xerox university microfilms - ohiolink etd center
TRANSCRIPT

INFORMATION TO USERS
This material was produced from a microfilm copy of the original document. While the most advanced technological means to photograph and reproduce this document have been used, the quality is heavily dependent upon the quality of the original submitted.
The following explanation of techniques is provided to help you understand markings or patterns which may appear on this reproduction.
1.The sign or "target" for pages apparently lacking from the document photographed is "Missing Page(s)". If it was possible to obtain the missing page{s) or section, they are spliced into the film along with adjacent pages. This may have necessitated cutting thru an image and duplicating adjacent pages to insure you complete continuity.
2. When an image on the film is obliterated with a large round black mark, it is an indication that the photographer suspected that the copy may have moved during exposure and thus cause a blurred image. You will find a good image of the page in the adjacent frame.
3. When a map, drawing or chart, etc., was part of the material being photographed the photographer followed a definite method in "sectioning" the material. It is customary to begin photoing at the upper left hand corner of a large sheet and to continue photoing from left to right in equal sections with a small overlap. If necessary, sectioning is continued again ~ beginning below the first row and continuing on until complete.
4. The majority of users indicate that the textual content is of greatest value, however, a somewhat higher quality reproduction could be made from "photographs" if essential to the understanding of the dissertation. Silver prints of "photographs" may be ordered at additional charge by writing the Order Department, giving the catalog number, title, author and specific pages you wish reproduced.
5. PLEASE NOTE: Some pages may have indistinct print. Filmed as received.
Xerox University Microfilms300 North Zeeb RoadAnn Arbor, Michigan 48106

77*2461MILLER, Larry Steven, 1948*
THE IMMUNOLOGIC ACTIVITY OF SUBCELLULAR FRACTIONS AND SOLUBLE PROTEIN FROM A MURINE FIBROSARCOMA,The Ohio State University, Ph.D., 1976 Health Sciences, immunology
Xerox University Microfilms, Ann Arbor, Michigan 48ioe
© Copyright "by Larry Steven Miller
1976

THE IMMUNOLOGIC ACTIVITY OF SUBCELLULAR FRACTIONS AND SOLUBLE PROTEIN FROM A MURINE FIBROSARCOMA
DISSERTATION
Presented in Partial Fulfillment of the Requirements for the Degree Doctor of Philosophy in the
Graduate School of the Ohio State University
ByLarry Steven Miller, B.S., M.S.
*****
The Ohio State University 1976
Reading Committee*Dr. Matthew C. Dodd Dr. Bruno J. Kolodziej Dr. Melvin S. Rheins
Approved By
Adviser Department of Microbiology

Dedicated to the memory of my great-grandfather, William T. Miller
ii

ACKNOWLEDGEMENTS
I want to extend my thanks and appreciation to the following people who have helped me in my graduate career*
Dr. Kolodziej and Dr. Rheins for their assistance and helpful suggestions in preparing this manuscript.
Dr. Dodd for providing me with the opportunity to conduct independent research and to teach as a graduate student. Also, for the many things he has done as my adviser.
My parents for their encouragement and help throughout my academic pursuits.
To my wife, Carolyn, who has helped me prepare this dissertation and has contributed greatly to the attainment of my graduate education.
This research was supported in part by grants from the Ohio Branch of the American Cancer Society and the National Institutes of Health,
iii

TABLE OF CONTENTS
PageACKNOWLEDGMENTS........................................... iiiV I T A ................................................... vLIST OF T A B L E S .............................. •........... viiLIST OF FIGURES.......................................... ixINTRODUCTION ........................................... 1LITERATURE REVIEW . . . . . . . . . . . . 4MATERIALS AND METHODS.............. 20RESULTS........................... ' .......................39DISCUSSION................. ; ............................85SUMMARY................................................. 9 6LITERATURE CITED .................................... 99

VITADecember 17, 19^8 . . . . Bom-Fremont, Ohio1972 ................... B.S., The Ohio State University,
Columbus, Ohio1972-197^ ............. i Graduate Teaching Associate,Department of Microbiology,
The Ohio State University, Columbus, Ohio
1972-197** ............... Research Associate, Department ofMicrobiology, The Ohio State University, Columbus, Ohio
197** . ............... M.S., The Ohio State University,Columbus, Ohio
I97A-I976 . . . . . . . . Graduate Teaching Associate,Department of Microbiology,The Ohio State University, Columbus, Ohio
1975 ................... Research Associate, Department ofNeurology, The Ohio State University, Columbus, Ohio
PUBLICATIONS"A Rapid Method for the Isolation of Plasma Membranes from Tumor Cells." Amer. Son. Microbiol. Abstr., p. 16**, 197***"A Rapid Method for the Isolation of Plasma Membranes from a Murine Fibrosarcoma Using an Aqueous Two-Phase Polymer System." Prep. Biochem., **i **89-**98, 197***"Parameters for Solubilizing Cell Surface Antigens from a Murine Fibrosarcoma Using Triton X-ll**." Fed. Proc.35*751. 1976."RNA-Mediated Lymphocyte Responsiveness to a Murine Fibrosarcoma Tumor as Measured by Microcytotoxicity and Lympho- blastogenesis." Amer. Soc. Microbiol. Abstr., p. 73, 1976.
v

*
FIELDS OF STUDY
Major Fieldi ImmunologyStudies in Immunology. Professors Matthew C. Dodd,
John H. Wallace, and Frank W. ChorpenningStudies in Pathogenic Microbiology. Professor Melvin
S . RheinsStudies in Cell Physiology. Professors Milton A.
Lessler and Clifford A. AngererStudies in Biochemistry. Professors Edward J.
Behrman, George A. Berber, Perry A. Frey, Elizabeth L. Gross, George A. Marzluf,Keith E. Richardson, John S. Rieske, Roy A. Scott, and Thomas E. Webb
Studies in Radioisotopes. Professors Louis Malspeis and Dennis R. Feller

1.2.
3-
5.
6 .
7.
8.
9.
10.
1 1 .
LIST OF TABLES
PageOrigin of genetic information specifyingnovel antigens on experimental tumors. 16Enzyme and ‘biochemical contents of various subcellular fractions. *f-5Titration of antisera from immune and tumor-bearing C3H mice by complement fixation. 49Treatment of interfacial material from tumor tissue with 2.5M NaCl. 50Results from skin testing and titration of antisera from rabbits and guinea pigs that were immunized with the HGF pellet from C3H normal or fibrosarcoma tissue. 53Comparison of antisera from rabbits immunizedwith interfacial material or HGF pellet fromC3H normal tissue or fibrosarcoma tissue. 55Comparison of antigenic fractions for the titration of antisera from rabbits that were immunized with interfacial material from the C 3H fibro sarc oma. 56The antigenic activity of subcellular fractions from the murine fibrosarcoma. 57Summary of the surface membrane marker activity, antigenic content, and immunoprotection for the various subcellular fractions from the CoH fibrosarcoma. 59The solubilization of the interfacial material with ImM Triton X-ll*f in 5mM Tris (pH 9*0) containing 2mM EDTA at 23° C for 1 hr. 70Titration of soluble protein in which theTriton X-114 has been removed by variousprocedures. 7 4*
vii

1 2 ,
13-
14.
15.
Page
Titration of rabbit anti-C^H normal tissue by passive hemagglutination using the tube and microplate tests. 75The skin test reactions of immune guinea pigswith soluble protein from the interfacialmaterial of the murine fibrosarcoma. 76The antibody activity of serum and IgG fraction from rabbits immunized with the HGF pellet from tumor and normal tissue. 79Titration of antigenic activity present in fractions which were eluted from the immuno- adsorbant by the column or the batch method. 83
viii

LIST OF FIGURES
1. The subcellular fractionation of the fibrosarcoma homogenate by using differential centrifugation and an an aqueous two-phase polymer system.
2. An outline for obtaining subcellular fractions from the homogenate by centrifugation at various speeds.
3. The alteration of enzyme marker activities in the supernatant fraction by increasing centrifugal forces.
4. The alteration of enzyme marker activities in the interfacial material fraction by increasing centrifugal forces.
5. The alteration of enzyme marker activities in the pellet fraction by increasing centrifugal forces
6 . The relative activity of various biochemical assays for the subcellular fractions.
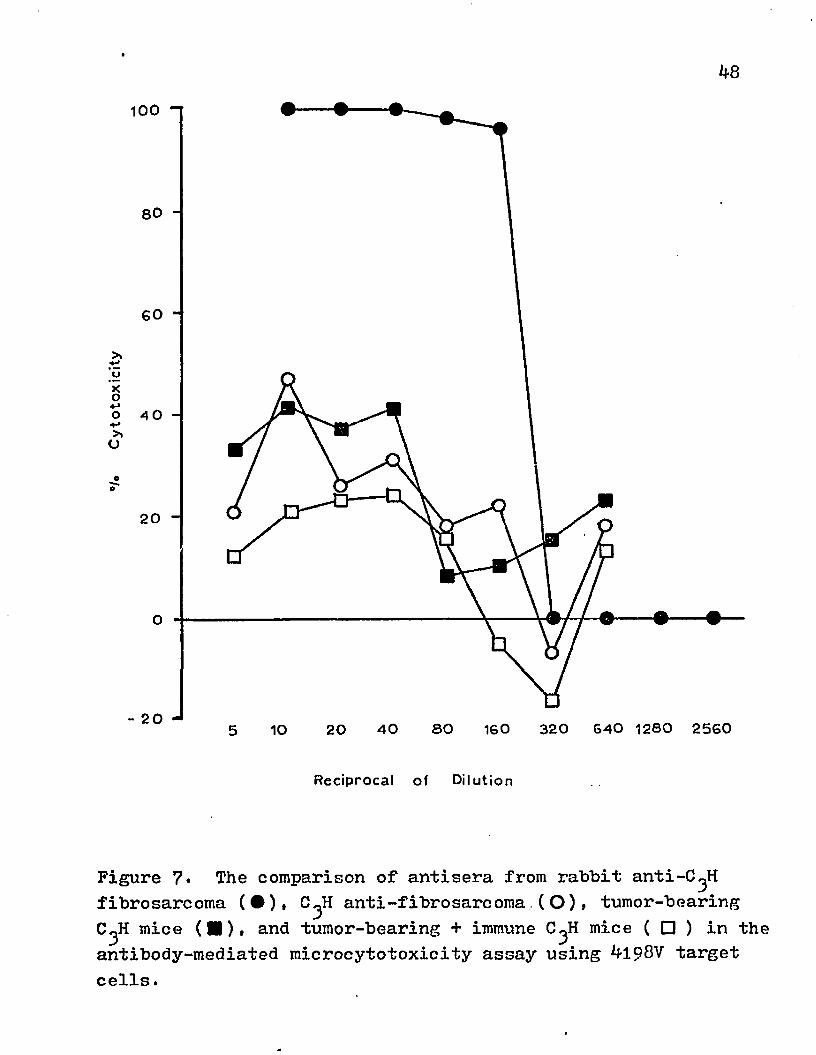
?. The comparison of antisera from rabbit anti-C^Hfibrosarcoma, C3H anti-fibrosarcoma, tumor-bearing C3H mice, and tumor-bearing + immune C3H mice in the antibody-mediated microcytotoxicity assay using 4198V target cells.
8. The appearance of palpable tumors in groups of mice that were injected with the HGF pellet and the other subcellular fractions.
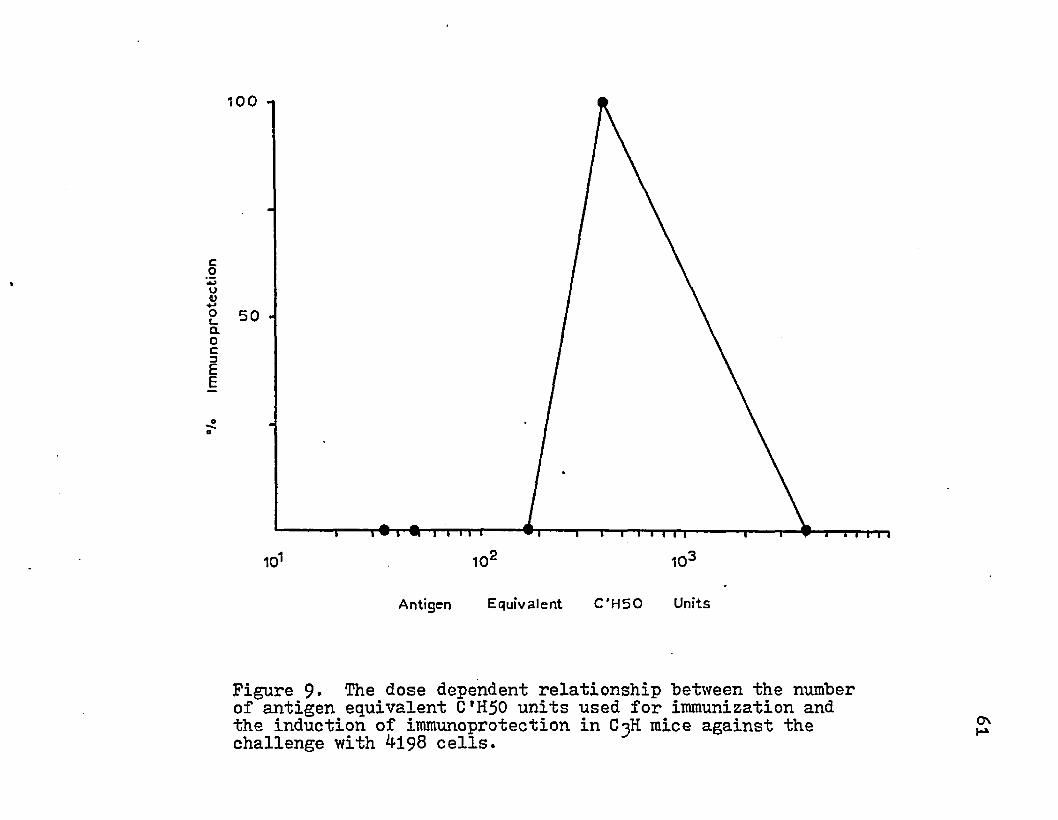
9* The dose dependent relationship between thenumber of antigen equivalent C*H50 units used for immunization and the induction of immuno- protection in C3H mice against the challenge with 4198 cells.
Page
23
40
41
42
43
46
48
60
61
ix

62
64
65
67
68
69
72
78
80
The effects of temperature and different media conditions on the antigenic activity of the HGF pellet from the fibrosarcoma tissue..The solubilization of interfacial material with Triton X-114 in 5raM Tris, 0.25M sucrose + 5mM Tris, or 0.15M NaCl + 5mM Tris at various pH conditions.The effects of Tris osmolality on the solubilization of interfacial material with ImM Triton X-114 at pH 9.0.The solubilization of the interfacial material with varying concentrations of Triton X-114 in 5mM Tris (pH 9 -G).The solubilization of interfacial material with ImM Triton X-114 in 5roM Tris (pH 9.0) at various temperatures.The solubilization of interfacial material with ImM Triton X-114 in Tris (pH y.0) at 23° C with various time exposures.The absorbance of various wavelengths by Triton X-114, bovine serum albumen, and Tris-buffered sucrose.Immunodiffusion analysis of the precipitate obtained from rabbit antiserum by salt fractionation with sodium sulfate.The isolation of antigens from normal tissue by affinity chromatography with IgG-conjugated Sepharose 4B.The isolation of antigens from tumor tissue by. affinity chromatography with IgG-conjugated Sepharose 4B.
x

INTRODUCTION
Malignant cells contain antigenic specificities which are not found in untransformed cells. The unique configurations are located at the cell surface and are important for lymphocyte recognition. Tumor tissue could "be disrupted and the various cellular components differentially fractionated by centrifugation. The subcellular fraction containing the most surface membrane from the ne :plastic tissue could be identified by enzyme and immunologic assays. Antibodies to tumor antigens would have a variety of applications in tumor immunology. Antisera could be produced in heterogeneic animals by injecting a surface membrane fraction. The absorption of these antisera with normal tissue will remove irrelevant antibody specificities and provide antibody which is specific for the tumor. Because of the specificity and sensitivity of immunologic reactions, this absorbed antisera could be used for the immunodiagnosis of cancer. This would provide a means for the early detection of cancer, the histologic identification of the tumor, and the localization of the tumor mass with radiolabelled antibody. This has been attempted with the carcinoembryonic
1

2
antigen in human carcinomas; however, the antisera to this antigen will give falsely positive results in patients with noncancerous diseases. Also, antibodies to the multiple membrane components could be produced to study the changes in the antigen mosaic of the cell surface during oncogenesis.
The neoplastic cell has been able to establish itself in the host by escaping the iiJhjiunological defense mechanisms of the body. Animals and cancer patients which have had the tumor excised or spontaneously regressed contain a serum factor which is cytotoxic for the tumor cells. This serum substance has been identified as an immunoglobulin and appears to play an important role in tumor regression. The absorbed heterologous antiserum could also be used in the immunotherapy of tumor-bearing animals and human patients.
The membrane in the subcellular fraction could be solubilized with a surfactant. Various conditions may effect the release of antigens from the membrane and may provide information about the association between various membrane components. These soluble antigens could also provide a means for assessing the cell-mediated response of the tumor-bearing animal or human patient by in vivo and in vitro assays. Furthermore, the blockade of tumoricidal responses in the tumor-bearing animal or patient appears to be the result of soluble antigen or antigen-antibody complexes. The solubilization of tumor specific antigens from neoplastic

3
tissue would permit the study of soluble antigen in the abrogation of immune responses. Knowledge of the mechanism by which cellular and humoral responses are rendered ineffective could be used to determine methods for producing a protective immunological response.
The isolation of a tumor specific antigen would allow the study of the oncogenic process at the molecular level. These unique specificities could be isolated with an immunoadsorbant. The isolation of cell surface antigens would permit an immunological and biochemical comparison of these determinants with normal tissue antigens. Information from these experiments would provide a better understanding of the molecular changes that occur during the oncogenic process. The development of batch immunoadsorbant procedures would provide the antigen in large quantities. Such material could be used for the production of specific immunoglobulins or for immunization against the tumor.

