zebrafish tenascin‐w, a new member of the tenascin family
TRANSCRIPT

Zebrafish Tenascin-W, a New Memberof the Tenascin Family
Philipp Weber,1 Dirk Montag,1 Melitta Schachner,1,2 Robert R. Bernhardt1
1 Department of Neurobiology, Swiss Federal Institute of Technology, Honggerberg, HPM,CH-8093 Zurich, Switzerland
2 Zentrum fur Molekulare Neurobiologie, Universitat Hamburg, Martinistrasse 52,D-20246 Hamburg, Germany
Received 3 September 1997; accepted 6 November 1997
also showed high levels of expression. At 3 days, ex-ABSTRACT: A cDNA clone encoding tenascin-pression by sclerotomal and neural crest cells contin-W, a novel member of the tenascin family, was isolatedued to be observed while expression in the somiticfrom a 20- to 28-h postfertilization (hpf) zebrafishmesoderm was decreased. In juvenile fish, tn-w wascDNA library on the basis of the conserved epidermalexpressed weakly by cells in the myosepta and, moregrowth factor-like domains represented in all tenascinstrongly, by presumably nonneuronal cells in the dor-molecules. An open reading frame of 2796 base pairssal root ganglia. In these tissues and at the same devel-encodes a mature protein consisting of heptad repeats,opmental stages, the expression of tn-w partially over-a cysteine-rich amino terminal region, 3.5 epidermallapped with the distribution of tn-c mRNA. In addi-growth factor–like repeats, five fibronectin type IIItion, tn-c was expressed in the central nervous systemhomologous repeats, and a domain homologous to fi-(CNS) and in the axial mesoderm, neither of which
brinogen. These domains are the typical modular ele- expressed tn-w at any of the age stages examined. Thements of molecules of the tenascin family. Sequence expression pattern of tn-w suggests an involvement incomparison demonstrated that TN-W shares homolo- neural crest and sclerotome cell migration and in thegies with the members of the tenascin family but is formation of the skeleton. Similar and possibly over-not a species homolog of any identified tenascin. The lapping functions could also be performed by tn-c,expression pattern of tn-w was analyzed by in situ which appears to have additional functions during thehybridization in 1-day-old embryos, in 3-day-old lar- development of the CNS. q 1998 John Wiley & Sons, Inc.vae, and in juvenile zebrafish. At 24–25 hpf, tn-w J Neurobiol 35: 1–16, 1998mRNA was expressed in the lateral plate mesoderm, Keywords: extracellular matrix; zebrafish; epidermalmost conspicuously in the presumptive sclerotome. growth factor–like repeats; fibronectin type III–likeMigrating cells of sclerotomal and neural crest origins repeats; neural crest
INTRODUCTION rect cell–cell contact, through gap junctions (forreview, see Bennett et al., 1991) or transmembranereceptors. To interact over longer distances, cellsCell–cell interactions have been shown to play cru-secrete recognition molecules that diffuse and bindcial roles in the development and preservation ofto receptors on distant target cells (Gingell, 1993).multicellular organisms (reviewed by Damsky etDiffusible recognition molecules may be depositedal., 1993; Truckwell et al., 1993; Vernadakis,into the extracellular matrix (ECM) and may set up1996). Interactions over short distances rely on di-gradients that influence cell migration and axonaloutgrowth (Hauzenberger et al., 1995; Varela-Echa-varria et al., 1997).Correspondence to: R. Bernhardt
q 1998 John Wiley & Sons, Inc. CCC 0022-3034/98/010001-16 Recognition molecules of the tenascin family are
1
8p3a 1935/ 8P3A$$1935 02-24-98 10:27:42 nbioa W: Neurobio

2 Weber et al.
Figure 1 Amino acid sequence and modular structure of the zebrafish TN-W protein. (A)Zebrafish TN-W sequence arranged in groups of homologous repeats. The hydrophobic regionis underlined by a bar (amino acids 12–26). All conserved cysteine residues (C) are highlightedwith filled triangles (cys-rich region) or asterisks (EGF domains) . The conserved tryptophan(W), leucine (L), and threonine (Y) residues used to align the FNIII domains are typed inbold. Numbers on the right indicate the amino acid positions, whereas numbers on the leftindicate the FNIII domains. (B) Schematic representation of the domain structure of zebrafishTN-W. The EGF domains (ellipses) , the FNIII domains (squares) , and the FG domain (hatchedcircle) are indicated.
secreted into the ECM. Three paralogs, distinct tad repeats, epidermal growth factor–like (EGF)repeats, fibronectin type III–like (FNIII) repeats,members of the family, have been identified so far
in mammals: tenascin-C (TN-C) (Bourdon et al., and a globular domain shared with fibrinogen (FG)(Erickson and Inglesias, 1984; Chiquet-Ehrismann1983; Erickson and Inglesias, 1984; Kruse et al.,
1985; Aufderheide et al., 1987; Faissner et al., et al., 1994; Chiquet-Ehrismann, 1995). All knownmembers of the tenascin family exist in several iso-1988; Garin-Chesa et al., 1989; Nishi et al., 1991),
tenascin-R (TN-R) (Kruse et al., 1985; Pesheva et forms owing to alternative splicing of FNIII repeats.TN-C and TN-R molecules can form multimers atal., 1989; Fuss et al., 1991; Carnemolla et al., 1996),
and tenascin-X (TN-X) (Morel et al., 1989; their cysteine-rich N-terminal part, which appears tobe stabilized by adjacent disulfide bridges (EricksonMatsumoto et al., 1992, 1994; Bristow et al., 1993;
Geffrotin et al., 1995). Orthologs, or species homo- and Inglesias, 1984; Pesheva et al., 1989; Norenberget al., 1992).logs, of mammalian TN-C are known in chicken
(Chiquet and Fambrough, 1984a, 1984b; Grumet et Tenascin-C, originally named myotendinous an-tigen (Chiquet and Fambrough, 1984a, 1984b), cy-al., 1985), amphibians (Onda et al., 1991), and fish
(Qiao et al., 1995; Tongiorgi et al., 1995a). Chicken totactin (Grumet et al., 1985), hexabrachion (Erick-son and Inglesias, 1984), and J1-200/220 (Kruse etalso possess a TN-R ortholog (Rathjen et al., 1991;
Norenberg et al., 1992, 1996) and yet another mem- al., 1985) is present in a large number of developingtissues including the nervous system (for review,ber of the tenascin family, TN-Y (Hagios et al.,
1996), which so far has no ortholog in other species. see Bartsch, 1996), but is absent from skeletal andheart muscles (Erickson and Bourdon, 1989). TN-Tenascins are large multimeric proteins con-
sisting of typical structural domains including hep- R, originally named janusin, J1-160/180, or re-
8p3a 1935/ 8P3A$$1935 02-24-98 10:27:42 nbioa W: Neurobio
Tenascin-W in Zebrafish 3
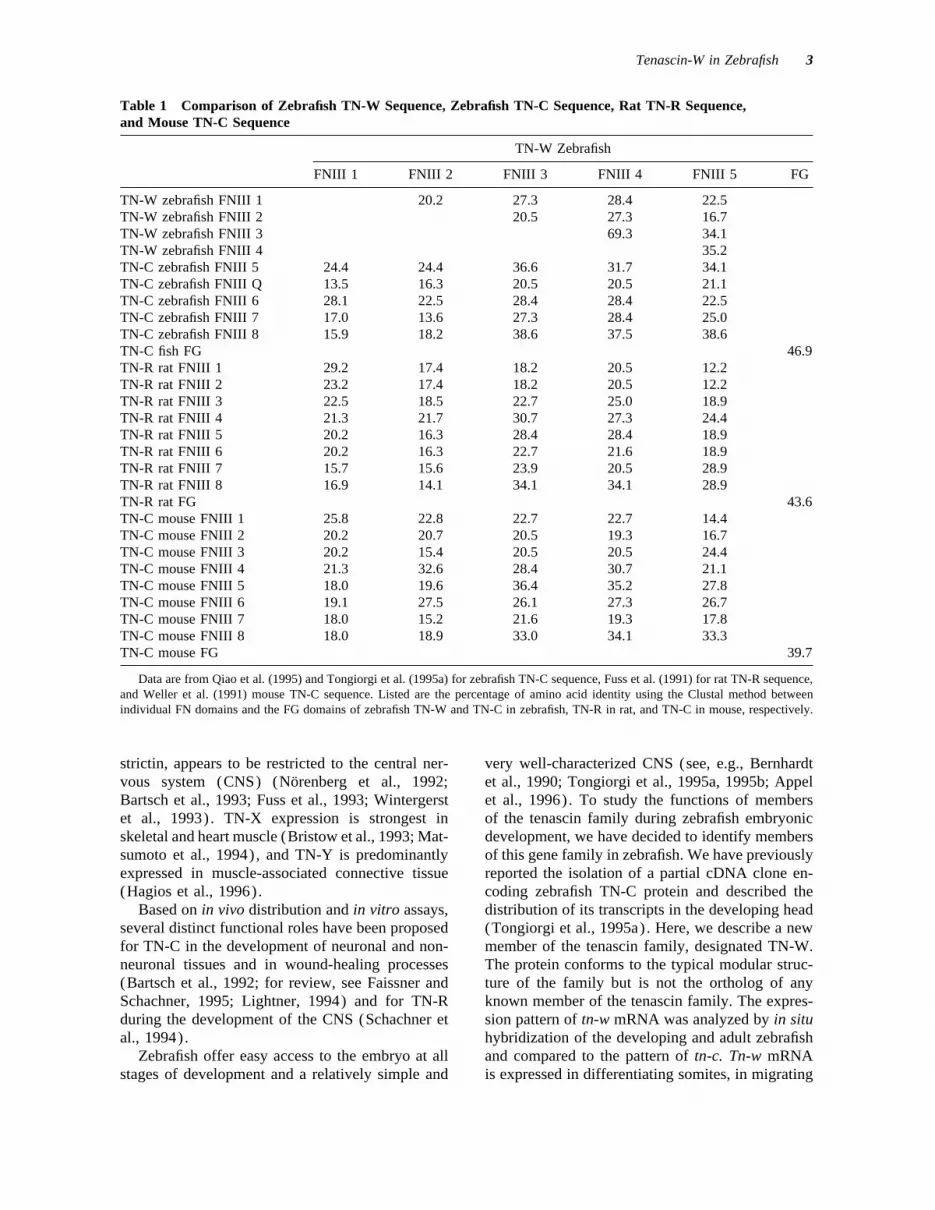
Table 1 Comparison of Zebrafish TN-W Sequence, Zebrafish TN-C Sequence, Rat TN-R Sequence,and Mouse TN-C Sequence
TN-W Zebrafish
FNIII 1 FNIII 2 FNIII 3 FNIII 4 FNIII 5 FG
