yac transgenic analysis reveals wilms' tumour 1 gene activity in the proliferating coelomic...
TRANSCRIPT

YAC transgenic analysis revealsWilms’ Tumour 1gene activity in theproliferating coelomic epithelium, developing diaphragm and limb
Adrian W. Moorea, Andreas Schedlb, Lesley McInnesa, Michael Doylec,Jacob Hecksher-Sorensena, Nicholas D. Hastiea,*
aMRC Human Genetics Unit, Western General Hospital, Crewe Road, Edinburgh EH4 2XU, UKbMax-Delbruck Centrum for Molecular Medicine, Robert-Ro¨ssle-Strasse 10, 13 122 Berlin-Buch, Germany
cResearch Institute of Molecular Pathology, Dr. Bohr-Gasse 7, A-1030 Vienna, Austria
Received 27 August 1998; revised version received 8 October 1998; accepted 8 October 1998
Abstract
Wilms’ Tumour Igene (WT1) is required for the correct development of the urogenital system. To examine its regulation and expression,we created several transgenic mouse lines containing ab-galactosidase reporter driven by the humanWT1 promoter. A 5 kb promoterweakly recapitulated a subset of the endogenousWt1 expression pattern. In contrast, 470 and 280 kb YAC transgenes reproduced thecorrect pattern with high activity and highlighted new expression sites.Wt1 is expressed in the septum transversum revealing how itsmutation causes diaphragmatic defects.Wt1 expression in the limb demarcates a zone between chondrogenic and apoptotic domains.Finally, Wt1 is expressed in mesenchymal cells derived from the coelomic epithelium. Based upon these and further data we discuss aWt1role in epithelial↔ mesenchymal transitions. 1998 Elsevier Science Ireland Ltd. All rights reserved
Keywords:Coelomic epithelium; Diaphragm; Epithelial to mesenchymal cell type transition; Septum transversum; Hepatic sinusoids;Interdigital; Mesenchymal to epithelial cell type transition; lacZ; Limb metanephros; Mouse embryo; Nephric cord; Podocyte; Generegulation; Rete testis Sertoli; Transgenic; Wilms’ Tumour I gene; Yeast artificial chromosome
1. Introduction
Wilms’ tumours or nephroblastomas are kidney tumoursarising from a single kidney precursor cell (metanephricblastema cell) in which developmental control has beenlost (Machin, 1984). Around 15% of Wilms’ tumourscarry a homozygous disruption or deletion of theWilms’Tumour 1gene (WT1). In addition to acting as a tumoursuppressorWT1 is involved in the development of the uro-genital system. Extensive in situ analysis has shown thatWT1 is highly expressed in the kidneys, gonads and themesothelium of mammals (Pritchard Jones et al., 1990; Pel-letier et al., 1991; Armstrong et al., 1992; Sharma et al.,1992; Mundlos et al., 1993; Park et al., 1993; Rackley etal., 1993), and the equivalent structures of other vertebrates
such as birds, reptiles and amphibians (Kent et al., 1995; DelRio-Tsonis et al., 1996; Carroll and Vize, 1996). Due to itsexpression at sites where cells are undergoing a transitionfrom a mesenchymal to an epithelial state, it has been sug-gested that the WT1 protein could be involved in this pro-cess (Pritchard Jones et al., 1990).
Further evidence thatWT1 is involved in urogenitaldevelopment has come from the discovery that in additionto sporadic Wilms’ tumours, different mutations ofWT1cangive rise to three different human congenital syndromes.Haploinsufficieny forWT1 in WAGR syndrome (Wilms’tumour, Aniridia, Genital abnormalities and mental Retar-dation) leads to genital abnormalities. A dominantWT1mutation in Denys-Drash syndrome and the disruption ofcorrect gene splicing in Frasier syndrome both lead to geni-tal abnormalities and in addition glomerular nephropathywhich causes renal failure (Hastie, 1994; Barbaux et al.,1997). Mice homozygous for aWt1 knockout allele die in
Mechanisms of Development 79 (1998) 169–184
0925-4773/98/$ - see front matter 1998 Elsevier Science Ireland Ltd. All rights reservedPII S0925-4773(98)00188-9
* *Corresponding author. Tel.: +44 131 3322471; fax: +44 131 3432620;e-mail: [email protected]

utero around E13–E14 with edema which is symptomaticof heart failure (Kreidberg et al., 1993). Both the kidneysand gonads of homozygous nullWt1mice fail to develop. Inthe developing renal system ofWt1 null mice, the meta-nephric blastema forms, but the ureteric bud does not branchfrom the mesonephric duct, and these cells undergo apop-tosis.
Expression of theWT1gene is tightly controlled duringembryonic development. Furthermore, aberrant expressionof the gene may have severe clinical effects. A Study ofWT1 transcriptional regulation is therefore important forseveral reasons. Firstly, mutations of regulatory elementsmay be of clinical importance. Secondly, determination ofregulatory elements will lead to the identification of factorsinteracting with the gene promoter and hence help elucidatethe regulatory cascade of whichWT1 is part. To date how-ever, WT1 transcriptional control has proven difficult tounravel. The putativeWT1promoter region is highly con-served between mice and humans (~73% identical) withlong stretches of complete identity between the two promo-ters. The promoters are ~70% GC rich, lack TATA orCCAAT boxes (Gessler and Bruns, 1993; Hofmann et al.,1993) and are more similar to those of ubiquitouslyexpressed, rather than developmentally regulated genes. Aputative promoter lying in the 5′ region of theWT1 genedisplays constitutive activity when tested in a large numberof cell lines (Hofmann et al., 1993; Fraizer et al., 1994;Fraizer et al., 1994; Rupprecht et al., 1994; Campbell etal., 1994; Wu et al., 1995). The binding of both tissue spe-cific and ubiquitous transcription factors: Sp1, NF-kB,WT1, GATA1, PAX8 and PAX2, to a number of elementsof the proximal region of theWT1promoter regulates geneactivity in cultured cells (Rupprecht et al., 1994; Wu et al.,1995; Dehbi and Pelletier, 1996; Cohen et al., 1997; Dis-cenza et al., 1997; McConnell et al., 1997; Zhang et al.,1997; Dehbi et al., 1998). To analyse the mechanism ofWT1 gene regulation and expression in vivo we havemade transgenic lines using three different reporter con-structs. A 5 kb fragment of the humanWT1promoter regionwas able to drive weak, but specific expression of a lacZreporter gene mimicking some of the endogenous expres-sion pattern ofWt1. In contrast a lacZ reporter gene insertedinto exon I of theWT1 gene locus on two different yeastartificial chromosomes (YACs) showed high levels ofexpression in transgenic mice in all the expected endogen-ousWt1sites. The high sensitivity of the lacZ reporter geneallowed us to identifyWt1 activity at sites which had notbeen previously identified, even following detailed in situinvestigation by several groups. The gene is expressed in themesenchymal cells derived from the proliferating coelomicepithelium during early development and marks structureswhich later derive from this. Expression ofWt1 within thecoelomic epithelium and sub-coelomic epithelial mesench-yme implies that the gene may be involved in an epithelialto mesenchymal cell type transition in addition to amesenchymal to epithelial one. Furthermore,Wt1 is ex-
pressed on the surface of the septum transversum, explain-ing why diaphragmatic defects are associated withWT1mutation. Finally,Wt1 is expressed in a domain duringlimb development which indicates that the gene may havea role in limiting the domains of digital chondrogenesis and/or interdigital apoptosis.
2. Results
2.1. Tissue specific expression of a 5 kb WT1promoterfragment in transgenic animals
We isolated 5 kb of the humanWT1promoter (USWT1)consisting of approximately 3 kb 5′ of theWT1transcriptioninitiation site,WT1 exon 1 and 1 kb of intron 1. USWT1spans sequences identified as a minimal promiscuousWT1promoter in transfected cell lines and includes transcriptionfactor binding sites proposed to have a role in the regulationof the WT1gene. A lacZ cassette, under the control of itsown translation initiation site, was next cloned intopUSWT1 to give pUSWt1LZ (Fig. 1A).
In order to evaluate the tissue specificity of the USWT1promoter in vivo, we used the USWt1LZ construct to gen-erate two transgenic lines (L21 and L52) and eight embryosat E12.5 which expressed the transgene transiently (T1–T8). Comparison of the expression pattern between eachof these transgenic embryos allowed any potential tissuespecific activity of USWT1 to be assessed.
First inspection of the embryos showed a relatively largedegree of variation between the expression pattern in eachof the embryos, indicating that the USWT1 promoter isweak and susceptible to position effects. However, a carefulcomparison indicated various domains of expression, whichare similar to the expression of the endogenousWt1 geneand recurred in several of the embryos (summarized inTable 1). These domains included the ventral spinalchord, the body wall musculature, the intercostal mesench-yme and/or the differentiating limb musculature. In fiveembryos transgene expression was also found in mesothelialtissues or on the surface of the umbilical hernia (Fig. 1B,C),which is an extension of the mesothelial gut mesentery. Inaddition, a large proportion of embryos showed transgeneexpression in the interdigital region and/or the marginalzones of the developing hand plate. This domain representsa bona fide, previously unidentified domain ofWt1activity(see below). Interestingly, no transgene expression could bedetected within the developing urogenital system, suggest-ing that at least one tissue specific element was missingfrom the construct.
2.2. Creation of YAC transgenic lines carrying the humanWT1 locus
As the USWT1 promoter was weak and unable to drivereporter gene expression in the developing urogenital sys-
170 A.W. Moore et al. / Mechanisms of Development 79 (1998) 169–184

