. The data was
collected through a series of rigorous experimentation for both the anaerobic
digestion and aquaculture modules of the IBS. The data obtained was used
to parameterise a coupled anaerobic digestion-hydrodynamic ecological
model. The resulting model adequately simulated key processes within the
IBS, which was further improved with a novel auto calibration algorithm.
The primary contribution of this work has been to develop an automatic
parameter calibration for the aquaculture component of the model.
Parameter calibration in aquaculture models has been time consuming as it
is basically a “trial and error” procedure. This thesis presents a significant
contribution in this area.
Summarising, the objectives of this PhD research are to
1) Develop a mathematical model for the anaerobic digestion system
using microbial kinetics. The outputs of this model can be used as an
input to the aquaculture model.
2) Use DYRESM CAEDYM as an existing model to test its suitability
for modelling the aquaculture component of the IBS comprising of
bioconversion stages of algae, zooplankton and fish. This model will
be modified to suit the operating conditions and dimensions
(morphometry) of the IBS.
3) Conduct a sensitivity analysis on the modified DYRESM CAEDYM
aquaculture model.

7
4) Develop a computer program for automated calibration and
parameter estimation for the aquaculture model.
1.4 Background
The 21st century has exhibited global concerns related to increasing
population, diminishing fossil energy, water, land resources and higher
levels of pollution. These have multiple effects on sustainable development
and maintaining the quality of life in the future. The IBS approach can
reduce the need for fossil fuel. Biogas technology will play a unique role as
it provides energy, nutrients and better sanitation. Large volumes of biogas
can provide electricity to the grid, local communities and industries
(Mansson 1998; Kranert M. 2000).
Wastewater (effluent) from livestock is not a pollutant necessarily but a
nutrient source which can be recycled. There through integrated farming
practices has been evidence of recycling effluent through agriculture,
horticulture and aquaculture in several Asian countries (Kumar and Crips
2012). Aquaculture is common in many developing countries and has been
adapted as a technology for treatment of wastewater (Islam 1996). Examples
are sewage fed fish culture in Munich, Germany and the “bheries” in
Kolkata, India (Kumar 2002). Integrated farming systems with aquaculture
as a module differ from the traditional extensive and intensive farming
systems as aquaculture is used as a tool for recycling wastewater and
recovering nutrients.
A major concern in the 21st century is environmental pollution from solid
wastes and wastewaters from mega-cities, intensive animal farms and
industries. The ‘waste’ which provides income through producing a
valuable product, in effect, becomes a ‘resource’ (Crips and Kumar 2003).
Nitrogen (N) and phosphorus (P) are resources and their bioavailability can
be optimised through aerobic or anaerobic digestion processes. The process
simply allows recycling the nutrient and water, prevents aquatic pollution

8
and produces valuable end products. The IBS approach can have a
multipurpose role in sustainable environmental protection as it cleans the
environment and can generate products of economic value at the same time.
The amount of wastewater generated has been increasing over the years
with the increase in human population. Large amounts of domestic sewage,
industrial effluents and solid wastes are being generated everyday which has
made treatment difficult. There are different processes of wastewater
treatment e.g. conventional activated sludge and trickling filter methods,
oxidation/waste stabilisation ponds, aerated lagoons and variation in
anaerobic treatment systems (Gopakumar, Ayyappan et al. 2000). However
while most of these are energy-based treatment processes, only a few of
them lead to any resource recovery e.g. root zone treatment, wetland system,
aquatic macrophyte and aquaculture. Knowledge of macrophytes being used
as an effective agent for removing nutrients from wastewater led to the
concept of treatment of domestic sewage through aquaculture. A more
developed process called the Up Flow Anaerobic Sludge Blanket (UASB)
has also been developed (Pearson 1987; Curtis 1992; Pearson 1996).
Wastewater treatment usually involves additional costs (e.g. energy usage).
If the treatment itself produces income, prevents pollution and complies
with the environmental standards, it increases the profitability and the
sustainability of the industry (Williams, Biswas et al. 2007). While treating
the organic waste in the sewage, aquaculture products (fish), aquatic plants
and agricultural products can be produced. The introduction of aquaculture
into the wastewater treatment industry to remove nutrients and release clean
effluent has proved to be successful in many different countries (Edwards
and Pullin 1990). Some examples of these are
• Pig-biogas-duckweed-cassava IBS in Vietnam
• Brewery wastes-duck-insect larvae-aquatic plants-earthworm IBS in
Samoa
• Compost toilet and graywater garden system in Fiji
• St. Petersburg Eco-House, Russia
• Pozo Verde Farm in Colombia

9
• Sewage-duckweed-fish-banana IBS in Bangladesh
• Rice-flower-fish IBS in China
1.5 Commercial Scale Integrated Biosystems
The uniqueness of an IBS is that it is capable of handling large volumes of
wastewater from a variety of industries. Wastewater treatment in an IBS
could be through a series of biological processes complementing each other.
In 2005, the South Australian Research & Development Institute (SARDI)
and the Environmental Biotechnology Cooperative Research Centre
(EBCRC) commenced a project, entitled “Commercial scale integrated
biosystems for organic waste and wastewater treatment for the livestock and
food processing industries”, for which this research forms a part. This
project was proposed to be set up at Roseworthy Campus, The University of
Adelaide, where there is a commercial pig and poultry unit, and
considerable land available.
Raw piggery effluent was chosen as the source of wastewater which is
available from the Roseworthy piggery. Pigs have a high efficiency of food
conversion and are capable of reproducing and sustaining themselves by
feeding on farm wastes and kitchen refuse (Gopakumar, Ayyappan et al.
2000). Pig waste has certain advantages over cow, horse, sheep and goat
waste for aquaculture because pigs have a limited capability to consume
roughage. As a result their excreta contain lesser amounts of cellulose,
hemicellulose and lignin which are difficult to decompose, as these
materials are not a large part of the feed mix for pigs (Flachowsky and
Hennig 1990). These organic compounds form a blanket at the bottom of the
pond, which becomes a maintenance problem. The waste produced by 20–
30 pigs per year is equivalent to 1 tonne of ammonium sulphate supplied to
the soil (Kumar and Sierp 2003).
The treatment of raw piggery effluent is planned through the following
stages of an IBS as shown in Figure 1.3.

10
1.5.1 Anaerobic Digestion
Anaerobic digestion of the raw piggery effluent is proposed to be conducted
in two stages. A primary thermophilic treatment conducted at 500 C
acidifies the raw piggery effluent. The purpose of this stage is to kill the
pathogens present. A secondary mesophilic treatment conducted at ambient
temperature generates methane rich biogas and effluent rich in nitrogen (N)
and phosphorus (P) which is in bio available forms to be utilised in culturing
micro algae, zooplankton and fish downstream.
The functions of the anaerobic digesters within the IBS project were to
• Receive, hold and anaerobically digest piggery effluent received from
storage sump;
• Generate biogas rich in methane during the anaerobic digestion process;
• Produce nutrients (N & P) in bioavailable form to be used in subsequent
aquaculture stages of the IBS downstream. N will be present in the form
of total ammonia nitrogen (TAN) (approximately 80% of total nitrogen,
TN) and P will be present in the form of soluble phosphorus (SP)
(approximately 70% of total phosphorus).
1.5.2 Microalgal Ponds
The functions of the algal ponds are to
• Receive and hold effluent from anaerobic digesters and reduce
concentrations of nutrients (N & P) by algal growth.
• Maximise algal growth.

11
1.5.3 Zooplankton and Fish Ponds
The functions of the zooplankton and fish ponds were to
• Receive, hold effluent from algal ponds and forward treated water from
these ponds to the horticulture facility.
• Act as an aerated pond and support zooplankton and fish growth.
• Convert nutrient load within algae into zooplankton and fish growth.
Figure 1.3 Flowchart of the IBS process
Raw
Piggery
Effluent
Two- Stage Anaerobic
Digestion
Microalgae
Zooplankton
Fish
Clean
Water
Raw
Piggery
Effluent

12
1.6 Organisation of the Thesis
A literature review is presented in Chapter 2 which describes different
mathematical models developed for both anaerobic digestion and
aquaculture systems. Chapter 2 provides background knowledge and
presents the research required that motivated this study. An investigation on
the existence of integrated models for the IBS is conducted. The literature
review also provides the different techniques used by researchers to develop
algorithms for automated parameter estimation and calibration in
wastewater modelling.
Chapter 3 presents the experimental methods employed to collect data from
the laboratory scale anaerobic digestion system. Experiments were also
conducted using the pilot scale Integrated Indoor Aquaculture System to
gain a better understanding into the bioconversion capability of microalgae
using anaerobically digested piggery effluent as the wastewater source. In
addition to this, Chapter 3 also describes the procedures employed for set up
and commissioning of the Pilot Scale Anaerobic Digestion and Integrated
Aquaculture Systems required for conducting future experiments on a pilot
plant scale.
The field data collected from the anaerobic digestion experiments were
assessed in Chapter 4 where a mathematical model was developed using
kinetic equations from literature.
Chapter 5 introduces the numerical model DYRESM (DYnamic REservoir
Simulation Model) CAEDYM (Computational Aquatic Ecological and
Dynamic Model) and the subsequent improvements to the source code that
were necessary to model the IBS.
A sensitivity analysis conducted on selected parameters in DYRESM
CAEDYM is presented in Chapter 6.
Chapter 7 describes the development of an algorithm using FORTRAN 90
for automated parameter calibration for the aquaculture model.

13
The thesis is concluded in Chapter 8 with summary and conclusions.
Recommendations and directions for further research are also presented.
1.7 Research Program
The sequence and achievements of the research undertaken is summarised
below.
• Literature Review (Chapter 2).
• Laboratory Work for the Anaerobic Digestion system (Chapter 3).
• Laboratory Work for bioconversion of algae (Chapter 3).
• Data Collection and development of a mathematical model for the
Anaerobic Digestion component of the IBS (Chapter 4).
• Application and modification of the numerical model DYRESM
CAEDYM to model the aquaculture stages of the IBS consisting of
the bioconversion stages of microalgae, zooplankton and fish
(Chapter 5).
• Sensitivity analysis on selected critical parameters in DYRESM
CAEDYM (Chapter 6).
• Development and incorporation of an automated parameter
estimation and calibration script in FORTRAN 90 for calibrating
parameters in DYRESM CAEDYM (Chapter 7).

14
2 Literature Review The main objectives of this research were to
1) develop a simple mathematical model for the anaerobic digestion
system,
2) use numerical modelling techniques to model the aquaculture
component of the IBS using an existing modelling software and to
test its suitability for modelling the IBS,
3) develop an algorithm for automated parameter estimation and
calibration for the aquaculture model.
This chapter is divided into three sections. The first two sections provide a
brief review of the different mathematical models developed for Anaerobic
Digestion, Integrated Aquaculture and the IBS processes. The last section
deals with the parameter calibration technique used for setting up an
automated parameter calibration as part of this PhD Research.
For this particular study these components form part of the IBS.
• Two Stage Anaerobic Digestion
• Bioconversion of algae
• Bioconversion of zooplankton
• Bioconversion of fish
2.1 Anaerobic Digestion
Anaerobic digestion is a complex biochemical process in which organic
compounds are mineralized to biogas, primarily consisting of methane and
carbon dioxide, through a series of reactions mediated by several groups of
micro organisms in the absence of oxygen. Anaerobic digestion has been
used for waste treatment (Chen 1983; Yu, Wilson et al. 1998; Parker 2005;
Lee, Suh et al. 2009; Ramirez, Volcke et al. 2009; Santos, López et al.
2010; Appels, Lauwers et al. 2011; Rajagopal, Rousseau et al. 2011; Yu,
Zhao et al. 2012), but at present the focus is also on generating energy from
the method and treating organic, municipal and food processing wastes.

15
2.1.1 Hydrolysis and Fermentation
Hydrolysis is the first step in the anaerobic digestion of most insoluble
organic wastes. It breaks down complex organic compounds (e.g.
carbohydrates, fats and proteins) into their monomers (simple sugars like
glucose). This process of breakdown of complex organic matter is
performed by extracellular enzymes, which are produced by both facultative
and anaerobic bacteria. The monomers produced as a result of hydrolysis
are then fermented to volatile fatty acids (VFA) like acetic, propionic,
butyric, valeric acids and alcohols, CO2, H2 and some lactic acid.
The significance of hydrolysis is that it is considered to be the rate limiting
step during anaerobic digestion of insoluble organic compounds (Eastman
and Ferguson 1981; Noike and Endo 1985; Yasui, Goel et al. 2008). The
rate limiting step or rate determining step (RDS) is the slowest step
occurring in a reaction. Temperature and pH are two factors affecting
hydrolysis. Hydrolysis rate of carbohydrates is generally faster than that of
proteins (Yu and Zheng 2003).
Carbohydrates such as starch and sugars are most commonly hydrolysed by
Bacteriodes, Clostridia, Butyrivibrio, Selemonas, Micrococcus and
Lactobacillus (Huang 1975; Poulsen and Peterson 1985; Budiastuti 2004;
Myint and Nirmalakhandan 2006; Myint, Nirmalakhandan et al. 2006).
Sugars are common energy sources for fermentative microorganisms.
Generally pyruvate is produced during the cell during this breakdown.
Pyruvate is then metabolized primarily to acetate, formate, hydrogen and
carbon dioxide. Other products such as propionate, butyrate, succinate,
ethanol and lactate can also be found (Thauer and Jungermann 1977).
Lactic acid is the most common product in the fermentation of sugars. In
natural fermentation processes homofermentative bacteria such as
Lactobacillus curratus and Lactobacillus plantarum initiate acidification of
the medium according to the following reaction (Lin and Sato 1986)
+− +→ HCHOHCOOCHOHC 22 36126 Eq. 2.1

16
Heterofermentative bacteria such as Lactobacillus buchneri and
Lactobacillus brevis convert glucose according to the following reaction
(Lin and Sato 1986)
+− +++→ HCOOHCHCHCHOHCOOCHOHC 22336126 2 Eq. 2.2
2.1.2 Acetogenesis and Homoacetogenesis
The fermentation products from hydrolysis e.g. propionic and butyric acids
and ethanol need to be converted to a simpler product, i.e. acetic acid before
being utilised by the methanogenic bacteria. The bacteria responsible for
this conversion are known as acetogenic bacteria or hydrogen producing
bacteria (Le Hyaric, Canler et al. 2010; Donoso-Bravo, Mailier et al. 2011;
Madsen, Holm-Nielsen et al. 2011; Salomoni, Caputo et al. 2011; Zhang,
Lee et al. 2011). The common alcohols and fatty acid degrading acetogens
are Acetobacterium, Acetobacter, Syntrophobacter, Syntrophomonas and
some Desulfovibrio species (McInerney and Bryant 1981; Budiastuti 2004).
Another group of acetogens known as H2-acetogenic and homoacetogenic
bacteria convert H2 and CO2 to acetate according to the reaction
OHCOOHCHHCO 2322 242 +→+ Eq. 2.3
Acetobacterium woodee and Clostridium aceticum are bacterial species
capable of performing the above reaction (Budiastuti 2004).
2.1.3 Methanogenesis
Methanogenesis is the final step in anaerobic digestion to produce methane
(CH4) and carbon dioxide (CO2) from acetate and hydrogen produced in
acetogenesis step. In all anaerobic digestion processes, methanogenesis is
carried out by methanogenic bacteria which are sensitive to oxygen and pH
(Zehnder 1978; Taconi, Zappi et al. 2008). Methanosarcina and
Methanotrix are two bacterial groups which can utilise acetic acid and are
found in abundance in anaerobic digesters (Zehnder 1978; Zhang, Lee et al.
2011).

17
There are two types of methanogenic bacteria i.e. aceticlastic methanogens
and H2 utilising methanogens (Zehnder 1978). The function of aceticlastic
methanogenic bacteria is carbon removal and they play an important role in
controlling the pH during the fermentation process by the removal of acetate
to form CO2 and CH4 (Mosey 1983). They are responsible for 60-70% of
methane produced in anaerobic digesters (McInerney and Bryant 1981)
according to the reaction given below
−− +→+ 3423 HCOCHOHCOOCH Eq. 2.4
The H2-utilising methanogenic bacteria are responsible for 30% of the total
methane produced in anaerobic digesters (McInerney and Bryant 1981). The
process involves the reduction of CO2 by H2 (McInerney and Bryant 1981)
according to the reaction
−+− +→++ 3422 34 HCOCHOHHHCO Eq. 2.5

18
The processes involved in anaerobic digestion described above are
summarised below in Figure 2.1.
Figure 2.1 Flow Chart of processes in anaerobic digestion (Pullammanappallil, Chynoweth et al. 2001)
CH4
Methanogenesis
Acetate H2 + CO2 Acetate
Fermentation Acetogenesis
Complex Polymers
Monomers
H2 + CO2 Acetate Propionate,
Butyrate
Hydrolysis
Fermentation Fermentative
Bacteria
H2 producing
fatty acid
oxidizing
Bacteria Methanogens
Methanogens Methanogens

19
Complex organic compounds are hydrolysed to form simpler polymers
(monomers). The monomers are further broken down into volatile fatty
acids (acetate, propionate, butyrate) and H2 and CO2 by fermentative
bacteria. Methanogenic bacteria further break down the volatile fatty acids
to produce biogas.
Anaerobic digestion is commonly used for effluent and sewage treatment. In
developing countries, farm-based anaerobic digestion systems offer the
potential for cheap, low-cost energy for cooking and lighting facilities.
Anaerobic digestion techniques can help reduce the emission of greenhouse
gasses by replacement of fossil fuels. Improvement in anaerobic digestion
can be accomplished by multiple ways, some of which are optimisation of
the process conditions, pretreatment of input effluent and increase of
process temperature.
2.2 Mathematical Models in Anaerobic Digestion
Understanding and application of anaerobic treatment has made significant
progress in the past 30 years and numerous mathematical models have been
developed (Lyberatos and Skiadas 1999). The complexities of anaerobic
treatment and less experience with the process compared with its aerobic
counterpart are reasons for variations among models and lag in
standardisation of an anaerobic model. A review of previous models was
conducted to examine their applicability and inclusion of the significant
phenomena in anaerobic treatment. Some of the different approaches used
by researchers were:
• Models assuming substrate inhibited Monod kinetics of the
methanogens. e.g. (Graef and Andrews 1974; Hill and Barth 1977);
(Kleinstreuer and Powegha 1982); (Moletta, Verrier et al. 1986);
(Smith, Bordeaux et al. 1988)).
• Models where the influence of pH and volatile fatty acids is taken into
account. e.g. (Hill 1982); (Bryers 1985).

20
• Models where the process is primarily controlled by the hydrogen
concentration in the reactor. e.g. (Mosey 1983); (Pullammanappallil,
Owens et al. 1991); (Costello, Greenfield et al. 1991a); (Costello,
Greenfield et al. 1991b).
• Complex models assuming inhibition of compounds. e.g. (Angelidaki,
Ellegaard et al. 1993); (Siegrist, Renggli et al. 1993).
The first dynamic model for an anaerobic process was developed by
Andrews (Andrews 1969). A major limitation of this model was that the
pH was assumed to be constant. Graef and Andrews (1974) removed this
limitation by considering physico chemical interactions among the liquid,
gas and biological phases. They used the modified version of Monod
kinetics to consider the inhibition of methane formers by non ionised
VFA. It was assumed that the utilisation of acetic acid by methane formers
was rate limiting, as a result their model included only one group of
bacteria. The work of Graef and Andrews (Graef and Andrews 1974) was
considered by Hill and Barth (Hill and Barth 1977) to develop a model to
simulate anaerobic digestion of animal waste. A second bacterial
population was added to consider the VFA production by acid formers and
VFA utilization by methane formers. Particulate hydrolysis was also
incorporated into their model. They further modified the Monod
expression to include inhibition of methane formers by both ammonia and
nitrogen and VFA. Their model predicts general trends of anaerobic
digestion of animal manure.
An extensive model that considered the biological phase of anaerobic
digestion of glucose has been developed (Mosey 1983). Two important
advancements in the model were consideration of
• four populations of bacteria
• role of hydrogen gas in the formation of intermediate products of
acetic, propionic and butyric acids, and in the conversion of
intermediate products of propionate and butyrate into acetic acid.

21
Harper and Pohland (Harper and Pohland 1986) and (Mosey 1983)
indicated that hydrogen concentration in the digester controls the course of
substrate utilisation. Numerous studies on analysis of the thermodynamics
of reactions in anaerobic digestion have been conducted (McInerney and
Bryant 1981), (McInerney and Beaty 1988), (Harper and Pohland 1986)
and (Thauer and Jungermann 1977). The effect of hydrogen partial
pressure on the production of acetic acid, propionic and butyric acids was
determined.
Mosey (Mosey 1983) investigated the regulatory role of hydrogen by
considering the metabolic pathways of the acid forming bacteria. He
developed a comprehensive mathematical model for the utilisation of
glucose via the Embden-Meyerhof pathway, which converts glucose to
pyruvate. Mosey (Mosey 1983) related the relative availability of NAD+
and NADH to ρH2 to develop mathematical expressions that can predict the
relative production of acetic, propionic and butyric acids from the
utilization of glucose.
Mosey (Mosey 1983) also considered the reaction stoichiometries to
determine the regulation factor for each VFA produced from the
biodegradation of carbohydrates, as well as for the utilisation of butyric
and propionic acids.
The work of Mosey (Mosey 1983) was a keystone in the development of
more advanced models. A number of researchers, (Rozzi and Merlini
1985), (Jones 1989) and (Jones & Hall 1989) have developed models,
based on four population model of (Mosey 1983) for the biological phase
and the model of (Graef and Andrews 1974) for the physio chemical
system. These models predict change of individual VFA species, pH, pH2,
and biogas production and composition as a function of time. (Costello,
Greenfield et al. 1991a), (Costello, Greenfield et al. 1991b) developed a
six population model with a major change of introducing lactic acid
bacteria into the model of (Mosey 1983).

22
The mathematical models mentioned above and their respective process
flowcharts are further described in Appendix A.
2.3 Primary and Secondary Production
Aquaculture is an ancient practice for rearing aquatic organisms (Stickney
2000). Algae, zooplankton, molluscs and fish are grown in natural or
experimental designed basins. Phytodepuration is an important practice used
to recover clean water from wastewater for re-use and its characteristic is to
depress eutrophication and pollution (Brix and Scheirup 1989; Di Termini,
Prassone et al. 2011). In this process water to be purified is put in ponds
with specific phytoplankton and plants, which are efficient in uptake of
nutrients, therefore acting as biological filters. As a result they naturally
purify the water.
In the scientific literature a few papers connected with the study of
modelling integrated aquaculture dynamics can be found which could be
due to the aquaculture system’s complexity and the difficulty to adequately
describe them. There are three different approaches to modelling primary
and secondary production of species (Hamilton and Schladow 1997).
1. The most common modelling approach has been exemplified by the
development and application of steady state, input-output models.
Generally nutrient concentrations are calculated from net inputs and
chlorophyll-a concentration is predicted by correlation with the
limiting nutrient, most often phosphorus. Factors that can affect
phytoplankton biomass, such as light, climate, biological interactions
and internal loading of nutrients are not considered. The assumption
that the lake is a continuously mixed system is very restrictive and
only applicable at certain times of the year, if at all. The
shortcomings of such approaches include an inability to make
predictions in the face of varying physical and biological conditions,
and a failure to offer insights into the determinants of changing
water quality (Hamilton and Schladow 1997).

23
2. The second approach referred to as ecological water quality
modelling, specifically addresses many of the biological and
chemical factors that are absent in the simple input-output models.
Such models represent ecological processes by time varying,
interdependent conservation equations, with rate coefficients that
require calibration (Di Toro, O' Connor et al. 1971; Jørgensen,
Kamp-Nielsen et al. 1975; Jørgensen, Mejer et al. 1978; Scavia
1980; Matsuoka, Goda et al. 1986; Miyanaga 1986). The physical
processes of transport and mixing within the water body have
generally been oversimplified. Interactions between physical
processes and the biological and chemical processes described by
these models are poorly represented. The predictive abilities of these
models are compromised (Schladow and Hamilton 1997).
3. The third approach has been the extension of hydrodynamic models
to include water quality components, either by combination with
simple input-output models or more recently with ecological models.
The most common approach for the hydrodynamics has been to use
a one dimensional (1D) model, with retention of variables in the
vertical dimension. Process based hydrodynamic models have also
been identified and reported in literature (Stefan and Ford 1975;
Imberger, Patterson et al. 1978). These have been coupled with
ecological models giving the water quality models MINLAKE
(Riley and Stefan 1988) and CE-QUAL (USCE, 1986) respectively.
The models differ most in the extent to which the individual
processes are described (Schladow and Hamilton 1997).
Hydrodynamic and water quality models have become more abundant and
sophisticated since the early models which were developed in the 1970s
because of an increased demand for better models and an improvement in
the world of computing technology (Jørgensen, Jørgensen et al. 1981;
Jørgensen, Kamp-Nielsen et al. 1986; Jørgensen 1995).
The hydrodynamic models typically fall into one of the two categories; the
first are the relatively simple advective-diffusive models that require little

24
input data but are of low resolution such as MINLAKE (Riley and Stefan
1988) and AQUASIM (Gal, Imberger et al. 2003). The second group
include models based on a turbulence closure scheme in which the vertical
transport is related to the turbulent kinetic energy such as DYRESM
(Balistrieri, Tempel et al. 2006)
2.4 Models developed for Primary and Secondary
Production
The hydrodynamic model DYRESM and the ecological model CAEDYM
were combined to simulate the IBS ponds at Roseworthy, South Australia
(Chapter 5), conduct a sensitivity analysis on selected parameters of
DYRESM CAEDYM (Chapter 6) and develop an auto calibration algorithm
(Chapter 7). To model the fixed depth ponds of the IBS, modifications to the
source code of DYRESM CAEDYM were done which are described in
Chapter 5. The model parameters of reduced depth, climatic conditions,
nutrient inflows and outflows and pH were used as inputs to the model.
2.4.1 The Hydrodynamic Model DYRESM
DYRESM (DYnamic REservoir Simulation Model) is a one dimensional
hydrodynamics model for predicting the vertical distribution of temperature,
salinity and density in large water bodies satisfying the one-dimensional
approximation. The one-dimensional approximation is valid when the forces
acting to destabilise a water body (wind stress, surface cooling or plunging
inflows) do not act over prolonged periods of time. The dynamics of many
water bodies are well described using this approximation provided time
scales of extreme events such as storms and floods are not long. The model
can predict seasonal and inter-annual variability of water bodies as well as
sensitivity testing to long term changes in environmental factors. DYRESM
can either run in isolation for purely hydrodynamic studies or it can be
coupled to CAEDYM (Computational Aquatic Ecosystem Dynamics
Model) for investigations involving biological and/or chemical processes.
This computer model parameterises the important physical processes
leading to temporal changes in the temperature, salinity and density
distributions in water bodies. The model relies on data obtained from both

25
the field and from the laboratory. The model can quantifiably verify the
thermal characteristics in systems over time scales ranging from several
weeks to tens of years.
The purpose of the model is to provide a quantitative description of the
interactions that occur between physical and ecological processes, and the
water quality consequences of these interactions. The model comprises of
subroutines for phytoplankton production and loss, nutrient cycling and
dissolved oxygen dynamics. At each sub daily time step and in each model
layer, the set of equations that describe these processes is solved.
Daily - Loop
Figure 2.2 Schematic Flow Chart of DYRESM (Robson and Hamilton 2004)
Input daily data
Internal Mixing
Surface Heat Fluxes
Wind Mixing
Inflows
Withdrawals
Overflows
Daily Predictions

26
Figure 2.3 DYRESM simulation process (Imerito 2007)
Initial profile file
(.pro)
Parameters file
(.par)
Meteorological file
(.met)
Morphometry file
(.par)
Inflow file
(.inf)
Outflow file
(.wdr)
Mixer file – optional
(.mix)
Reference File
[NetCDF]
Simulation File
[NetCDF format]
DYRESM
Input Output

27
Flow chart of the DYRESM process is shown in Figure 2.2 and the
DYRESM simulation process is shown in Figure 2.3. The data from the
DYRESM input files e.g. meteorological, morphometry, inflow, outflow
and artificial mixing (optional) files is stored in a NetCDF reference file,
which along with the initial profile and parameters file forms a simulation
file in NetCDF format. This simulation file forms an input to the DYRESM
program.
DYRESM is based on a Lagrangian layer scheme and the layers are
adjusted to stay within user defined limits. The water body is modelled by a
series of horizontal layers of uniform property but variable thickness. The
layer positions change as inflow, outflow, evaporation and rainfall affect the
stored volume, and layer thicknesses change as the layers are moved
vertically to accommodate volume changes. An advantage of this layer
scheme is that it lends itself to the vertical structure of the water body. The
layers are counted from bottom to top.
Limits are set on the individual layer thicknesses and volumes. The upper
and lower limits are set to ensure that adequate resolution is achieved and
excessive number of layers is not used. To ensure that the layer structure is
stable, the individual layer densities are checked from top down. If the
upper layer density is higher than the layer immediately below, the two
layers are amalgamated and the layer properties conserved by appropriate
governing phenomenon. This process is repeated for all layers.
The density of water varies with the water temperature which causes
thermal stratification in the water body. When the temperature of the water
body decreases, the density usually decreases till the water temperature
reaches 4 0C. When salinity increases the density also increases. The density
of water (kg/m3) in a layer given its temperature (0C), salinity (psu),
pressure (bars) is given by the equation of state for density of salt water
(Imerito 2007).

28
( ) ( )
⎟⎟⎠
⎞⎜⎜⎝
⎛−
=
),,(1
0,,,,
PSTKPSTPST ρ
ρ Eq. 2.6
where ( ) 22/30,, DSCSBSAST +++=ρ Eq. 2.7
where A, B, C and D are polynomial functions of temperature.
To ensure stability of the layer structure, the individual layer densities are
calculated from top to bottom. If the upper layer density is higher than the
immediate layer below, the two layers are mixed and the layer properties are
conserved. This process is repeated for all layers culminating in a stable
structure.
The surface heat, mass and momentum exchange are the primary driving
mechanisms for DYRESM. The majority of the energy for heating, mixing
and stratifying the lake is obtained from these surface exchanges. The
surface exchanges include heating due to short wave radiation penetration
into the lake, and fluxes at the surface due to evaporation, sensible heat (i.e.
convection of heat from the water surface to the atmosphere), long wave
radiation and wind stress.
The density variations occurring in water bodies inhibits vertical motion
while horizontal density variations are minimised by lateral and longitudinal
convection, which occurs at a faster rate than vertical advection. DYRESM
provides a one-dimensional hydrodynamic platform for CAEDYM.

29
Atmospheric Surface
Long Wave Solar Long Wave Sensible and
Radiation Radiation Radiation Latent Heat
Euphotic
Depth
Attenuation
Figure 2.4 Surface energy flux exchanges
The meteorological data used by DYRESM can be in either daily or sub-
daily time step. Shortwave radiation Qsw is input directly into the model.
Shortwave radiation penetrates according to the Beer-Lambert law such that
( ) ( ) xswx
AettQttQ η−= 2121 ,, Eq. 2.8
where x is the depth below the water surface (measured down from the
surface of the reservoir)
ηA is the attenuation coefficient
The shortwave energy per unit area entering layer j through its upper face is
1−−=Δ jjj QQQ Eq. 2.9
or
( )jjA Zjj eQQ Δ−−=Δ
η1 Eq. 2.10
where, ηA is the attenuation coefficient of the jth layer of thickness ∆zj, and
1−−=Δ jjj zzz (z0 = 0; zN = H) Eq. 2.11

30
where, H is the depth of the water.
The penetrative short wave radiation has a wavelength of < 700 nm.
Radiation between 400-700 nm is defined as Photosynthetically Active
Radiation (PAR).
The net longwave radiation can be input into the model as incoming
longwave, net longwave or cloud cover data. The sensible heat loss from
the surface of the water body for the period ∆t can be calculated as (Imerito
2007)
( ) TTTUCCQ SaaPASS Δ−= ρ Eq. 2.12
where, CS is the sensible heat transfer coefficient for wind speed at 10 m
reference height above the water surface ( = 1.3 x 10-3)
ρA is the density of air in kg m-3
CP is the specific heat of air at constant pressure (= 1003 J kg-1K-1)
Ua is the wind speed at the standard reference height of 10 m in ms-1
Temperatures are either both in Celsius or both in Kelvin
The heat loss due to evaporation is given by (Imerito 2007)
( )[ ]⎭⎬⎫
⎩⎨⎧ Δ−= TTeeULC
PQ ssaaEALlh ρ
622.0,0min Eq. 2.13
where, P is the atmospheric pressure in hectopascals,
CL is the latent heat transfer coefficient (=1.3 x 10-3) for wind speed at a
reference height of 10 m,
ρA is the density of air in kg m-3,
LE is the latent heat of evaporation of water (=2.453 x 106 J kg-1),
Ua is the wind speed in ms-1 at a reference height of 10 m,
ea is the vapour pressure of air (hectopascals),
ea is the saturation vapour pressure at the water surface temperature TS
(hectopascals)

31
No condensation effects occurring in the numerical model, hence Qlh ≤ 0.
Thus, the total non penetrative energy density deposited in the surface layer
during the period ∆t is given by
lhswlwpennon QQQQ ++=− Eq. 2.14
The change in mass in the surface layer (layer number N) due to latent heat
flux is calculated as
( )v
NlhN L
AQlhM
−=Δ Eq. 2.15
where, AN is the surface area of the surface layer and LV is the latent heat of
vaporisation of water.
The change in surface layer mass due to rainfall is
( )d
hNNn NtRArainM Δ
=Δ ρ Eq. 2.16
where, Rh is the daily total rainfall in mm,
Nd is the number of seconds in a day
Therefore, the total mass change of the surface layer for the period ∆t is
( ) ( )rainMlhMM nnn Δ+Δ=Δ Eq. 2.17
The current model accounts for changes due to evaporation and
precipitation of pure water.
Surface Layer Mixing in DYRESM is undertaken with three specific
mechanisms: convective overturn, stirring and shear. The kinetic and
potential energies of the individual layers, starting at the surface layer, are

32
used to determine if the adjacent layers will mix. If there is sufficient energy
available, the layers will mix; any excess energy will be used to determine if
subsequent layers will mix. The amalgamation of layers ceases when there
is insufficient energy left, which is carried over to the next time step.
Turbulent Kinetic Energy (TKE) is introduced into the surface mixed layer
(SML) through the following processes:
Convective mixing: twAKE NNpconv Δ= −3*1ρη Eq. 2.18
Wind Stirring: tuAKE NNsstirr Δ= −3*1ρη Eq. 2.19
Shear Mixing: ( )211
1
2 −−
−
+= NN
NN
NNkshear UU
MMMM
KEη Eq. 2.20
where, ηp, ηs and ηk are efficiency coefficients,
ρN is the layer density,
Ai is the layer surface area,
Mi is the layer mass,
Ui is the layer speed,
∆t the model time – step
For water quality simulations using CAEDYM, bottom stress magnitudes
need to be calculated. This requires the epilimnion (top most layer in a
thermally stratified lake) and hypolimnion (dense, bottom layer of water in a
thermally stratified lake) water speeds to be determined, UE, and UH,
respectively. The epilimnion velocity, UE, is equal to the surface layer
velocity, UN, calculated as
( ) tzuU
NN Δ
Δ=
2*
Eq. 2.21

33
The velocity in the hypolimnion is calculated by
HEEHEH
EEH hhUhh
hhUU ≥<
⎩⎨⎧
= ;&; Eq. 2.22
where, hH and hE are the thicknesses of the hypolimnion and epilimnion
respectively.
Using the drag coefficient at the bottom, CD,bottom , the magnitudes of the
bottom stresses for the epilimnion and hypolimnion can be directly
calculated as
2, EEbottomDE UC ρτ = Eq. 2.23
2, HHbottomDH UC ρτ = Eq. 2.24
Inflows into DYRESM may be either surface (e.g. river or stream) or
subsurface (e.g. groundwater or pipe inflow) flows. The inflow to the water
body is modelled by inserting the volume into the layer of equal density,
taking into account the associated entrainment as the inflow passes through
the different layers. The layer structure is set within the limits set by the
user. Outflow occurs by the combination of withdrawals and overflow. The
required quantity of water is removed from the layer adjacent to the outlet.
If the amount of water exceeds the volume of the layer, then water is taken
from successive layers above the outlet until the required volume is
removed from the water column.
DYRESM can model two types of destratification systems-bubble plume
diffusers and surface mechanical mixers. Bubble plume diffusers consist of
a perforated pipe beneath the surface of water, through which compressed
air is pumped. As the air rises through the water column, it entrains fluid
and mixes the water column. DYRESM uses the simple buoyant plume
equations assuming that the plumes are circular and non-interacting.

34
The initial buoyancy flux is calculated as
71.0
⎟⎟⎠
⎞⎜⎜⎝
⎛=
diff
airairdiff PP
QQ Eq. 2.25
where, Qair is the free air flow rate of the compressor,
Pair is the air pressure (usually assumed as 101.3 kPa)
Pdiff is the pressure at the level of the diffuser due to both the atmosphere
and the depth of water,
Qdiff is the diffuser air flow rate.
The initial volumetric flow rate of entrained water is calculated as (Fisher,
List et al. 1979)
3/53/115
6 zBLbQ RPπ
α= Eq. 2.26
where, B is the buoyancy flux (m4s-3),
z is the bottom layer thickness (m),
b1 is a constant (= 4.7 (Fisher, List et al. 1979))
LR is the plume aspect ratio (assumed to be a constant of 0.1),
α is an entrainment coefficient (within the range 0.04-0.14).
The combined buoyancy flux of the air bubbles and entrained water is
calculated as
Pi
Piii QggQB ⎟⎟
⎠
⎞⎜⎜⎝
⎛ −−=
ρρρ
Eq. 2.27
where, ρi is the density of the current layer,
QP is the flow rate of the entrained volume.
The flow rate of the entrained volume in layer I is calculated as
( )1
3/51
3/53/115
6−
+−= − iPiiiRP QzzBLbQ πα Eq. 2.28

35
The development and process parameterisations of DYRESM have been
described extensively in literature (Imberger 1981; Hamilton, Schladow et
al. 1995; Hamilton and Schladow 1997; Schladow and Hamilton 1997;
Imerito 2007).
2.4.2 The Aquatic Ecological Model CAEDYM
CAEDYM (Hipsey, Romero et al. 2006) is an ecological model which
simulates the C, N, P, DO, and Si cycles along with inorganic suspended
solids, phytoplankton, and optional biotic compartments such as
zooplankton, fish, bacteria and others (Figure 2.5). CAEDYM can be linked
to hydrodynamic models like 1D DYRESM (Imerito 2007), DYRIM (a
quasi 2-D Lagrangian river floodplain model) and the 3D ELCOM (Hodges
and Dallimore 2001). The coupling between CAEDYM and the
hydrodynamic driver is dynamic.
Numerous optional biological and other state variables can also be
configured as per the requirements of the user. CAEDYM is more advanced
than traditional N-P-Z models as it can resolve species or group specific
ecological interactions. CAEDYM operates on any sub-daily time step and
is generally run at the same time interval as the hydrodynamic driver. The
user can specify if the same simulation is for freshwater, estuaries or coastal
water.
The major biogeochemical state variables in CAEDYM are given in Figure
2.5. The user can customize the input requirements by using a simple
configuration file. An input file for parameters can also be adjusted without
having to adjust the source code. However changes to the source code might
be required for specific variables of interest to the user.
Consequently CAEDYM was the most appropriate choice for simulating the
phytoplankton, zooplankton fish and nutrient dynamics in the aquaculture
section of the Integrated Biosystems. An extensive description of CAEDYM
can be found in (Hipsey, Romero et al. 2006).