TN-W zebrafish FNIII 1 20.2 27.3 28.4 22.5TN-W zebrafish FNIII 2 20.5 27.3 16.7TN-W zebrafish FNIII 3 69.3 34.1TN-W zebrafish FNIII 4 35.2TN-C zebrafish FNIII 5 24.4 24.4 36.6 31.7 34.1TN-C zebrafish FNIII Q 13.5 16.3 20.5 20.5 21.1TN-C zebrafish FNIII 6 28.1 22.5 28.4 28.4 22.5TN-C zebrafish FNIII 7 17.0 13.6 27.3 28.4 25.0TN-C zebrafish FNIII 8 15.9 18.2 38.6 37.5 38.6TN-C fish FG 46.9TN-R rat FNIII 1 29.2 17.4 18.2 20.5 12.2TN-R rat FNIII 2 23.2 17.4 18.2 20.5 12.2TN-R rat FNIII 3 22.5 18.5 22.7 25.0 18.9TN-R rat FNIII 4 21.3 21.7 30.7 27.3 24.4TN-R rat FNIII 5 20.2 16.3 28.4 28.4 18.9TN-R rat FNIII 6 20.2 16.3 22.7 21.6 18.9TN-R rat FNIII 7 15.7 15.6 23.9 20.5 28.9TN-R rat FNIII 8 16.9 14.1 34.1 34.1 28.9TN-R rat FG 43.6TN-C mouse FNIII 1 25.8 22.8 22.7 22.7 14.4TN-C mouse FNIII 2 20.2 20.7 20.5 19.3 16.7TN-C mouse FNIII 3 20.2 15.4 20.5 20.5 24.4TN-C mouse FNIII 4 21.3 32.6 28.4 30.7 21.1TN-C mouse FNIII 5 18.0 19.6 36.4 35.2 27.8TN-C mouse FNIII 6 19.1 27.5 26.1 27.3 26.7TN-C mouse FNIII 7 18.0 15.2 21.6 19.3 17.8TN-C mouse FNIII 8 18.0 18.9 33.0 34.1 33.3TN-C mouse FG 39.7
Data are from Qiao et al. (1995) and Tongiorgi et al. (1995a) for zebrafish TN-C sequence, Fuss et al. (1991) for rat TN-R sequence,and Weller et al. (1991) mouse TN-C sequence. Listed are the percentage of amino acid identity using the Clustal method betweenindividual FN domains and the FG domains of zebrafish TN-W and TN-C in zebrafish, TN-R in rat, and TN-C in mouse, respectively.
strictin, appears to be restricted to the central ner- very well-characterized CNS (see, e.g., Bernhardtet al., 1990; Tongiorgi et al., 1995a, 1995b; Appelvous system (CNS) (Norenberg et al., 1992;
Bartsch et al., 1993; Fuss et al., 1993; Wintergerst et al., 1996). To study the functions of membersof the tenascin family during zebrafish embryonicet al., 1993). TN-X expression is strongest in
skeletal and heart muscle (Bristow et al., 1993; Mat- development, we have decided to identify membersof this gene family in zebrafish. We have previouslysumoto et al., 1994), and TN-Y is predominantly
expressed in muscle-associated connective tissue reported the isolation of a partial cDNA clone en-coding zebrafish TN-C protein and described the(Hagios et al., 1996).
Based on in vivo distribution and in vitro assays, distribution of its transcripts in the developing head(Tongiorgi et al., 1995a). Here, we describe a newseveral distinct functional roles have been proposed
for TN-C in the development of neuronal and non- member of the tenascin family, designated TN-W.The protein conforms to the typical modular struc-neuronal tissues and in wound-healing processes
(Bartsch et al., 1992; for review, see Faissner and ture of the family but is not the ortholog of anyknown member of the tenascin family. The expres-Schachner, 1995; Lightner, 1994) and for TN-R
during the development of the CNS (Schachner et sion pattern of tn-w mRNA was analyzed by in situhybridization of the developing and adult zebrafishal., 1994).
Zebrafish offer easy access to the embryo at all and compared to the pattern of tn-c. Tn-w mRNAis expressed in differentiating somites, in migratingstages of development and a relatively simple and
8p3a 1935/ 8P3A$$1935 02-24-98 10:27:42 nbioa W: Neurobio

4 Weber et al.
cDNA Library
The lambda ZAP II zebrafish (20–28 hpf) polyA/ cDNAlibrary with a complexity of more than 1.0 1 107 recom-binants was provided by Dr. D. J. Grunwald (Universityof Utah, Salt Lake City) .
Cloning and Sequencing
The 444-bp-long cDNA insert of clone pGEX-EGF/S(Xiao et al., 1996), which encodes the 4.5 EGF repeatsof rat TN-R (Fuss et al., 1991), was radioactively labeled(Random Priming Kit; Boehringer Mannheim) and usedas a probe to screen the cDNA library (as described bySambrook et al., 1989). Replica filters (Pall; Biodyne)were hybridized under low-stringency conditions at 427Cin 30% formamide, 61 SSPE, 51 Denhardt’s solution,0.5% sodium dodecyl sulfate (SDS), and 100 mg/mLsonicated herring sperm DNA (Boehringer Mannheim).Filters were washed three times for 20 min at 427C in11 SSPE, 0.2% SDS, and exposed to autoradiography.Positive clones were purified by repeated plating at lowdensity and further screening. Single plaques of positivelambda clones were further amplified. Lambda-DNA wasisolated (QIAGEN) and subcloned into pBluescript KS0(Stratagene).
To obtain the 5* part of zebrafish tn-w cDNA, rapidamplification of cDNA ends (RACE) was performedwith a 5 * RACE Kit (Boehringer Mannheim) according
Figure 2 Northern blot analysis of the expression of tn- to the manufacturer’s instructions on total RNA isolatedw and tn-c mRNAs at early and late larval stages. RNA from 4-day-old zebrafish (RNeasy Mini Kit; QIAGEN).from 3-day-old (1, 3) and 14-day-old zebrafish (2, 4) Reverse transcription (AMV reverse transcriptase; Boeh-was hybridized with a 1795-bp cDNA probe specific for ringer Mannheim) of total RNA was performed usingthe EGF, all FNIII, and a part of the FG domain-coding primer SP1 (5 * CTTTGGATGGTAGGACAGGA 3 * ;sequence of the zebrafish tn-w gene (lanes 1 and 2), or annealing temperature 557C, 30 cycles) . The purifiedthe 1.1-kb fragment of the tn-c zebrafish cDNA subclone (High Pure PCR Product Purification Kit; Boehringer9 (Tongiorgi et al., 1995a) (lanes 3 and 4). The size of Mannheim) primary transcript was amplified using ana RNA marker in kilobases is indicated at the left margin. oligo dT-anchor primer and the nested primer SP2
(5* AGTATCGGCTCCTCTCACAG 3 * ; annealing tem-perature 607C, 30 cycles) . The resulting fragment wassubcloned into pBluescript KS0. The nucleotide se-cells of neural crest and sclerotome origin, and inquences were determined by sequencing from bothcells contributing to the skeleton.strands with the T7-Sequencing Kit (Pharmacia) usingsynthetic oligonucleotides as primers. Sequence analysiswas performed with DNA Strider 1.2 and the proteinalignment of the deduced amino acid sequence was gener-MATERIALS AND METHODSated with MegAlign (DNA Star) using the Clustalmethod.
Animals
Zebrafish embryos were collected from our breeding col- RNA Preparation and Northernony, maintained according to standard procedures (West- Blot Analysiserfield, 1993) at 28.57C. After hatching, larvae were keptat 28.57C in small breeding tanks where they developed Total RNA from 3- and 14-day-old zebrafish was isolated
using the RNeasy Kit (QIAGEN). RNA was separatedinto adult fish. The histological analysis focused on threeage groups: (a) embryos at 24–25 h postfertilization by electrophoresis in a 1.5% agarose gel containing 7%
formaldehyde and transferred onto Hybond-N mem-(hpf); (b) 3-day-old, recently hatched larvae; and (c)juvenile fish 10–15 mm in body length (age approxi- branes (Amersham). Hybridization was carried out with
the following 32P-labeled cDNA probes labeled to 108mately 6–8 weeks) .