tem, we decided to use yeast artificial chromosomes (YACs)spanning theWT1 locus as transgenes. We hoped that aYAC derived transgene would contain all regulatory ele-ments required for expression of theWT1gene and wouldshield these elements from position effects. Two differentconstructs were used: WT470, a 470 kb YAC spanning thehumanWT1 locus, and WT280, which was obtained bytruncation of WT470 at a position of approximately 10 kbdownstream of theWT1 termination site (Menke et al.,1998). To follow transgene expression a lacZ reportergene was introduced into both WT470 and WT280 byhomologous recombination in yeast (Fig. 2A) (see Section
4. This recombination resulted in an in frame fusion ofWT1exon 1 and the lacZ gene giving rise to WT470LZ andWT280LZ, respectively.
WT470LZ and WT280LZ YAC DNA was isolated asdescribed by Schedl et al. (1993). Seven transgenic founderswere produced by pronuclear microinjection, five of whichtransmitted the transgene to their offspring and were used toestablish transgenic lines (lines A, E and H contain WT470LZ; lines C and D contain WT280LZ). The extent ofYAC transgene integration in each line was determined byhybridizing EcoRI digested genomic Southern blots withprobes spanning different regions along the WT470LZ
Fig. 1. Restriction map and expression pattern of USWT1LZ. (A) USWT1 consists of around 3 kb 5′ of theWT1transcription initiation site,WT1exon 1 and1 kb of intron 1. The construct used by Hewitt et al., 1996 in the creating of transgenic mice driving reporter gene expression spans the region between thetwo PstI restriction sites in USWt1, that of Cohen et al., 1997 contains slightly less 5′ sequence. LacZ, under the control of its own translation initiationcodon, was inserted into theSacII site of pUSWT1 to give pUSWT1LZ. (B) T4 and (C) T5 vibratome sections of transient transgenic embryos carryingUSWT1LZ show USWt1LZ transgene activity in a subset of regions of endogenousWt1activity. T4 hadb-galactosidase activity in the ventral spinal cord,intercostal mesenchyme, mesothelial surface of the liver and the umbilical hernia. T5 had lacZ activity in the ventral spinal cord and slight expression in themesothelial lining of the pleural cavity.
171A.W. Moore et al. / Mechanisms of Development 79 (1998) 169–184

and WT280LZ YACs (Fig. 3 and Section 4. A PCR basedapproach was used to estimate the number o copies of trans-gene integrated in each line (Fig. 3F and Section 4).
The extent of transgene integration in each line is sum-marized in Fig. 2B.
Line A (WT470LZ): FISH analysis mapped the transgeneto chromosome 4, in the region from A3 through B (data notshown). Line A contained one copy of a complete trans-gene. Southern analysis indicated two copies of the LVAsuggesting a second partial integration of the 3′ end of WT470LZ.
Line E (WT470LZ): using Southern analysis it was onlypossible to make out bands corresponding toWT1exons l, 2and 3, indicating that WT470LZ was deleted 3′ from WT1intron 3 in line E. In addition, this line was positive for L800implying that either a single YAC deleted for the 3′ regionof WT1or two separate disrupted YACs had integrated. Dueto the absence of exon 10 integration in this line, a PCRbased copy number analysis was impossible. However,comparison of signal intensities on Southern blots suggestedthe integration of a single copy.
Line H (WT470LZ): transgene integration in this lineoccurred in the vicinity of the telomere of chromosome 4(data not shown). PCR analysis indicated that nine copies oftheWT1locus are integrated. Using the short vector arm as aprobe, more than nine fragments were detected. In contrast,no signal was obtained when hybridising with the long vec-tor arm. These data suggested that WT470LZ had integratedin a tandem array containing differing YAC breakpoints 3′of the SVA. A similar type of YAC transgene integrationhas been previously reported (Schedl et al., 1996).
Line C (WT280LZ): contained one copy of the integratedtransgene truncated 5′ of 1.4CIS.
Line D (WT280LZ): two copies of theWT1locus and theLVA were present, however the transgene was truncatedbetween the 1.4CIS andWt1.
2.3. YAC reporter gene expression mimicked theendogenous Wt1 specific expression pattern
Transgene expression in lines A and H was strong andmimicked that of the endogenous murineWt1 gene in alltissues (Table 2). For example, the expression of lacZ in lineA exactly mirrored that of the endogenous gene in the devel-oping metanephros (Fig. 4A). The lacZ reporter wasexpressed in the metanephric mesenchyme and then up-regulated as this condensed around the branches of the ure-teric and remained on in the glomerulus. The WT470LZtransgene in lines A and H was also active in the correctWt1 specific expression pattern in the developing gonads(Fig. 4E,F), both in the gonadal mesenchyme and theretetestis/rete ovarii. In the Sertoli cells of the developing testis(E12.5–E18.5) however, although endogenousWt1 isexpressed at a very high level, expression of the transgenewas weak. During this period the level of expression of thetransgene in the foetal Sertoli cells increased over time sothat by E18.5b-galactosidase activity was quite high.
Transgene expression was also seen in the correctWt1specific pattern in the nervous system. Theb-galactosidaseexpression was first detected at around E12.5 in the ventralhalf of the lumbar region of the spinal cord adjacent to theventricular zone and suculus limitans (Fig. 4G). At E15.5the domain was now in the medial ventral part of the spinalcord (Fig. 4H). Expression of lacZ was also seen in the roofof the fourth ventricle of the brain (Fig. 4I). In lines A and Htwo regions of ectopic expression of the reporter alsooccurred. These were in the dorsal spinal cord at the levelof the hind brain and in the developing corpus striatum (datanot shown). Ectopic transgene reporter expression has fre-quently been reported at very similar sites due to crypticelements present in lacZ (Meyer-Puttlitz et al., 1995) and/orproduced by chimeric constructs (Russo et al., 1988).
Unlike lines A and H, lines C, D and E showed only a
Table 1
b-Galactosidase expression in USWT1LZ transgenic embryos
Wt1 L21 L52 T1 T2 T3 T4 T5 T6 T7 T8
Mesothelium + + – – – + + + – – +Umbilical hernia –2 + – – + – – + – – +Body wall musculature + + – – – – + – – – –Intercostal mesenchyme + + – – + – – – – – +Limb mesenchyme +3 + – – + – + + + + –Interdigital +3 + + – + – – + – – +Tongue +1 + – – + – – – – + –Ventral spinal cord + – – – – + + + – – +Lower jaw – + – – + – – – – – –Palate – + – – – – + – – + –Notochord – + – – – + – – – – –
An illustration of the domains of expression ofWt1, the endogenousWt1 gene at E12.5. This expression data was collected using either in situ mRNAhybridisation or immunohistochemistry with exception of the tongue (1) where expression was shown by RT-PCR (Armstrong et al., 1992). (2)Wt1expression has not been reported in the umbilical hernia but this is an extension of theWt1 positive gut mesentary. (3) Domains of expression previouslyunreported but described in this paper. L21 and L52,b-galactosidase expression driven by the inserted transgene in E12.5 embryos of the transgenic lines L21and L52. T1–T8b-galactosidase expression driven by the inserted transgene in E12.5 embryos of the eight transient transgenic embryos.
172 A.W. Moore et al. / Mechanisms of Development 79 (1998) 169–184

