
1
Mobile &Transposable Elements
JJuummpipinng g
GGeenneess

2
Ethnobotany
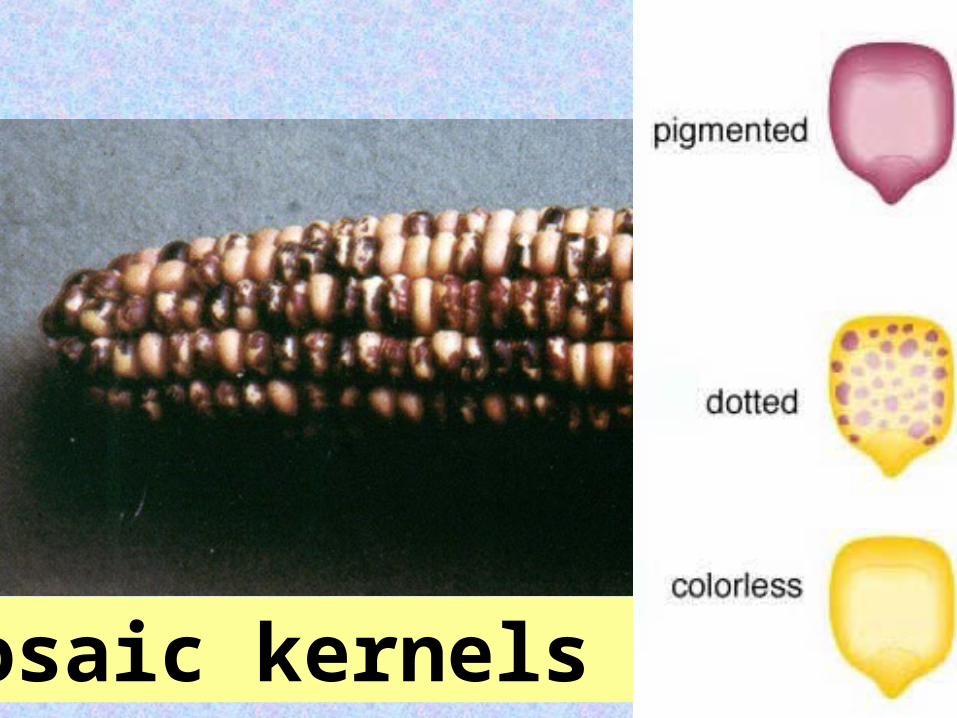
3mosaic kernels

4
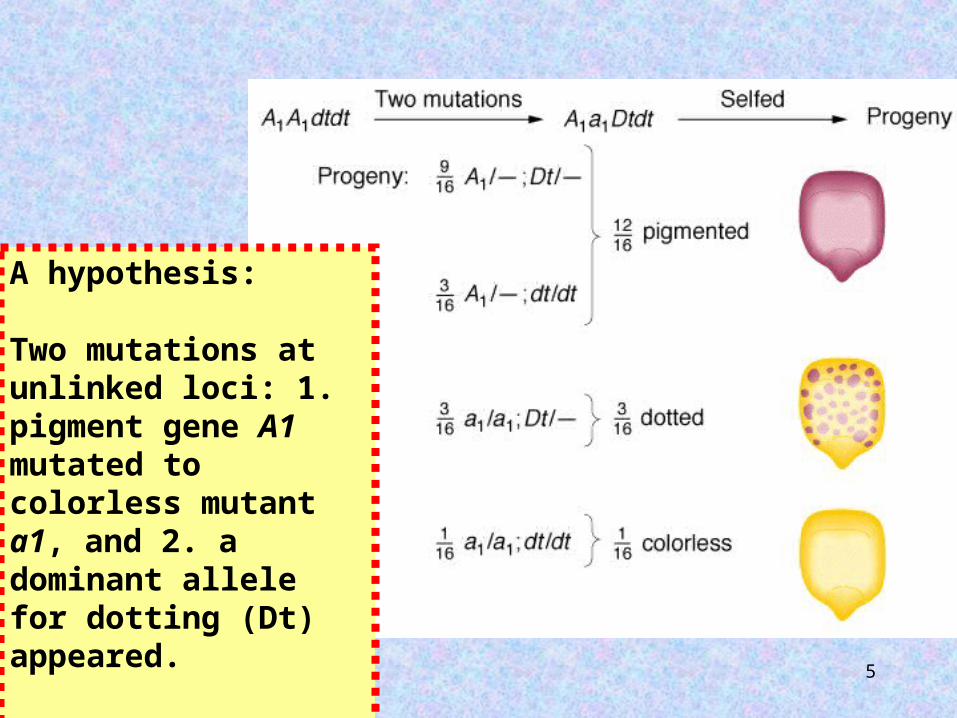
1938: Marcus Rhoades reported odd phenotypic ratios in corn.Self pollination
of a pigmented corn kernel yielded:
12 : 3 : 1
pigmentedpigmented : dotted : colorless
5
A hypothesis:
Two mutations at unlinked loci: 1. pigment gene A1 mutated to colorless mutant a1, and 2. a dominant allele for dotting (Dt) appeared.
The presence of the Dt allele caused spots of pigment to appear.