LITERATURE REVIEW
At the end of the 19th century, some successes had been achieved in immunizing against infectious diseases. Surgical excision of tumors was only occasionally successful (112). Attempts to induce resistance ‘against tumors by vaccination was pursued because rats in which an implanted tumor had regressed demonstrated the ability to resist the growth of a second implanted tumor (38). Ehrlich proposed the Athrepsia Hypothesis which stated that the resistance was effective against not only the original tumor but also those of other histological types (68). Cancer patients given injection of alcoholic extracts from malignant tissue after surgery were free of the neoplasia for four years (108). These achievements were attributed to "the altered chemical nature" of the malignant cells but not normal tissue (108). In 1922, these early hopes of producing panimmunity to a wide variety of tumors were discredited when neoplastic tissue was shown to induce immunity to embryonic (33* 99) and normal tissue (73* 91)• Furthermore, cytotoxic activity could not be evoked when the autochthonous tumor was used for immunization (113). Woglom commented that this was

"precisely the difference from fighting a traitor from within the body and repelling an invader from without"(113)* Thus, these early investigations in eliciting tumor immunity failed to achieve specificity. The immunologic response was directed against heterogeneic differences (33» **3) rather than the tumor specific antigen.
Many subsequent attempts were made to prove the presence of antigens specific for tumors (61), But results were erratic and the activity to normal tissue was still occasionally present in the tumor antisera. The existence of tumor specific antigens was viewed with much skepticism until inbred lines of animals became available. These animals were produced by repeated consanguineous matings until the animal stock was genetically homogenous.In 19^3» Gross induced a sarcoma in C^H inbred mice with methylcholanthrene. The mice in which the tumor regressed were resistant to challenge with this tumor but not with a syngeneic carcinoma (4-8). This tentative identification was repeated in other experiments (39) but the specificity was still questionable. The possibility of tumor rejection being due to individual genetic differences was discarded when it was shown that mice immunized with a methylcholanthrene induced fibrosarcoma would evoke resistance to challenge with tumor but not to skin grafts (9*0 * Inbred animals could also be immunized by tumor ligation, treatment

6
with irradiated cells, or injection of suboptimal doses of cells.
These landmark discoveries opened the door to further investigations regarding the nature of this specific immunity in other animal and human tumors. At present, the resistance to tumor growth is generally attributed to cellular immunity (3)• Histological examination of the tumor mass during the rejection process displayed the presence of lymphoid cells (?6). Furthermore, lymphoid cells from immune animals transferred resistance (30, 82) and the various lymphoid tissues were shown not to be equivalent in transferring immunity (82). Adoptive protection was dependent on the number of lymphocytes injected concurrently with tumor cells. However, cells from immune animals were found to be ineffective against an established progressive tumor even in high numbers (36). These immune lymphocytes appeared 3-10 days after the injection of tumor cells (82) and protective immunity was maintained beyond 80 days. However, adoptive transfer of syngeneic and allogeneic lymphoid cells provided protection for a shorter duration of time (^5 days and 15 days, respectively) (82). When extract from leukocytes of immunized animals were mixed with tumor cells and injected intraperitoneally, regression of malignant tissue occurred (31» 107)• Lymphocytes from tumor-bearing animals also protected normal mice against tumor challenge. This concomitant immunity which was present

7
in animals with a progressive tumor raised questions as to the importance of cellular immunity in tumor graft rejection (30)* Thus, cell-mediated immunity provided resistance to a small tumor inoculum and the tumor-bearing animal demon strated a cellular response to the malignancy.
Further evidence for the importance of the cellular response in tumor cell destruction comes from experiments with immunosuppressed animals. When heterologous antilymphocyte serum was injected into tumor immunized C^H mice, the ability to resist a challenge with the malignant cells was abrogated. This serum was shown not to react with the C^H carcinoma cells (37)• Furthermore, irradiation of immune cells before their transfer abolished their ability to protect (86). This evidence supported an immune surveillance mechanism for tumor cell destruction in which lymphocytes are the effector cells.
The resistance to neoplastic growth may also be a result of nonlymphoid cells. The injection of a large number of macrophages from normal Wistar rats into syngeneic rats bearing a small tumor retarded the growth of the tumor (13). Furthermore, there is evidence that macrophages destroy tumor cells byi (i) an increased potential to destroy foreign cells, (ii) specific recognition of the target cell, and (iii) enhanced "firing” so there is increased dissemination of toxic factors (5) • • Spleen and lymph node cells from syngeneic donors which were irradiated

8
with 3000R were less effective in promoting tumor cell destruction than were similarly treated hybrid and allogeneic cells. This was a result of the ability of such cells to detect only minor differences in surface antigens. Allogeneic inhibition was more effective in protecting against tumor challenge due to the ability to recognize more antigeneic dissimilarities (83)• Therefore, allogeneic inhibition produced by a radiation-resistant population of cells from normal animals provided evidence for a non-immune surveillance mechanism (37t 83)•
Studies with lymphocytes from human cancer patients have demonstrated the immunological phenomena discussed with experimental tumor systems. Patients with melanoma, osteosarcoma, breast carcinoma, colon carcinoma, ovarian carcinoma, lung carcinoma and sarcoma demonstrated positive skin test, cytotoxicity, and lymphoblastogeneic responses (18, 53* 76, 77) which revealed the presence of concomitant immunity. Also, adoptive immunity has been attempted in humans using lymphocytes from patients who showed no recurrence of a similar type tumor. After five years, no evidence of malignant mass or graft versus host reaction was seen with the allogeneic lymphocytes (79)• However, when immunosuppressive therapy was given before injecting lymphocytes from the donor, there were complications from graft versus host reaction and antibodies to allogeneic antigens (*)•).

Thus, an effective cellular response was produced against the tumor. Although concomitant immunity to the tumor was present in human patients, it did not appear to "be protective against a progressive neoplastic mass.
Antibodies may play an important role in the rejection of a malignancy by the host. Serum from animals in which the tumor had regressed can also specifically destroy syngeneic tumor target cells (18, U-7, 50, 70) . In animals with progressive tumors, the antibody titer was significantly lower (37p 4-7> 70). Cytotoxic antibody for tumor specific antigens has been demonstrated in isologous and heterologous antisera (1). Although the tumor-bearing animal or patient demonstrated both a cellular and humoral response, the tumor still progresses. This paradox was clarified when it was observed that sera from tumor-bearing animals blocked cell-mediated cytoxicity (18, 23, 50), and lymphoblasto- genesis (76, 106). Furthermore, a progressive tumor prevented the rejection of homografts or heterografts by the host (78)• Thus, the active immune response to the tumor specific antigens was present, but this may be abrogated by soluble tumor specific antigen (76, 115) or antigen-antibody complexes (115)•
The presence of an immune response which is specific for the malignant cells indicates that there must be present antigens exclusive to the tumor on the cell surface. Initial

10
experiments were performed with methylcholanthrene-induced tumors. When tumor cells were injected into a number of syngeneic mice, the inoculum could be rejected. When these mice were challenged with the same tumor, no growth resulted (39» 48, 49). Pinal proof of the existence of tumor specific antigens on chemically-induced tumors was obtained when tumor-immunized mice demonstrated the ability to reject the tumor but not a skin graft from a syngeneic donor (94). Tumor specific antigens induced by other chemical carcinogens have been demonstrated by immunodiffusion (33)t graft rejection (49, 105)» target cell-lymphocyte rosette formation (114), and cytotoxicity assays (100). Tumors induced with the same or a different carcinogenic agent have distinct identities and immunization against one does not provide protection against the other (39» 48). Carcinogens may induce neoplasia by direct action on the DNA because alteration of RNA or proteins by these chemicals would not be heritable. Gene function can be disrupted by the interaction of a DNA strand with only a few molecules of these chemicals. These changes in informational sequences may effect a site for cellular regulation (17) or activate an oncogene (35)• Therefore, the tumor specific antigens produced by carcinogens are a result of the direct action of a chemical on random sites of the cellular DNA.

11
Unique surface specificities can also be induced with DNA and RNA viruses (88). Tumor specific antigens of viral- induced tumor are not associated with viral antigens (55* ^5* 102, 103); however, the viral genome may code for other novel specificities on the cell surface (52) . Viral-induced tumor antigens do not show the highly individualized characteristics of chemically-induced tumor. Neoplastic cells transformed by the same oncogenic virus demonstrate cross reaction with each other (29, 51* 52, 102), even if the tumors were produced in different species (52). DNA viruses may integrate their genome into the cellular DNA at a complementary site; however, RNA viruses cannot incorporate their information directly into the host's DNA strands. The RNA must first be transcribed into DNA by a reverse transcriptase which is derived from viral information. This exogenous nucleic acid is then integrated into the nuclear material at the appropriate site. These integrated exogenous genetic materials may be activated at a later time to produce a transformed cell(35* 93) • Thus, genetic information from an extrinsic source can induce common tumor specific determinants.
Spontaneous tumors arising in animals and human patients have also demonstrated antigenic specificities present only on malignant cells by cytotoxicity assays (53)* radiolabelled antibody (29), immunofluorescence (87, 95)* immunodiffusion (27), passive hemagglutination (^5* 57)* and complement

12
fixation (32). In addition, these tumors demonstrate cross reactivity between tumors of the same histological type but not with dissimilar types of neoplastic tissue (32, 53f 57#87, 95)• Furthermore, there is evidence that the Gross virus is the causative agent of spontaneous lymphomas in AKR mice (109)• The murine mammary tumors provide an example of a spontaneous tumor produced by an oncogenic virus (12). In humans, Burkitt's lymphoma has been tentatively attributed to Epstein-Barr virus (96). In addition, extracts from human osteosarcoma or lung carcinoma can transform a human embryonic cell line (87). Thus, the antigenic similarity and reports of known oncogenic agents in spontaneous tumors may be evidence for their viral etiology (32, 53, 87).
Histological examination of neoplastic cells revealed that their appearance resembled that of embryonic tissue (9)• Perchloric extracts from colon carcinomas demonstrated antigenic determinants that are shared with embryonic tissue (11, ^6). Furthermore, heterologous antiserum to a fetal tissue extract gave positive results with melanoma tissue but not adult tissues or carcinoembryonic antigen (6). Peripheral blood lymphocytes from cancer patients destroy not only the tumor target cells but also embryonic cells (40).The presence of embryonic and fetal antigens on experimental tumors has also been identified (25, 3^, 87)* Fetal and embryonic antigens are spontaneously released from the

13
neoplastic cells and induce antibody production. Serum from multiparous females demonstrate the ability to block cell- mediated cytotoxicity to fetal target cells (11, 25)• These early antigens are found also in low concentrations on normal tissues (20, 5**t 75)* Therefore, there’is histological and immunological evidence that malignancies are dedifferentiated cells.
Other shifts in the surface antigen mosaic are also produced as a result of transformation. Neoplastic cells may also show a loss in certain antigenic specificities that were present on normal untransformed cells. Antisera from rabbits immunized with hamster kidney carcinoma were absorbed with liver, lung, and sheep erythrocytes and reacted with only tumor tissue. When this antiserum was further absorbed with kidney tissue, the activity of the antisera was not decreased. When heterologous antiserum to normal kidney was prepared in the same manner and then absorbed with these normal tissues, activity was still present to kidney tissue (111). The loss of tissue specific antigens as a result of neoplastic transformation has been described in other experimental animal tumors (8, 58, 85) and human colon carcinoma (21).
In summary, the carcinogenic process causes alterations in the antigenic mosaic of the cell surface. The tumor specific antigen may represent an altered normal surface

14
antigen or an entirely novel specificity induced "by viral and chemical agents. Antigens that are also present on normal tissue may be present in greater quantity in tumor tissue (antigen amplification) or disappear (antigenic deletion). However, others may remain relatively constant in concentration (51)• These shifts in the surface antigens may provide answers to help solve the mystery of the carcinogenic process.
The appearance of novel antigenic specificities on neoplastic cells implies that there is present in the cellular DNA extrinsic or intrinsic information coding for these phenotypes (Table 1). Different theories have been proposed to explain the transformation process. The Warburg Hypothesis stated that the respiratory system in normal cells is irreversibly injured and this is compensated by increased glycolysis. Although there may be dramatic shifts in the biochemical pathways in malignant cells, there are normal tissues that display higher rates of glycolysis than neoplastic tissue. The Greenstein Hypothesis purported that the biochemical pathways converge or are simplified to a common enzymatic pattern but more recent evidence suggests that enzymatic patterns in malignant cells are divergent. Observations also demonstrated the possibility that carcinogenesis may be associated with the loss of proteins essential for growth regulation

15
(Deletion Hypothesis) or enzymes in catabolism (Catabolic Deletion Hypothesis). These concepts have been incorporated into a more general theory on control mechanisms in which neoplastic cells arise from abnormal regulation of cellular metabolism due to random gene mutation or epigenetic information. The somatic mutation theory proposes that carcinogenesis proceeds in normal cells which undergo heritable gene mutations and this results in the loss of negative feedback on systems required for cell division.The mechanism may involve random error in nucleic acid replications errors induced by chemical, biological, or physical agents; or a loss of genetic material during segregation (93)• A Viral Theory of Carcinogenesis has been more recently proposed. This is based on evidence that there are nucleic acid sequences in the cellular DNA from transformed cells which are homologous with the viral genome. Chemical and physical induction of tumors are integrated into this theory as components that may activate a latent virus (92). This theory has been further extended into oncogene and provirus theories. The oncogene theory states that cellular DNA obtained information for oncogenic virus at an early time in evolution; whereas, the provirus theory postulates that the acquisition of this new genetic information is a result of recent infection by an exogenous oncogenic virus (35)•
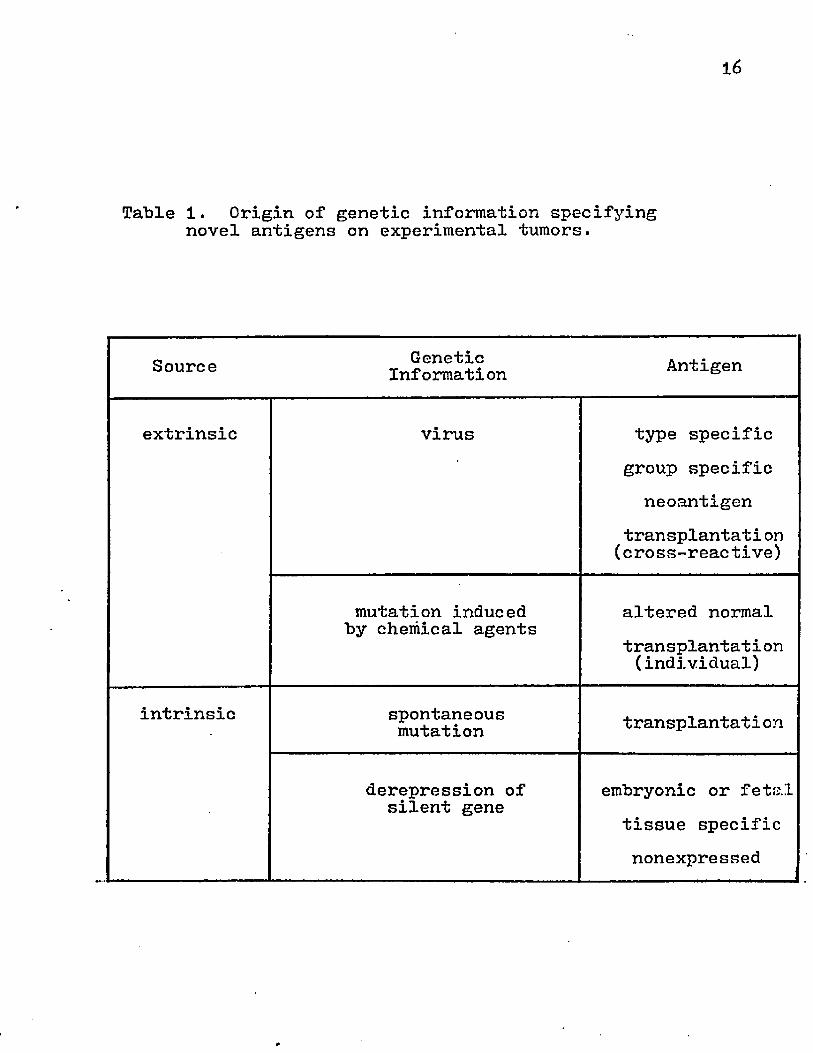
16
Table 1. Origin of genetic information specifying novel antigens on experimental tumors.
Source GeneticInformation Antigen
extrinsic virus type specificgroup specific
neoantigentransplantati on (cross-reactive)
mutation induced by chemical agents
altered normaltransplantati on (individual)
intrinsic spontaneousmutation transplantati on
derepression of silent gene embryonic or fetal
tissue specific nonexpressed