subset of theWt1 expression pattern (Table 2). Line Eexpressed the reporter gene in the developing metanephrosin a Wt1specific expression pattern whilst C and D did not.Line C showed expression ofb-galactosidase in all correctWt1expression domains with the exception of the metane-phros. Some ectopic lacZ expression was found in the per-ipheral nervous system of this line. Due to the truncation ofthe WT280LZ transgene in lines C and D, it was impossibleto conclude whether upstream or downstream regions wererequired for properWt1expression in the metanephros. Wetherefore performed an additional injection experiment,using WT280LZ as a construct. Three transiently expressingembryos were isolated at embryonic day E17.5 and thedeveloping kidneys stained forb-galactosidase activity.
All of these embryos showed theWt1 specific expressionpattern, in particular with high levels within the proximalpart of the comma, S-shaped body and glomeruli (Fig. 4B–D, respectively). Hence, WT280LZ contains all regulatoryelements required for the correct expression ofWt1 in thesestructures. In addition, truncation of WT280LZ 5′ of theWt1gene appeared to be responsible for the failure of the trans-gene in lines C and D to drive reporter gene expressionwithin the metanephros. As at least 15 kb upstream ofWT1were present in line C (see Figs. 2B and 3D), elementscontrolling metanephros specific expression are likely to besome distance 5′ of the WT1gene.
2.4. Analysis of reporter gene expression identified newdomains of Wt1 activity
The developmental expression pattern ofWt1 has beenextensively studied in mammals and other vertebrates.However, the high sensitivity of theb-galactosidase reportergene driven by a YAC transgenic construct allowed us toidentify previously unreported domains ofWt1expression.
2.5. Wt1 marks the first formation of the nephrogenic cord
The high sensitivity of the lacZ reporter gene allowed usto analyse the developmental onset ofWT1 expression ingreat detail. First expression of the lacZ reporter gene inlines A and H was detected in a six somite stage embryocaudal to the last somite formed (Fig. 5A). As the nextsomites formed theb-galactosidase expressing single cellswere seen laying at the junction between the splanchnopleu-ric and somatopleuric mesoderm. They marked the firstpronephric formation. of the nephrogenic cord (Fig. 5B).By 15–18 somites the nephrogenic cord had given rise toa mesonephric condensation of cells (Fig. 5C,D). The num-ber of forming mesonephric cord mesenchyme cells expres-sing lacZ extended caudally as the tail elongated concurrentwith somitogenesis. A transverse section of an embryo atthis stage showed that at the most caudal end of the meso-nephric cord cells were forming at the junction between thesplanchnopleuric and somatopleuric mesoderm immedi-ately adjacent to the aorta (Fig. 5E). In different embryosit was possible to see transgene expression either more cau-dal to the latest formed somite or up to two somites rostral tothis. Therefore the differentiation of the mesonephric tissuewas at least to some degree independent of somitogenesis.By the 22 somite stage embryo at the rostral end of meso-nephric ridge transgene expression had spread into the cellson the edge of the coelomic cavity which were overlayingthe mesonephric tissue (Fig. 5F).
2.6. Wt1 is expressed in the septum transversum andhepatic sinusoids
Following the first WT470LZ expression in the nephro-genic cord a second domain ofb-galactosidase positive cells
Fig. 2. Creation of the WT470LZ and WT280LZ YAC constructs andsummary of the state of transgene integration in the YAC transgeniclines. (A) A schematic representation of the creation of WT470LZ fromWT470. Exactly the same protocol was followed to make WT280LZ fromWT280. (i) A cartoon of the WT470 YAC. TheWT1gene is representedby an open arrow pointing in the direction of gene transcription The boxesrepresent the vector arms with a dot representing centromeric sequences.(ii) A more detailed map of the region surroundingWT1exon1 in WT470.WT1exon 1 is represented by a black bar,WT1ATG indicates the positionof the WT1 translation start site. (iii) A map of the lacZ targeting vectorpWT-lac. In this construct lacZ (open bar) and leu2 (hatched bar) havebeen inserted in frame into theXhoI restriction site ofWt1 exon 1 (blackbar). (iv) Recombination in yeast between theWT1 locus carried onWT470 and the two regions of pWT-lac homologous toWT1exon1 andintron1 inserts lacZ and leu2 into the YAC to give WT470LZ. A filledarrow represents the targetedWT1 locus which now encoded a WT1exon1/lacZ fusion protein. (B) A schematic representation of the extentof transgene integration in each YAC transgenic line. (i) Cartoons ofWT470LZ and WT280LZ (symbols as in (A)) showing the position ofthe molecular probes used for Southern analysis. (iii) A representationof which probes detected sequence integration in transgenic lines A, Eand H (containing WT470LZ) and lines C and D (containingWT280LZ). A black bar indicates sequence interaction.
173A.W. Moore et al. / Mechanisms of Development 79 (1998) 169–184

appeared caudal to the embryonic heart. This domain ofexpression was on the surface of the septum transversum(Fig. 5C,D) and represented a hitherto unknown region ofWt1 expression. We carried outWt1 whole mount RNA in
situ hybridisation on E9.5 day mouse embryos and con-firmed that this was indeed a bona fide expression domainof Wt1(Fig. 5G). From E10.5 WT470LZ was also expressedon the endothelial cells lining the sinusoids (Fig. 5J). These
Fig. 3. Southern analysis of the extent and number of copies of transgene insertion in the YAC transgenic lines. Southern analysis ofEcoRI digested genomicDNA from each transgenic line (lines A, E, H, C and D), YAC DNA, human genomic and wildtype mouse genomic DNA controls. (A) Blot hybridised withWT33, WT1 cDNA. The lane containing human DNA represents the normal pattern of hybridisation for theEcoRI digestedWT1gene probed with its owncDNA. The numbers up the left hand side of the blot represent theWT1exons represented by each of these bands. Insertion of lacZ into theWT1 locus inWT470 or WT280 added an additionalEcoRI site between WT1 exons 1 and 2 such that the endogenous exon 1 and 2 band is split into two (*exon1,**exon2). The human cDNA also hybridised to the endogenous mouseWt1gene, but with reduced efficiency; this hybridisation gave bands A–F seen veryweakly in the wildtype mouse control lane and more strongly in lane E. In line E only bands fromWT1exons 1–3 are present, the rest of the gene has beenlost. In line D all theWT1 specific bands are present but are extremely weak on this blot. (B–F) Blot hybridized with SVA, L800, 1.4CIS and LVA,respectively. (F) Estimation of the transgene copy number in each transgenic line. The ratio of human derived to mouse derived PCR products gives anindication of the number of humanWT1loci carried in the line (see Section 4. Samples were removed during the exponential stage of a PCR reaction at 25(lane 1) 28 (lane 2) and 30 (lane 3) cycles. The figure shows the relative amount of human and mouse derived product at each of these stages. Line Acontained one copy of a transgenicWT1 locus, line H9, line C1 and line D2. It is was not possible to carry out this copy number analysis for line E as thetransgene is deleted forWT1exon 10. The control shown represents PCR carried out on DNA from a non-transgenic mouse.
174 A.W. Moore et al. / Mechanisms of Development 79 (1998) 169–184

cells are derived from the epithelial surface of the septumtransversum (Le Douarin, 1975).
2.7. Wt1is expressed in the mesenchyme derived fromsomatopleuric and splanchnopleuric proliferating coelomicepithelium
Between E9.5 and E 10.5 there was rapid expansion oftransgene expression. The somatopleuric and splanchno-pleuric coelomic epithelium formed and expressed thetransgene This epithelium began to proliferate and giverise to a sub-epithelial mesenchymal tissue layer (Bannisteret al., 1995).Wt1was expressed in the mesenchyme derivedfrom coelomic epithelia throughout the embryo. For exam-ple, Fig. 5 shows the beginnings of the formation of the sub-coelomic epithelial layer in the parietal pericardium atE10.5 (Fig. 5H) and the subsequent cells formed from theepithelium at E12.5 (Fig. 5I).
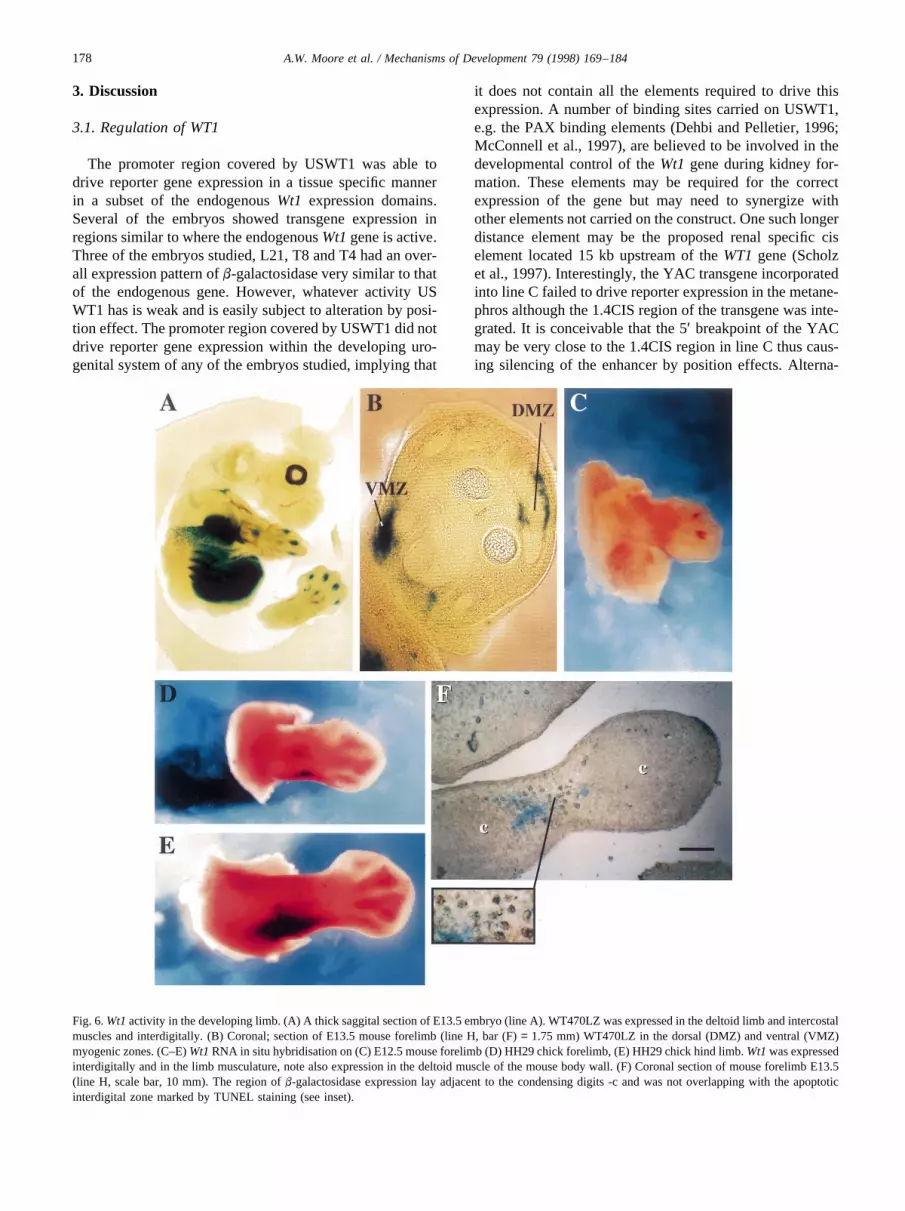
2.8. Wt1expression identified in the limb
At E11.5 WT470LZ expression was switched on at thebase of the limb where it joins the torso. This expression waswithin the developing limb musculature and is connected tothe domain ofWt1 expression in the body wall muscle,confirmed byWt1 in situ hybridisation (data not shown).By E12.5 additional lacZ expression was found in the ante-rior and posterior marginal zones and the base of themesenchyme web between the third and fourth digits.Over the next 24 hb-galactosidase expression first appearedbetween the second and third digit and then spread in a lineeither side of the condensing digits. Simultaneously expres-sion began in the webs between the first and second digitand the fourth and fifth digits. By E13.5 zones of WT470LZexpression ran alongside each developing digit (Fig. 6A).There was also a number ofb-galactosidase positive cellsaround the developing metatarsi and in the dorsal ventralmyogenic zones (Fig. 6A,B). At E15.5 thin domains of
expression sat in the remnants of the interdigital web andran alongside each digit (data not shown).
Reporter gene expression within the interdigital regionswas also seen in animals carrying the active USWt1LZtransgene (see Table 2). In addition, expression ofWT1has been reported in the limb of the alligator (Kent et al.,1995). To check that the limb represents a true expressiondomain of the mouseWt1gene we carried out RNA in situhybridisation analysis of E12.5 mouse embryos (Fig. 6C)and Hamburger-Hamilton stage 29 chick limbs (Fig. 6D,E).Wt1 expression was evident in both the interdigitalmesenchyme and limb musculature in both of these species.The evolutionary conservation of these expression domainsin mouse, chick and possibly the alligator implies that theymay be of functional significance.
Interdigital regions are known to be sites of apoptosiswithin the developing embryo. AsWT1has been implicatedin both suppression and induction of apoptosis (Mahes-waran et al., 1995; Englert et al., 1995; Algar et al., 1996;Menke et al., 1997), we attempted to determine the relation-ship between the WT470LZ expressing cells and thoseundergoing apoptosis. Limbs from E12.5 line H mouseembryos were stained with X-gal for reporter gene activity,microtome sectioned and subjected to TUNEL analysis todetect apoptotic nuclei. TheWt1 expressing cells did notoverlap with the apoptotic cells and lay immediately adja-cent to the region of the digits which are undergoing chon-drogenesis (Fig. 6F). These data suggest thatWt1may play arole in specifying the region of the digit which is betweenthe chondrogenic and apoptotic zones. To investigatefurther the role ofWt1 in limb formation we examinedlimbs from wildtype andWt1 knockout animals at E13.5.We were unable to detect any obvious differences in thepatterns of apoptosis, chondrigenesis and limb muscle for-mation was evident between wild type and knockoutembryo limbs (data not shown). Hence any defects causeby the loss of function of Wt1 in limb development is notrevealed at this stage.
Table 2
b-Galactosidase expression in WT470LZ, WT280LZ and USWt1LZ transgenic embryos
A H E C DWt1 USWT1 WT470 WT470 WT470 WT280 WT280
LZ LZ LZ LZ LZ LZ
Mesothelium + + + + +2 + +2
Musculature + + + + + + –Mesonephros + – + + + + +Metanephros + – + + + – –1
Gonad + – + + – – –2
Interdigital + + + + – + –Tongue + + + + – + –Spinal cord + + + + – – –Fourth ventricle + – + + – + –
An illustration of the domains of expression ofWt1. USWT1, an amalgamation of all theb-galactosidase expression data from L21,L52 and T1–T8.(A,H,E,C,D) the five YAC transgenic lines. Notes: (1) Expression ofb-galactosidase driven by WT280LZwas seen in 3/3 transient transgenic embryosanalysed. (2)b-Galactosidase expression levels in these were reduced compared to the other YAC transgenic lines. In line D transgene expression inmesothelial tissues was in the pericardium only.
175A.W. Moore et al. / Mechanisms of Development 79 (1998) 169–184