6
Barbara McClintock1902-1992

7
Barbara McClintock1902-1992

8
9
10

11
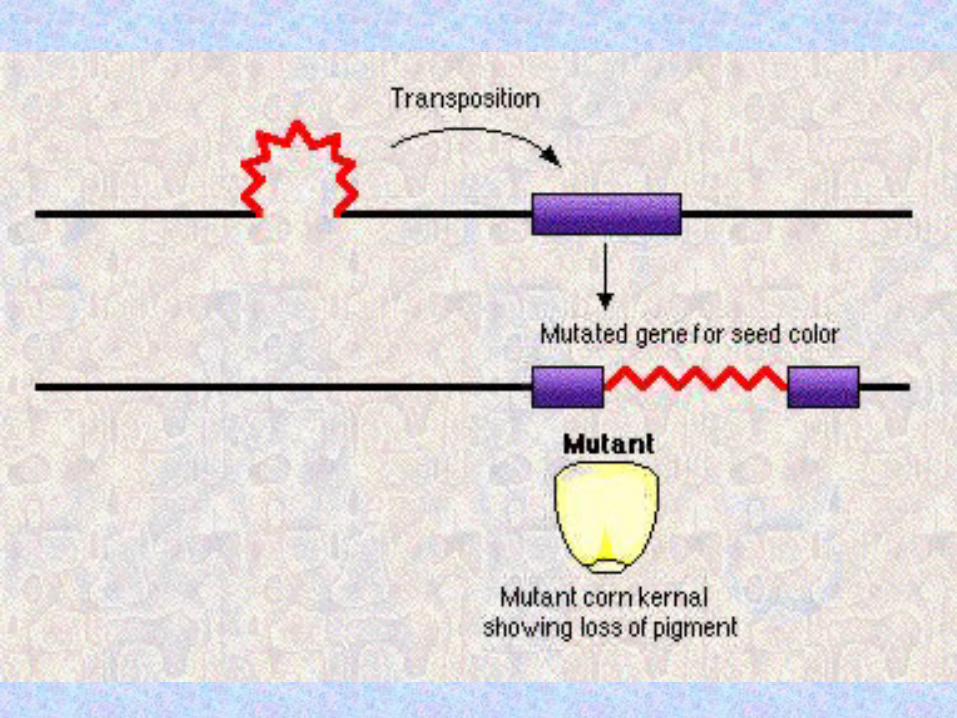
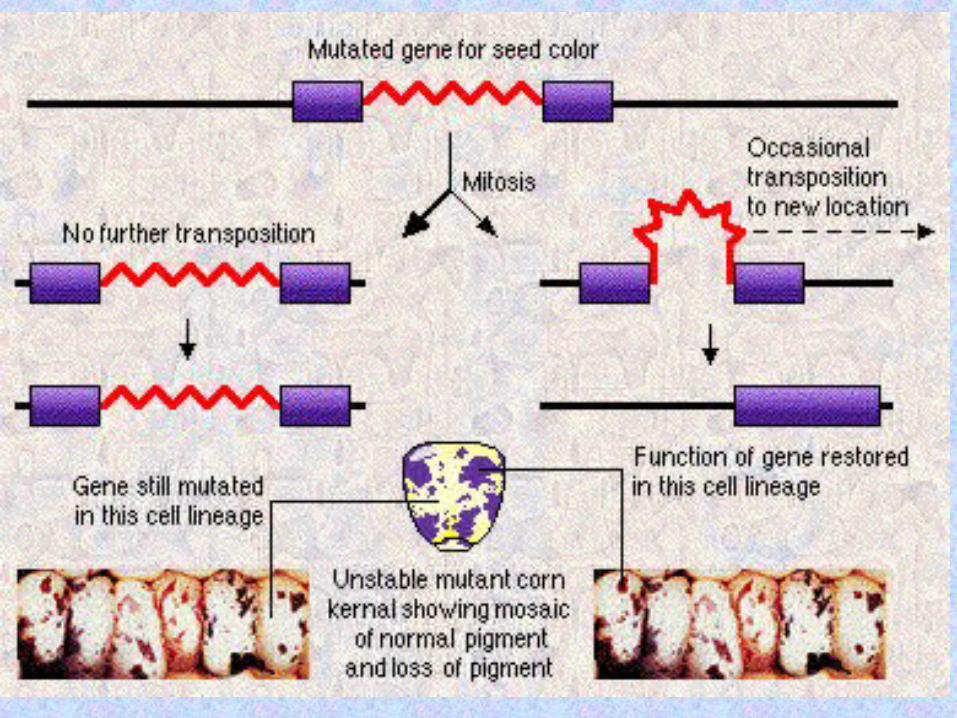
TranspositionTransposition = the = the movement of genetic movement of genetic information from one information from one chromosomal location, chromosomal location, the the donor sitedonor site, to , to another, the another, the tartargget et sitesite. .

12
DNA sequences that can DNA sequences that can change their genomic change their genomic location location intragenomically intragenomically either either autonomouslautonomously or y or non-autonomouslnon-autonomouslyy are are called called transtranspposable osable elementselements. .

13
“copy-and-paste”“cut-and-paste”

14
Retrotransposons can be divided into five orders on the basis of their mechanistic features, sequence organization, and reverse transcriptase phylogeny: LTR retrotransposons, DIRS-like elements, Penelope-like elements, LINEs, and SINEs.
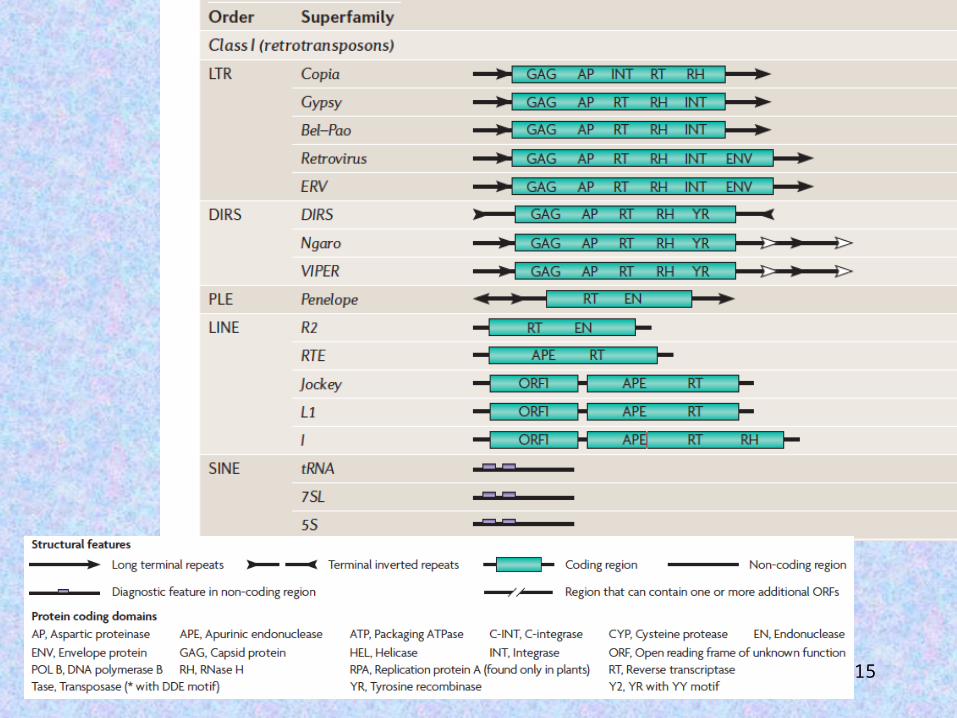
15