36
Figure 2.5 Overview of CAEDYM state variables showing the water column, benthic and sediment components (Hipsey, Romero et al. 2006)

37
2.4.2.1 Phytoplankton (Algae) Model in CAEDYM
There are seven phytoplankton groups configurable within CAEDYM.
These are given in Table 2.1. Phytoplankton biomass is represented in terms
of chlorophyll-a (µg Chla L-1) or in terms of carbon (mg C L-1) depending
on the configuration set by the user. An overview of phytoplankton
dynamics as modelled by CAEDYM is shown in Figure 2.6.
Table 2.1 Description of the seven phytoplankton groups configurable within CAEDYM
Group Identifier Description
1 DINOF Dinoflagellates
2 CYANO Freshwater
Cyanobacteria
3 NODUL Marine/Estuarine
Cyanobacteria
4 CHLOR Chlorophytes
5 CRYPT Cryptophytes
6 MDIAT Marine/Estuarine
diatoms
7 FDIAT Freshwater
Diatoms

38
Figure 2.6 Schematic of phytoplankton dynamics within CAEDYM (Hipsey, Romero et al. 2006)
The phytoplankton groups modelled with CAEDYM were chlorophytes
represented by Scenedesmus, freshwater cyanobacteria represented by
Anabaena circinalis and freshwater diatoms represented by Nitzchia. Total
Respiration
DIC loss
only
Grazing
IC:IN:IP same as
Algae
Mortality
IC:IN:IP same as algae
Algae
IC:IN:IP fixed or variable
ISi fixed
Atmosphere
Water Column
Sediments
Growth
Resuspension
Settling
Vertical Migration
Excretion

39
chlorophyll-a concentration represents the biomass of each phytoplankton
group.
The generalized growth rate (µg) is a function of PAR, phosphorus,
nitrogen, silica, carbon and temperature and is represented by
( ) ( ) ( ) ( ) ( )[ ] ( )TfCfSifPfNfIfMAXg ,,,,minµµ = Eq. 2.29
where, f(I), f(N), f(P) and f(Si) represent limitation by PAR, nitrogen,
phosphorus and silica respectively,
f(C) is used for explicitly modelling the internal carbon,
f(T) is the temperature function
µMAX is the maximum growth rate at 200 C (day-1) in the absence of
significant limitation by light or nutrients.
Light limitation is modelled by the Webb Model (Hipsey, Romero et al.
2006), which ignores photo-inhibition and is given by
( ) ⎟⎟⎠
⎞⎜⎜⎝
⎛−
−= kII
eIf 1 Eq. 2.30
where, I is the incoming irradiance,
Ik is the light intensity at which the photosynthetic rate is numerically
equivalent to µMAX if light saturation behaviour was absent.
Michaelis-Menten kinetics is used to simulate nutrient limitation of
phytoplankton growth. For phosphorus limitation, the Michaelis-Menten
term is
( )PKPO
POPf+
=4
4 Eq. 2.31
where, KP is the half-saturation constant for the effect of phosphorus on the
growth rate.

40
For nitrogen limitation, the Michaelis-Menten term is
( )NKNONH
NONHNf
++
+=
34
34 Eq. 2.32
where, KN is the half-saturation constant for the effect of nitrogen on the
growth rate.
Maximum growth for phytoplankton occurs at optimum temperature Topt.
For T < TS (standard/ optimum temperature), the temperature limitation to
growth is calculated by
( ) 20−= TvTf Eq. 2.33
where , ν is a non-dimensional temperature multiplier.
For T > TS, the temperature limitation equation is
( ) ( ) bvvTf aTkT +−= −−20 Eq. 2.34
where, k, a and b are unknown which can be solved using the following
boundary conditions
T = TS ; f(T) = 1
T = Topt; ( ) 0=∂
∂
tTf
T = Tmax; f(T) = 0
The respiration of phytoplankton uses a lumped term that includes
respiration, excretion and natural mortality. The respiration term R is
calculated as
20−= T
rvkR Eq. 2.35

41
where, kr is a respiration rate coefficient.
The environment described for the IBS is freshwater, so there is no
limitation by salinity. The respiration term can be adjusted to account for
grazing. Numerous improvements and changes to the source code were
required for CAEDYM to effectively model the aquaculture stages of the
IBS. These are discussed in detail in Chapter 5.
DYRESM accounts for dissolved oxygen dynamics in the water column
according to the following processes
• Air/ water surface exchange
• Phytoplankton photosynthesis and respiration
• Sediment chemistry
• Nitrification
Atmospheric exchange is based on the model of Wanninkhof (1992) given
by,
( )waterairOO CCkF −=22
Eq. 2.36
where, FO2 is the flux of oxygen across the air – water boundary (g m-2 s-1)
kO2 is the oxygen transfer coefficient (ms-1)
Cwater is the oxygen concentration in the surface waters near the interface
(gm-3)
Cair is the concentration of oxygen in the air phase near the interface (gm-3)

42
2.4.2.2 Zooplankton Model in CAEDYM
Up to five generic zooplankton groups are configurable within CAEDYM
(state variables ZOOP1 – ZOOP5). All of these groups have the same
functionality (Figure 2.7).
Figure 2.7 Schematic of zooplankton dynamics within CAEDYM (Hipsey, Romero et al. 2006)
Grazing
Faecal pellets
to water
column as
POM
Faecal
pellets to
sediments
Respiration
Excretion
Mortality
Egestion
Zooplankton IC:IN:IP fixed
Atmosphere
Water Column
Sediments
Messy Feeding
food returned as POM
Predation

43
Zooplankton graze on phytoplankton, detritus, bacteria and on other
zooplankton. The total amount of carbon grazed can be calculated as
( ) ( ) ( )ZPOCBZAfTfgZG MAXC ,,,= Eq. 2.37
where, gmax is the grazing rate coefficient (mg consumed C (mg zooplankton
C)-1 day-1),
f(T) is a function for temperature dependence of grazing.
Predation of zooplankton is generally due to consumption by fish and by
other zooplankton. The total loss of zooplankton through grazing by fish
and other zooplankton is given by
( ) ( ) ( ) ( ) ( )zfFGWZ
N
ffXkz
ZGWZ
N
kzXz
GRZZ ZFfFGZZfZGZFZf
Z
F
Z
Z
,,,, ∑∑ +=
Eq. 2.38
where, X is the nutrient of interest: C, N, P.
The above formula sums over each zooplankton group k, and each fish
group f, in order to determine the total removal of zooplankton group.
Respiration is modelled by using a respiration rate coefficient, kzRz, for each
zooplankton class, z, and assigning the usual temperature dependence
( ) zT
zRzzDIC ZvkZR 20−= Eq. 2.39
Zooplankton losses through mortality, excretion (as liquid) and egestion
(faecal pellets) are modelled according to:
( ) ( )zCzEzzDOCL ZGkZE = Eq. 2.40
( ) ( ) zT
zMzzCzFzzPOCL ZkZGkZE 20−+= ν Eq. 2.41
where, kzEz and kzFz are fractions of the grazed food that are lost to excretion
and faecal pellets respectively,

44
kzMz is the mortality rate coefficient (day-1).
2.4.2.3 Fish Model in CAEDYM
Up to three generic fish groups are configurable within CAEDYM with state
variables FISH1-FISH3. All of these three groups have similar functionality,
although unlike zooplankton, fish are capable of recruiting from different
fish classes. Therefore the groups can be used to model several species or
different size/age classes of a single species. An overview of fish dynamics
as captured by CAEDYM is shown in Figure 2.8.
A literature review conducted did not provide any available work done
using DYRESM CAEDYM to model fish growth in ponds.
Fish can potentially graze on phytoplankton, detritus, on zooplankton, other
fish, on benthic macroinvertebrates and macroalgae and macrophytes. The
total amount of carbon grazed for each fish group is calculated as
( ) ( ) ( ) fFFDF
TFMAXfC FSGMPCGZBVFPOCZAfTfgFG
fff,,,,,,,,1=
Eq. 2.42
where, gMAX is the grazing rate coefficient (mg consumed C (mg fish C)-1
day-1),
𝑓!!!! 𝑇 is a function for temperature dependence of grazing
The fish model allows for predation by birds and by other fish groups. For
grazing by other fish, the relationship is given by
( ) ( ) ( )kfFGWFf
N
kxf
GRZF FFfFGFFf
f
Fx
f,,,0 ∑= Eq. 2.43
The total predatory loss is the sum of that by other fish and by birds.
Respiration is modelled by using a respiration rate coefficient, kfRf, for each
fish class, f, and assigning temperature dependence

45
( ) fT
fRffDIC FkFR 20−= ν Eq. 2.44
Fish losses through mortality, excretion and egestion are modelled
according to
( ) ( )fCfEffDOCL FGkFE = Eq. 2.45
EDOCL Ff( ) = k fEfGC Ff( ) Eq. 2.46
( ) ( ) ( )[ ] ( ) fDOff
TfMffCfFffPOCL SfDOfFkFGkFE 120 1++= −ν Eq. 2.47
where, kfEf and kfFf are fractions of the grazed food that are lost to excretion
and faecal pellets respectively,
kfMf is the mortality rate coefficient (day-1).

46
Figure 2.8 Schematic of fish dynamics within CAEDYM (Hipsey, Romero et al. 2006)
Grazing
Faecal pellets
to water
column as
POM
Faecal
pellets to
sediments
Respiration
Excretion
Mortality
Messy Feeding
food returned as POM
Predation
Egestion
Fish 1 IC:IN:IP fixed
Fish 2 IC:IN:IP fixed
Fish 3 IC:IN:IP fixed
Atmosphere
Water Column
Sediments

47
2.5 Techniques used in Water Quality Models Calibration
Mathematical model calibration is the process of determining the parameters
appearing in the equations of a model such that results produced by the
model agree closely with a set of measured data, in the context of the
selected objective function(s) (Ostfeld and Salomons 2005). The basic
approach in mathematical calibration is usually the same; an objective
function is designed to measure the agreement between measured (field)
data and the mathematical model simulated data for a particular set of
coefficients chosen by the user. The objective function ensures that small
values represent close agreement. The calibration process, which is a
constrained minimisation technique, adjusts the coefficient values within the
feasible domains to minimise the objective function.
It is an accepted fact that a mathematical model prediction should not be
deterministic, most probable representation, but should also explicitly
include an estimate of uncertainty. Mathematical models are characterised
by a certain degree of uncertainty, which results from both uncertainties in
modelled processes and observation errors, and the structural and numerical
errors of the mathematical model. Good modelling practice requires the
modeller to provide an evaluation of the confidence of the model predictions
(Ratto, Tarantola et al. 2001). This enables the researcher to assess the
uncertainties associated with the outcome (response) of the model itself.
Uncertainty Analysis (UA) and Sensitivity Analysis (SA) are pre requisites
for model building in any field where models are used. Model uncertainty
can be accounted for by the application of parametric uncertainty
methodologies and conditioning model predictions on observations.
Realistic assessment of these various sources of uncertainty is important for
science based decision making and will help direct resources towards model
structural improvements and uncertainty reduction (Blasone, Vrugt et al.
2008).
There has been an increase in number of methods in recent years to derive
meaningful uncertainty bounds on model predictions. Some of the methods
to represent model parameter, state and prediction uncertainty include

48
classical Bayesian (Kuczera and Parent 1998; Thiemann, Trosset et al.
2001; Vrugt, Gupta et al. 2003), pseudo-Bayesian (Beven and Binley 1992;
Freer, Beven et al. 1996), set-theoretic (Klepper, Scholten et al. 1991),
multiple criteria (Gupta, Sorooshian et al. 1998; Yapo, Gupta et al. 1998;
Boyle, Gupta et al. 2000; Madsen 2000; Madsen 2003; Vrugt, Gupta et al.
2003), sequential data assimilation (Moradkhani, Hsu et al. 2005; Vrugt,
Diks et al. 2005) and multi model averaging methods (Georgakakos, Seo et
al. 2004; Vrugt and Robinson 2007). These methods differ in their
assumptions and how the different sources of error are being treated.
Proper calibration and validation of a model require huge amounts of data
representing the various variables of interest. This involves a large number
of model parameters. Therefore, rigorous parameterisation and reduction of
the parameter space are essential to facilitate the calibration process and
make it more robust. Over parameterisation must be avoided to ensure a
higher degree of credibility to the model prediction (Andersen, Refsgaard et
al. 2001).
Refsgaard (1997) suggested assessing the parameter values from field data
as much as possible and fixing spatial patterns of parameters to simplify the
calibration process. Refsgaard (Refsgaard 1997) also suggested reduction in
the dimensionality of the parameter space by means of a sensitivity analysis
on the model output(s). By conducting a sensitivity analysis, the parameters
that do not influence the model response can be identified and fixed to their
prior values (Christiaens and Feyen 2001; Mertens, Madsen et al. 2004;
Muleta and Nicklow 2005). Sonnenborg used a method to decrease the
number of calibration parameters by fixing some of the pre-estimated
parameters by using a simpler model. He used the calibration results found
under steady state conditions to constrain the parameter space of a transient
model (Henriksen, Troldborg et al. 2003; Sonnenborg, Christensen et al.
2003). Refsgaard (1997) also pointed out that not only time validation but
also a multi site validation should be performed. If the model is used with
different discretisation scales, the dependency of the processes on the
different modelling scales should also be tested (Vázquez and Feyen 2007).

49
Data requirements can be an obstacle for a proper model validation because
spatially distributed time series observations of all the variables are rarely
available. This is one of the main reasons for distributed models being
calibrated and validated against discharge data (Andersen, Refsgaard et al.
2001; Engeland, Braud et al. 2006; McMichael, Hope et al. 2006). Madsen
proposed a framework for use of multiple criteria to measure the model
performance, which is crucial in calibration and validation of distributed
models. His method allowed modellers to include the different variables of
interest (multi-variable criteria), their spatial variability (multi-site criteria)
and the different error functions applied for evaluating the performance of
the simulated variables (multi-response criteria) (Madsen 2003). Inspite of
this, there hasn’t been much work done on integrating/ incorporating the
different methods together. Few studies have used multi-variable and multi-
site data to calibrate and validate an integrated, distributed model
(Refsgaard 1997; Madsen 2003; Vázquez and Feyen 2007). The last
component of this project aims at addressing this gap in research by
developing an auto calibration script for DYRESM CAEDYM which is a
multi-variable model presented in Chapter 7.
There are various difficulties in comparing uncertainty analysis techniques
in mathematical modelling of water bodies. Most techniques are different in
their philosophies with respect to parameter distribution, likelihood function
and /or objective function. Different concepts on objective functions lead to
different results depending on whether to maximise or minimise the
objective function. Due to the complexity of mathematical models running
hydrological processes, a higher number of model runs are needed to
calibrate and assess the uncertainty, due to the larger dimensionality of the
parameter space. There is a need to apply more efficient calibration and
uncertainty assessment procedures to minimise the computational burden.
2.5.1 GLUE
The Generalized Likelihood Uncertainty Estimation (GLUE) method
inspired by (Hornberger and Spear 1981) is a method for investigating the
uncertainties in the water resources and environmental modelling (Beven

50
and Binley 1992; Beven 1993). It has been widely studied by researchers
and applied in determining hydrological modelling uncertainty (Vogel,
Batchelder et al. ; Freer, Beven et al. 1996; Aronica, Hankin et al. 1998;
Cameron, Beven et al. 1999; Aronica, Bates et al. 2002; Freer, McMillan et
al. 2004; Montanari 2005; Heidari, Saghafian et al. 2006; Mantovan and
Todini 2006; McMichael, Hope et al. 2006). This method maps the
uncertainty in the modelling process onto the parameter space and operates
within the context of Monte Carlo (MC) analysis coupled with Bayesian
estimation and propagation of uncertainty. The GLUE technique calls for
rejecting the concept of a unique global optimum parameter set within some
particular model structure; instead it recognizes the acceptability within a
model structure of different parameter sets that are similarly good in
producing fit model predictions (Blasone, Vrugt et al. 2008). This concept is
defined as equifinality and is directly addressed by the computation of
different sets of parameters within a pseudo-Bayesian Monte Carlo
framework. The outputs of GLUE procedure are parameter distributions
within the limits of available observational data and associated uncertainty
bounds.
The GLUE methodology has been applied for uncertainty assessment in
environment modelling, including rainfall-runoff modelling (Beven and
Binley 1992; Freer, Beven et al. 1996; Lamb, Beven et al. 1998),
groundwater modelling (Feyen, Beven et al. 2001), unsaturated zone
modelling (Mertens, Madsen et al. 2004), flood inundation modelling
(Pappenberger, Beven et al. 2005; Pappenberger, Beven et al. 2007) and
distributed hydrological modelling (Mertens, Madsen et al. 2004;
McMichael, Hope et al. 2006; Choi and Beven 2007). The reason for GLUE
technique being implemented in different modelling scenarios is explained
by its conceptual simplicity, relative ease of implementation and use, and its
ability to handle different error structures and models without major
modifications to the method itself.
The GLUE procedure is based upon making a large number of runs of a
given model with different sets of factor values chosen randomly. The

51
acceptability of each run is evaluated against observed values, and if the
acceptability is below a certain threshold, the run is considered to be ‘non-
behavioural’ and that parameter combination is removed from further
analysis. Outputs from the retained runs are likelihood weighted and ranked
to form a cumulative distribution of the output variable. Different sets of
initial values, boundary conditions or model structures can also be
considered.
A GLUE analysis consists of the following three steps:
I. A large number of parameter sets are randomly sampled from the
distribution. A threshold value is assigned. Each parameter set is then
assessed as being either “behavioural” or “non behavioural” through a
comparison of the likelihood measure.
II. Each behavioural parameter set is given a “likelihood weight”
according to
( )
( )∑=
= N
kk
ii
L
Lw
1
θ
θ Eq. 2.48
where N is the number of behavioural parameter sets,
L(θ) is a generalized likelihood measure,
wi is the likelihood weight.
III. The most commonly used likelihood measure, Nash Sutcliffe
coefficient (NS) (Nash and Sutcliffe 1970; Freer, Beven et al. 1996;
Beven and Freer 2001), is calculated as
( )( )
∑
∑
=
−
=
⎟⎠⎞
⎜⎝⎛ −
−
−=n
tt
n
tt
Mt
t
i
i
ii
yy
yyNS
1
21
2
1θ
Eq. 2.49
where n is the number of observed data points

52
yti and ytiM (θ) represent the observation and model simulation with
parameters
θ at time ti respectively
𝑦 is the average value of the observations.
The Nash-Sutcliffe model efficiency coefficient (E or NS) is commonly
used to assess the predictive power of hydrological discharge models.
However, it can also be used to quantitatively describe the accuracy of
model outputs for other things than discharge (such as nutrient loadings,
temperature, concentrations etc). NS efficiencies can range from -∞ to 1. An
efficiency of 1 (E = 1) corresponds to a perfect match between model and
observations. An efficiency of 0 indicates that the model predictions are as
accurate as the mean of the observed data, whereas an efficiency less than
zero (-∞ < E < 0) occurs when the observed mean is a better predictor than
the model. Essentially, the closer the model efficiency is to 1, the more
accurate the model is. In the GLUE approach, factors are never considered
independently but as sets of values.
GLUE methodology was applied for setting up an auto calibration program
using DYRESM CAEDYM which is described in Chapter 7 in detail. The
reasons to choose GLUE over other calibration routines were the ease of
implementation of the GLUE routine with DYRESM CAEDYM. The
author is proficient in programming in FORTRAN 90, in which DYRESM
CAEDYM has also been coded, so implementation of GLUE subroutines
was not difficult. The use of other complex calibration software was
beyond the scope of this PhD research.
2.6 Summary
A study of modelling the bioconversion of phytoplankton, zooplankton and
fish has been reported in literature (Jørgensen, Jørgensen et al. 1981;
Schladow 1984; Hamilton and Schladow 1997; Schladow and Hamilton
1997; Bruce, Hamilton et al. 2006; Spillman, Imberger et al. 2007; Burger,
Hamilton et al. 2008; Trolle, Skovgaard et al. 2008). Schladow (1984) and
Trolle et al. (2008) discuss modelling phytoplankton growth in large water

53
bodies, while Bruce et al. (2006) discloses modelling of zooplankton growth
and nutrient dynamics in lakes. There is minimal literature available for
modelling fish using DYRESM CAEDYM. These models have been
developed in individual water bodies but not as part of an IBS where the
output of effluent from the algal pond forms an input to the zooplankton
pond and an output of effluent from the zooplankton pond forms an input to
the fish pond. The aquaculture model DYRESM CAEDYM has been
applied extensively to ponds of large depth to model phytoplankton,
zooplankton and fish (Schladow and Hamilton 1997; Trolle, Skovgaard et
al. 2008). Application of the DYRESM CAEDYM model to ponds of
shallow depth (~1 m) has not been reported in literature. Modelling of the
IBS ponds of 1m depth reported in this thesis will help close the gap in the
literature.
There is minimal literature available relating use of an auto calibration
routine to calibrate the parameters used in CAEDYM. The conventional
process of “trial and error” of calibrating parameters is tedious and
inefficient. More investigation is required to incorporate an automatic
calibration program into DYRESM CAEDYM to provide an efficient and
effective method of model parameter calibration and validation. This PhD
research will enable the use of a software routine to automatically calibrate
the parameters used in CAEDYM when the simulated data is modelled
against real time data.

54
3 Experimental Methods and Data
Collection
3.1 Introduction
The aim of this PhD research is to develop a mathematical model for a
commercial scale IBS and to test the suitability of DYRESM CAEDYM as
a control and management tool for the IBS. In order to achieve this,
microcosm experiments were conducted to develop further research into the
IBS. A pilot plant anaerobic digestion system and an aquaculture mesocosm
facility were developed which replicated the functioning of an IBS at a
laboratory level. After completing successful trials of the mesocosm facility,
a design of the commercial scale IBS was developed with additional
facilities for research and development. The commercial scale IBS however
could not be constructed at the time this research was conducted due to
certain administrative and financial restrictions which were beyond the
control of the researcher. Therefore experimental data collected from both
the pilot plant and the mesocosm facility (pilot IBS) were used for
developing a model and assessing its suitability for the commercial scale
IBS.
In this Chapter, Stage 1 experiments used for data collection and anaerobic
digestion are reported. The anaerobic digestion experiments were
undertaken to provide data which were the inputs for the development of the
aquaculture model (described in Chapter 5). Approximately 18 months
(from February 2005 – September 2006) were required for the set up and
installation of the two stage anaerobic digestion system and the Integrated
Aquaculture System. These systems were the backbone of the research work
to be discussed in later chapters.
This chapter is organised as follows.
• Section 3.2 describes the set up, installation and commissioning of
the pilot scale two stage anaerobic digestion system.

55
• Section 3.5 discusses the set up and installation of the pilot scale
Integrated Aquaculture system (mesocosm facility) for the
aquaculture stages of the IBS.
• Sections 3.6 and 3.7 describe the experiments conducted for the
bioconversion of algae.
• Finally section 3.8 describes the batch scale anaerobic digestion
experiments which generated data for input into the aquaculture
model.
3.2 Set up of the Pilot Scale Two-Stage Anaerobic
Digestion System
3.2.1 Equipment: Reactors and Digesters
A pilot scale two-stage anaerobic digestion system was set up in the
Breakwell laboratory at the Roseworthy Campus, The University of
Adelaide, South Australia. Active involvement in the construction and set
up of the anaerobic digestion system with the post doctoral researchers was
important as it provided an opportunity to understand the anaerobic
digestion stage of the IBS process. The main objectives of this pilot scale
system were to conduct experiments to maximise throughput of raw piggery
effluent, whilst maintaining high biogas generation.
Parameters measured from the experiments conducted were
• Chemical Oxygen Demand (COD)
• Soluble COD (SCOD)
• Total COD (TCOD)
• Total Kjeldahl Nitrogen (TKN) and Total Ammonia Nitrogen (TAN)
• Total and Soluble Phosphorus (TP, SP)
• Volatile Fatty Acids (VFA)
• Total and Volatile Solids (TS, VS)
• Organic Loading Rate (OLR)
• Biogas composition

56
• Biogas volume
• Temperature
• pH
These parameters were necessary to record as they formed an input to the
anaerobic digestion and aquaculture models. COD, TAN, TP, SP and biogas
volume were used for the development of the anaerobic digestion model
while TAN, SP and pH formed inputs and outputs from the aquaculture
model DYRESM CAEDYM.
The two-stage anaerobic digestion system which comprised a thermophilic
and mesophilic digesters is shown in Figure 3.2 and Figure 3.3.
Thermophilic Digesters: Two 200 L stainless steel tanks were used as
thermophilic digesters for the experiments. A microprocessor based
controller was used to ensure a constant operational temperature in the
reactors. A centrifugal pump was connected to the reactors to enable
pumping of piggery effluent and hourly recirculation of acidifying effluent
within the reactors. In addition, hourly temperatures and gas production in
the reactors were continuously monitored and recorded by a data logger
interfaced with a computer.
Mesophilic Digesters: Four 225 L plug flow type digesters (polydigesters)
were used to further digest the effluent under ambient conditions. These
digesters had an inlet and outlet located at each end for conveying the
influent and effluent respectively. The hourly temperatures and gas
production in the digesters were monitored and recorded hourly using a data
logger.
pH was measured in both digesters using a pH measuring device (Thermo
Scientific, Eutech Instruments, PC 650). Temperature was measured using
RTD thermocouples (Temperature Controls Pty. Ltd.). Gas production in
the digesters was recorded using positive displacement type gas meters
comprising a float switch and solenoid valve.
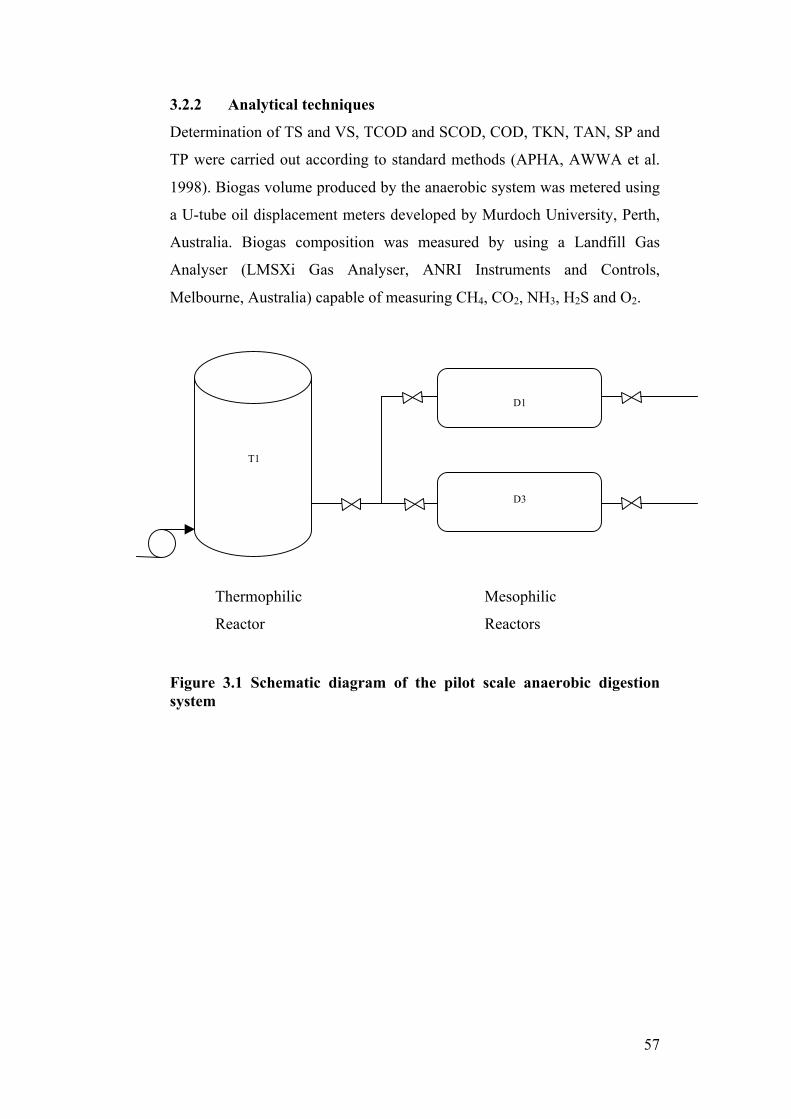
57
3.2.2 Analytical techniques
Determination of TS and VS, TCOD and SCOD, COD, TKN, TAN, SP and
TP were carried out according to standard methods (APHA, AWWA et al.
1998). Biogas volume produced by the anaerobic system was metered using
a U-tube oil displacement meters developed by Murdoch University, Perth,
Australia. Biogas composition was measured by using a Landfill Gas
Analyser (LMSXi Gas Analyser, ANRI Instruments and Controls,
Melbourne, Australia) capable of measuring CH4, CO2, NH3, H2S and O2.
Figure 3.1 Schematic diagram of the pilot scale anaerobic digestion system
T1
D1
D3
Thermophilic
Reactor
Mesophilic
Reactors

58
Figure 3.2 Thermophilic (stainless steel) digesters in the pilot scale
research facility
Figure 3.3 Mesophilic (polybag) digesters in the pilot scale research facility

59
3.3 Pilot Plant Stage 1 Experiments
The set up, installation and stage 1 experiments with the pilot scale
anaerobic digestion system were conducted from March 2005 to August
2005.
3.3.1 Acidogenesis
The acidification experiments using raw piggery effluent as the source of
wastewater were carried out in two 200 L reactors (T1 & T2) using 100 L as
the working volume. The reactors were set at a temperature of 38±2.5 0C
(T1) and 55±2.5 0C (T2) and were fed daily with raw piggery effluent based
on a constant hydraulic retention time of 6.7 days. The experiments were
continued for a period of 3-4 HRTs to achieve operational steady state and
stability. Due to the high variability in the content of solids in the piggery
effluent, the organic loading rate in the reactors fluctuated within a range of
1.5-5.5 g COD/L/day. Process performance was assessed by measurement
of pH (measured offline), biogas production and composition, total and
volatile solids reduction, and COD reduction. Nutrient quality was assessed
by analysing TKN, TAN, TP and SP in the acidified effluent in the reactors.
3.3.2 Methanogenesis
Anaerobic digestion of the pre-acidified reactor effluent was conducted in
two 225 L polybag digesters under ambient conditions. At start up, the
digesters were inoculated with 20% v/v anaerobic sludge (from Bolivar
Wastewater Treatment Plant, South Australia), 10% v/v of acidified effluent
and 80% mains water. Thereafter, the digesters were fed daily with the
acidified effluent from the acidogenic reactors based on a 15 day HRT. One
pair of the digesters was fed effluent from the reactor at 55 0C and the other
pair from the reactor at 38 0C. Measurements were made on pH,
temperature, gas volume and biogas composition. Nutrient analysis was
conducted on the digested effluent samples. The experiments ran for a
period of 28 days.

60
3.3.3 Results
Acidogenic Stage:
During the 28 days of operation, solids content in both the thermophilic (T1)
and mesophilic (T2) acidogenic reactor effluents steadily increased. This
was due to the daily addition of the piggery effluent into the original dilute
sample of effluent fed into the reactors during start up.
Effluent characteristics in the piggery effluent in T1 and T2 are shown in
Table 3.1.
Total and soluble COD levels in both the acidogenic reactors stabilised by
day 8 although there were constant fluctuations due to variations in the
COD of raw piggery effluent (12-36 g/L). By day 28, the SCOD
concentration in the effluent from T1 was higher than that in T2 indicating a
higher conversion of the COD to VFA. TKN in T1 and T2 were around
3400 and 3900 mg TKN/L respectively by day 8.
Table 3.1 Effluent characteristics in T1 and T2
Both T1 and T2 show a relatively stable pH during the 28 days of operation.
The TCOD decreased 47% while the SCOD increased 46% due to increased
solubilisation. TAN and SP increased by approximately 3 and 6 times
respectively due to conversion of VFA’s into bioavailable forms of nitrogen
and phosphorus.
Process Day pH SCOD TCOD TKN TAN SP TPg/L g/L mg/L mg/L mg/L mg/L
T1#(55#0C) 1 8.1 5 14 #, 500 15 #,8 8 6 13 3360 1223 46 8017 8 4.6 8.8 #, 1273 27 5628 7.4 7.3 7.4 #, 1667 100 #,
T2#(38#0C) 1 7.9 5 14 #, 500 15 #,8 7.6 7 11 3920 1284 49 7717 7.6 4.1 7.6 #, 1466 50 5328 7.3 7.2 9.7 #, 1658 114 ,#

61
Over the first 17 days of operation, both T1 and T2 showed a stable pH
during the acidification of piggery effluent as shown in Figure 3.4. A slight
decrease in pH in both reactors is observed for the remainder of the period
probably due to production of VFA’s. Slightly higher pH in T1 may be due
to marginally higher ammonia concentration from day 13 onwards. Higher
ammonia concentration may have resulted from degradation of the organic
nitrogen compounds at thermophilic temperatures.
Figure 3.4 pH profile during acidogenesis in T1 and T2 treating piggery effluent
By day 28, TAN and SP in both the reactors had stabilised at around 1700
mg/L and 100 mg PO4-P/L respectively (Figure 3.5 and Figure 3.6). The
sudden spike in the value of SP at the end of the experimental period could
be due to an increase in the rate of degradation of the effluent to a soluble
form of phosphorus.
0
2
4
6
8
10
0 10 20 30
pH
Day
pH
T1 (55 deg C)
T2 (38 deg C)
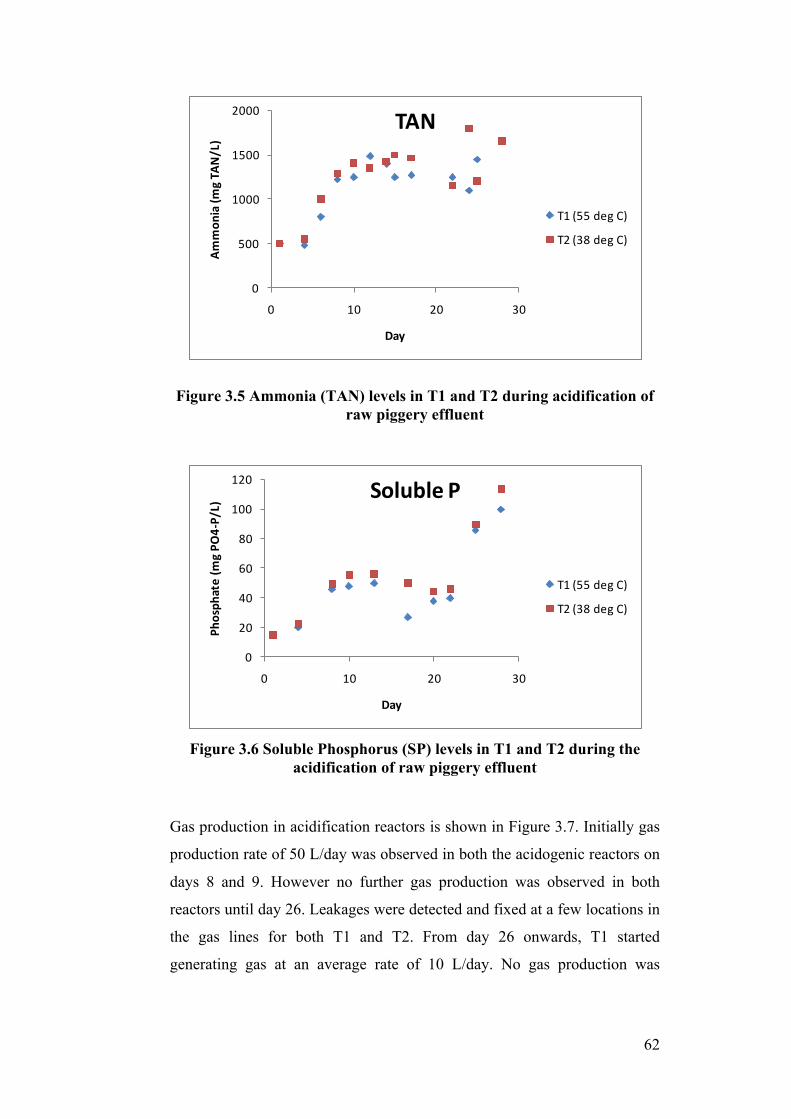
62
Figure 3.5 Ammonia (TAN) levels in T1 and T2 during acidification of raw piggery effluent
Figure 3.6 Soluble Phosphorus (SP) levels in T1 and T2 during the
acidification of raw piggery effluent
Gas production in acidification reactors is shown in Figure 3.7. Initially gas
production rate of 50 L/day was observed in both the acidogenic reactors on
days 8 and 9. However no further gas production was observed in both
reactors until day 26. Leakages were detected and fixed at a few locations in
the gas lines for both T1 and T2. From day 26 onwards, T1 started
generating gas at an average rate of 10 L/day. No gas production was
0
500
1000
1500
2000
0 10 20 30
Ammon
ia (m
g TA
N/L)
Day
TAN
T1 (55 deg C)
T2 (38 deg C)
0
20
40
60
80
100
120
0 10 20 30
Phosph
ate (m
g PO
4-‐P/L)
Day
Soluble P
T1 (55 deg C)
T2 (38 deg C)

63
observed in T2, so the gas production bags were replaced at the end of the
experimental run to ensure that there were no further gas leakages.
Figure 3.7 Gas production during anaerobic acidification of piggery effluent in T1 (55 0C) and T2 (38 0C)
(Source: Technical Advisory Committee Report (August 2005), EBCRC)
Methanogenic Stage:
Effluent characteristics in the two 200 L polydigesters treating the acidified
piggery effluent is presented in Table 3.2. TCOD levels in polydigesters 1
and 2 were 4.8 and 3.1 g/L. Assuming that the polydigesters were
approaching a steady state, polydigester 1 (receiving influent from T1)
showed a COD reduction of 28%, while polydigester 2 (receiving influent
from T2) showed a COD reduction of 61%. This result is supported by the
higher gas production rate measured in polydigester 2.

64
Table 3.2 Effluent characteristics in the polydigesters operating at ambient temperature
TAN and SP in both polydigesters stabilised around 1700 mg/L and 22 mg
PO4-P/L respectively. Gas production in polydigesters 1 and 2 is shown in
Figure 3.8. By day 28, polydigesters 1 and 2 produced biogas at an average
rate of 2.4 and 5.7 L/day and contained approximately 30% CO2. Lower gas
production rate in polydigester 1 (fed with the influent from T1) was
probably due to an inhibition arising from high concentrations of VFA or
maybe due to a higher degradation occurring in T1.
Process Day pH SCOD TCOD TKN TAN SPg/L g/L mg/L mg/L mg/L
Polydigester,1 1 7.6 0.4 1 280 477(Ambient
Temperature)25 7.5 2 4.8 1703 22
Polydigester,2 1 7.6 0.9 1.8 420 410(Ambient
Temperature)25 7.5 2.1 3.1 1678 23

65
Figure 3.8 Gas production during anaerobic digestion of acidified piggery effluent in polydigesters 1 and 2 operating under ambient
conditions
(Source: Technical Advisory Committee Report (August 2005), EBCRC)
3.4 Pilot Plant Stage 2 Experiments
The pilot plant stage 2 experiments were conducted over two time periods.
Period 1 (65 days) acidification experiments were conducted at 6.7 day
HRT at 38 0C and 55 0C. Period 2 (47 days) acidification experiments were
conducted at 4 day HRT at 38 0C and 55 0C. The acidified reactor effluent
was transferred to the methanogenic stage of digestion within the
mesophilic polydigesters during periods 1 and 2. The polydigesters were
operating at 23 day HRT at ambient temperature conditions. These
experiments were conducted from August 2005 to November 2005.
3.4.1 Methods
Acidogenesis
Acidification experiments using piggery effluent were carried out in 200 L
reactors using a working volume of 100 L. The experiments were conducted

66
in the reactors T1 and T2 at 38 ± 2.7 and 55 ± 2.5 0C respectively. Both the
reactors were fed daily with piggery effluent based on a constant HRT. The
experiments were conducted at two HRT; 6.7 and 4 days for a period of up
to 6-8 HRT to obtain a reliable data set. The organic loading rate in the
reactors fluctuated between 0.64-5.44 g COD/L/day due to high variability
in solid and organic content of the raw piggery effluent. Process
performance was assessed by measurement of pH, biogas production
including methane production, solids reduction and COD reduction.
Nutrient quality was assessed by analysing TKN, TAN, SP and TP in the
acidified effluent.
Methanogenesis
Methanogenesis of the acidogenic reactor effluent was conducted in four
225 L polybag digesters (D1, D2, D3 & D4) under ambient conditions. The
digesters were initially started up using anaerobic sludge (source of sludge)
and were fed daily with the effluent from reactors based on a 23 day HRT.
Two of the digesters (D1 and D3) were fed using effluent from the
thermophilic reactor operating at 55 0C (T2) and the remaining two digesters
(D2 and D4) were fed from the mesophilic reactor operating at 38 0C (T1).
Temperature, pH and biogas volume were continuously measured during the
process. Physio-chemical analyses were performed on the digested effluent
samples using a composite sample from each digester. Biogas samples were
analysed using the online gas analyser (Gas Data LMSXi).

