
2752 Bromatologia EQ4 1

1. GENERALIDADES
1.1 Flujos de materia y energía
1.2 Reservas energéticas
1.3 Combustibles circulantes
1.4 Gasto energético
1.5 Mecanismos de control
1.6 Regulación del peso corporal
2. BASES DEL METABOLISMO ENERGÉTICO
2.1 Balance energético
2.2 Tasa metabólica
2.3 Métodos de medición del metabolismo energético
2.4 Combustibles exógenos, reservas endógenas y su utilización
2.5 El ATP, la moneda energética de cambio
2.6 Calorimetría indirecta
2.7 Oxígeno y producción de ATP
2.8 Conclusiones
3. FLUJOS POSABSORTIVOS
3.1 Consumo de oxígeno
3.2 Medición de la tasa de recambio
3.3 Producción de glucosa
3.4 Utilización de la glucosa
3.5 Aminoácidos
3.6 Producción de urea
3.7 Lípidos
3.8 Algunos efectos hormonales
3.9 Conclusiones

4. FLUJOS EN AYUNO
4.1 Aspectos generales
4.2 Flujos de combustibles en ayuno de 3 a 4 días
4.3 Flujos de combustibles en ayuno prolongado
4.4 Control de flujos en ayuno
4.5 Algunos efectos hormonales
4.6 Conclusiones
5. APORTE EXÓGENO DE COMBUSTIBLES
5.1 Vaciamiento gástrico
5.2 Termogénesis prandial o Costo energético de la ingestión (CEI)
5.3 Entrada exógena de carbohidratos
5.4 Entrada exógena de lípidos
5.5 Entrada exógena de aminoácidos
5.6 Algunos efectos hormonales
5.7 Conclusiones
Conclusiones

1. GENERALIDADES

1.1 Flujos de materia
Cualquier ser vivo puede ser visto como un sistema deestado estable por el cual entra y sale materia singarantizar que la salga sea la misma que entró. Lo mismosucede en aquél organismo que ya no crece, sólo que acorto plazo como cuando se ingiere agua y salesminerales.
En la misma proporción que entra el agua ésta sale, encambio otras sustancias más complejas ricas en energíaquímica sufren transformaciones a sustancias más sencillasde manera que la energía fluye, esto se observa comoenergía contenida y energía gastada.

Entradas Salidas
Carbohidratos (CHO)
Lípidos (LIP)
Proteínas (PRO)
Oxígeno (O2)
MATERIA
Bióxido de Carbono
Agua
Productos nitrogenados
Energía química en PRO,
LIP y CHO
ENERGÍA
Síntesis orgánicas
Contracción muscular
Transporte activo
Ciclos de sustrato
Producción de calor
Entradas y salidas de materia y energía

Las entradas son el contenido cuali y
cuantitativo del alimento ingerido más O2.
La salida es de urea y amonio por lo
general del catabolismo oxidativo.
La tasa del flujo catabólico (valor por
unidad de tiempo) determinada por el
gasto energético que pueden aumentar de
manera considerable como cuando hay
ejercicio o hace frío. La ingesta calórica se
determina cuantitativamente por el gasto
energético previo. Un adulto normal y casi
sedentario ingiere y gasta de 2 000 a 2 500
Kcal al día.

1.2 Reservas energéticas
La diferencia entre los objetos de entrada, es el lugar por
donde lo hacen, como el O2 que lo hace por la nariz y otros
son ingeridos.
Se puede ayunar por días ingiriendo agua y sales minerales
pero no dejar de respirar sin daño cerebral porque el cuerpo
tiene reservas de energía pero no de O2.

Compuesto Ingestión Reservas Oxidación
g Cal Cal g Cal
Carbohidratos 250 1 000 2 000 1 000 50
Lípidos 110 1 000 125 000 1 000 0.8
Proteínas 125 500 40 000 500 1.25
Tabla adaptada de una figura de trabajo de Ravussin y Swinburn (1992)
que considera una dieta diaria con 40% de la energía de carbohidratos,
40% de lípidos y 20% de proteínas
Aquí, la entrada exógena y la utilización calórica de los 3 componentes
están perfectamente balanceadas pero se observa la diferencia entre la
reserva de lípidos y de carbohidratos.

Reservas internas:
Carbohidratos: El glucógeno es un polímero de glucosa para reserva con
agua y que se encuentra en músculo de 30-40% y en hígado en 5-7% de su
peso. Se puede usar después de glucólisis. Los órganos que llevan glucosa a
la sangre son el hígado y la corteza renal.
Lípidos: Su reserva se encuentra distribuida por todo el cuerpo como
grasa subcutánea, perirrenal, gonadal, peritoneal, etc. El peso de la grasa
aumenta en mujeres y con la edad. Cuando se oxidan dan más calorías que
los carbohidratos.
Proteínas: Representan el 30-40% del peso corporal en una persona
normal y por ello su reserva no se encuentra disponible en su totalidad al
comprometer la vida del individuo.

1.3 Combustibles circulantes
Un combustible biológico es una sustancia con cierta energía en sus
enlaces y que se puede liberar por oxidación después de otras
reacciones mediadas por encimas. Otros metabolitos al no ser
oxidados sirve en la síntesis de otros en las vías metabólicas como el
ATP que no se almacena.
Combustibles circulantes:
a) Glucídicos: glucosa, lactato, piruvato.
b) Lipídicos: ác. Grasos libres, glicerol, cuerpos cetónicos.
c) Protídicos: aminoácidos.

a) Manejo de complejos glucídicos: La glucosa es la única que
varía poco en condiciones fisiológicas, así la glucemia la
regula el sistema de control del cuerpo, La glucosa se
libera en orina cuando rebasa los 200 mg/dL de sangre.
De manera exógena la glucosa viene de carbohidratos y
entra por intestino, las otras hexosas tienen vida circulante
breve y se oxidan o almacenan en glucógeno. La síntesis de
glucosa en sangre se da por gluconeogénesis en hígado o
en corteza renal por circunstancias de emergencia.
Los precursores gluconeogénicos son el lactato, piruvato,
aminoácidos y el glicerol.

El uso de la glucosa depende de su captación por las células
en su paso por la membrana gastando energía y se da un
transporte pasivo o facilitado por la insulina, en tal caso el
tejido muscular y adiposo dependen de la insulina. El
cerebro humano en reposo usa 120 g de celulosa/día en
forma constante y los músculos de 30-120 sólo cuando hay
actividad motora. Cuando no se puede contar sólo con la
glucosa se oxidan lípidos.
La glucosa entra en la célula, se fosforila en posición 6 y
entra en la síntesis de glucógeno o la glucólisis donde se
producen 2 moléculas de piruvato en citoplasma y 2 ATP
por molécula de glucosa.