16
When a transposable element is When a transposable element is inserted into a host genome, a inserted into a host genome, a small segment of the host DNA small segment of the host DNA (usually 4-12 bp) is (usually 4-12 bp) is duplicated at the insertion duplicated at the insertion site. site.
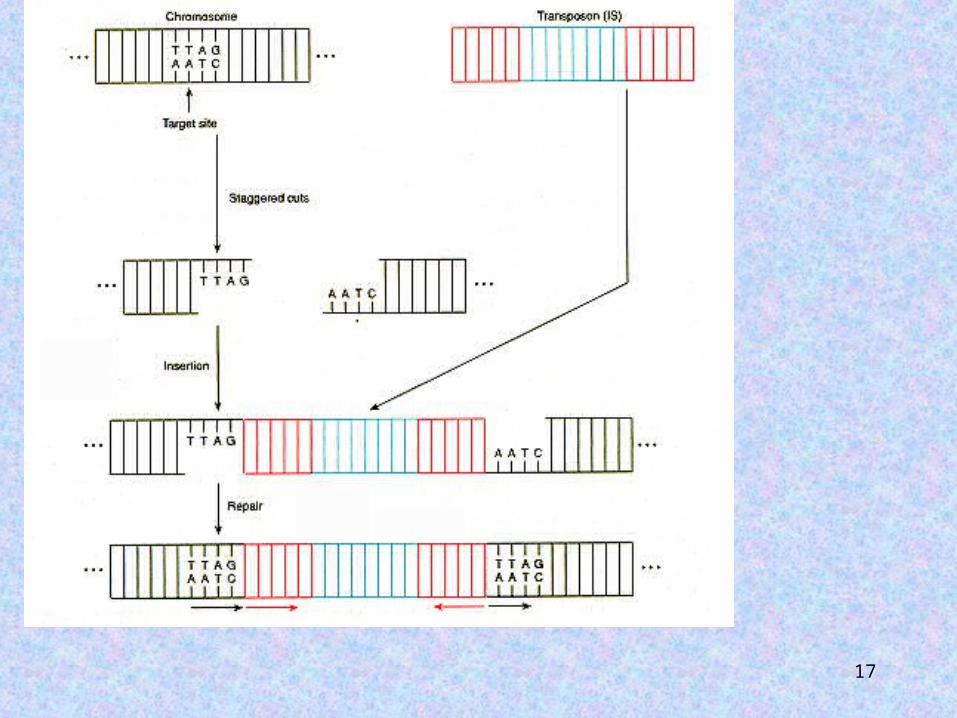
17

18

19
TranspositionTransposition = the = the movement of genetic movement of genetic information from one information from one chromosomal location, chromosomal location, the the donor sitedonor site, to , to another, the another, the tartargget et sitesite. .

20
DNA sequences that can DNA sequences that can change their genomic change their genomic location location intragenomically intragenomically either either autonomouslautonomously or y or non-autonomouslnon-autonomouslyy are are called called transtranspposable osable elementselements. .

21
Classifications of Transposable Elements

22
Transposition may be replicative or conservative. Replicative transposition will result in two copies of the element, one at the donor site and one at the target site. Following conservative transposition the transposable element will only be found at the target site, with no change in copy number.
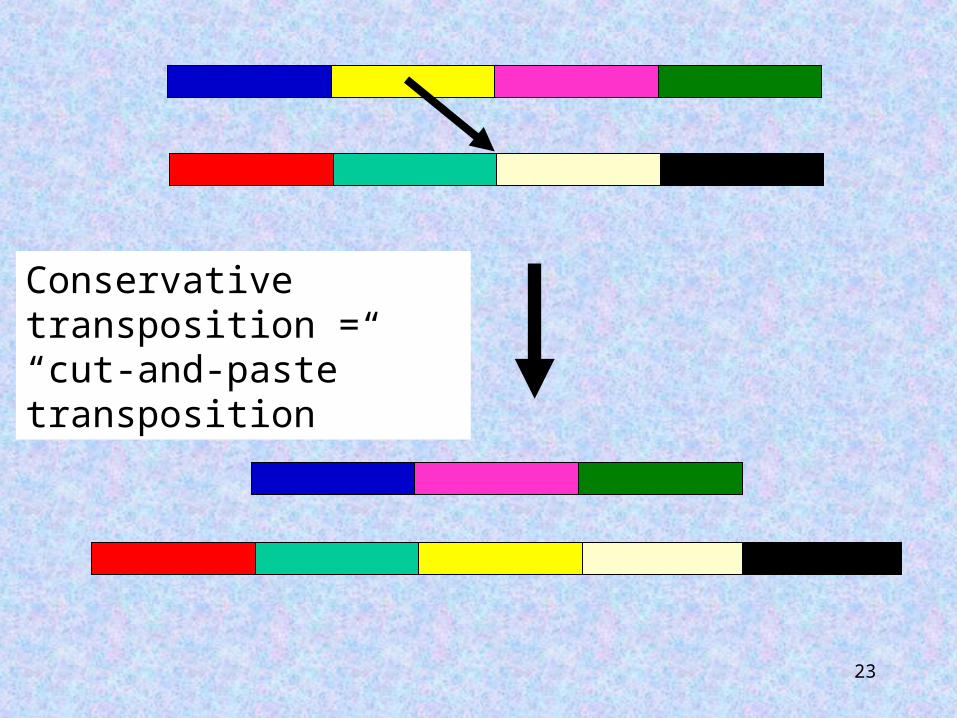
23
Conservative transposition = “cut-and-paste” transposition
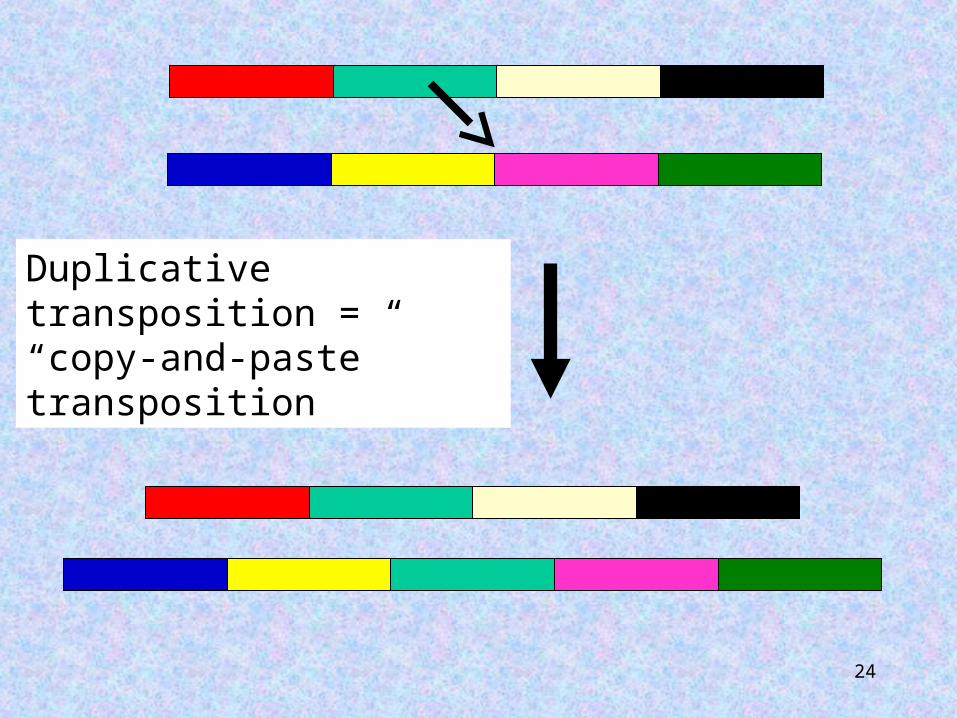
24
Duplicative transposition = “copy-and-paste” transposition