8p3a 1935/ 8P3A$$1935 02-24-98 10:27:42 nbioa W: Neurobio

Tenascin-W in Zebrafish 5
cpm/mg by random priming (Feinberg and Vogelstein, terminated by a stop codon at position 2923 and1983): a 1795-bp EcoRI fragment of the tn-w zebrafish encodes a protein of 932 amino acid residues withcDNA encoding the 3.5 EGF, 5 FNIII repeats, and part a predicted molecular mass of 132 kD. The stopof the FG domain and the 1.1-kb fragment of the tn-c codon is followed by 606 bp of 3 * untranslatedzebrafish cDNA subclone 9 (Tongiorgi et al., 1995a). sequence, a polyadenylation signal at position 3532,
and a polyA/ stretch.In Situ Hybridization
Analysis of the Protein StructureDigoxigenin uridine triphosphate (Boehringer Mann-of TN-Wheim) –labeled riboprobes were synthesized by in vitro
transcription as described (Doerries et al., 1993), usingThe analysis of the predicted amino acid sequencethe 1795-bp EcoRI fragment of a tn-w zebrafish cDNAof TN-W shows that the protein consists of differentsubclone coding for 3.5 EGF repeats, all 5 FNIII repeats,domains [Fig. 1(A)] . The hydrophobicity profileand part of the FG domain [Fig. 1(A)] , or the 1.1-kb(Kyte and Doolittle, 1982) indicates a hydrophobicfragment of the tn-c zebrafish cDNA subclone 9 (Tongi-region at the amino terminus (residues 11–26) [Fig.orgi et al., 1995a). In situ hybridization on embryonic1(A)] . Residue 27 is predicted to be the first aminowhole mounts and on cryosections of fresh-frozen tissue
was performed as described previously (Appel et al., acid of the mature protein according to the method1996). In addition, some embryos reacted for in situ of von Heijne (1988). The putative signal peptidehybridization as whole-mount preparations were subse- is followed by an amino terminal region (aa 27–quently embedded in a water-soluble plastic-embedding 146) which contains eight cysteines [Fig. 1(A)] inmedium (Durcupan; Fluka, Switzerland) and semithin- a spacing similar to that in TN-C and TN-R (Chi-sectioned (Tongiorgi et al., 1995b). Photomicrographs
quet-Ehrismann, 1990; Norenberg et al., 1992; Fusswere taken with a Zeiss Axiophot microscope equippedet al., 1993). The cysteine-rich segment of TN-Wwith Nomarski optics.may allow the assembly of monomers intomultimers, as has been shown for TN-C and TN-R(Erickson and Inglesias, 1984; Spring et al., 1989;RESULTSNorenberg et al., 1992; Fuss et al., 1993). One in-complete EGF segment of 12 amino acids and threeIsolation of a Zebrafish cDNA Clonecomplete EGF repeats of 31 amino acids each (aaEncoding a New Member of the147–251) follow the cysteine-rich stretch. The EGFTenascin Familyrepeats are followed by five FNIII repeats (aa 252–698) with characteristically conserved tryptophan,A 20- to 28-hpf zebrafish cDNA library was
screened for tenascin-related molecules using a leucine, tyrosine, and threonine residues, and a do-main homologous to fibrinogen (aa 699–932).probe encoding the 4.5 EGF domains of rat TN-R
(Fuss et al., 1991). Screening of 6 1 105 recombi- These are all typical modular elements of the mole-cules belonging to the tenascin family [Fig. 1(B)] .nants using low-stringency hybridization yielded 10
positive clones. Four of these contained sequencesrelated to tenascin. The inserts of three of the clones Analysis of the Relationship of TN-Wwere identical and also contained in the fourth to Other Tenascin Proteinsclone. The analysis of this latter clone revealed apartial cDNA encoding a novel member of the ten- Comparison of the entire deduced amino acid se-
quence of TN-W with the sequences of the otherascin family. To obtain the missing 5* part of thiscDNA, a 5 * RACE approach was applied to total known members of the tenascin family demon-
strates relatively low degrees of homology. TheRNA isolated from 4-day-old zebrafish. In a dupli-cate set of experiments, a cDNA fragment of 890 highest homology (35.9% amino acid identity) is
observed to the partial sequence of zebrafish TN-bp was amplified encoding the amino terminal endand 126 bp of the 5* untranslated sequence. The C, which encompasses the most 3* 4.5 FNIII do-
mains. The comparison to the entire sequences ofcombined sequence represents a cDNA for tn-w of3591 bp (European Molecular Biology Laboratories TN-R of rat and chicken reveals 31.7% and 31.1%
identity, respectively. Homology to mouse TN-C isaccession no. AJ001423). The ATG codon at nucle-otide position 127 is flanked by the most conserved 29.1%, to human TN-C 21.4%, to mouse TN-X
20.3%, to chicken TN-Y 20.3%, and to human TN-nucleotides of the consensus sequence for transla-tion initiation sites in eukaryotes (Kozak, 1991). X 15.5%. These homologies are considerably lower
than those found between zebrafish TN-C and itsThe following open reading frame of 2796 bp is
8p3a 1935/ 8P3A$$1935 02-24-98 10:27:42 nbioa W: Neurobio
6 Weber et al.
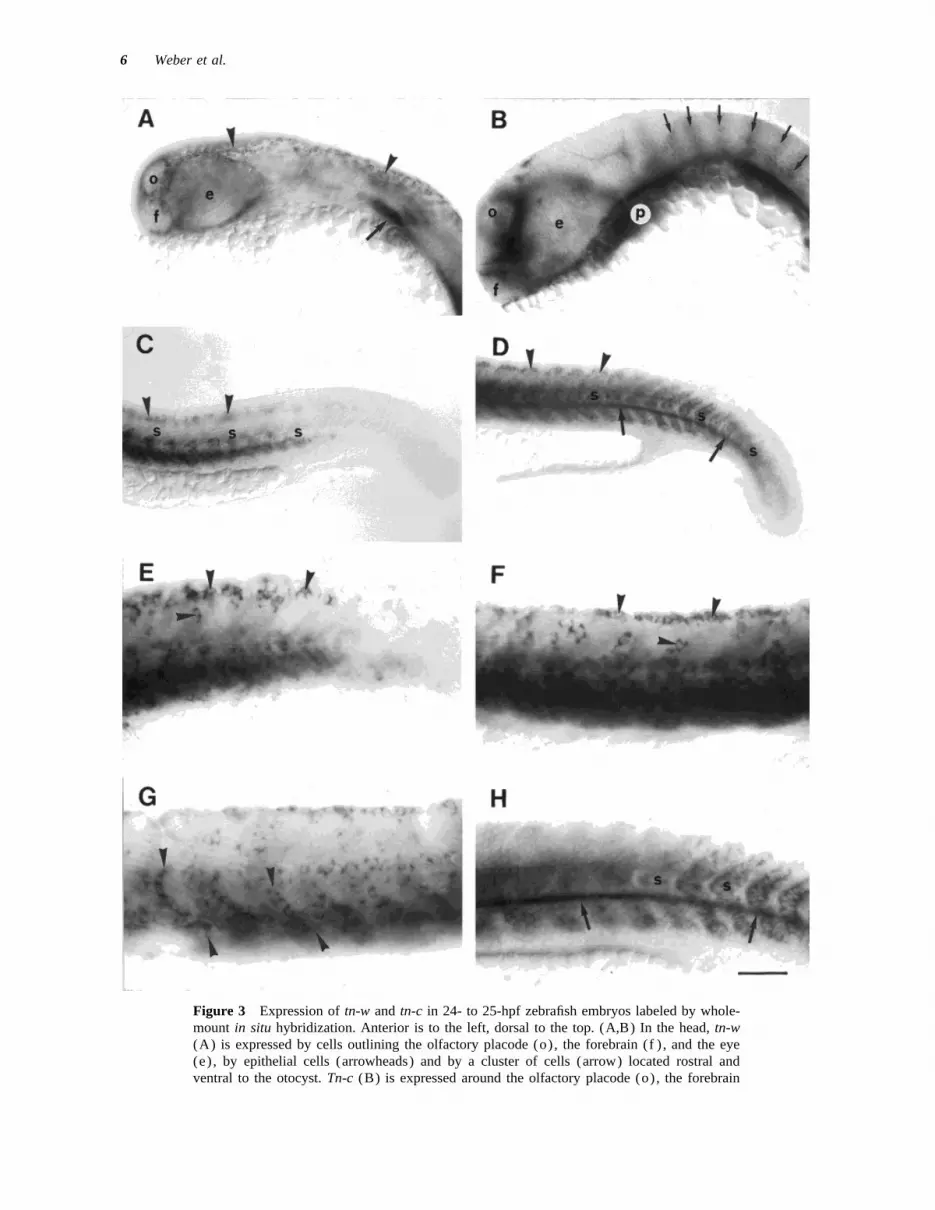
Figure 3 Expression of tn-w and tn-c in 24- to 25-hpf zebrafish embryos labeled by whole-mount in situ hybridization. Anterior is to the left, dorsal to the top. (A,B) In the head, tn-w(A) is expressed by cells outlining the olfactory placode (o), the forebrain (f ) , and the eye(e) , by epithelial cells (arrowheads) and by a cluster of cells (arrow) located rostral andventral to the otocyst. Tn-c (B) is expressed around the olfactory placode (o), the forebrain
8p3a 1935/ 8P3A$$1935 02-24-98 10:27:42 nbioa W: Neurobio

Tenascin-W in Zebrafish 7
orthologs in chicken or mammals (57.3–60.5%) rat TN-R, and 39.7% to mouse TN-C) (Table 1).The FG domain is the most conserved module of(Tongiorgi et al., 1995a). Other recognition mole-
cules in the zebrafish and their orthologs in mam- the tenascin molecules (Erickson, 1994). The TN-C orthologs in fish and human show an amino acidmals and chicken show levels of amino acid identi-
ties that vary between approximately 40% and 60% identity of 71.4%. This comparison also argues thatTN-W is not the homolog of any other known tenas-(e.g., L1) (Tongiorgi et al., 1995b), (e.g., N-CAM)
(Bernhardt et al., 1996). cin molecule.The notion that TN-W is not directly related to
any other known tenascin is strengthened by the Northern Blot Analysissequence alignment of the EGF repeats of TN-Wwith those of other tenascin molecules. Correspond- Total RNA from 3- and 14-day-old zebrafish was
subjected to Northern blot analysis. After hybridiza-ing EGF repeats tend to be highly conserved be-tween the orthologs of each of the different tenascin tion with a 1795-bp cDNA probe corresponding to
all EGF, all FNIII, and part of the FG domain ofproteins (e.g., 73.0–94.6% for TN-C in mammalsand in chicken; not shown). In contrast, it was not zebrafish tn-w, a strong signal of about 4.3 kb was
detectable with total RNA from 3-day-old zebrafish,possible to identify EGF repeats in TN-W that obvi-ously corresponded to particular repeats in other and to a much lower extent with total RNA from
14-day-old zebrafish (Fig. 2) . In the preparationtenascins. However, the cysteines, presumably im-portant for the protein structure (Chiquet-Ehris- from 3-day-old larvae, the tn-w mRNA formed a
smear. This likely reflects a large amount of mRNAmann, 1990), are absolutely conserved.The individual FNIII domains of TN-W are also loaded and, possibly, some degradation. In the prep-
aration from 14-day-old larvae, less tn-w mRNAnot closely related to any particular FNIII domain inother members of the tenascin family. The maximal was present and only the band at 4.3 kb was visible,
arguing against the existence of additional relatedamino acid identity is 38.6% between FNIII do-mains three and five of zebrafish TN-W and FNIII messages. After hybridization with a 1.1-kb cDNA
fragment of zebrafish tn-c (Tongiorgi et al., 1995a),domain eight of zebrafish TN-C (Table 1). Thelow levels of identity do not allow us to determine a signal of about 6.0 kb was detectable with total
RNA from 3-day-old zebrafish, whereas no signaldefinitively whether the linear arrangement of theFNIII domains of TN-W is similar to that of other was detectable with total RNA from 14-day-old ze-