La glucólisis anaerobia es fuente de energía de varios tejidos
como las células sanguíneas, piel, médula renal, córnea,
cristalino, etc. Y como producto final de este proceso esta el
lactato. En los tejidos donde se lleva a cabo la glucólisis aerobia el
piruvato puede formar acetil-CoA para el ciclo de los CAT para
la síntesis de 36 ATP que en cuanto al alimento consumido sirve
como precursor en la síntesis de ácidos grasos, lipogénesis. El
lactato puede oxidarse a piruvato para ser combustible de las
mitocondrias.

b) Manejo de combustibles lipídicos: La mayoría de los lípidos
pasa del alimento a la sangre como quilomicrones que se
sintetizan en el epitelio intestinal, la mayoría son triglicéridos
que después de hidrolizarse en capilares del tejido adiposo
vuelve a sintetizarse en las células adiposas, además libera 1
molécula de glicerol y 3 de ácidos grasos libres (AGL) que
pasan a la sangre unidas a la albúmina para actuar como fuente
de energía de diversos tejidos. En el caso de los músculos,
además de la glucososa necesitan otro combustible como la
grasa entre sus fibras.

La oxidación de una molécua de ácido palmítico rindepara la síntesis de 129 moléculas de ATP, por ello lamayoría de los tejidos usan a los lípidos en lipólisiscomo fuente de energía, excepto los que dependen dela glucólisis anaerobia y el sistema nervioso. El hígadocapta los AGL canalizándolos para síntesis de TG y la boxidación mitocondrial donde se produce el AcCoAque pasará a CAT asegurando las necesidadesenergéticas del órgano, a partir de 2 moléculas deAcCoA se pueden producir cuerpos cetónicos que sonutilizados por el cerebro para ahorrar energía duranteel ayuno.

c) Manejo del combustible protéico: Los aminoácidos obtenidos
de la digestión total de las proteínas ingeridas, son absorbidos y
como una parte de las proteínas endógenas (enzimas digestivas,
de secreción…) se digieren, se reabsorben de 50-70 g de
aminoácidos/día. Parte de los aminoácidos re sintetizan
proteínas y parte va a sangre, éste recambio se refleja en el
gasto energético en reposo que disminuye durante el ayuno
puede impedir la acumulación de proteínas y péptidos
peligrosos para la salud permitiendo la concentración de
enzimas reguladoras del flujo en las vías metabólicas. Este
recambio se ve afectado por la intervención de hormonas.

1.4 Gasto energético
De los 3 combustibles, en común está la síntesis de ATP que se
puede dar en citoplasma o por acoplamiento entre oxidación y
fosforilación del ADP en mitocondrias. El gasto energético es
inversamente proporcional al peso total y que depende de la
temperatura ambiente, actividad, alimentación y factores
genéticos. Cuando los 3 combustibles se oxidan necesitan O2
inspirado y liberan CO2, a esto se le conoce como cociente
respiratorio y no vale lo mismo para los 3 combustibles.

Por calorimetría indirecta se puede medir los gases y que
compuestos han sido usados en un lapso de tiempo y aumenta
por un mayor uso de glucosa y glucógeno. Tal gasto energético se
determina considerando el metabolismo total como el 100% en
vida sedentaria:
Metabolismo de reposo= metabolismo basal + costo de
termorregulación (70%).
Metabolismo de actividad (20%).
Termogénesis inducida por dieta o costo energético de la
ingestión o acción dinámica específica del alimento (10%).

1.5 Mecanismos de control
El gasto de energía necesita ajustes metabólicos de del flujo
óptimo de los combustibles:
a) Control de gasto energético: Aunque 2 personas coman lo
mismo puede que una adelgace y otra no por factores
metabólicos genéticos. Hay un control neuroendócrino de éste
gasto, el sistema simpatoadrenal y las hormonas tiroideas hacen
ineficiente la producción de energía útil produciendo más calor
(termogénesis). La triyodotironina plasmática disminuye en el
ayuno.

a) Controles en los flujos de combustibles: La secreción de
insulina es controlada por l sistema nervioso autónomo por
hormonas y combustibles circulantes debido a glucemia y la
respuesta a la glucosa. La insulina actúa en la reserva de glucosa
y la inhibición de el uso total de glucosa o grasas. Así la
hipoglucemia se trata con hormonas contrarreguadoras como
adrenalina y glucagon. De manera que aumenta la producción
hepática y renal de glucosa.

1.6 Regulación del peso corporal
Se puede dar al regular la ingesta del alimento al no ingerir
más de lo que se gasta pues las calorías sobrantes se depositan
como grasa, por ello se usa el índice metabólico corporal:
peso en Kg/estatura en m2 = 20-27 como normal que varían
con los factores genéticos. Se ha encontrado que la leptina
inhibe la ingesta y estimula el metabolismo. Incluso se ha
asociado a la obesidad con las actividades termogénica y
lipolítica des sistema simpático.

GENERALIDADES
Flujos de materia y energía
Entrada y salida de materia y
estabilidad.
Reservas energéticas
Origen y agrupación
endo y exógena: LIP, CHO y PRO.
Combustibles circulantes
Producidos en tejidos y que
viajan en sangre: glucosa,
aminoácidos y ácidos libres.
Gasto energético
Relación molar de
O2 adquirido y
CO2 eliminado
con relación a síntesis de
ATP.
Mecanismos de control
Tipos de control
energético como gasto y
flujo.
Regulación del peso corporal
Ingesta regulada de alimento
valioso o chatarra y
acumulación de TM.

2. BASES DEL METABOLISMO
ENERGÉTICO

2.1 Balance energético
Es la relación entre la cantidad de energía obtenida por
ingestión, la absorción de CHO, LIP y PRO en cierto tiempo,
y la energía gastada. Este dato se calcula con la cantidad de
energía de entrada y el cambio de peso, así el aumento de
peso indica mayor entrada que salida y la energía en su
mayoría se adquiere de carbohidratos. Tomando en cuenta los
3 combustibles.