25
DNA-mediated and RNA-mediated transposable elements:
1.Class I transposable elements (retrotransposons).2.Class II transposable elements (DNA transposons).
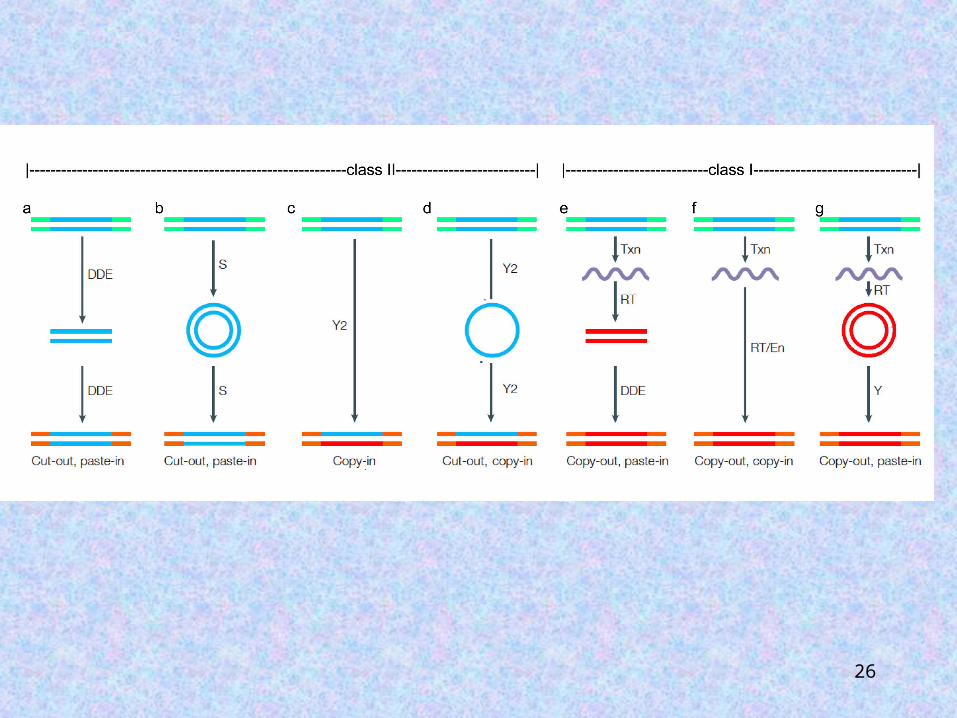
26

27
Autonomous and nonautonomous transposable elements Autonomous transposable elements encode all the components of the transposition machinery.
Nonautonomous transposable elements appropriate the transposition machinery of autonomous transposable elements.

28
Active and fossil transposable elements
A transposable element is defined as active if it contains all the necessary sequence elements for either autonomous or nonautonomous transposition.
Active elements may be rendered defective by different types of mutation, in which case they are referred to as fossil transposable elements.

29
Active and fossil transposable elements
A transposable-element family may contain different combinations of active autonomous, active nonautonomous, fossil autonomous, and fossil nonautonomous transposable elements.
For example, the human genome contains approximately 50,000 fossil autonomous and 200,000 fossil nonautonomous DNA transposons.
Intriguingly, the human genome seems to contain NO active DNA transposons.

30
According to the numbers and kinds of genes they contain, DNA-mediated transposable elements are divided into insertion sequences and transposons.

31

32
Found in Escherichia coli and Shigella dysinteria.
Length = 770 nucleotides, including two inverted terminal repeats, 23 bp each.
Contains two out-of-phase reading frames, insA and insB, from which a single protein is produced by translational frameshifting at a run of adenines.
The N-terminal is an inhibitor of transposition; the C-terminal is a transposase, an enzyme that catalyzes the insertion of transposable elements into insertion sites.

33
Carriers and non-carriers of the insertion sequence can be separated by centrifugation because the carriers are heavier.

34
galE = UDP-galactose 4-epimerase galT = galactose-l-phosphate uridylyltransferase galK = galactokinase galM = mutarotase
Galactose (gal) operon
galE galT galK galM
INSERTION SEQUENCES (IS)Insertion sequences were first discovered in the gal operon of E. coli.

35
Insertion of an IS affects only the transcription of the genes downstream from the insertion. For example, if the IS occurs in the galT gene, the galT, galK and galM genes will be disrupted, but galE will not be.This phenomenon is known as a POLARPOLAR mutation.
Galactose (gal) operon
galE galT galK galM
IS

36

37

38

39

40
Stupidity
41
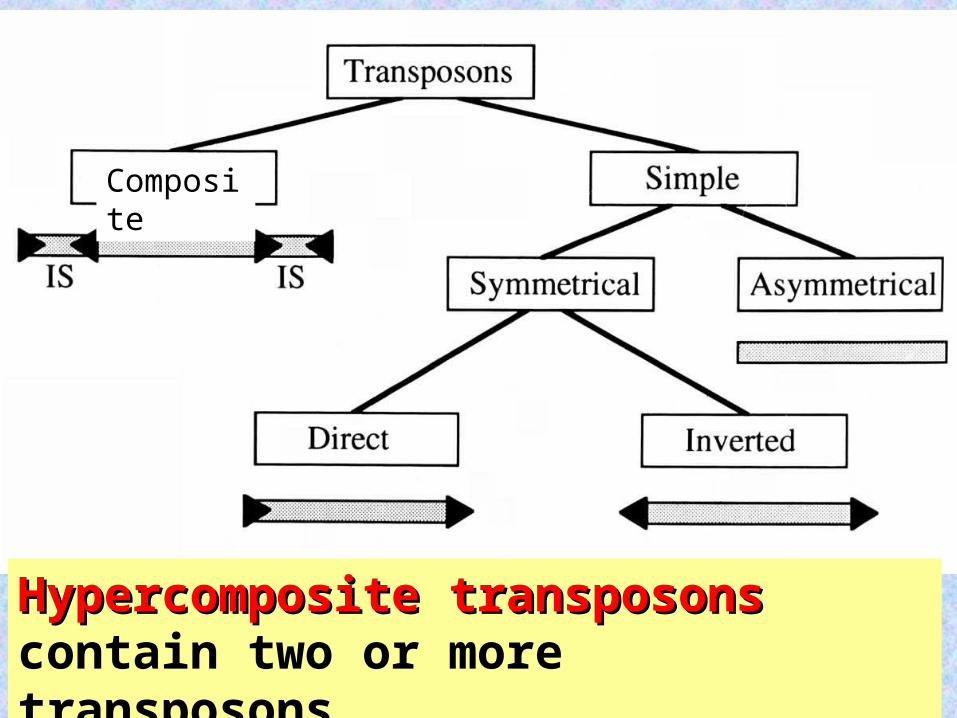
Hypercomposite transposonsHypercomposite transposons contain two or more transposons.
Composite

42
(symmetrical-inverted) Tn3 from E. coli confers streptomycin resistance. tnpR and bla are transcribed on one strand; tnpA on the other. Tn3 is flanked by 38-bp-long inverted repeats.
(asymmetrical) Tn554 from Staphylococcus aureus lacks terminal repeats and contains 8 protein-coding genes. Three of the genes are transcribed as a unit and encode transposases (tnpA, tnpB, and tnpO). The spc and ermA genes confer spectinomycin and erythromycin resistance, respectively.