brafish (Fig. 2) . The absence of cross-reactivity be-tenascin molecules. As shown in Table 1, a compar-ison of the five FNIII domains of TN-W and the tween the two probes demonstrates that they recog-
nize two distinct mRNAs.eight constitutively expressed FNIII domains ofrat TN-R and mouse TN-C may indicate a similar-ity in their order. We also compared the amino Expression of tn-w mRNA in theacid sequences between the five FNIII domains Zebrafishwithin TN-W. Domains three and four show highamino acid identity (69.3%), suggesting the dupli- We analyzed the expression pattern of tn-w in 1-
day-old zebrafish embryos, in 3-day-old larvae, andcation of a single ancestral domain. The otherthree domains are not highly related to each other in juvenile zebrafish. Below, we compare this pat-
tern to that of tn-c with which tn-w shares some(Table 1).The FG domain of TN-W also shows only rela- structural and sequence homologies (see above),
suggesting the possibility that tn-w and tn-c maytively low amino acid identity to the FG domainsof other tenascins (46.9% to fish TN-C, 43.6% to serve similar functions. For a detailed description
(f ) , and the eye (e) , in the paraxial mesoderm (p) and at the rhombomere boundaries (arrows).(C,D) In the trunk and tail, tn-w (C) is expressed in the ventral somites (s) shortly aftersegmentation and also by presumptive neural crest cells along the dorsal midline (arrowheads) .Tn-c (D) is expressed throughout the somites (s) prior to segmentation, in the hypochord(arrows), and neural crest (arrowheads) . (E,F) Tn-w (E) and tn-c (F) are expressed by neuralcrest cells (arrowheads) . (G,H) Tn-w (G) is expressed by presumptive sclerotome cells whichextend in single file from the ventral edge of the somite toward the horizontal myoseptum(arrowheads) . Tn-c (H) is expressed throughout the somite (s) and in the hypochord (arrows).Scale bar: (A–D) Å 100 mm; (E–H) Å 5 mm.
8p3a 1935/ 8P3A$$1935 02-24-98 10:27:42 nbioa W: Neurobio

8 Weber et al.
of the expression of tn-c in the embryonic head andtrunk, the reader is also referred to Tongiorgi et al.(1995a).
In situ hybridization for tn-w in whole-mountpreparations of 24- to 25-hpf embryos revealed ex-pression in the head and in the trunk (Fig. 3) . Inthe head, the mRNA was detected in cells sur-rounding the olfactory placodes and the forebrain,and in large cells scattered throughout the epidermis[Fig. 3(A)] . The tn-w –expressing epidermal cellswere confined to the head and most anterior trunk,but were not found more posteriorly [compare Fig.3(A) and 3(C)] . Just ventral and slightly anteriorto the otocyst, a bilateral cluster of densely packedcells, possibly destined to contribute to the gillarches (Kimmel et al., 1995), was strongly labeledby the tn-w probe [Fig. 3(A)] .
The expression pattern of tn-c in the embryonichead (Tongiorgi et al., 1995a) differed in severalaspects from that of tn-w. Expression of tn-c mRNAwas detected in the brain, concentrated at the rhom-bomere boundaries of the hindbrain [Fig. 3(B)]and in three distinct domains of the fore- and mid-brain (not shown) (see Tongiorgi et al., 1995a).Tn-c, in contrast to tn-w, was expressed in paraxialmesoderm, which may contribute to the craniumand pharyngeal arches (Schilling and Kimmel,
Figure 4 Expression of tn-w and tn-c in the trunk of1994; Tongiorgi et al., 1995a) but not in epidermal24- to 25-hpf embryos labeled by in situ hybridizationcells [Fig. 3(B)] . The only overlap in the expres-on cryosections (A,B) or by whole-mount in situ hybrid-sion of tn-c and tn-w mRNAs in the head was inization followed by plastic embedding and semithin sec-
the cluster of cells probably contributing to the gilltioning (C,D). In (A,B), the spinal cord (sc) and noto-
arches [compare Fig. 3(A) and 3(B)] and in cells chord (n) are indicated. (A,C) Tn-w is expressed by neu-surrounding the olfactory placodes and the fore- ral crest cells in both the medial ( large arrowheads) andbrain. The latter have previously been found to be the lateral (small arrowhead) migratory pathways, theof neural crest origin and to contribute to the forma- sclerotome (arrows), and the somite (s) . (B,D) Tn-c istion of the cartilage of the head skeleton (Tongiorgi expressed by neural crest cells (arrowheads) , and the
hypochord (h), in the somite, concentrated around theet al., 1995a).prospective horizontal myoseptum (broken bar) , and inIn the trunk of 24- to 25-hpf embryos labeled bythe sclerotome (arrows). Expression is also detected inwhole-mount in situ hybridization, expression of tn-the floor plate region of the spinal cord (asterisks) .w was detected in the somites, including the sclero-Scale bar: in (B) Å 50 mm for (A,B); in (D) Å 20 mmtome, and in neural crest cells [Fig. 3(C,E,G)] . Thefor (C,D).
expression levels in the somitic mesoderm showed apronounced caudal to rostral gradient, reflecting thecontinuing addition of new somites to the caudal (Morin-Kensicki and Eisen, 1997) were located at
the level of the notochord and more ventrally, oftentrunk: tn-w was not expressed in the unsegmentedmesoderm of the tail. In the most recently formed aligned to form segmentally repeated bands which
extended dorsoventrally [Fig. 3(G)] .somites of the caudal trunk, expression levels werelow. The older somites of the midtrunk region ex- Expression of tn-c was also detected in the meso-
derm and in neural crest cells [Fig. 3(D,F,H)] .pressed high levels of tn-w mRNA [Fig. 3(A)] .Presumptive neural crest cells (Raible et al., 1992) However, while tn-w expression was restricted to
the somitic mesoderm derived from the lateral plate,were labeled in small, segmentally reiterated clus-ters near the dorsal edge of the spinal cord and expression of tn-c was very prominent in a deriva-
tive of the axial mesoderm, the hypochord [Fig.scattered more ventrally at midspinal cord level[Fig. 3(C,E)] . Presumptive sclerotome cells 3(D,H)]. Hypochord cells are arranged in a single
8p3a 1935/ 8P3A$$1935 02-24-98 10:27:42 nbioa W: Neurobio

Tenascin-W in Zebrafish 9
Figure 5 Expression of tn-w and tn-c in the heads of 3-day-old zebrafish larvae. In situhybridization on cryosections. The dark appearance of pigment cells (some indicated by whitedots) in the skin and retina is due to pigment, not staining of mRNA. (A) Tn-w expression isrestricted to skeletal elements (arrowheads) in the forming skull and the jaw and to the cornea(c) . (B) Tn-c is expressed by skeletal elements in the jaw (arrowheads) , the brain (b) , andretina (asterisks) . Scale bar: in (B) Å 100 mm for (A,B).
row, just ventral to the notochord and extending On comparable cross sections hybridized withthe tn-c probe, strong labeling was detected in somi-from the tail bud to the head. These cells may later
become incorporated into the descending aorta tic cells concentrated adjacent to the notochord, inthe region of the horizontal myoseptum [Fig.which forms ventral to the notochord (R. R. Bern-
hardt, unpublished observations) . In the somitic 4(B,D)] . The sections also confirmed the promi-nent expression of tn-c by the hypochord [Fig.mesoderm, expression of tn-c also showed a caudal
to rostral gradient: expression was relatively low in 4(B)] and some expression by presumptive sclero-tome cells [Fig. 4(D)] . In addition, they revealedthe tail bud and increased in the transition zone from
unsegmented to segmented mesoderm. Expression tn-c expression by cells located at the ventral mid-line of the spinal cord. These are the midline floorwas maximal in the midtrunk somites [Fig. 3(D)]
and decreased again in more rostral somites (not plate cells and lateral cells that have previously beendefined as lateral floor plate cells (Bernhardt et al.,shown). Neural crest cells labeled by the tn-c
probe showed a distribution pattern similar to that 1992). The tn-c transcript levels in the floor platecells were relatively low and more readily detectedobserved after hybridization with the tn-w probe
[compare Fig. 3(D,F) to 3(C,E)] . Presumptive in the relatively thick cryosections than in semithinsections [Fig. 4(B,D)] .sclerotome cells also expressed tn-c, although
less conspicuously than tn-w [compare Fig. 3(F,H) The examination of the cross sections also con-firmed that some of the cells expressing tn-w andto 3(E,G)] .