17
The tumor-bearing animal also has demonstrated an immune response to the tumor specific antigens. However, the successful neoplastic cell has evaded these destructive mechanisms. Several possible mechanisms by which this may be achieved have been discovered. Cells which are more sensitive to the effects of antibody-mediated cytotoxicity probably have a higher antigen density on their cell surface. Tumor cells which resist the cytotoxic activity of one antiserum may become sensitive when exposed to antisera from several animals. Antibodies in these antisera may be directed to different antigenic components on the cell surface. Thus, antigen density on the cell surface may play a critical role in antibody-mediated cytotoxicity (84). Antigen concentration may also limit the effectiveness of lymphocytes to express cytotoxicity (59)• Antigen density also fluctuates with the cell cycle and has been shown to be highest during the G1 phase, and at its lowest levels during S, G2, M, and D (19)* Another mechanism by which a neoplastic cell may escape immune destruction is by antigenic modulation. In murine leukemias, the thymus leukemia antigen may be suppressed in immune hosts and the cell reverts to the original phenotype when this negative pressure is removed (16). Soluble antigens from tumor cells may neutralize the immune destruction of target cells. It has been demonstrated that soluble antigens will block

cell-mediated cytotoxicity of target cells and this is probably due to the prevention of recognition of surface antigen by lymphocytes because of steric hindrance (1*0 .The afferent blockade by soluble antigen is only temporary (1 *0 . Injection of solubilized tumor antigens will also induce enhancement of malignant growth (60, 76, 110, 115)* However, antigens that have been solubilized from tumor cells also demonstrate the ability to induce immunopro- tection within a narrow dosage range. The use of low or high amounts of soluble antigen will not produce a protective response (90). Neoplastic cells can also shed surface antigens in vivo. This property is shared by embryonic tissue, and may be an evolutionary mechanism designed to protect the fetal graft from immune destruction. However, the malignant cell which is embryonic in nature also enjoys the shelter that is provided by this evolutionary device (2). Thus, mechanisms exist in which the afferent and efferent arms of the immune response are blocked.
The host defenses against neoplastic cells may operate by two types of surveillance mechanisms* (i) immune in which lymphocytic cells are responsible for the recognition and destruction of endogenously arising aberrant cells, and (ii) nonimmunological in which removal of malignant or other foreign cells is based on contactual phenomena and is sim

19
ilar to allogeneic inhibition. The ability of neoplastic cells to escape immune surveillance may be attributed to tolerance, enhancement, or antigenic loss. Nonimmune surveillance may be evaded by adoptive changes or large colony size (6*0 .

MATERIALS AND METHODS
Cell Lines* A polyoma-induced fibrosarcoma (4198) in C^H mice was grown as a monolayer culture in Eagle's Minimum Essential Medium (MEM) containing 10# fetal calf serum (K.C. Biologicals) and incubated at 37°C in 5fo C02* A 1110170 genic variant of the 4198 cell line (4198V) and a fibroblast cell line (4094) were maintained in a similar manner. Confluent monolayers were detached with 0.25# trypsin 1*200 (Nutritional Biochemicals Corp.) in calcium-magnesium free phosphate-buffered saline. These cells were harvested with the culture medium and seeded into glass culture flasks at a concentration of 200 cells/mm . Additional culture medium was added to the flask to a depth of 3-4 mm. Cell lines were grown for approximately 20 subcultures and then dis7 carded. A fresh sample of cells stored in 9# dimethyl sulfoxide (DMS0) at -120°C was quickly thawed, washed in MEM + 10# FCS + 200 units Penicillin + 200 ug Streptomycin, and then seeded into a flask with media containing antibiotics. The cells from the second subculture were placed in ampules and stored in 9# DMS0 at -120°C.
Tumor Tissue* The fibrosarcoma cell monolayer was harvested, washed twice in Seligman's Balanced Salt Solution
20

21
(SBSS) , and resuspended to 2.5-5*0x10^ cells/ml. C^H/COX male mice 5-10 wks of age were injected subcutaneously with 0.1 ml of the **198 cell suspension. With this number of cells palpable tumors of 1.0-1.5 cm diameter were present on day 16. The tumor-bearing animal was killed by cervical dislocation and exsanguinated. The area of incision was washed with 70# ethanol, the skin was cut with sterile scissors, and the tumor tissue was cut from the leg with a sterile scapel. The excised tissue was placed in cold 0.15 M NaCl + 0.01 M phosphate at pH 7*2 (PBS). The necrotic tissue was dissected from the pinkish-white tissue and discarded. The remaining tumor tissue was placed in cold MEM + 200 units Penicillin + 200 ug Streptomycin and stored not more than 2** hrs at **°C. Liver, muscle, kidney and spleen tissue were collected in the same manner and kept separate from the tumor tissue.
Homogenizationt The tissue was drained, weighed, minced, and pushed through 102 mesh nylon screen (7**u) . Cold 0.25 M sucrose + 0.01 M Tris(hydroxylmethyl)aminoethane + 0.001 M MgSO^ at pH 7.2 (STM) was added to the disaggregated tissue to make the cell suspension 10# (w/v). The cell suspension was then homogenized by nitrogen cavitation in which the cell material was equilibriated with nitrogen at 600 psi for 30 min and then returned to atmospheric pressure. The homo- genate was collected in a chilled sidearm flask containing an adequate amount of 0,1 M cefchylene diaminetetraacetic acid (EDTA) at pH 7*2 to make the suspension 0.001 M and allowed

c22
to stand for 15 min at *J-°C. A small amount of the homo- genate was saved for analysis and the remainder was fractionated according to the protocol described below.
Subcellular Fractionation! The homogenate was centrifuged at various speeds for 15 min at 4°C in either 50 ml or 250 ml polycarbonate tubes or bottles using a Sovall RC2-B centrifuge containing an SS3^ or GSA rotor (Figure 1). The pellet was partitioned in an aqueous two-phase polymer system according to the method described by Miller et al. (81). The polymer system consisted of phosphate-buffered dextran T500 (Pharmacia) and polyethylene glycol 6000 (Union Carbide). In summary, the pellet was mixed with this polymer in a polycarbonate centrifuge tube and centrifuged at 1000 xg for 15 min at 4°C. During this centrifugation, the polymer phases separated and cellular material collected at the interface bewteen these phases and at the bottom of the centrifuge tube. The interfacial material and pellet were collected and washed with chilled 0.25 M sucrose + 0.01 M Tris at pH7.2 (ST). These samples were saved for analysis.
The supernatant remaining from the previous low speed centrifugation was centrifuged at 16,000 xg for 15 min at C. This intermediate centrifugal force (IGF) pellet was
resuspended in ST medium and saved for analysis. The supernatant was centrifuged at 113»000 xg for 60 min at ^°C. The supernatant was collected and saved for analysis. The pellet was resuspended in cold 0.01 M Tris at pH 7.2 using 5 strokes
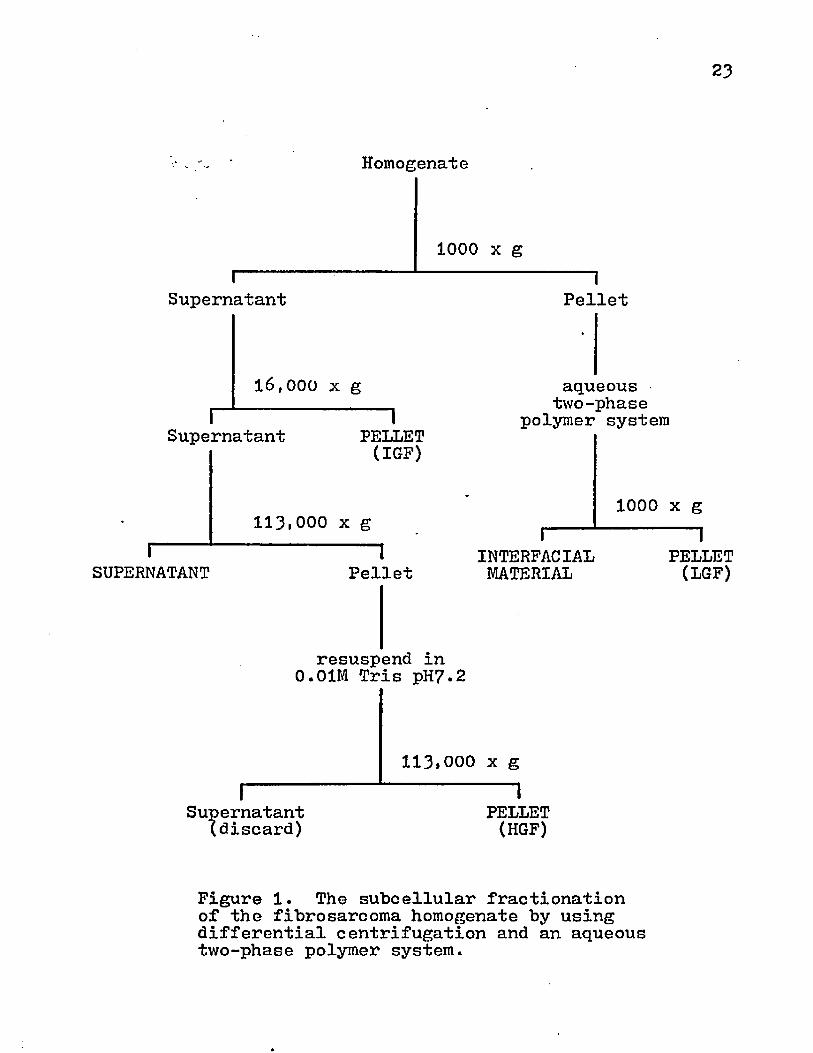
23
Homogenate
1000 x g
Supernatant
16,000 x g
Supernatant 1PELLET(IGF)
113.000 x gI-----
SUPERNATANT 1Pellet
Pellet
aqueous two-phase
polymer system
I----INTERFACIALMATERIAL
resuspend in 0.01M Tris pH?.2
I--------Supernatant(discard)
113,000 x g 1PELLET(HGF)
1000 x g 1PELLET(LGF)
Figure 1. The subcellular fractionation of the fibrosarcoma homogenate by using differential centrifugation and an aqueous two-phase polymer system.

24
of a Dounce homogenizer with a loose-fitting pestle (Type B) on ice and centrifuged again. This high centrifugal force (HGF) pellet was collected and resuspended in ST medium. The supernatant was discarded.
Enzyme and Biochemical Assays» This analysis was performed using previously described methods (81). The protein content was determined by the method of Lowry et al. (71) using bovine serum albumin as a standard. The DNA was determined by Burton's Method (22) using a deoxyribonucleate standard. The 51-mononucleotidase (EC 3*1*3*5** orthopho- phoric monoester phosphohydrolase) activity was assayed according to the method described by Lesko et al. (6?) and was used as enzyme marker for plasma membrane. The amount of inorganic phosphate released from adenosine monophosphate was measured by the Lowry and Lopez method (72). Cytochrome oxidase (EC 1.9*3*l*i cytochrome: oxygen oxidoreductase) was used as an enzyme marker for mitochondria and was assayed by the procedure described by Cooperstein and Lazarow (26). The presence of endoplasmic reticulum was determined by DPNH diaphorase (EC 1.6.4*, NADH: cytochrome c oxidoreductase) activity as described by Wallach and Kamat (110).
Immunization with Subcellular Fractions: Outbred albinorabbits (2.0-3*0 kg) were injected with cell fraction protein in complete or incomplete Freund's Adjuvant (FCA-FIA) according to the following schedule.
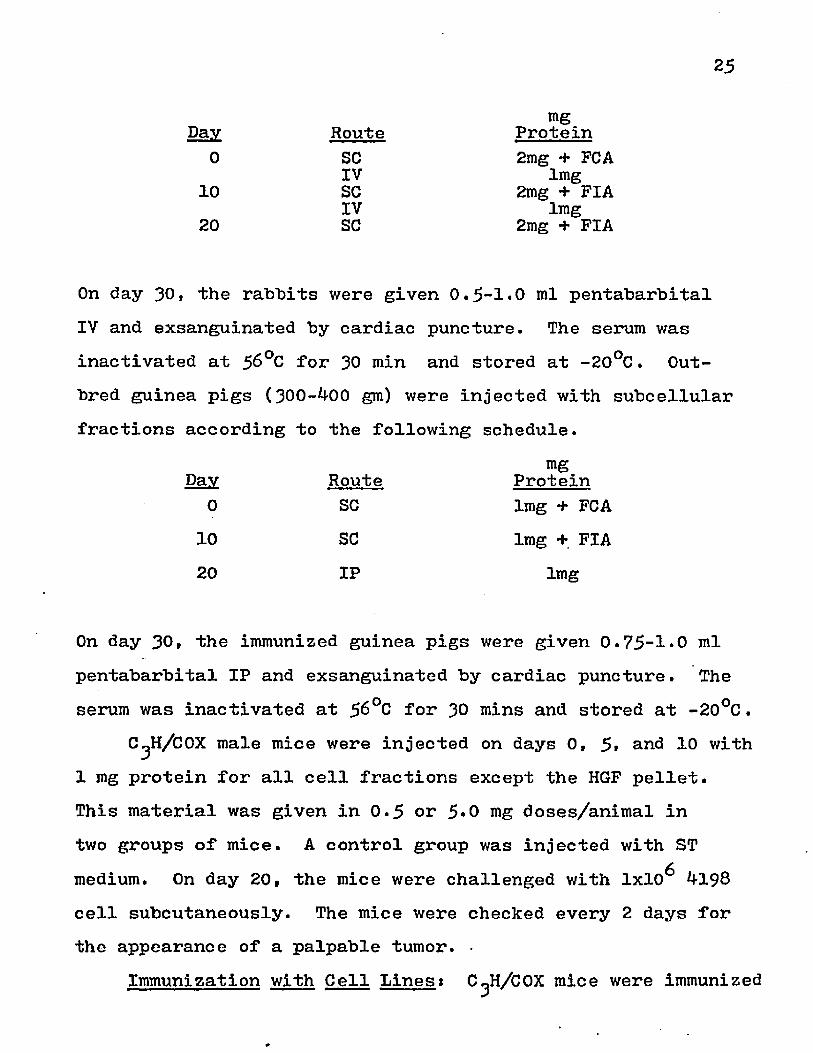
25
mgDay Route Protein
0 SC 2mg + FCAIV lmg
10 SC 2mg + FIAIV lmg20 SC 2mg + FIA
On day 30, the rabbits were given 0.5-1*0 ml pentabarbital IV and exsanguinated by cardiac puncture. The serum was inactivated at 56°C for 30 min and stored at -20°C. Out- bred guinea pigs (300-^00 gm) were injected with subcellular fractions according to the following schedule.
mgDay Route Protein
0 SC lmg + FCA10 SC lmg + FIA20 IP lmg
On day 30, the immunized guinea pigs were given 0.75-1*0 ml pentabarbital IP and exsanguinated by cardiac puncture. The serum was inactivated at $6°C for 30 mins and stored at -20°C.
C^H/COX male mice were injected on days 0, 5t arid 10 with 1 mg protein for all cell fractions except the HGF pellet.This material was given in 0-5 or 5*0 mg doses/animal in two groups of mice. A control group was injected with ST medium. On day 20, the mice were challenged with 1x10^ *1-198 cell subcutaneously. The mice were checked every 2 days for the appearance of a palpable tumor. •
Immunization with Cell Lines: C^H/COX mice were immunized

26
with the more antigenic variant of the polyoma-induced fibrosarcoma (ifl98V). This was accomplished by three intraperitoneal injections on days 0, 5> and 10 with 1x10** cells. On day 15* the mice were exsanquinated.Serum samples were inactivated at 56°C for 30 min and stored at -20°C. Guinea pigs were immunized with *H98 cells according to the following schedule.
Day # Cells0 1x10 7 + FCA5 1x107 + FIA
10 1x107
Serum Absorption! Rabbit antisera diluted 1*5 with PBS were absorbed by the addition on interfacial material from normal C^H tissue. This subcellular fraction was pelleted at 1000 xg for 15 min in a polycarbonate tube and the supernatant was discarded. Three volumes of diluted antiserum were mixed with the pellet and allowed to incubate for 1 hr at 37°C. This material was centrifuged at5000 xg for 10 min at b°C in a Sovall RC2-B centrifuge witha SS3** rotor. The supernatant was collected and absorbed one additional time. This antiserum was then centrifuged at10,000 xg for 10 min at ^°C.
Microcytotoxicity Assays The *H98V cells were harvested and suspended to 1x10** cells/ml in MEM + 10^ FCS + 200 units
RouteSCSCIP