43
Composite transposon Composite transposon Tn9 from Escherichia coli contains two copies of IS1 flanking the cat gene, which encodes a chloramphenicol-resistancechloramphenicol-resistance protein.

44
Some mobile elements can transpose themselves in all cells; others are cell-specific.
Tc1 elements in the nematode Caenorhabditis elegans and P elements in Drosophila melanogaster are usually mobile only in germ cells.

45
Transposition of many Transposition of many elements is elements is regulated by regulated by developmental stagedevelopmental stage. .
From an evolutionary From an evolutionary point of view, the point of view, the developmental timing of developmental timing of transposition is transposition is particularly important, particularly important, because it affects the because it affects the propagation of the propagation of the transposable element to transposable element to future generations. future generations.

46
LINE-1 transposable elements in mammals are particularly active during leptoteneleptotene and zygotenezygotene, when DNA-strand breakages occur. This offers an opportunity for transposable elements to insert themselves into new sites.

47
Genomic locations of target sites for transposition:
Exclusive genomic preference: In the vast majority of cases IS4 incorporates itself in the galactosidase operon of Escherichia coli, and thus each bacterium contains mostly one copy of IS4.
Complete randomness: Bacteriophage Mu transposes itself at random within the genome.
Intermediate genomic preference: 40% of all Tn10 transposons in E. coli are found in the lacZ gene, which constitutes a minute fraction of the host genome.

48
Genomic locations of target sites for transposition:
Affinity for a particular nucleotide composition: IS1 favors AT-rich sites.
Affinity for a particular sequence: IS630 has a special affinity for 5'—CTAG—3' sequences.
Chromosomal preference: TRIM elements in Drosophila miranda exhibit a preference for the Y chromosome.

49
Hotspots for P element insertion in the X chromosome of Drosophila melanogaster

50
Genomic locations of target sites for transposition:
The DIRS-1 transposable element in the slime mold Dictyostelium discoideum

51
Genomic locations of target sites for transposition:
Self affinity: DIRS-1 preferentially inserts itself into other DIRS-1 sequences. D. discoideum contains, on average, ~40 intact copies of DIRS-1 and ~300 fragments.
oldest oldestnewest
Active DIRS

52
Species specificity:
DIRSDIRS in Dictyostelium discoideum only.
marinermariner moves from species to species, even if the species belong to different taxonomic kingdoms.

53
Retroelements are sequences that contain a gene for reverse transcriptase, which catalyzes the synthesis of cDNA from an RNA template.
Not all retroelements possess the intrinsic capability to transpose. Therefore, not all retroelements are transposable elements.
Retroelements that transpose do so by retroposition.

54
Central Dogma

55
Modified Central Dogma
56
Not the Central Dogma
57
That would have been nice…

58
That would have been nice…
Joanna Masel

59

60
Retroelements
Retroelements are DNA or RNA sequences that contain a gene encoding the enzyme reverse transcriptase, which catalyzes the synthesis of DNA from an RNA template. The resulting DNA molecule is called complementary DNA (cDNA). Not all retroelements are transposable or mobile.
Retroelements can be divided into three categories: (1)transposable elements that move within a genome by replicative RNA-mediated transposition (but may also move intergenomically)(2) mobile nontransposable elements that only move intergenomically(3) non-mobile elements

61
Masayori InouyeMasayori InouyeRutgers UniversityRutgers University

62
Retrons are widely distributed among bacterial species.
Within each bacterial species retrons tend to be rare.
Natural populations of retron-carrying genomes possess a single retron copy, either in the intergenic part of the genome or inside a prophage (a viral genome that had became integrated into the bacterial chromosome).

63
multicopy single-stranded DNA (msDNA)

64
TERT genes Human telomeres consist of the sequence TTAGGG tandemly repeated many thousand times. Because of asymmetrical DNA replication, a few of these repeats are lost from the tips of the chromosomes each replication cycle.

65
TERT genes Telomerases are nucleoproteins whose function is to add DNA-sequence repeats to the 3' end of the DNA strands in the telomeres at the ends of linear eukaryotic chromosomes.
The de novo addition of TTAGGG repeats by the enzyme telomerase partially or wholly compensates for telomere shortening.
Telomerases in all eukaryotic species share at least two components essential for catalytic activity: a telomerase reverse transcriptase protein (TERT) and a telomerase RNA.
The TERT encoding gene is a retroelement. In humans, this retroelement is located on chromosome 5.

66
Retroplasmids
Extragenomic DNA and RNA molecules (plasmids) are frequently detected in fungal mitochondria. They can be divided into:
- Genomically derived plasmids (similar in sequence to the mitochondrial sequence)- Autonomously-replicating true plasmids that exhibit no sequence similarity with the host mitochondrial genome.

68
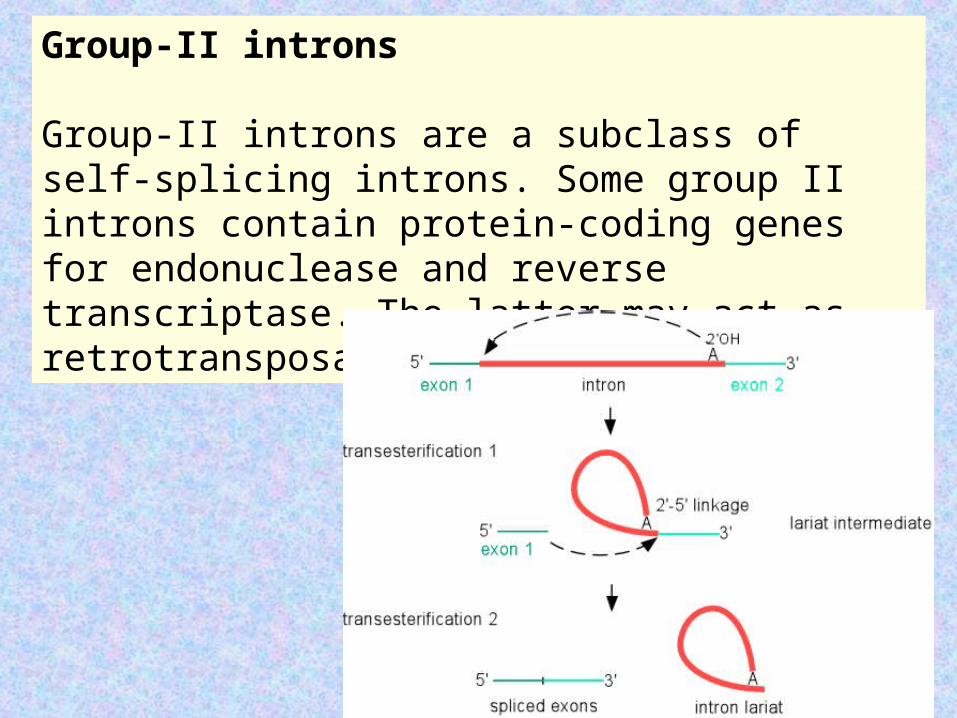
Group-II introns
Group-II introns are a subclass of self-splicing introns. Some group II introns contain protein-coding genes for endonuclease and reverse transcriptase. The latter may act as retrotransposable genetic elements.