2.2 Tasa metabólica
Representado por el sistema entrada (exógena o endógena) –
salida (gasto) y deben estar balanceados a corto plazo para que el
organismo permanezca estable. Medido como tasa metabólica
(TM)en gasto por unidad de tiempo es variable. Westerterp lo ve
como:
TM de sueño.
TM del despertar.
Costo energético de ingestión.
Costo energético de la actividad.
TM basal

Los cambios entre TM y TMB se dan por fluctuaciones del
CEA. Un factor de variación de TM es la T ambiente, pues
cuando baja se produce más calor y con más uso de energía. Para
medirlo se usa laTM Std. que está por debajo de:
laT ambiente
En reposo muscula y en estado de alerta (despierto)
En condiciones postabsorbativas (por la mañana)

Según Rolfe y Brown en 1997 Los procesos en TMB se dan así:
Síntesis de proteínas 25 a 30 %
ATPasa Na+/K+ 19 a 28 %
ATPasa Ca2+ 4 a 8 %
ATPasa de la actomiosina 2 a 8 %
Gluconeogénesis 3 %
Otras (síntesis de RNA y ADN, ciclos de
sustrato, escape de protones)
No definido

También influye el peso del individuo de forma inversamente
proporcional. En 1901 Voit dijo que el metabolismo en
condiciones de ayuno era proporcional a la superficie corporal.
Otro factor que influye es el porcentaje de grasa que varía según
el sexo y la edad debido a que el metabolismo de las células
adiposas es menor que el de otras por tener menor cantidad de
citoplasma, incluso se puede hacer una medición en la que no
haya grasa.

2.3 Métodos de medición del metabolismo energético
Se usa el de calorimetría directa o indirecta. La directa mide
pérdida de calor total en un espacio aislado dando el calor que se
produce en el cuerpo en un tiempo determinado por lo que se
debe tener en cuenta el calor almacenado y no es un método de
corto plazo.
La indirecta es más empleado y mide el intercambio de gases
durante la respiración ya que los CHO y LIP liberan CO2 en la
combustión, aunque las PRO además liberan N2 por la orina.

2.4 Combustibles exógenos, reservas y su utilización
De los 3 combustibles, no podemos digerir y absorberlos en
todas sus presentaciones en el tubo digestivo, sólo el 93-96%. La
energía metabolizable = combustibleo absorbido – energía
perdida en orina, de forma tal que se pierde 3-4% de las calorías
ingeridas en la orina diario. Atwater propuso que a partir de la
energía metabolizable se pueden usar factores como 4.9 y 4
Cal/g de CHO, LIP y PRO digeribles.

2.5 El ATP, la moneda de intercambio energético
La energía puede contenerse en moléculas cualquiera que
después reaccionarán con otras, la energía se desprenderá en la
síntesis de ATP de ADP + Pi endergónicamente lo contrario a la
hidrólisis.
ADP + Pi + H2O ATP + H+
En esta reacción como en otras, se pierde energía útil. Si 1 mol
de glucosa rinde a 688 Cal y la formación de 1 ATP 7.4 Cal, la
eficiencia de la reacción es de 38.2%.

Es como se considera que la relación entre el O2 consumido y el
ATP que se formó es de 3, y el almacenaje de glucógeno es de
2.1 ATP por cada residuo de glucosa, en cambio la lipogénesis
consume 11 ATP por c/molécula de tripalmitina sintetizada. La
gluconeogénesis a partir de lactato usa 6 ATP para c/molécula de
glucosa, así el costo total de la gluconeogénesis es de 4 ATP.
Aún así la síntesis de proteínas cuesta más de 4-6 ATP por
c/enlace peptídico con eficiencia del 10%

El recambio de proteínas endógenas consume el 20% de la TMB
hasta 3 veces más que las ingeridas y su gasto energético es de 1
Cal/g aproximadamente y se da en un órgano específico. Donde
surgen más problemas para calcular este gasto es en el ciclo de
los sustratos por el tipo de reacción que presenta, ya que en un
sentido ni siquiera se produce ATP.
Para el caso de la TMB, las bombas, que transportan iones
consumen el 17% de lo que usa el músculo esquelético, más el
35% en hígado sin contar el 80-85% que se consume cuando se
reabsorbe el Na renalmente.

2.6 Calorimetría
Para determinar la cantidad de calorías producidas, gastadas por
alguien y el papel de los 3 combustibles, se usa la calorimetría
indirecta.
A continuación se observa en la tabla el O2 consumido y la
producción de CO2 que se elimina según los 3 combustibles de
los que hemos hablado. Su tasa de intercambio lo denotamos
comoVO2 yVCO2.

Combustible Cal / L O2 Cal / L CO2 CR
CHO (glucosa) 5.01 5.01 1.00
CHO (glocógeno y
sacarosa)
5.04 5.04 1.00
LIP 4.69 6.63 0.71
PRO 4.48 5.58 0.80
CR = Cociente respiratorio (VCO 2/VO2)
Calorías producidas por litro de oxígeno consumido y litro de dióxido de carbono
producido.
Para el cálculo adecuado y con menos errores de las calorías que se
producen, es mejor mezclar las 3 macromoléculas de los combustibles y
denotar el O2 que interviene. Cuando se toma en intervalos inadecuado
pueda haber cambios de pH que afecten la determinación.

La acidosis se da por exceso de ácido láctico durante el ejercicio
y para compensar esto, el organismo eliminará un exceso de
CO2. Las proteínas oxidadas reflejan a las de entrada y se mide su
tasa de oxidación con el N2 urinario = 6.25g proteína. Entonces
si queremos medir la TPE según los 3 combustibles con éste
método calorimétrico (Cal/min) se usa:
TPE = 3.91 VO2 +1.10 VCO2 – 1.93 N

Al usar CR como factor en relación a los 3 combustibles existenciertos inconvenientes, los metabolitos que actúan lo hacen enotras vías metabólicas, así en la lipogénesis es:
9C6H12O6 + 8O2 2C12H32O2 + 22CO2 +22H2O CR=2.75
Al oxidarse el mismo palmitato:
2C10H32O2 + 46O2 32CO2 + 32H2O CR=0.696
La suma de las dos ecuaciones:
9C6H12O6 + 54O2 54O2 + 54H2O CR=1.00

O sea, el CR > 1.00 si la lipogénesis rebasa la oxidación de LIP y
< 1.00 si excede la lipogénesis o acelerada.
C6H10O2 + 5O2 4C4H8O3 CR = 0
Así la urea en orina resulta de la oxidación de proteínas y no de la
glucosa.