Penicillin + 200 ug Streptomycin. The cell suspension was dispensed in 10 ul aliquots into Falcon 303^ plates. The plates were incubated at 37°C in C02 for -24- hrs to permit cell attachment. The medium was then discarded by inverting and shaking the plate. The cells were washed once with MEM. Dilutions of antisera were made in MEM +10# FCS + 200 units Penicillin + 200 ug Streptomycin and 10 ul were dispensed into the appropriate wells. After incubating at 37°C in C02 for 60 min , the serum dilutions were discarded and the cells were washed twice with MEM. Guinea pig complement was diluted 1*5 in cold MEM + 10# FCS + 200 units Penicillin + 200 ug Steeptomycin and 10 ul were added to the plate. The plates were incubated for an additional 30-60 min and the medium containing complement was discarded. The cells were washed twice with 10 ul of Hank's Balanced Salt Solution and the adherent cells were fixed by added acetone alcohol (1*1) for 30-60 min . The fixative agent was discarded and the plates were allowed to air dry for 5 min • The cells v/ere then stained with crystal violet and rinsed with tap water. The number of cells in each well was counted with an American Optical microscope using a grid ocular lens.
Quantitative Complement Fixation Assay* The C H 5 0 assay for titrating serum or antigen was performed according to the method described by Levine (69)• Sheep erythrocytes were

28
aseptically collected in acid citrate dextrose. The cells were allowed to age two weeks before use and were discarded after 3 months. One milliliter of the sheep erythrocytes was diluted to 15 ml with PBS and centrifuged at 700 xg for 10 min at room temperature. This washing procedure was repeated one additional time with cold PBS and then chilled with complement fixation buffer or CFB (8.16 g NaCl, 1.21 g Tris, 0.122 g MgSO^.7 H^O, 0.0^85 g CaClg.2 HgO, 1 g gelatin pH 7*^ at 4°C) . The sheep erythrocytes were adjusted to a volume of 7 ml. The concentration of the cell suspension was adjusted to 1x10 cells/ml by lysing 0.1 ml of the erythrocyte .-suspension in 1.^ ml of 0.1# Na2C0^ and reading the absorbance at 5*H nm in a Gilford Spectrophotometer. The cells were diluted with CFB until the absorbance was 0.680. This standardized cell suspension was made fresh daily.
Guinea pig complement was titrated in the following manner. Reconstituted lyophilized guinea pig complement (GIBC0) was made to the following reciprocal dilutions 10,25. 75. 100 , 125 , 150 , 175. 200 , 225, 250, 275 . 300 , 325 ,
350, 375. and 4-00 with cold CFB and kept on ice. Four-tenths ml of cold PBS and 0.2 ml of the various complement dilutions were added to chilled agglutination tubes and incubated at 37°C for 30 mins. During this incubation, 1 ml of sensitizer (It1000 in the CFB) was added to 1 ml of the standardized sheep erythrocyte suspension in 0.25 ml aliquots ’

29
every 15 sec with mixing. This was incubated at 37°C for 15 min -, diluted with 18 ml of CFB, and stored at *f°C. After the initial incubation of the complement, 0.2 ml of the sensitized red cells was added to each complement dilution and saline control. This was allowed to incubate at 37°C for 30 min and the agglutination tubes were then centrifuged for 1 min in Serofuge. The optical density (0D) of the supernatant was read at *H3 ran in a Gilford Spectrophotometer using a rapid sampler attachment. The dilution of complement that corresponds to 50% lysis was calculated using the von Krogh equation K=log ;x (K=dilutionyielding 50% hemolysis, x=dilution of complement, y=percent
°° lowest°dilution • and *=<>.158333 for lxlO9 cells). One C'H50 unit of complement was defined as the dilution ofguinea pig complement that resulted in 50% lysis of 1x10^rbc suspension after incubation at 37°C for 30 min . TwoC*H50 units of complement (usually 1*55-1*60 dilution) wereused in all antibody or antigen titrations.
Antibody was titrated in C*H50 assay by the following method. Two-fold dilutions of heat-inactivated serum in 0.2 ml CFB were made and added to this was 0.2 ml of an antigen which was diluted to a concentration that was not anticomplementary. Then, 0.2 ml of guinea pig complement containing 2 C'H50 units was added. These reagents were mixed and incubated at 37°C for 30 min . Next, 0.2 ml of sensitized

30
standardized sheep erythrocytes was added and incubated at 37°C for 30 min . The control tubes for antigen, antibody, complement, and CFB were always included in the assay. The agglutination tubes were centrifuged for 1 min in a Serofuge and the absorbance of the supernatant in each tube was read at 413 nm with a Gilford Spectrophotometer using a rapid sampler accessory. The dilution of antiserum giving 50% hemolysis was determined using the von Krogh equation (x= reciprocal of the antibody dilution). The C'H 50 units/ml was calculated by multiplying the number of C'H50 units/0.2 ml by 5.
The quantitative titration of antigen by the C*H50 assay was performed in the same manner. Dilutions of anti- gen were made in CFB, 0.2 ml of an antibody dilution containing 2 C*H50 units/0.2 ml, and 0.2 ml of complement was added. The remainder of the titration was performed according to the method described for the antiserum titration.The C'H50 units/mg protein were calculated by multiplying the number of C*H50 units/0.2 ml by 5 and dividing by the mg antigen protein/ml. The protein concentration was determined by the method of Lowry et al. (71).
Elution of Antibody from Tumor Tissue with 2.5 M NaCli Interfacial material from tumor tissue was washed with PBS and incubated with 9 volumes 2.5 M NaCl + 0.01 M phosphate buffer pH 7.2 at 37°C for 60 min . This was centrifuged at

31
10,000 xg for 10 min and the supernatant was collected.The pellet was extracted an additional time. The combined supernatants were concentrated to a volume of 1-2 ml on a Diaflo with 20 psi Ng using a PM10 membrane at 4°C.
Treatment with DNase and RNase: A stock solutionof 10 mg/ml containing DNase 2x crystallized and RNase crystalline 5x (Nutritional Biochemical Corp.) was prepared and stored at -20°C. Before use, this was thawed and diluted 1:10 with PBS. One milligram of DNase or RNase was used for every 100 ug of DNA present in a sample and allowed to incubate for 60 min at 37°C. The cellular material was centrifuged at 1000 xg for 10 min .The supernatant was discarded. This digested material was washed three times with PBS and then stored at -20°C.
Solubilization with Triton X-11*H The cellular material was pelleted by centrifugation. The supernatant was discarded and the pellet was resuspended with Trisat pH 9*0 to a protein concentration of lmg/ml. Five microliters of Triton X-ll^ (Sigma) were added for every 10 ml of protein solution (ImM Triton). This material was stirred for 30 mir at 23°C. A 0.1 ml of this mixture was collected and saved for protein analysis. This solution was then centrifuged at 113,000 xg for 6o min at ^°C in a Beckman L-2 Ultracentrifuge using an SW27 rotor. The supernatant was considered to be soluble and 0.1 ml was saved for

32
protein determination. The pellet was also saved for protein analysi s.
Detergent Removal* Several methods were examined for the removal of Triton from the solubilized protein, (i) Gel Filtration* A 2.5 x 60 cm column was packed with course Sephadex G-25 in PBS +0.2^ NaN^ at 4°C. The void column was determined with Blue Dextran 2000 (2mg/ml). Ten milliliter samples of solubilized membrane were applied to the gel bed. The flow rate was adjusted to 13ml/hr with a Buchler peristaltic pump and 6.$ ml fractions were collected with a Buchler Fractomat. (ii) Ammonium Sulfate Precipitation*The solubilized material was brought to 0.28 g ( N H ^ ) p e r ml at 0°C. The solid (NH^JgSO^ was added in small aliquots and was dissolved by stirring with a glass rod. This material was extracted twice with 3 volumes of chilled ether and the top phase of ether was discarded. The bottom phase was centrifuged at 1131000 xg for 6o min at ^-°C. The pellet was resuspended in 10 ml PBS and stored at -20°C. (iii)Ether Extraction* An equal volume of ether was mixed with soluble protein by inversion and the top phase was discarded. The bottom phase was extracted an additional time with ether and stored at -20°C. (iv) Chloroform-methanol Extraction* Chloroform-methanol solution (2*1) was mixed with the solubilized material by inversion. The top phase was collected and extracted again with the chloroform-ethanol mixture.

The top phase was then stored at -20°C. (v) Micelle Dispersion* Micelles of Triton X-114- were dispersed by adding 1 vol of 95% ethanol and 3 volumes ethylene glycol to 5 vol of solubilized protein at ^°C (*H). The detergent was removed by concentration on a Diaflo with a PM10 membrane using 20 psi nitrogen at This was diluted with 0.15 MNaCl + 0.01 M Tris at pH 7.2, concentrated to the original volume, and stored at -20°C.
Antigen-coated Erythrocytes* Ten milliliters of rabbit erythrocytes were collected in 2 ml acid citrate dextrose and washed three times in PBS. The.erythrocyte suspension was adjusted to 50%> with PBS. Bis-diazotized benzidine (BDB) was prepared by mixing **5 ml of 0.2 M HC1 with 0.23 g benzidine. Five milliliters of triple distilled water were added to 0.175 g NaNOg. The BDB and sodium nitrite were cooled to 0°C in an ice bath. Then, 5 ml of chilled sodium nitrite were slowly added to the cooled benzidine solution over a 1 min interval with stirring. The reaction was allowed to proceed at 0°C for 30 min with occasional shaking. Two milliliters of the diazonitized benzidine were pipetted into screw cap .vials and immersed in a dry-ice acetone bath. The yellow-brown solution was stored at -20°C. The antigen was coupled to the washed rabbit erythrocytes by adding reagents to an agglutination tube in the following order*1 ml antigen (0.3-5 mg protein), 0.1 ml rbc, and 1 ml BDB

34
(1*25 in phosphate "buffer). This solution was mixed and allowed to stand at 23°C for 15 min with occasional stirring. This was centrifuged at 4°C for 2 min in a Serofuge and resuspended in 2.5 ml of cold 0.15M phosphate "buffer containing 1% heat-inactivated normal rabbit serum. The washing procedure was repeated and the, antigen-coated erythrocytes were then resuspended to a 1% solution with cold PBS.
Immunodiffusioni A 1 fo suspension of Ion Agar in PBS + 0.2fo NaN^ was made by heating the solution until all the
pagar was dissolved. The hot agar was poured into 60 cm disposable Petri dishes and allowed to cool at room temperature. These agar plates were stored at 4°C until needed. Wells were cut in the agar with a No.O cork bore (a center- to-center distance of 1.5 cm) and the agar plugs were removed by aspiration. The wells were then filled with the antigen or antibody solution. The plates, were placed at 4°C and observed for lines of precipitation over the next week.
Treatment of Serum with a Low pH Buffer* Serum from tumor-bearing mice was collected by cutting open the chest cavity of ether anesthetized mice, and snipping the chambers on the right side of the heart. The blood was collected from the chest cavity with a Pasteur pipette. The blood was allowed to clot at 37°C for 30-60 min and was then

35-4 •
refrigerated from 4 hrs to overnight. The serum was then diluted 1:15 with cold 0.1 M glycine buffer at pH 2.8 andstirred for 20-30 min at 4°C . The treated serum was thendiluted with 3 vol of PBS and concentrated to the original volume on a Diaflo with an XM 100 membrane using 10 psi nitrogen. This material was diluted with 9 vol PBS andconcentrated to the original volume.
Preparation of 1'mmunoadsorbants: (i) Isolation of Serum lgG* IgG was isolated by salt fractionation. Serum was diluted with an equal volume of 0.2 M phosphate buffer pH 8.0. Anhydrous powder Na^SO^ , was added in small aliquots with stirring at 23°C until the concentration was 0.18 gm/ml. This mixture was allowed to stand at room temperature for 15 min and then was centrifuged at 10,000 xg for 15 min at 23°C. The supernatant was discarded and the pellet was resuspended to 40# the original volume in the 0.2 M phosphate buffer and dissolved in the serum solution. The centrifugation step was repeated and the supernatant was discarded. The pellet was resuspended in 20# of the original volume with the phosphate buffer. Next, 0.12 gm Na^SO^ was added for every milliliter of solution and centrifuged. This pellet was resuspended in PBS and dialyzed against 20 vol PBS for 24 hrs at 4°C with stirring, (ii) Activation of Sepharose 4B with Cyanogen Bromide (24): Preswollen Sepharose 4B gel was washed with 5 vol double

distilled water on a sintered glass filter with aspiration.The packed gel "beads were then resuspended in an equal volume of double distilled water and a volume of 2 M Na^CO^ was slowly added with stirring. The stirring rate was increased and 0.05 vol cyanogen bromide (CNBr) in acetonitrile (2mg/ml) was added. This activation of Sepharose was done in a fume hood since CNBr is a potent lacrimator. The CNBr and Sepharose 4B are stirred for 1-2 min , poured into a sintered glass funnel, and washed with 5~10 vol of the following chilled solutions* 0.1 M NaHCO^ at pH 9.5 double distilled water, and 0.1 M bicarbonate buffer + 0.5 M NaCl at pH 8.3. The CNBr-activated Sepharose 4B was filtered to a moist cake and transferred to a 1 oz. French square bottle with a spatula. (iii) Conjugation of Rabbit IgG to CNBr-Activated Sepharose 4b* The rabbit IgG that was obtained by salt fractionation was diluted in 0.5 M NaCl + 0.1 M bicarbonate buffer at pH8.3 (coupling buffer) to make the concentration 10 mg protein/ ml as measured by the absorbance at 280 nm (1 mg rabbit IgG is equivalent to 1.46 0D). An equal volume of the diluted protein was added to the activated gel and allowed to mix for 20 hrs at 4°C using a magnetic stirrer. The IgG conjugated gel was filtered on a sintered glass filter and the effluent was collected in a test tube for spectrophotometer analysis to determine the amount of protein conjugated. The

gel was washed with 5 vol of 0.5 M NaCl + 0.1 M bicarbonate buffer at pH 8.3 and then the moist cake was placed in a 1 oz. French square bottle. One volume of 1M ethanolamine pH9.0 was stirred with the gel to block any remaining CNBr active sites. An additional washing was done with the coupling buffer. Adsorbed protein was eluted with five alternate washings of 2-3 vol 1.0 M NaCl + 0.1 M acetate buffer at pH 4.0 and 2-3 vol 1.0 M NaCl + '0.1 M bicarbonate buffer at pH 8.0. After these extensive washings, the IgG conjugated Sepharose 4B was washed once with PBS +0.2^ NaN^ and then stored in this medium at 4°C . (iv) Affinity Chromatography1 A K 9/15 column (Pharmacia) was packed with the IgG-conjugated Sepharose 4B. PBS containing 0.2f* NaN^ was used as the eluant. A 1-2 ml of a soluble protein sample was applied to the gel bed and allowed to run at a flow rate of 6 ml/hr. Aliquots of 2.5 nil were collected on a Buchler Fractomat and the absorbance of the eluant was monitored at 280 mm with a Perkin Elmer Spectrophotometer. The transmittance was recorded on a Kipp Zonen Model B10 chart recorder. The adsorbed protein was eluted by using 2.5 M NaCl + 0.01 M P0^ at pH 7.2 or 0.1 M glycine buffer at pH 2.8. The eluted material was adjusted to a pH of 7.0 and was desalted by diluting with double distilled water or PBS+NaN^ and concentrating on a Diaflo with a PM 10 membrane using 20 psi nitrogen.

38
A batch procedure was also attempted. The gel was drained to a moist cake and transferred to a 1 oz. French square bottle. Two milliliters of the soluble protein were added and stirred at ^°C for 18-2^ hrs with a magnetic stirrer. The eluant was harvested by suction on a sintered glass filter and saved. The adsorbed protein was eluted by adding 1 vol of chilled 0.1 M glycine buffer and stirring at 4°C for 30 min . The dissociated protein was then harvested, and the pH was adjusted to 7*0. This was diluted with 9 vol of PBS and concentrated to the original volume with a Diaflo using a PM 10 membrane. The protein concentration of all fractions obtained by these affinity chromatography procedures was measured by the method of Lowry et al.(?l). All samples were stored at -20°C.