69
Group-II introns
Group-II introns can integrate into the homologous position of an intronless allele of the same gene (homing), and at much lower frequencies into other sites (retroposition).
Retroposition

70
Twintrons are introns-within-introns excised by sequential splicing reactions.
Group II twintrons have presumably been formed by the insertion of a group II intron into an existing group II intron.

71
Group-III introns are short ORF-less introns found in a small number of protist eukaryotes, such as Euglena gracilis. They appear to be group-II introns from which the central ORF-containing portion has been removed.
Thus, group-III introns are essentially nonautonomous group-II introns.
Group III twintrons are known.

72
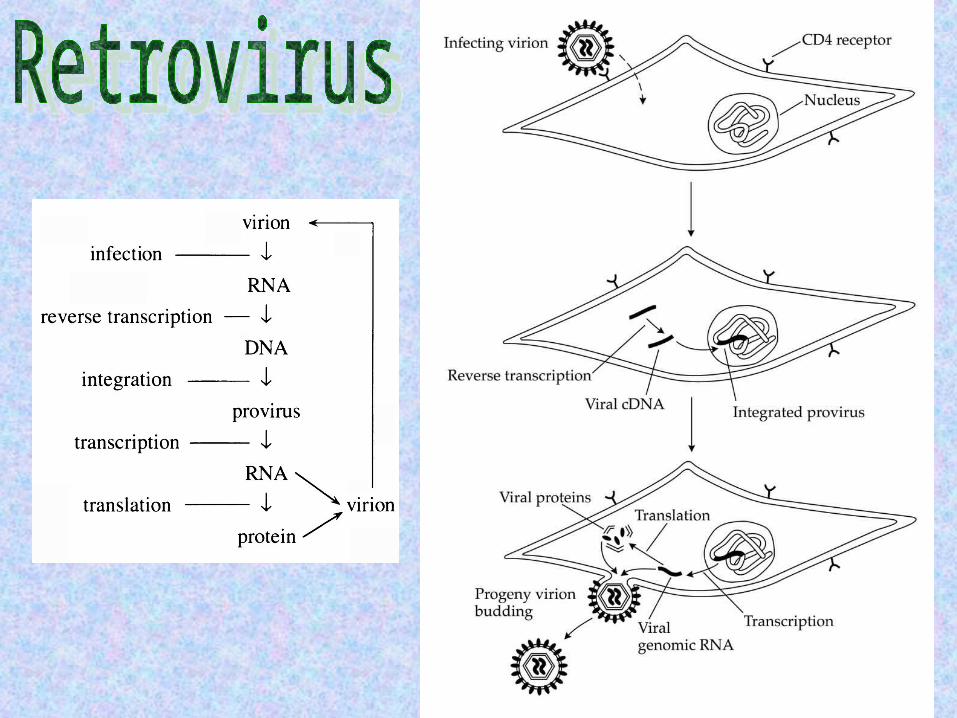
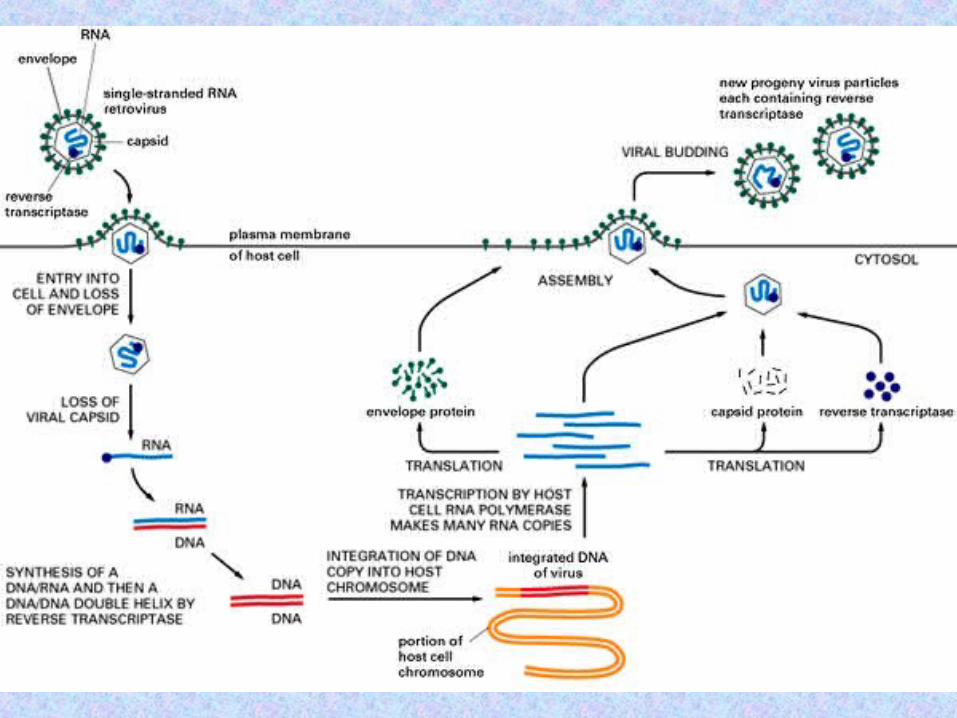
Retrotransposons are transposable elements that use RNA-mediated transposition, but do not construct virion particles, i.e., they lack the env (envelope) gene, and so, unlike retroviruses, cannot independently transport themselves across cells.
Initially, the retrotransposons were divided into LTR retrotransposons and non-LTR retrotransposons (or retroposons) according to whether or not their coding sequences were flanked by long terminal repeats (LTRs).
Subsequent evolutionary studies indicated that while most non-LTR retrotransposons constitute a monophyletic group, the LTR-retrotransposons are paraphyletic.
Some LTR-retrotransposons have secondarily acquired env-like reading frames that may enable them to move from cell to cell (i.e., they are in practice viruses).