2.7 Oxígeno y producción de ATP
El pozo total de un adulto es de 84 mmoles y una tasa de
recambio de 1300 mmoles casi igual al peso corporal, de forma
que se hace imposible el almacén de ATP como con la
fosfocreatina de músculos y nervios.
La DGº (energía libre) en la hidrólisis de ATP a ADP + Pi in vitro
= -7.3Cal/mol y bajo condiciones de células de homeotermo
hasta -12.5 Cal / mol.
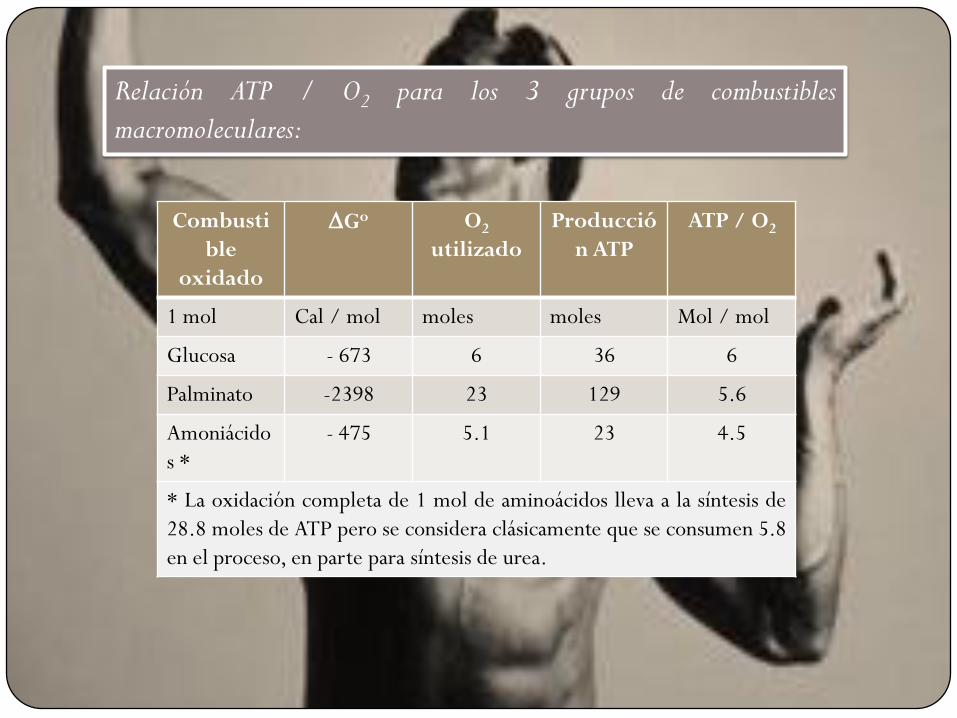
Combusti
ble
oxidado
DGo O2
utilizado
Producció
n ATP
ATP / O2
1 mol Cal / mol moles moles Mol / mol
Glucosa - 673 6 36 6
Palminato -2398 23 129 5.6
Amoniácido
s *
- 475 5.1 23 4.5
* La oxidación completa de 1 mol de aminoácidos lleva a la síntesis de
28.8 moles de ATP pero se considera clásicamente que se consumen 5.8
en el proceso, en parte para síntesis de urea.
Relación ATP / O2 para los 3 grupos de combustibles
macromoleculares:

De tomar como válido el dato de -12.5 Cal / mol la eficiencia de
cada grupo para conservar la energía es de 67%. El ATP / O2 =
P/O depende de que la relación NADH + H+ + 1/2O2
NAD + H2O se acople a la síntesis de 3 ATP que FADH2 +
1/2O2 FAD + H2O a la de 2ATP.
Entonces la eficiencia de la síntesis de ATP depende del gradiente
electroquímico de protones que atraviesen la membrana que
disminuye según las células de la apertura.

Así el metabolismo se puede dividir en reacciones de
acoplamiento y desacoplamiento:
Oxidación de sustratos reducción de NAD a NADH
Oxidación de NADH en la cadena de transporte electrónico
producción del gradiente protónico por la membrana interna.
Paso de protones al interior de la mitocondria por sintasa del
ATP síntesis de ATP.
Síntesis de proteínas, transportes activos, contracción muscular
hidrólisis de ATP.

De desacoplamiento:
Degradación de proteínas.
Paso pasivo de iones y protones a favor de gradientes electroquímicos.
Relajación muscular.
No todo el O2 es para síntesis de ATP. En muchos órganos, las
mitocondrias presentan un escape pasivo de protones desacoplando la
oxidación de la fosforilación, el escape es casi el 20% de TMB y por
ello se relaciona con un P/O mecánico máximo sin considerar el
escape y otras reacciones desacoplantes. Dando P/O=2.5 en
mitocondrias que respiran con NADH y de 1.5 para las que lo hacen
con succinato, el P/O efectivo es de 1.8-2.0 si solo se toma el
consumo de O2.

Estimación de la proporción de la disipación de energía libre en
la molécula de glucosa:
Glucólisis 3%
Oxidación de sustratos 10%
Cadena respiratoria 10% escape de protones 15%
Síntesis y transporte de ATP 4%
ATPasa Na+/K+
Síntesis de proteínas 14%
Resto 33%

BASES DEL METABOLISMO
Balance energético
Relación entre
cantidad de energía obtenida y la gastada.
Tasa metabólica
“Caja negra”
de sistema entrada-salida de energía
Métodos de
medición del
metabo-lismo
energé-tico
Métodos para
determinar calor total y
gastado.
Combustibles exógenos, reservas
endógenas y su utilización
Fuentes de
energía: LIP; PRO y CHO.
Sus reservas no son iguales.
El ATP, moneda de energética de cambio
Modo de energía
común en todo
proceso.
Calori-metría
indirecta
Estimación de calorías producidas en relación al tiempo.
O2 y produ-cción
de ATP
Relación entre la
síntesis de ATP y la entrada de O2.

3. Flujos posabsortivos

3. Flujos posabsortivos
Se considero adecuado empezar a analizar los datos
cuantitativos acerca de los flujos de oxigeno y
combustibles en el humano en la condición postabsortiva
por diferentes razones
Es la condición en la cual se encuentra el organismo después de
haberse absorbido desde el intestino a la sangre todos los
componentes energéticos.
Se conforma a los requisitos para estimar los valores de la tasa
metabólica basal
Es la condición basal que permite apreciar los cambios de las
variables en el caso de la aplicación de otras condiciones

A la sangre no llega ningún combustible exógeno. Por lo
tanto, el gasto energético queda enteramente a cargo de las
reservas.

3.1 Consumo de oxigeno
El consumo de oxigeno medido por calorimetría
indirecta refleja mejor la tasa de producción de energía
(TPE) que los valores de VCO2 para estimar en que
medida es suficiente estimar la TPE con base en el VO2
sin tomar en cuenta el VCO2 se trabajo con datos, el cual
demuestra que el calculo de la oxidación de CHO y LIP
basado en dos evaluaciones por calorimetría indirecta, de
30 minutos de cada una, con un intervalo de 15 a 20
minutos.

Lo interesante es que las mujeres obesas presentaron menor
VO2 reportado al peso pero valores mayores al reportarlo al
PSG. Esto muestra la importancia que tiene el porcentaje de
grasa en los cálculos de la tasa metabólica.