67
3.4.2 Results
Acidogenic Reactors:
Period 1 (Day 1-65):
Performance parameters of the acidogenic reactors T1 and T2 during period
1 (Day 1-65) are shown in Table 3.3 and Table 3.4
Table 3.3 Performance parameters of acidogenic reactor T1 (38 0C) during period 1 (Days 1-65)
Table 3.4 Performance parameters of acidogenic reactor T2 (55 0C) during period 1 (Days 1-65)
Reactor T1Parameters Min Max Avg
Total Solids (g/L) 3.2 7.9 6Volatile Solids (g/L) 1.7 5.2 3.6Soluble COD (g/L) 2.3 8.3 5.8Total COD (g/L) 5.6 13.5 7.6TKN (mg/L) 2400 3900 2900TAN (mg/L) 540 1700 1100
Soluble P (mg/L) 42 92 73Total P (mg/L) 80 119 98
VFA as COD (g/L) 0.3 2.2 1Weekly biogas (L) 0 472 114% CH4 in biogas 28 41.5 32.4
Reactor T2Parameters Min Max Avg
Total Solids (g/L) 4.8 8.9 6.1Volatile Solids (g/L) 1.9 6 3.5Soluble COD (g/L) 1.3 7.4 5Total COD (g/L) 3.8 8.6 6.7TKN (mg/L) 2400 3600 2900TAN (mg/L) 560 1700 1200
Soluble P (mg/L) 28 100 62Total P (mg/L) 60 133 93
VFA as COD (g/L) 0.26 1.5 0.76Weekly biogas (L) 0 551 149% CH4 in biogas 41.5 60 49

68
Performance of the acidification process is measured by the increased
production of VFA’s, as well as solubilisation of organic compounds which
is indicated by an increase in the concentration of soluble COD. During
period 1 (65 day experimental period), both the thermophilic and mesophilic
reactors showed a variable trend in pH during the acidification of the
piggery effluents as shown in Figure 3.9. Initially pH decreased due to
acidification of the effluent, the methanogenic activity following it resulted
in an upward trend in the pH value. The pH increase can be explained by a
drop in the organic loading rate (due to the dilute nature of the raw piggery
effluent) that led to sustained methanogenic activity in both the reactors.
The percentage of soluble COD in both reactors T1 and T2 were 78% and
74% (expressed as a percentage of Total COD in the effluent). Total COD
in the raw piggery effluent was approximately 18.2 g/L of which 68% was
present in the soluble form. The OLR during this period was approximately
2.7 g COD/L/day. Reduction in TCOD and SCOD was calculated to be 53%
and 58% respectively for reactor T1 and 60% and 63% for reactor T2.
VFA levels in reactors T1 and T2 remained around 1 g/L due to VFA’s
being consumed during the process which indicates a stable methanogenic
activity in the reactors as shown in Figure 3.10. The VFA level in both the
reactors reached concentrations over 2 g/L on day 53 due to an increase in
the OLR from 0.64 to 2.4 g COD/L/day over a 2-week period from day 44.
The VFA level then stabilised to below 1 g COD/L/day due to re-
establishment of methanogens even as the OLR remained steady at 2.4 g
COD/L/day.
TKN levels in both reactors reached 2900 mg/L, which was approximately
88% of the TKN level in the raw piggery effluent. TAN levels in reactors
T1 and T2 were 38% and 41% of the TKN in the effluent. Soluble P levels
in reactors T1 and T2 were 74% and 66% of the Total P. Trends of TKN,
TAN and phosphorus in the reactor effluent are shown in Figure 3.11,
Figure 3.12, Figure 3.13 and Figure 3.14.

69
Average weekly biogas production in T1 and T2 were approximately 114 L
and 149 L as shown in Figure 3.15. The yield of biogas was calculated to be
0.40 L/g COD in T1 and 0.52 L/g COD based on the average loading rate of
2.7 g COD/L/day. The average methane composition in the biogas samples
were 32.4% v/v and 49% v/v. The lower percentage of methane is probably
due to introduction of dissolved oxygen through daily feeding that can
inhibit the growth of methanogens. A marginal higher methane percentage
in T2 was probably because of a lower percentage of dissolved oxygen in
the effluent at a higher temperature.
Period 2 (Day 65-112):
Performance parameters of the acidogenic reactors T1 and T2 during period
2 (Day 65-112) are shown in Table 3.5 and Table 3.6.
Table 3.5 Performance parameters of acidogenic reactor T1 (38 0C) during period 2 (Day 65-112)
Reactor T1Parameters Min Max Avg
Total Solids (g/L) 4.7 6.4 5.9Volatile Solids (g/L) 2.4 3.9 3.5Soluble COD (g/L) 1.5 10.9 5.7Total COD (g/L) 6.1 24 10.7TKN (mg/L) 2500 3200 2800TAN (mg/L) 530 1900 1200
Soluble P (mg/L) 84 119 94Total P (mg/L) 129 153 136
VFA as COD (g/L) 0.21 2.4 1Weekly biogas (L) 94 454 257% CH4 in biogas 45 60 53

70
Table 3.6 Performance parameters of acidogenic reactor T2 (55 0C) during period 2 (Day 65-112)
Over the 47 day experimental period, both thermophilic and mesophilic
reactors showed stable pH values between 7.2 and 7.4. SCOD in both
reactors T1 and T2 were 53% and 68% of the TCOD in the effluent. Total
COD in the raw piggery effluent was approximately 20.4 g/L of which 41%
was present in the soluble form. The average OLR during the period was
approximately 5.1 g COD/L/day. The reduction in SCOD and TCOD was
calculated to be 31% and 72% respectively for reactor T1 and 18% and 51%
for reactor T2.
VFA levels in reactors T1 and T2 remained around 1 g/L, which showed
that there was a stable VFA consumption process and stable methanogenic
activity in the reactors. However, VFA levels in reactor T1 reached a
concentration of 2.4 g/L on day 95 and T2 reached 3.9 g/L on day 102 due
to an increase in the organic loading rate (OLR) from 1.8 to 8.3 g
COD/L/day from day 86 onwards (Figure 3.10).
TKN levels in reactors T1 and T2 reached 2800 and 2900 mg/L, which was
approximately 60% of the TKN level in the raw piggery effluent. TAN
levels in reactors T1 and T2 were 38% and 41% of the TKN in the effluent.
This is similar to TAN levels in the raw piggery effluent which ranged from
30% to 39% of TKN. SP levels in reactors T1 and T2 were 69% and 72% of
Reactor T2Parameters Min Max Avg
Total Solids (g/L) 4.5 10.6 7.6Volatile Solids (g/L) 2.3 7.3 4.8Soluble COD (g/L) 2 11.3 6.8Total COD (g/L) 5 17.9 10TKN (mg/L) 2400 4000 2900TAN (mg/L) 580 2000 1300
Soluble P (mg/L) 75 107 87Total P (mg/L) 113 129 121
VFA as COD (g/L) 0.24 2.3 1.1Weekly biogas (L) 112 601 304% CH4 in biogas 48 58 52

71
TP in the effluent. This is similar to SP levels in the raw piggery effluent,
which was calculated to be 68% of TP. Trends of TKN, TAN and P in the
reactor effluent are shown in Figure 3.11, Figure 3.12, Figure 3.13 and
Figure 3.14.
Average weekly biogas production in T1 and T2 were 257 L and 304 L
respectively as shown in Figure 3.15. Biogas yield was calculated to be 0.36
L/g COD in T1 and 0.43 L/g COD in T2 based on the average loading rate
of 5.1 g COD/L/day. The average methane compositions in the biogas
samples were 53 % v/v for T1 and 52 % v/v for T2 respectively.
Figure 3.9 pH profile during acidogenesis in reactors T1 and T2
treating raw piggery effluent
5
6
7
8
9
10
0 20 40 60 80 100 120
pH
Day
pH
T1 (38 deg C)
T2 (55 deg C)

72
Figure 3.10 VFA profile during acidogenesis in reactors T1 and T2
treating raw piggery effluent
Figure 3.11 TKN profile during acidogenesis in reactors T1 and T2
treating raw piggery effluent
0
1
2
3
4
5
0 20 40 60 80 100 120
Total V
FA (g COD/
L)
Day
VFA
T1 (38 deg C)
T2 (55 deg C)
0
1000
2000
3000
4000
5000
0 20 40 60 80 100 120
Nitrogen
(mg TKN/L)
Day
TKN
T1 (38 deg C)
T2 (55 deg C)

73
Figure 3.12 Ammonia nitrogen profile during acidogenesis in reactors
T1 and T2 treating raw piggery effluent
Figure 3.13 TP profile during acidogenesis in reactors T1 and T2
treating raw piggery effluent
0
500
1000
1500
2000
2500
0 20 40 60 80 100 120
Ammon
ia (m
g TA
N/L)
Day
TAN
T1 (38 deg C)
T2 (55 deg C)
0
50
100
150
200
0 20 40 60 80 100 120
Total Pho
sphate (m
g PO
4-‐P/L)
Day
Total P
T1 (38 deg C)
T2 (55 deg C)

74
Figure 3.14 SP profile during acidogenesis in reactors T1 and T2
treating raw piggery effluent
Figure 3.15 Biogas production and methane composition during acidogenesis in reactors T1 and T2 treating raw piggery effluent
(Source: Technical Advisory Committee Report (November 2005),
EBCRC)
0
50
100
150
200
0 20 40 60 80 100 120
Soluble Ph
ospate (m
g PO
4-‐P/L)
Day
Soluble P
T1 (38 deg C)
T2 (55 deg C)

75
Polydigesters:
Period 1 (Days 1-65):
Performance parameters of polydigesters D1-D4 treating the acidified
reactor effluent during Period 1 (Days 1-65) are shown in Table 3.7 and
Table 3.8.
Table 3.7 Performance parameters of Polydigesters D1 and D3 (ambient temperature) during Period 1 (Days 1-65)
Table 3.8 Performance parameters of Polydigesters D2 and D4 (ambient temperature) during Period 1 (Days 1-65)
Polydigesters D1 D3Parameters Min Max Avg Min Max Avg
Total Solids (g/L) 2.7 3.7 3 1.7 3.5 2.4Volatile Solids (g/L) 1.1 1.8 1.3 0.71 1.8 1.1Soluble COD (g/L) 2.3 5.3 3.9 1 4 2.8
TKN (mg/L) 1500 2400 1900 830 2400 1500TAN (mg/L) 550 1700 1100 170 860 630
Soluble P (mg/L) 22 47 32 16 25 20VFA as COD (g/L) 0 0.84 0.42 0 0.64 0.24Weekly biogas (L) 3 58 26 10 52 30% CH4 in biogas 72 74 74 63 70 67
Polydigesters D2 D4Parameters Min Max Avg Min Max Avg
Total Solids (g/L) 2.6 3.5 3 1.8 3.4 2.5Volatile Solids (g/L) 1.1 1.7 1.3 0.74 1.8 1.3Soluble COD (g/L) 1.9 4.1 3.5 1.6 5 3.5
TKN (mg/L) 1700 2200 2000 740 2100 1495TAN (mg/L) 580 1700 1100 260 1200 710
Soluble P (mg/L) 23 47 38 20 41 29VFA as COD (g/L) 0 0.72 0.39 0 0.92 0.33Weekly biogas (L) 4 87 45 18 81 49% CH4 in biogas 72 79 76 67 72 69

76
Period 2 (Days 65-112):
Performance parameters of Polydigesters D1 to D4 treating the acidified
reactor effluent during Period 2 (Days 65-112) are shown in Table 3.9 and
Table 3.10.
Table 3.9 Performance parameters of Polydigesters D1 and D3 (ambient temperature) during Period 2 (Days 65-112)
Table 3.10 Performance parameters of polydigesters D2 and D4 (ambient temperature) during period 2 (Days 65-112)
During period 1, pH in polydigesters D1-D4 remained between 6.9 and 8
(Figure 3.16). The average VFA in the effluent was between 0.33 and 0.42 g
COD/L, with an increase to 0.85-1.4 g COD/L towards the end of period 1
(Figure 3.17). Approximately 42-58 % of the TKN in the digested effluent
Polydigesters D1 D3Parameters Min Max Avg Min Max Avg
Total Solids (g/L) 3.8 4.3 4 3.4 4.1 3.8Volatile Solids (g/L) 1.9 2.1 2 1.8 2.1 1.8Soluble COD (g/L) 4.3 11 6 3.5 11.1 6.3
TKN (mg/L) 2500 2800 2700 2200 2700 2400TAN (mg/L) 640 1700 1200 600 1800 1100
Soluble P (mg/L) 53 67 58 35 79 57VFA as COD (g/L) 0.66 1.3 1 0.68 2 1.2Weekly biogas (L) 59 509 181 49 293 123% CH4 in biogas 71 76 73 66 74 71
Polydigesters D2 D4Parameters Min Max Avg Min Max Avg
Total Solids (g/L) 2.6 3.5 3 1.8 3.4 2.5Volatile Solids (g/L) 1.1 1.7 1.3 0.74 1.8 1.3Soluble COD (g/L) 1.9 4.1 3.5 1.6 5 3.5
TKN (mg/L) 1700 2200 2000 740 2100 1495TAN (mg/L) 580 1700 1100 260 1200 710
Soluble P (mg/L) 23 47 38 20 41 29VFA as COD (g/L) 0 0.72 0.39 0 0.92 0.33Weekly biogas (L) 4 87 45 18 81 49% CH4 in biogas 72 79 76 67 72 69

77
was present in the form of TAN in D1 and D3 while in D2 and D4 TAN
constituted about 47-55 % of TKN (Figure 3.18 and Figure 3.19). Soluble P
values were in the range 25-50 mg PO4-P/L (Figure 3.20). Biogas yield in
D1 and D3 were approximately 0.05 and 0.06 L/g COD while it was
approximately 0.1 L/g COD in both D2 and D4 (Figure 3.21).
During period 2, pH in polydigesters D1-D4 remained between 7 and 7.5
(Figure 3.16). The average VFA in the effluent was between 0.81 and 1.2 g
COD/ L. The average VFA level in D1 and D3 was slightly higher than in
D2 and D4 towards the end of period 2 (Figure 3.17). Approximately 44-
46% of the TKN in the digested effluent was present in the form of TAN in
D1 and D3 while in D2 and D4 TAN constituted about 46-50% of TKN
(Figure 3.18 and Figure 3.19). SP values were in the range 65-80 mg PO4-
P/L (Figure 3.20). Biogas yield in D1 and D3 was 0.24 and 0.17 L/g COD
respectively while it was 0.34 and 0.32 L/g COD in D2 and D4 (Figure
3.21).
Figure 3.16 pH profile during anaerobic digestion in polydigesters D1,
D2, D3 & D4 treating acidified piggery effluent from the acidogenic reactors
6.5
7
7.5
8
8.5
0 20 40 60 80 100 120
pH
Day
pH
D1
D2
D3
D4

78
Figure 3.17 VFA profile during anaerobic digestion in polydigesters D1,
D2, D3 & D4 treating acidified piggery effluent from the acidogenic reactors
Figure 3.18 TKN profile during digestion in polydigesters D1, D2, D3 &
D4 treating acidified piggery effluent from the acidogenic reactors
0
0.5
1
1.5
2
0 20 40 60 80 100 120
Total V
FA (g COD/
L)
Day
VFA
D1
D2
D3
D4
0
500
1000
1500
2000
2500
3000
0 20 40 60 80 100 120
Nitrogen
(mg TKN/L)
Day
TKN
D1
D2
D3
D4

79
Figure 3.19 Ammonia profile during digestion in polydigesters D1, D2,
D3 & D4 treating acidified piggery effluent from the acidogenic reactors
Figure 3.20 Soluble P profile during digestion in polydigesters D1, D2,
D3 & D4 treating acidified piggery effluent from the acidogenic reactors
0
500
1000
1500
2000
2500
0 20 40 60 80 100 120
Ammon
ia (m
g TA
N/L)
Day
TAN
D1
D2
D3
D4
0
25
50
75
100
0 20 40 60 80 100 120
Phosph
ate (m
g PO
4-‐P/L)
Day
Soluble P
D1
D2
D3
D4

80
Figure 3.21 Biogas production and methane composition during
digestion in polydigesters D1, D2, D3 & D4 treating acidified piggery effluent from the acidogenic reactors
(Source: Technical Advisory Committee Report (November 2005),
EBCRC)
3.4.3 Conclusions
The set up and installation of the pilot plant anaerobic digestion system
provided an opportunity to be involved with a team of post doctoral
researchers and anaerobic digestion experts. This helped in understanding
the concepts of the anaerobic digestion process, in particular the kinetics of
raw piggery effluent digestion in the two stages; a thermophilic stage
followed by a methanogenic stage. It also gave the researcher an
opportunity to apply the principles of engineering while assisting in setting
up and commissioning the pilot plant. The hands on approach during start

81
up and troubleshooting of the system provided ample opportunity to learn
about the process in depth.
Nutrient analysis conducted according to standard methods (APHA,
AWWA et al. 1998), as part of the pilot scale project, on the raw and
anaerobically digested piggery effluent provided the necessary training to
conduct the nutrient assays when the laboratory scale anaerobic digestion
experiments were set up (Section 3.8), as well as giving baseline data.
The data collected from these anaerobic digestion experimental runs
provided valuable information for use as an input to the aquaculture
component of the IBS model. The values of COD, TKN, TAN, SP and P
were used as inputs in developing the anaerobic digestion component of the
model and also as inputs to DYRESM CAEDYM for the aquaculture stages
of the model.
During Stage 1 of the experiments, in the thermophilic stage of anaerobic
digestion both T1 and T2 showed a relatively stable pH during the 28 days
of operation. The TCOD decreased 47% while SCOD increased 46% due to
increased solubilisation. TAN and SP increased by approximately 3 and 6
times respectively due to conversion of VFA’s into bioavailable forms of N
and P. Gas production in the thermophilic reactors was initially 50 L/day but
this declined over the period, possibly due to leakages detected in the
system. In the polydigesters (methanogenic stage), a maximum COD
reduction of 61% was observed in polydigester 2 along with a higher gas
production rate. TAN and SP in both the polydigesters stabilised around
1700 mg/L and 22 mg PO4-P/L respectively. Biogas production in
polydigesters 1 and 2 were at an average rate of 2.4 and 5.7 L/day
containing approximately 30% CO2.
Stage 2 of the experiments was conducted in two periods. pH values
remained stable for both thermophilic and mesophilic reactors between 7.2
and 7.4. The reduction in soluble and total COD ranged between 31-53%
and 58-72% for T1 and 18-60% and 51-63% for T2 respectively. TAN
levels in T1 and T2 averaged 38% and 41% of the TKN value in the

82
effluent. Soluble P levels in T1 and T2 were 69-74% and 66-72% of total P
in the effluent. Average weekly biogas production was between 114-257 L
and 149-304 L for T1 and T2 respectively. The methanogenic digesters
(polydigesters D1-D4) recorded a pH between 6.9-8. Approximately 42-
58% of the TKN in the digested effluent was present in the form of TAN.
Soluble P values were in the range 25-80 mg PO4-P/L. Weekly biogas
output in the polydigesters ranged from 45-181 L.
The values of COD, TAN, Soluble P and biogas production will be used in
the development of the anaerobic digestion model (Chapter 4). The values
of TAN and Soluble P will form input data to DYRESM CAEDYM
(Chapter 5).
3.5 Set up of the Pilot Scale Integrated Aquaculture
System
A pilot scale integrated aquaculture infrastructure (mesocosm facility) was
set up in the Breakwell Laboratory building of Roseworthy Campus for the
culture of microalgae, zooplankton and fish using digested piggery effluent
from the anaerobic system. The system works on an indoor gravity flow
consisting of a series of tanks of varying sizes to provide a range of
hydraulic retention times. The integrated aquaculture system consisting of a
series of tanks is shown in Figure 3.23. Associated infrastructure was
established including controlled lighting, pumping and pipe work to enable
conveyance of digested piggery effluent and water to the system.
Experiments relating to bioconversion of microalgae using digested piggery
effluent were conducted once the system was commissioned in early 2007.

83
Figure 3.22 Schematic diagram of the pilot scale Integrated Aquaculture System
Figure 3.23 Clear Perspex tanks set up at differential heights for micro-algal culture while blue fibre glass tanks are used for fish culture. These tanks are part of the indoor integrated aquaculture system (mesocosm)
at Roseworthy Laboratory
Microalgae
Zooplankton
Fish
Anaerobically digested
piggery effluent

84
3.6 Bioconversion of piggery effluent to algae (280 L
working volume)
3.6.1 Objective
The aim of this experiment was to evaluate the bioconversion of algae
Chlorella sp. at a fixed working volume of 280 L and a nutrient
concentration of 30 mg/L/day of TAN. This was achieved by the addition of
anaerobically digested piggery effluent as the nutrient source. The data of
TAN, SP and pH obtained from these experiments were used to model the
zooplankton component of DYRESM CAEDYM for the IBS model.
3.6.2 Materials and Methods
Three 400 L photo bioreactors were used each with a total working volume
of 280 L. The reactors were filled with 280 L of dechlorinated mains water.
The reactors were inoculated with Chlorella sp. (10%), which was obtained
from the microalgal laboratory at the University of Adelaide. The
experiment was conducted under controlled fluorescent light (45000 Lux)
and 15:9 hours photoperiod (light: dark) at 24 0C. The concentration of
nutrient in the reactors were maintained above 30 mg/L TAN by daily
addition of anaerobically digested piggery effluent. The effluent had a TAN
of 1646 mg/L and soluble orthophosphate of 55.2 mg/L. Samples of culture
were collected daily. The samples were analysed for total algal cell density,
and nutrient concentrations (ammonia and soluble orthophosphate). Nutrient
analysis was conducted according to the APHA et al., Standard Methods
(1998). The cell density was measured using a haemocytometer and a
binocular microscope. Once the cell density reached 2.5 million cells/mL,
the algal culture was diluted to 1.0 million cells/mL by harvesting the algae
and making up the volume to 280 L by addition of dechlorinated water. pH,
temperature and DO were monitored daily. pH was maintained above 9.5 to
minimize risk of infestation by rotifers into the system. The algae removed
from these reactors were used as an inoculum for the start up of three other
algal reactors.

85
Equilibrium equations for calculating unionized ammonia:
Unionized ammonia = )(101
2.1 pHpka
iaTotalAmmonx−+
Eq. 3.1
Where pka = 0.902 + ⎟⎠
⎞⎜⎝
⎛+T2.273
2730
T = temperature in 0C (Emerson 1975; Whitehouse 2006)
3.6.3 Results
From the equilibrium equations it was calculated that to maintain a
unionized ammonia concentration greater than 3.0 mg/L (minimum
threshold value), the TAN in the algal reactors (with a working volume of
280 L) would have to be greater than 30 mg/L.
The mean cell density of the algae in the three reactors over a period of 60
days is shown in Figure 3.24. The mean cell density of the algal culture
reached 2.5 million cells/ml on day 10, 22, 29 and 34. The algae were
harvested on these particular days in the afternoon by removing 110 L of the
algal culture and the cell density was reduced to 1.5 million cells/ml to keep
them growing continuously. The mean cell density however reduced to 1.5
million/ml after day 40. This was probably due to the removal of the biofilm
which had developed on the interior surface of the reactor. The biofilm
accelerated the removal of particulate matter present in the piggery effluent,
thus reducing turbidity and facilitating cell growth and nutrient
consumption.
The mean ammonia concentration measured as TAN is shown in Figure
3.25. It is evident that the ammonia concentration was reduced by 30
mg/L/day. After day 40 the ammonia concentration did not decline by the
same amount.
The mean soluble phosphorus concentration which was in the range
between 2-3 mg/L/day is shown in Figure 3.26. After day 40, there was an

86
increase in the phosphorus concentration above 3 mg/L which rose steadily
till day 60.
The decrease in ammonia and phosphorus reduction could be due to the
removal of the biofilm which decreased the rate of consumption of nutrients
by the algal culture.
The mean pH of the algal cultures is shown in Figure 3.27. The pH was
maintained above 9.5 during the run to prevent growth of rotifers by
addition of pelletized NaOH. Rotifers are known to feed on algae, organic
detritus, dead bacteria and protozoans (Lionard, Azemar et al. 2005) and
thus if left uncontrolled can graze on the algae present in the tanks which is
not desired.
Figure 3.24 Mean cell density in 280 L algal culture
0.000.501.001.502.002.503.00
0 10 20 30 40 50 60 70
Cell D
ensity (M
/ml)
Days
Cell Density
Cell Density (M/ml) (280 L)

87
Figure 3.25 Mean TAN concentration in 280 L algal culture
Figure 3.26 Mean SP concentration in 280 L algal culture
05101520253035
0 10 20 30 40 50 60 70
Conc. o
f Ammon
ia(TAN
mg/L)
Days
Ammonia
Ammonia (280 L)
0
1
2
3
4
5
0 10 20 30 40 50 60 70
Conc. o
f Pho
spho
rus
(mg/L)
Days
Phosphorus
Phosphorus (280 L)
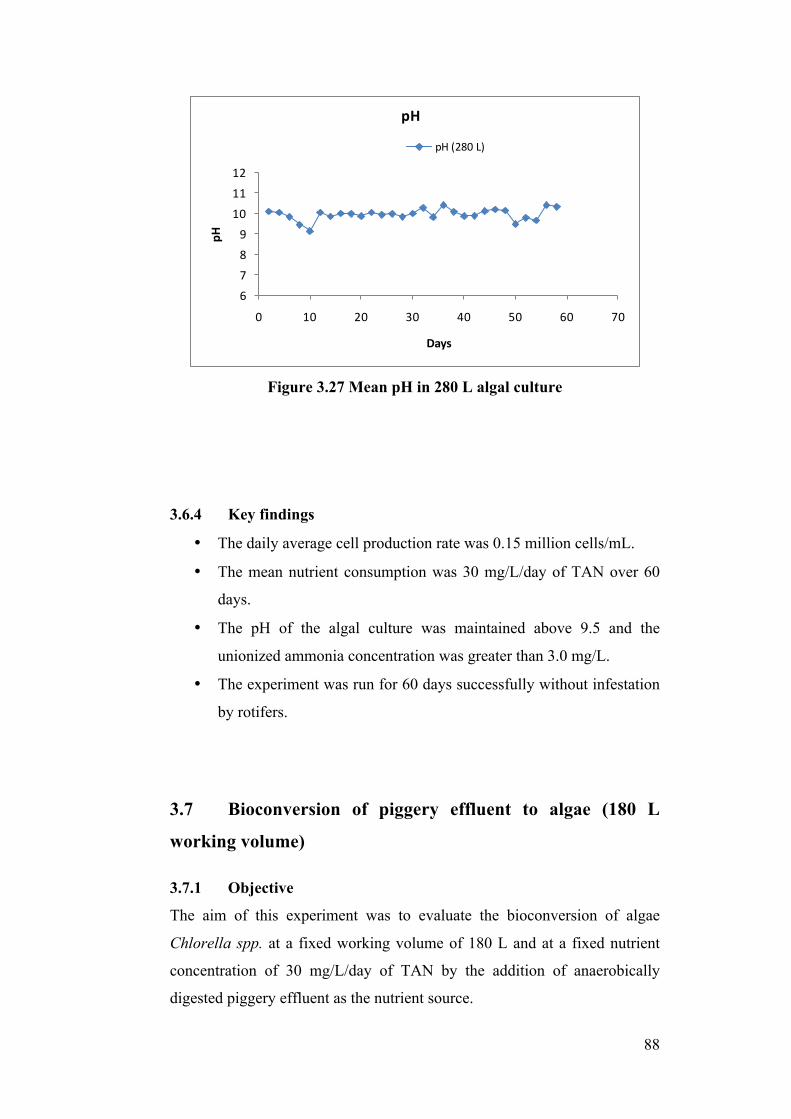
88
Figure 3.27 Mean pH in 280 L algal culture
3.6.4 Key findings
• The daily average cell production rate was 0.15 million cells/mL.
• The mean nutrient consumption was 30 mg/L/day of TAN over 60
days.
• The pH of the algal culture was maintained above 9.5 and the
unionized ammonia concentration was greater than 3.0 mg/L.
• The experiment was run for 60 days successfully without infestation
by rotifers.
3.7 Bioconversion of piggery effluent to algae (180 L
working volume)
3.7.1 Objective
The aim of this experiment was to evaluate the bioconversion of algae
Chlorella spp. at a fixed working volume of 180 L and at a fixed nutrient
concentration of 30 mg/L/day of TAN by the addition of anaerobically
digested piggery effluent as the nutrient source.
67
89
1011
12
0 10 20 30 40 50 60 70
pH
Days
pH
pH (280 L)

89
3.7.2 Materials and Methods
Three 400 L photo bioreactors were used each with a total working volume
of 180 L. The reactors were filled with 180 L of dechlorinated mains water.
The reactors were inoculated with 10% density of Chlorella spp. (1.0 x 106
cells/mL). The experiment was conducted under controlled fluorescent light
(45000 Lux) and 15:9 hours photoperiod (light: dark) at 24 0C. The
concentration of nutrient in the reactors were maintained above 30 mg/L
TAN by addition of anaerobically digested piggery effluent daily. The
effluent had a TAN of 1650 mg/L and soluble orthophosphate of 55.6 mg/L.
Samples of culture were collected daily. The samples were analysed for
total algal cell density, and nutrient concentrations (ammonia and soluble
orthophosphate). Nutrient analysis was conducted according to the APHA et
al., Standard methods (1998). The cell density was measured using a
haemocytometer and a binocular microscope. Once the cell density reached
2.0 million cells/mL, the algal culture was diluted to 1.0 million cells/mL by
harvesting the algae and making up the volume to 280 L by addition of
dechlorinated water. pH, temperature and DO were monitored daily. pH was
maintained above 9.5 to minimize risk of infestation by rotifers into the
system.
3.7.3 Results
The mean cell density of the algae in the three reactors over a period of 14
days is shown in Figure 3.28. The mean cell density of the algal culture
reached a peak of 2.0 million cells /mL on day 9. The cell density reduced to
1.0 million cells/mL by harvesting the algae on day 9 by removing 90 L of
the algal culture to ensure their continuous growth.
The mean ammonia concentration measured as TAN is shown in Figure
3.29. The nutrient consumption was less than 15.0 mg/L/day during the first
few days of the run which explains the decrease in cell density of the culture
during that time. Once the cell density started to increase, the nutrient
consumption also increased to an average of 25 mg/L/day. However this
was less when compared to the nutrient consumption of 30 mg/L/day when
the cell culture was maintained at a working volume of 280 L.

90
The mean SP concentration of the three algal reactors is shown in Figure
3.30. During the run of 14 days, the phosphorus concentration was between
(2-3) mg/L/day.
The mean pH of the algal cultures is shown in Figure 3.31. The pH was
maintained above 9.5 during the run to prevent growth of rotifers.
Figure 3.28 Mean cell density for 180 L algal culture
0.00
0.50
1.00
1.50
2.00
2.50
0 5 10 15
Cell Den
sity (M/m
l)
Days
Cell Density
Cell Density (M/ml) (180 L)

91
Figure 3.29 Mean TAN for 180 L algal culture
Figure 3.30 Mean SP for 180 L algal culture
02468
10121416
0 2 4 6 8 10 12 14 16
Conc. O
f Ammon
ia
(TAN m
g/L)
Days
Ammonia
Ammonia (180 L)
0
1
2
3
4
5
0 5 10 15
Conc. O
f Solub
le Pho
spho
rus
(mg/L)
Days
Phosphorus
Phosphorus (180 L)

92
Figure 3.31 Mean pH for 180 L algal culture
3.7.4 Key findings
• The average daily cell production rate was 0.11 million cells/mL.
• The mean nutrient consumption was 25 mg/L/day of Total Ammonia
Nitrogen (TAN).
• The experiment ran successfully for 14 days.
• It was observed that higher cell density of 2.5 million cells/mL and
greatest nutrient reduction of 30 mg/L/day of TAN occurred when
the working volume of algae culture was 280 L.
Table 3.11 Comparison of data between experiments 1 and 2
Data Experiment 1 Experiment 2
Working Volume (L) 280 180
Cell Production Rate
(million cells/ mL) 0.15 0.11
Mean Nutrient
Consumption
(mg/L/day)
30 25
6
7
8
9
10
11
12
0 2 4 6 8 10 12 14 16
pH
Days
pH
pH (180 L)

93
3.8 Lab Scale Anaerobic Digestion Experiments
3.8.1 Introduction
Small scale anaerobic digestion experiments were conducted in the
laboratory to collect data for the development of a mathematical model for
the anaerobic digestion system. These experiments were conducted in batch
mode to understand the degradation of raw piggery effluent over time and to
measure TAN, SP, COD and pH to use as input data to the aquaculture
model DYRESM CAEDYM. The results of the experiments conducted and
the development of the anaerobic digestion model is described in Chapter 4.
3.8.2 Materials & Methods
Raw piggery effluent was collected from the Roseworthy Piggery,
Roseworthy and chilled at 40C in a cool room to slow down microbial
activity, thereby reducing any biodegradation. The characteristics of the
piggery effluent used for the anaerobic digestion experiments are
represented in Table 3.12 and Table 3.13.
Table 3.12 Characteristics of the raw piggery effluent from the Roseworthy piggery
Parameter Range
COD (g/L) 10 – 40
TS (g/L) 7 – 8
VS (g/L) 3 – 4
TAN (mg/L) 700 – 1000
TKN (mg/L) 3000 –
4000
SP (mg/L) 70 – 80
TP (mg/L) 180 – 200
Nitrate (mg/L) < 2.5
pH 7.5 – 8.0

94
Table 3.13 Characteristics of the raw piggery effluent data
Parameter Value
COD (g/L) 14.6
TAN (mg/L) 751.5
SP (mg/L) 76.1
pH 7.8
The experiment was carried out in 1.0 L batch reactors (Schott Bottles) as
shown in Figure 3.32. The laboratory scale reactors used in this study were
made of glass and were covered with aluminium foil on the exterior to
minimize the effect of light on the contents inside the bottle. Temperatures
of 370 C, 450 C and 550 C were chosen for the experiments. The experiment
was run with triplicates of each temperature. Temperatures were maintained
using a thermostat on a hot plate heater with a magnetic stirrer. The
magnetic stirrer was used to maintain a well mixed culture in the Schott
Bottles with continuous stirring. 1.0 L piggery effluent was used as the
mixed culture for the experiments. It consisted of 70% raw piggery effluent
obtained from the cool room and 30% semi digested piggery effluent as
inoculum to initiate the digestion process. The measurements of COD, TAN
and SP were conducted according to Standard Methods (APHA, AWWA et
al. 1998). Samples were collected at the start of the experiment, on day 7,
and then every two days till no significant change in COD, TAN and P was
observed. The biogas produced in the batch reactor was passed through a
Schott Bottle containing potassium hydroxide (KOH) solution to absorb the
carbon dioxide (CO2) in the biogas. Methane gas (CH4) was collected and
measured by the downward displacement of water.

95
Figure 3.32 Experimental apparatus for the laboratory study of anaerobic digestion.
3.9 Conclusions
This chapter has discussed the set up, installation and commissioning of the
two stage anaerobic digestion system and the Integrated Aquaculture system
which are the two main components of the IBS process. The set up and
methods for data collection for small scale batch anaerobic digestion
experiments were also discussed. The data collected from these experiments
will be analysed for the development of an anaerobic digestion model in
Chapter 4.
The pilot scale Integrated Aquaculture system provided useful information
on the growth of microalgae by consuming nutrients (N & P) available in
anaerobically digested piggery effluent. A higher working volume (280 L)
yielded a higher microalgal cell concentration and a higher reduction in the
TAN and P values. This result is in agreement with other studies conducted
Sampling
Port
Raw Piggery
Effluent
KOH
Solution
Hot Plate with
Magnetic Stirrer
Water

96
on aquaculture systems (Molina Grima, Fernández et al. 1999; Ugwu,
Aoyagi et al. 2008).
The data to be used for modelling the IBS in the following chapters is
shown in Table 3.14. The values of TAN, SP, COD and CH4 will be used
for the anaerobic digestion model development while TAN, SP and pH will
be used for developing the aquaculture model of the IBS.
The values of TAN, soluble phosphorus (SP) and pH obtained from the
anaerobic digestion experiments were used as inputs into the aquaculture
component of the IBS model i.e. DYRESM-CAEDYM.
Table 3.14 Data from experiments to be used in IBS model development
Data Value
TAN 1600 mg/L
SP 150 mg/L
COD 7000 mg/L
CH4 150 mL
pH 7.5

97
4 Development of an Anaerobic Digestion
Model
4.1 Introduction
The aim of this chapter is to present a simple model of the biochemical
processes for the anaerobic digestion module of the IBS. The original aim of
this study was to use a simple methanogenesis model and to determine the
correlation between methane production and release of phosphorus (P); and
total ammonia nitrogen (TAN).
There has been numerous studies conducted on the relationship between
COD, CH4 production, TAN and P (Hill and Barth 1977; Chen and
Hashimoto 1978; Kleinstreuer and Powegha 1982; Mueller 1982; Mosey
1983; Moletta, Verrier et al. 1986; Pullammanappallil, Owens et al. 1991;
Costello, Greenfield et al. 1991a; Costello, Greenfield et al. 1991b;
Angelidaki, Ellegaard et al. 1993; Romli 1993; Pullammanappallil,
Chynoweth et al. 2001). These publications show that COD decrease in
effluents is compensated by increase in CH4 production, TAN and P.
Once raw data obtained from the experiments in Chapter 3 was analysed, it
was found that P and TAN were not correlated to methane production. A
new approach to modelling the anaerobic digestion system was required
which involved separate microbial kinetics for TAN, P and methane
production.
Conventional anaerobic digesters operate on a continuous basis. Death
kinetics are not incorporated in continuous digester models as washout is
present swamping the death effects. However in batch processes there is no
washout, therefore death kinetics need to be implemented. The initial
assumption was that the increase in TAN and P were correlated to methane
production with a corresponding decrease in COD.
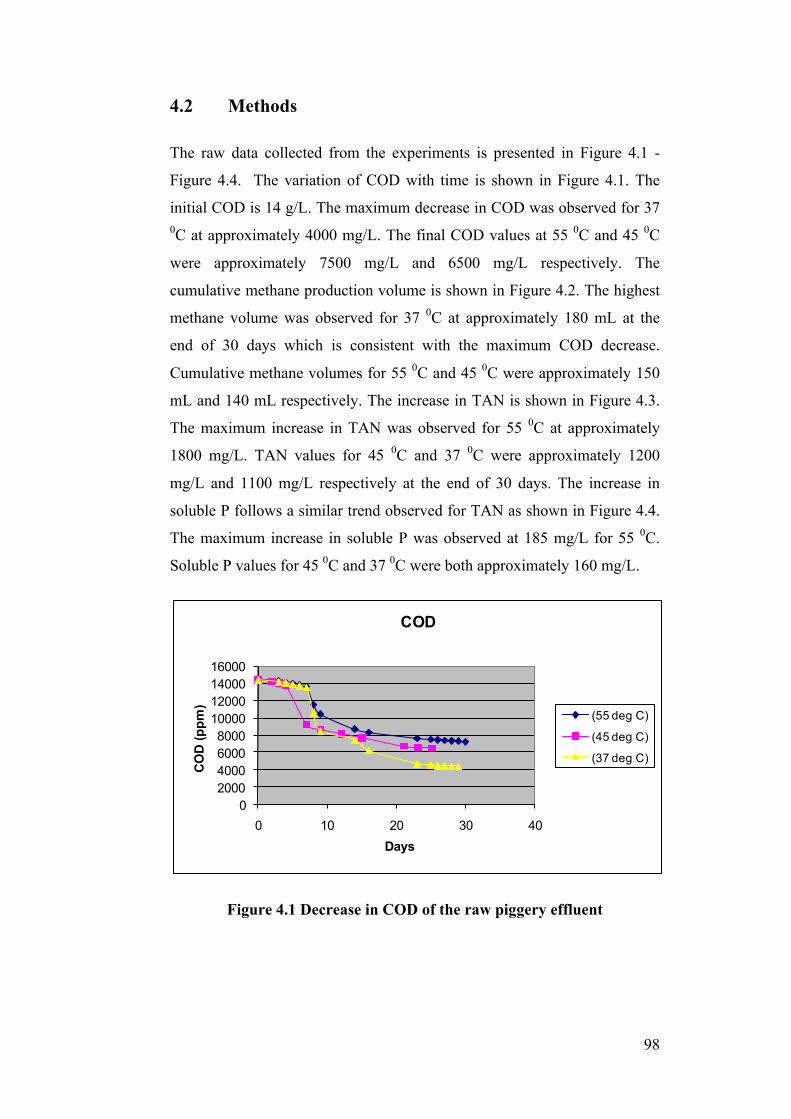
98
4.2 Methods
The raw data collected from the experiments is presented in Figure 4.1 -
Figure 4.4. The variation of COD with time is shown in Figure 4.1. The
initial COD is 14 g/L. The maximum decrease in COD was observed for 37 0C at approximately 4000 mg/L. The final COD values at 55 0C and 45 0C
were approximately 7500 mg/L and 6500 mg/L respectively. The
cumulative methane production volume is shown in Figure 4.2. The highest
methane volume was observed for 37 0C at approximately 180 mL at the
end of 30 days which is consistent with the maximum COD decrease.
Cumulative methane volumes for 55 0C and 45 0C were approximately 150
mL and 140 mL respectively. The increase in TAN is shown in Figure 4.3.
The maximum increase in TAN was observed for 55 0C at approximately
1800 mg/L. TAN values for 45 0C and 37 0C were approximately 1200
mg/L and 1100 mg/L respectively at the end of 30 days. The increase in
soluble P follows a similar trend observed for TAN as shown in Figure 4.4.
The maximum increase in soluble P was observed at 185 mg/L for 55 0C.
Soluble P values for 45 0C and 37 0C were both approximately 160 mg/L.
Figure 4.1 Decrease in COD of the raw piggery effluent
02000400060008000
10000120001400016000
0 10 20 30 40
COD
(ppm
)
Days
COD
(55 deg C)
(45 deg C)
(37 deg C)

99
Figure 4.2 Cumulative methane output
Figure 4.3 Increase in TAN of the raw piggery effluent
020406080
100120140160180200
0 10 20 30 40
Cum
.Met
hane
(ml)
Days
Methane
(55 deg C)
(45 deg C)
(37 deg C)
0200400600800
100012001400160018002000
0 10 20 30 40
TAN
(ppm
)
Days
TAN
(55 deg C)
(45 deg C)
(37 deg C)

100
Figure 4.4 Increase in soluble P of the raw piggery effluent
The original intention of modelling the anaerobic digestion process as it
occurred was to correlate TAN and soluble P with methane production,
which can be predicted from COD reduction. Scaled plots for COD, TAN,
Soluble P and CH4 production are shown in Figure 4.5. This allows data to
be compared on one set of axes. TAN, Soluble P and CH4 production plots
were normalized and inversed while the COD plot was just normalized to
standardise all data on the same scale.
Normalization was done by dividing the parametric values by the highest
value. It was observed that COD and CH4 production were related as they
followed a similar trend, mainly below the straight line in Figure 4.5. On the
contrary, processes involving TAN and Soluble P were similar, but
generally above the straight line in Figure 4.5. The straight line is merely
used to compare the experimental data. This indicated that both TAN/
Soluble P and COD/ CH4 followed two different sets of processes which
were not correlated. The straight line indicated that COD and CH4 behaved
similarly (i.e. were related) but TAN and Soluble P were different
(unrelated) to COD reduction (but appeared similar to each other).
Therefore an alternate approach to modelling the anaerobic digestion
process was required to be adopted which is described below.
020406080
100120140160180200
0 10 20 30 40
P (p
pm)
Days
Soluble P
(55 deg C)
(45 deg C)
(37 deg C)

101
Figure 4.5 Scaled/Normalized Plot for Anaerobic Digestion of Piggery Effluent
4.2.1 Development of Model Equations for the Anaerobic Digestion
Process
The equations defining organism growth processes are listed below.
Organism change:
Organism Growth Rate = f (substrate, temperature) Eq. 4.1
The Death Rate function is defined by a function similar to Organism
Growth Rate.
Organism Death Rate = f (substrate, temperature) Eq. 4.2
Amount of Product = [Organism Growth Rate] x [Product Constant]
Eq. 4.3
00.10.20.30.40.50.60.70.80.9
1
0 10 20 30day
55 C
Sc. COD
Sc. TAN
Sc. P
Sc. CH4
StrLine

102
Organism Change:
=dtdO
[OrganismInflow] + [Organism Growth Rate] – [Organism Death Rate]
- [OrganismOutflow] Eq. 4.4
Substrate Change:
dtdS = [SubstrateInflow] + [[Organism Death Rate] x [Substrate Returned
Constant]] – [[Organism Growth Rate] x [Substrate Use Constant]] – [SubstrateOutflow] Eq. 4.5
Equations 4.4 and 4.5 have been derived from mass balance of parameters.
The batch data was fitted using “minimisation of errors squared” technique,
which essentially minimises the sum of the square of the errors. Each set of
batch data had 7 constants for the averages obtained for temperatures of 37 0C (mesophilic), 45 0C (intermediate, top end of mesophilic) and 55 0C
(thermophilic). These constants are listed below.
• Initial Substrate Concentration
• Initial Bacterial Concentration
• Growth Rate (U)
• Death Rate (D)
• Substrate Use Constant
• Substrate Returned Constant
• Methane Volume (Gas Constant)
As the initial substrate concentration, initial bacterial concentration,
substrate use constant, substrate returned constant and methane volume (gas
constant) should not change with temperature only the growth rate and death
rate constants were assumed to vary with temperature (Chen and Hashimoto
1978; Chen 1983).