RESULTS
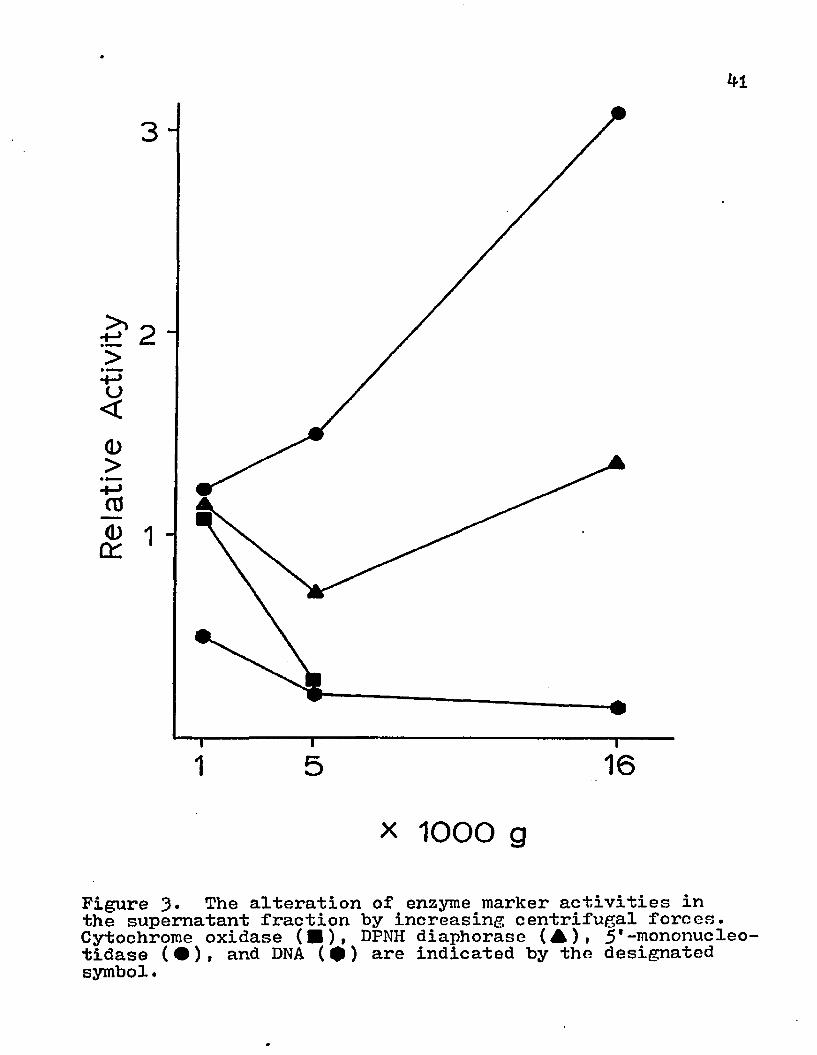
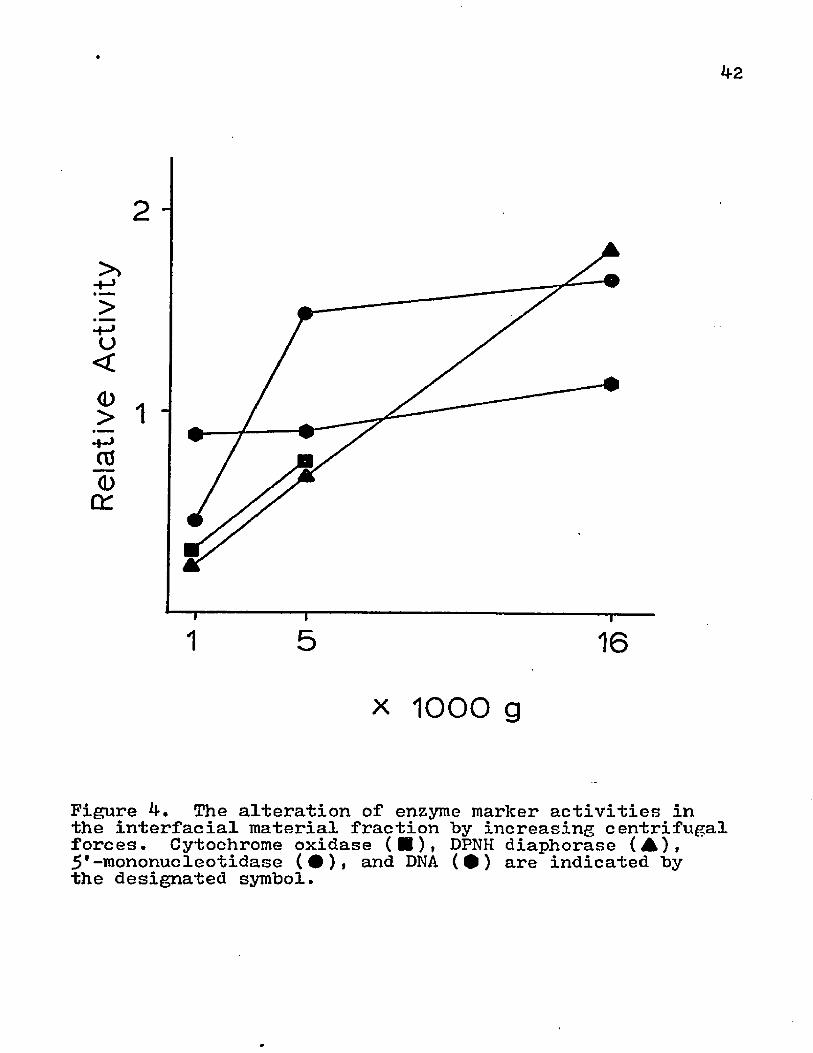
The effects of various centrifugational forces on the subcellular contents of the supernatant, interfacial material, and pellet were investigated. The homogenate was centrifuged at 1000, 50°0» or 20,000 xg for 15 min . The supernatant was saved for analysis and the pellet material was partitioned on an aqueous two-phase polymer system (Fig. 2). The redistribution of various subcellular organelles by altering the centrifugal forces is depicted in Figs. 3i and 5* With higher centrifugation speed, the supernatant displayed a decrease in the amounts of endoplasmic reticulum, mitochondria, and nuclei; whereas, the surface membrane activity was increased (Fig. 3)• The cellular material that was removed from the supernatant was transferred to the interfacial material (Fig. k) or the pellet (Fig. 5)• The DNA content of the interfacial material remained relatively constant for the various centrifugal speeds, but there was a marked transfer of nuclei to the pellet. The interfacial material obtained more of the mitochondria and endoplasmic reticulum from the supernatant than the pellet. Even though there was a marked increase in
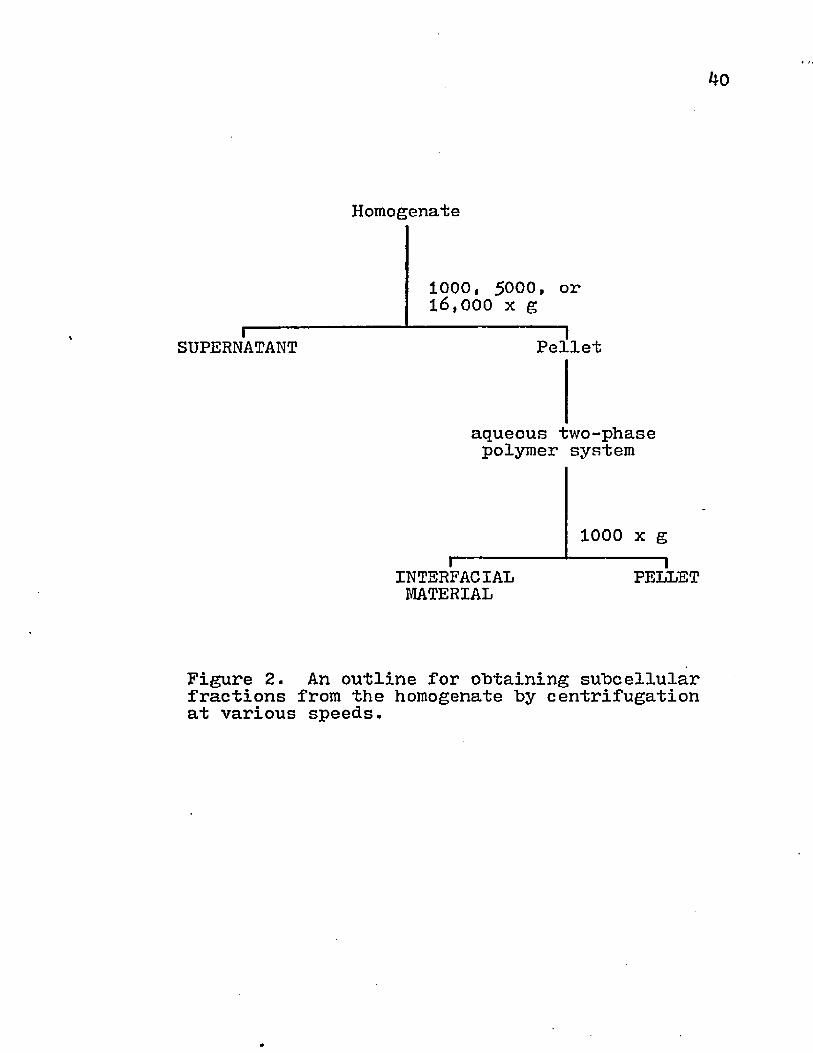
4o
i-----SUPERNATANT
Homogenate
1000, 5000, or16,000 x g 1Pellet
aqueous two-phase polymer system
I------INTERFACIALMATERIAL
1000 x g 1PELLET
Figure 2. An outline for obtaining subcellular fractions from the homogenate by centrifugation at various speeds.
kl
o<<D>+->aj
DC
3
1
1 16x 1000 g
Figure 3* The alteration of enzyme marker activities in the supernatant fraction by increasing centrifugal forces. Cytochrome oxidase (■), DPNH diaphorase (A)» 5'-mononucleotidase (•)» and DNA (•) are indicated by the designated symbol.
42
>>>u<<D>* MW
l | * J
rgd>Cd
1 16X 1000 g
Figure 4. The alteration of enzyme marker activities in the interfacial material fraction by increasing centrifugal forces. Cytochrome oxidase (■)• DPNH diaphorase (A),5'-mononucleotidase (•), and DNA (•) are indicated by the designated symbol.

o<(D>+->J?<L>
OH
1 16
x 1000 g
Figure 5* The alteration of enzyme marker activities in the pellet fraction by increasing centrifugal forces. Cytochrome oxidase (■)» DPNH diaphorase (▲), 5*-mononucleotidase (#), and DNA (0 ) are indicated by the designated symbol.

Zi4
surface membrane content in supernatant with higher centrifugal forces, this enzyme activity was also present in the interfacial material and pellet.
Since the supernatant retained 5'-mononucleotidase activity after centrifugation at 16,000 xg for 15 min , it was centrifuged at 113,000 xg for 6o min. to determine if this surface membrane marker activity could be sedimented.This ultracentrifugation removed most of the surface membrane vescicles from the supernatant (Table 2).
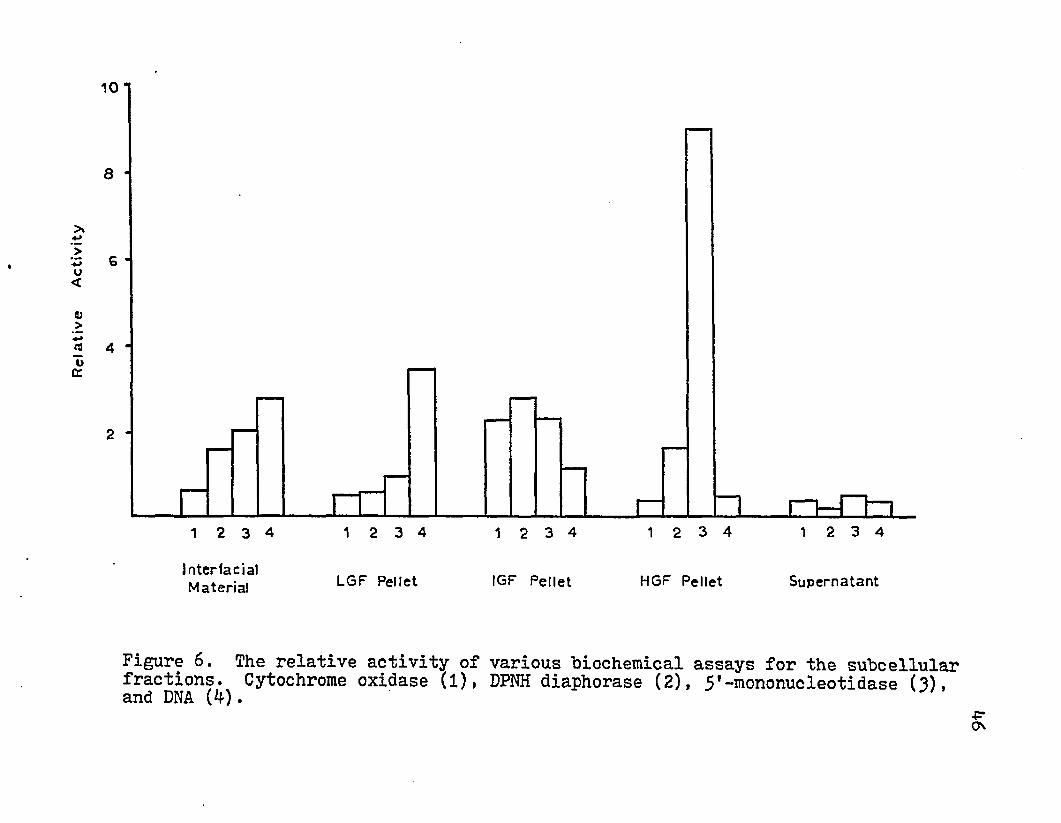
The above results were used to construct a protocol for subcellular fractionation by using the aqueous two-phase polymer system and differential centrifugation (Fig. 1). The results of various enzyme and biochemical assays are given in Table 2. The amount of mitochondria (cytochrome oxidase) activity and rough endoplasmic reticulum (DPNH diaphorase) were greatest in the IGF (16,000 xg) pellet. Nuclei were found in the greatest quantity in the LGF pellet but also appeared in the interfacial material and IGF pellet. The interfacial material, IGF pellet, and HGF pellets contained significantly increased 5*-mononucleotidase activity (surface membrane marker). The HGF contained almost a nine-fold increase in activity. ( Fig. 6) and provided 1-2 mg protein pergwetwttissue. The supernatant contained negligible amounts of the enzymatic and biochemical markers.
Xenogeneic and syngeneic animals were immunized with tumor antigens and the antisera were compared for antibody-
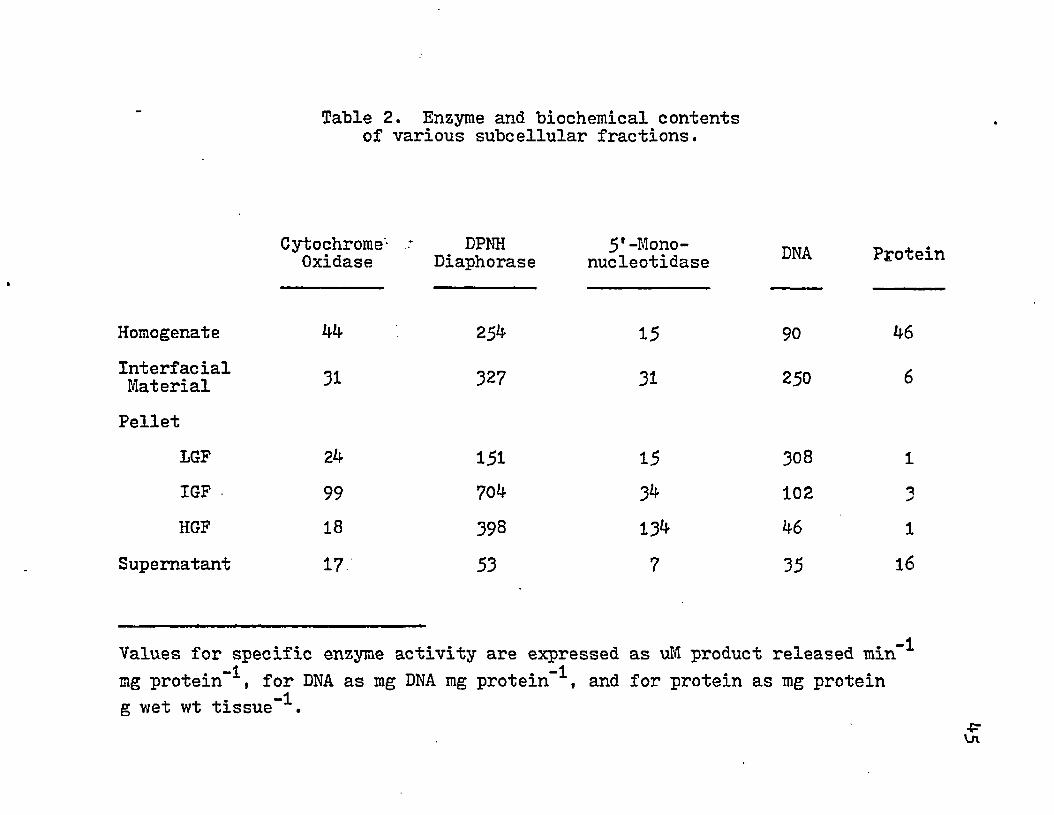
Table 2. Enzyme and biochemical contents of various subcellular fractions.
InterfacialMaterial
Cytochrome- DPNH 5' -Mono- _ . .Oxidase Diaphorase nucleotidase rroxein
Homogenate 44 254 15 90 46
31 327 31 250 6Pellet
LGF 24 151 15 308 1IGF 99 704 34 102 3HGF 18 398 134 46 1
Supernatant 17 53 7 35 16
Values for specific enzyme activity are expressed as uM product released min-1 -1mg protein , for DNA as mg D M mg protein , and for protein as mg protein
-1g wet wt tissue .
Rel
ativ
e A
ctiv
ity
10 1
8
6 •
j = L1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4
m-n-11 2 3 4
Inter facial Material LGF Pellet IGF Pellet HGF Pellet Supernatant
Figure 6. The relative activity of various 'biochemical assays for the subcellular fractions. Cytochrome oxidase (1), DPNH diaphorase (2), 5'-mononucleotidase (3), and DNA (^).
•p*ON

mediated cytotoxicity with 4198 target cells (Fig. 7). The rabbit anti-C^H fibrosarcoma tissue was highly cytotoxic (100#) and had a titer of 1*160. The immune isologous serum did not achieve the high cytotoxicity (40-47#) nor the titer (lt4o) that was attained with the heterologous antisera. Antisera from both syngeneic C^H mice immunized with 4198V and tumor-bearing animals had equivalent titers (li4o) and attained 40-47# cytotoxicity in this assay. When sera from mice with a progressive tumor were first incubated with the target cells and then the immune syngeneic serum was added, the percent cytotoxicity was lower than the immune serum alone but the titer remained 1*40.
When the isologous antiserum was titrated in a C'H50 assay, no complement-fixing activity was detected with immune serum, but it was found at a low dilution in the serum of tumor-bearing C^H mice (Table 3)■
Sera from mice with a progressive tumor may contain immune complexes. These sera were treated with a glycine buffer at pH 2.8 which will dissociate antigen-antibody complexes. Ultrafiltration v/ith an XM100 membrane was used to remove antigen (mol wt <100,000) but retain the antibody (mol wt ^ 150,000). This treated serum did not display complement-fixing activity (Table 3).
An attempt was made to determine if circulating antibody was present on the fibrosarcoma material from a subcellular fraction. The interfacial material was incubated with 2,5 M
48100
80 -
60 -
o
0
20
3 2 0 6 4 0 1 2 8 0 2 5 6 04 0 8 0 1 6 02010
Reciprocal of Dilution
Figure 7* The comparison of antisera from rabbit anti-C^H fibrosarcoma (•)» C^H anti-fibrosarcoma.(O ), tumor-bearing C^H mice (■)> and tumor-bearing + immune C^H mice ( □ ) in the antibody-mediated microcytotoxicity assay using 4198V target cells.