Es interesante examinar el VO2 de los diferentes órganos respecto
a su peso relativo en el organismo.
El cerebro, corazón, riñones e hígado suman un 5.5% del peso
corporal y consumen mas del 50% del oxigeno.
Los músculos esqueléticos en reposo tienen un consumo muy bajo
por unidad de masa muscular.
El músculo cardiaco usa mas consumos de oxigeno.

3.2 Medición de la tasa de recambio
Las técnicas comúnmente empleadas para determinar los
flujos de diferentes metabolitos utilizan marcadores
radiactivos.
Los metabolitos que se inyectan o se ingieren pueden estar
marcados en uno o mas de uno de sus carbonos.

También se presenta el problema de las reaccionesintracelulares que recirculan una marca entre variosmetabolitos.
Otro método para medir la suerte de un metabolito através de cierto órgano o grupo es la determinación desu concentración arterial y la de la vena que drena dicharegión.
La diferencia entre las dos concentraciones multiplicadapor el flujo de sangre (o de plasma) regional puededeterminar el flujo neto del metabolito.

3.3 Producción de glucosa
La glucosa es el combustible cuya concentración sanguínea seencuentra mas estrictamente regulada.
En la condición postabsotiva la entrada de glucosa a la sangre sedebe principalmente a la producción hepática mediante dos víasmetabólicas.
La glucogenenólisis
La gluconeogénesis

Entre los a.a los que mas importancia cuantitativa tiene parala glucogeogenesis son la ALA y la GLN. Por transaminaciónla ALA puede dar piruvato y de aquí glucosa.
La participación relativa de los sustratos gluconeogenicoscirculantes en condiciones postasortivos es del orden de 30 a40% para L y piruvato, un 10 a 15 % para GOL y un 50%para AA, de los cuales la ALA y la GLN representan lamitad.
La tasa gluconeogenica será del orden de 4 a 5μmol/Kg.min. para una producción hepática yrenal total de G de 11 a 13 μmol/Kg.min.

3.4 Utilización de la glucosa
Es evidente que mantener una glucemia del orden de 4.7mmolar (85mg/dl) es necesario que la utilización de laG(RdG) tenga el mismo valor que su producción.
La glucosa puede entrar a las células por un transportefacilitado dependiente de insulina o por transporte pasivo.Los principales tejidos dependientes de insulina son elmuscular y el adiposo.

El problema de la utilización no oxidativa de la glucosa en la
glucólisis se asocia con aquel de la producción anaeróbica de ATP
en este proceso. Una serie de tejidos son dependientes mas bien
del ATP sintetizado en el citosol.

3.5 Aminoácidos
Los músculos son los principales proveedores de AA a lasangre. Los únicos AA cuyo flujo neto es desde la sangre a losmúsculos son la serina y la cisteina.
En condiciones de alimentación normal en individuos sanos,la síntesis será cuantitativamente igual a la proteolisis poraporte exógeno de nuevos AA.

La masa muscular contiene mas del 50% del pozo corporal total
de AA y el hígado es el único órgano que tiene las enzimas del
ciclo de la urea.
La producción muscular neta de AA se ha estimado midiendo la
diferencia arteriovenosa a través de una pierna o un antebrazo.

3.6 Producción de urea
El nitrógeno resultante de la desafinación de los AA se elimina,
en la condición postabsortiva, en un 90% bajo forma de urea,
compuesto que se sintetiza únicamente en el hígado.
Las enzimas del ciclo de la urea se encuentran principalmente en
la región periportal del hígado.

En la misma región hepática están también localizadas
mayoritariamente las enzimas de la gluconeogenesis,
implicadas en la síntesis de glucosa a partir de los esqueletos
desaminados de los aminoácidos.
Un aspecto importante en la producción de urea es que un
20% sale a la luz del intestino secretada en la bilis, el jugo
pancreático y por la pared intestinal a todo lo largo del
intestino delgado.

3.7 Lípidos
Los depósitos corporales de lípidos están compuestos
principalmente por triglicéridos y están distribuidos en
diferentes agrupaciones en tejido adiposo.

Su deposito relativo a la masa corporal es mayor en mujeres
que en varones.
Los quilomicrones circulantes resultantes de los lípidos
ingeridos, así como las lipoproteínas de muy densidad
producidas por el hígado, liberan a nivel de ciertos tejidos,
principalmente en los capilares del tejido adiposo y los de
los músculos esqueléticos, ácidos grasos y glicerol, bajo la
acción de la lipoproteinlipasa.

Los tres ácidos grasos liberados de cada molécula de triglicérido
atraviesan el endotelio, penetran en los adipocitos son activados a
acil CoA y se esterifican con una molécula de GOL fosfato a TG
de reserva.

Los TG contenidos en los adipocitos pueden ser hidrolizados
bajo la acción de la lipasa sensible a hormonas intracelular.
Se puede decir que la RaGOL no da cuenta necesariamente
de la tasa lipolítica.
Los TG plasmáticos e intracelulares participan en cierta
medida a la oxidación lipídica total.
Podría haber reesterificación extrahepática importante.

3.8 Algunos efectos hormonales
En la condición postabsortiva no se puede determinar el efecto de
algunas hormonas sin acompañar su administración con otras
perfusiones.
Esto es evidente en el caso de la insulina la cual administrada sola
induce hipoglucemia, por lo que es necesaria la perfusión
simultanea de glucosa.

En estudios se demostró que en condiciones de hipoglucemia
moderada, gluconeogenesis tiene mayor participación en la
glucorregulación, siendo el principal factor para la prevención de
una mayor caída de la glucemia.

3.9 Conclusiones
La medición del VO2 estima con bastante fidelidad, aunqueno exactamente, la TM. Dado el gasto energético muchomayor del PSG que el de la MA, es preferible referirse la TMal PSG, sobre todo cuando se utilizan sujetos conproporciones muy variables de MA.
Los flujos de los combustibles circulantes se estimanmediante el uso de marcadores de ciertos átomos de susmoléculas. Los problemas de la metodología son muchos yespecíficos para cada caso.

Flujos posabsortivos
Consumo
de oxigeno
Lípidos
Medición de
la tasa de
recambio
Producción
de glucosa
Utilización de
la glucosa
Aminoácidos
Producción
de urea
Algunos
efectos
hormonales
dos vías
metabólicas
Los músculos
son los
principales
proveedores
de AA a la
sangre
•utilizan
•marcadores
•radiactivos.
Para el buen
funcionamiento
del organismo
no se puede
determinar el
efecto de
Algunas
hormonas
sin acompañar
su administración
con otras perfusiones
producción
anaeróbica de ATP
El nitrógeno
resultante de
la desafinación
de los AA se
elimina
un 90% bajo
forma de urea
El músculo
cardiaco
usa
mas consumo
de oxigeno
determinar los
flujos
de diferentes
metabolitos
por
glucogenenólisisgluconeogénesis
El nitrógeno resultante
de la desafinación de
los AA se elimina,
en la condición
postabsortiva,
en un 90% bajo
forma de urea
para

4. Flujos en ayuno

4.1 Aspectos generales.
Los datos experimentales de humanos ayunados suscitan una seriede problemas.
La falta del aporte exógeno de combustible hace necesaria lautilización de las reservas endógenas.
Al ahorro de AA necesarios para la gluconeogenesis se puedellevar a cabo al reducir el consumo de glucosa.