Fig. 4. Expression of WT470LZ and WT280LZ mimics that of the endogenousWt1 gene. Both WT470LZ and WT280LZ were expressed in the correctspatiotemporal sequence during nephrogenesis as is illustrated in a transient E17.5 WT280LZ embryo (A–C) and an E15.5 line H (WT470LZ) embryo (D).b-galactosidase was expressed in the proximal part of the (A) comma, (B) S-shape bodies and the (C) podocytes of the mature glomerulus, (P)-distal(D)axis isin the vertical plane. Scale bar, (D)= 30 mm. (D) E 15.5 metanephros and adrenal gland (labelled A) (line H), scale bar, 100 mm. WT470LZ was expressedin the metanephric mesenchyme and upregulated in the mesenchyme undergoing nephrogenesis. It was not expressed in the ureteric duct itself. Expression ofWT470LZ within the developing testes of an (E) E15.5 embryo (line A, scale bar, 100 mm) and (F) E17.5 embryo (line H, scale bar, 50 mm). Note the highlevel of transgene expression in the rete testis. The level of expression of the transgene in the foetal Sertoli cells increased over time, compare E15.5 to E17.5.Labels in (F): arrowheads, germ cells; arrows, Sertoli cells. Expression of the WT470LZ transgene mimicked the endogenous expression pattern ofWt1in thenervous system. (G,H) Transverse section of the spinal cord in the lumbar region of the mouse embryo. The dorsal-ventral axis of the cord is the verticalplane of the frame: dorsal, up; ventral, down. (G) E12.5, (line H), (H) E15.5, (line H). Scale bar, (G,H,) 225 mm. (I) Saggital section of a E13.5 embryo line Hshowing lacZ expression in the roof of the fourth ventricle (R IV). Cb, cerebellar primordium; Cp, choroid plexus. Scale bar, 250 mm.
176 A.W. Moore et al. / Mechanisms of Development 79 (1998) 169–184

Fig. 5. WT470LZ expression during development highlights the nephrogenic cord, septum transversum and sub-coelomic epithelial mesenchyme. (A) Sixsomite embryo (line A) expression of WT470LZ (arrowhead) could be seen in the nephrogenic cord forming caudal to the sixth somite. (B) In a 10 somiteembryo. (C) 15–18 somite embryo (line A) the mesonephric cord (M) is forming and this expressed WT470LZ as did the pronephric region (P). In additionthe septum transversum -S clearly expressed the transgene. (D) A sagittal section through an embryo 20–22 somite stage (line A, scale bar, (H) 285 mm).Open arrow cranial surface of the septum transversum, closed arrow lateral surface. (E) A transverse section through nephrogenic cord of a 20–22 somiteembryo at the most caudal level of the mesonephros (line H, scale bar, 90 mm). Mesonephric cord cells are forming at the junction between thesplanchnopleuric (Sp) and somatopleuric (Sm) mesoderm immediately adjacent to the dorsal aorta DA. (F) A transverse section through more cranialintermediate mesoderm of a 20–22 somite stage embryo (line A, scale bar, 35 mm). The domain of transgene expression now has abutted the coelomic cavity;MD, mesonephric duct. (G) Wt1 RNA in Situ hybridisation on an E9.5 embryo clearly showsWt1expression in the mesonephric mesoderm and the septumtransversum. Expression in the head is probably due to probe trapping. (H–I) Expression of WT470LZ in both the coelomic epithelium and sub coelomicepithelial mesenchyme of the parietal pericardium but not the somatopleuric mesenchyme of an (H) E10.5 and (I) E12.5 embryo (both panels line H, scalebar, 35 mm). Labels in panel H: PP, parietal pericardium; V, ventricle; At, Atrium. (J) E12.5. the transgene was expressed in the epithelial lining on thesinusoid (S) and the mesothelial coat of the liver; H, hepatic cord (Line H, scale bar, 35 mm).
177A.W. Moore et al. / Mechanisms of Development 79 (1998) 169–184
3. Discussion
3.1. Regulation of WT1
The promoter region covered by USWT1 was able todrive reporter gene expression in a tissue specific mannerin a subset of the endogenousWt1 expression domains.Several of the embryos showed transgene expression inregions similar to where the endogenousWt1gene is active.Three of the embryos studied, L21, T8 and T4 had an over-all expression pattern ofb-galactosidase very similar to thatof the endogenous gene. However, whatever activity USWT1 has is weak and is easily subject to alteration by posi-tion effect. The promoter region covered by USWT1 did notdrive reporter gene expression within the developing uro-genital system of any of the embryos studied, implying that
it does not contain all the elements required to drive thisexpression. A number of binding sites carried on USWT1,e.g. the PAX binding elements (Dehbi and Pelletier, 1996;McConnell et al., 1997), are believed to be involved in thedevelopmental control of theWt1 gene during kidney for-mation. These elements may be required for the correctexpression of the gene but may need to synergize withother elements not carried on the construct. One such longerdistance element may be the proposed renal specific ciselement located 15 kb upstream of theWT1 gene (Scholzet al., 1997). Interestingly, the YAC transgene incorporatedinto line C failed to drive reporter expression in the metane-phros although the 1.4CIS region of the transgene was inte-grated. It is conceivable that the 5′ breakpoint of the YACmay be very close to the 1.4CIS region in line C thus caus-ing silencing of the enhancer by position effects. Alterna-
Fig. 6.Wt1activity in the developing limb. (A) A thick saggital section of E13.5 embryo (line A). WT470LZ was expressed in the deltoid limb and intercostalmuscles and interdigitally. (B) Coronal; section of E13.5 mouse forelimb (line H, bar (F)= 1.75 mm) WT470LZ in the dorsal (DMZ) and ventral (VMZ)myogenic zones. (C–E)Wt1RNA in situ hybridisation on (C) E12.5 mouse forelimb (D) HH29 chick forelimb, (E) HH29 chick hind limb.Wt1was expressedinterdigitally and in the limb musculature, note also expression in the deltoid muscle of the mouse body wall. (F) Coronal section of mouse forelimb E13.5(line H, scale bar, 10 mm). The region ofb-galactosidase expression lay adjacent to the condensing digits -c and was not overlapping with the apoptoticinterdigital zone marked by TUNEL staining (see inset).
178 A.W. Moore et al. / Mechanisms of Development 79 (1998) 169–184