103
4.3 Results
The results for the model at 55 0C are presented in this section. The results
for temperatures 45 0C and 37 0C are presented in Appendix B.
4.3.1 Methane Model
The initial step was to fit the proposed model to the average of three runs at
each temperature as shown in Table 4.1
Table 4.1 Fitted r2 values for CH4
Since U and D were the only constants expected to be influenced by
temperature these were adjusted to minimise the r2, giving the values in
Table 4.2.
Table 4.2 Refitted Growth and Death Rates for CH4 with r2 values
Temp Substr Bact SubstUse U D SustrRet Const Fitted r2
37 1.19 0.0000079 0.000562 0.889 0.957 0.00093 413 0.9917245 1.02 0.0000203 0.000862 0.992 0.947 0.00146 419 0.9785855 1.10 0.0000110 0.000765 0.910 0.905 0.00148 416 0.98971
Average 1.11 0.0000130 0.000729 0.930 0.936 0.00129 416 0.98667
Temp U D r2
37 0.774 0.702 0.9839945 1.159 1.230 0.9757055 0.844 0.860 0.98925
Average 0.98298

104
Figure 4.6 Modelled methane data at 55 0C
The modelled methane plot in Figure 4.6 shows the simulated CH4 levelling
off after 30 days of simulation but the measured data seems to be increasing
slightly after that time period. During initial start up, the simulated CH4
increases almost instantly whereas in the measured data there is an initial
lag phase for approximately 3 days during which there is negligible CH4
production.
0.0
0.2
0.4
0.6
0.8
1.0
0 5 10 15 20 25 30
Day
Methane 55 -‐ normalised
Substr
Bact
Grwth
Death
Cum CH4
Measured

105
Figure 4.7 Temperature Response for Growth and Death Rates in CH4 modelling
The temperature response plot for CH4 in Figure 4.7 shows the response
plots for Death and Growth Rates almost superimposed on each other.
Growth and Death Rate is maximum for 45 0C and minimum for 37 0C, with
the value of Death Rate being slightly higher than that of the Growth Rate
for 45 0C.
4.3.2 TAN Model
Table 4.3 Fitted r2 values for TAN
Table 4.4 Refitted Growth and Death Rates for TAN with r2 values
y = -‐0.0044x2 + 0.4114x -‐ 8.3828R² = 1
y = -‐0.0057x2 + 0.5351x -‐ 11.265R² = 1
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
0 10 20 30 40 50 60
D-‐De
ath
U-‐Growth
Temperature (oC)
Temperature Response -‐ CH4
U
D
Poly. (U)
Poly. (D)
Temp Substr Bact SubstUse U D Const Fitted r2
37 3.21 1.19E-05 0.00879 0.243 2.049 182 0.9615845 3.21 2.60E-05 0.01060 0.308 2.863 228 0.9968155 3.13 5.25E-05 0.00846 0.231 1.735 182 0.99201
Average 3.18 3.01E-05 0.00928 0.261 2.215 197 0.97920
Temp U D r2
37 0.160 1.304 0.9651745 0.295 2.651 0.9966155 0.313 2.588 0.99175
Average 0.98451
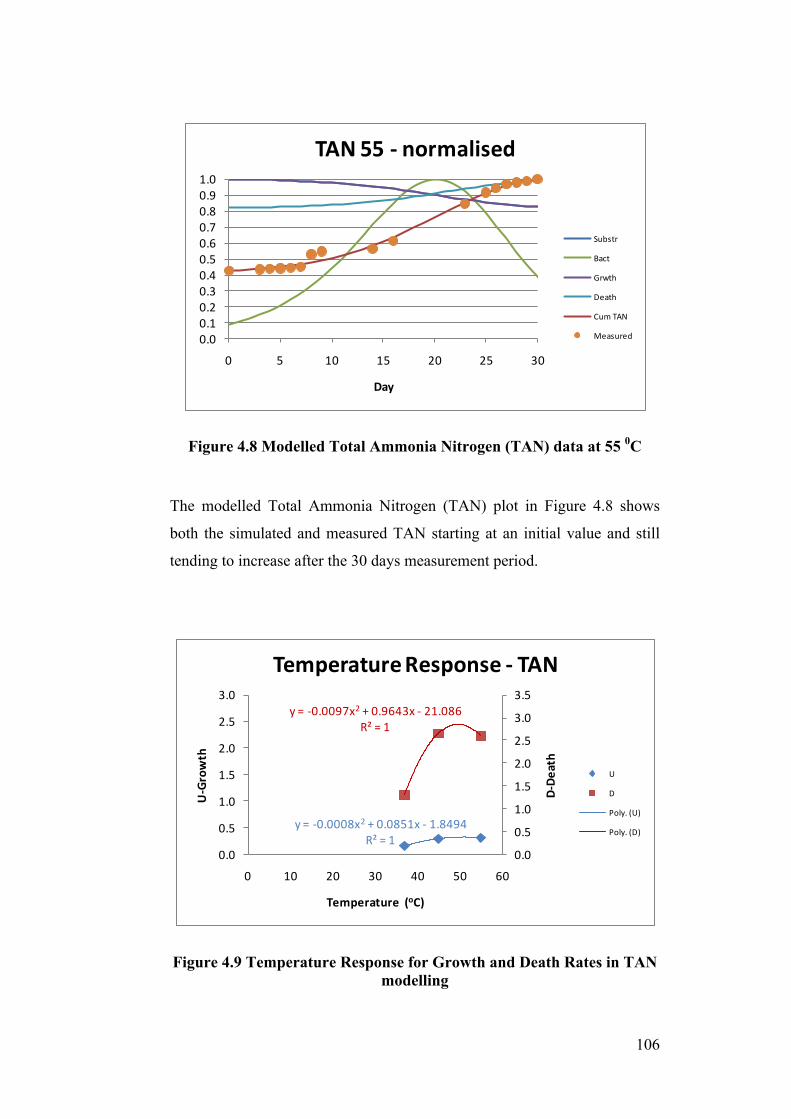
106
Figure 4.8 Modelled Total Ammonia Nitrogen (TAN) data at 55 0C
The modelled Total Ammonia Nitrogen (TAN) plot in Figure 4.8 shows
both the simulated and measured TAN starting at an initial value and still
tending to increase after the 30 days measurement period.
Figure 4.9 Temperature Response for Growth and Death Rates in TAN modelling
0.00.10.20.30.40.50.60.70.80.91.0
0 5 10 15 20 25 30
Day
TAN 55 -‐ normalised
Substr
Bact
Grwth
Death
Cum TAN
Measured
y = -‐0.0008x2 + 0.0851x -‐ 1.8494R² = 1
y = -‐0.0097x2 + 0.9643x -‐ 21.086R² = 1
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
0.0
0.5
1.0
1.5
2.0
2.5
3.0
0 10 20 30 40 50 60
D-‐De
ath
U-‐Growth
Temperature (oC)
Temperature Response -‐ TAN
U
D
Poly. (U)
Poly. (D)

107
The temperature response plot for TAN in Figure 4.9 shows visibly different
response plots for Death and Growth Rates. The values for Growth Rates
were significantly lower than that of Death Rates for the corresponding
temperature. The Death Rate value at 45 0C being the maximum of the three
temperatures but Growth Rate continues to increase, although at a decreased
rate.
4.3.3 P Model
Table 4.5 Fitted r2 values for Soluble P
Table 4.6 Refitted Growth and Death Rates for Soluble P with r2 values
Figure 4.10 Modelled Soluble P data at 55 0C
Temp Substr Bact SubstUse U D Const Fitted r2
37 2.41 2.86E-05 0.00254 0.233 0.617 25.6 0.9955545 2.63 5.16E-05 0.00284 0.193 0.669 24.1 0.9962655 2.56 8.46E-05 0.00508 0.207 0.961 30.4 0.99766
Average 2.53 5.49E-05 0.00349 0.211 0.749 26.7 0.99590
Temp U D r2
37 0.216 0.862 0.9924945 0.215 0.911 0.9956455 0.202 0.705 0.99316
Average 0.99376
0.00.10.20.30.40.50.60.70.80.91.0
0 5 10 15 20 25 30
Day
P 55 -‐ normalised
Substr
Bact
Grwth
Death
Cum P
Measured

108
The modelled Soluble Phosphorus (P) plot in Figure 4.10 is similar in trend
to the TAN plot in Figure 4.8. Both the simulated and measured P data
commences at an initial value and tends to increase to the end of the 30 day
measurement period while the simulated P has levelled off.
Figure 4.11 Temperature Response for Growth and Death Rates in P modelling
The temperature response plot for P in Figure 4.11 shows the response plots
in the vicinity of each other. Growth and Death Rates at 45 0C have the
maximum value.
4.4 Comparison of model data vs. measured data
CH4 model:
The comparison of modelled and measured CH4 data is shown in Table 4.7.
There is close agreement between the modelled and measured data. Initially,
the measured CH4 volume is slightly less than the predicted (modelled)
values; however towards the end of the experimental period the measured
CH4 volume is higher than the modelled data.
y = -‐7E-‐05x2 + 0.0055x + 0.1052R² = 1
y = -‐0.0015x2 + 0.1278x -‐ 1.8347R² = 1
0.0
0.2
0.4
0.6
0.8
1.0
0.0
0.2
0.4
0.6
0.8
1.0
0 10 20 30 40 50 60
D-‐De
ath
U-‐Growth
Temperature (oC)
Temperature Response -‐ P
U
D
Poly. (U)
Poly. (D)

109
Table 4.7 Comparison of modelled CH4 and measured CH4 data
TAN model:
The comparison of modelled and measured TAN is shown in Table 4.8. Both
the modelled TAN and measured TAN have a close relationship. Similar to
the CH4 data, the measured TAN data is slightly less than the modelled
TAN data at the start of the experimental period. At the end of the
experimental period, the measured TAN data is almost the same as the
modelled TAN data.

110
Table 4.8 Comparison of modelled TAN and measured TAN data

111
P model:
The comparison of measured soluble P data and modelled soluble P data is
shown in Table 4.9. Initially the measured P data is higher than the predicted
(modelled) P data but by the end of the experiment, the data is similar.
Table 4.9 Comparison of modelled P and measured P data

112
4.5 Discussion
The methane model developed in this thesis is markedly different to other
published models (Andrews 1969; Chen and Hashimoto 1978; Hill 1982;
Kleinstreuer and Powegha 1982; Mosey 1983; Moletta, Verrier et al. 1986;
Angelidaki, Ellegaard et al. 1993; Romli 1993; Siegrist, Renggli et al.
1993). These previously published models assume microbial growth as the
limiting step. Microbes grow on substrate and production of biogas in turn is
proportional to the microbial growth on substrates. The initial assumption
that methanogensis was the limiting step in the process appears to be
incorrect. From the results we know that methane production is proportional
to COD reduction. However soluble P and TAN were not proportional to
COD.
The volume of methane obtained was lower than conventional anaerobic
digestion processes. It is known from literature that
1g of COD reduced ~ 0.5 L of CH4 (Chen 1983; (Hill and Barth 1977; Hill
1982)
The average COD reduction in our experimental set up was 7 g. According
to the above equation, approximately 3.5 L of CH4 should be produced but
the average CH4 obtained was approximately 150 mL. This is markedly
lower than the expected value. This could be due to the following reasons.
• One of the reasons for reduced CH4 was leaks in the system.
Considerable volumes of CH4 could have been lost through minute
gaps in the transfer tubes and Schott bottle caps. Approximately 30%
of the methane is estimated to be lost due to leaks in the system.
• Raw piggery effluent obtained from the piggery could have
undergone a significant percentage of anaerobic digestion process
while stored in the piggery sump for 2-3 days before collection.
Therefore only a small part of the COD reduced accounted for the
CH4 production while the remainder must have been transformed
into some other product which we are unaware of at this stage.

113
Further investigation into this was beyond the scope of this PhD
research.
• Presence of other electron acceptors in the system (e.g. oxygen,
sulphate, nitrate, oxidized metals etc) will result in a lower methane
yield because oxidation by bacteria is energetically favoured over
methanogenesis.
The initial lag phase for 3 days during which there was negligible CH4
production could be due to the presence of oxygen in the headspace, which
remained after CO2 was absorbed. The continued production of CH4 even
after 30 days could be due to the fact that some of the less degradable
substances become available as substrates at that time.
A number of differences have been observed in temperature response as
well. Anaerobic digestion microbes normally develop best around human
body temperature (37 0C). Our results indicated that temperature responses
are highest for 45 0C. For CH4, both the growth and death rates peak at
45 0C, growth rate being slightly less than death rate. For soluble P, death
rate peaks at 45 0C, while growth rate is noticeably less than death rate. In
both these cases, the microbes are expiring faster than they are growing. For
TAN death rate peaks at 45 0C, growth rate is almost same as death rate
with growth rate increasing slightly.
4.6 Further Work
This model provides a basis for initiating a mathematical model for the
anaerobic digestion component of the IBS. Further work, listed below,
needs to be done in this area.
• The validity of the model should be checked at different input
concentrations of piggery effluent over a wider range. This could not
be accomplished during this research due to the limited availability
of the pilot scale anaerobic digestion system. It is recommended that

114
the model should be validated against data obtained from the pilot
scale anaerobic digestion system for a period of 2 years at least.
• The model should be applied to the large scale IBS, when it is
constructed.
• The model needs to be validated against different wastes and
effluents. The kinetic constants for these different wastes need to be
determined.

115
5 Modelling Commercial Scale Integrated
Biosystems
5.1 Introduction
A mathematical model was developed for the anaerobic digestion
component of the IBS in Chapter 4. The purpose of the anaerobic digestion
model is to predict the values of total ammonia nitrogen (TAN) and soluble
phosphorus (P) at the end of the anaerobic digestion process of raw piggery
effluent. The values of TAN and P (presented in Chapter 3) form the basis
of input to the dynamic model DYRESM CAEDYM, which is used to
model the aquaculture stages of the IBS which will be discussed in detail in
this chapter.
The aquaculture component of the IBS consists of the bioconversion stages
of phytoplankton, zooplankton and fish. Each stage is able to accept the
input effluent from the previous stage and the nutrient load is effectively
reduced so the wastewater forms an appropriate input to the subsequent
stage. Phytoplankton (microalgae) growth is the first component in the
aquaculture stage of the IBS receiving the anaerobically digested piggery
effluent. The role of the phytoplankton is to utilise the nutrients present in
the wastewater, thereby reducing the percentage of nitrogen and phosphorus
in the piggery effluent. The proposed IBS contains two phytoplankton
ponds. The function of the first pond is to receive the piggery wastewater
which is rich in nutrients (N and P) and initiate a high bioconversion of
algae. The function of the second pond is to further reduce the nutrients in
the wastewater before it is transferred to the zooplankton pond. The second
algal pond, in effect, acts as a buffer so that the nutrient levels in the
wastewater are brought down to a threshold level (< 10 mg/L) acceptable
for the zooplankton to survive. High nutrient level in the wastewater will
cause the zooplankton to die, as they are highly sensitive organisms
(Lampert and Taylor 1985).

116
Zooplankton is the second component in the aquaculture stage of the IBS.
Zooplankton play an important role in water body dynamics, as grazers that
control phytoplankton and bacterial populations, as a food source for higher
trophic levels and in the excretion of dissolved nutrients (Bruce, Hamilton et
al. 2006). Zooplankton grazing on phytoplankton can transfer more than
50% of carbon fixed by algae to higher trophic levels (Scavia 1980).
Zooplankton excretion strongly influences trophic dynamics in ecosystems
by contributing inorganic N and P for primary and bacterial production
(Lehman 1980; Wen and Peters 1994; Gilbert 1998). The factors controlling
zooplankton growth include temperature, zooplankton and phytoplankton
biomass and species composition, internal nutrient ratios and mixing
regimes (Hudson and Taylor 1996; Hudson, Taylor et al. 1999).
In the IBS, the population of zooplankton is critical as this will determine
the bioconversion of algae resulting in reduction of algae and nutrients in
the wastewater. The impact of zooplankton grazing on phytoplankton
biomass and the significance of this impact has been the focus of many
investigations (Hodgkin and Rippingale 1971; Rippingale and Hodgkin
1974a; Rippingale and Hodgkin 1974b; Rippingale and Hodgkin 1977;
Griffiths and Caperon 1979; Stearns, Litaker et al. 1987; Svensson and
Stenson 1991; Cyr and Pace 1992; Rippingale 1994). There has been ample
research conducted on the role of zooplankton in controlling the population
of algae through grazing (Harris 1986; De Melo, France et al. 1992; Boon,
Bunn et al. 1994). There is evidence from past studies that the depletion of
phytoplankton biomass is dependent on the density and size of the
zooplankton grazers (Martin 1970; Gamble 1978; Lampert and Taylor 1985;
Vanni 1987; Turner and Granelli 1992), diurnal variation in zooplankton
feeding rates (McAllister 1971; Mackas and Bohrer 1976; Lampert and
Taylor 1985; Peterson, Painting et al. 1990) and phytoplankton abundance
(Mullin and Brooks 1970; Frost 1972; Reeve and Walter 1977; Ambler
1986; Durbin and Durbin 1992).
Numerous models have previously been used to evaluate different aspects of
zooplankton dynamics in lakes (Scavia 1980; Lunte and Leucke 1990;

117
Krivtsov, Goldspink et al. 2001; Chen, Ji et al. 2002; Hongping and Jianyi
2002; Ji, Chen et al. 2002; Håkanson and Boulion 2003; Rukhovets,
Astrakhantsev et al. 2003; Bruce, Hamilton et al. 2006), reservoirs (Osidele
and Beck 2004; Romero, Antenucci et al. 2004), and estuaries (Griffin,
Herzfeld et al. 2001; Robson and Hamilton 2004). These models range from
simple mass balances to complex simulation tools using a large number of
parameters in hydrodynamic processes (Robson and Hamilton 2004). Some
models provide more detailed information on the spatial and temporal
changes in the nutrients between different trophic levels in a large water
body which is possible with the availability of field or laboratory data.
These models can be used to predict how the fluxes change in response to
the different environmental factors (Bruce, Hamilton et al. 2006)
Fish ponds form the final component in the aquaculture stage of the IBS as
fish are sensitive to nutrient pollution. The function of the fish ponds is to
receive the wastewater from the zooplankton ponds and promote fish
growth by bioconversion of fish utilising the zooplankton and nutrients from
the zooplankton pond, leaving cleaner water which can be reused in the
piggery or other livestock enterprises for flushing or used in other
enterprises like horticulture.
The development of the mathematical model for the aquaculture stage of the
IBS is discussed in detail in the subsequent sections.
5.1.1 Model Description
The dynamic DYRESM CAEDYM model developed at the Centre for
Water Research, University of Western Australia (www.cwr.uwa.edu.au),
which has previously been used for mathematical modelling of water bodies
(Hamilton and Schladow 1997; Bruce, Hamilton et al. 2006; Burger,
Hamilton et al. 2008) was used for this study. DYRESM is a one
dimensional model which resolves around vertical distribution of
temperature, salinity and density in lakes and reservoirs based on a dynamic
Lagrangian structure. It simulates the lake as horizontally uniform layers
that expand and contract in response to heat, mass and momentum
exchanges (Imberger 1981; Gal, Imberger et al. 2003). DYRESM has been

118
coupled to the ecological model CAEDYM which can simulate up to seven
phytoplankton groups, five species of zooplankton, three species of fish,
dissolved oxygen (DO), organic and inorganic nitrogen, phosphorus and
carbon, using a series of partial differential equations that are characterized
by rate constants (Robson and Hamilton 2004). These rate constants are
defined by the user and vary in the model in response to other
environmental variables (e.g. temperature, DO, pH). The theoretical
framework for DYRESM – CAEDYM and its applications to lake, estuarine
and water bodies are disclosed in multiple citations (Hamilton and
Schladow 1997; Robson and Hamilton 2004; Romero, Antenucci et al.
2004).
DYRESM CAEDYM has been previously used to model natural water
bodies (e.g. lakes, ponds, rivers etc) which have significant depths (e.g. >10
m). There isn't any information available where DYRESM CAEDYM has
been used to model phytoplankton, zooplankton and fish growth and
bioconversion in artificial ponds of depth less than 5 m (e.g. the IBS used in
this PhD study). This focus in this chapter is to assess the suitability of
DYRESM CAEDYM as a modelling tool for an IBS.
5.2 Methods
5.2.1 Research Site
The commercial scale IBS was proposed to be constructed at the
Roseworthy Campus of The University of Adelaide, Roseworthy (340 31‘
60S, 1380 43‘ 60E) situated 51 km north of Adelaide, South Australia.

119
Figure 5.1 Schematic of the proposed commercial scale IBS
Experimental data from the pilot scale anaerobic digestion system and
mesocosm aquaculture set up were used as inputs for DYRESM CAEDYM.
These inputs were then used in the model set up for the proposed
commercial scale IBS and it was tested to assess the suitability of DYRESM
CAEDYM as an effective modelling tool for the IBS. The outputs from the
model would then provide guidance for the operational control and
management for the proper functioning of the commercial scale IBS system.
5.2.2 Alterations to the original source code of DYRESM CAEDYM
Application of DYRESM CAEDYM to evaluate aquatic management
strategies have been widely applied to lakes, reservoirs, rivers, estuaries and
coastal zones. These water bodies cover a significant area and have a large
depth. The program for DYRESM CAEDYM is developed in order to
Algae Pond
Zooplankton
Pond
Fish Pond
Anaerobically digested
piggery effluent
Clean
Water

120
succesfully simulate water bodies of depths greater than 5 m. Lake Rotorua,
(area 79 km2, mean depth 10.8 m) is the shallowest water body modelled
using DYRESM CAEDYM reported in literature (Burger, Hamilton et al.
2008).
The ponds proposed to be constructed as part of the IBS had an area of 100
m2 and mean depth 1.0 m. Hence the initial trial simulations crashed as the
model could not successfully run simulations for such a shallow depth.
After discussions with other researchers using the same model, a need to
modify the original source code of DYRESM CAEDYM was identified.
5.2.2.1 Compilation of the source code
During a visit to the Centre for Water Research (CWR), University of
Western Australia (UWA), Perth in November 2007, the source code for the
model was obtained from the UWA researchers. The source code was
compiled and built using Intel Visual FORTRAN 10.1 on Microsoft Visual
Studio 2005 making use of instructions posted on the CWR Model User’s
Forum. The purpose of compilation of the source code was to create an
executable code to run the model. This executable code could be re created
multiple times by altering the source code which would be suitable for
running simulations for the large scale IBS. The new executable code was
evaluated by running the test files to ensure that there was no error in
compilation.
5.2.2.2 Breakpoints
It was observed that the program crashed while executing the subprogram
dyconsts.f90. The dyconsts.f90 file contains parameter constants required to
compile the model simulations successfully.
In the dyconsts.f90 file the following parameters were changed.
• AREA_HT_DELTA_Z: This parameter deals with the z-grid spacing
used for determining the coefficients alpha and beta used to create
piecewise smooth function of area vs. height.
• COEFFS_TBL_DELTA_Z: This parameter deals with the z-grid
spacing used in determining the area height function coefficients.

121
• INTERP_DELTA_Z: This parameter deals with the z-grid spacing used
for interpolating cumulative volumes and layer surface areas.
• MIN_GRID_THICK: This parameter deals with the minimum allowed
grid thickness for surface layer re grid.
These parameters represent the size of the discrete intervals describing the
grid array that relates grid heights to sediment area. Data from the
morphology file and the above mentioned parameters are interpolated
during simulations to produce a finer grid, which can be used in DYRESM
to relate the different layers to a specific sediment area. The formation of
the simulation grid is dependent on the depth of the water body which is to
be simulated. As the depth of the water body is decreased, the value of the
above parameters is also decreased which gives a higher vertical grid
resolution necessary to fit in the shallow depth. This is necessary to run the
simulations successfully.
5.2.3 Input Data
5.2.3.1 Meteorological Data
Meteorological data is one of the key driving forces to run DYRESM
CAEDYM. Daily meteorological data required as input to the DYRESM
CAEDYM model was taken from the Roseworthy Automatic Weather
Station [AWS Bureau of Meteorology Station 023122] situated at the
Roseworthy Campus, The University of Adelaide. Data included short wave
radiation, daily averages of air temperature, total daily rainfall and wind
speed. The closest cloud cover observations were taken at Rosedale, 10 km
to the east. Daily averages of relative humidity (RH) and air temperature
were used to derive the mean daily water vapour pressure input to the model
(Antenucci and Imerito 2002).
The meteorological data input file requires vapour pressure data as one of
the essential input parameters. The meteorological data file obtained from
the Roseworthy AWS had data for relative humidity and air temperature
instead of vapour pressure. Hence vapour pressure was calculated using the

122
formula given in the DYRESM User Science Manual (Imerito 2007). The
details of the equation are given below.
Vapour Pressure Based on Relative Humidity and Air Temperature:
⎥⎥⎦
⎤
⎢⎢⎣
⎡⎟⎟⎠
⎞⎜⎜⎝
⎛+⎟⎟⎠
⎞⎜⎜⎝
⎛
+⎟⎠
⎞⎜⎝
⎛= cbqqahe
D
Da
*303.2exp100
Eq. 5.1 (T.V.A. 1972)
where
• ea = vapour pressure [hPa]
• h = relative humidity of air [%]
• qD = dry bulb air temperature [0C]
and the coefficients for over water calculations are
a = 7.5
b = 237.3
c = 0.7858
Vapour Pressure is one of the inputs in the meteorological file required to
run the DYRESM simulations. Since this data was not available from the
Roseworthy Agricultural Weather Station [AWS Station 023122], the above
formula was used to calculate vapour pressure using relative humidity and
air temperature.
The daily meteorological data collected over a period from 1st June 2006 till
31st May 2007 is presented in Figure 5.2.

123
Figure 5.2 Meteorological data comprising short wave radiation, air temperature, rainfall, wind speed, cloud cover and vapour pressure
(clockwise, starting from left)
5.2.3.2 Inflow and Outflow Data
The phytoplankton growth model had one inflow of 0.2 m3 day-1 of
anaerobically digested piggery effluent. The zooplankton growth model had
0
50
100
150
200
250
300
350
400
450
13 May 2006 10 October 2006 9 March 2007 6 August 2007
SW (W/m
2)
Date
0
5
10
15
20
25
30
35
40
13 May 2006 10 October 2006 9 March 2007 6 August 2007
Air Temperature (0C)
Date
00.0050.010.0150.020.0250.030.0350.040.0450.05
13 May 2006 10 October 2006 9 March 2007 6 August 2007
Rainfall (m)
Date
0
2
4
6
8
10
12
14
13 May 2006 10 October 2006 9 March 2007 6 August 2007
Wind Speed (m/s)
Date
0
0.2
0.4
0.6
0.8
1
1.2
13 May 2006 10 October 2006 9 March 2007 6 August 2007
Cloud Cover (%
)
Date
0
5
10
15
20
25
30
13 May 2006 10 October 2006 9 March 2007 6 August 2007
Vapour Pressure (mb)
Date

124
one inflow of 0.2 m3 day-1 from the algae pond. The fish growth model had
one inflow of 0.2 m3 day-1 from the zooplankton pond.
Water quality variables for the model inflow in each of the phytoplankton,
zooplankton and fish models included daily estimates of water temperature
(0C), dissolved oxygen (mg L-1), PO4 (mg L-1), NH4 (mg L-1) and pH.
The outflow data consisted of withdrawal of 0.2 m3 day-1 of wastewater
from each of the phytoplankton, zooplankton and fish ponds.
5.2.3.3 Initial Profile
The initial profile consisted of temperature and salinity at depths of 0.2 m,
0.4 m, 0.6 m, and 0.8 m in the water column in each of the phytoplankton,
zooplankton and fish ponds.
5.2.3.4 CAEDYM Water Quality parameters
The dynamics of three phytoplankton groups, represented by equivalent
chlorophyll-a concentration, were simulated in the model; chlorophytes,
cyanobacteria and freshwater diatoms. Phytoplankton parameters for these
groups were assigned based on literature values (Lewis, Brookes et al. 2004;
Robson and Hamilton 2004).
The dynamics of one zooplankton group, ZOOP1, represented by equivalent
gm C m-3, were simulated in the model. Zooplankton parameters for these
groups were assigned based on literature values (Bruce, Hamilton et al.
2006).
The dynamics of one fish group, FISH1, represented by equivalent gm C m-
3, were simulated in the model.

125
5.3 Results
Model simulations were conducted over a one year period from 1st June
2006 till 31st May 2007.
5.3.1 Temperature
Figure 5.3 Simulated pond temperature (all ponds are the same temperature)
The variation of temperature in a pond is shown in Figure 5.3, since the
algal, zooplankton and fish ponds all had same temperature as a result of
being shallow. The average temperature of wastewater in all the ponds
reached a maximum of approximately 400 C during summer and a minimum
of 100 C in winter. This output appears reasonable with the air temperatures
during the year where the summer temperatures reach a maximum of 35 0C
and the winter temperatures reach a minimum of 5 0C.
0
5
10
15
20
25
30
35
40
45
24-‐Mar-‐06 02-‐Jul-‐06 10-‐Oct-‐06 18-‐Jan-‐07 28-‐Apr-‐07 06-‐Aug-‐07
Tempe
rature (d
egree C)
Day
Temperature

126
5.3.2 Phytoplankton growth in algal pond 1
Figure 5.4 Simulated chlorophyte growth in algal pond 1
0
10
20
30
40
50
60
70
80
90
100
24-‐Mar-‐06 02-‐Jul-‐06 10-‐Oct-‐06 18-‐Jan-‐07 28-‐Apr-‐07 06-‐Aug-‐07
Chloroph
yte (ug/L)
Day
Chlorophyte
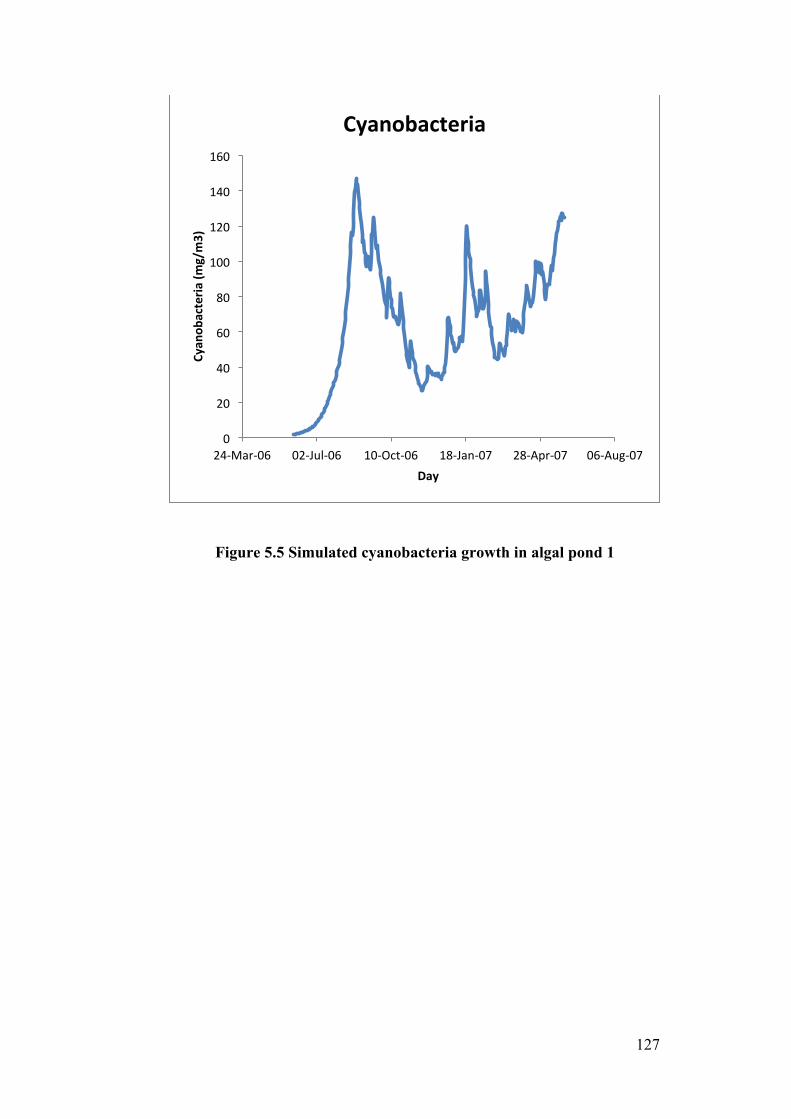
127
Figure 5.5 Simulated cyanobacteria growth in algal pond 1
0
20
40
60
80
100
120
140
160
24-‐Mar-‐06 02-‐Jul-‐06 10-‐Oct-‐06 18-‐Jan-‐07 28-‐Apr-‐07 06-‐Aug-‐07
Cyan
obacteria
(mg/m3)
Day
Cyanobacteria

128
Figure 5.6 Simulated freshwater diatoms growth in algal pond 1
Simulated chlorophyte growth in algal pond 1 is shown in Figure 5.4. The
chlorophytes initially grow to a maximum of 90 µg (chlorophyll-a) L-1 in
the first three months of simulation followed by a sharp decrease and almost
nil growth in summer followed by a steady concentration of 30 µg
(chlorophyll-a) L-1 for the remainder of the simulation period. Simulated
cyanobacteria growth in algal pond 1 is shown in Figure 5.5. The growth of
cyanobacteria spikes up to a maximum of 200 mg m-3 after the first month
of simulation coinciding with the decrease in concentration of the
chlorophytes. Cyanobacteria growth decreases for a brief period following
the spike and increases at a steady rate for the remainder of the simulation
period.
Growth of fresh water diatoms in algal pond 1 is shown in Figure 5.6, which
is almost negligible for the year. It is evident from the simulation plots that
cyanobacteria tend to dominate algal pond 1 throughout the entire
simulation period.
0
0.05
0.1
0.15
0.2
0.25
0.3
0.35
0.4
0.45
0.5
24-‐Mar-‐06 02-‐Jul-‐06 10-‐Oct-‐06 18-‐Jan-‐07 28-‐Apr-‐07 06-‐Aug-‐07
Fresh Water Diatoms (mg/m3)
Day
Fresh Water Diatoms

129
Figure 5.7 Nutrient Profile in Algal Pond 1
The nutrient concentration in algal pond 1 is shown in Figure 5.7. The Total
Ammonia Nitrogen (TAN) concentration is denoted as NH4 concentration.
The NH4 concentration of the anaerobically digested piggery effluent which
forms an influent into the first algal pond of the IBS is 1600-1700 mg/L.
The average NH4 concentration in the algal pond 1 at the end of the
simulation period is approximately equal to 200 mg/L; therefore a reduction
of ~88% is obtained in the first module of the IBS. The soluble phosphate
concentration, denoted by PO4, is approximately 90 mg/L at the end of the
simulation period, whereby a reduction of ~40% is obtained, as the soluble
phosphate concentration in the anaerobically digested piggery effluent is
approximately 150 mg/L. The total phosphate, TP, concentration is
overlapped with the PO4 concentration, which implies that majority of the
phosphate in the effluent is in the soluble form after anaerobic digestion has
occurred in the piggery effluent. Both the nitrate (NO3) and the Total
Nitrogen (TN) concentration is showing an increasing trend at the end of the
simulation period, the values of TN being approximately 570 mg/L and NO3
being 360 mg/L as seen from the above figure.
0"
100"
200"
300"
400"
500"
600"
13/05/2006" 22/07/2006" 30/09/2006" 09/12/2006" 17/02/2007" 28/04/2007" 07/07/2007"
Nutrie
nt(Con
centra-o
n((m
g/l)(
Date(
Nutrient(Profile(in(Algal(Pond(1(
NH4"
NO3"
TN"
TP"
PO4"
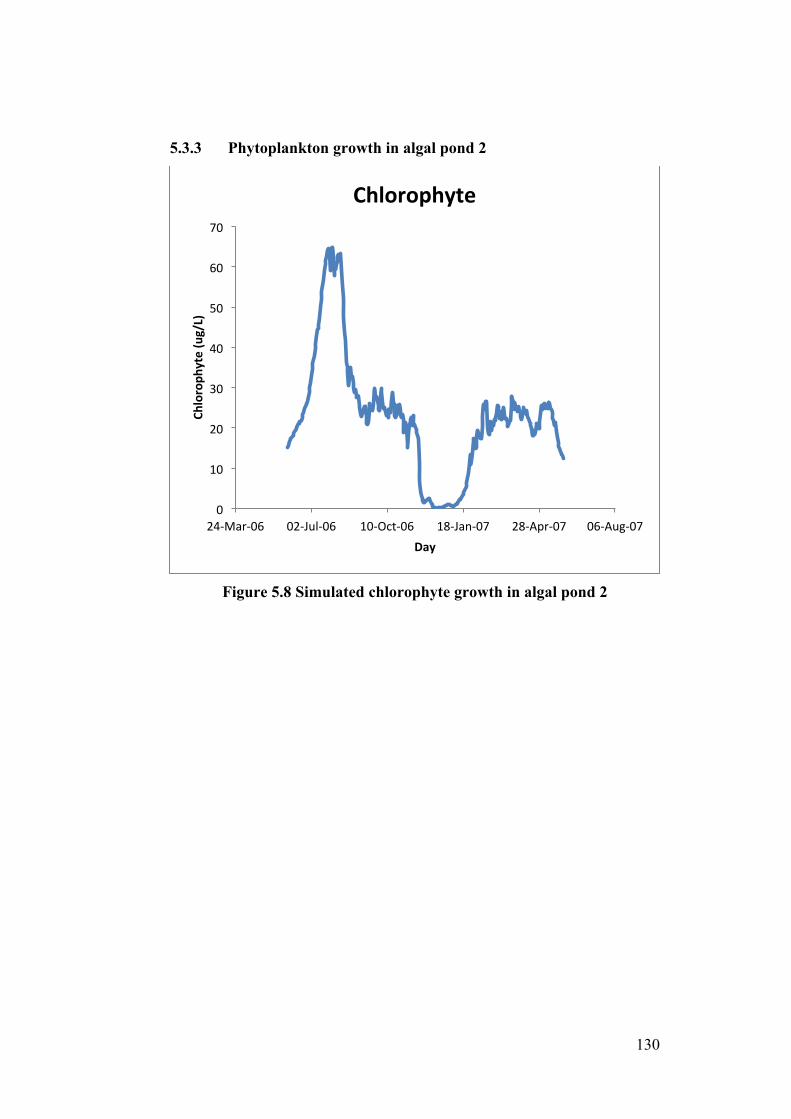
130
5.3.3 Phytoplankton growth in algal pond 2
Figure 5.8 Simulated chlorophyte growth in algal pond 2
0
10
20
30
40
50
60
70
24-‐Mar-‐06 02-‐Jul-‐06 10-‐Oct-‐06 18-‐Jan-‐07 28-‐Apr-‐07 06-‐Aug-‐07
Chloroph
yte (ug/L)
Day
Chlorophyte