4.2 Flujos de combustibles en ayuno de 3 a 4 días
A las 24 horas de ayuno total, el contenido de glucógeno hepáticollega a solo un 14% del inicial o sea a unos 40 μmol deglucosa/g de hígado.
En un ayuno de 3 a 4 días la gluconeogenesis proveea la circulación unos 5 a 8 μmol G/Kg. min. Lacontribución renal aumenta a 20-25% del total.

La excreción de nitrógeno urinario aumenta en un 30 al 40
% a valores del orden de 8mmol/Kg. Min. A diferencia de
la situación postabsortiva, donde un 90% del nitrógeno de
encuentra en la urea, después de 3 días de ayuno la urea
contiene solo un 80% del nitrógeno.
Esto se debe a la mayor tasa de captación de GLN por los
riñones, con mayor liberación de amonio en la orina y de G
por la gluconeogenesis a la sangre.

4.3 Flujos de combustibles en ayuno
prolongado.
Se pudiera pensar que el metabolismo en ayuno prolongado no es
mas que la continuación de lo que ocurre en los primeros días de
ayuno.
La diferencia es sin embargo importante ya que no es posible
asegurar la supervivencia si no se reduce la perdida de proteínas.

En ayuno corto la respuesta metabólica esta dirigida
principalmente a mantener la producción hepática y renal de
glucosa, mientras que la prolongación del ayuno determina un
cambio de estrategia en el sentido de minimizar el catabolismo
proteico.

La disminución del consumo de AA en la gluconeogenesis en
ayuno prolongado se demuestra por la caída en la tasa de
excreción de nitrógeno en la orina.

4.4 control de flujos en ayuno
El ayuno tiene por lo menos dos efectos hormonales importantes:
la reducción en los niveles plasmáticos de la insulina que se da aun
después del ayuno nocturno, y la de la triyodotironina.
La inulina baja después de 2 a 3 días de ayuno de unos 70 a 15-
20pmol/l.

La respuesta de la insulina a la administración de glucosa es
mas lenta y la respuesta hipoglucemia a la administración de
insulina es menor.
Los niveles bajos pero constantes de la glucemia se
mantienen por la intervención de las hormonas mal llamadas
contrarreguladoraas que proporcionan los niveles
sanguíneos de los sustratos para la gluconeogenesis y
estimulan la lipólisis y la cetogénesis.

4.5 algunos efectos hormonales
Las principales diferencias entre el ayuno nocturno y de tres
días son la caída de los niveles de T3 y de la RaG y el
incremento de la proteolisis.
Además hay un efecto en el bloqueo beta adrenérgico sobre
algunas variables metabólicas en ayuno de 12 y 48 horas.

En comparación con la situación postabsortiva en condiciones
de hipoglucemia e hipoinsulinemia por el ayuno de tres días, el
antagonista beta adrenérgico disminuyó tanto la lipólisis como
la participación de los LIP al gasto energético.

4.6 conclusiones En un varón con IMC normal y pesando 70 Kg la MA representa
unos 10 Kg, es decir, unos 8 Kg de triglicéridos de reserva.
Esta reserva aseguraría un gasto energético de 72,000 Cal y, por tanto, cubría teóricamente las necesidades de un metabolismo de reposo durante unos 50 días en falta de aporte exógeno de combustibles.
El verdadero problema del ayuno consiste en asegurar mediante la gluconeogenesis una RaG que garantice el nivel de glucemia compatible con las necesidades de los diferentes órganos y tejidos.
La utilización proporcional de AA en ayuno prolongado no baja mucho y sigue siendo un 7 al 10% de la TM, a pesar de la reducción de la tasa proteolítica.

Flujos en ayuno
Aspectos generalesFlujos de
combustibles
en ayuno
de 3 a 4 días
Flujos de
combustibles
en ayuno
prolongado
control de
flujos en ayunoalgunos
efectos
hormonales
no es posible
asegurar la
supervivencia si no
se reduce la
perdida de
proteínas
La disminución del consumo de AA
en la gluconeogenesis se demuestra por
la caída en la tasa de excreción
de nitrógeno en la orina
El glucógeno
hepático probé
la glucosa necesaria
para el funcionamiento
del cuerpo
Para el buen
funcionamiento
del organismo
La falta del aporte
exógeno de
combustible
hace necesaria
la utilización
de las reservas
endógenas
mayor tasa de captación de GLN por los riñones
con mayor liberación de amonio en la orina
y de G por la gluconeogenesis a la sangre.
reducción en los niveles
plasmáticos de la insulina
y la de la triyodotironina.
Los niveles bajos pero constantes de la glucemia se
mantienen por la intervención de las hormonas
mal llamadas contrarreguladoras
La diferencia
entre el ayuno
nocturno y de tres
días es la caída de
los niveles de T3
y de la RaG y el
incremento
de la proteólisis.
El antagonista
beta adrenérgico
disminuyó tanto
la lipólisis como
la participación
de los LIP al
gasto energético.

APORTE EXÓGENO DE
COMBUSTIBLES

GENERALIDADES
La ingestión diaria de calorías de los
seres humanos puede ser muy variable
en cuanto a cantidad, composición
relativa de los tres tipos de macro
nutrientes y distribución de la toma de
alimento a lo largo del día. En lo que a
los macro nutrientes se refiere, antes se
pensaba que el organismo es
energéticamente ciego, que las calorías
obtenidas contribuyen de manera
indiferenciada al balance energético ,
sin importar su origen.

MÁS GENERALIDADES….
Los datos experimentales más recientes se han enfocado
sobre todo en estudiar los efectos de la composición de
las dietas más que de la energía total de ésta, primero
por la preocupación del problema de obesidad y,
segundo, porque se averiguo que el manejo endógeno
de los tres tipos de combustibles puede influir
importantemente en el metabolismo energético.
El balance energético se realiza mediante la regulación
por separado del balance de carbohidratos, lípidos y
proteínas.
VACIAMIENTO GÁSTRICO
Un primer dato relacionado con la toma
de alimento es el paso químico del
estómago al duodeno. Los alimentos
líquido se vacían más rápidamente que
los sólidos.
En el humano se estima que un alimento
líquido se vacía al duodeno con una tasa
aproximada de 200 Cal/h.
La relativa constancia calórica de la tasa
de vaciamiento gástrico se interpreta ser
el resultado de la retroalimentación
ejercida por quimiorreceptores
intestinales presentes aun en el yeyuno.