tively, another element mapping further upstream may berequired for properWT1 expression in the metanephros.Future transgenic experiments with YACs carrying selec-tive deletions of the 1.4CIS region and other putative controlelements will allow us to test their importance forWT1developmental regulation in vivo.
Hewitt et al. (1996) and Cohen et al. (1997) have reportedthe generation of 10 transgenic lines using about 2 kb of theWT1 promoter linked to a lacZ reporter gene. They wereunable to detect transgene expression in any of these linesand speculated that the minimalWT1promoter had adoptedan ‘inactive’ configuration in the transgenic situation. Theconstructs used by Hewitt and Cohen contained less exon 1sequence than USWT1 and no sequences from intron 1 (Fig.1A). It is possible that these additional sequences on ourconstruct contained vital elements for the binding of basaland tissue specific transcription factors; however no specificfactor binding sites have so far been ascribed to this region.We have carried out a sequence analysis using ‘Signal Scan’(Prestridge, 1991) for transcription factor binding sites con-tained within exon 1 3′ of the PstI site at which both theHewitt and Cohen constructs end through the 260 bp ofintron 1 which have been sequenced (Gessler and Bruns.1993); this leaves 800 bp unanalysed. This region containsnumerous short motifs capable of binding factors such asCAC binding proteins, NF proteins and Sp1; however therepetition and shortness of these sites makes it difficult toascribe an individual function to any element. It is possiblethat the extra DNA inserted in the USWT1LZ transgenicanimals, as opposed to that in the other studies, has helped to‘open up’ a promoter which is weak in activity. The promo-ter is then activated to an extent where transcription fromthe locus is now detectable. This opening may simply be dueto the extra amount of DNA inserted or alternatively it couldbe due to a specific insulator element encoded on USWT1but missing from the other constructs.
3.2. Why is WT470LZ not expressed at a high level inembryonic Sertoli cells?
In this study we have demonstrated that WT470LZexpression in transgenic mice closely mimics expressionof the endogenousWt1 gene. The one exception whichwas found was in the Sertoli cells of the developing testis,where transgene expression was very weak (Fig. 4E,F). Theexpression level ofb-galactosidase increased over time,perhaps due to the steady accumulation of the lacZ geneproduct. This weak expression of WT470LZ could possiblybe explained by negative effects of the inserted lacZ gene onany enhancer for Sertoli cell specific expression carried onthe YAC transgene. However we believe that reduction inreporter gene expression in the embryonic Sertoli cells ismore likely to be due to species specific differences betweenthe mouse and human regulatory elements at theWT1locus.
DNA sequences which play a role in sexual differentia-tion are often highly divergent between species, e.g.SRY
(Whitfield et al., 1993). The male to female sex reversalobserved in some DDS patients together with the recentreport that WT1 acts as a cofactor in the regulation of theMullerian Inhibiting Substance(MIS) gene promoter clearlydemonstrate thatWT1 has a role in sexual determination/differentiation processes (Nachtigal et al., 1998). Perhapsthose regulatory DNA sequences required to drive geneexpression in the sex specific Sertoli cells have undergonespecies specific divergence. A similar inability of a humanderived transgene to drive lacZ reporter gene expression inSertoli cells has been seen with YAC transgenes spanningthe humanSOX9locus (Wunderle et al., 1998), another geneinvolved in sex determination.
3.3. Wt1 marks the first formation of the nephrogenic cordand coelomic epithelium
In the WT470LZ transgenic mouse, the first cells to showexpression of lacZ are immediately caudal to the last somiteof a six somite embryo and mark the first formation of thenephrogenic cord. In the chick the first formation of inter-mediate mesoderm has been observed at the eight somitestage, lateral to the sixth somite (Saxen, 1987). In humans,the first formation of the pronephric tissue occurs in anembryo of eight to nine somites and may extend craniallyas far as the fourth somite (Hamilton et al., 1952). The use ofWt1 as a marker for the very first formation of the nephro-genic cord may help to define the origin of this tissue.
As the volume of nephrogenic cord increases in the meso-nephric ridge it comes to lie adjacent to the coelomic cavity.At this point flattened epithelial-like cells, which are expres-singWt1, now abut the nephrogenic cord (Fig. 5F). It wouldbe interesting to see if, at this point, cells which line thecoelom and which expressWt1 also express mesothelialmarkers such as cytokeratins (LaRocca and Rheinwald,1984) whilst those which do not expressWt1 have a moremesenchymal nature. If this were the case, then the acquisi-tion of Wt1expression would be concurrent with a mesench-ymal to epithelial cell type transition as has previously beenproposed (Pritchard Jones et al., 1990). A further interestingpossibility could be that the coelomic epithelium itself inthis region of the embryo is derived from migrating cellsfrom the nephrogenic cord rather than a mesenchymal toepithelial cell type transition in the cells abutting the coe-lom. Further experiments using diI labelled cells injectedinto the presumptive intermediate mesoderm or the surfaceof the coelomic cavity could clarify this question.
3.4. The involvement of Wt1 in mesenchymal to epithelialand epithelial to mesenchymal type transitions
From E10.5 WT470LZ/WT1 is expressed in the coelomicepithelium as it proliferates and undergoes an epithelial tomesenchymal cell type transition. This proliferation givesrise to theWt1 positive sub-coelomic epithelial mesench-yme. The coelomic epithelium produces cells destined for
179A.W. Moore et al. / Mechanisms of Development 79 (1998) 169–184

all major mesenchymal populations of the trunk (Gruen-wald, 1942). It is not a truly differentiated epithelial layerbut is more similar to the neuroepithelium. It is a pseudos-tratitied epithelium with an inner germinal surface contain-ing cells which have apical epithelial specialisation but nobasal lamina. Both generalized and localized regions ofmesenchyme are produced by the coelomic epithelium.General mesenchyme gives rise to the smooth muscle andconnective tissues of the gut, respiratory tract, reproductiveand urinary tracts among others. Localized populationsinclude the formation of the genital ridges and the spleen.Regions such as the genital ridges, spleen, gut mesenteriesand the smooth muscle of the female reproductive tract areWt1 positive (Pritchard Jones et al., 1990; Pelletier et al.,1991). Hence there seems to be a lineage relationshipbetweenWt1 expression in the coelomic epithelium as itproliferates in the early embryo and expression in thesestructures later in development.Wt1may be down regulatedin these structures upon cell differentiation, or it may remainexpressed into adult life as occurs in the female reproduc-tive tract. Expression ofWT1in the sub-coelomic epithelialmesenchyme may also explain the occurrence of extrarenalWilms’ tumours. These are found in the retroperitonealspace and gonads (Aterman, 1989).WT1positive mesench-yme derived from the coelomic epithelium may have thesame developmental potential as the metanephric mesench-yme and hence give rise to these tumours.
Specific regions of the coelomic epithelium in addition togiving rise to mesenchyme, also produce specialized epithe-lial cells. In lower vertebrates, e.g. fish, birds and reptiles thepodocyte cells of the glomerulus are a continuation of thecoelomic epithelial layer (Kent, 1987). This is also the casein the external pronephric glomeruli of some higher verte-brates, e.g. humans (Hamilton et al., 1952). The epithelialcells surrounding the primordial germ cells (Sertoli cellsand granulosa cells) expressWt1 and may also derivefrom the coelomic epithelium (Bannister et al., 1995).Both podocytes and Sertoli/granulosa cells, althoughappearing morphologically epithelial in nature, expressmesenchymal markers (Ho¨lthofer et al., 1984; Guraya,1995). Hence these cells types could potentially representprogeny of the coelomic epithelial layer partially trans-formed into mesenchyme.
In contrast, the podocytes of the mesonephric and meta-nephric glomeruli are not derived from the coelomic epithe-lium, but from mesenchymal cells within the mesonephriccord and metanephric blastema, respectively (Saxen, 1987).During metanephric nephrogenesis, epithelial structuresform via a mesenchymal to epithelial cell type transitionwithin the metanephric mesenchyme for whichWt1 isbelieved to be essential (Kreidberg et al., 1993). If thismesenchymal to epithelial cell type transition is terminal,as in the epithelium of the nephron tubule, then the gene isdown regulated. However cells destined to form podocytesexpressWt1 throughout nephrogenesis. Podocytes formfrom cells at the proximal part of the S-shaped body
which have already undergone a mesenchymal to epithelialcell type transition.
They now reacquire mesenchymal characteristics as theydifferentiate (Holthofer et al., 1984) Perhaps continuedexpression of theWt1 gene is required for this final partialepithelial to mesenchymal cell type transition, reminiscentof that of the coelomic epithelium into the subcoelomicepithelial mesenchyme. Interestingly, in keeping with thistheory, Denys-Drash syndrome patients often have under-developed podoctyes (Jadresic et al., 1990).
Wt1may also be associated with the ability of cells to flipback and forth between an epithelial and mesenchymal celltype in the gonad. The ability of Sertoli cells to differentiateinto testicular duct cells and vice versa has been reported ina variety of organisms including the Chondrichthys, Tele-osts and amphibians (Grier, 1993). In the axolotlWt1 isexpressed in both cell types at the regions where this trans-formation is occurring; hence it may have a role in thisprocess (Del Rio-Tsonis et al., 1996). Elsewhere in thegonadWt1 may be marking other cells which undergo amesenchymal to epithelial cell type transition. There isstrong transgene expression in therete testis(male) (Fig.4E) andrete ovarii (female). One theory proposes that therete testis/rate ovarii derive from the mesonephric tubules(Satoh, 1991). However the mesonephric tubules do notexpress Wt1 and the rete testis/rate ovarii express itstrongly. Given their expression of Wt1, we would speculatethat the rete testis/rate ovarii have formed via a mesenchy-mal to epithelial cell type transition within the Wt1 positivestroma of the gonad.
What is not clear at this time, is the degree to which themesenchymal cells derived from the coelomic epitheliumand those in the kidney and gonad which undergo amesenchymal to epithelial cell type transition are related.The genital ridges appear to form via localized proliferationof the coelomic epithelium (Gruenwald, 1942), but thenephrogenic cord does not (Fig. 5E). In fact, as raisedabove, it is possible that the coelomic epithelium couldform from the nephrogenic cord. Certainly, as the glomeruliof lower vertebrates project into the coelomic cavity (Kent,1987), the nephrogenic mesenchyme and sub-coelomic epi-thelia mesenchyme may be evolutionarily related.
3.5. Wt1 activity in the septum transversum is required forproper diaphragm formation
The diaphragm is formed from a number of structures allof which expressWt1 in their coelomic epithelium and sub-epithelial mesenchyme. The structures which combine toform the diaphragm are the pars diagrammata of the septumtransversum, the dorsal mesentery of the oesophagus andthe somatopleuric mesenchyme dorsal to the mesonephricridge (Bannister et al., 1995). In theWt1null mouse embryothe diaphragm does not form correctly resulting in herniawith the lungs entering the abdominal cavity (Kreidberg etal., 1993). Similarly, somaticWT1 mutations in patients
180 A.W. Moore et al. / Mechanisms of Development 79 (1998) 169–184