131
Figure 5.9 Simulated cyanobacteria growth in algal pond 2
0
20
40
60
80
100
120
140
160
180
24-‐Mar-‐06 02-‐Jul-‐06 10-‐Oct-‐06 18-‐Jan-‐07 28-‐Apr-‐07 06-‐Aug-‐07
Cyan
obacteria
(mg/m3)
Day
Cyanobacteria

132
Figure 5.10 Simulated fresh water diatoms growth in algal pond 2
Chlorophyte growth in algal pond 2 is shown in Figure 5.8. It follows a
similar trend to that as in algal pond 1. The chlorophyte grows initially to a
maximum of 60 µg (chlorophyll-a) L-1 in the first three months of
simulation, then drops off to almost nil growth during the summer months,
and then increases to approximately 22 µg (chlorophyll-a) L-1 which it
maintains till the end of the simulation. Figure 5.9 shows the cyanobacteria
growth in algal pond 2, which is also similar to that in algal pond 1. Here
the cyanobacteria concentration reaches a maximum of 150 mg m-3. Figure
5.10 shows the growth of freshwater diatoms in algal pond 2, which is again
negligible. Cyanobacteria tend to dominate algal pond 2, similar to algal
pond 1.
0
0.05
0.1
0.15
0.2
0.25
0.3
0.35
0.4
0.45
0.5
24-‐Mar-‐06 02-‐Jul-‐06 10-‐Oct-‐06 18-‐Jan-‐07 28-‐Apr-‐07 06-‐Aug-‐07
Fresh Water Diatoms (mg/m3)
Day
Fresh Water Diatoms
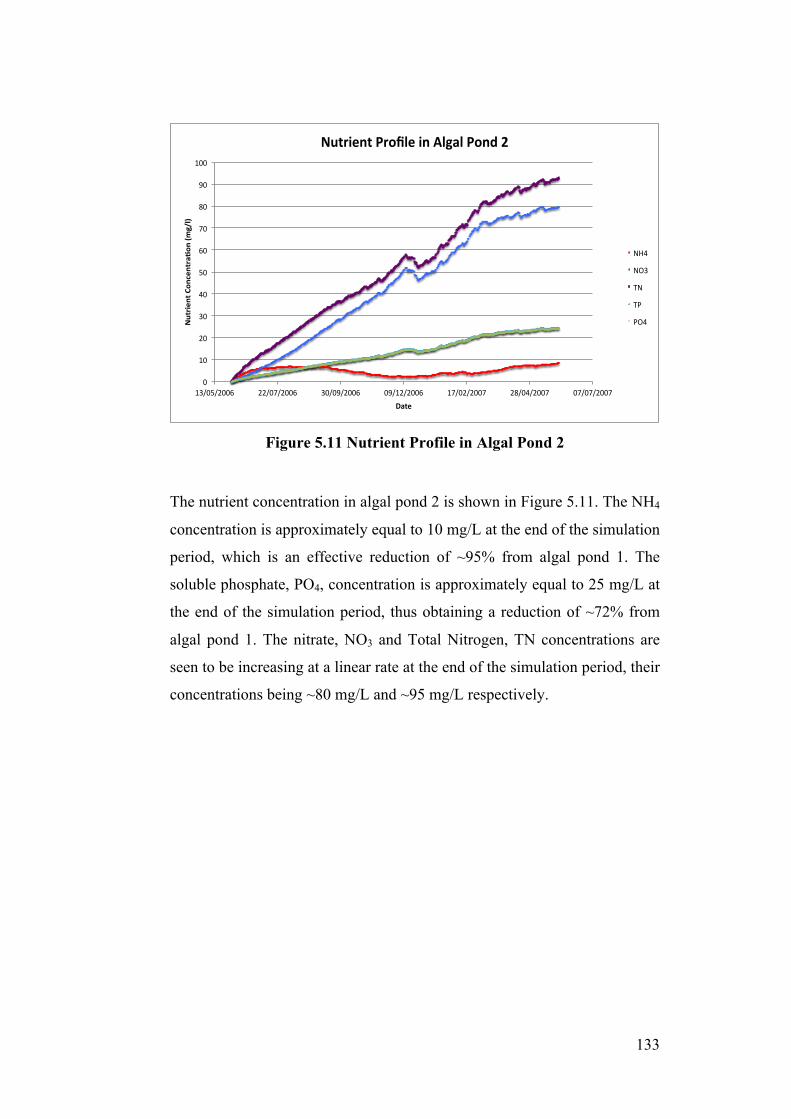
133
Figure 5.11 Nutrient Profile in Algal Pond 2
The nutrient concentration in algal pond 2 is shown in Figure 5.11. The NH4
concentration is approximately equal to 10 mg/L at the end of the simulation
period, which is an effective reduction of ~95% from algal pond 1. The
soluble phosphate, PO4, concentration is approximately equal to 25 mg/L at
the end of the simulation period, thus obtaining a reduction of ~72% from
algal pond 1. The nitrate, NO3 and Total Nitrogen, TN concentrations are
seen to be increasing at a linear rate at the end of the simulation period, their
concentrations being ~80 mg/L and ~95 mg/L respectively.
0"
10"
20"
30"
40"
50"
60"
70"
80"
90"
100"
13/05/2006" 22/07/2006" 30/09/2006" 09/12/2006" 17/02/2007" 28/04/2007" 07/07/2007"
Nutrie
nt(Con
centra-o
n((m
g/l)(
Date(
Nutrient(Profile(in(Algal(Pond(2(
NH4"
NO3"
TN"
TP"
PO4"

134
5.3.4 Zooplankton growth
Figure 5.12 Simulated zooplankton growth in zooplankton pond
Zooplankton growth is shown in Figure 5.12. Peaks in zooplankton biomass
occurred at periodic intervals. The biomass peak in the zooplankton growth
occurred after a decrease in chlorophyte growth. This pattern can be
explained by a phenomenon called the Lotka-Volterra predator-prey cycle
(Krebs 1985) where the prey biomass increases and a subsequent crash,
followed by a predator biomass increase. This predator-prey pattern has
been widely reported in literature for the past few years.
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
24-‐Mar-‐06 02-‐Jul-‐06 10-‐Oct-‐06 18-‐Jan-‐07 28-‐Apr-‐07 06-‐Aug-‐07
Zoop
lank
ton (g C/m
3)
Day
Zooplankton

135
Figure 5.13 Nutrient Profile in Zooplankton Pond
The nutrient concentration in the zooplankton pond is shown in Figure 5.13.
The NH4 concentration starts of at approximately 0.5 mg/L during the initial
stages of the simulation, then reduces to almost 0 mg/L in the middle of the
simulation period and then increases to 0.5 mg/L towards the end of the
simulation period. A reduction of ~95% is obtained in the zooplankton pond
for NH4 concentration. The PO4 concentration increases almost linearly
during the period of the simulation and attains a value of approximately 7.5
mg/L, an effective reduction of ~70% since being transferred from algal
pond 2. The NO3 concentration also remains steady at a value of ~2.1 mg/L.
0"
1"
2"
3"
4"
5"
6"
7"
8"
13/05/2006" 22/07/2006" 30/09/2006" 09/12/2006" 17/02/2007" 28/04/2007" 07/07/2007"
Nutrie
nt(Con
centra-o
n((m
g/l)(
Date(
Nutrient(Profile(in(Zooplankton(Pond(
NH4"
NO3"
PO4"

136
5.3.5 Fish growth
Figure 5.14 Simulated fish growth in fish pond
Fish growth is shown in Figure 5.14. Initially there was a decline in fish
growth for the first three months; growth then increased to 0.6 g (C) m-3
after which there was a decrease during the summer months, and finally fish
growth increased to a maximum of 0.8 g (C) m-3 before stabilizing till the
end of the simulation run.
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
24-‐Mar-‐06 02-‐Jul-‐06 10-‐Oct-‐06 18-‐Jan-‐07 28-‐Apr-‐07 06-‐Aug-‐07
Fish (g C/m
3 )
Day
Fish

137
Figure 5.15 Simulated zooplankton growth in fish pond
The amount of zooplankton in the fish pond is shown in Figure 5.15. The
zooplankton quantity drops dramatically and stays at a minimum level for
the remainder of the simulation period. This output is in agreement and
corresponds with the fish growth as shown in Figure 5.14, where fish
growth shows an increase at the end of the simulation period.
0
0.05
0.1
0.15
0.2
0.25
24-‐Mar-‐06 02-‐Jul-‐06 10-‐Oct-‐06 18-‐Jan-‐07 28-‐Apr-‐07 06-‐Aug-‐07
Zoop
lank
ton (g C/m
3 )
Day
Zooplankton

138
Figure 5.16 Nutrient Profile in Fish Pond
The nutrient concentration in the fish pond is shown in Figure 5.16. The
NH4 concentration increases to approximately 0.2 mg/L for a brief period at
the start of the simulation and remains at 0 mg/L for the maximum period of
simulation, before finally rising to approximately 0.1 mg/L towards the end
of the simulation, thereby achieving an effective reduction of ~80 % from
the zooplankton pond. The NO3 concentration is also almost 0 mg/L during
the simulation period and slightly increases to 0.25 mg/L at the end. The
PO4 concentration increases linearly similar to that in the zooplankton pond,
and attains an approximate value of 3.25 mg/L at the end of the simulation
period, thus achieving a reduction of ~56.7%.
0"
0.5"
1"
1.5"
2"
2.5"
3"
3.5"
13/05/2006" 22/07/2006" 30/09/2006" 09/12/2006" 17/02/2007" 28/04/2007" 07/07/2007"
Nutrie
nt(Con
centra-o
n((m
g/l)(
Date(
Nutrient(Profile(in(Fish(Pond(
NH4"
NO3"
PO4"

139
5.4 Discussion
The hydrodynamic and ecological modelling software DYRESM CAEDYM
has been extensively applied to model phytoplankton, zooplankton and fish
growth in water bodies having large depths and surface areas especially
reservoirs, lakes, estuaries and ponds. This study reported in Chapter 5 has
applied the model to simulate phytoplankton, zooplankton and fish growth
in artificially made shallow ponds with a mean depth of 1.0 m. The
parameters used in the source code of the model were altered to successfully
run the model for the IBS ponds. This is a significant achievement in
modelling water bodies of depth <5 m using DYRESM CAEDYM which
has not been done before. The simulated data obtained from this modelling
exercise is based on data obtained from pilot scale and mesocosm
experiments. The model does not incorporate any control management
measures. However the results obtained from this modelling study would
help us in deciding the required management and control measures which
need to be incorporated in order to obtain favourable results.
The phytoplankton, zooplankton and fish growth was simulated using
weather data for the period 1st June 2006 to 31st May 2007. A year’s worth
of meteorological data was used to obtain credible model outputs. The pilot
scale and mesocosm experiments were conducted in extremely temperature
controlled environments. The meterological data provided seasonal
variation input into the model for simulating the IBS which is exposed to
ambient conditions.
The growth of chlorophyte (Chlorella) occurred during the start of the
simulation period in June 2006 but declined as the simulation period
progressed. Chlorophytes reached a maximum concentration of 90 µg (chl-
a) L-1 and 60 µg (chl-a) L-1 in the first and second algal ponds respectively.
The decrease in chlorophyte growth is dominated by cyanobacteria. During
the summer period, there is practically negligible growth of chlorophytes.
The growth of cyanobacteria (Anabaena circinalis) occurs during periods
when there is permanent stratification, which normally does not occur in

140
commercial IBS with effective management and control measures in place.
It is evident from Figure 5.5 that the growth of cyanobacteria occurs during
late spring and summer when there is high insolation. The cyanobacteria
concentration reaches a maximum of 200 mg m-3 and 150 mg m-3 in the first
and second algal ponds respectively. The cyanobacteria concentration
remains constant for the rest of the simulation period.
Cyanobacteria have a higher affinity for nitrogen and phosphorus than other
photosynthetic organisms and can fix atmospheric nitrogen (Boon, Bunn et
al. 1994; Bergman, Gallon et al. 1997; Mur, Skulberg et al. 1999; Ute 2003;
Fiore, Neilan et al. 2005; Hense and Beckmann 2006; Marino and Howarth
2009; Hense and Burchard 2010; Jonathan P 2011; Paerl, Xu et al. 2011).
Cyanobacteria have a growth rate less than that of green algae; however, at
very low light intensities their growth rate is higher. When the turbidity of
the water body is high or at low CO2/ high pH, they have a better chance of
out-competing other species in the water environment, especially when the
concentrations of nitrogen and phosphorus is limited (Mur, Skulberg et al.
1999). Cyanobacteria synthesise and secrete large quantities of
polysaccharides from their cells. This protective coating enables them to
withstand stress brought about by lack of water and high temperatures.
Available phosphorus appears to be a critical factor to stimulate rapid
growth of cyanobacteria during the summer months. Cyanobacteria often
exploit a combination of these features to dominate the water body
(Kardinaal and Visser 2005). Cyanobacterial blooms can flood the water
environment with the biotoxins which they produce. These toxins modify
zooplankton communities and interfere with the development of fish.
Freshwater diatom growth in the IBS ponds was almost negligible. In open
water bodies, the condition that causes the diatom blooms to end is a lack of
silicon. Unlike other nutrients, this is the only major requirement of diatoms
so it is not regenerated in the plankton ecosystem as efficiently as nitrogen
or phosphorus. As the nutrients (N and P) decline along gradients, silicon is
usually the first to be diminished (followed by N and P) (Egge and Aksnes

141
1992). It was found that diatom dominance of mesocosm communities was
directly related to the availability of silicic acid (Egge and Aksnes 1992).
Zooplankton and fish growth occurred at periodic intervals. This is due to
lack of chlorophyte growth, which provides food for zooplankton. A
number of studies have been conducted on modelling zooplankton and fish
growth in water bodies (Vojtƒõch 1987; Levine, Borchardt et al. 1999;
Carlotti, Giske et al. 2000; Ray, Berec et al. 2001; Håkanson and Boulion
2003; Hunt and Matveev 2005; Freund, Mieruch et al. 2006; Mitra and
Flynn 2006; Jefferson T 2010). There is limited literature available on
modelling of zooplankton and fish growths using DYRESM CAEDYM.
Table 5.1 Nutrient Concentrations in different stages of the IBS
The nutrient concentrations in different stages of the IBS is shown in Table
5.1. An overall reduction of 99.99% for NH4 is obtained while a reduction
of 97.88% for PO4 is obtained. These nutrient reduction data are extremely
ideal which would be expected of an IBS. The NH4 concentrations in both
the zooplankton and fish ponds is extremely low, which is suitable for the
growth of zooplankton and fish as these organisms are highly sensitive to
ammonia concentrations in effluent.
Nutrient (mg/l)
Anaerobic Digestion
Algal Pond 1
Algal Pond 2
Zooplankton Pond Fish Pond
NH4 1600 200 10 0.5 0.1% reduction - 88 95 95 80
PO4 150 90 25 7.5 3.25% reduction - 40 72 70 56.7

142
5.4.1 Limitations of the model
The model developed for the aquaculture component of the IBS has certain
limitations.
• The IBS model developed is hypothetical. The commercial scale IBS
could not be constructed at the time this PhD study was conducted.
As a result, the data for validating the model was not available to the
researcher. This proved to be a considerable set back in determining
the validity of the model.
• Parameters which could account for effective management and
control of the IBS has not been accounted for in this model as this
model has used inputs based on experiments conducted in the pilot
and mesocosm stages.
• DYRESM CAEDYM requires both inflow and outflow volumes for
the model to run successfully. The values of inflow and outflow
need to be manually set by the user. However in real life modelling
situations, it is desirable for the user to input only the inflow volume
and a desired depth of the ponds. The outflow volume, water quality
parameters and the bioconversion rates should be automatically
calculated using the input values. Also this model accounts for
controlling the input and output volumes initially by manual
adjustments which is a tedious task.
• DYRESM CAEDYM is primarily designed to model reservoirs,
lakes and water bodies having depths greater than 5m. In order to
model ponds with depths lesser than that, like that in the IBS, the
source code of the model needs to be altered to make it suitable for
the simulations. This is a tedious task and the person needs to have
sufficient programming skills to compile the source code to be able
to run the simulations. As there was only elementary computer
programming skills present at the time of this study, this research
mainly focussed on testing the suitability of this modelling package
for a commercial scale IBS and ways by which it could be modified
to effectively manage and control the IBS.

143
5.4.2 Comparison with algal data obtained from Bolivar Wastewater
Treatment Plant
Bolivar Wastewater Treatment Plant (WWTP) is one of Adelaide’s major
wastewater treatment plants, situated (Latitude: 34º 41' 28.21" S; Longitude:
138º 35' 12.67" E) 26.7 kms north of Adelaide. The treatment plant
processes on average 135 million litres of household and industrial
wastewater every day. The plant has the capacity to handle the wastewater
treatment needs of 1.3 million people. The wastewater passes through
primary screening, primary clarifier, activated sludge treatment, secondary
clarifier, maturation lagoons, and a final filtration unit. Primary effluent is
pumped to a new activated sludge process for secondary treatment. The
activated sludge treatment is known as a suspended growth biological
treatment process because active microorganisms are maintained in
suspension in the wastewater. Treated wastewater or effluent is decanted
from the surface of the settling clarifier tank and then flows by gravity to
stabilisation lagoons. The six-stabilisation lagoons cover a total area of 347
hectares, providing a nominal detention time of about 30 days, for further
treatment by microorganisms, including algae. The lagoons are
approximately 1.2 m deep. These maturation lagoons are significantly
different in size and the water retention time also differs accordingly. The
lagoon/s, which receive the wastewater directly from clarifiers, have the
highest level of nutrients. The conditioning of effluent includes symbiotic
activity by various micro-organisms. This enables continued nitrification
and de-nitrification in the effluent water, growth of algae and establishment
of an eco-system within the stabilisation lagoons. The maturation lagoons
receive wastewater containing an average ammonia concentration of 5-7
mg/l. During winter period ammonia levels will be about 9 mg/l. The
available information indicates no heavy metal content is present in the
wastewater. A significant proportion of the final effluent from the lagoons is
pumped to the Dissolved Air Floatation and Filtration (DAFF) plant to meet
the irrigation needs of market garden crops in the Virginia area through a
pipe line (Virginia Pipeline Scheme).

144
Due to its proximity to Roseworthy, the climatic conditions of both sites are
relatively similar. In the absence of data for validating the IBS model, raw
data on algal succession was obtained from Mr. Amos (Adelaide University)
for comparison with the simulated algal data for the IBS. The algal growth
data for chlorophytes (Chlorella), cyanobacteria (Microcystis flos aquae)
and fresh water diatoms (Nitzchia) for the period September 1999 to January
2003 is shown in Figure 5.17.
Figure 5.17 Algal succession data obtained from Bolivar Wastewater
Treatment Plant
Figure 5.18 Algal succession data obtained from Bolivar Wastewater
Treatment Plant for the period 2000 - 2001
0
500000
1000000
1500000
2000000
2500000
3000000
3500000
4000000
24/07/1998 6/12/1999 19/04/2001 1/09/2002 14/01/2004
Algae(Cells/ml)
Date
Algal Growth
Chlorella
Microcystis flos aquae
Nitzchia
0
500000
1000000
1500000
2000000
2500000
3000000
3500000
4000000
1/10/00 20/11/00 9/1/01 28/2/01 19/4/01 8/6/01
Algae(Cells/ml)
Date
Algal Growth
Chlorella
Microcystis flos aquae
Nitzchia

145
The algal succession data in Figure 5.17 shows that there is a greater
dominance of cyanobacteria in the wastewater pond as compared to
chlorophyte or fresh water diatoms. The concentration of fresh water diatom
(Nitzchia) is almost negligible while chlorophyte (Chlorella) concentration
reaches a maximum of 500000 cells/ mL between the period 1998-1999
after which cyanobacteria (Microcystis flos aquae) concentration dominates
the pond with a maximum concentration of approximately 4000000 cells/
mL till the end of the data collection period. Algal succession data for one
year (2000-2001) is shown in Figure 5.18. Chlorella and Nitzchia have
negligible concentration during this period. The concentration of
Microcystis flos aquae increases to a maximum during the months of
January 2001 to March 2001, as the conditions for cyanobacteria growth are
favourable during summer. These results assure the researcher that the data
obtained from simulations using DYRESM CAEDYM are similar to the
algal succession data obtained from the Bolivar WWTP, hence determining
the authenticity of the simulations conducted. It is hence proven in both the
modelling outputs and the data obtained from the Bolivar WWTP that
cyanobacteria dominate with reduced growth of chlorophytes, while
freshwater diatoms show negligible growth.
5.5 Typical Outputs from established IBS
5.5.1 Central Institute of Freshwater Aquaculture, India
An aquaculture sewage treatment plant (ASTP) comprising duckweed and
fish culture was designed and set up by the Central Institute of Freshwater
Aquaculture, Kausalyaganga, Bhubaneswar, Orissa, India, in collaboration
with the Public Health Engineering Department, Government of Orissa,
under a project on "Aquaculture as a tool for utilisation and treatment of
domestic sewage" funded by the National River Conservation Directorate,
Ministry of Environment and Forests, Government of India. The ASTP
comprises a set of duckweed ponds where algae and duckweed are used for
the removal of nutrients and for the reduction in BOD and COD levels,
followed by fish and marketing ponds. Duckweed culture before the fish

146
ponds assists in removal of heavy metals and other chemical residues that
otherwise get into the human food chain through cultured fish. The waste to
be treated contained BOD5 levels of about 100 mg/L with a total retention
period of 5 days, with the final effluent BOD5 level brought down to 15-20
mg/L, meeting the required standards of different parameters for discharge
into natural waters.
Table 5.2 Nutrient and Plankton outputs from Central Institute of Freshwater Aquaculture system
From Table 5.2, it is evident that there is a significant increase in the net
plankton levels in both the duckweed and fish ponds. The phytoplankton
and zooplankton percentages are approximately 97% and 99% in the
duckweed ponds and 95% in the fish ponds. There is also a significant
reduction in ammonium-N and phosphate-P when compared with the source
effluent and the outlet effluent. This shows that the IBS set up at the Central
ParameterSource
EffluentDuckweed
Ponds Fish PondsOutlet
EffluentAmmonium-N
(µg N/l)Nitrate-N (µg N/l)Nitrite-N (µg N/l)
Phosphate-P (mg/l)BOD5 (mg/l)COD (mg/l)
Net Plankton (no./l)
Phytoplankton (%) Nil 0.75-97.83 4.22-95.43 Nil
Zooplankton (%) Nil 1.56-99.25 4.57-95.78 Nil
Nil 263-25079 37-18624 Nil
140-152 72-86 30-36 18-22
180-200 160-172 58-64 32-52
0.56-5.48 0.28-12.22 0.35-14.40 0.24-4.46
0.20-5.50 0.10- 4.0 0.10-4.6 0.04-2.60
21.9-556.6 9.8-339.38 5.51-177.5 7.8-156.8
0.58-54.0 0.11-34.26 0.56-63.30 0.29-11.26

147
Institute of Freshwater Aquaculture has been successful in reducing the
nutrient levels in the effluent and at the same time increasing the
bioconversion growth rates of phytoplankton and zooplankton.
5.5.2 IBS set up in France
A pilot plant (experimental lagoon system) on a small scale was developed
for biotreatment of swine manure with the production of algae, zooplankton
and fish (Sevrin-Reyssac 1998). The objective of this plant was to test the
efficiency of an IBS in a temperate climate and produce valuable biomasses.
An experimental lagoon system with a total area of 2100 m2 was set up 100
km east of Paris. It consisted of two algal ponds, two zooplankton ponds and
a fish pond working in series and in a closed recycling circuit. Swine
manure containing ammonia and phosphates as main nutrients were used by
microalgae, which are in turn consumed by zooplankton, and the latter being
consumed by fish. The functioning of this system was studied for a year.
It was observed that during winter there was a reduction in algal
productivity and the algal production collapsed in summer. The average
algal productivity was 0.68 g d.m m-2 d-1 during winter, which was due to
reduced removal of nutrients from the effluent (less than 200 mg d.m L-1).
The algal productivity in summer was between 6-9 g d.m m-2 d-1 for only a
few days and then there was a collapse due to predation by rotifers.
During winter, the low algal productivity can be explained by inorganic
carbon limitation due to high pH values. Zooplankton (daphnids) production
was abundant in winter but collapsed in summer due to a crash in the algal
production. Fish production was also reduced during summer. The
concentration of rotifers in the algal ponds was very high after algal
collapse.

148
These outputs are similar to that obtained from the simulated studies for the
proposed commercial IBS. The comparison of data between this system in
France and the commercial scale IBS is valid due to the following reasons;
the pilot plant in France is a lagoon system with an area of 2100 m2, with a
maximum fish pond size of 250 m2; the climatic conditions at both the
experimental sites in France and Roseworthy are temperate; and the
experimental set up in France has very little effective control and
management tools in place, which is similar to the set up used for our
modelling studies. The aim of this PhD research is to determine the
suitability of DYRESM CAEDYM as a modelling tool to model the
commercial scale IBS and determine what effective control and
management tools need to be incorporated into the IBS. This is also similar
to the pilot plant study in France where the initial simulations were
conducted on the system with little control and management. From the
results obtained, the researchers at the experimental site in France were able
to incorporate certain control and management strategies to obtain
favourable results, which is actually the aim for this PhD study as well.
After conducting multiple experiments, the authors of the pilot plant set up
in France, discovered that injection of carbon dioxide into the algal ponds
with slight agitation (mixing) helped improve the algal productivity. The
zooplankton ponds required gentle aeration at higher temperatures. Similar
strategies need to be implemented in the current IBS project which would
overcome the problems of low algal, zooplankton and fish productivity.

149
Figure 5.19 Diagram of the pilot-scale IBS for recycling swine manure in France
5.6 Conclusions
The mathematical model (DYRESM CAEDYM) developed for the
commercial scale IBS provides information on the chlorophyte, zooplankton
and fish concentrations when anaerobically digested piggery effluent was
fed through the aquaculture module of the IBS from the anaerobic digestion
system. The aim of this chapter is to investigate the suitability of DYRESM
CAEDYM as a modelling tool for a commercial IBS. The results obtained
from this modelling study would also provide information on the parameters
which need to be controlled and managed for the efficiency of the system.
The maximum chl a concentration predicted by the model is approximately
90 µg chl a L-1 and 60 µg chl a L-1 in algal ponds 1 and 2 respectively
(Figure 5.4 & Figure 5.8). The chlorophyte growth is for a short period
during the first three months of the simulation. Both the algal ponds are
dominated by cyanobacteria with the maximum cyanobacteria concentration
of 200 mg m-3. Freshwater diatom concentration is almost negligible. The
Algal Raceways
Fish Pond
Zooplankton
Pond
Tank with manure Recycled Water
Intermediate
Basin

150
results obtained from this study are in agreement with those found in
previous studies (Jørgensen, Jørgensen et al. 1981; Chen, Ji et al. 2002;
Hongping and Jianyi 2002; Havens, James et al. 2003; Freund, Mieruch et
al. 2006; Guven and Howard 2006; Håkanson, Bryhn et al. 2007; Fragoso
Jr, Marques et al. 2008), and confirm the dominance of cyanobacteria in
water bodies over chlorophyte growth.
Nutrient profile in the IBS ponds is shown in Figure 5.7, Figure 5.11, Figure
5.13, Figure 5.16 and Table 5.1. In algal pond 1, the concentrations of NH4
and PO4 were ~200 mgL-1 and ~90 mgL-1 respectively. In algal pond 2, the
concentrations of NH4 and PO4 were ~10 mgL-1 and ~25 mgL-1 respectively.
There was a slight dip in the NH4 concentration during the summer period.
In the zooplankton pond, concentrations of NH4 and PO4 were ~0.5 mgL-1
and ~8 mgL-1 respectively. NH4 concentration during the summer months is
almost negligible with a sudden spike in the autumn period. Concentration
of PO4 increased linearly at the end of the simulation period. In the fish
pond, concentrations of NH4 and PO4 were ~0.1 mgL-1 and ~3.25 mgL-1
respectively. The increase in concentrations of NH4 and PO4 is in agreement
with the reduction in bioconversion rates in phytoplankton, zooplankton and
fish. The crash in algal growth is directly related to the linear increase in the
nutrient concentrations.
The comparative IBS examples sourced from sites in India and France show
that IBS has been successfully implemented in different parts of the world
comprising tools for better control and management of ponds. The use of
mixing (agitation) and aeration assist in mixing the ponds and the effluent
uniformly which minimises stratification in ponds and thus reduces the
growth of cyanobacteria, and in turn improves the growth of phytoplankton,
zooplankton and fish. The model DYRESM CAEDYM could incorporate
the use of mixers and aeration in the IBS ponds to overcome the problems
of algal crashes in summer.
The parameters which can be controlled for managing the commercial scale
IBS in an effective way would be parameters related to inflow and outflow
volumes and flow rates of effluent, retention time of the effluent, nutrient

151
loads, rates of mixing and aeration within the ponds and control of biomass
conversion for primary, secondary and tertiary productions. These
management strategies could also be used to operate an IBS with a variety
of different effluents to its maximum capacity and construct an IBS with
better module design.
The outputs from the modelling studies presented above were studied in a
sensitivity analysis presented in Chapter 6 in order to identify those critical
parameters which cause a significant change in the phytoplankton output
when subject to a variation. An auto calibration program is developed in
Chapter 7 which will be used to validate the model with real time data by
adjusting the parameters identified in the sensitivity analysis in Chapter 6.

152
6 Sensitivity Analysis
6.1 Introduction
A sensitivity analysis is used to determine how “sensitive” a model is to
changes in values of the parameters of the model and to changes in the
structure of the model. Parameter sensitivity is usually performed as a series
of tests in which the modeller sets different parameter values to see how a
change in the parameter causes a change in the dynamic behaviour of the
output. By showing how the model behaviour responds to changes in
parameter values, sensitivity analysis is a useful tool in model building as
well as in model evaluation, since it helps the modeller to understand the
dynamics of a system. Experimenting with a wide range of values can offer
insight into the behaviour of a system in any situation. Discovering that the
system behaviour greatly changes for a change in a parameter value can
identify a leverage point in the model- i.e. a parameter whose specific value
can significantly influence the behaviour mode of the system.
A sensitivity analysis also helps to build confidence in the model by
studying the uncertainties that are often associated with parameters in
models. Many parameters in system dynamics models represent quantities
that are very difficult, or even impossible to measure to a great deal of
accuracy in the real world. Also, some parameter values change in the real
world. Therefore, when building a system dynamics model, the modeller is
usually at least somewhat uncertain about the parameter values he chooses
and must use estimates. Sensitivity analysis allows determination of what
level of accuracy is necessary for a parameter to make the model
sufficiently useful and valid. If the tests reveal that the model is insensitive,
then it may be possible to use an estimate rather than a value with greater
precision. A sensitivity analysis can also indicate which parameter values
are reasonable to use in the model. If the model behaves as expected from
real world observations, it gives some indication that the parameter values
reflect, at least in part, the “real world.”

153
In Chapter 5 the suitability of DYRESM CAEDYM as a mathematical
model for the proposed commercial IBS was tested but the model could not
be validated with real time data because the IBS was not built at the time
this research was conducted. The purpose of developing the model was to
provide a quantitative description of the interactions that occur between
physical and ecological processes. DYRESM CAEDYM is a complex
model which has numerous interactive variables. The hydrodynamic
component DYRESM doesn’t require any site specific validation data,
however the ecological component CAEDYM requires calibration with field
data from the system which is being modelled. To minimise effort, it is
important to determine those parameters which the model result is most
sensitive to, then the calibration process should be directed towards these
parameters. Sensitivity studies form the basis for calibrating the parameters
for model validation. As the number of parameters which need to be
calibrated is large, a rigorous calibration method involving manual input of
parameters is not desired as this is time consuming. An automatic iterative
calibration program has been developed in Chapter 7 which automatically
validates the model with field data by adjusting the identified critical
parameters. A sensitivity analysis is conducted in this chapter to show how
sensitive the model outputs are to the selected parameters, which will assist
both the calibration discussed in Chapter 7 and the design and operation of
the aquaculture system.

154
6.2 Methods
Multiple studies have been conducted in the past regarding sensitivity
analysis for water quality models (Jørgensen, Jørgensen et al. 1981;
Jørgensen, Kamp-Nielsen et al. 1986), including DYRESM CAEDYM
(Schladow and Hamilton 1997). The assigned parameter range selected for
this sensitivity analysis study were obtained from Schladow and Hamilton
(1997) (Schladow and Hamilton 1997). The assigned values for the
parameters are the same values used in developing the model for the
proposed commercial IBS discussed in Chapter 5. These parameters were
also selected for the Auto Calibration routine to be discussed in Chapter 7.
Sensitivity analysis was conducted on the algal component of the IBS. The
sensitivity of model results to changed values of the parameters was
quantified with reference to chlorophyll a (Chl-a) growth. Chl-a growth was
considered to be one of the most important outputs for this research as it is
well correlated with biomass production in the proposed large scale IBS.
For the sensitivity analysis, the model was run with each of the parameters
in Table 6.2 using the assigned value, the minimum (-10% of assigned
value) and the maximum (+10% of the assigned value) while the other
parameters were kept fixed at the assigned value. The model was run for a
period of 365 days, starting from 1st June 2006 to 31st May 2007.
The working formulae for the values chosen are shown below.
Minimum Value = Assigned Value – (10% of Assigned Value) Eq. 6.1
Maximum Value = Assigned Value + (10% of Assigned Value) Eq. 6.2
% Range = 100 – [(Minimum Value/Maximum Value)*100] Eq. 6.3
The assigned values were selected from Schladow and Hamilton (1997).
The minimum and maximum values were computed as 10% of the assigned
values to show a distribution range of these parameters and their effects on
the chl-a output.

155
Table 6.1 Parameters used in sensitivity analysis (Schladow and Hamilton 1997)
Parameter Definition UnitsAssigned
RangeAssigned
Value
P max
Maximum phytoplankton
growth rateday -1 1.3 – 3.6 1.52
kr
Phytoplankton respiration coefficient
day -1 0.05 – 0.17 0.079
vR
Phytoplankton temperature multiplier
1.02 – 1.14 1.08
IPmin
Minimum phytoplankton
internal Pmg P (mg Chl a)-1 0.1 – 1.0 0.3
IPmax
Maximum phytoplankton
internal Pmg P (mg Chl a)-1 1.0 – 5.0 2
UPmax
Maximum rate of
phytoplankton P uptake
mg P (mg Chl a)-1 0.05 – 1.0 0.3
INmin
Minimum phytoplankton
internal Nmg N (mg Chl a)-1 1.5 – 4.0 3
INmax
Maximum phytoplankton
internal Nmg N (mg Chl a)-1 8.0 – 15.0 9
UNmax
Maximum rate of
phytoplankton N uptake
mg N (mg Chl a)-1 0.5 – 10.0 1.5
KP
Half saturation constant for
phytoplankton P uptake
mg L-1 0.0125 – 0.025 0.0125
KN
Half saturation constant for
phytoplankton N uptake
mg L-1 0.01 – 0.2 0.014
Ik
Parameter for initial slope of P-
I curveµE m-2 s-1 100 – 500 254

156
Table 6.2 Parameters used in the sensitivity analysis study
ParameterAssigned
Value -ve 10% +ve 10%
Pmax 1.52 1.368 1.672kr 0.079 0.0711 0.0869vR 1.08 0.972 1.188IPmin 0.3 0.27 0.33IPmax 2 1.8 2.2UPmax 0.3 0.27 0.33INmin 3 2.7 3.3INmax 9 8.1 9.9UNmax 1.5 1.35 1.65KP 0.0125 0.01125 0.01375KN 0.014 0.0126 0.0154Ik 254 228.6 279.4

157
6.3 Results
6.3.1 Maximum Phytoplankton Growth Rate (Pmax)
Figure 6.1 Chlorophyll-a response to change in Pmax values
During the early part of the simulation, which occurs in winter, a 10%
increase in Pmax , resulted in an increase in Chl-a, peaking 10 days or 25%
earlier and at a level 10% higher than the assigned value, while a 10%
decrease in Pmax resulted in a decrease in Chl-a peaking 20 days or 28.5%
later and at a level 20% lower than the assigned value. Through the middle
of the simulation period, simulated values were quite small, however the
minimum chlorophyll-a value fell 5% below the assigned value for the
remaining days of the simulation. The initial spike in Chl-a output could
possibly be due to the low water temperature in winter and a high
availability of nutrients at the start of the simulation. The Chl-a output was
almost negligible during the summer months because of the spike in growth
in cyanobacteria which suppresses the growth of chlorophytes as shown in
Chapter 5.
0
20
40
60
80
100
120
0 100 200 300 400
Chloroph
yll-‐a (u
g/L)
Simulation Day No.
Chlorophyte
Assigned Value
minus 10%
plus 10%

158
6.3.2 Phytoplankton Respiration Coefficient (kr)
Figure 6.2 Chlorophyll-a response to change in kr values
The effects observed in this case were similar to the previous one, but in the
opposite direction. An increase of 10% in the kr value resulted in a 5%
decrease in the Chl –a peak value and the peak occured 5 days or 5% after
the Chl-a peak for the assigned value. A decrease of 10% in the kr value
resulted in the Chl-a peaking at 18% higher and at 5 days or 5% earlier than
for the Chl-a peaks at the assigned value. During the middle of the
simulation period, which occurs in the summer months, simulation
differences were almost negligible. Towards the final simulation period, the
maximum value of kr yielded a Chl-a output which was 5 % lower than the
assigned value, while the minimum value of kr did not show any significant
differences than the assigned values.
0
20
40
60
80
100
120
0 100 200 300 400
Chloroph
yll-‐a (u
g/L)
Simulation Day No.
Chlorophyte
Assigned Value
minus 10%
plus 10%

159
6.3.3 Phytoplankton Temperature Multiplier (vR)
Figure 6.3 Chlorophyll-a response to change in vR values
An increase in 10% in the vR value resulted in a 50% increase in the Chl-a
peak with the peak occurring 20 days earlier or 40% before the Chl-a peak
at the assigned value. A 10% decrease in the vR value resulted in a 70%
reduction in the Chl-a peak and the peak occurred 210% later, in mid
summer compared to the peak at the assigned value. The minimum value of
vR resulted in extremely low value in Chl-a (96% lower than the peak at the
assigned value) which continued for half of the simulation period. Towards
the second half of the simulation period, the maximum vR value resulted in
almost negligible values in Chl-a till the end of the simulation, while the
minimum vR value yielded a higher Chl-a output than the assigned value
until the last few weeks. The final simulation values for both maximum and
minimum values of vR were much lesser than the assigned values which
will cause difficulty in running the IBS ponds for a consecutive second year.
0
20
40
60
80
100
120
140
0 100 200 300 400
Chloroph
yll-‐a (u
g/L)
Simulation Day No.
Chlorophyte
Assigned Value
minus 10%
plus 10%

160
6.3.4 Minimum Phytoplankton Internal P (IPmin)
Figure 6.4 Chlorophyll-a response to changes in IPmin values
A 10% increase and decrease in the IPmin value does not result in any
significant change in the Chl- a output. The Chl-a peaks occur almost at the
same value and time as that of the assigned value. There is a dip in the Chl-a
output (50% drop from the instantaneous value before the dip) for the
minimum value around the 110th day into the simulation, which is recovered
almost completely after a couple of days.
0
20
40
60
80
100
120
0 100 200 300 400
Chloroph
yll-‐a (u
g/L)
Simulation Day No.
Chlorophyte
Assigned Value
minus 10%
plus 10%

161
6.3.5 Maximum Phytoplankton Internal P (IPmax)
Figure 6.5 Chlorophyll-a response to changes in IPmax values
Again there is very little difference for the prediction of different values of
IPmax, when compared to that of IPmin. A 10% increase and decrease in the
IPmax value does not result in any significant change in the Chl- a output.
The Chl-a peaks occur at the same value and time as that of the assigned
value. A dip in the minimum value of the parameter, similar to the one for
IPmin , is observed in this case as well. The dip is for a short period ~2 days.
0
20
40
60
80
100
120
0 100 200 300 400
Chloroph
yll-‐a (u
g/L)
Simulation Day No.
Chlorophyte
Assigned Value
minus 10%
plus 10%