COSTO ENERGÉTICO DE LA INGESTIÓN (CEI)
Un 10% de la tasa metabólica diaria de un adulto moderadamente activo se
debe al costo energético de la ingestión. Se determinó la existencia de dos
fases del CEI llamadas: “termogénesis facultativa”, aparentemente inducida
por la activación del sistema simpatoadrenal y “termogénesis obligada”,
producida por la digestión, absorción y puesta en reserva de los
combustibles exógenos.
Las entradas sensoriales iniciadas por la ingestión activa la oxidación bajo
control neurológico; las concentraciones de insulina y de noradrenalina
aumentan en los primeros 10 min de la ingestión para disminuir después.

Se recomienda que los estudios acerca del CEI se hagan
durante 5 a 6 horas. Su valor es proporcional al peso
magro e inversamente proporcional a la masa adiposa.
También se correlaciona con la cantidad de calorías
ingeridas, pero sólo hasta un valor de saturación.
Disminuye algo con la edad, posiblemente debido a una
resistencia a la estimulación simpática. El CEI difiere
según la hora del día, siendo más bajo por la noche.
ENTRADA EXÓGENA DE CARBOHIDRATOS
Si en el caso postabortivo o en el
ayuno se puede hablar de la Ra de la
entrada endógena a la circulación, en
el caso del flujo de origen exógeno
por absorción intestinal este valor es
mucho más difícil de estimar. La tasa
de entrada a la sangre portal de un
metabolito como la glucosa dependerá
del tiempo que transcurrió desde la
ingestión, de su concentración en la
luz del intestino, de su utilización por
el epitelio intestinal, etc

Como un dato muy general, una toma normal de alimento mixto
tarda en absorberse unas 5 o 6 horas y se necesitan unas 8 horas
para almacenar y oxidar todos los metabolitos ingeridos.
Las principales rutas que puede tomar la glucosa exógena son:
oxidación, síntesis de glucógeno hepático y muscular, producción
de lactato, lipogénesis y producción de aminoácidos no esenciales.
En cualquier caso, la administración de carbohidratos induce la
secreción de insulina la cual facilita la entrada de glucosa a los
tejidos dependientes de esta hormona, principalmente el
muscular, el cardiaco y el adiposo. La acción de la insulina se
ejerce a través de unos transportadores membranales específicos
llamados GLUT-4 y, en menor grado GLUT-1.

Las diferencias cuantitativas en cuanto a la
distribución de la glucosa exógena puede
depender de la vía de entrada, intravenosa
o por absorción intestinal. Puede diferir
el flujo de entrada, la temporalidad de la
secreción de insulina y la suerte hepática
de la glucosa.
El exceso de glucosa de origen exógeno
determina una hiperglucemia en forma
de campana, con pendiente ascendente
más abrupta que la descendente,
indicando que la absorción intestinal
primero rebasa la captación celular para
luego quedar atrás.

La hiperglucemia induce una hiperinsulinemia y disminución
en los niveles circulantes del glucagón; todo esto inhibe en
cierta medida la producción hepática de glucosa. Sin embargo
el control de la secreción de estas dos hormonas comprende
una fase cefálica importante
La tasa de recambio del glucógeno hepático es directamente
proporcional a su concentración, sugiriendo que éste es un
mecanismo de regulación del contenido de la reserva.
La síntesis de glucógeno hepático a partir de la glucosa
exógena sólo representa un 40 a 50% de la síntesis total; lo
demás se hace por vía gluconeogénica.

Lo que resulta cuantitativamente claro en cuanto al
camino metabólico de la glucosa exógena es que:
1.-Un 40 a un 50% está captada por los músculos que la
usan principalmente en la síntesis de glucógeno.
2.- La síntesis hepática directa de glucógeno representa
sólo un 40 a 50% del total
3.- Los niveles de insulina y la respuesta tisular a la
hormona influyen importantemente en las vías de
utilización pero mucho menos en las de la utilización de
fructosa.

ENTRADA EXÓGENA DE LÍPIDOS
La mayor parte de la grasa
ingerida se absorbe como ácidos
grasos y monoglicéridos. Sólo
los ácidos grasos de cadena
menor de 12 carbonos pasan a
los capilares intestinales; el resto
se reesterifica en el epitelio
intestinal, se asocia con unas
proteínas para formar
quilomicrones que pasan a los
capilares linfáticos y por ende a
la cava.

La ingestión de un alimento mixto conteniendo 41% de
sus calorías en lípidos, hace que los quilomicrones
plasmáticos lleguen a la concentración máxima en 4 a 5
horas. La actividad de la lipo proteinlipasa en la pared
de los capilares del tejido adiposo aumenta y disminuye
la de la lipo proteinlipasa muscular. La captación de
áciods grasos sólo se ve incrementada en los adipositos.

ENTRADA EXÓGENA DE AMINOÁCIDOS
En el manejo de los aminoácidos, exógenos o
endógenos, resalta el papel de los aminoácidos
esenciales. Su proporción total en las proteínas
corporales es de un 50%, lo que implica que su tasa de
recambio es de unos 100 a 150g al día. No obstante,
sus requerimientos diarios son de sólo unos 6g al día.
Esto indica un reciclaje extenso y eficiente,
principalmente en el hígado.

•La absorción intestinal de aminoácidos depende mucho
de las características de las proteínas ingeridas.
•Los aminoácidos exógenos afectan proporcionalmente
más la síntesis que la degradación proteica muscular.
•La cantidad de proteínas ingeridas afecta, como es de
esperarse, el flujo de aminoácidos y la excreción de
nitrógeno.

ALGUNOS EFECTOS HORMONALES
La hiperinsulinemia no afecta el transporte de la
glucosa a través de la barrera hematoencefálica ni se
metabolismo cerebral, de manera que los cambios en
el metabolismo de los carbohidratos provocados por
estas metodologías no involucran al cerebro.