with Denys–Drash Syndrome can also lead to diaphrag-matic hernia (Devriendt et al., 1995) suggesting thatWT1is required for the correct formation of the diaphragm. It isnot clear how mutations in Wt1 cause these diaphragm mal-formations. There is a possible association ofWt1 expres-sion with the ability of cells to undergo proliferation (Menkeet al., 1996; Algar et al., 1996). Hence it is tempting tospeculate thatWT1activity is required for proper prolifera-tion of the tissues which will form the diaphragm, and thatdiaphragm hernia may be due to insufficient closure of thepleuro peritoneal canals. Alternatively tissues destined toform the diaphragm may undergo apoptosis, similar tothat occurring in the metanephric blastema ofWt1knockoutmice (Kreidberg et al., 1993). This second possibility seemsunlikely however, as we did not detect an increase in thenumber of apoptotic nuclei either within the coelomicepithelium or sub-coelomic epithelial mesenchyme inWt1null embryos (data not shown).
The endothelial cells of the hepatic sinusoid also expressWT470LZ and by inferenceWt1 (Fig. 5J). These cells areepithelial cells which are originally derived from the coe-lomic epithelial surface of the septum transversum (LeDouarin, 1975). The foetal liver is a domain of haematopoi-esis; hence the expression ofWt1in the endothelial lining ofthe hepatic sinusoids may be of interest given a potentialrole for the gene both in normal haematopoiesis and ingenesis of leukaemia (King Underwood et al., 1996).
3.6. The Wt1 gene is expressed in the developing limb
WT470LZ expression also led us to identify a so farundescribed domain ofWt1 expression in the limb. Thisexpression occurs between the developing digits immedi-ately adjacent to the zone of chondrogenesis. PerhapsWt1plays a role in delineating these domains. Interestingly,when cell death in the interdigital regions of chick embryosis blocked a default pathway for these cells is chondrogen-esis (Zou and Niswander, 1996). In correlation with thisfinding, loss ofWT1 expression in some Wilms’ tumourscan lead to the aberrant differentiation of metanephric blas-temal stem cells into cartilage (Pritchard Jones and Fleming,1991) as well as muscle (Miyagawa et al., 1998).
Analysis of theWt1 knockout mouse at E13.5 did notshow any obvious abnormalities in limb patterning. How-ever, asWt1 expression during limb development is verytransient, a knock-out related phenotype may be very subtle.Alternatively, a limb phenotype may not be obvious at thestage of development when homozygous null embryos die.
The observed interdigital expression ofWt1described inthis study may however be of clinical relevance, as a strongcorrelation between sporadic Wilms’ tumours and exostosesin patients has been noted (Hartley et al., 1994). Exostosesare benign enchondromata consisting of cartilage cappedosseous projections especially from the long bones of apatient. They are rarely present at birth, but develop withinthe first decade of life. The expression pattern of WT470LZ
laying alongside the developing bones of the limb raises theinteresting possibility that failure ofWT1 activity may beinvolved in exostoses development. In addition, a zone ofWT1expression surrounds the developing ribs as they formimplying that the gene may play similar function in thisprocess (data not shown).EXT2involved in hereditary mul-tiple exostoses lies at 11p12 (Wuyts et al., 1996) and at leastone patient with Wilms’ tumour and exostoses carries adeletion encompassing both ETX2 and WT1. However, dis-ruption of EXT2 is unable to explain exostoses formationcoincident with a number of sporadic Wilms’ tumour cases.
4. Experimental procedure
4.1. Construct design
A l sub-library was constructed by partial Sau3A diges-tion of YAC yIE5 and subsequent cloning intoBamHIdigested l EMBL3 vector (Fantes et al., 1995; Menke etal., 1998). l phages covering part of theWT1 locus wereidentified by hybridisation with WT33 as a probe (Call etal., 1990). A 5 kbBamHI-SalI fragment was isolated fromthe l library clone lWT4 and inserted into the plasmid vectorpUC18 to give pUSWT1. A 3.7 kbHindIII- BamHI lacZcassette isolated from pCH110 (Pharmacia) was clonedinto theSacII site upstream of theWT1start codon to givepUSWT1LZ (Fig. 1A).
WT470 and WT280 are both derived from yIE5 (Menkeet al., 1998). The lacZ targeting vector pWT-lac (plasmidvector pUC 18) was constructed by inserting the lacZ cas-sette from pCHI 10 (Pharmacia) in frame into theXhoI of a1.5 kb PstI-SalI fragment containingWT1exon 1 isolatedfrom the l library clone lWT4. After insertion of a selectablemarker (LEU2) downstream of lacZ, yeast containingWT470 or WT280 were transformed and clones analysedby Southern analysis for homologous recombination events(Fig. 2A).
4.2. Trangenics
Transgenic embryos were generated according to (Schedlet al., 1993; Hogan et al., 1994). Embryos were genotypedeither by Southern analysis using either ILZ or WT33 as aprobe of by PCR. The PCR primers were: HumanWT1specific 5′ACTTCACTCGGGCCTTGATAG3′, 5′GTGG-AGAGTCAGACTTGAAAG3′ bacterial lacZ specific5′AGCTCTGGCACACGTGTCAG3′, 5′GAATGGACGC-TGATAGATGAATTG3′. A PCR based approach wasused to estimate the number of copies of transgene inte-grated in each YAC transgenic line (Fig. 3F). PCR primers5′TGAAAAGCCCTTCAGCTGTC3′ and 5′GGAGTTT-GGTCATGTTTCTCT3′ amplify human and mouse exon10 with equal efficiency to give rise to a 107 bp product.Subsequently, digestion withBstUI distinguishes humanand mouse derived products. Following digestion the PCR
181A.W. Moore et al. / Mechanisms of Development 79 (1998) 169–184