162
6.3.6 Maximum Rate of Phytoplankton P Uptake (UPmax)
Figure 6.6 Chlorophyll-a response to changes in UPmax values
A 10% increase and decrease in the IPmax value does not result in any
significant change in the Chl- a output. The Chl-a peaks occur at the same
value and time as that of the assigned value.
0
20
40
60
80
100
120
0 100 200 300 400
Chloroph
yll-‐a (u
g/L)
Simulation Day No.
Chlorophyte
Assigned Value
minus 10%
plus 10%

163
6.3.7 Minimum Phytoplankton Internal N (INmin)
Figure 6.7 Chlorophyll-a response to changes in INmin values
A 10% increase in INmin value resulted in a 10% decrease in the peak value
of Chl-a where the peaking occurred 15% later than the peaking at the
assigned value, while a 10% decrease in INmin value resulted in a similar
magnitude peak to that of the peak at the assigned value, however the peak
occurred 10 days earlier and lasted for longer. During the summer months,
there was negligible Chl-a output. For the remainder of the simulation
period, the maximum and minimum values of INmin are 5% lower and
higher (respectively) than the Chl-a output for the assigned value, and this
difference disappears towards the end of the simulation period.
0
20
40
60
80
100
120
0 100 200 300 400
Chloroph
yll-‐a (u
g/L)
Simulation Day No.
Chlorophyte
Assigned Value
minus 10%
plus 10%

164
6.3.8 Maximum Phytoplankton Internal N (INmax)
Figure 6.8 Chlorophyll-a response to changes in INmax values
A 10% increase and decrease in the INmax value does not result in any
significant change in the Chl- a output. The Chl-a peaks occur at the same
value and time as that of the assigned value.
0
20
40
60
80
100
120
0 100 200 300 400
Chloroph
yll-‐a (u
g/L)
Simulation Day No.
Chlorophyte
Assigned Value
minus 10%
plus 10%

165
6.3.9 Maximum Rate of Phytoplankton N Uptake (UNmax)
Figure 6.9 Chlorophyll-a response to changes in UNmax values
A 10% increase in UNmax yields a similar Chl-a peak to the one obtained for
the assigned value, however the peak happens approximately 5 days earlier
and persists for a longer time. A decrease in 10% in UNmax value has a Chl-a
peak which is 10% lower than the Chl-a peak for the assigned value
occurring 10 days later. There is some difference in the startup phase in
autumn for each of the three values of the parameter before a final
convergence in the Chl-a output towards the end of the simulation.
0
20
40
60
80
100
120
0 100 200 300 400
Chloroph
yll-‐a (u
g/L)
Simulation Day No.
Chlorophyte
Assigned Value
minus 10%
plus 10%

166
6.3.10 Half Saturation Constant for Phytoplankton P Uptake (KP)
Figure 6.10 Chlorophyll-a response to changes in KP values
A 10% increase and decrease in the KP value does not result in any
significant change in the Chl- a output, except for the abrupt dip at 110th
day. The Chl-a peaks occur at the same value and time as that of the
assigned value.
0
20
40
60
80
100
120
0 100 200 300 400
Chloroph
yll-‐a (u
g/L)
Simulation Day No.
Chlorophyte
Assigned Value
minus 10%
plus 10%

167
6.3.11 Half Saturation Constant for Phytoplankton N Uptake (KN)
Figure 6.11 Chlorophyll-a response to changes in KN values
A 10% increase and decrease in the KN value does not result in any
significant change in the Chl- a output. The Chl-a peaks occur at the same
value and time as that of the assigned value. Here there is a similar dip to
that of KP in the Chl-a output, but in this case the dip is observed for the
maximum value.
0
20
40
60
80
100
120
0 100 200 300 400
Chloroph
yll-‐a (u
g/L)
Simulation Day No.
Chlorophyte
Assigned Value
minus 10%
plus 10%
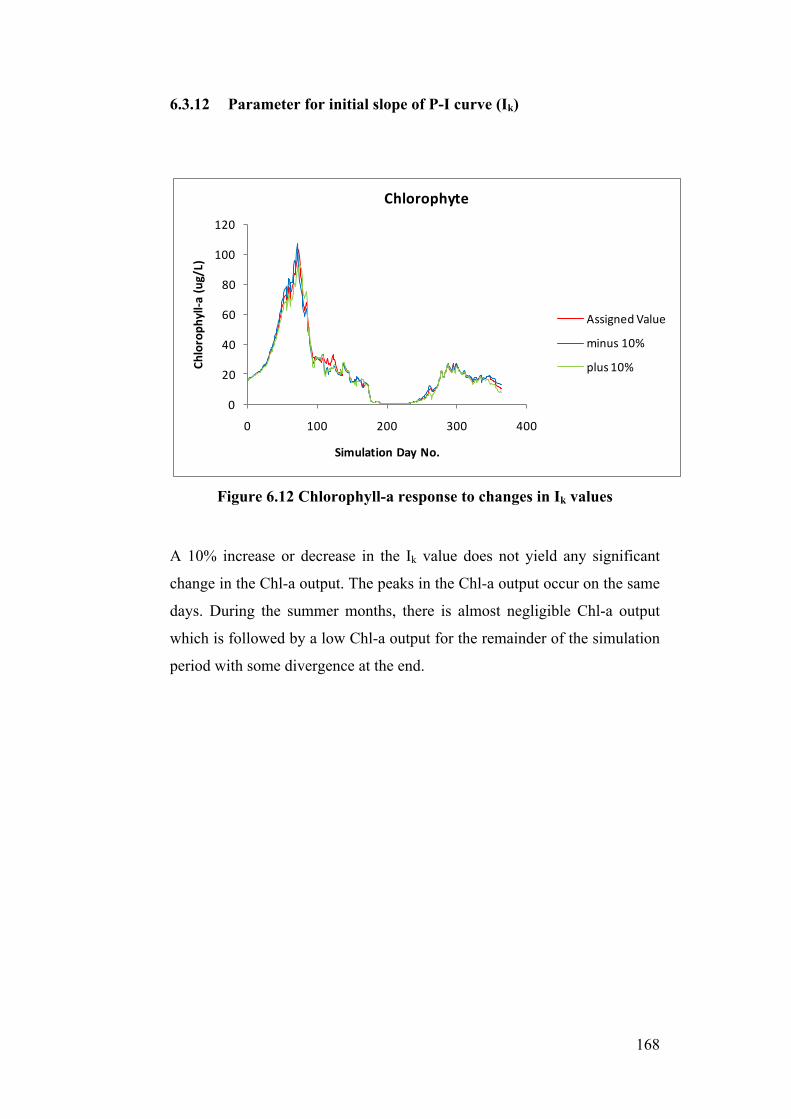
168
6.3.12 Parameter for initial slope of P-I curve (Ik)
Figure 6.12 Chlorophyll-a response to changes in Ik values
A 10% increase or decrease in the Ik value does not yield any significant
change in the Chl-a output. The peaks in the Chl-a output occur on the same
days. During the summer months, there is almost negligible Chl-a output
which is followed by a low Chl-a output for the remainder of the simulation
period with some divergence at the end.
0
20
40
60
80
100
120
0 100 200 300 400
Chloroph
yll-‐a (u
g/L)
Simulation Day No.
Chlorophyte
Assigned Value
minus 10%
plus 10%

169
6.4 Discussion
Sensitivity analysis is the study of how the variation (uncertainty) in the
output of a mathematical model can be qualitatively or quantitatively
measured, to different sources of variation in the input of the model. It is a
technique for systematically changing parameters in a model to determine
the effects of such changes. In more general terms uncertainty and
sensitivity analysis investigate the robustness of a study when the study
includes some form of mathematical modelling. Sensitivity analysis can be
useful to mathematical modellers for a range of purposes including:
• decision making or the development of recommendations for
decision makers (e.g. testing the robustness of a result);
• enhancing communication from modellers to decision makers (e.g.
by making recommendations more credible, understandable,
compelling or persuasive);
• increased understanding or quantification of the system (e.g.
understanding relationships between input and output variables); and
• model development (e.g. searching for errors in the model).
The outputs (Figure 6.1 to Figure 6.12) show that the model results for
chlorophyll-a are sensitive to a relatively small subset of parameters.
Parameters for which chlorophyll-a is highly sensitive are those that directly
alter growth rates i.e. maximum phytoplankton growth rate (Pmax),
phytoplankton respiration coefficient (kr) and phytoplankton temperature
multiplier (vR), or indirectly affect growth rates through their ability to
utilise nitrogen (INmin and UNmax). These results are in agreement with the
results presented in (Hamilton and Schladow 1997) where chl-a was highly
sensitive for maximum phytoplankton growth rate and phytoplankton
respiration rate. However in Hamilton and Schladow (Hamilton and
Schladow 1997) chl-a is sensitive to IPmin and UPmax i.e. parameters that
indirectly affect growth rates through their ability to utilise phosphorus as
opposed to INmin and UNmax in the IBS ponds. This could be due to the IBS
system being a nitrogen limited system.

170
The percentage change in parameter input compared to the percentage
change in Chl-a output is shown in Table 6.3.
Table 6.3 Comparison of percentage variation in input parameter and output Chl-a
Parameters which exhibit significant percentage deviations in output with
respect to change in input are Pmax, kr, vR, INmin and UNmax. All these
parameters have an assymetric effect i.e. moving one way has significantly
ParameterAssigned
Value % Range% change in input
% change in output
-10 -20
10 10
-10 18
10 -5
-10 -70
10 50
-10 ~0
10 ~0
-10 ~0
10 ~0
-10 ~0
10 ~0
-10 ~0
10 -10
-10 ~0
10 ~0
-10 -10
10 ~0
-10 ~0
10 ~0
-10 ~0
10 ~0
-10 ~0
10 ~0Ik 254 18.2
KP 0.0125 18.2
KN 0.014 18.2
INmax 9 18.2
UNmax 1.5 18.2
UPmax 0.3 18.2
INmin 3 18.2
IPmin 0.3 18.2
IPmax 2 18.2
kr 0.079 18.2
vR 1.08 18.2
Pmax 1.52 18.2

171
less effect than moving the other way. This shows that there is an optimum
somewhere between the two points.
6.5 Conclusions and Further Work
This study provided an overview of a selected group of parameters which
are most sensitive to chlorophyll-a output. This technique can be applied to
the commercial IBS, when constructed, in future. The sensitivity of different
outputs can be tested against a combination of different parameters for the
IBS which can assist the modeller in deciding which parameters need to be
altered to expect a different output, and which parameters can be altered
without expecting any change in the output. This mechanism would provide
effective control and management of the IBS from a modeller’s perspective.
Further investigation needs to be done on the response of different outputs
e.g. cyanobacteria, freshwater diatoms, zooplankton, fish growth etc with
changes in the parameters selected for this study. This is useful in
identifying the parameters which are most sensitive to a particular output.
However when the number of parameters increases along with a variety of
outputs to be assessed, a manual method of altering parameter values to
observe the change in output is time consuming and more of a “trial and
error” approach. Also this sensitivity study relies on using the minimum,
maximum and mean values of the parameters selected without observing
any changes in the intermediate values, i.e. observing any changes in the
output during incremental changes to the selected parameter. This
incremental change is required when field data needs to be validated against
the simulated data. It is obvious that manually inputting the values with
incremental changes would be an extremely time consuming and frustrating
process. Therefore a need for development of an efficient automatic
calibration process is required, which has been developed in Chapter 7.

172
7 Automated Parameter Estimation and
Calibration
7.1 Introduction
Automatic calibration is a mathematical technique employed by scientists,
engineers and mathematical modellers to calibrate and validate a
mathematical model to obtain realistic data. The fundamental way is to alter
the variable parameters of a model and compare the outputs with the
obtained field (actual) data. The parameter value which gives the closest fit
(agreement) of the field data to the simulated data is chosen to represent that
value. In simple models, which require minimum number of parameters,
calibration and validation can be done manually as long as it doesn’t
consume much time. However in complex models, like DYRESM
CAEDYM, where a large number of parameters, either singularly or in
conjunction, have a profound effect on the outputs, manual calibration is
lengthy, tedious and an inefficient process. Often, this would lead to
erroneous and inconsistent results, which would misguide the modellers and
operational engineers. Therefore a method of automated calibration which
includes model validation and parameter estimation by using automated
computer programs is very useful in these circumstances. This provides the
advantage of running a more sophisticated process, reduces the chances of
errors and false estimations and is less tedious and taxing as opposed to the
conventional manual methods.
In Chapter 6 a sensitivity analysis study was conducted on selected
parameters for the proposed large scale IBS model. The sensitivity analysis
yielded valuable information about the response of chlorophyll-a output to
fixed positive and negative variations to those parameters. This paved a way
for calibrating the model with real time data. However there is a need for
effectively calibrating a model like DYRESM CAEDYM which has been
discussed in Section 6.5. This chapter introduces a method to calibrate the
proposed large scale IBS model using an automatic calibration program

173
which fits the simulated model output to the field data by varying the
parameters selected.
7.2 Methods
7.2.1 Incorporating Monte Carlo and GLUE calibration in
DYRESM CAEDYM
A calibration routine involving Bayesian Monte Carlo method and GLUE
was set up to enable automated parameter calibration in DYRESM
CAEDYM. Both the Bayesian Monte Carlo and GLUE method have been
discussed in Chapter 2. The GLUE procedure explicitly recognizes the
equivalence or near equivalence of different parameter sets or model
structures in the representation of hydrological responses (Beven and Binley
1992). The parameters selected for the calibration were
1) Maximum Potential Growth Rate of Phytoplankton (Pmax)
2) Parameter for initial slope of P_I curve (IK)
3) Half Saturation Constant for Nitrogen (KN)
4) Half Saturation Constant for Phosphorus (KP)
5) Internal Minimum Phosphorus (IPmin)
6) Internal Maximum Phosphorus (IPmax)
7) Maximum Rate of Phytoplankton Phosphorus Uptake (UPmax)
8) Internal Minimum Nitrogen (INmin)
9) Internal Maximum Nitrogen (INmax)
10) Maximum Rate of Phytoplankton Nitrogen Uptake (UNmax)
11) Respiration Rate (kr)
These parameters were determined using initial simulation studies to limit
the bounds of the parameter space to reduce over parameterization. Ranges

174
for the above parameters were obtained from literature (Hamilton and
Schladow 1997; Schladow and Hamilton 1997). Due to the unavailability of
real time field data, pseudo field data was generated with random values for
demonstration purpose for this calibration routine to function properly.
7.2.2 Analysis of the auto calibration program
The calibration program comprises of a main program which has several
subroutines embedded within it. These subroutines perform individual tasks
for the calibration routine to function properly. The sequence of the program
execution is explained below:
1. The program first aligns the numerical values in the water quality
parameters file to match that of an 80 character set. This is done so
that specific parameter values can be inserted at the appropriate
fields in the water quality parameters file, without causing the
DYRESM CAEDYM program to crash. It has been observed that
insertion of parameter values at different fields in the parameters file
causes the model to crash. Subroutine ReformatDYCD.f90 transfers
the parameters from “WQParameters.dat” file to “dycd.dat” file
according to the 80 character alignment. The new “dycd.dat” file is
used for running the model simulations. This alignment needs to be
done only once before the start of the initial simulations.
2. The main program Driver.f90 consists of three subroutines
• datamod.f90
• master_run.f90
• glue_sub.f90
The main program Driver.f90 is executed in the following sequence:
First the number of simulations to be conducted is entered. For
demonstration purposes the number of simulations is set at 1000. Large
number of simulations requires longer computing times. A randomised
value (within the estimated limits) is selected for each of the 11
parameters selected for calibration. These values are stored in the new
“dycd.dat” file which will be used for CAEDYM simulations. A nested

175
loop performs this function using the datamod.f90 and the num_ascii.f90
subroutines. This subroutine places the parameter at the exact position in
the “dycd.dat” file. It is extremely important to place these values at the
exact position else CAEDYM will crash during the simulations.
The next part of the code is the subroutine master_run.f90. This
subroutine calls for the program DYRESM CAEDYM to run.
The final part of the code is the GLUE (Monte Carlo) calibration,
executed by the glue_sub subroutine. Once DYRESM CAEDYM
simulations are completed, GLUE calibration is executed. Chlorophyte
and cyanobacteria concentration outputs are chosen for demonstration
purposes. CAEDYM stores the simulated chlorophyte and cyanobacteria
data in “CHLOR.ITS” and “CYANO.ITS” files as Integrated Time
Series (ITS) files. A pseudo field data is randomly created for
demonstration of the program. Both the pseudo field and the simulated
data files are opened and GLUE calibration is performed where the
Nash-Sutcliffe coefficient (NS) is calculated. The numerical values of
the parameter along with the NS value for that particular parameter
combination is written to a text file for later comparison.
The above sequence is repeated for the total number of simulations (i.e.
1000 in this case). For each simulation, the NS values are recorded with
the parameter value combination. At the end of the simulation period,
the parameter combination which gives the NS value closest to 1.0 will
give the best fit with the field data. A flowchart describing the above
sequence is shown in Figure 7.1.
The program code written in FORTRAN 90 is shown in Appendix E. A
GLUE calibration output of 10 simulations is shown in Appendix F.

176
7.2.3 Flowchart of the GLUE program
Figure 7.1 FlowChart of the Monte Carlo & GLUE Calibration script
NO
YES
Is Count = N
Sort NS results to choose best parameter
combination, θi
END
Generate model simulations for every θ
Run GLUE calibration script
Calculate Nash Sutcliffe (NS)
coefficients
Count = Count + 1
Generate random values for the selected
parameters, θ, within the specified
feasible range
Count = 0
START
INPUT: DYRESM CAEDYM Program,
Selected Model Parameters, θ
Total No. of Iterations, N
Field Data

177
7.3 Results
7.3.1 GLUE Calibration for Chlorophyte growth
Nash-Sutcliffe (NS) coefficients computed for each of the 1000 iterations
for chlorophyte calibration is shown in Figure 7.2. Each data point
represents an NS value. The scatter data points range from -3.5 to
approximately 1. The majority of the data points are in between 1 and -1.
Figure 7.2 Nash Sutcliffe Coefficients for GLUE Calibration
-‐4
-‐3
-‐2
-‐1
0
1
0 200 400 600 800 1000 1200
Nash -‐Sutcliffe
coe
fficie
ntNS
No. of Iterations
Nash Sutcliffe

178
Table 7.1 shows 10 random simulations chosen from the 1000 simulations
performed to demonstrate the GLUE calibration. The highest value of NS
was 0.903704. This means that if the model is run with the parameter values
used in simulation run no. 10 it will give 90.37% agreement with the field
data. This parameter combination will be the validated data set.
Table 7.1 GLUE Calibration results for 10 random simulations
Figure 7.3 shows the model (simulated) and field data before GLUE
calibration.
Figure 7.4 shows the model (simulated) and field data after GLUE
calibration. There is a close agreement between the model (simulated) and
field data once the GLUE calibration script has been applied. The parameter
set for simulation no. 10 as discussed above gave the best NS coefficient,
hence it has been used as the validation data in the GLUE calibration.
Runs Pmax IK KP KN INmin INmax UNmax IPmin IPmax UPmax kr NS
1 1.6916 168.76 0.0115 0.0108 2.9488 10.406 1.0426 0.3649 2.7350 0.3453 0.0893 0.5255
2 1.5978 206.13 0.0124 0.0130 2.5194 6.702 1.0081 0.3282 1.1005 0.4977 0.085 0.8715
3 0.5846 177.91 0.0110 0.0157 2.9517 7.270 1.2204 0.4289 1.2571 0.3336 0.095 - 1.079
4 1.3099 248.48 0.0120 0.0136 2.0501 11.923 1.7405 0.3344 1.8494 0.2858 0.094 - 0.157
5 1.4327 209.31 0.0179 0.0169 2.5443 10.924 1.0936 0.3828 2.4322 0.3798 0.0969 0.4035
6 1.1459 200.65 0.0136 0.0102 2.1284 12.879 1.9878 0.2067 1.4837 0.3807 0.0937 0.0389
7 1.2831 206.47 0.0110 0.0153 2.4674 9.7179 0.1247 0.2601 1.5325 0.4538 0.0845 0.8236
8 1.0641 276.47 0.0176 0.0161 2.3192 9.346 1.0351 0.4792 1.0798 0.2792 0.0842 - 0.376
9 1.3235 294.22 0.0108 0.0156 2.9931 14.014 1.4117 0.4079 1.7869 0.3526 0.0913 0.6438
10 1.5238 239.20 0.0110 0.0155 2.2686 7.6804 1.0117 0.2583 2.8496 0.2192 0.0881 0.9037

179
Figure 7.3 Comparison between simulated (model) and field data before
GLUE calibration for chlorophyte growth
Figure 7.4 Comparison between simulated (model) and field data after GLUE calibration for chlorophyte growth
0
10
20
30
40
50
60
70
0 50 100 150 200 250 300 350 400
Chloroph
yll
(ugL
-‐1)
No. of Days
Before Calibration
Simulated Data Field Data
0
10
20
30
40
50
60
70
0 50 100 150 200 250 300 350 400
Chloroph
yll
(ugL
-‐1)
No. of Days
After calibration
Simulated Data Field Data

180
Table 7.2 shows the lower and upper bounds, the initial values used for the
model simulation and the calibrated values of the parameters which give an
NS value of 0.903704.
Table 7.2 Numerical Values of the Calibrated Parameters
Calibrated Parameter
Lower Bound Upper Bound
Initial Value
Calibrated Value
Pmax 0.3 2.5 1.52 1.5238Ik 150 300 254 239.2KP 0.01 0.02 0.0125 0.011KN 0.01 0.02 0.014 0.0155INmin 2 3 3 2.2686INmax 5 15 9 7.6804UNmax 1 2 1.5 1.0117IPmin 0.2 0.5 0.3 0.2583IPmax 2 3 2 2.8496UPmax 0.2 0.5 0.3 0.2192kr 0.08 0.1 0.079 0.0881
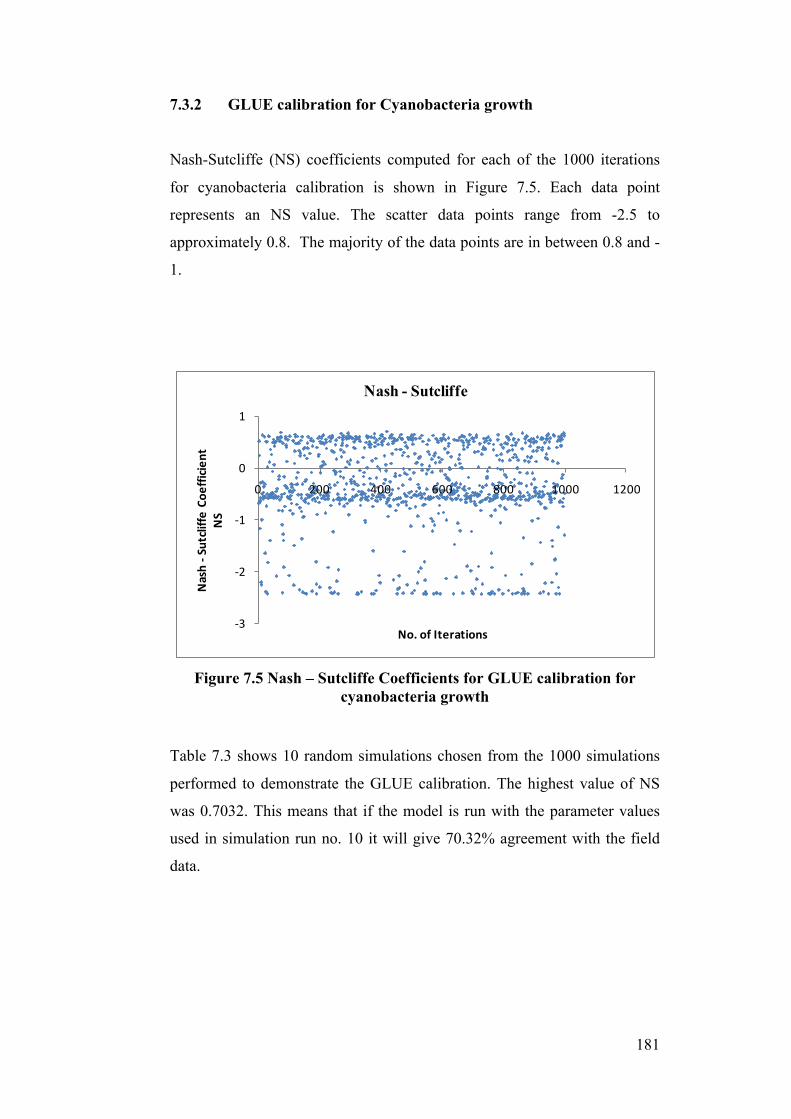
181
7.3.2 GLUE calibration for Cyanobacteria growth
Nash-Sutcliffe (NS) coefficients computed for each of the 1000 iterations
for cyanobacteria calibration is shown in Figure 7.5. Each data point
represents an NS value. The scatter data points range from -2.5 to
approximately 0.8. The majority of the data points are in between 0.8 and -
1.
Figure 7.5 Nash – Sutcliffe Coefficients for GLUE calibration for
cyanobacteria growth
Table 7.3 shows 10 random simulations chosen from the 1000 simulations
performed to demonstrate the GLUE calibration. The highest value of NS
was 0.7032. This means that if the model is run with the parameter values
used in simulation run no. 10 it will give 70.32% agreement with the field
data.
-‐3
-‐2
-‐1
0
1
0 200 400 600 800 1000 1200
Nash -‐Sutcliffe
Coe
fficie
ntNS
No. of Iterations
Nash - Sutcliffe

182
Table 7.3 GLUE Calibration Results for 10 random simulations
Figure 7.6 shows the model (simulated) and field data before GLUE
calibration.
Figure 7.7 shows the model (simulated) and field data after GLUE
calibration. There is a close agreement between the model (simulated) and
field data once the GLUE calibration script has been applied. The parameter
set for simulation no. 10 as discussed above gave the best NS coefficient,
hence it has been used as the validation data in the GLUE calibration.
Run
s Pma
x I
K K
P K
N INmi
n INma
x UNma
x IPmi
n IPma
x UPma
x k
r N
S 1 0.670
2 66.5
7 0.006
0 0.002
9 2.511
7 4.753
2 0.996
3 0.122
6 0.879
9 0.079
7 0.068
5 0.520
9 2 0.835
3 31.8
0 0.004
9 0.003
9 2.952
4 4.825
0 0.837
7 0.129
4 0.643
2 0.050
0 0.063
1 0.635
9 3 0.872
9 78.8
1 0.006
5 0.003
7 2.175
9 3.639
5 0.675
0 0.067
4 0.496
9 0.116
5 0.067
9 - 0.603
6
4 0.438
6 57.1
6 0.003
9 0.000
9 2.546
4 3.514
9 0.826
1 0.111
0 0.791
4 0.069
6 0.068
7 - 0.312
3 5 0.8826
75.8
7 0.006
7 0.001
0 2.179
0 3.364
5 0.968
0 0.089
4 0.739
9 0.064
2 0.075
9 0.510
1 6 0.554
3 72.3
4 0.005
7 0.004
7 2.418
3 4.192
5 0.940
2 0.140
4 0.369
8 0.083
1 0.067
2 0.506
8 7 0.533
5 66.7
4 0.004
5 0.001
6 2.675
7 3.336
6 0.838
1 0.104
8 0.836
2 0.140
7 0.074
5 0.012
8 8 0.809
2 45.7
2 0.006
1 0.000
7 2.736
2 3.474
3 0.287
9 0.071
1 0.741
9 0.071
5 0.071
1 - 1.831
9 9 0.708
1 44.3
6 0.004
9 0.003
2 2.093
1 4.645
3 0.606
9 0.128
3 0.863
9 0.102
4 0.067
8 0.614
5 1
0 0.750
0 62.5
0 0.005
2 0.001
0 2.01
5 4.002
0 0.751
1 0.122
4 0.600
0 0.118
3 0.074
0 0.703
2
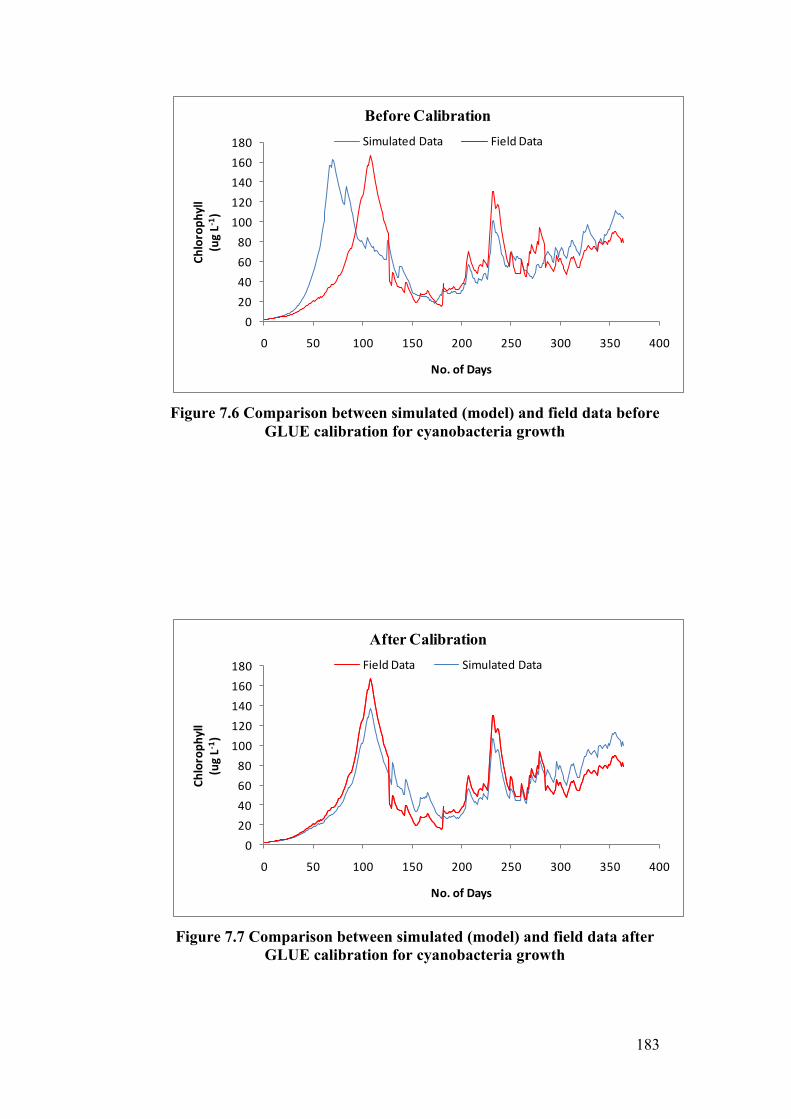
183
Figure 7.6 Comparison between simulated (model) and field data before
GLUE calibration for cyanobacteria growth
Figure 7.7 Comparison between simulated (model) and field data after
GLUE calibration for cyanobacteria growth
020406080100120140160180
0 50 100 150 200 250 300 350 400
Chloroph
yll
(ug L-‐1 )
No. of Days
Before CalibrationSimulated Data Field Data
020406080100120140160180
0 50 100 150 200 250 300 350 400
Chloroph
yll
(ug L-‐1 )
No. of Days
After CalibrationField Data Simulated Data

184
Table 7.4 shows the lower and upper bounds, the initial values used for the
model simulation and the calibrated values of the parameters which give an
NS value of 0.7032. The simulated data obtained by this calibrated value
parameter set gave the best fit to the field data for cyanobacteria output.
Table 7.4 Numerical Values of the Calibrated Parameters
7.4 Discussion and Conclusions
In recent years there has been an increase in the use and application of
distributed, physically-based, water quality and integrated hydrological
models. There have been multiple issues raised regarding how to properly
calibrate and validate these models and also assess the uncertainty of the
estimated parameters and the spatially-distributed responses. These issues
remain quite unexplored. For complex models, rigorous parameterisation,
reduction of the parameter space and the use of effective and efficient
algorithms are essential to facilitate the process of calibration and make it
more robust.
In this study, the GLUE methodology based on random Monte Carlo
simulations was applied to the DYRESM CAEDYM model for
demonstration purposes. 1000 random Monte Carlo simulations were
Calibrated Parameter
Lower Bound Upper Bound
Initial Value
Calibrated Value
Pmax 0.25 1 0.52 0.75Ik 20 100 54 62.5KP 0.002 0.007 0.003 0.0052KN 0 0.005 0.014 0.001INmin 2 3 2.5 2.015INmax 3 5 4 4.002UNmax 0.25 1 0.5 0.7511IPmin 0.05 0.15 0.1 0.1224IPmax 0.3 0.9 0.6 0.6UPmax 0.05 0.15 0.1 0.1183kr 0.06 0.08 0.07 0.074

185
conducted using parameters selected from the sensitivity analysis study in
Chapter 6. Large number of simulations would have required increased
computational times which would have not have been feasible and within
the time frame of this research work. After each simulation, the Nash-
Sutcliffe (NS) coefficient was computed. Multiple adjustments were made
to the data files in CAEDYM to be able to read the data for the
corresponding parameter. Program codes were written in FORTRAN 90 to
execute the above procedure. Pseudo field data was generated to run the
GLUE calibration module.
Chlorophyte and cyanobacteria growth output was used to demonstrate
GLUE calibration. Out of the 1000 iterations, the best NS coefficient
obtained was 0.9037 and 0.7032 for chlorophyte and cyanobacteria
respectively. Figure 7.4 and Figure 7.7 show a close agreement between the
simulated and pseudo field data after executing the GLUE calibration
program, which proves that the program can be used for model validation
purposes in future. This is definitely an improvement in the field of model
validation for DYRESM CAEDYM, as opposed to the previous “trial and
error” method which was time consuming and inaccurate. The complexity
of parameter interactions and their influence on the output in DYRESM
CAEDYM is enormous (as a combination of parameters has a profound
effect on a single output), and obtaining a “near perfect” parameter
combination using manual adjustments is extremely difficult. Thus the
calibration program as demonstrated in this chapter assists in overcoming
these challenges.
However, as much as the calibration process is automated, the user’s insight
and knowledge cannot be replaced. For example, while using the GLUE
method to calibrate DYRESM CAEDYM, the user decides which
parameters to calibrate by understanding which have the most influence on
model predictions for a specific case, through preliminary simulations and
acquired experience. Understanding the driving forces that work in each
simulation is critical to avoid the problem of over parameterisation and
unnecessary lengthy computational times.

186
7.5 Further Work
There is reasonable scope for further work to be conducted in this field.
• Simulations should be run with a larger number of iterations
(~100000) to obtain a better fit with the field data. Larger number of
simulations would require increased computing times, but would
provide a better accuracy in calibrating parameters. The chances of
obtaining a parameter set closest to the field data would be increased
significantly if the number of iterations are increased.
• Other outputs from the model e.g., nutrient data should also be
calibrated using the above procedure for effective model validation.
The main program is capable of including more parameters for
calibration. Additional parameters would require increased
computing times but would provide more accuracy in model outputs.

187
8 Summary and Conclusions
8.1 General Discussions and Conclusions
The aim of this PhD research was to develop a mathematical model for a
commercial scale IBS based on the suitability of DYRESM CAEDYM as a
modelling package and to use the results obtained from this modelling
exercise for deciding the required control and management of the IBS.
The objectives of this research were to
1. Obtain chemical parameters for piggery effluent from a two stage
anaerobic digestion system to serve as an input to the aquaculture
model.
2. Use numerical modelling techniques to test the suitability of
DYRESM CAEDYM as a modelling tool to model a commercial
scale IBS with a depth of 1 m.
3. Conduct a sensitivity analysis on the model parameters.
4. Use computer programming techniques to enable an automated
parameter estimation and calibration of the mathematical model.
8.2 Summary of the Research Results
This thesis has demonstrated the suitability of developing a mathematical
model for the IBS utilising piggery effluent as the nutrient source and has
provided results in the following key areas:
8.2.1 Laboratory Experiments and Development of Anaerobic
Digestion Model
The set up, installation and commissioning of the pilot scale two stage
anaerobic digestion system was conducted for a period of approximately 12
months. The main objective of this two-stage anaerobic digestion system
(comprising of a thermophilic system followed by a mesophilic digestion
system) was to conduct experiments to maximise throughput of raw piggery
effluent, whilst maintaining a high biogas generation. The experiments were

188
conducted in stages, with each stage having different hydraulic retention
times (HRT). The thermophilic (acidogenic) stage had HRT of 6.7 days,
while the mesophilic (methanogenic) stage had HRT of 15 days. Data for
pH, total solids, volatile solids, soluble COD, TKN, TAN, soluble P, VFA,
biogas volume and % CH4 in biogas was collected regularly. The set up,
installation and experiments conducted with the pilot plant system provided
an opportunity to understand the kinetics of raw piggery effluent digestion,
which paved a way for the development of an anaerobic digestion model.
A pilot scale integrated aquaculture (mesocosm) facility was set up in an
indoor climate controlled environment utilising anaerobically digested
piggery effluent for the culture of microalgae, zooplankton and fish.
Experiments were conducted relating to bioconversion of microalgae using
digested piggery effluent. The working volume of the microalgae (Chlorella
spp.) culture was kept at 280L and 180L, and the nutrient concentration was
fixed at 30 mg/L/day. Cell density, pH, TAN and phosphorus measurements
were taken daily. These experiments provided a better understanding of the
nutrient interactions and biomass conversions between digested piggery
effluent and microalgae which proved valuable during the simulations of the
aquaculture model DYRESM CAEDYM.
Batch scale anaerobic digestion experiments were conducted in the
laboratory for the development of a mathematical model. These experiments
were conducted in a batch mode to understand the degradation of raw
piggery effluent over a period of time and to obtain nutrient parameters of
TAN, soluble P, COD and pH which were necessary inputs to DYRESM
CAEDYM.
Analysis of the raw data from the batch scale experiments showed that TAN
and P were not correlated to methane production as initially suspected. A
new approach to modelling the anaerobic digestion system was utilised
which involved separate microbial kinetics for TAN, P and methane
production. Data outputs proved that both TAN/ Soluble P and COD/ CH4
followed two different sets of processes which were not correlated.

189
Microbial equations were developed which were used to fit the batch data
using “minimisation of errors squared” technique. The methane model
developed was remarkably different to other published models. It was
observed that methanogenesis was not the rate limiting step. Methane
production was found to be proportional to COD reduction, while P and
TAN were observed not to be proportional to COD.
8.2.2 Modelling the Aquaculture Component of the IBS
DYRESM CAEDYM was chosen as the appropriate modelling tool to
model the aquaculture stages of the IBS, consisting of bioconversion stages
of algae, zooplankton and fish. The ponds in the IBS were designed for a
depth of 1.0 m.
Initial simulations did not prove to be successful as the source code for
DYRESM CAEDYM had been originally written to simulate deep water
bodies ( > 5 m). The source code for this software was obtained from Centre
for Water Research (CWR), University of Western Australia (UWA), Perth
and compiled using Intel Visual Fortran Compiler 10 on Microsoft Visual
Studio 2005. On executing the program in debug mode with multiple
breakpoints, the parameters which caused the program to crash at depths
<5m were identified. These parameters were
• AREA_HT_DELTA_Z
• COEFFS_TBL_DELTA_Z
• INTERP_DELTA_Z
• MIN_GRID_THICK
Lower values for these parameters were set and the program re-compiled.
The software was able to then successfully simulate IBS ponds of depth 1.0
m.
0.2 m3 day-1 of anaerobically digested piggery effluent was passed through
each of the stages of the IBS. There was an overall 99.99% reduction in
NH4 concentration in the system. Chlorophyte growth peaked to a maximum
of 90 µg (chl-a) L-1 in the algal pond 1 and 60 µg (chl-a) L-1 in algal pond 2

190
followed by a sharp decrease. Cyanobacteria dominated both the algal ponds
1 and 2 with a maximum concentration of 200 mg m-3 and 150 mg m-3
respectively. Freshwater diatoms did not have any significant growth in
either pond due to low availability of silicon. Zooplankton growth exhibited
short spikes of periodic growth followed by a rapid decline and peaked to a
maximum of 0.6 g C m-3. Fish growth varied over the entire simulation
period and reached a maximum value of 0.8 g C m-3. The results obtained
here were different to those that would have been expected of an IBS, as in
an IBS there is effective management and control mechanisms put in place
which prevent the dominance of cyanobacteria and facilitate the increased
bioconversion rates of phytoplankton, zooplankton and fish, while the
simulations conducted on the proposed commercial scale IBS did not have
any control mechanisms incorporated into the model. A comparison study
with the pilot plant lagoon system in France also showed phytoplankton
growth crashes in summer due to dominance of rotifers as this system also
did not have any control mechanisms. However the researchers at the site in
France discovered that incorporation of certain control strategies, such as,
injection of carbon dioxide and slight agitation (mixing) improved the algal
productivity. The comparative IBS examples sourced from sites in India and
France show that IBS has been successfully implemented in different parts
of the world comprising tools for better control and management of ponds.
The use of mixing (agitation) and aeration assist in mixing the ponds and the
effluent uniformly which minimises stratification in ponds and thus reduces
the growth of cyanobacteria, and in turn improves the growth of
phytoplankton, zooplankton and fish. The model DYRESM CAEDYM
could incorporate the use of mixers and aeration in the IBS ponds to
overcome the problems of algal crashes in summer.
The parameters which can be controlled for managing the commercial scale
IBS in an effective way would be parameters related to inflow and outflow
volumes and flow rates of effluent, retention time of the effluent, nutrient
loads, rates of mixing and aeration within the ponds and control of biomass
conversion for primary, secondary and tertiary productions. These
management strategies could also be used to operate an IBS with a variety

191
of different effluents to its maximum capacity and construct an IBS with
compatible module design.
8.2.3 Sensitivity Analysis
A sensitivity analysis was conducted on selected parameters of the model.
These parameters were subject to minimum, maximum and mean values and
simulations were run with these values. For each simulation, the output was
observed with reference to changes in values of chlorophyll-a concentration.
Chlorophyll a growth was considered to be one of the most important
outputs for this research as it was well correlated with biomass production
in the proposed commercial scale. IBS Parameters for which chlorophyll-a
was highly sensitive were those that directly altered growth rates i.e.
maximum phytoplankton growth rate (Pmax), phytoplankton respiration
coefficient (kr) and phytoplankton temperature multiplier (vR), or indirectly
affect growth rates through their ability to utilize phosphorus ( IPmin and
UPmax) , nitrogen (INmin and UNmax) and light penetration properties (Ik).
8.2.4 Automated Parameter Estimation and Calibration
Parameter calibration for DYRESM CAEDYM has been a tedious and time
consuming process involving manual input of values for every simulation. A
“trial and error” process has been used in the past, which resulted in
unsatisfactory results for model validation.
A calibration program, written in FORTRAN 90, involving Bayesian Monte
Carlo method and GLUE was set up to enable automated parameter
calibration in DYRESM CAEDYM. Due to the unavailability of real time
field data, pseudo field data was generated with random values for this
calibration routine to function properly. The model was calibrated by
minimizing the phytoplankton growth output parameter (chlorophyll-a in
case of chlorophyte calibration and mg m-3 in case of cyanobacteria
calibration).