Cross sections through 24- to 25-hpf embryos at tn-c were located in the migratory pathways of neu-ral crest and sclerotome cells. Neural crest cellsthe anterior and midtrunk levels, both of fresh-fro-
zen tissue processed for in situ hybridization [Fig. migrate along a medial (at the interface of somiteand spinal cord) or a lateral (subepidermal) path-4(A)] and of embryos sectioned only after whole-
mount staining and plastic embedding [Fig. 4(C)] , way (Raible et al., 1992); sclerotome cells migratein the medial pathway (Morin-Kensicki and Eisen,showed that tn-w expression levels were highest in
bilateral clusters of cells located ventral to the noto- 1997). Therefore, labeled cells at the level of thenotochord may be sclerotomal, while those locatedchord. These clusters appeared segmentally reiter-
ated in the posterior halves of each somite and are dorsolaterally to the spinal cord are undoubtedly ofneural crest origin. Tn-w –expressing neural cresttherefore not equally obvious in all cross sections
[Fig. 4(A,C)] . On the basis of their distribution, cells were detected in both pathways, but tn-c–ex-pressing cells appeared confined to the medial path-they can be identified as sclerotome (Morin-Ken-
sicki and Eisen, 1997). Low levels of tn-w tran- way [compare Fig. 4(C) and 4(D)].Sections through the heads of 3-day-old larvaescripts were observed in cells located laterally in
the somite, while no expression was detected in the (Fig. 5) showed that the expression patterns ob-served in whole-mount preparations of 24- to 25-spinal cord.
8p3a 1935/ 8P3A$$1935 02-24-98 10:27:42 nbioa W: Neurobio
10 Weber et al.
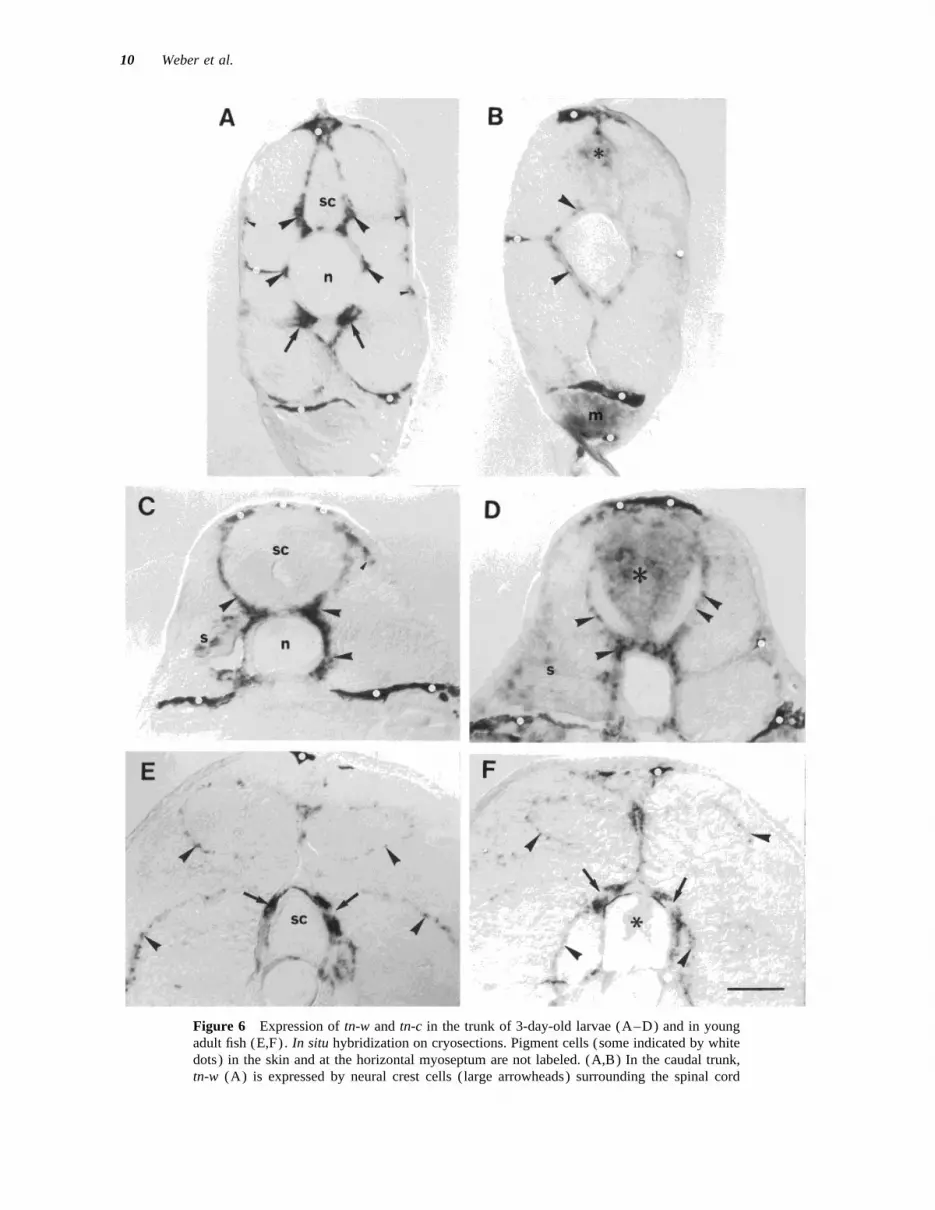
Figure 6 Expression of tn-w and tn-c in the trunk of 3-day-old larvae (A–D) and in youngadult fish (E,F). In situ hybridization on cryosections. Pigment cells (some indicated by whitedots) in the skin and at the horizontal myoseptum are not labeled. (A,B) In the caudal trunk,tn-w (A) is expressed by neural crest cells ( large arrowheads) surrounding the spinal cord
8p3a 1935/ 8P3A$$1935 02-24-98 10:27:42 nbioa W: Neurobio

Tenascin-W in Zebrafish 11
hpf embryos had not fundamentally changed. Tn-w mRNAs were decreased compared to the larvae.The exception were cells in the dorsal root gangliawas expressed by cells in the forming skull capsule
and in the forming jaw. Cells in the epidermis over- (DRGs), derivatives of the neural crest, that ex-pressed high levels of tn-w and tn-c. These cellslying the skull and in the cornea were also labeled
by the tn-w probe [Fig. 5(A)] . Tn-c, in contrast to have not been identified definitively, though theirrelatively small size suggests that they may be non-tn-w, was expressed in the brain and retina [Fig.
5(B)] (Bernhardt et al., 1996). While cells in the neuronal. In the somites, expression of both mRNAswas restricted to mesenchymal cells of the segmen-jaw also expressed tn-c, there was no conspicuous
expression in the forming skull. Tn-c could not be tal myosepta [Fig. 6(E,F)] . No other structuresshowed detectable levels of expression.detected in the epidermis or cornea.
In situ hybridization on cryosections throughthe trunk of 3-day-old larvae also showed expres-sion patterns of tn-w and tn-c that were very simi- DISCUSSIONlar to those in the 24- to 25-hpf embryos, exceptthat expression levels in the somites were mark- We describe a new tenascin-like molecule in zebra-
fish, TN-W. It is clearly different from TN-C, whichedly decreased. Tn-w continued to be expressedprominently by cells located at the interface of was previously identified in zebrafish and which
is the ortholog of TN-C in mammals and chickenthe somite and spinal cord and notochord [Fig.6 (A,C) ] . Some labeled cells were also detected (Tongiorgi et al., 1995b). TN-W is composed of
3.5 EGF repeats, 5 FNIII domains, and a domainunderneath the epidermis, in the lateral path ofneural crest migration [Fig. 6 (A,C) ] . At the me- homologous to fibrinogen. While these modules are
typical for all members of the tenascin family, nodial surface of the somite and just ventral to thenotochord, there were strongly labeled scleroto- known tenascin contains as few of them as TN-W.
Because the protein differs in its structure from allmal cells [Fig. 6 (A)] .Weak tn-c expression was still detectable in pre- other tenascins, and because it shows only very low
levels of homology to any known tenascin, TN-Wsumptive neural crest and sclerotomal cells adjacentto the spinal cord and notochord, but there was no is considered a novel member of the tenascin family.
While the other tenascins contain several EGFevidence of neural crest cells labeled in the lateralpath of migration [Fig. 6(B)] . Tn-c was also ex- and FNIII repeats that are highly homologous to
each other, only FNIII repeats three and four of TN-pressed ventrally in the tail, caudal to the cloaca,in the intermediate mass mesoderm [Fig. 6(B)] . W are clearly related to each other. The similarities
of individual domains are probably due to duplica-The hypochord was no longer detected. Tn-c contin-ued to be expressed in the spinal cord. In the caudal tions of sequences derived from an ancestral gene.