73
LTR retrotransposons
Non-LTR retrotransposons
74

75

76

77
Integrase
78

79
1. HepaDNAviruses, e.g., hepatitis B virus
Pararetroviruses:Pararetroviruses:

80
2. Caulimoviruses, e.g., cauliflower mosaic virus
Pararetroviruses:Pararetroviruses:

81

82
Pararetroviruses are not transposable elements

83
84

85
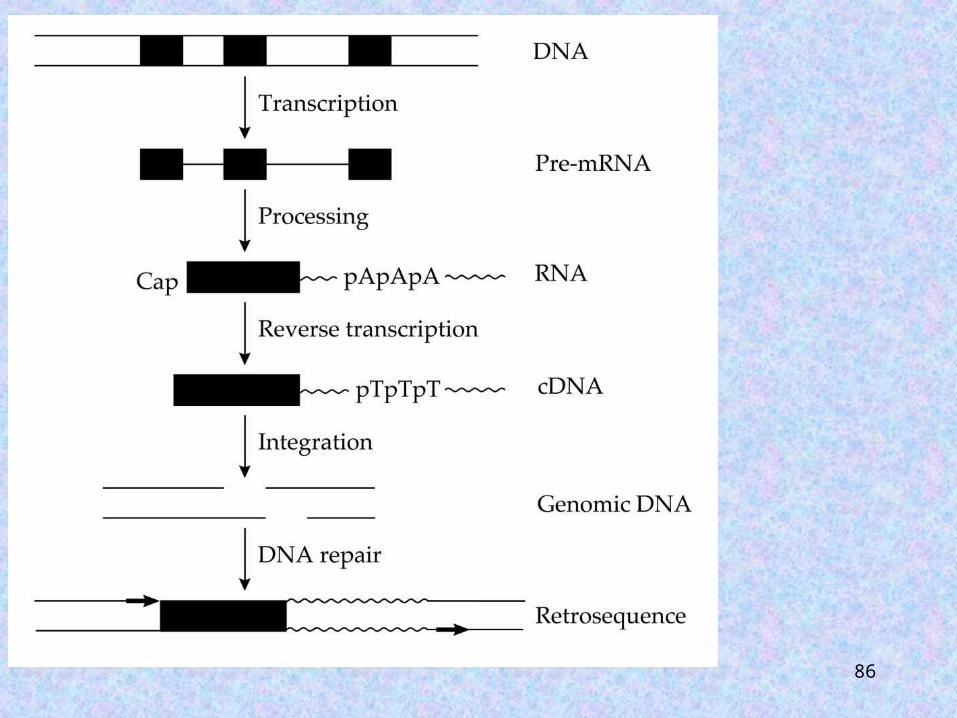
RETROSEQUENCES
Restrosequences (or retrotranscripts) are genomic sequences that have been derived through the reverse transcription of RNA and subsequent integration of the resulting cDNA into the genome.
Retrosequences lack the ability to produce reverse transcriptase, and have been produced through the use of a reverse transcriptase from a retroelement.
86

87
Diagnostic features of mRNA derived retrosequences
1. lack of introns
2. precise boundaries coinciding with the transcribed regions
3. stretches of poly(A) at the 3’ end
4. short direct repeats at both ends
5. truncations
6. posttranscriptional modifications

88

89
manymany
manymany
fewfew

90
Retrosequences:RetrogenesRetrogenes
Processed genesSemiprocessed genes
RetropseudogenesRetropseudogenesProcessed pseudogenesSemiprocessed
pseudogenes

91

92
93
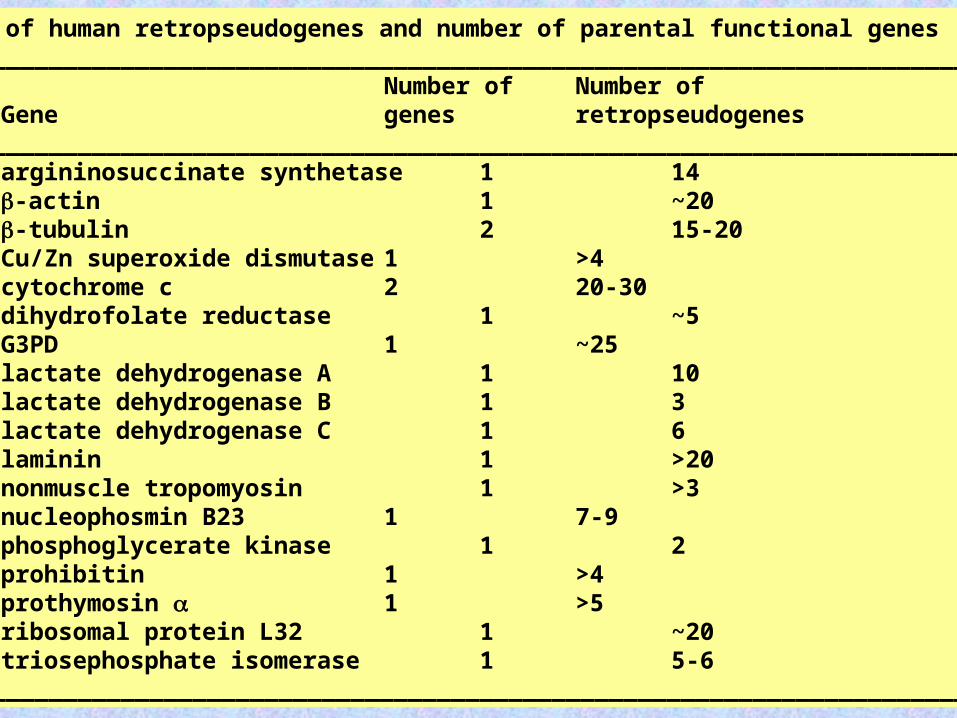
Number of human retropseudogenes and number of parental functional genes__________________________________________________________________________
Number of Number ofGene genes retropseudogenes
__________________________________________________________________________argininosuccinate synthetase 1 14-actin 1 ~20-tubulin 2 15-20Cu/Zn superoxide dismutase 1 >4cytochrome c 2 20-30dihydrofolate reductase 1 ~5G3PD 1 ~25lactate dehydrogenase A 1 10lactate dehydrogenase B 1 3lactate dehydrogenase C 1 6laminin 1 >20nonmuscle tropomyosin 1 >3nucleophosmin B23 1 7-9phosphoglycerate kinase 1 2prohibitin 1 >4prothymosin 1 >5ribosomal protein L32 1 ~20triosephosphate isomerase 1 5-6
__________________________________________________________________________

94
Genes tend to “bombard” Genes tend to “bombard” the genome with dead copies the genome with dead copies of themselves. of themselves.
The The “Vesuvian”“Vesuvian” paradigm paradigm