192
For demonstration purposes, chlorophyte and cyanobacteria growth were
calibrated using the GLUE methodology incorporated into DYRESM
CAEDYM. 1000 Monte Carlo simulations were run using selected
parameters whose range was obtained from literature. For chlorophyte
calibration, a particular set of parameters gave 90.37% agreement (NS value
of 0.9037) with the field data for chlorophyte calibration. For cyanobacteria
calibration, a different set of parameters gave 70.32% agreement (NS value
of 0.7032) with the field data for cyanobacteria calibration.
8.3 Summary
The results presented in this thesis describe the outputs from the anaerobic
digestion experiments, involving both the two-stage pilot and the batch scale
set up, the suitability of DYRESM CAEDYM to model the aquaculture
stages of the IBS and the auto calibration program using FORTRAN 90 to
automatically validate the DYRESM CAEDYM with field data. Validating
the suitability of modelling IBS ponds using DYRESM CAEDYM and the
auto calibration program has closed the gap identified in the literature
review.
8.4 Recommendations for further studies
The mathematical model developed for the aquaculture component of the
IBS using DYRESM CAEDYM as the mathematical modelling tool
represents qualitative data only. Data for validating this model was not
available to the researcher due to the commercial outdoor IBS facility not
being constructed within the stipulated time at Roseworthy, South Australia.
Once the outdoor ponds have been constructed, the IBS facility should be
run on a continuous basis for a period of couple of years and data should be
collected regularly. The data collected should be validated against the
proposed IBS aquaculture model and the automated calibration routine run
several times to minimize the error between the simulated and field data.

193
Further investigation needs to be done on the response of different outputs
e.g. cyanobacteria, freshwater diatoms, zooplankton, fish growth etc in a
effectively controlled and managed system with changes in the parameters
selected for this study and additional parameters as well. This is useful in
identifying the parameters which are most sensitive to a particular output.
However when the number of parameters increases along with a variety of
outputs to be assessed, a manual method of altering parameter values to
observe the change in output is time consuming and more of a “trial and
error” approach. Also this sensitivity study relies on using the minimum,
maximum and mean values of the parameters selected without observing
any changes in the intermediate values, i.e. observing any changes in the
output during incremental changes to the selected parameter.
The automated calibration routine should be tested against other model
outputs e.g. nutrients, temperature, pH, DO etc to enable it to be used for
calibrating different outputs as desired by the modeller. The number of
iterations should also be increased to a larger value (~>100000) to obtain a
better precision in the values of the parameters calibrated. However this is
something that the modeller needs to decide because a balance needs to be
maintained between data precision and extensive computational time.
In recent years there has been an increase in the use and application of
distributed physically-based, water quality and integrated hydrological
models with the increase in the demand for efficiently managing the
precious water resources. There have been multiple issues raised regarding
how to properly calibrate and validate these models and also assess the
uncertainty of the estimated parameters and the spatially-distributed
responses. These issues remain quite unexplored. For complex models,
rigorous parameterisation, reduction of the parameter space and the use of
effective and efficient algorithms are essential to facilitate the process of
calibration and make it more robust. To efficiently calibrate, validate and
use a mathematical model, an automated calibration method is required
which eliminates the conventional techniques of calibrating parameters of
the model with a “trial and error” approach and introduces a sophisticated

194
automated method which reduces the scope of computational errors and
provides more realistic outputs which are of valuable guidance to the
process engineers. Water recycling and re-use applications require an
effective method of management, optimisation and control of the process
parameters and system which can be accomplished by the use of a robust
mathematical model. The robust models come with proper calibration and
validation. This study delivers valuable information and innovative method
to calibrate and validate the use of model.
The use of automated calibration method has high applicability particularly
in the development of mathematical models for managing the performance
of wastewater recycling technology which is being highly sought by the
modern world in order to minimise the impact on diminishing precious
water resources. This calibration technique has also demonstrated that for a
complex aquaculture model like DYRESM CAEDYM where manually
validating the parameters is an unwieldy task, automatic calibration routine
using GLUE methodology is an effective way to validate the model which
reduces the risks of computational errors.

195
9 References
Ambler, J. W. (1986). "Effect of food quantity and quality on egg production of
Acartia tonsa Dana from East Lagoon, Galveston, Texas." Estuarine, Coastal and
Shelf Science 23(2): 183-196.
Andersen, J., J. C. Refsgaard, et al. (2001). "Distributed hydrological modelling of
the Senegal River Basin -- model construction and validation." Journal of
Hydrology 247(3-4): 200-214.
Andrews, J. F. (1969). "Dynamic model of the anaerobic digestion model." J.
Sanit. Engng. 1: 95 - 116.
Angelidaki, I., L. Ellegaard, et al. (1993). "A mathematical model for dynamic
simulation of anaerobic digestion of complex substrates: focussing on ammonia
inhibition." Biotechnology and Bioengineering 42: 159 - 166.
APHA, AWWA, et al. (1998). Standard Methods for the Examination of Water
and Wastewater. Washington DC, USA.
Appels, L., J. Lauwers, et al. (2011). "Anaerobic digestion in global bio-energy
production: Potential and research challenges." Renewable and Sustainable
Energy Reviews 15(9): 4295-4301.
Aronica, G., P. D. Bates, et al. (2002). "Assessing the uncertainty in distributed
model prediction using observed binary pattern information with GLUE."
Hydrological Processes 16: 2001 - 2016.
Aronica, G., B. Hankin, et al. (1998). "Uncertainty and equifinality in calibrating
distributed roughness coefficients in a flood propagation model with limited data."
Advances in Water Resources 22(4): 349-365.

196
Balistrieri, L. S., R. N. Tempel, et al. (2006). "Modeling spatial and temporal
variations in temperature and salinity during stratification and overturn in Dexter
Pit Lake, Tuscarora, Nevada, USA." Applied Geochemistry 21(7): 1184-1203.
Bergman, B., J. R. Gallon, et al. (1997). "N2 Fixation by non-heterocystous
cyanobacteria." FEMS Microbiology Reviews 19(3): 139-185.
Beven, K. (1993). "Prophecy, reality and uncertainty in distributed hydrological
modelling." Advances in Water Resources 16(1): 41-51.
Beven, K. and J. Freer (2001). "Equifinality, data assimilation, and uncertainty
estimation in mechanistic modelling of complex environmental systems using the
GLUE methodology." Journal of Hydrology 249(1-4): 11-29.
Beven, K. J. and A. Binley (1992). "The future of distributed models: model
calibration and uncertainty prediction." Hydrological Processes 6: 279 - 298.
Blasone, R.-S., J. A. Vrugt, et al. (2008). "Generalized likelihood uncertainty
estimation (GLUE) using adaptive Markov Chain Monte Carlo sampling."
Advances in Water Resources 31(4): 630-648.
Boon, P. I., S. E. Bunn, et al. (1994). "Consumption of cyanobacteria by
freshwater zooplankton: implications for the success of 'Top - down' control of
cyanobacterial blooms in Australia." Australian Journal of Marine and Freshwater
Research. 45(5): 875 - 887.
Boyle, D. P., H. V. Gupta, et al. (2000). "Towards improved calibration of
hydrological models: combining the strengths of manual and automatic methods."
Water Resources Research 36: 3663 - 3674.
Brix, H. and H. H. Scheirup (1989). "The use of aquatic macrophytes in water
pollution control." Ambio 18: 100 -107.

197
Bruce, L. C., D. Hamilton, et al. (2006). "A numerical simulation of the role of
zooplankton in C, N and P cycling in Lake Kinneret, Israel." Ecological
Modelling 193(3-4): 412-436.
Bryers, J. D. (1985). "Structured modelling of the anaerobic digestion of biomass
particulates." Biotechnology and Bioengineering. 27: 638 - 649.
Budiastuti, H. (2004). Intensification of Single Stage Continously Stirred Tank
Anaerobic Digestion Process using Carriers. School of Environmental Science.
Perth, Western Australia, Murdoch University.
Burger, D. F., D. P. Hamilton, et al. (2008). "Modelling the relative importance of
internal and external nutrient loads on water column nutrient concentrations and
phytoplankton biomass in a shallow polymictic lake." Ecological Modelling
211(3-4): 411-423.
Cameron, D. S., K. J. Beven, et al. (1999). "Flood frequency estimation by
continous simulation for a gauged upland catchment (with uncertainty)." Journal
of Hydrology 219: 169 - 187.
Carlotti, F., J. Giske, et al. (2000). 12 - Modeling zooplankton dynamics. ICES
Zooplankton Methodology Manual. H. Roger, W. Peter, L. Jürgenet al. London,
Academic Press: 571-667.
Chen, C., R. Ji, et al. (2002). "A model study of the coupled biological and
physical dynamics in Lake Michigan." Ecological Modelling 152(2-3): 145-168.
Chen, Y. R. (1983). "Kinetic analysis of anaerobic digestion of pig manure and its
design implications." Agricultural Wastes 8(2): 65-81.
Chen, Y. R. and A. G. Hashimoto (1978). "Kinetics of methane formation."
Biotechnology and Bioengineering Symposium. 8: 269.

198
Choi, H. T. and K. Beven (2007). "Multi-period and multi-criteria model
conditioning to reduce prediction uncertainty in an application of TOPMODEL
within the GLUE framework." Journal of Hydrology 332(3-4): 316-336.
Christiaens, K. and J. Feyen (2001). "Analysis of uncertainties associated with
different methods to determine soil hydraulic properties and their propagation in
the distributed hydrological MIKE SHE model." Journal of Hydrology 246(1-4):
63-81.
Costello, D. J., P. F. Greenfield, et al. (1991a). "Dynamic modelling of a single -
stage high - rate anaerobic reactor - I. Model derivation." Water Resources 25:
847 - 858.
Costello, D. J., P. F. Greenfield, et al. (1991b). "Dynamic modelling of a single -
stage high - rate anaerobic reactor - II. Model verification." Water Resources 25:
859 - 871.
Crips, S. and M. S. Kumar (2003). Environmental and other impact of
aquaculture. In Aquaculture: Farming fish and Shellfish. L. J. S. a. S. P.C.,
Blackwell Publications, U.K.
Curtis, T. P., D.D. Mara, S.A. Silva (1992). "The effect of sunlight on faecal
coliforms in ponds: Implications for research and design." Water Science and
Technology 26(78): 1729-1738.
Cyr, H. and M. L. Pace (1992). "Grazing by zooplankton and its relationship to
community structure." Canadian Journal of Fisheries and Aquatic Sciences. 49(7):
1455 - 1465.
De Melo, R., R. France, et al. (1992). "Biomanipulation: hit or myth?" Limnology
Oceanography 37: 192 - 207.
Di Termini, I., A. Prassone, et al. (2011). "On the nitrogen and phosphorus
removal in algal photobioreactors." Ecological Engineering 37(6): 976-980.

199
Di Toro, D. M., D. J. O' Connor, et al. (1971). "A Dynamic Model for the
Phytoplankton Population in the Sacramento San Joaquin Delta." Adv. Chem. Ser.
106: 131 - 180.
Donoso-Bravo, A., J. Mailier, et al. (2011). "Model selection, identification and
validation in anaerobic digestion: A review." Water Research 45(17): 5347-5364.
Durbin, A. G. and E. G. Durbin (1992). "Seasonal change in size frequency
distribution and estimated age in the marine copepod Acartia hudsonica during a
winter - spring diatom bloom in the Narragansett Bay. ." Limnology
Oceanography 37: 379 - 392.
Eastman, J. A. and J. F. Ferguson (1981). "Solubilization of Particulate Organic
Carbon during the Acid Phase of Anaerobic Digestion." Journal of the Water
Pollution Control Federation 53: 352 - 366.
Edwards, P. and R. S. V. Pullin (1990). General Discussion on wastewater - fed
aquaculture. . Proceedings of the International Seminar on Wastewater
Reclamation and Reuse for Aquaculture, Calcutta, India.
Egge, J. K. and D. L. Aksnes (1992). "Silicate as regulating nutrient in
phytoplankton competition." Marine Ecology Progress Series 83: 281-289.
Emerson, K. e. a. (1975). "Aqueous Ammonia Equilibrium Calculations: Effect of
pH and temperature. ." J. Fish. Res. Board Can. 32(12).
Engeland, K., I. Braud, et al. (2006). "Multi-objective regional modelling."
Journal of Hydrology 327(3-4): 339-351.
Feyen, L., K. Beven, et al. (2001). "Stochastic capture zone delineation within the
generalized likelihood uncertainty estimation methodology; conditioning on head
observations." Water Resources Research 37(3): 625 - 638.

200
Fiore, M. d. F. t., B. A. Neilan, et al. (2005). "Characterization of nitrogen-fixing
cyanobacteria in the Brazilian Amazon floodplain." Water Research 39(20): 5017-
5026.
Fisher, H. B., E. G. List, et al. (1979). Mixing in Inland and Coastal Waters.,
Academic Press.
Flachowsky, G. and A. Hennig (1990). "Composition and digestibility of
untreated and chemically treated animal excreta for ruminants--A review."
Biological Wastes 31(1): 17-36.
Fragoso Jr, C. R., D. M. L. M. Marques, et al. (2008). "Modelling spatial
heterogeneity of phytoplankton in Lake Mangueira, a large shallow subtropical
lake in South Brazil." Ecological Modelling 219(1-2): 125-137.
Freer, J., K. J. Beven, et al. (1996). "Bayesian estimation of uncertainty in runoff
prediction and the value of data: an application of the GLUE approach." Water
Resources Research 32(7): 2161 - 2173.
Freer, J. E., H. McMillan, et al. (2004). "Constraining dynamic TOPMODEL
responses for imprecise water table information using fuzzy rule based
performance measures." Journal of Hydrology 291(3-4): 254-277.
Freund, J. A., S. Mieruch, et al. (2006). "Bloom dynamics in a seasonally forced
phytoplankton–zooplankton model: Trigger mechanisms and timing effects."
Ecological Complexity 3(2): 129-139.
Frost, B. W. (1972). "Effects of size and concentration of food particles on the
feeding behaviour of the marine planktonic copepod Calanus pacificus. ."
Limnology Oceanography. 17(6): 805 - 815.
Gal, G., J. Imberger, et al. (2003). "Simulating the thermal dynamics of Lake
Kinneret." Ecological Modelling 162(1-2): 69-86.

201
Gamble, J. C. (1978). "Copepod grazing during a declining spring phytoplankton
bloom in the northern North Sea. ." Marine Biology. 49: 303 - 315.
Georgakakos, K. P., D.-J. Seo, et al. (2004). "Towards the characterization of
streamflow simulation uncertainty through multimodel ensembles." Journal of
Hydrology 298(1-4): 222-241.
Gilbert, P. M. (1998). "Interactions of top - down and bottom - up control in
planktonic nitrogen cycling." Hydrobiologia 491: 91 - 100.
Gopakumar, K., S. Ayyappan, et al. (2000). Present Status of Integrated Fish
Farming in India and Wastewater Treatment Through Aquaculture Proceedings of
the National Workshop on Wastewater Treatment and Integrated Aquaculture.
Adelaide, South Australian Research & Development Institute (SARDI). 22 - 37.
Graef, S. P. and J. F. Andrews (1974). "Stability and control of Anaerobic
Digestion." Journal WPCF 46: 667 - 682.
Griffin, S. L., M. Herzfeld, et al. (2001). "Modelling the impact of zooplankton
grazing on phytoplankton biomass during a dinoflagellate bloom in the Swan
River Estuary, Western Australia." Ecological Engineering 16(3): 373-394.
Griffiths, F. B. and J. Caperon (1979). "Description and use of an improved
method for determining estuarine zooplankton grazing rates on phytoplankton. ."
Marine Biology 54: 301 - 309.
Gupta, H. V., S. A. Sorooshian, et al. (1998). "Toward improved calibration of
hydrologic models:multiple and noncommensurable measures of information."
Water Resources Research 34: 751 - 763.
Guven, B. and A. Howard (2006). "Modelling the growth and movement of
cyanobacteria in river systems." Science of The Total Environment 368(2-3): 898-
908.

202
Håkanson, L. and V. V. Boulion (2003). "Modelling production and biomasses
of herbivorous and predatory zooplankton in lakes." Ecological Modelling 161(1-
2): 1-33.
Håkanson, L., A. C. Bryhn, et al. (2007). "On the issue of limiting nutrient and
predictions of cyanobacteria in aquatic systems." Science of The Total
Environment 379(1): 89-108.
Håkanson, L. and V. V. Boulion (2003). "Modelling production and biomasses of
herbivorous and predatory zooplankton in lakes." Ecological Modelling 161(1-2):
1-33.
Hamilton, D. P., G. Schladow, et al. (1995). "Controlling the indirect effects of
flow diversions on water quality in an Australian reservoir." Environment
International 21(5): 583-590.
Hamilton, D. P. and S. G. Schladow (1997). "Prediction of water quality in lakes
and reservoirs. Part I -- Model description." Ecological Modelling 96(1-3): 91-
110.
Harper, S. R. and F. G. Pohland (1986). "Recent Development in Hydrogen
Measurement during Anaerobic Biological Wastewater Treatment."
Biotechnology and Bioengineering 28: 585 - 602.
Harris, G. P. (1986). Phytoplankton Ecology - Structure, Function and
Fluctuation. London, Chapman and Hall.
Havens, K. E., R. T. James, et al. (2003). "N:P ratios, light limitation, and
cyanobacterial dominance in a subtropical lake impacted by non-point source
nutrient pollution." Environmental Pollution 122(3): 379-390.
Heidari, A., B. Saghafian, et al. (2006). "Assessment of flood forecasting lead
time based on generalized likelihood uncertainty estimation." Stochastic
Environmental Research and Risk Assessment 20(5): 363 - 380.

203
Henriksen, H. J., L. Troldborg, et al. (2003). "Methodology for construction,
calibration and validation of a national hydrological model for Denmark." Journal
of Hydrology 280(1-4): 52-71.
Hense, I. and A. Beckmann (2006). "Towards a model of cyanobacteria life
cycle—effects of growing and resting stages on bloom formation of N2-fixing
species." Ecological Modelling 195(3-4): 205-218.
Hense, I. and H. Burchard (2010). "Modelling cyanobacteria in shallow coastal
seas." Ecological Modelling 221(2): 238-244.
Hill, D. T. (1982). "A comprehensive dynamic model for animal waste
methanogenesis." Transactions of the ASAF 25: 1374 - 1380.
Hill, D. T. and C. L. Barth (1977). "A dynamic model for simulation of animal
waste digestion." Journal WPCF 10: 2129 - 2143.
Hipsey, M. R., J. R. Romero, et al. (2006). Computational Aquatic Ecosystem
Dynamics Model v2.3 Science Manual, Centre for Water Research, University of
Western Australia, Australia.
Hodges, B. and C. Dallimore (2001). Estuary and Lake Computer Model,
ELCOM, Science Manual, code version 1.5.0, Centre for Water Research,
University of Western Australia.
Hodgkin, E. P. and R. J. Rippingale (1971). "Interspecies conflict in estuarine
copepods." Limnology Oceanography 16: 573 - 576.
Hongping, P. and M. Jianyi (2002). "Study on the algal dynamic model for West
Lake, Hangzhou." Ecological Modelling 148(1): 67-77.

204
Hornberger, G. M. and R. C. Spear (1981). "An approach to the preliminary
analysis of environmental systems." Journal of Environmental Management 12: 7
- 18.
Huang, A. A. (1975). "Enzymatic Hydrolysis of Cellulose to Sugar."
Biotechnology and Bioengineering Symposium. 5: 245 - 252.
Hudson, J. J. and W. D. Taylor (1996). "Measuring regeneration of dissolved
phosphorus in planktonic communities." Limnol. Oceanogr. 41: 1560 - 1565.
Hudson, J. J., W. D. Taylor, et al. (1999). "Planktonic nutrient regeneration and
cycling efficiency in temperate lakes." Nature 400: 659 - 661.
Hunt, R. J. and V. F. Matveev (2005). "The effects of nutrients and zooplankton
community structure on phytoplankton growth in a subtropical Australian
reservoir: An enclosure study." Limnologica - Ecology and Management of Inland
Waters 35(1-2): 90-101.
Imberger, J. (1981). "The influence of stream salinity on reservoir water quality."
Agricultural Water Management 4(1-3): 255-273.
Imberger, J., J. C. Patterson, et al. (1978). "Dynamics of Reservoirs of Medium
Size." Journal of Hydrology, Division, ASCE 104(725 - 743).
Imerito, A. (2007). Dynamic Reservoir Simulation Model v4.0 Science Manual,
Centre for Water Research, University of Western Australia, Australia.
Islam, M. S. e. a. (1996). "Faecal contamination of a fish culture farm where
duckweed grown in hospital wastewater are used as fish feed." Proceedings of the
5th Annual Scientific Conference, ASCON, Dhaka, Bangladesh: 49.
Jefferson T, T. (2010). "Zooplankton community grazing impact on a bloom of
Alexandrium fundyense in the Gulf of Maine." Harmful Algae 9(6): 578-589.

205
Ji, R., C. Chen, et al. (2002). "Influences of suspended sediments on the
ecosystem in Lake Michigan: a 3-D coupled bio-physical modeling experiment."
Ecological Modelling 152(2-3): 169-190.
Jonathan P, Z. (2011). "Nitrogen fixation by marine cyanobacteria." Trends in
Microbiology 19(4): 162-173.
Jørgensen, S. E. (1995). "State of the art of ecological modelling in limnology."
Ecological Modelling 78(1-2): 101-115.
Jørgensen, S. E., L. A. Jørgensen, et al. (1981). "Parameter estimation in
eutrophication modelling." Ecological Modelling 13(1-2): 111-129.
Jørgensen, S. E., L. Kamp-Nielsen, et al. (1975). "A submodel for anaerobic mud-
water exchange of phosphate." Ecological Modelling 1(2): 133-146.
Jørgensen, S. E., L. Kamp-Nielsen, et al. (1986). "Examination of the generality
of eutrophication models." Ecological Modelling 32(4): 251-266.
Jørgensen, S. E., H. Mejer, et al. (1978). "Examination of a lake model."
Ecological Modelling 4(2-3): 253-278.
Kardinaal, W. E. A. and P. M. Visser (2005). "Dynamics of Cyanobacterial
Toxins: Sources of Variability in Microcystin Concentrations. ." Harmful
Cyanobacteria.
Kleinstreuer, C. and T. Powegha (1982). "Dynamic simulator for anaerobic
digestion process." Biotechnology and Bioengineering. 24: 1941 - 1951.
Klepper, O., H. Scholten, et al. (1991). "Predicition uncertainty in an ecological
model of the Oosterchelde Estuary." Journal of Forecasting 10: 191 - 209.

206
Kranert M., H. K. (2000). Anaerobic Digestion of Organic Wastes - Process
Parameters and Balances in Practice. Material Flow Analysis of Integrated
Biosystems.
Krebs, C. J. (1985). Ecology: The Experimental Analysis of Distribution and
Abundance. New York, Harper & Row.
Krivtsov, V., C. Goldspink, et al. (2001). "Expansion of the model [`]Rostherne'
for fish and zooplankton: role of top-down effects in modifying the prevailing
pattern of ecosystem functioning." Ecological Modelling 138(1-3): 153-171.
Kuczera, G. and E. Parent (1998). "Monte Carlo assessment of parameter
uncertainty in conceptual catchment models: the Metropolis algorithm." Journal of
Hydrology 211(1-4): 69-85.
Kumar, M. S. (2002). Integrated Farming for Sustainable Primary Industry:
“Water and Nutrient Recycling through Integrated Aquaculture” Proceedings of
the Infom 2000 National workshop on Integrated Food Production and Resource
management. “Integrated biosystem for sustainable Development”. W. a. Pillai-
McGrarry: 51 - 62.
Kumar, M. S. and S. Crips (2012). Environmental and Other Impact of
Aquaculture, Lucas, J.S. and Southgate P.C. Blackwell Publications, UK.
Kumar, M. S. and M. Sierp (2003). Integrated Wastewater Treatment and
Aquaculture Production. Adelaide, Rural Industries Research and Development
Corporation: 1 - 62.
Lamb, R., K. Beven, et al. (1998). "Use of spatially distributed water table
observations to constrain uncertainty in a rainfall-runoff model." Advances in
Water Resources 22(4): 305-317.

207
Lampert, W. and B. E. Taylor (1985). "Zooplankton grazing in a eutrophic lake:
implications of diel vertical migration." Ecology. 66: 68 - 82.
Le Hyaric, R., J.-P. Canler, et al. (2010). "Pilot-scale anaerobic digestion of
screenings from wastewater treatment plants." Bioresource Technology 101(23):
9006-9011.
Lee, M.-Y., C.-W. Suh, et al. (2009). "Variation of ADM1 by using temperature-
phased anaerobic digestion (TPAD) operation." Bioresource Technology 100(11):
2816-2822.
Lehman, J. T. (1980). "Release and cycling of nutrients between planktonic algae
and herbivores." Limnol. Oceanogr. 25: 620 - 632.
Levine, S. N., M. A. Borchardt, et al. (1999). "The Impact of Zooplankton
Grazing on Phytoplankton Species Composition and Biomass in Lake Champlain
(USA-Canada)." Journal of Great Lakes Research 25(1): 61-77.
Lewis, D. M., J. D. Brookes, et al. (2004). "Numerical Models for management of
Anabaena Circinalis." Journal of Applied Phycology 16: 457 - 468.
Lin, C. and K. Sato (1986). "Methanogenic Digestion Using Mixed Substrate of
Acetic, Propionic and Butyric Acids." Water Research 20(3): 385 - 394.
Lionard, M., F. Azemar, et al. (2005). "Grazing by meso- and microzooplankton
on phytoplankton in the upper reaches of the Schelde estuary." Estuarine, Coastal
and Shelf Science 64(4): 764-774.
Lunte, C. C. and C. Leucke (1990). "Trophic interactions of leptodora in Lake
Mendota." Limnol. Oceanogr. 35: 1091 - 1100.
Lyberatos, G. and I. V. Skiadas (1999). "Modelling of Anaerobic Digestion - A
Review." Global Nest: the Int. J 1(2): 63 - 76.

208
Mackas, D. and R. Bohrer (1976). "Flouresecence analysis of zooplankton gut
contents and investigation of diel feeding patterns. ." Journal of Experimental and
Marine Biology and Ecology. 25: 77 - 85.
Madsen, H. (2000). "Automatic calibration of a conceptual rainfall-runoff model
using multiple objectives." Journal of Hydrology 235(3-4): 276-288.
Madsen, H. (2003). "Parameter estimation in distributed hydrological catchment
modelling using automatic calibration with multiple objectives." Advances in
Water Resources 26(2): 205-216.
Madsen, M., J. B. Holm-Nielsen, et al. (2011). "Monitoring of anaerobic digestion
processes: A review perspective." Renewable and Sustainable Energy Reviews
15(6): 3141-3155.
Mansson, T., Foo, E.L. (1998). Swedish efforts in integrating bio-fuels as
alternative fuels for transportation in buses, lorries and cars. Proceedings of the
Internet Conference on Integrated Biosystems.
Mantovan, P. and E. Todini (2006). "Hydrological forecasting uncertainty
assessment: Incoherence of the GLUE methodology." Journal of Hydrology 330(1
- 2): 368 - 381.
Marino, R. W. and R. Howarth (2009). Nitrogen Fixation. Encyclopedia of Inland
Waters. E. L. Editor-in-Chief:  Gene. Oxford, Academic Press: 65-72.
Martin, J. H. (1970). "Phytoplankton - zooplankton relationships in Narragansett
Bay. 4. The seasonal importance of grazing. ." Limnology Oceanography. 15: 413
- 418.
Matsuoka, Y., T. Goda, et al. (1986). "An eutrophication model of Lake
Kasumigaura." Ecological Modelling 31(1-4): 201-219.

209
McAllister, C. D. (1971). Some aspects of nocturnal and continuous grazing by
planktonic herbivores in relation to production studies. . Fisheries Research Board
of Canada.
McInerney, M. and M. Bryant (1981). "Basic Principles of Bioconversions in
Anaerobic Digestion of Methanogenesis." Biomass Conversion Processes for
Energy and Fuels: 227 - 296.
McInerney, M. J. and P. Beaty (1988). "Anaerobic Community Structure from a
Non Equilibrium Thermodynamic Perspective." Canadian Journal of
Microbiology 34: 487 - 493.
McMichael, C. E., A. S. Hope, et al. (2006). "Distributed hydrological modelling
in California semi - arid shrublands: MIKE SHE model calibration and
uncertainty estimation." Journal of Hydrology 317(3 - 4): 307 - 324.
Mertens, J., H. Madsen, et al. (2004). "Including prior information in the
estimation of effective soil parameters in unsaturated zone modelling." Journal of
Hydrology 294(4): 251-269.
Mitra, A. and K. J. Flynn (2006). "Accounting for variation in prey selectivity by
zooplankton." Ecological Modelling 199(1): 82-92.
Miyanaga, Y. (1986). "Modelling of stratified flow and eutrophication in
reservoirs." Ecological Modelling 31(1-4): 133-144.
Moletta, R., D. Verrier, et al. (1986). "Dynamic modelling of anaerobic
digestion." Water Research 20(4): 427-434.
Molina Grima, E., F. G. A. Fernández, et al. (1999). "Photobioreactors: light
regime, mass transfer, and scaleup." Journal of Biotechnology 70(1-3): 231-247.

210
Montanari, A. (2005). "Large sample behaviours of the generalized likelihood
uncertainty estimation (GLUE) in assessing the uncertainty of rainfall - runoff
simulations." Water Resources Research 41(W08406).
Moradkhani, H., K.-L. Hsu, et al. (2005). "Uncertainty assessment of hydrologic
model states and parameters; sequential data assimilation using the particle filter."
Water Resources Research 41(5): 1 - 17.
Mosey, F. E. (1983). "Mathematical modelling of the anaerobic digestion process:
regulatory mechanisms for the formation of short chain volatile acids from
glucose." Water Science Technology 15: 209 - 232.
Mueller, D. K. (1982). "Mass balance model for estimation of phosphorus
concentration in reservoirs." Water Resources Bulletin 18: 377 - 382.
Muleta, M. K. and J. W. Nicklow (2005). "Sensitivity and uncertainty analysis
coupled with automatic calibration for a distributed watershed model." Journal of
Hydrology 306(1-4): 127-145.
Mullin, M. M. and E. R. Brooks (1970). "The effect of concentration of food on
body size, cumulative ingestion, and rate of growth of the marine copepod
Calanus helgolandicus. ." Limnology Oceanography. 15: 748 - 755.
Mur, L. R., O. M. Skulberg, et al. (1999). "Cyanobacteria in the Environment."
Toxic Cyanobacteria in Water: A Guide to their Public Health Consequences,
Monitoring and Management.(World Health Organisation).
Myint, M. and N. Nirmalakhandan (2006). "Evaluation of First Order, Second
Order and Surface Limiting Reactions in Anaerobic Hydrolysis of Cattle
Manure." Environmental Engineering Science 23(6): 966 - 976.
Myint, M., N. Nirmalakhandan, et al. (2006). "Anaerobic Fementation of Cattle
Manure: Modelling of Hydrolysis and Acidogenesis." Water Research In Press,
Corrected Proof.

211
Nash, J. E. and J. V. Sutcliffe (1970). "River flow forecasting through conceptual
models part I -- A discussion of principles." Journal of Hydrology 10(3): 282-290.
Noike, T. and G. Endo (1985). "Characteristic of Carbohydrate Degradation and
the Rate Limiting Step in Anaerobic Digestion." Biotechnology and
Bioengineering 27(10): 1482 - 1489.
Osidele, O. O. and M. B. Beck (2004). "Food web modelling for investigating
ecosystem behaviour in large reservoirs of the south-eastern United States: lessons
from Lake Lanier, Georgia." Ecological Modelling 173(2-3): 129-158.
Ostfeld, A. and S. Salomons (2005). "A hybrid genetic--instance based learning
algorithm for CE-QUAL-W2 calibration." Journal of Hydrology 310(1-4): 122-
142.
Paerl, H. W., H. Xu, et al. (2011). "Controlling harmful cyanobacterial blooms in
a hyper-eutrophic lake (Lake Taihu, China): The need for a dual nutrient (N
& P) management strategy." Water Research 45(5): 1973-1983.
Pappenberger, F., K. Beven, et al. (2007). "Grasping the unavoidable subjectivity
in calibration of flood inundation models: A vulnerability weighted approach."
Journal of Hydrology 333(2-4): 275-287.
Pappenberger, F., K. Beven, et al. (2005). "Uncertainty in the calibration of
effective roughness parameters in HEC-RAS using inundation and downstream
level observations." Journal of Hydrology 302(1-4): 46-69.
Parker, W. J. (2005). "Application of the ADM1 model to advanced anaerobic
digestion." Bioresource Technology 96(16): 1832-1842.
Pearson, H. W., D.D. Mara, L.R. Cawley, H.M. Arridge, S.A. Silva (1996). "The
performance of an innovative tropical experiment waste stabilization pond system
operating at high organic loadings." Water Science and Technology 33(7): 63-73.

212
Pearson, H. W., D.D. Mara, S.W. Mills, D.J. Smallman (1987). "Physico-
chemical parameters influencing faecal bacterial survival in waste stabilization
ponds." Water Science and Technology 11(12): 145-152.
Peterson, W. T., S. Painting, et al. (1990). "Diel variation in gut pigment content,
diel vertical migration and estimates of grazing impact for copepods in the
southern Benguella upwelling region in October 1987." Journal of Plankton
Research. 12: 259 - 281.
Poulsen, O. M. and L. W. Peterson (1985). "A Standard Formula for the
Determination of the Initial Rate of Hydrolysis of Carboxymethylcellulase."
Biotechnology and Bioengineering 27: 409 - 414.
Pullammanappallil, P., D. P. Chynoweth, et al. (2001). "Stable performance of
anaerobic digestion in the presence of high concentration of propionic acid."
Bioresource Technology 78: 165 - 169.
Pullammanappallil, P., J. M. Owens, et al. (1991). "Dynamic model for
conventionally mixed anaerobic digestion reactors." AIChE Annual
Meeting(277c): 43 - 53.
Rajagopal, R., P. Rousseau, et al. (2011). "Combined anaerobic and activated
sludge anoxic/oxic treatment for piggery wastewater." Bioresource Technology
102(3): 2185-2192.
Ramirez, I., E. I. P. Volcke, et al. (2009). "Modeling microbial diversity in
anaerobic digestion through an extended ADM1 model." Water Research 43(11):
2787-2800.
Ratto, M., S. Tarantola, et al. (2001). "Sensitivity analysis in model calibration:
GSA-GLUE approach." Computer Physics Communications 136(3): 212-224.

213
Ray, S., L. Berec, et al. (2001). "Optimization of exergy and implications of body
sizes of phytoplankton and zooplankton in an aquatic ecosystem model."
Ecological Modelling 140(3): 219-234.
Reeve, M. R. and M. A. Walter (1977). "Observations on the existence of lower
threshold and upper critical food concentrations for the copepod Acartia tonsa
Dana." Journal of Experimental and Marine Biology and Ecology. 29: 211 - 221.
Refsgaard, J. C. (1997). "Parameterisation, calibration and validation of
distributed hydrological models." Journal of Hydrology 198(1-4): 69-97.
Riley, M. J. and H. G. Stefan (1988). "Minlake: A dynamic lake water quality
simulation model." Ecological Modelling 43(3-4): 155-182.
Rippingale, R. J. (1994). "A calanoid copepod Gladioferans imparipes, holding
onto surfaces." Hydrobiology. 292/293: 351 - 360.
Rippingale, R. J. and E. P. Hodgkin (1974a). "Predation effects on the distribution
of a copepod." Australian Journal of Marine and Freshwater Research. 25: 81 - 91.
Rippingale, R. J. and E. P. Hodgkin (1974b). "Population growth of a copepod
Gladioferens imparipes Thomson." Australian Journal of Marine and Freshwater
Research. 25: 351 - 360.
Rippingale, R. J. and E. P. Hodgkin (1977). "Food availability and salinity
tolerance in a brackish water copepod. ." Australian Journal of Marine and
Freshwater Research. 28: 1 - 7.
Robson, B. J. and D. P. Hamilton (2004). "Three-dimensional modelling of a
Microcystis bloom event in the Swan River estuary, Western Australia."
Ecological Modelling 174(1-2): 203-222.

214
Romero, J. R., J. P. Antenucci, et al. (2004). "One- and three-dimensional
biogeochemical simulations of two differing reservoirs." Ecological Modelling
174(1-2): 143-160.
Romli, M. (1993). "Modelling and Verification of a Two Stage Anaerobic
Wastewater Treatment System." Chemical Engineering.
Rozzi, A. and S. Merlini (1985). "Development of a Four Population Model of the
Anaerobic Degradation of Carbohydrate." Environmental Technology Letters 6:
10 - 19.
Rukhovets, L. A., G. P. Astrakhantsev, et al. (2003). "Development of Lake
Ladoga ecosystem models: modeling of the phytoplankton succession in the
eutrophication process. I." Ecological Modelling 165(1): 49-77.
Salomoni, C., A. Caputo, et al. (2011). "Enhanced methane production in a two-
phase anaerobic digestion plant, after CO2 capture and addition to organic
wastes." Bioresource Technology 102(11): 6443-6448.
Santos, M. a. d. l. Å. M. n., J. Å. S. L√≥pez, et al. (2010). "Modelling the
anaerobic digestion of wastewater derived from the pressing of orange peel
produced in orange juice manufacturing." Bioresource Technology 101(11): 3909-
3916.
Scavia, D. (1980). "An ecological model of Lake Ontario." Ecological Modelling
8: 49-78.
Schladow, S. G. (1984). "The upper mixed zone of a salt gradient solar pond: Its
dynamics, prediction and control." Solar Energy 33(5): 417-426.
Schladow, S. G. and D. P. Hamilton (1997). "Prediction of water quality in lakes
and reservoirs: Part II - Model calibration, sensitivity analysis and application."
Ecological Modelling 96(1-3): 111-123.