Table 3* Titration of antisera from immune and tumor-bearing C«H mice by c omplement fixati on. ^
Serum fromi C*H50 UNITS/ml
Immune C^H Mice 0
Tumor-bearing C^H Miceuntreatedtreated with a low pH buffer
21
0

Table k. Treatment of interfacial material from tumor tissue with 2.5M NaCl.
Treated■*Supernatant 0
Pellet 153*
Untreated 184+
* C'H50 Units/ml + C*H50 Units/mg protein

51
NaCl for 60 min at 37°C and the eluted material was concentrated on a Diaflo with a XM100 membrane. The eluant did not display any complement fixation activity (Table *0 and no lines of precipitation could be detected when it was tested by immunodiffusion with goat anti-mouse 2fG or IgG. However, this method may not have been efficient in dissociating the antibody from the surface membrane. This high ionic strength treatment did not significantly alter the antigenic activity of the interfacial material. Other methods of eluting antigen-antibody complexes can be deleterious to labile antigen. When the serum of the tumor- bearing mice was assayed for the presence of tumor antigen in the serum, no activity was detected. The lack of activity may possibly be due to the fact that the absorbed rabbit antisera detected an antigen specificity different from the one that might be circulating in the serum.
Rabbits and guinea pigs were immunized with the HGF pellet from normal and tumor tissue. Before the animals were exsanguinated, they were given an intradermal injection with 100 ug protein of the immunogen. The skin test was consid- ered positive if the induration was 2 .‘5 mm or >78 • % 111111 after *J-8 hrs. The rabbits and guinea pigs gave positive skin tests (Table 5)• The area of induration was greater for the tumor HGF than the normal. This may represent a qualitative difference in the determinants present in these

52
antigenic sources. The difference also appears in the serum titration of the immunized animal (Table 5)•
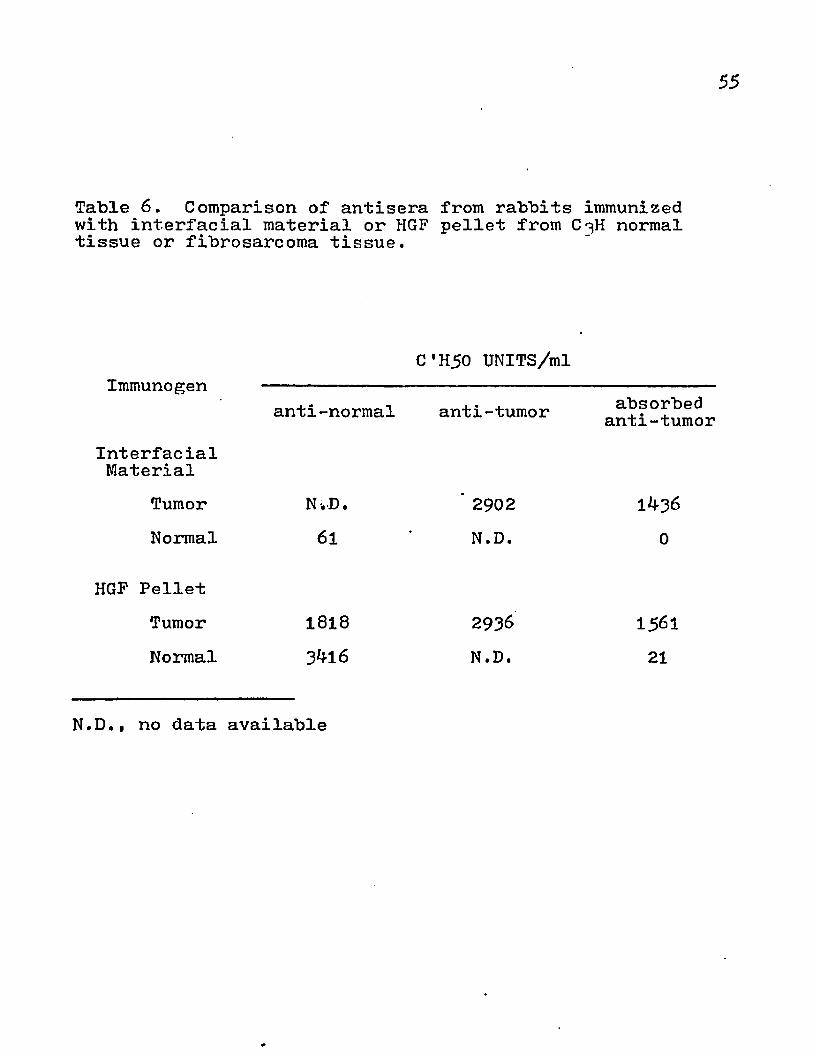
The specific activity of the various enzyme assays revealed that the HGF pellet and interfacial material would he suitable for injection because of their high content of plasma membrane and negligible contamination by other cellular organelles. The LGF pellet and IGF pellet were not used, although there was 5'-mononucleotidase activity present because of their high content of other marker enzyme activities. When rabbits were immunized with interfacial material or HGF pellet from the fibrosarcoma, the titers of the antisera were comparable for both groups of animals (Table 6). Furthermore, absorption of these antisera with interfacial material from normal tissues (liver, kidney, muscle) removed the reactivity to normal tissue but not tumor tissue. The quantitative differences between unabsorbed and absorbed sera may be due to the qualitative difference in the antigens present in the normal and tumor cells. There were also quantitative differences in the amount of normal antigens present in tumor and normal tissues.
The effects of antigenic fractions on the titer of antisera from rabbits that were immunized with tumor interfacial material were compared. Higher complement fixation activity was obtained with the homogenate rather than the purified HGF pellet (Table 7).
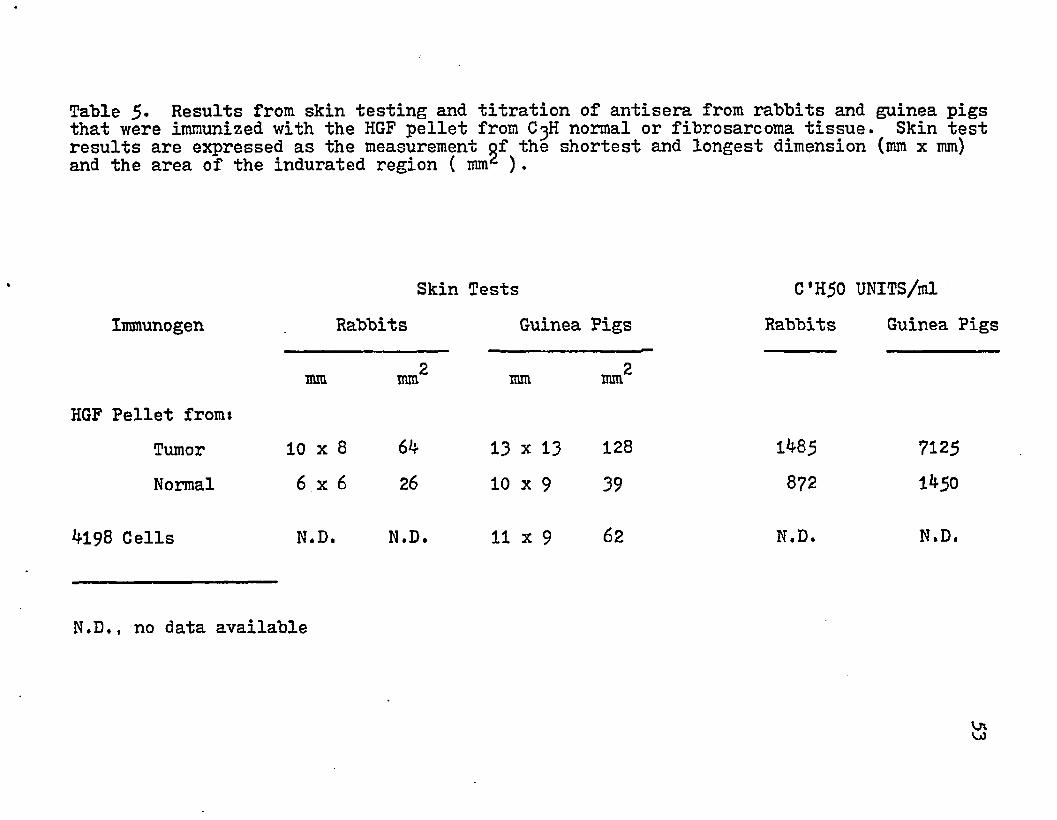
Table 5- Results from skin testing and titration of antisera from rabbits and guinea pigs that were immunized with the HGF pellet from C3H normal or fibrosarcoma tissue. Skin test results are expressed as the measurement of the shortest and longest dimension (mm x mm) and the area of the indurated region ( mm2 ).
ImmunogenSkin Tests
Rabbits Guinea PigsC'H50 UNITS/ml
Rabbits Guinea Pigs
mm 2mm mm mm2mmHGF Pellet from*
Tumor 10 x 8 64 13 x 13 128 1485 7125Normal 6 x 6 26 10 x 9 39 872 1450
4198 Cells N.D. N.D. 11 X 9 62 N.D. N.D.
N.D., no data available

5**
The antigenic content of the various subcellular fractions were quantitated by the C'H50 assay. Rabbit antisera were used to assay for normal and tumor-defined antigens (Table 8). The HGF pellet contained the highest activity for both normal and tumor antigen. This fraction provided a five-fold increase in tumor specific antigen. The presence of cell-mediated response was determined by skin testing.All the fractions gave a positive response and the areas of induration were comparable to that produced by fibrosarcoma cells.
The titration of the subcellular fractions with absorbed rabbit anti-tumor sera demonstrated the presence of tumor-defined antigens in the various subcellular fractions. The presence of tumor specific transplantation antigen was assayed by immunizing groups of C^H mice with the various subcellular fractions and then challenging with 1x10^ tumor cells. No difference in tumor appearance was noted for immunized or control animals except for with the low dose of HGF pellet (Table 9)• This quantity of HGF pellet delayed the appearance of a palpable tumor (Fig. 8). High doses of this subcellular fraction did not result in protection. The percent of protection and the quantity of antigenic equivalent C'H50 units injected demonstrated a dose-response relationship (Fig. 8). Injection of suboptimal and superthreshold amounts of C H 5 0 units of antigen provided no protection. An
55
Table 6. Comparison of antisera from rabbits immunized with interfacial material or HGF pellet from C^H normal tissue or fibrosarcoma tissue.
Immunogen
InterfacialMaterial
TumorNormal
C'H50 UNITS/ml
anti-normal anti-tumor absorbed anti-tumor
N-.D.61
2902
N.D,
HGF PelletTumor 1818 2936 15^1Normal 3bl6 N.D. 21
N.D,, no data available

56
Table 7* Comparison of antigenic fractions for the titration of antisera from rabbits that were immunized with interfacial material from the C^H fibrosarcoma.
C*H50 UNITS/ml
Homogenate 6144HGF Pellet 2902

57
Table 8. The antigenic activity of subcellular fractions from the murine fibrosarcoma.
C'H50 UNITS/mgAntigen
HomogenateXnterfacialMaterialPellet
LGFIGFHGF
SupernatantM 98 CellsSerum from Tumor-bearing C3H Mice
Control
Rabbit a n t i ^ H Normal Tissue
97130
2k
12710682k
N.D.
N.D.
N.D.
Absorbed Rabbit anti-C^H Tumor Tissue
5526
1713626920
N.D.
0
N.D.
Skin Test
2mm
138iko
10 7 N.D.162
N.D.113
N.D.
0
N.D., no data available