95
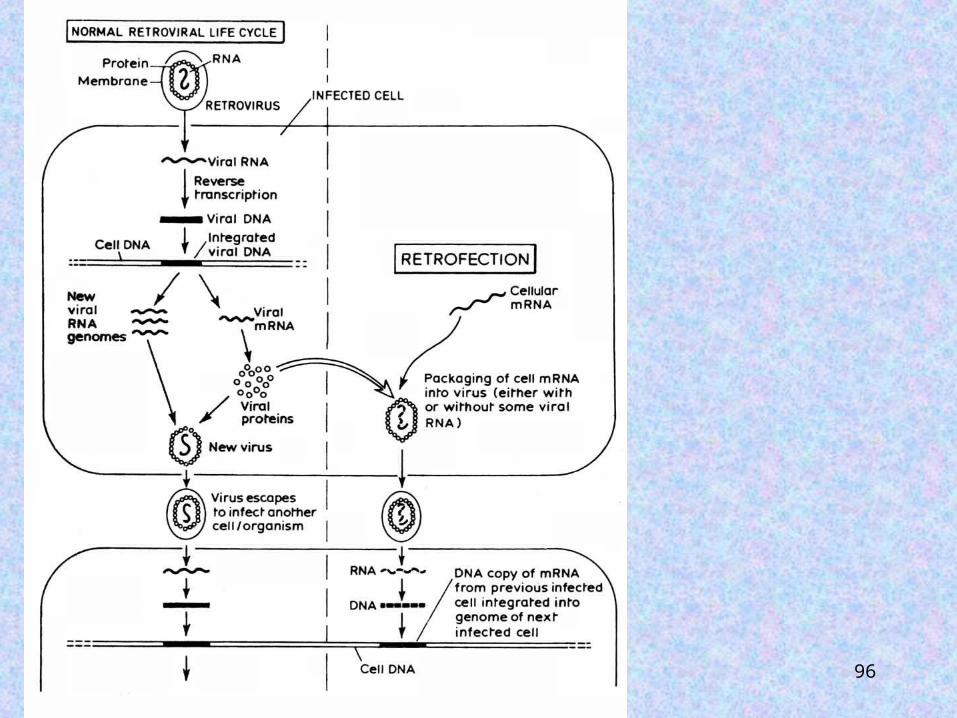
How do you get How do you get reverse transcribed reverse transcribed sequences to become sequences to become incorporated into the incorporated into the germline genome if germline genome if the gene itself is the gene itself is notnot transcribed in transcribed in the germline?the germline?
96

97
Pseudogenes are affected by two evolutionary processes:
Compositional assimilation: The accumulation of mutations which obliterate the similarity between the pseudogene and its functional paralogue. The nucleotide composition of the pseudogene will come to resemble its surroundings, eventually “blending” into it.
Abridgment: Due to the excess of deletions over insertions, pseudogenes become increasingly shorter compared to the functional gene.

98
It takes on average 400 million years for a mammalian retropseudogene to lose half of its length.
Mammals are ~200 million years old and, therefore, the mammalian genome is expected to contain reptilian pseudogenesreptilian pseudogenes. These ancient pseudogenes have by now become unrecognizable.

99
Mammalian processed pseudogenes are created at a much faster rate than the rate by which they are obliterated by deletion.

100
Processed pseudogenes are abundant in mammals.
Processed pseudogenes are rare in amphibians, rarer in birds, and even rarer in Drosophila.

101
Creation rates
Deletion rates
102
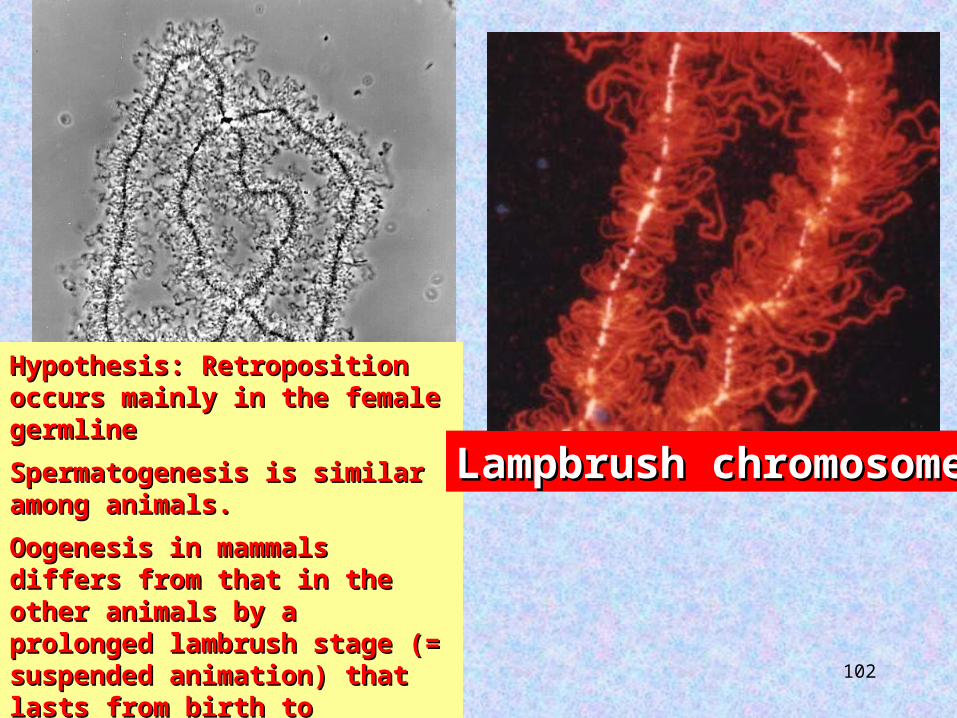
Hypothesis: Retroposition Hypothesis: Retroposition occurs mainly in the female occurs mainly in the female germlinegermline
Spermatogenesis is similar Spermatogenesis is similar among animals. among animals.
Oogenesis in mammals Oogenesis in mammals differs from that in the differs from that in the other animals by a other animals by a prolonged lambrush stage (= prolonged lambrush stage (= suspended animation) that suspended animation) that lasts from birth to lasts from birth to ovulation (up to 40 years ovulation (up to 40 years in humans).in humans).
Lampbrush chromosomeLampbrush chromosome

103
Creation ratesCreation rates are determined by the length of the suspended animation during oogenesis
20-40 years in humans.
2-4 months in amphibians.
Less than 3 weeks in birds.
Less than 1 day in Drosophila.

104
Prediction:
Retrosequences should be found in highhigh numbers on the X chromosomeX chromosome, in intermediateintermediate numbers on autosomesautosomes, and be rarerare on the Y Y chromosomechromosome.

105
Prediction: Retrosequences should be found in highhigh numbers on the X chromosomeX chromosome, in intermediateintermediate numbers on autosomesautosomes, and be rarerare on the Y Y chromosomechromosome.
Density of Processed Pseudogenes in Human Chromosomes(from Bischof et al. 2006) Mean Density Density inDensity in in Autosomes X-chromosome Y-chromosome 2.28 ± 0.40 3.01 0.74

106
Differences among organisms in numbers of retropseudogenes can also be due to deletion ratedeletion rate.
DNA loss in Drosophila is ~75 times faster than that in mammals.
This high rate may explain the dearth of pseudogenes in Drosophila.