215
Sevrin-Reyssac, J. (1998). "Biotreatment of swine manure by production of
aquatic valuable biomasses." Agriculture Ecosystems & Environment 68: 177-
186.
Siegrist, H., D. Renggli, et al. (1993). "Mathematical modelling of anaerobic
mesophilic sewage sludge treatment." Water Science Technology 27: 25-36.
Smith, P. H., F. M. Bordeaux, et al. (1988). "Biological Production of Methane
from Biomass." In: Methane from Biomass. A treatment approach: 291 - 334.
Sonnenborg, T. O., B. S. B. Christensen, et al. (2003). "Transient modeling of
regional groundwater flow using parameter estimates from steady-state automatic
calibration." Journal of Hydrology 273(1-4): 188-204.
Spillman, C. M., J. Imberger, et al. (2007). "Modelling the effects of Po River
discharge, internal nutrient cycling and hydrodynamics on biogeochemistry of the
Northern Adriatic Sea." Journal of Marine Systems 68(1-2): 167-200.
Stearns, D. E., W. Litaker, et al. (1987). "Impacts of zooplankton grazing and
excretion on short-interval fluctuations in chlorophyll a and nitrogen
concentrations in a well-mixed estuary." Estuarine, Coastal and Shelf Science
24(3): 305-325.
Stefan, H. and D. E. Ford (1975). "Temperature Dynamics in Dimictic Lakes."
Journal of Hydrology, Division, ASCE 11058: 97 - 114.
Stickney, R. (2000). Encyclopaedia of Aquaculture., John Wiley & Sons Inc.:
1063.
Svensson, J.-E. and J. A. E. Stenson (1991). "Herbivoran impact on
phytoplankton community structure." Hydrobiology 224: 77 - 84.
T.V.A. (1972). Heat and mass transfer between a water surface and atmosphere.
Water Resouces Research Laboratory Report 14.

216
Taconi, K. A., M. E. Zappi, et al. (2008). "Methanogenesis under acidic pH
conditions in a semi-continuous reactor system." Bioresource Technology 99(17):
8075-8081.
Thauer, R. K. and K. Jungermann (1977). "Energy Conservation in Chemotrophic
Anaerobic Bacteria." Bact. Rev. 41(1): 100 - 108.
Thiemann, M., M. Trosset, et al. (2001). "Bayesian recursive parameter estimation
for hydrological models." Water Resources Research 37(10): 2521 - 2535.
Trolle, D., H. Skovgaard, et al. (2008). "The Water Framework Directive: Setting
the phosphorus loading target for a deep lake in Denmark using the 1D lake
ecosystem model DYRESM-CAEDYM." Ecological Modelling 219(1-2): 138-
152.
Turner, J. T. and E. Granelli (1992). "Zooplankton feeding ecology: grazing
during enclosure studies of phytoplankton blooms from the west coast of Sweden.
." Journal of Experimental and Marine Biology and Ecology. 157(1): 19 - 31.
Ugwu, C. U., H. Aoyagi, et al. (2008). "Photobioreactors for mass cultivation of
algae." Bioresource Technology 99(10): 4021-4028.
Ute, M. (2003). "Cyanobacteria associations in shallow polytrophic lakes:
influence of environmental factors." Acta Oecologica 24, Supplement 1(0): S11-
S23.
Vanni, M. J. (1987). "Effects of nutrients and zooplankton size on the structure of
a phytoplankton community." Ecology. 68: 624 - 635.
Vázquez, R. F. and J. Feyen (2007). "Assessment of the effects of DEM gridding
on the predictions of basin runoff using MIKE SHE and a modelling resolution of
600 m." Journal of Hydrology 334(1-2): 73-87.

217
Vogel, R. M., R. Batchelder, et al. "Appraisal of the generalized likelihood
uncertainty estimation (GLUE) method." Water Resources Research in press.
Vojtƒõch, V. l. (1987). "Interactions between algae and zooplankton in a
continuous cultivation system." Ecological Modelling 39(1-2): 33-43.
Vrugt, J. A., C. Diks, et al. (2005). "Improved treatment of uncertainty in
hydrologic modelling: combining the strengths of global optimization and data
assimilation." Water Resources Research 41(1): 1 - 17.
Vrugt, J. A., H. V. Gupta, et al. (2003). "Effective and efficient algorithm for
multi objective optimization of hydrologic models." Water Resources Research
39(8): 1214.
Vrugt, J. A., H. V. Gupta, et al. (2003). "A Shuffled Complex Evolution
Metropolis algorithm for optimization and uncertainty assessment of hydrologic
model parameters." Water Resources Research 39(8): 1201.
Vrugt, J. A. and B. A. Robinson (2007). "Treatment of uncertainty using
ensemble methods: comparison of sequential data assimilation and Bayesian
model averaging." Water Resources Research 43(W01411).
Warburton, K. (2001). Recycled organics, integrated biosystems and the role of
planning. 2nd National Recycled Organics Conference and Technology Expos,
The University of Queensland, Gatton Recycled Organics Consortium.
Warburton, K., D. Ramage, et al. (2002). Integrated Biosystems for Sustainable
Development. Proceedings of the InFoRM 2000 National Workshop on Integrated
Food Production and Resource Management., Rural Industries Research and
Development Corporation (RIRDC).
Wen, Y. H. and R. H. Peters (1994). "Empirical models of phosphorus and
nitrogen excretion rates by zooplankton." Limnol. Oceanogr. 39: 1669 - 1679.

218
Whitehouse, A. E., Straughan, J.R. (2006) Ammonia-The Rest of the Story.
Williams, C., T. Biswas, et al. (2007). "Use of recycled water from intensive
primary industries to grow crops within integrated biosystems." Agricultural
Science 21(2): 34 - 36.
Yapo, P. O., H. V. Gupta, et al. (1998). "Multi-objective global optimization for
hydrologic models." Journal of Hydrology 204(1-4): 83-97.
Yasui, H., R. Goel, et al. (2008). "Modified ADM1 structure for modelling
municipal primary sludge hydrolysis." Water Research 42(1-2): 249-259.
Yu, H., Q. and X. J. Zheng (2003). "High Rate Anaerobic Hydrolysis and
Acidogenesis of Sewage Sludge in a Modified Upflow Reactor." Water Science
and Technology 48(4): 69 - 75.
Yu, H., F. Wilson, et al. (1998). "Kinetic analysis of an anaerobic filter treating
soybean wastewater." Water Research 32(11): 3341-3352.
Yu, L., Q. Zhao, et al. (2012). "Experimental and modeling study of a two-stage
pilot scale high solid anaerobic digester system." Bioresource Technology 124(0):
8-17.
Zehnder, A. J. B. (1978). "Ecology of Methane Formation." Water Pollution
Microbiology 2: 349 - 376.
Zhang, L., Y.-W. Lee, et al. (2011). "Anaerobic co-digestion of food waste and
piggery wastewater: Focusing on the role of trace elements." Bioresource
Technology 102(8): 5048-5059.
Zhang, Z. S. (1990). "Wastewater-fed aquaculture." Proceedings of the
International Seminar on wastewater reclamation and resuse for aquaculture,
Calcutta, India, 6-9 Dec.

219
Appendix A Anaerobic Digestion Models
Models that assume substrate inhibited Monod kinetics of the
methanogens
Graef & Andrews Model (1974):
The (Graef and Andrews 1974) model involves the acetoclastic
methanogens. The conversion of fatty acids into biogas is considered
limiting. Volatile fatty acids are expressed as acetic acid and the
methanogens composition is assumed to be C5H7NO2. The overall reaction
is represented as
4227533 92.092.0032.0032.0 CHCONOHCNHCOOHCH ++→+
Eq A.1
(Andrews 1969) assumed the Monod kinetics with substrate inhibition. The
equation is given by
i
S
KI
SK
++=1
maxµµ Eq A.2
where µ is the specific growth rate
µmax is the maximum specific growth rate
KS is the half velocity constant
S is the concentration of growth limiting substrate
Ki is the inhibition constant
I is the inhibitor concentrator
No experimental verification of the Graef & Andrews model has been done
till date.

220
Hill & Barth Model (1972)
The model considers hydrolysis, acidogenesis and ammonia inhibition.
Block Diagram of (Hill and Barth 1977) mathematical model (Lyberatos and Skiadas 1999)
Kleinstreuer & Powegha Model (1982)
This model involves hydrolysis of biodegradable solids, acetogenesis and
methanogenesis, dependent on pH and temperature.
Process stages of anaerobic digestion given by (Kleinstreuer and Powegha 1982) (Lyberatos and Skiadas 1999)
Insoluble
Inorganics
Soluble
Organics
Volatile
Organic
Acids
CO2
Extracellular
Enzymes
Acid
Formers
Methane
Formers
CH4 NH3
Fats/Lipids
Carbohydrates
Proteins
Acetogenic
Bacteria
Methanogenic
Bacteria
Soluble
Organic
Compounds
Acetate
H2,
CO2
CH4
+
CO2

221
Moletta et al., (1986) Model:
This model involves acidogenesis step that forms acetate from glucose and
an inhibition by undissociated acetic acid.
Flow chart of (Moletta, Verrier et al. 1986) model (Lyberatos and Skiadas 1999)
Smith et al., (1988) Model:
This model assumes a slow and fast hydrolysis steps. Acidogenesis of the
soluble intermediates and methanogenesis are also taken into account.
Flow chart of (Smith, Bordeaux et al. 1988) model (Lyberatos and Skiadas 1999)
Acidogenic
bacteria Methanogenic
bacteria Easily
fermentable
organics
(glucose
equivalent)
Organic
Acids
(acetate
equivalent)
CH4
VFAs
Rapidly degradable
biomass
Slowly degradable
biomass
Acidogenic
Bacteria
Methanogenic
Bacteria
Soluble
Organic
Matter
CH4
+
CO2

222
Models where the influence of pH and volatile fatty acids (VFAs) is
taken into account
Hill’s Model (1982):
This model was especially developed for describing digestion of manure
and animal waste. The model considers hydrolysis, acidogenesis and
ammonia inhibition. It assumes that methanogenesis depends on total fatty
acids. The bacterial groups participating in the digestion process are
• Acidogenic – form a mixture of acetic, propionic and butyric acids.
• Hydrogenogenic – convert propionic and butyric acid into acetic acid
and H2.
• Homoacetogenic – produces acetate from H2 and CO2.
• H2 – methanogenic – reduces CO2 into CH4.
• Acetate – methanogenic – converts acetic acid into biogas.
Inhibition occurs due to total fatty acid concentration.
Flow Chart of (Hill 1982) model (Lyberatos and Skiadas 1999)
Glucose
Acidogenic bacteria
Hydrogenogenic bacteria
Homoacetogenic bacteria
Acetate methanogenic
bacteria H2 methanogenic
bacteria
CH4 + CO2 CH4 + H2O
H2 + CO2 Acetate
Propionate Butyrate

223
Bryer’s Model (1985):
Bryer’s Model (Bryers 1985) considers volatile fatty acids as the key
parameter, but also considers the influence of other parameters such as pH.
Flow Chart of (Bryers 1985) Model (Lyberatos and Skiadas 1999)
Insoluble Organic Matter
Amino acids, simple sugars Fatty acids
Acid forming bacteria
Propionate
Propionate utilizing bacteria
Methanogenic bacteria Acetate H2
CH4

224
Models where the process is primarily controlled by the H2
concentration in the reactor
Mosey’s Model (1983):
(Mosey 1983) considered the H2 partial pressure as the key regulatory
parameter of the anaerobic digestion of glucose. The model considers four
bacterial groups to participate in the conversion of glucose to CO2 and CH4.
• The acid forming bacteria which ferment glucose to produce a mixture
of acetate, propionate and butyrate.
• The acetogenic bacteria convert the propionate and butyrate to acetate.
• The acetoclastic methane bacteria convert acetate to CO2 and CH4.
• The hydrogen utilizing methane bacteria reduce CO2 to CH4.
Mosey found that the hydrogen partial pressure also influences the
acetogenic growth rate, since high values inhibit the generation of propionic
and butyric acids. According to the Mosey model, a sudden increase in the
organic loading rate is expected to cause an accumulation of VFAs since
acetogens grow at a slower rate than the acidogens.

225
Pullammanappallil et al., (1991) Model:
Based on the work of Mosey, followed the model of Pullammanappallil
(Pullammanappallil, Owens et al. 1991) allowed description of the gas
phase and acetoclastic inhibition by undissociated fatty acids.
Flow Chart of (Mosey 1983) and (Pullammanappallil, Owens et al. 1991) models (Lyberatos and Skiadas 1999)
H2 + CO2 Acetate
Propionate Butyrate
Glucose
Acid forming bacteria
Propionic acid and
Butyric acid bacteria
Acetoclastic methane
bacteria
H2 utilizing methane
bacteria
CH4 + CO2 CH4 + H2O

226
Costello et al., (1991a, 1991b) Model:
Costello (Costello, Greenfield et al. 1991a), (Costello, Greenfield et al.
1991b) assumed that the glucose is first converted into acetic acid, butyric
and lactic acids, followed by conversion of lactate into propionate and
acetate by another bacterial group.
The anaerobic ecosystem model developed by (Costello, Greenfield et al. 1991a) (Lyberatos and Skiadas 1999)
CH4
CH4, CO2
H2, CO2
H2, CO2
H2, CO2
CH4, CO2, H2
Glucose
Acid forming bacteria
acetate butyrate lactate
H2, CO2
H2, CO2
LIQUID PHASE GAS PHASE
Glucose LIQUID PHASE
butyric
bacteria
propionic
bacteria
aceticlastic bacteria H2 utilizing bacteria
Acid forming bacteria
Lactic acid bacteria

227
Complex models assuming inhibition of compounds
Angelidaki’s Model (1993):
The model of Angelidaki (Angelidaki, Ellegaard et al. 1993) considers
hydrolysis, acidogenesis, acetogenesis and methanogenesis as the key stages
in anaerobic digestion. This model adequately describes the steps in the
anaerobic digestion of manure. Digesters fed with manure exhibit a self
regulation of pH, attributed to the generated ammonia. Free ammonia is
assumed to inhibit methanogenesis, acetic acid is assumed to inhibit
acetogenesis, and total VFA is assumed to inhibit acidogenesis. The
maximum specific growth rate of the bacteria and the degree of ionization
of ammonia are assumed to depend on the pH and the temperature.
Whenever free ammonia inhibits methanogenesis, acetic acid is
accumulated. This causes an inhibition to acetogenesis, and a consequent
accumulation of propionic and butyric acids leading to inhibition of
acidification. The model describes the behaviour of manure fed digesters
very well. VFA accumulation reduces the pH, causing a decrease in the free
ammonia concentration and the inhibition of methanogenesis. The process is
self regulatory unless the magnitude of the disturbance is larger than the
system can withstand. After that stage, the pH drops and the digester fails to
operate effectively.
Insoluble Carbohydrates
Soluble Carbohydrates
Acidogens
Acetogens
Propionate Butyrate
CH4
Acetate

228
Flow Chart of (Angelidaki, Ellegaard et al. 1993) (Lyberatos and Skiadas 1999)
Siegriest Model (1993):
Siegriest’s model (Siegrist, Renggli et al. 1993) is a more complicated
model that takes into account ammonia inhibition, lysis and hydrolysis of
cell biomass, description of a physical chemical system of pH level,
including the main buffer systems.
Flow Chart of (Siegrist, Renggli et al. 1993) Model (Lyberatos and Skiadas 1999)
acetate
CH4
+
H2O
CH4
+
CO2
Propionate
oxidizing
acetogens
Hydrogen
utilizing
methanogens
Acetoclastic
methanogens
acetate propionate acetate
Fatty acids
oxidizing
acetogens
Acidogens H2
+
CO2
Fatty acids
Biopolymers
Amino acids
and sugars

229
Appendix B Data from Batch Scale Anaerobic Digestion
Experiments
Modelled Total Ammonia Nitrogen (TAN) data at 37 0C
Modelled Soluble Phosphorus (P) data at 37 0C
0.00.10.20.30.40.50.60.70.80.91.0
0 10 20 30 40
Title
TAN 37 -‐ normalised
Substr
Bact
Grwth
Death
Cum TAN
Measured
0.00.10.20.30.40.50.60.70.80.91.0
0 5 10 15 20 25 30
Day
P 37 -‐ normalised
Substr
Bact
Grwth
Death
Cum P
Measured

230
Modelled methane data at 37 0C
Modelled Total Ammonia Nitrogen (TAN) data at 45 0C
0.0
0.2
0.4
0.6
0.8
1.0
0 5 10 15 20 25 30
Day
Methane 37 -‐ normalised
Substr
Bact
Grwth
Death
Cum CH4
Measured
0.00.10.20.30.40.50.60.70.80.91.0
0 10 20 30 40
Day
TAN 45 -‐ normalised
Substr
Bact
Grwth
Death
Cum TAN
Measured
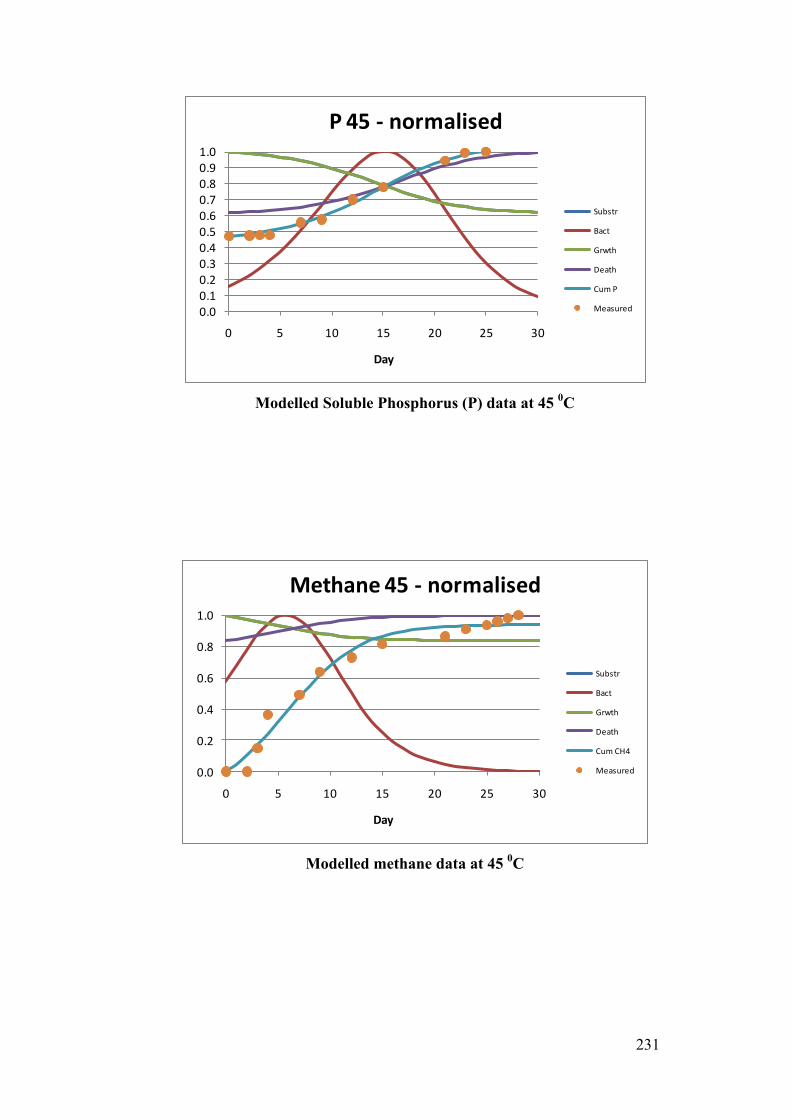
231
Modelled Soluble Phosphorus (P) data at 45 0C
Modelled methane data at 45 0C
0.00.10.20.30.40.50.60.70.80.91.0
0 5 10 15 20 25 30
Day
P 45 -‐ normalised
Substr
Bact
Grwth
Death
Cum P
Measured
0.0
0.2
0.4
0.6
0.8
1.0
0 5 10 15 20 25 30
Day
Methane 45 -‐ normalised
Substr
Bact
Grwth
Death
Cum CH4
Measured

232
Sensitivity Analysis conducted on the methane model at 37 0C, 45 0C &
55 0C
55 CSubstr Bact SubstUse U D SustrRet Const r2 7 14 21 28 r2 7 14 21 281.10 0.0000134 0.00105 0.745 0.700 0.00191 405 0.9889 48.8 116.5 144.1 148.50.99 -0.2160 28.2 48.6 57.7 60.8 -121.8% -42.1% -58.3% -60.0% -59.1%1.21 -0.9538 80.1 227.8 250.9 251.5 -196.4% 64.3% 95.6% 74.2% 69.3%
0.0000121 0.9858 44.5 110.8 141.3 146.8 -0.3% -8.7% -4.9% -1.9% -1.2%0.0000147 0.9860 52.9 121.6 146.5 150.2 -0.3% 8.6% 4.4% 1.7% 1.1%
0.00095 0.9397 47.9 107.1 126.7 129.3 -5.0% -1.8% -8.0% -12.1% -13.0%0.00116 0.9143 49.5 126.3 164.7 172.2 -7.5% 1.6% 8.4% 14.3% 15.9%
0.671 0.5459 34.9 71.8 90.8 96.6 -44.8% -28.5% -38.3% -37.0% -35.0%0.820 0.4000 67.6 177.1 203.2 204.7 -59.5% 38.6% 52.0% 41.0% 37.9%
0.630 0.8116 58.3 151.2 176.8 178.7 -17.9% 19.5% 29.8% 22.7% 20.3%0.770 0.8051 40.7 86.5 109.1 115.2 -18.6% -16.5% -25.7% -24.3% -22.4%
0.00172 0.9763 49.1 120.5 152.5 158.2 -1.3% 0.7% 3.4% 5.8% 6.5%0.00210 0.9827 48.5 113.4 137.9 141.5 -0.6% -0.5% -2.7% -4.3% -4.7%
365 0.9549 43.9 105.0 129.9 133.8 -3.4% -9.9% -9.9% -9.9% -9.9%446 0.9516 53.7 128.3 158.7 163.6 -3.8% 10.1% 10.1% 10.1% 10.1%
Maximum 0.9889
Variables Used From Model Percentage Increase
Cumulative Methane - Day Day
45 CSubstr Bact SubstUse U D SustrRet Const r2 7 14 21 28 r2 7 14 21 281.09 0.0000169 0.001 0.748 0.701 0.00195 410 0.9643 57.7 117.8 134.4 136.50.98 -100.0% -100.0% -100.0% -100.0% -100.0%1.20 -100.0% -100.0% -100.0% -100.0% -100.0%
0.0000152 -100.0% -100.0% -100.0% -100.0% -100.0%0.0000186 -100.0% -100.0% -100.0% -100.0% -100.0%
0.00090 -100.0% -100.0% -100.0% -100.0% -100.0%0.00110 -100.0% -100.0% -100.0% -100.0% -100.0%
0.673 -100.0% -100.0% -100.0% -100.0% -100.0%0.823 -100.0% -100.0% -100.0% -100.0% -100.0%
0.631 -100.0% -100.0% -100.0% -100.0% -100.0%0.771 -100.0% -100.0% -100.0% -100.0% -100.0%
0.001755 -100.0% -100.0% -100.0% -100.0% -100.0%0.002145 -100.0% -100.0% -100.0% -100.0% -100.0%
369451
Maximum 0.9643
Cumulative Methane - Day Day
Variables Used From Model Percentage Increase
37CSubstr Bact SubstUse U D SustrRet Const r2 7 14 21 28 r2 7 14 21 281.12 0.0000117 0.00106 0.745 0.702 0.0019 408 0.9904 47.6 127.1 162.7 167.91.01 -0.0310 27.7 52.5 65.8 70.8 -103.1% -41.8% -58.7% -59.6% -57.8%1.23 -0.5358 79.2 247.1 272.5 273.0 -154.1% 66.3% 94.4% 67.5% 62.6%
0.0000105 0.9873 43.2 120.5 159.9 166.4 -0.3% -9.3% -5.2% -1.7% -0.9%0.0000129 0.9882 52.0 133.0 165.1 169.4 -0.2% 9.2% 4.7% 1.5% 0.9%
0.00095 0.9303 47.6 116.8 140.2 142.8 -6.1% 0.0% -8.1% -13.8% -14.9%0.00117 0.9149 49.2 139.1 186.3 194.4 -7.6% 3.3% 9.4% 14.5% 15.8%
0.671 0.6013 33.8 77.0 102.5 110.5 -39.3% -29.0% -39.4% -37.0% -34.2%0.820 0.5327 66.5 195.2 227.0 228.5 -46.2% 39.5% 53.6% 39.5% 36.1%
0.632 0.8702 56.9 164.8 196.5 198.5 -12.1% 19.4% 29.7% 20.8% 18.2%0.772 0.8381 39.8 94.3 124.9 133.1 -15.4% -16.4% -25.8% -23.3% -20.8%
0.00171 0.9821 47.9 131.5 172.7 179.4 -0.8% 0.6% 3.5% 6.1% 6.8%0.00209 0.9821 47.4 123.7 155.3 159.6 -0.8% -0.5% -2.7% -4.5% -5.0%
367 0.9558 42.9 114.3 146.3 151.0 -3.5% -10.0% -10.0% -10.0% -10.0%449 0.9644 52.4 139.9 179.0 184.8 -2.6% 10.0% 10.0% 10.0% 10.0%
Maximum 0.9904
Variables Used From Model Percentage Increase
Cumulative Methane - Day Day

233
Appendix C DYRESM CAEDYM INPUT FILES
A.1 DYRESM CONFIGURATION
A.2 DYRESM PARAMETER

234
A.3 PHYSICAL DATA AND LAKE MORPHOMETRY
A.4 INITIAL PROFILE

235
Appendix D Nutrient Data in the IBS ponds
NH4 concentration in algal pond 1
PO4 concentration in algal pond 1
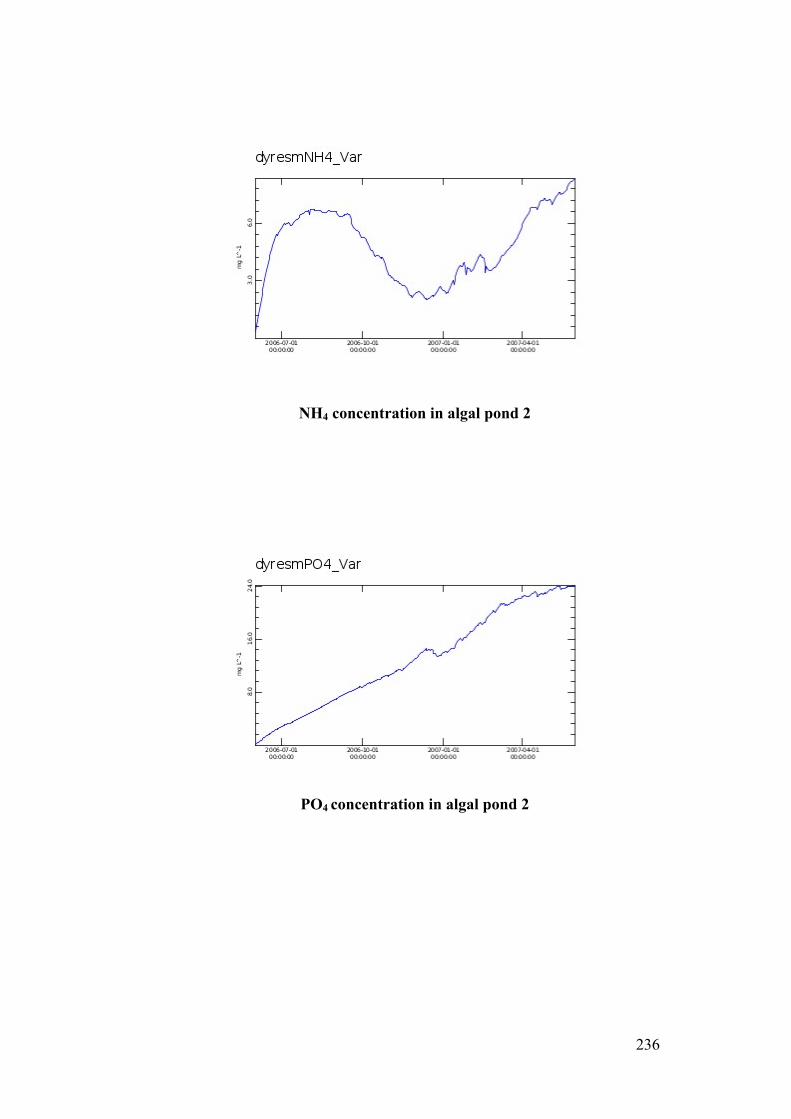
236
NH4 concentration in algal pond 2
PO4 concentration in algal pond 2

237
NH4 concentration in zooplankton pond
PO4 concentration in zooplankton pond

238
NH4 concentration in fish pond
PO4 concentration in fish pond

239
Profile / Contour Plots:
Temperature
Simulated temperature profile in algal ponds 1 & 2
Chlorophyte:
Simulated chlorophyte growth profile in algal pond 1

240
Simulated chlorophyte growth profile in algal pond 2
Cyanobacteria:
Simulated cyanobacteria growth profile in algal pond 1

241
Simulated cyanobacteria growth profile in algal pond 2
Freshwater Diatoms:
Simulated freshwater diatoms growth profile in algal pond 1
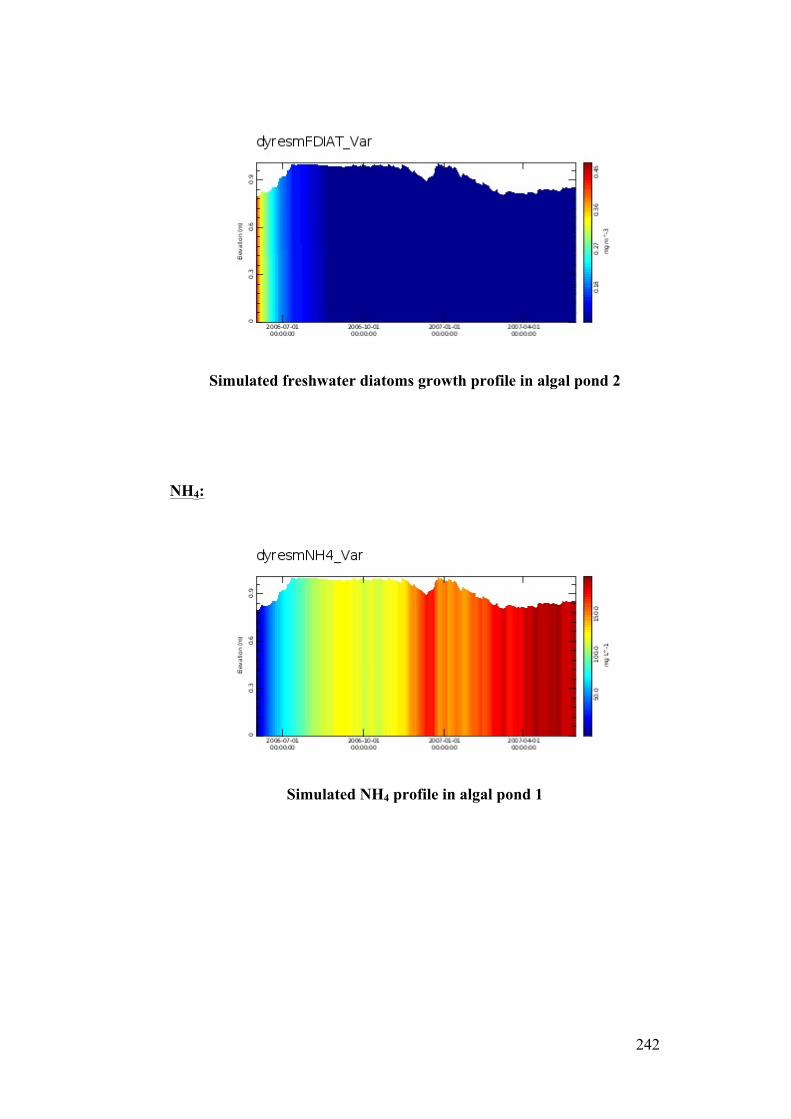
242
Simulated freshwater diatoms growth profile in algal pond 2
NH4:
Simulated NH4 profile in algal pond 1

243
Simulated NH4 profile in algal pond 2
PO4:
Simulated PO4 profile in algal pond 1

244
Simulated PO4 profile in algal pond 2
pH:
Simulated pH profile in algal pond 1

245
Simulated pH profile in algal pond 2
Zooplankton:
Simulated zooplankton growth profile in zooplankton pond

246
Simulated NH4 profile in zooplankton pond
Simulated PO4 profile in zooplankton pond

247
Simulated pH profile in zooplankton pond
Fish:
Simulated fish growth profile in fish pond

248
Simulated NH4 profile in fish pond
Simulated PO4 profile in fish pond

249
Simulated pH in fish pond

250
Appendix E Program Code for Auto Calibration of
DYRESM CAEDYM Reformat DYCD.f90
program reformat_dycd
integer::I,J
character(80)::adummy,ndummy
open(unit=123,file="dycd.dat")
open(unit=345,file="dycd.dat.new")
do
read(123,100,end=500)adummy
100 Format(a80)
print *,len(trim(adummy))
do I=1,80
ndummy(I:I)=" "
ndummy(I:I)=adummy(I:I)
end do
write(345,100)ndummy
end do
500 close(123)
end program reformat_dycd

251
Datamod.f90
subroutine datamod(x,idec,irec,ioffset)
implicit none
integer::irec,idec,ioffset,I
real,INTENT(IN)::x
character::adummy(81)
character::Out(9)
open(unit=123,file="dycd.dat",ACCESS='DIRECT',RECL=81,FORM='FO
RMATTED')
call num_ascii(x,idec,Out)
read(123,200,REC=irec)adummy
200 format(81a1)
Do I=1,9
adummy(I+ioffset:I+ioffset)=Out(I:I)
End do
write(123,200,REC=irec)adummy
999 close(123)
end subroutine datamod

252
Program Driver.f90
Program driver
Implicit none
Real:: value,val_increment,rand
Integer :: i,idec,ioffset,irec,J
do I = 1, 1000
Do J = 1, 11
if(J.eq.1) Then
idec=2
ioffset=5
irec=15
value=0.25 + rand()*(1.0-0.25)
write(*,*)"Pmax ",value
call datamod(value,idec,irec,ioffset)
Endif
if(J.eq.2) Then
idec=0
ioffset=4
irec=41
value=20 + rand()*(100-20)
write(*,*)"IK ",value
call datamod(value,idec,irec,ioffset)
Endif
if(J.eq.3) Then
idec=4
ioffset=5
irec=67
value=0.002 + rand()*(0.007-0.002)

253
write(*,*)"KP ",value
call datamod(value,idec,irec,ioffset)
Endif
if(J.eq.4) Then
idec=4
ioffset=5
irec=83
value=0.0 + rand()*(0.005-0.0)
write(*,*)"KN ",value
call datamod(value,idec,irec,ioffset)
Endif
if(J.eq.5) Then
idec=4
ioffset=5
irec=131
value=2 + rand()*(3-2)
write(*,*)"INmin ",value
call datamod(value,idec,irec,ioffset)
Endif
if(J.eq.6) Then
idec=4
ioffset=5
irec=139
value=3 + rand()*(5-3)
write(*,*)"INmax ",value
call datamod(value,idec,irec,ioffset)
Endif
if(J.eq.7) Then
idec=2
ioffset=4

254
irec=147
value=0.25 + rand()*(1-0.25)
write(*,*)"UNmax ",value
call datamod(value,idec,irec,ioffset)
Endif
if(J.eq.8) Then
idec=4
ioffset=5
irec=155
value=0.05 + rand()*(0.15-0.05)
write(*,*)"IPmin ",value
call datamod(value,idec,irec,ioffset)
Endif
if(J.eq.9) Then
idec=4
ioffset=5
irec=163
value=0.3 + rand()*(0.9-0.3)
write(*,*)"IPmax ",value
call datamod(value,idec,irec,ioffset)
Endif
if(J.eq.10) Then
idec=2
ioffset=4
irec=171
value=0.05 + rand()*(0.15-0.05)
write(*,*)"UPmax ",value
call datamod(value,idec,irec,ioffset)
Endif

255
if(J.eq.11) Then
idec=4
ioffset=5
irec=271
value=0.06 + rand()*(0.08-0.06)
write(*,*)"kr ",value
call datamod(value,idec,irec,ioffset)
Endif
End do
call system("master_run")
call glue_sub()
End do
End

256
Subroutine num_ascii.f90
Subroutine num_ascii(x,idec,Out)
Implicit None
Integer::I,J,K,Irange,Idec,Ipos,Inum,Istart,Ineg
Real,intent(in) ::x
Real ::xx,y
Character::Out(9)
Irange=10000000
xx=x
Do I = 1,9
Out(I:I)=" "
End do
K=1
Inum=9
Istart = 0
Ineg=0
If (xx .lt. 0.0)then
Ineg = 1
xx=Abs(xx)
Endif
Ipos=10**(Idec)
xx=xx*10**Idec
Do I=1,Inum
J = 0

257
y = xx - INT(xx/Irange)*Irange
J = (xx - y)/Irange
If (J .gt. 0 .and. Istart .eq. 0)then
Istart=1
If(Ineg .eq. 1) then
Out(K:K)="-"
K = K + 1
Endif
Endif
If(Istart .eq. 1)then
xx = xx - J*Irange
Out(K:K)=Char(J+48)
K = K + 1
Endif
If(Ipos .eq. Irange)Then
If(Ineg .eq. 1 .and. Istart .eq. 0) then
Out(K:K)="-"
K = K + 1
Endif
Out(K:K)="."
Istart = 1
K = K + 1
Endif
Irange=Irange/10
End do
Return
End

258
Subroutine Glue_sub.f90
Subroutine glue_sub
Implicit none
integer::i,j,p,q,u,v
real(8)::x,y,errcl,sumerrcl=0.0,avgcly,totalcl=0.0,sumvarcl=0.0,NScl
real(8)::dummy1,dummy2,dummy3
real(8):: varcl
totalcl=0
!This part of the program is for chlorophyte calibration
open(unit=123,file="fldcyano.txt")
do i=1,364
read(123,*),y
totalcl=totalcl+y
!print*,y
end do
close(123)
avgcly=totalcl/364
sumerrcl=0
sumvarcl=0
open(unit=222,file="Files/CYANO.ITS")
open(unit=333,file="fldcyano.txt")
do j=1,364
read(222,*),dummy1,x
read(333,*),y
errcl=(x-y)**2
sumerrcl=sumerrcl+errcl
varcl=(y-avgcly)**2
sumvarcl=sumvarcl+varcl
!print*,x,y

259
end do
NScl=1-(sumerrcl/sumvarcl)
write(*,345)NScl
345 format(f10.6," ")
close(222)
close(333)
return
end subroutine glue_sub

260
Appendix F GLUE Calibration Numerical Outputs
Pmax 0.3000495
IK 162.7549
KP 1.6013525E-02
KN 1.8916111E-02
INmin 2.967956
INmax 6.896898
UNmax 1.514976
IPmin 0.3194025
IPmax 2.262906
UPmax 0.4230537
kr 8.1790954E-02
-1.138279
Pmax 1.532858
IK 237.3345
KP 1.8095667E-02
KN 1.5919188E-02
INmin 2.511713
INmax 13.76634
UNmax 1.995085
IPmin 0.4178635
IPmax 2.966611
UPmax 0.2891307
kr 8.8521019E-02
-0.222198
Pmax 2.278895
IK 247.9498
KP 1.9015342E-02
KN 1.9615330E-02
INmin 2.164713
INmax 13.57987

261
UNmax 1.906845
IPmin 0.2882079
IPmax 2.936244
UPmax 0.3243934
kr 8.6169131E-02
-2.921868
Pmax 1.432764
IK 209.3145
KP 1.7897844E-02
KN 1.6891414E-02
INmin 2.544273
INmax 10.92407
UNmax 1.093630
IPmin 0.3828454
IPmax 2.432260
UPmax 0.3798648
kr 9.6898548E-02
0.403511
Pmax 1.383240
IK 265.9270
KP 1.0716619E-02
KN 1.1918589E-02
INmin 2.223608
INmax 12.80367
UNmax 1.083967
IPmin 0.2544030
IPmax 2.616697
UPmax 0.3737425
kr 9.4786234E-02
0.733218

262
Pmax 0.9910892
IK 215.8265
KP 1.4119846E-02
KN 1.9088892E-02
INmin 2.992306
INmax 10.77058
UNmax 1.163922
IPmin 0.4095565
IPmax 2.334848
UPmax 0.3367347
kr 8.1524365E-02
-0.795741
Pmax 0.5833541
IK 176.1786
KP 1.4417774E-02
KN 1.0372999E-02
INmin 2.504163
INmax 9.674148
UNmax 1.579016
IPmin 0.4022338
IPmax 2.093411
UPmax 0.2200110
kr 9.6654058E-02
-1.099454
Pmax 1.157521
IK 178.6295
KP 1.1653987E-02
KN 1.9585462E-02
INmin 2.990804
INmax 6.135645
UNmax 1.870639
IPmin 0.3866753

263
IPmax 2.672653
UPmax 0.3842137
kr 9.1754116E-02
0.815829
Pmax 0.6319873
IK 188.2522
KP 1.7989391E-02
KN 1.5930453E-02
INmin 2.896652
INmax 7.717200
UNmax 1.197950
IPmin 0.2811444
IPmax 2.396368
UPmax 0.2287791
kr 9.2961669E-02
-1.080720
Pmax 1.691624
IK 168.7633
KP 1.1512727E-02
KN 1.0888514E-02
INmin 2.948863
INmax 10.40636
UNmax 1.042604
IPmin 0.3649918
IPmax 2.735089
UPmax 0.3453635
kr 8.9309938E-02
0.525529

264