These duplications are thought to have occurredspinal cord, expression was now by cells locatedmainly in the dorsal grey matter and no longer by during the evolution of the tenascin gene family
and to have given rise to the increased structuralcells at the ventral midline [Fig. 6(B)] . More ante-riorly, cells throughout the spinal grey matter ex- complexity (Hagios et al., 1996). Because only one
such duplication is apparent in TN-W, it appearspressed tn-c [Fig. 6(D)] .In situ hybridization on cryosections through ju- possible that its structure may be reminiscent of the
hypothetical ancestor.venile fish showed that expression levels of both
(sc) , by presumptive neural crest cells located subepidermally (small arrowheads) , and bysclerotome cells (arrows). Labeled cells (arrowheads) at the notochord (n) level may be ofneural crest or sclerotome origin. Tn-c (B) is expressed by neural crest cells and/or sclerotomalcells surrounding the notochord (arrowheads) , by cells in the dorsal spinal cord (asterisks) ,and by cells in the intermediate mass mesoderm (m). (C,D) In the anterior trunk, tn-w (C) isexpressed by presumptive neural crest and/or sclerotome cells (arrowheads) surrounding thespinal cord and notochord, and by a few cells in the somite (s) . Tn-c (D) is expressed bypresumptive neural crest cells and sclerotomal cells (arrowheads) , and weakly in the somite(s) . Expression is prominent in the spinal cord (asterisks) . (E,F) In the trunk of juvenile fish,tn-w (E) and tn-c (F) are expressed in the septal mesenchyme (arrowheads) and in the dorsalroot ganglia (arrows). Very weak expression of tn-c is detected in the spinal cord (asterisks) .Scale bar: in (F) Å 50 mm for (A–D) and 100 mm for (E,F).
8p3a 1935/ 8P3A$$1935 02-24-98 10:27:42 nbioa W: Neurobio

12 Weber et al.
The expression patterns of the mRNAs encoding Eisen, 1994; Jesuthasan, 1996). It is unclear howneural crest cells distinguish between the two path-TN-W and TN-C suggest that both are of impor-
tance primarily during early stages of development. ways which converge near the dorsal midline. Inthe chicken, a role of the dermamyotome has beenNorthern blot analysis indicates the down-regula-
tion of both mRNAs during larval development. suggested (Oakley et al., 1994), and in zebrafishthe early lateral somite appears to repel neural crestThis is confirmed by in situ hybridization, which
shows that the expression of both tn-w and tn-c is cells (Jesuthasan, 1996). The observation that tn-w but not tn-c is expressed by presumptive neuralmost prominent in embryos and early larvae but is
reduced in juvenile fish. In situ hybridization dem- crest cells in the lateral pathway indicates that theexpression of a specific combination of ECM mole-onstrates an overlap of the expression of the two
mRNAs in some cell populations, consistent with cules may contribute to the choice of one or theother pathway.the possibility that the two molecules may serve
similar functions. In other cell populations, how- A role of ECM molecules in neural crest cellmigration is implied by experiments that interfereever, only tn-w or tn-c mRNA is expressed, indica-
tive of specific roles of the two proteins. with the best-studied class of receptors for ECMcomponents, the integrins (Lallier and Bronner-Fra-Both tn-w and tn-c are expressed in the neural
crest and in the sclerotome and by presumptive mi- ser, 1993). In the case of TN-C, the hypothesis ofits role in neural crest migration is supported bygratory cells derived from the two cell populations.
There are some subtle differences, however. Expres- antibody perturbation experiments in the chicken(Bronner-Fraser, 1988; Halfter et al., 1989). TN-Csion of tn-w by sclerotomal cells appears to be at
higher levels and for a longer time period than that may contribute to cell migration, including neuronalmigration (Chuong et al., 1987; Husmann et al.,of tn-c. Presumptive neural crest cells located sub-
epidermally appear to express tn-w but not tn-c. The 1992), by influencing cell shape, an effect that ismodified by fibronectin and a chondroitin sulphatemajor migratory route of neural crest and sclero-
tome cells is defined by lineage studies showing proteoglycan to which TN-C has been reported tobind (Hoffman and Edelman, 1987; Hoffman et al.,that in the zebrafish trunk, neural crest cells delami-
nate from the dorsal aspect of the spinal cord and 1988; Chiquet-Ehrismann et al., 1988; Faissner etal., 1990). These functions could not be identifiedstart to migrate ventrally along a medial pathway
at the interface of somite and spinal cord/notochord so far by experiments in which the expression ofTN-C was mutated by inactivation of its gene inat around 16 h of development. The cells will con-
tribute to the peripheral nervous system (Raible et mice (Saga et al., 1992; Forsberg et al., 1996),probably owing to compensatory mechanisms.al., 1992; Raible and Eisen, 1994; Jesuthasan,
1996). Sclerotome cells originate at the ventrome- Should a mammalian homolog of TN-W exist, thenit would appear possible that such a molecule coulddial edge of the somite and migrate dorsally along
the same medial pathway as the neural crest cells, have substituted for some functions of the missingTN-C, at least in the neural crest cells.starting also around 16 h of development (Morin-
Kensicki and Eisen, 1997). These studies have left Cells in the pharyngeal arches and in the neuro-cranium were also found to express tn-w and tn-c.it unclear for how long the migration of the two
cell populations continues. The finding of tn-w – Again, the patterns of expression, while similar,were not completely identical. Paraxial mesodermand tn-c–expressing cells in the migratory pathways
in 3-day-old larvae indicates that the migration con- in the region between the eye and the otocyst whichcontributes mesenchyme to the pharyngeal archestinues at least until the third day of development.
This conclusion is also supported by the observation (Schilling and Kimmel, 1994) expressed tn-c butnot tn-w. Expression levels of tn-c were conspicu-that identified spinal ganglia are first detected at this
time (Bernhardt et al., 1990) or even later (Shul- ously down-regulated by cells in the forming skullof 3-day-old fish, as opposed to the prominent ex-man-Weis, 1968). The expression patterns of tn-w
and tn-c in the trunk are consistent with the hypothe- pression of tn-w. Despite these differences, the ex-pression of both mRNAs by cells contributing tosis that their deposition in the ECM may contribute
to define the medial migratory pathway. the skeleton suggests that another common functionof the two proteins may be in cartilage formation,A second, lateral pathway of neural crest cell
migration exists at the interface of epidermis and as previously proposed for TN-C (Vaughan et al.,1987; Mackie et al., 1987; Tongiorgi et al., 1995a).somite. Prospective pigment cells enter this pathway
4–5 h after the first neural crest cells have entered The two molecules may have different functionsin the differentiation of the somitic mesoderm. Inthe medial pathway (Raible et al., 1992; Raible and
8p3a 1935/ 8P3A$$1935 02-24-98 10:27:42 nbioa W: Neurobio

Tenascin-W in Zebrafish 13
addition to its very prominent expression in the pre- sites of expression. Antibody perturbation experi-ments and manipulations at the mRNA level maysumptive sclerotome, tn-w is expressed at lower lev-
els throughout the somite, particularly in the lateral allow us to define the roles of tn-c and tn-w inzebrafish development.regions. In contrast, tn-c is most strongly expressed
in the vicinity of the forming horizontal myoseptumwhere the first muscle cells, the muscle pioneers, The authors thank Robert Riggleman, Kathryn Helde,differentiate (Felsenfeld et al., 1991). Different re- and David Jonah Grunwald for providing the zebrafishgions of the somitic mesoderm in fish, as in other cDNA library; Tiziana Flego for technical assistance; and
Kathrin Mannigel for animal care.vertebrates, differ in their responses to signals thatregulate mesoderm differentiation (e.g., van Eedenet al., 1996, and references therein) . The signalsthat govern the regional differentiation originate at REFERENCESleast in part from the notochord and involve signal-ing by the transcription factor sonic hedgehog
APPEL, C., GLOOR, S., SCHMALZING, G., SCHACHNER, M.,(Johnson et al., 1995). In the chicken, this resultsand BERNHARDT, R. R. (1996). Expression of the
in the differential activation of specific transcription Na,K-ATPase beta 3 subunit during development offactors of the pax family (Johnson et al., 1995). the zebrafish central nervous system. J. Neurosci. Res.Because pax proteins have been reported to influ- 46:551–564.ence tn-c promoter activity (see Jones et al., 1992), AUFDERHEIDE, E., CHIQUET-EHRISMANN, R., and EKBLOM,it is possible that such a signaling cascade may P. (1987). Epithelial-mesenchymal interactions in the
developing kidney lead to expression of tenascin in themanifest itself in the differential expression of ten-mesenchyme. J. Cell Biol. 105:599–608.ascins. In zebrafish ntl mutants, which lack a differ-
BARTSCH, U. (1996). The extracellular matrix moleculeentiated notochord (Halpern et al., 1993), tn-c ex-tenascin-C: expression in vivo and functional character-pression in the somite indeed appears to be affectedization in vitro. Progr. Neurobiol. 49:145–168.(E. Tongiorgi, personal communication).