products were Southern blotted and hybridised with 5′TG-CATGTTGTGATGGCGGAC3′ which hybridises withequal efficiency to mouse (59 bp) and human (107 bp) spe-cific bands. The ratio of human (1× transgene) derived tomouse (2× endogenous gene) products can be elucidatedusing PhosphorImager visualisation and the ‘ImageQuant’programme (Molecular Dynamics). This ratio multiplied bytwo gives an approximation of the number of copies of thetransgene inserted into the genome in each line. FISH ana-lysis, to determine the integration site of a transgene, wascarried out as described in (Fantes et al., 1992) using cosmicB2.1 as a probe (Fantes et al., 1995).
4.3. Histological analysis of embryos
The day of plug was designated E0.5. Embryos wereisolated or individual tissues dissected in PBS and stainedas whole mounts forb-galactosidase activity (Hogan et al.,1994), wax embedded, microtome sectioned and counterstained or processed for vibratome sectioning via standardmethods. Alternatively dissected tissues were cryosectionedand then stained forb-galactosidase activity as described byPrice (1994). Detection of apoptotic cells (TUNEL analysis)was carried out using an in situ cell death detection kit, POD(Boehringer Mannheim) in accordance with manufacturersinstructions. Substrate for POD was provided by a VEC-TOR VIP kit (Novocastria).
4.4. Molecular probes
For Southern analysis: WT33-WT1 cDNA (Call et al.,1990), SVA-YAC short vector probe, 1.4 kbSalI/StuI frag-ment isolated from pYAC4, LVA-YAC long vector armprobe, 1.1 kbEcoRI/Pst1 fragment isolated from pBR322;L800: an 800 bp single copy clone isolated from WT470DNA which was isolated from the region of theEagI sitelying around 100 kb downstream ofWT1 (Fantes et al.,1995) via a linking library approach (unpublished data),1.4CIS (Scholz et al., 1997) the 1.4 kb fragment proposedto act as a cis-regulatory element laying 15 kb 5′ of theWT1gene isolated via PCR of WT470LZ DNA; ILZ: 3.6 kbHindIII– BamHI fragment of UYI (Lowings et al., 1992)containing the lacZ cassette. Whole mount RNA in situhybridisation was carried out as detailed by (Wilkinson,1992). MouseWT1 RNA in situ probe PKS2-610bp frag-ment of Wt1 3′ UTR (Mundlos et al., 1993), ChickWT1RNA in situ probe C17 (Kent et al., 1995).
The authors wish to thank Veronica van Heyningen forhelpful discussions; in addition Norman Davidson and team,Brendan Doe and team, Liz Graham and Donald Hay andteam for excellent technical support. A.M. was supported bya Medical Research Council (UK) Studentship, A.S. by aWellcome Trust International Fellowship, M.D. by a Well-come Trust Vacation Scholarship and N.H. is an Interna-tional Research Scholar of the Howard Hughes MedicalInstitute.
References
Algar, E.M., Khromykh, T., Smith, S.I., Blackburn, D.M., Bryson, G.J.,Smith, P.J., 1996. A WT1 antisense oligonucleotide inhibits prolifera-tion and induces apoptosis in myeloid leukaemia cell lines. Oncogene12, 1005–1014.
Armstrong, J.F., Pritchard Jones, K., Bickmore, W., Hastie, N.D., Bard,J.B.L., 1992. The expression of the Wilms’ tumour gene, WT1, in thedeveloping mammalian embryo. Mech. Dev. 40, 85–97.
Aterman, K.., 1989. Extrarenal nephroblastomas. J. Cancer Res. Clin.Oncol. 115, 409–417.
Bannister, L.H., Berry, M.M., Collins, P., Dyson, M., Dussek, J.E., Fergu-son, M.W.J., (1995). Grays’ Anatomy. Churchill Livingstone, NewYork.
Barbaux, S., Niaudet, P., Gubler, M.-C., Grunfeld, J.-P., Jaubert, F.,Kuttenn, F., Fekete, C.N., Souleyreau-Therville, N., Thibaud, E.,Fellous, M., McElreavey, K., 1997. Donor splice-site mutations inWT1 are responsible for Frasier syndrome. Nat. Genet. 17, 467–470.
Call, K.M., Glaser, T., Ito, C.Y., Buckler, A.J., Pelletier, J., Haber, D.A.,Rose, E.A., Kral, A., Yeger, H., Lewis, W.H., et al., (1990). Isolationand characterisation of a zinc finger polypeptide gene at the humanchromosome 11 Wilms’ tumor locus. Cell 60, 509–520.
Campbell, C.E., Huang, A., Gurney, A.L., Kessler, P.M., Hewitt, J.A.,Williams, B.R., 1994. Antisense transcripts and protein binding motifswithin the Wilms tumour (WT1) locus. Oncogene 9, 583–595.
Carroll, T.J., Vize, P.D., 1996. Wilms’ tumor suppressor gene is involvedin the development of disparate kidney forms: evidence from expressionin the Xenopuspronephros. Dev. Dynam. 206, 131–138.
Cohen, H.T., Bossone, S.A., Guoming, Z., McDonald, G.A., Sukhatme,V.P., 1997. Sp1 is a critical regulator of theWilms’ tumour-1gene. J.Biol. Chem. 272, 2901–2913.
Dehbi, M., Hiscott, J., Pelletier, J., 1998. Activation of the wt1 Wilms’tumour suppressor gene by NF-kB. Oncogene 16, 2033–2039.
Dehbi, M., Pelletier, J., 1996. PAX8-mediated activation of the wt1 tumorsuppressor gene. EMBO J. 15, 4297–4306.
Del Rio-Tsonis, K., Covarrubias, L., Kent, J., Hastie, N.D., Tsonis, P.A.,1996. Regulation of the Wilms’ Tumour Gene during spermatogenesis.Dev. Dynam. 207, 372–381.
Devriendt, K., Deloof, E., Moerman, P., Legius, E., Vanhole, C., deZegher, F., Proesmans, W., Devlieger, H., 1995. Diaphragmatic herniain Denys-Drash syndrome. Am. J. Med. Genet. 57, 97–101.
Discenza, M.T., Dehbi, M., Pelletier, J., 1997. Overlapping DNA recogni-tion motifs between Spl and a novel trans-acting factor within thewt1 tumour suppressor gene promoter. Nucleic Acids Res. 25, 4314–4322.
Englert, C., Hou, X., Maheswaran, S., Bennett, P., Ngwu, C., Re, G.G.,Garvin, A.J., Rosner, M.R., Haber, D.A., 1995. WT1 suppresses synth-esis of the epidermal growth factor receptor and induces apoptosis.EMBO J. 14, 4662–4675.
Fantes, J.A., Bickmore, W.A., Fletcher, J.M., Ballesta, F., Hanson, I.M.,van Heyningen, V., 1992. Submicroscopic deletions at the WAGRlocus, revealed by nonradioactive in situ hybridization. Am. J. Hum.Genet. 51, 1286–1294.
Fantes, J.A., Oghene, K., Boyle, S., Danes, S., Fletcher, J.M., Bruford,E.A., Williamson, K., Seawright, A., Schedl, A., Hanson, I., et al.,(1995). A high-resolution integrated physical, cytogenetic, and geneticmap of human chromosome 11: distal p13 to proximal p15.1. Genomics25, 447–461.
Fraizer, G.C., Wu, Y.J., Hewitt, S.M., Maity, T., Ton, C.C., Huff, V.,Saunders, G.F., 1994. Transcriptional regulation of the human Wilms’tumor gene (WT1). Cell type-specific enhancer and promiscuouspromoter. J. Biol. Chem. 269, 8892–8900.
Gessler, M., Bruns, G.A., 1993. Sequence of the WT1 upstream regionincluding the Wit-1 gene. Genomics 17, 499–501.
Grier, H.J., (1993). Comparative organization of Sertoli cells including theSertoli cell barrier. In: Russell, L.D., Griswold, M.D. (Eds.), The SertoliCell. Cache River Press, Clearwater, FL, pp. 704–739.
182 A.W. Moore et al. / Mechanisms of Development 79 (1998) 169–184