La hiperinsulinemia en condiciones de euglucemia
inhibe la lipólisis. La oxidación de lípidos y la
producción hepática de glucosa y aumenta la
utilización de la glucosa por las tres vías: la síntesis de
glucógeno, oxidación y producción de lactato. El
incremento en las primeras dos vías se hace,
prácticamente en paralelo, pero el de la tasa de
síntesis del glucógeno es mayor que el de la oxidación

Con insulinemia basal, la hiperglucemia incrementa la RdG. Lo
que expande el pozo de glucosa-6-fosfato lo que, a su vez,
induce mayor sínteesis de glucógeno y oxidación de la glusosa.
En hiperglucemia e hiperinsulinemia, tanto la glucosa como la
insulina tienen, además efectos intracelulares, la sintetasa de
glucógeno está estimulada no sólo por la insulina sino de manera
alosterica por el mayor nivel de glucosa-6-fosfato. En cambio, la
actividad de la piruvatodeshidrogenasa ya no aumenta durante la
hiperinsulinemia, por lo que no se incrementa más la oxidación
de glucosa.
La hiperglucemia sola no cambia mucho la tasa de oxidación de
los lipidos si no está acompañada de hiperinsulinemia

Varios de los trabajos confirman el efecto
negativo que tiene los lípidos sobre la oxidación
de la glucosa, otros autores demostraron que lo
inverso también ocurre, en el sentido de que la
oxidación de lípidos está inhibida por la
disponibilidad intracelular de la glucosa.

CONCLUSIONES
La ingestión determina aumento del gasto energético
(CEI), aumento diferente cuantitativamente en función de
la prevalencia de uno u otro de los macro nutrientes
(PRO>CHO>LIP). El CEI tiene una fase facultativa
inicial, no dependiente de los gastos energéticos debidos a
la absorción, la digestión y la puesta en reserva, que podría
estar determinada por la fase cefálica de la secreción de
insulina y de la activación del sistema nervioso central.

La síntesis de glucógeno hepático a partir de la glucosa absorbida se
hace por dos vías:
-La directa a partir de la misma molécula de glucosa
-La indirecta, por vía gluconeogénica, a partir de piruvato y/o
lactato que se originan por glucólisis intra y/o extra hepática.
Los quilomicrones fresultantes de la absorción de los ácidos grasos de
cadena larga pueden ser fuentes de ácidos grasos libres por acción de
la liopoproteinlipasa adiposa o de ácidos grasos captados
directamente por otro tejidos, a ´partir de las mismas proteínas
circulantes.
La ingestión de un alimento rico en proteínas aumenta la síntesis
proteica y disminuye la proteólisis, pero también aumenta la
oxidación de aminoácidos.

La ingestión diaria de calorías de los seres humanos puede ser muy variable en
cuanto a cantidad, composición relativa de los tres tipos de macro nutrientes y
distribución de la toma de alimento a lo largo del día.
Los alimentos líquidos se vacían más rápidamente que
los sólidos.
la administración de
carbohidratos induce la
secreción de insulina la
cual facilita la entrada
de glucosa a los tejidos
dependientes de esta
hormona,
principalmente el
muscular, el cardiaco y
el adiposo
La mayor parte de la grasa
ingerida se absorbe como
ácidos grasos y
monoglicéridos. La ingestión
de un alimento mixto
conteniendo 41% de sus
calorías en lípidos, hace que
los quilomicrones
plasmáticos lleguen a la
concentración máxima en 4
a 5 horas
La absorción intestinal
de aminoácidos depende
mucho de las
características de las
proteínas ingeridas.
Los aminoácidos
exógenos afectan
proporcionalmente más
la síntesis que la
degradación proteica
muscular
APORTE EXÓGENO DE
COMBUSTIBLES
ENTRADA EXÓGENA
DE CARBOHIDRATOS
ENTRADA EXÓGENA DE
LÍPIDOS
ENTRADA
EXÓGENOADE
AMINOÁCIDOS
La oxidación de lípidos
y la producción
hepática de glucosa y
aumenta la utilización
de la glucosa por las
tres vías: la síntesis de
glucógeno, oxidación y
producción de lactato
ALGUNOS EFECTOS
HORMONALES

Conclusiones

Conclusión:
En este caso, me costó más entender ciertos procesos y
reacciones pero al final puedo concluir la importancia de
tomar en cuenta las entradas y salidas de alimento con
relación a la energía recibida y la que se gasta, la cual
dependerá del organismo y su metabolismo junto con las
disposiciones genéticas predispuestas. Por ello se debe abarcar
más ampliamente el área bioquímica.

El manejo adecuado del control energético enfocado y aplicado a
nuestra población podría disminuir las tasas de obesidad en
cualquier nivel y las de desnutrición mejorando la calidad de
vida. Además de poder hacer un uso más conciente de las
aplicaciones de la energía y encontrar métodos o formas más
sencillas y precisas para medir el estado calórico corporal ya que
el cuerpo en su totalidad lleva a cabo diversas reacciones, como
oxidaciones, fosforilaciones, hidrólisis, etc. Que a veces
interfieren en determinaciones.

Los seres humanos tenemos un aporte energético que
proviene de una dieta variada en cuanto a tipos de alimentos y
cantidades. Cada tipo de macronutrientes tiene un
metabolismo diferente dentro del cuerpo humano, pero sin
embargo todas esas rutas metabólicas conllevan a un mismo
fin, la producción de energía. Las rutas metabólicas tienen
interacciones entre ellas, lo que significa que al metabolizar
un tipo de macronutriente se altera el metabolismo de otro.
Conclusión:

Conclusión personal
Loreto Pérez Ana Laura.
Comprender el metabolismo del ser humano es difícil yse han realizado muchos estudios que han durado años,pero es de suma importancia conocer los factores queintervienen en el metabolismo, el enfoque analítico es laherramienta que nos permite distinguir si los valoresestán en un rango establecido o no y estos nosdemuestran si existe alguna alteración fisiológica, estopuede ser de gran ayuda para el tratamiento deenfermedades o de alguna manera prevenirlas.

Conclusión personal
Rojas Morales Alberto
El metabolismo energético son todas las vías y rutas que utiliza elmetabolismo, para mantener en equilibrio y buen funcionamiento, de elrendimiento energético optimo, para la supervivencia del organismo.
El almacenamiento, distribución, uso y eliminación de todos los recursosque tiene el cuerpo humano se ven implicados en este proceso cuyameta principal es el de obtener energía suficiente para mantener la vidadel propio organismo, si bien muchos de estos procesos llevados alextremo implican la destrucción de las estructuras del mismo cuerpo esde maravillarse que el mismo organismo sea el que de la orden y la llevea cabo de tal manera que nada se desperdicia y mantiene la vida aun enlas peores circunstancias y siempre en un optimo desempeño.
La importancia del estudio de las reacciones bioquímicas del organismollevado acabo por diversos autores es simplemente descubrir todosaquellos procesos y mecanismo que mantienen y lograr llevar a cabo lasustentación de la vida diaria del ser humano su comprensión es unaverdadera arma contra las enfermedades fisiológicas y metabólicas dellos seres vivos