BARTSCH, S., BARTSCH, U., DOERRIES, U., FAISSNER, A.,Cells in the head epidermis and in the corneaWELLER, A., EKBLOM, P., and SCHACHNER, M. (1992).
express tn-w but not tn-c, while cells of the hypo- Expression of tenascin in the developing and adult cer-chord express tn-c only. We do not know what the ebellar cortex. J. Neurosci. 12:736–749.role of the protein in these cells may be. The most BARTSCH, U., PESHEVA, P., RAFF, M., and SCHACHNER,striking difference in the expression patterns of the M. (1993). Expression of janusin (J1-160/180) in thetwo mRNAs is that only tn-c transcripts can be retina and optic nerve of the developing and adult
mouse. Glia 9:57–69.detected in the CNS. We have previously suggestedBENNETT, M. V. L., BARRIO, L. C., BARGIELLO, T. A.,that this expression may contribute to the consolida-
SPRAY, D. C., HERTZBERG, E., and SAEZ, J. C. (1991).tion of regional identities in the brain (Tongiorgi etGap junctions: new tools, new answers, new questions.al., 1995a). Furthermore, expression of tn-c by theNeuron 6:305–320.floor plate may be speculated to contribute to axonal
BERNHARDT, R. R., CHITNIS, A. B., LINDAMER, L., andpathfinding near the ventral midline of the spinalKUWADA, J. Y. (1990). Identification of spinal neurons
cord. in the embryonic and larval zebrafish. J. Comp. Neurol.In the CNS of mammals and in chicken, two 302:603–616.
members of the tenascin family are expressed: tn-c BERNHARDT, R. R., NGUYEN, N., and KUWADA, J. Y.and tn-r. Whether another tenascin paralog exists (1992). Growth cone guidance by floor plate cells inin the fish, expressed in a pattern similar to tn-r in the spinal cord of zebrafish embryos. Neuron 8:869–
882.mammals and in the chicken, remains to be seen.BERNHARDT, R. R., TONGIORGI, E., ANZINI, P., andThe present data argue, however, for the possibility
SCHACHNER, M. (1996). Increased expression of spe-that there may be even more diversity in the tenascincific recognition molecules by retinal ganglion cellsfamily than has hitherto been anticipated. In fish,and by optic pathway glia accompanies the successfulas in birds and mammals, the different tenascinsregeneration of retinal axons in adult zebrafish.share partly overlapping patterns of expression andJ. Comp. Neurol. 376:253–264.
could possibly serve some overlapping functions. BOURDON, M. A., WIKSTRAND, C. J., FURTHMAYR, H.,However, it is also apparent that during zebrafish MATTHEWS, T. J., and BIGNER, D. D. (1983). HumanCNS development, tn-c is in a position to play a glioma-messenchymal extracellular matrix antigen de-specific role which cannot be duplicated by tn-w. fined by monoclonal antibody. Cancer Res. 43:2796–This is generally true for the different members of 2805.
BRISTOW, J., KIAN TEE, M., GITELMANN, S. E., MELLON,the tenascin family, all of which show some specific
8p3a 1935/ 8P3A$$1935 02-24-98 10:27:42 nbioa W: Neurobio

14 Weber et al.
S. H., and MILLER, W. L. (1993). Tenascin-X: a novel coproteins are immunochemically related to tenascin.Differentiation 37:104–114.extracellular matrix protein encoded by the human XB
gene overlapping P450c21B. J. Cell Biol. 122:265– FAISSNER, A., KRUSE, J., KUHN, K., and SCHACHNER, M.278. (1990). Binding of the J1 adhesion molecules to extra-
cellular matrix constituents. J. Neurochem. 54:1004–BRONNER-FRASER, M. (1988). Distribution and functionof tenascin during cranial neural crest development in 1015.the chick. J. Neurosci. Res. 21:135–147. FAISSNER, A. and SCHACHNER, M. (1995). Tenascin and
janusin: glial recognition molecules involved in neuralCARNEMOLLA, B., LEPRINI, A., BORSI, L., GERMANO, Q.,URBINI, S., and ZARDI, L. (1996). Human tenascin-R. development and recognition. In: Neuroglia. H. Ketten-
mann and B. R. Ransom, Eds. Oxford University Press:J. Biol. Chem. 271:8157–8160.New York, pp. 422–426.CHIQUET, M. and FAMBROUGH, D. M. (1984a). Chick
myotendinous antigen. I. A monoclonal antibody as a FEINBERG, A. P. and VOGELSTEIN, B. (1983). A techniquefor radiolabeling DNA restriction endonuclease frag-marker for tendon and muscle morphogenesis. J. Cell
Biol. 98:1926–1936. ments to high specific activity. Anal. Biochem. 132:6–13.CHIQUET, R. and FAMBROUGH, D. M. (1984b). Chick my-
otendinous antigen. II. A novel extracellular glycopro- FELSENFELD, A. L., CURRY, M., and KIMMEL, C. B.(1991). The fub-1 mutation blocks initial myofibriltein complex consisting of large disulfide-linked sub-
units. J. Cell Biol. 98:1937–1946. formation in zebrafish muscle pioneer cells. Dev. Biol.148:23–30.CHIQUET-EHRISMANN, R. (1990). What distinguishes ten-
ascin from fibronectin? FASEB J. 4:2598–2604. FORSBERG, E. H., FROHLICH, L., MEYER, M., EKBLOM, P.,ASZODI, A., WERNER, S., and FASSLER, R. (1996). SkinCHIQUET-EHRISMANN, R. (1995). Tenascins, a growingwounds and severed nerves heal normally in mice lack-family of extracellular matrix proteins. Experientiaing tenascin-C. Proc. Natl. Acad. Sci. USA 93:6594–51:853–862.6599.CHIQUET-EHRISMANN, R., HAGIOS, C., and MATSUMOTO,
FUSS, B., POTT, U., FISCHER, P., SCHWAB, M. E., andK. (1994). The tenascin gene family. Perspect. Dev.SCHACHNER, M. (1991). Identification of a cDNANeurobiol. 2:3–7.clone specific for the oligodendrocyte-derived repul-CHIQUET-EHRISMANN, R., KALLA, P., PEARSON, C. A.,sive extracellular matrix molecule J1-160/180. J. Neu-BECK, K., and CHIQUET, M. (1988). Tenascin inter-rosci. Res. 29:299–307.feres with fibronectin action. Cell 53:383–390.
FUSS, B., WINTERGERST, E. S., PESHEVA, P., andCHUONG, C.-M., CROSSIN, K. L., and EDELMAN, G. M.SCHACHNER, M. (1993). Characterization of the neural(1987). Sequential expression and differential functionrecognition molecule janusin (J1-160/180). Biochem.of multiple adhesion molecules during the formationSoc. Trans. 144:197–198.of cerebellar cortical layers. J. Cell Biol. 104:331–
342. GARIN-CHESA, P., MELAMED, M. R., and RETTIG, W. J.(1989). Immunohistochemical analysis of human neu-DAMSKY, C., SUTHERLAND, A., and FISHER, S. (1993).ronectin expression in normal, reactive, and neoplasicExtracellular matrix 5: adhesive interactions in earlytissues. J. Histochem. Cytochem. 37:1767–1776.mammalian embryogenesis, implantation, and placen-
tation. FASEB J. 7:1320–1329. GEFFROTIN, C., GARRIDO, J. J., TREMET, L., and VAIMAN,M. (1995). Distinct tissue distribution in pigs of tenas-DOERRIES, U., BARTSCH, U., NOLTE, C., ROTH, J., andcin-X and tenascin-C transcripts. Eur. J. Biochem.SCHACHNER, M. (1993). Adaptation of a non-radioac-231:83–92.tive in situ hybridization method to electron micros-
copy—detection of tenascin messenger RNAs in GINGELL, D. (1993). Contact signalling and cell motility.Symp. Soc. Exp. Biol. 47:1–33.mouse cerebellum with digoxigenin-labelled probes
and gold-labelled antibodies. Histochemistry 99:251– GRUMET, M., HOFFMAN, S., CROSSIN, K. L., and EDEL-
262. MAN, G. M. (1985). Cytotactin, an extracellular matrixprotein of neural and non-neural tissues that mediatesERICKSON, H. P. (1994). Evolution of the tenascin fam-
ily—implications for function of the C-terminal fi- glia-neuron interaction. Proc. Natl. Acad. Sci. USA82:8075–8079.brinogen-like domain. Perspect. Dev. Neurobiol. 2:9–
19. HAGIOS, C., KOCH, M., SPRING, J., CHIQUET, M., andCHIQUET-EHRISMANN, R. (1996). Tenscin-Y: a proteinERICKSON, H. P. and BOURDON, M. A. (1989). Tenascin:
an extracellular matrix protein prominent in specialized of novel domain structure is secreted by differentiatedfibroblasts of muscle connective tissue. J. Cell Biol.embryonic tissues and tumours. Annu. Rev. Cell Biol.
5:71–92. 134:1499–1512.HALFTER, W., CHIQUET-EHRISMANN, R., and TUCKER,ERICKSON, H. P. and INGLESIAS, J. L. (1984). A six-
armed oligomer isolated from cell surface fibronectin R. P. (1989). The effect of tenascin and embryonicbasal lamina on the behaviour and morphology of neu-preparations. Nature 311:267–269.ral crest cells in vitro. Dev. Biol. 132:14–25.FAISSNER, A., KRUSE, J., CHIQUET-EHRISMANN, R., and
MACKIE, E. (1988). The high-molecular-weight J1 gly- HALPERN, M. E., HO, R. K., WALKER, C., and KIMMEL,
8p3a 1935/ 8P3A$$1935 02-24-98 10:27:42 nbioa W: Neurobio

Tenascin-W in Zebrafish 15
C. B. (1993). Induction of muscle pioneers and floor Molecular Cloning: A Laboratory Manual. ColdSpring Harbor Laboratory Press: Cold Spring Harbor,plate is distinguished by the zebrafish no tail mutation.