58
intermediate dose of antigen resulted in no appearance of tumor on day 16 and was able to delay the appearance of a palpable tumor 12 days beyond that of the control groups.This correlation can further be extended to the specific activity of the surface membrane marker (Table 9)•
The effects of various conditions on the loss of antigenicity were also studied. Tumor HGF pellet was stored at various temperatures (4°C and -20°C) and in different media (PBS, ST medium, and PBS + glycerol). Storage at 4°C appeared to be less deleterious to the antigen than -20°C (Fig. 10). The samples stored at -20°C were thawed every time a titration was performed and this probably contributed to antigen destruction. Comparison of the three media demonstrated that PBS did not help to preserve the antigenicity as well as the ST medium. Both sucrose and glycerol are cryo- protective agents and their stabilizing effect was demonstrated in this experiment. An additional aspect of the experiment was the importance of adding a bacteriostatic agent to the medium. All the media contained 0.2JS sodium azide. If this was omitted, bacterial contamination rapidly destroyed the antigen activity in the samples stored at 4-°C.
Various parameters may effect the release of protein from the lipid matrix. Solubilization with a nonionic detergent was performed under the following standard conditions. The protein concentration was adjusted to 1 mg/ml in 0.005 M
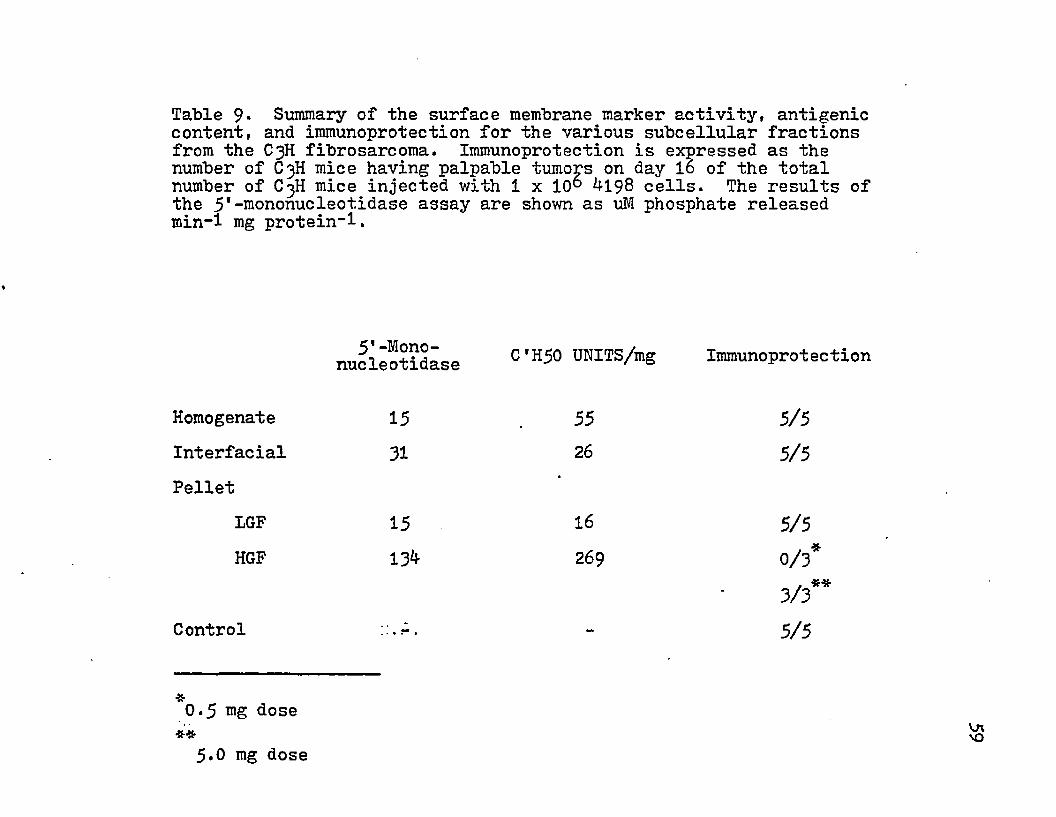
Table 9* Summary of the surface membrane marker activity, antigenic content, and immunoprotection for the various subcellular fractions from the C3H fibrosarcoma. Immunoprotection is expressed as the number of C3H mice having palpable tumops on day 16 of the total number of C3H mice injected with 1 x 10° M 98 cells. The results of the 5*-mononucleotidase assay are shown as uM phosphate released min-1 mg protein-1.
C'H50 UNITS/mg Immunoprotectionnucleotidase
Homogenate 15 55 5/5Interfacial 31 26 5/5Pellet
IGF 15 16 5/5HGF 13^ 269 0/3*
. **3/3Control ::.A, - 5/5
0.5 mg dose 5.0 mg dose
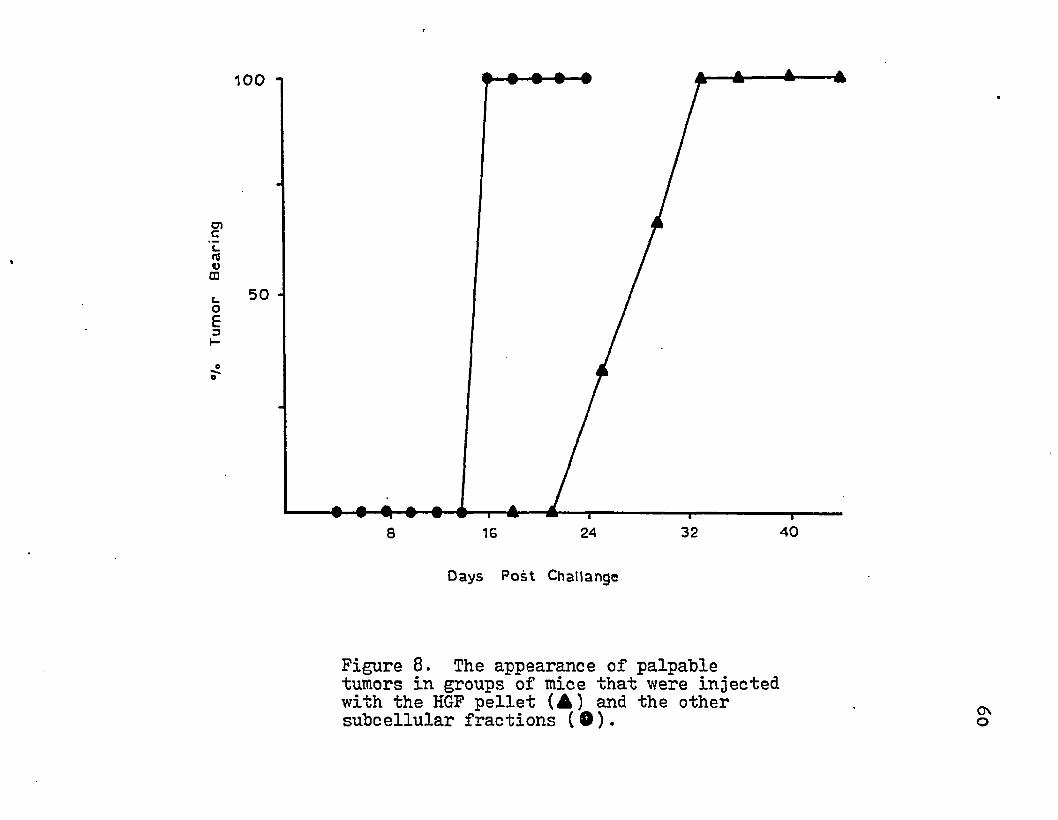
Tum
or
Bea
ring
100 n
50 ■
0o
4032248 16
Days Post Challange
Figure 8. The appearance of palpable tumors in groups of mice that were injected with the HGF pellet (A) and the othersubcellular fractions (®). o
Imm
uno
prot
ectio
n
100 •
5 0 -
oo
Antigen Equivalent C 'H 50 Units
Figure 9* The dose dependent relationship between the number of antigen equivalent C*H50 units used for immunization and the induction of immunoprotection in C3H mice against the challenge with ^198 cells.

100 1
80 -
Time (days)
0\Figure 10. The effects of temperature ( 4° C, open symbols and -20° C, closed Msymbols) and different media conditions ( • ST, BPBS, and A PBS + glycerol) on the antigenic activity of the HGF pellet from the fibrosarcoma tissue.

Tris buffer at pH 9.0 (80). Triton X-11*J- was added to a final concentration of ImM (62) and this mixture was stirred for 30 min : at 23°C. After solubilization, a protein sample was collected for protein determination and then centrifuged at 113*000 xg for 60 min at 4°C. The supernatant contained the soluble material and was also assayed for protein content. The percent solubilization was calculated by dividing the mg protein/ml of the supernatant by mg protein/ml of the sample before centrifugation and multiplying by 100. The pellet from this ultracentrifugation was also measured for protein. The effects of pH, osmolality, detergent concentration, temperature, and time were investigated.
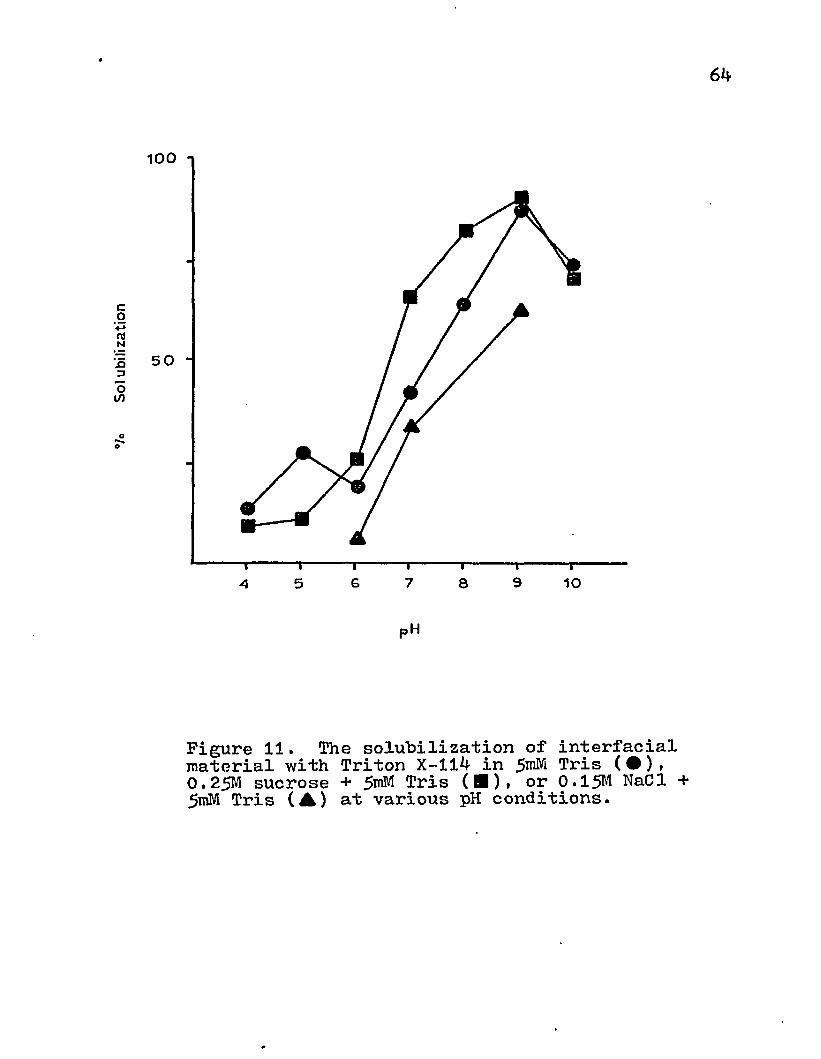
As the pH was increased, more protein was released from the interfacial material (Fig. 11). The optimum solubilization appeared at pH 9*0. This relationship was also demonstrated with Tris-buffered 0.25 M sucrose and 0.15 M NaCl.
There was a quantitative difference in the amount of protein extracted from the subcellular fraction with varying concentrations of Tris. A significant decrease in solubilization occurred with increasing concentrations of Tris (Fig. 12). The addition of sucrose to the Tris buffer did not appear to enhace or decrease the amount of solubilization for a given concentration of Tris although it did increase the osmolality of the medium.
The release of protein from the interfacial material
Sol
ubili
zatio
n
6i\
100 i
5 0
ao
95 7 10G 8A
pH
Figure 11. The solubilization of interfacial material with Triton X-114 in 5 ^ Tris ( • )( 0.25M sucrose + Tris (■). or 0.15M NaCl + 5mM Tris (A) at various pH conditions.

Sol
ubili
zatio
n
65
1 0 0 *i
5 0
oo
100 150 200 25 0 3 0 0 3 5 05 0
miliiosmolais
Figure 12. The effects of Tris osmolality on the solubilization of interfacial material with ImM Triton X-ll^- at pH 9*0. Tris (#) and 0.25M sucrose + Tris (■)•

66
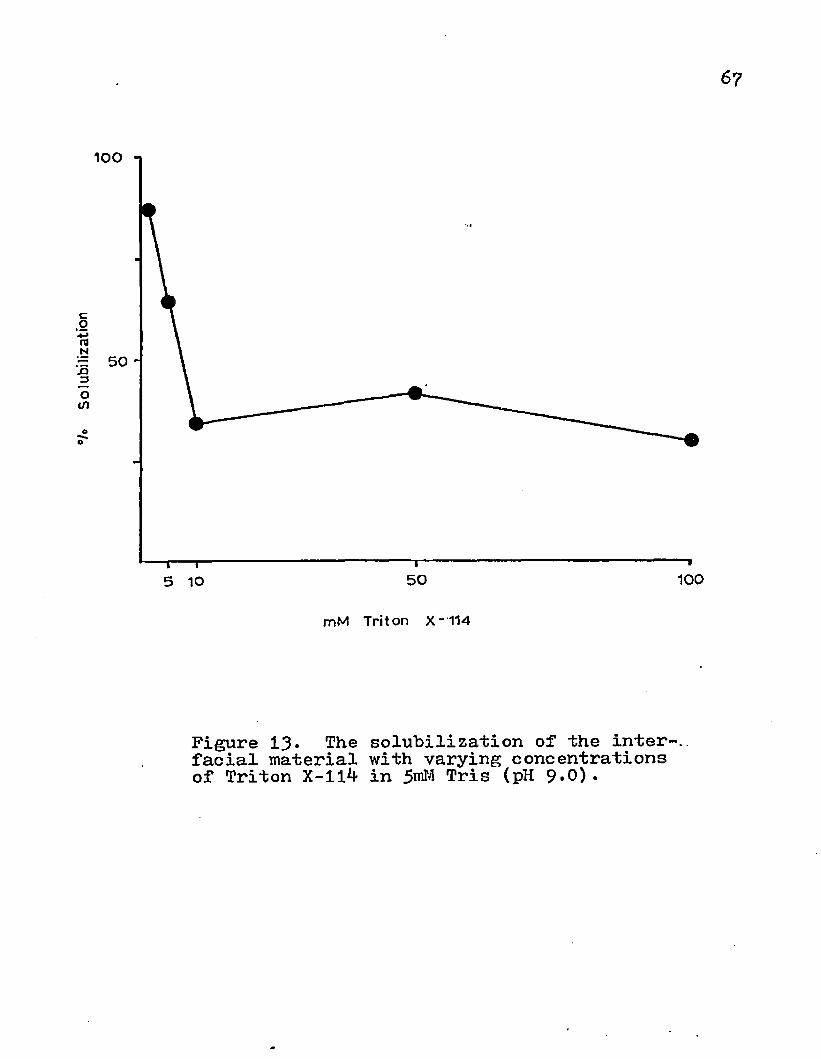
membranes was also dependent on Triton X-114 concentration. As increasing amounts of this nonionic detergent were added to a constant amount of protein, the percent solubilization decreased id the 1-10 mM range and remained below 41# with 10, 50, and 100 mM (Fig. 13).
The time and temperature parameters were also investigated. Increases in temperature produced minimal changes in the amount of protein solubilized with ImM Triton X-114 at pH 9.0 (Fig. 14). The extraction of proteins from the lipid matrix was complete within 60 min (Fig. 15)•
Divalent cations may contribute to the stability of membranes. These cations may prevent the extraction of proteins from the matrix and so the solubilization medium containing 2mM EDTA was used to chelate the divalent cations. This concentration of EDTA had little effect on the amount of protein released with ImM Triton X-114 at pH 9-0 (Table 10) .
There has been noted that the presence of nucleic acid in the membrane sample can interfere with the solubilization by Triton X-114 (60) . The interfacial material was treated with DNase and RNase to remove these nucleic acids from the sample. This treatment did not alter the antigenic activity. When this sample was used for solubilization, no changes in the amount of protein released were noted.
The protein content was measured by reading the
®/o
Sol
ubili
zatio
n67
IOO
50
1005 0
mM Triton X “'114
Figure 13. The solubilization of the interfacial material with varying concentrations of Triton X-ll^ in Tris (pH 9*0)•

°/o
Sol
ubili
zatio
n68
100
50
2010 3 0 4 0 5 0
Temp (C °)
Figure 14. The solubilization of interfacial material with ImM Triton X-ll^ in 5roM Tris (pH 9.0) at various temperatures.
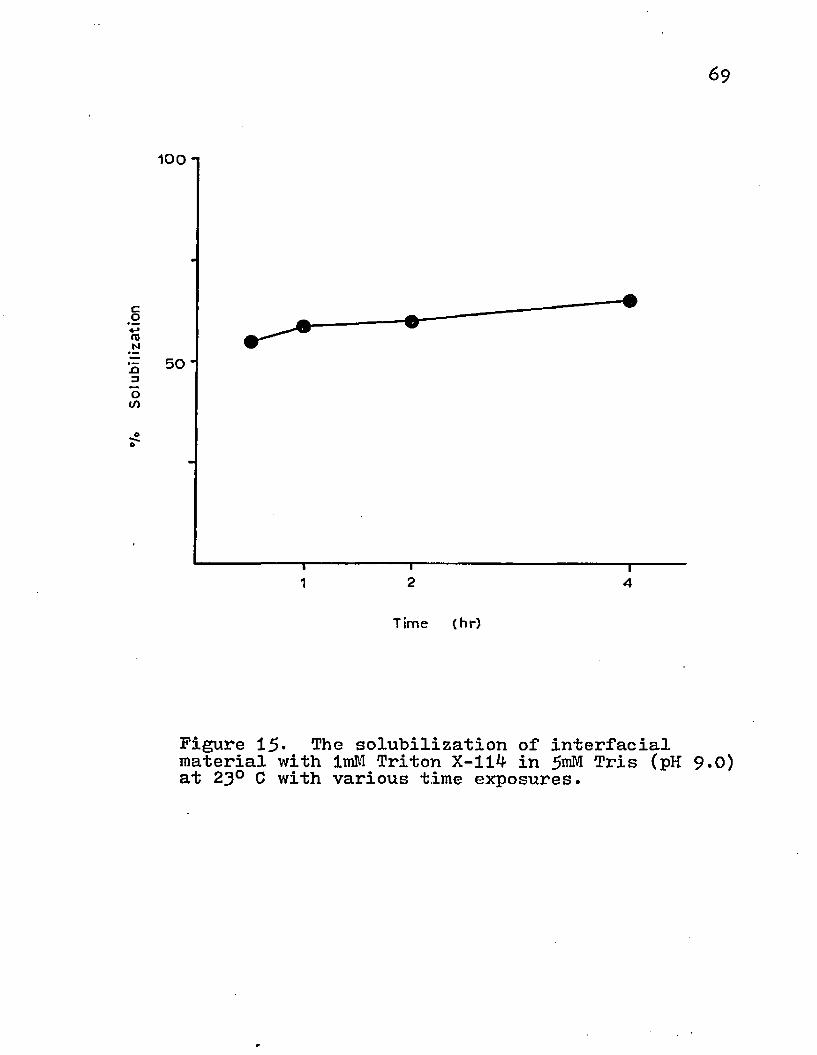
Sol
ubili
zatio
n
69
1001
50 ”
O
21 4
Time (hr)
Figure 15* The solubilization of interfacial material with imM Triton X-114- in jJmM Tris (pH 9.0) at 23° C with various time exposures.