Gruenwald, P., 1942. Common traits in the development and structure ofthe organs originating from the coelomic wall. J. Morphol. 70, 353–387.
Guraya, S.S., 1995. The comparative cell biology of accessory somatic (orSertoli) cells in the animal testis. Int. Rev. Cytol. 160, 163–220.
Hamilton, W.J., Boyd, J.D., Mossman, H.W., 1952. Human Embryology.Heffer, Cambridge, UK.
Hartley, A.L., Birch, J.M., Blair, V., Jones, P.M., Gattamaneni, H.R.,Kelsey, A.M., 1994. Second primary neoplasms in a population-basedseries of patients diagnosed with renal tumours in childhood. Med.Pediatr. Oncol. 22, 318–324.
Hastie, N.D., 1994. The genetics of Wilms’ tumor: a case of disrupteddevelopment. Annu. Rev. Genet. 28, 523–558.
Hewitt, J.A., Kessler, P.M., Campbell, C.E., Williams, B.R., 1996. Tissue-specific regulation of the WT1 locus. Med. Pediatr. Oncol. 27, 456–461.
Hofmann, W., Royer, H.D., Drechsler, M., Schneider, S., Royer Pokora,B., 1993. Characterization of the transcriptional regulatory region of thehuman WT1 gene. Oncogene 8, 3123–3132.
Hogan, B.L., Beddington, R., Constantini, F., Lacy, E., 1994. Manipula-tion of the Mouse Embryo. Cold Spring Harbour Laboratory Press,Plainview, NY.
Holthofer, H., Miettinen, A., Lehto, V.-P., 1984. Expression of vimentinand cytokeratin types of intermediate filament proteins in developingand adult human kidneys. Lab. Invest. 50, 552–559.
Jadresic, L., Leake, J., Gordon, I., Dillon, M.J., Grant, D.B., Pritchard, J.,Risdon, R.A., Barrat, T.M., 1990. Clinicopathologic review of twelvechildren with nephropathy, Wilms tumor and genital abnormalities(Drash syndrome). J. Pediatr. 117, 717–725.
Kent. G.C., 1987. Comparative Anatomy of the Vertebrates. Times Mirror/Mosby, St. Louis, Toronto.
Kent, J., Coriat, A.M., Sharpe, P.T., Hastie, N.D., van Heyningen, V.,1995. The evolution of WT1 sequence and expression pattern in thevertebrates. Oncogene 11, 1781–1792.
King Underwood, L., Renshaw, J., Pritchard Jones, K., 1996. Mutations inthe Wilms’ tumor gene WT1 in leukemias. Blood 87, 2171–2179.
Kreidberg, J.A., Sariola, H., Loring, J.M., Maeda, M., Pelletier, J.,Housman, D., Jaenisch, R., 1993. WT-1 is required for early kidneydevelopment. Cell 74, 679–691.
LaRocca, P.J., Rheinwald, J.G., 1984. Coexpression of simple epithelialkeratins and vimentin by human mesothelium and mesothelioma in vivoand in culture. Cancer Res. 44, 2991–2999.
Le Douarin, N.M., 1975. An experimental analysis of liver development.Med. Biol. 53, 427–455.
Lowings, P., Yavuzer, U., Goding, C.R., 1992. Positive and negative ele-ments regulate melanocyte-specific promoter. Mol. Cell Biol. 12, 3653–3662.
Machin, G.A., 1984. Persistent renal blastema as a precursor of Wilms’tumor. In: Pochedly, C., Baum, E.S. (Eds.), Wilms’ Tumor: Clinical andBiological Manifestations. Elsevier, New York, pp. 215–250.
Maheswaran, S., Englert, C., Bennett, P., Heinrich, G., Haber, D.A., 1995.The WT1 gene product stabilizes pS3 and inhibits p53-mediatedapoptosis. Genes Dev. 9, 2143–2156.
McConnell, M.J., Cunliffe, H.E., Chua, L.J., Ward, T.A., Eccles, M.R.,1997. Differential regulation of the human Wilms tumour suppressorgene (WT1) promoter by two isoforms of PAX2. Oncogene 14, 2689–2700.
Menke, A., McInnes, L., Hastie, N.D., Schedl, A., 1998. The Wilms’tumour suppressor WT1 Approaches to gene function. Kidney Int. 53,1512–1518.
Menke, A.L., Riteco, N., van Ham, R.C., de Bruyne, C., Rauscher, F.J.,van der Eb, A.J., Jochemsen, A.G., 1996. Wilms’ tumor I splice variantshave opposite effects on the tumorigenicity of adenovirus-transformedbaby-rat kidney cells. Oncogene 12, 537–546.
Menke, A.L., Shvarts, A., Riteco, N., Van, H.R., Van, d.E., A.J., Jochem-sen, A.G., (1997). Wilms’ tumor 1-KTS isoforms induce p53-indepen-dent apoptosis that can be partially rescued by expression of theepidermal growth factor receptor or the insulin receptor. Cancer Res.57, 1353–1363.
Meyer-Puttlitz, B., Lin, J.H., Sun, T.T., Margolis, R.K., 1995. Ectopicexpression of a bacterial lacZ gene in the limbic system of transgenicmice. NeuroReport 6, 1674–1678.
Miyagawa, K., Kent, J., Moore, A., Charlieu, J.P., Little, M.H., William-son, K.A., Kelsey, A., Brown, K.W., Hassam. S., Briner, J., Hayashi, J.,Hirai, H., Yazaki, Y., van Heyningen, V., Hastie, N.D., (1998). Loss ofWT1 function leads to ectopic myogenesis in Wilms’ tumour. Nat.Genet. 18, 15–16.
Mundlos, S., Pelletier, J., Darveau, A., Bachmann, M., Winterpacht, A.,Zabel, B., 1993. Nuclear localization of the protein encoded by theWilms’ tumor gene WT1 in embryonic and adult tissues.Development 119, 1329–1341.
Nachtigal, M.W., Hirokawa, Y., Enyeart-VanHouten, D.L., Flanagan, J.N.,Hammer, G.D., Ingraham, H.A., 1998. Wilms’ Tumor 1 and Dax-1modulate the orphan nuclear receptor SF-1 in sex-specific geneexpression. Cell 93, 445–454.
Park, S., Schalling, M., Bernard, A., Maheswaran, S., Shipley, G.C.,Roberts, D., Fletcher, J., Shipman, R., Rheinwald, J., Demetri, G., etal., (1993). The Wilms tumour gene WT1 is expressed in murine meso-derm-derived tissues and mutated in a human mesothelioma. Nat. Genet.4, 415–420.
Pelletier, J., Schalling, M., Buckler, A.J., Rogers, A., Haber, D.A.,Housman, D., 1991. Expression of the Wilms’ tumor gene WT1 inthe murine urogenital system. Genes Dev. 5, 1345–1356.
Prestridge, D.S., 1991. Signal scan: a computer program that scans DNAsequences for eukaryotic transcription elements. CABIOS 7, 203–206.
Price, J., 1994. In: Stern, C.D., Holland, P. (Eds.), Essential Developmen-tal Biology: A Practical Approach. IRL Press, Oxford, pp. 190–197.
Pritchard Jones, K., Fleming, S., 1991. Cell types expressing the Wilms’tumour gene (WT1) in Wilms’ tumours: implications for tumourhistogenesis. Oncocene 6, 2211–2220.
Pritchard Jones, K., Fleming, S., Davidson, D., Bickmore, W., Porteous,D., Gosden, C., Bard, J., Buckler, A., Pelletier, J., Housman, D., et al.(1990). The candidate Wilms’ tumour gene is involved in genitourinarydevelopment. Nature 346, 194–197.
Rackley, R.R., Flenniken, A.M., Kuriyan, N.P., Kessler, P.M., Stoler,M.H., Williams, B.R., 1993. Expression of the Wilms’ tumor suppressorgene WT1 during mouse embryogenesis. Cell Growth Differ. 4, 1023–1031.
Rupprecht, H.D., Drummond, I.A., Madden, S.L., Rauscher, F.J.,Sukhatme, V.P., 1994. The Wilms’ tumor suppressor gene WT1 isnegatively autoregulated. J. Biol. Chem. 269, 6198–6206.
Russo, A.F., Crenshaw, E.B., Lira, S.A., Simmons, D.M., Swanson, L.W.,Rosenfeld, M.G., 1988. Neuronal expression of chimeric genes in trans-genic mice. Neuron 1, 311–320.
Satoh, M., 1991. Histogenesis and organogenesis of the gonad in humanembryos. J. Anat. 177, 85–107.
Saxen, L., 1987. Organogenesis of the Kidney. Cambridge UniversityPress, Cambridge, London.
Schedl, A., Larin, Z., Montoliu, L., Thies, E., Kelsey, G., Lehrach, H.,Schutz, G., 1993. A method for the generation of YAC transgenic miceby pronuclear microinjection. Nucleic Acids Res. 21, 4783–4787.
Schedl, A., Ross, A., Lee, M., Engelkamp, D., Rashbass, P., vanHeyningen, V., Hastie, N.D., 1996. Influence of PAX6 gene dosageon development: overexpression causes severe eye abnormalities. Cell86, 71–82.
Scholz, H., Bossone, S.A., Cohen, H.T., Akella, U., Strauss, W.M.,Sukhatme, V.P., 1997. A far upstream cis-element is required forWilms’ tumor-1 (WT1) gene expression in renal cell culture. J. Biol.Chem. 272, 32836–32846.
Sharma, P.M., Yang, X., Bowman, M., Roberts, V., Sukumar, S., 1992.Molecular cloning of rat Wilms’ tumor complementary DNA and astudy of messenger RNA expression in the urogenital system and thebrain. Cancer Res. 52, 6407–6412.
Whitfield, L.S., Lovell Badge, R., Goodfellow, P.N., 1993. Rapid sequenceevolution of the mammalian sex-determining gene SRY. Nature 364,713–715.
183A.W. Moore et al. / Mechanisms of Development 79 (1998) 169–184

Wilkinson, D.G., 1992. Whole mount in situ hybridisation of vertebrateembryos. In: Wilkinson, D.G. (Ed.), In Situ Hybridisation: A PracticalApproach. IRL Press, Oxford, pp. 75–83.
Wu, Y., Fraizer, G.C., Saunders, G.F., 1995. GATA-1 transactivates theWT1 hematopoietic specific enhancer. J. Biol. Chem. 270, 5944–5949.
Wunderle, V.M., Critcher, R., Hastie, N., Goodfellow, P.N., Schedl, A.,1998. Deletion of long range regulatory elements upstream causes cam-pomelic dysplasia. Proc. Natl. Acad. Sci. USA, 95, 10649–10654.
Wuyts, W., Van Hul, W., Wauters, J., Nemtsova, M., Reyniers, E., VanHul, E.V., De Boulle, K., de Vries, B.B., Hendrickx, J.. Herrygers, I.,Bossuyt, P., Balemans, W., Fransen, E., Vits, L., Coucke, P., Nowak,
N.J., Shows, T.B., Mallet, L., van den Ouweland, A.M., McGaughran.J., Halley, D.J., Willems, P.J., (1996). Positional cloning of a geneinvolved in hereditary multiple exostoses. Hum. Mol. Genet. 5, 1547–1557.
Zhang, X., Xing, G., Fraizer, G.C., Saunders, G.F., 1997. Transactivationof an intronic hematopoietic-specific enhancer of the human Wilms’tumor I gene by GATA-1 and c-Myb. J. Biol. Chem. 272, 29272–29280.
Zou, H., Niswander, L., 1996. Requirement for BMP signaling in inter-digital apoptosis and scale formation. Science 272, 738–741.
184 A.W. Moore et al. / Mechanisms of Development 79 (1998) 169–184