Cell 75:99. NY.MATSUMOTO, K. I., ISHIHARA, N., ANDO, A., INOKO, H.,HAUZENBERGER, D., KLOMINEK, J., BERGSTROM, S. E.,
and SUNDQVIST, K. G. (1995). T lymphocyte migra- and IKEMURA, T. (1992). Extracellular matrix proteintenascin-like gene found in human MHC class III re-tion: the influence of interactions via adhesion mole-
cules, the T cell receptor, and cytokines. Crit. Rev. gion. Immunogenetics 36:400–403.Immunol. 15:285–316. MATSUMOTO, K. I., SAGA, Y., IKEMURA, T., SAKAKURA,
T., and CHIQUET-EHRISMANN, R. (1994). The distribu-HOFFMAN, S., CROSSIN, K. L., and EDELMAN, G. M.(1988). Molecular forms, binding functions, and de- tion of tenascin-X is distinct and often reciprocal to
that of tenascin-C. J. Cell Biol. 125:483–493.velopmental expression patterns of cytotactin and cyto-tactin-binding proteoglycan, an interactive pair of ex- MOREL, Y., BRISTOW, J., GITELMAN, S. E., and MILLER,tracellular matrix molecules. J. Cell Biol. 106:519– W. L. (1989). Transcript encoded on the opposite532. strand of the human steroid 21-hydroxylase/comple-
ment component C4 gene locus. Proc. Natl. Acad. Sci.HOFFMAN, S. and EDELMAN, G. M. (1987). A proteogly-can with HNK-1 antigenic determinants is a neuron- USA 86:6582–6586.associated ligand for cytotactin. Proc. Natl. Acad. Sci. MORIN-KENSICKI, E. M., and EISEN, J. S. (1997). Sclero-USA 84:2523–2527. tome development and peripheral nervous system seg-
mentation in embryonic zebrafish. DevelopmentHUSMANN, K., FAISSNER, A., and SCHACHNER, M. (1992).Tenascin promotes cerebellar granule cell migration 124:159–167.and neurite outgrowth by different domains in the fi- NISHI, T., WEINSTEIN, J., GILLESPIE, W. M., and PAULSON,bronectin type-III repeats. J. Cell Biol. 116:1475– J. C. (1991). Complete primary structure of porcine1486. tenascin: detection of tenascin transcripts in adult sub-
maxillary glands. Eur. J. Biochem. 202:643–648.JESUTHASAN, S. (1996). Contact inhibition/collapse andpathfinding of neural crest cells in the zebrafish trunk. NORENBERG, U., HUBERT, M., and RATHJEN, F. G.Development 122:381–389. (1996). Structural and functional characterization of
tenascin-R (restrictin) , an extracellular matrix glyco-JOHNSON, S. L., AFRICA, D., WALKER, C., and WESTON,J. A. (1995). Genetic control of adult pigment stripe protein of glial cells and neurons. Int. J. Neurosci.development in zebrafish. Dev. Biol. 167:27–33. 14:217–231.
JONES, F. S., CHALEPAKIS, G., GRUSS, P., and EDELMAN, NORENBERG, U., WILLE, H., WOLFF, J. M., FRANK, R.,G. M. (1992). Activation of the cytotactin promoter and RATHJEN, F. G. (1992). The chicken neural extra-by the homeobox-containing gene Evx-1. Proc. Natl. cellular matrix molecule restrictin: similarity withAcad. Sci. USA 89:2091–2095. EGF-, fibronectin type III-, and fibrinogen-like motifs.
Neuron 8:849–863.KIMMEL, C. B., BALLARD, W. W., KIMMEL, S. R., ULL-
MANN, B., and SCHILLING, T. F. (1995). Stages of em- OAKLEY, R. A., LASKY, C. J., ERICKSON, C. A., and TOS-
bryonic development of the zebrafish. Dev. Dynam. NEY, K. W. (1994). Glycoconjugates mark a transient203:253–310. barrier to neural crest migration in the chicken embryo.
Development 120:103–114.KOZAK, M. (1991). An analysis of vertebrate mRNAsequences: intimations of translational control. J. Cell ONDA, H., POULIN, M. L., TASSAVA, R. A., and CHIU,Biol. 115:887–903. I.-M. (1991). Characterization of a newt tenascin
cDNA and localization of tenascin mRNA during newtKRUSE, J., KEILHAUER, G., FAISSNER, A., TIMPL, R., andSCHACHNER, M. (1985). The J1 glycoprotein—a novel limb regeneration by in situ hybridization. Dev. Biol.
148:219–232.nervous system cell adhesion molecule of the L2/HNK-1 family. Nature 316:146–148. PESHEVA, P., SPIESS, E., and SCHACHNER, M. (1989). J1-
160 and J1-180 are oligodendrocyte-secreted nonper-KYTE, J. and DOOLITTLE, R. F. (1982). A simple methodfor displaying the hydropathic character of a protein. missive substrates for cell adhesion. J. Cell Biol.
109:1765–1778.J. Mol. Biol. 157:105–132.LALLIER, T. and BRONNER-FRASER, M. (1993). Inhibition QIAO, T., MADDOX, B. K., and ERICKSON, H. P. (1995).
A novel alternative splice domain in zebrafish tenascin-of neural crest cell attachment by integrin antisenseoligonucleotides. Science 259:692–695. C. Gene 156:307–308.
RAIBLE, D. W. and EISEN, J. S. (1994). Restriction ofLIGHTNER, V. A. (1994). Tenascin: does it play a role inepidermal morphogenesis and homeostasis? J. Invest. neural crest cell fate in the trunk of the embryonic
zebrafish. Development 120:495–503.Dermatol. 102:273–277.MACKIE, E. J., THESLEFF, I., and CHIQUET-EHRISMANN, R. RAIBLE, D. W., WOOD, A., HODSDON, W., HENION, P. D.,
WESTON, J. A., and EISEN, J. S. (1992). Segregation(1987). Tenascin is associated with chondrogenic andosteogenic differentiation in vivo and promotes chon- and early dispersal of neural crest cells in the embry-
onic zebrafish. Dev. Dynam. 195:29–42.drogenesis in vitro. J. Cell Biol. 105:2569–2579.MANIATIS, T., FRITSCH, E. F., and SAMBROOK, J. (1982). RATHJEN, F. G., WOLFF, J. M., and CHIQUET-EHRISMANN,
8p3a 1935/ 8P3A$$1935 02-24-98 10:27:42 nbioa W: Neurobio

16 Weber et al.
R. (1991). Restrictin: a chick neural extracellular ma- derivatives, and in discrete domains of the embryoniczebrafish brain. J. Neurobiol. 28:391–407.trix protein involved in cell attachment co-purifies with
VAN EEDEN, F. J., GRANATO, M., SCHACH, U., BRAND,the cell recognition molecule F11. DevelopmentM., FURUTANI-SEIKI, M., HAFFTER, P., HAM-113:151–164.MERSCHMIDT, M., HEISENBERG, C. P., JIANG, Y. J.,SAGA, Y., YAGI, T., IKAWA, Y., SAKAKURA, T., and AI-
KANE, D. A., KELSH, R. N., MULLINS, M. C., ODEN-ZAWA, S. (1992). Mice develop normally without ten-THAL, J., WARGA, R. M., and NUSSLEIN-VOLLHARD, C.ascin. Genes Dev. 6:1821–1831.(1996). Genetic analysis of fin formation in the zebra-SCHACHNER, M., TAYLOR, J., BARTSCH, U., and PESHEVA,fish, Danio rerio. Development 123:255–263.P. (1994). The perplexing multifunctionality of ja-
VARELA-ECHAVARRIA, A., TUCKER, A., PUSCHEL, A. W.,nusin, a tenascin-related molecule. Perspect. Dev. Neu-and GUTHRIE, S. (1997). Motor axon subpopulationsrobiol. 2:33–41.respond differentially to the chemorepellents netrin-1SCHILLING, T. F. and KIMMEL, C. B. (1994). Segment andand semaphorin D. Neuron 18:193–207.cell type lineage restrictions during pharyngeal arch
VAUGHAN, L., HUBER, S. M., CHIQUET, M., and WIN-development in the zebrafish embryo. DevelopmentTERHALTER, K. H. (1987). A major, six armed glyco-120:483–494.protein from embryonic cartilage. EMBO J. 6:349–SHULMAN-WEIS, J. (1968). Analysis of the development353.of the nervous system of the zebrafish, Brachydanio.
VON HEIJNE, G. (1988). Transcending the impenetrable:I. The normal morphology and development of the
how proteins come to terms with membranes. Biochim.spinal cord and ganglia of the zebrafish. J. Embryol.
Biophys. Acta 947:307–333.Exp. Morphol. 19:109–119. WESTERFIELD, M. (1993). The Zebrafish Book. Univer-
SPRING, J., BECK, K., and CHIQUET-EHRISMANN, R. sity of Oregon Press: Eugene, OR.(1989). Two contrary functions of tenascin: dissection WINTERGERST, E. S., FUSS, B., and BARTSCH, U. (1993).of the active site by recombinant tenascin fragments. Localization of janusin mRNA in the central nervousCell 59:325–334. system of the developing and adult mouse. Eur. J.
TONGIORGI, E., BERNHARDT, R. R., and SCHACHNER, M. Neurosci. 5:299–310.(1995b). Zebrafish neurons express two L1-related XIAO, Z.-C., TAYLOR, J., MONTAG, D., ROUGON, G., andmolecules during early axonogenesis. J. Neurosci. Res. SCHACHNER, M. (1996). Distinct functions of recombi-42:547–561. nant tenascin-R domains on neuronal cell adhesion,
TONGIORGI, E., BERNHARDT, R. R., ZINN, K., and growth cone guidance, neuronal polarity, and interac-SCHACHNER, M. (1995a). Tenascin-C mRNA is ex- tion with the neuronal immunoglobulin superfamily ad-
hesion molecule F3/11. Eur. J. Neurosci. 8:766–782.pressed in cranial neural crest cells, in some placodal
8p3a 1935/ 8P3A$$1935 02-24-98 10:27:42 nbioa W: Neurobio