Table 10. The solubilization of the interfacial material with ImM Triton X-114- in 5mM Tris (pH 9.0) containing 2mM EDTA at 23° C for 1 hr.
% Solubilization
Solubilization Medium 57Solubilization Medium + 2mM EDTA 63

absorbance at 280 nm in an attempt to replace the Lowry assay. Triton X-114 was found to interfere with protein absorbancy by this method in comparison to the Lowry assay. The OD of Triton X-114 at various wavelengths closely paralleled that of the protein (Pig. 16) and was attributed to the presence of a phenolic ring in the chemical structure of this nonionic surfactant. This ring structure absorbs the same wavelengths as phenylalanine, tyrosine, and tryptophan residues in proteins. The optical density of a ImM solution of Triton X-114 could not be adjusted to zero because of clouding effects at 23°C. This turbidity could be removed by cooling the detergent to 4°C.
Since the presence of detergent might have adversly affected the various immunological assays in which intact cells were used, several methods were examined for reducing the concentration of Triton X-114 in the soluble antigen preparation (Table 11). Gel filtration with Sephadex G-25 resulted in a slight reduction of antigen activity. The extraction procedures reduced the biological activity drastically. This may be attributed to the organic nature of these solvents and their ability to denature proteins. Precipitation with '(NHji gSO and ether resulted in a three-fold increase in antigen activity per mg protein. This salt frac tionation procedure selectively precipitated the antigens from this mixture. Treatment of soluble protein containing
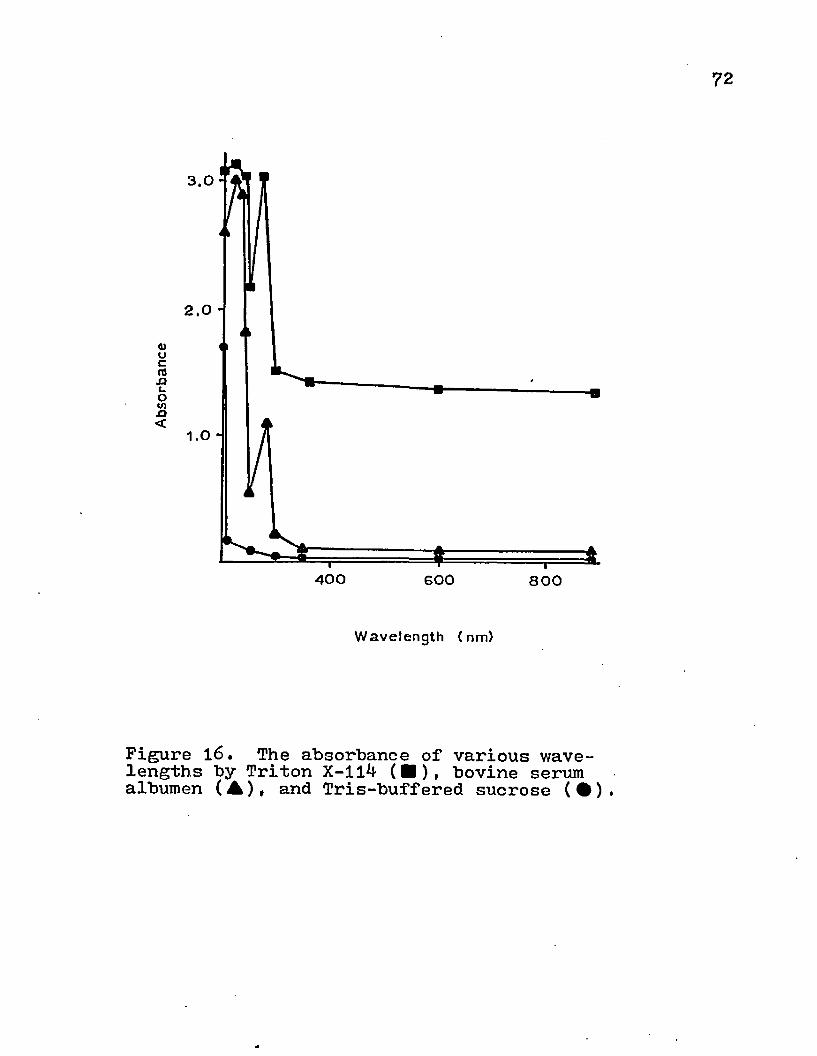
72
«)caJJ3uOV)n<
4 0 0 6 0 0 8 0 0
Wavelength (nm)
Figure 16. The absorbance of various wavelengths by Triton X-114 (■), bovine serum albumen (A)t and Tris-buffered sucrose (•).

73
detergent with Tris-buffered 10$ ethanol and 30$ ethylene glycol has been shown to remove 95$ of the surfactant (*H'); however, this procedure resulted in a loss of 15$ of the antigenic activity. In addition, the supernatant and pellet from the solubilized interfacial material were analyzed for C*H50 units/mg. More than 9^$ of the antigenic protein was released by the detergent treatment and less than 6$ was contained in the sediment (Table 11).
Antigen-coated erythrocytes were used to titer heterologous antisera to normal mouse tissue. The soluble protein was attached to the surface of erythrocytes with BDB and the tube and microplate methods for testing passive hemagglutination were used for this titration. For both methods, the 1»25 dilution of BDB resulted in the highest hemagglutination activity (Table 12). Furthermore, the microplate assay was more sensitive than the tube method.
The soluble antigen also demonstrated activity in cell- mediated responses. Guinea pigs previously immunized with the interfacial material were skin tested with the extracted mater-
i
ial. The soluble protein produced an area of induration comparable in size to the immunogen (Table 13); however, the detergent containing materials produced a greater response than the sample with the detergent removed. The area of induration of the detergent control might explain this difference in size.

74
Table 11. Titration of soluble protein in which the Triton X-114 has been removed by various procedures.
C ’H50 UNITS/mgRabbit anti-C3H Normal Tiaaue Rabbit anti-C3Hlissue Fibrosarcoma
Sephadex G-25 269Extraction with:
Ether 140Chloroform-methanol
(NH4)2S0^ Ppt. + ether
Untreated Soluble Protein
Pellet from the C ent ri fugat i on
1*5
1055
Ethylene Glycol + 228 247Ethanol
338
21

75
Table 12. Titration of rabbit anti-C^H normal tissue by passive hemagglutination using the tube and microplate tests. Soluble protein from normal C^H tissue was conjugated to rabbit erythrocytes witn various dilutions of bis-diazotized benzidine.
BDB Titer ofDilution Antiserum
Tube Method It 10 01*25 It 6^
1i 50 1i 32
Microplate Method 1*10 01 125 112561*50 1:128
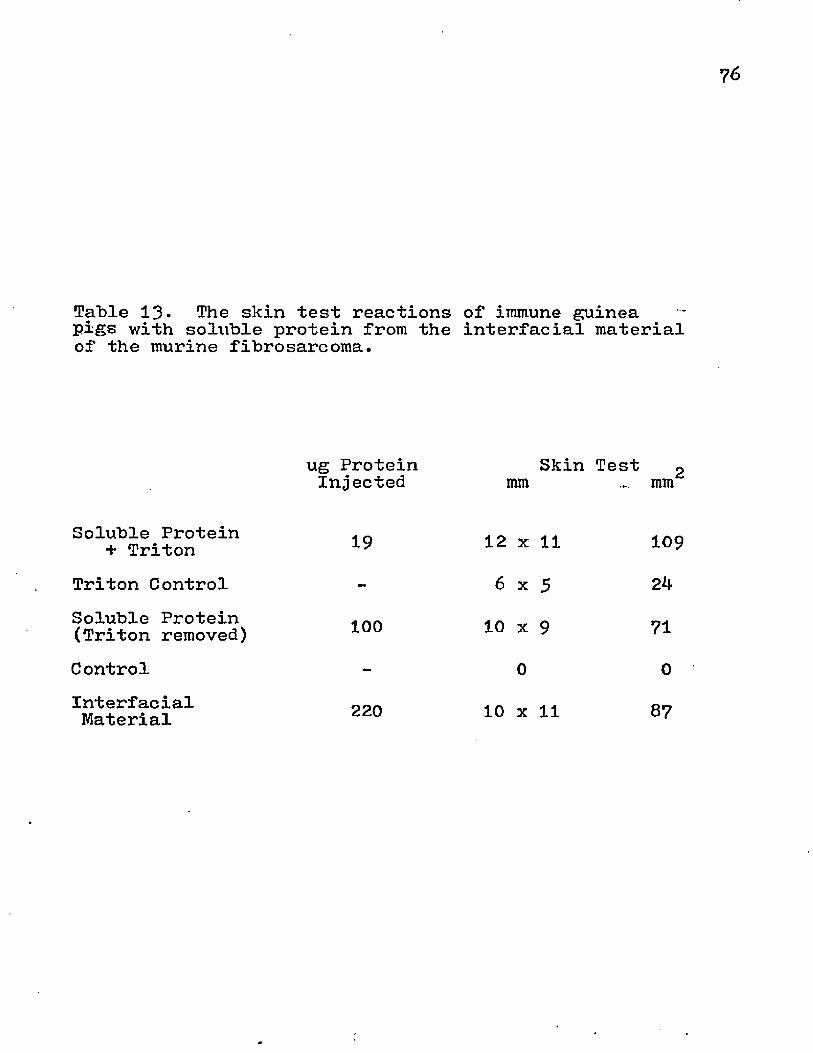
Table 13. The skin test reactions of immune guinea pigs with solxible protein from the interfacial material of the murine fibrosarcoma.
Soluble Protein + Triton
Soluble Protein (Triton removed)
InterfacialMaterial
ug Protein Skin Test gInjected mm mm
19 12 x 11 109
Triton Control ~ 6 x 5 2^
100 10 x 9 71Control - 0 0
220 10 x 11 87

The rabbit antisera which were produced to the C^H normal and tumor tissue were fractionated with sodium sulfate to obtain the igG. Immunodiffusion analysis showed that the IgG was not contaminated by other gammaglobulins but did contain other serum constituents (Pig. 17)* IgG was titrated and demonstrated the ability to fix complement. The IgG from tumor antisera was absorbed with che interfacial material from normal tissue and displayed specificity for only the tumor material and not normal tissue (Table 1*0 .
The rabbit anti normal tissue o.r absorbed rabbit antitumor IgG were conjugated to a Sepharose matrix. The absorbance of the IgG was measured before and after conjugation and approximately 86% of the protein was conjugated to the CNBr-activated Sepharose *KB.
Affinity chromatography was investigated as a method for isolating normal or tumor specific antigens. When soluble antigens from normal tissue were applied to a column packed with rabbit anti normal tissue Sepharose, an initial fraction came through the gel bed (Fig. 18). After the absorbance had returned to the baseline, 2.5 M NaCl was added. This was done to increase the ionic strength and elute any material which may have nonspecifically adsorbed to the gel matrix. A small change in optical density was noticed with this treatment. This was the result of mixing
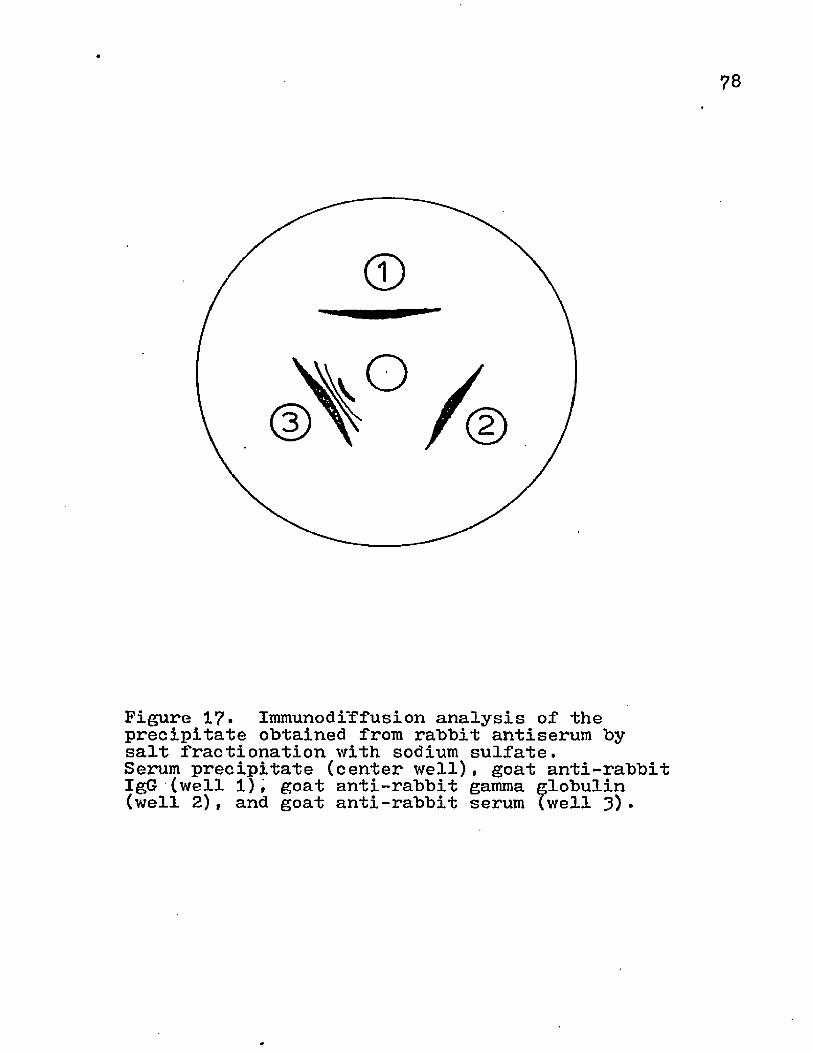
7 8
Figure 17. Immunodiffusion analysis of the precipitate obtained from rabbit antiserum by salt fractionation with sodium sulfate.Serum precipitate (center well), goat anti-rabbit IgG (well 1), goat anti-rabbit gamma globulin (well 2), and goat anti-rabbit serum (well 3)•

79
Table 1^. The antibody activity of serum and IgG fraction from rabbits immunized with the HGF pellet from tumor and normal tissue.
Rabbit anti-normal Serum IgG
Rabbit anti-tumor Serum IgG
Absorbed rabbit anti-tumor IgG
C'H50 UNITS/ml Tumor Antigen Normal Antigen
181830^5
35161720456 0
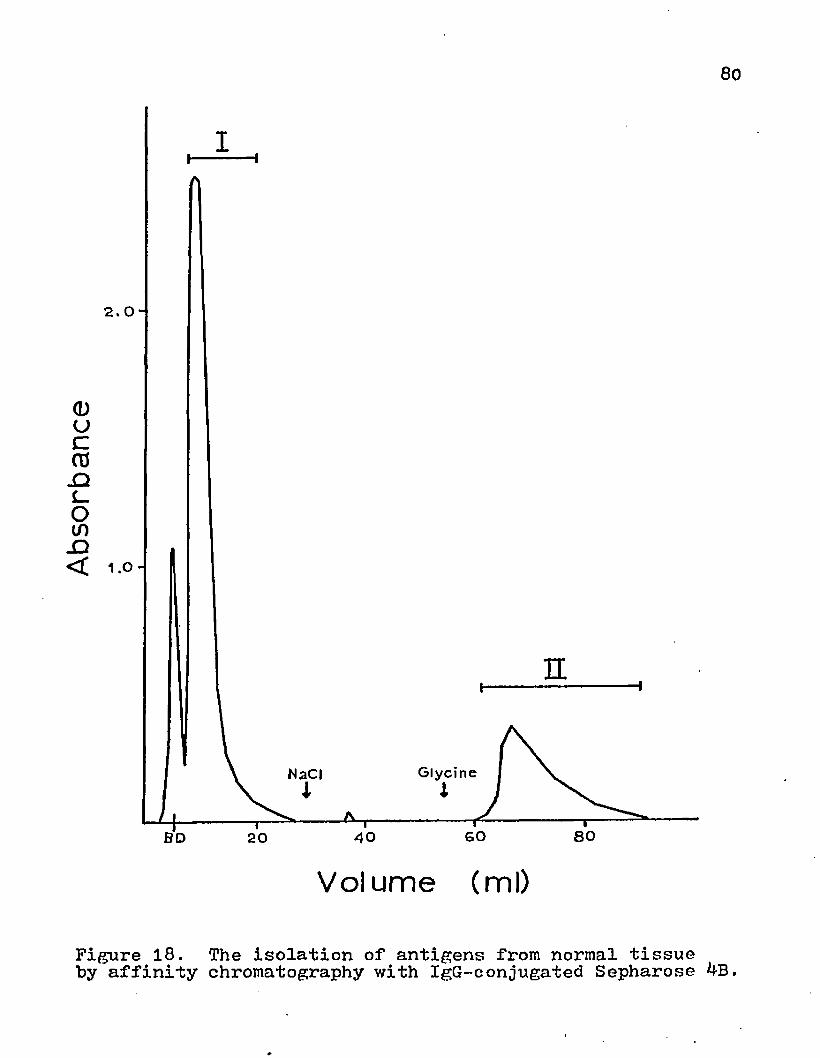
Abs
orba
nce
80
2.0 -
GlycineNaCI
8 0GO4 0BD 20
Volume (ml)
Figure 18. The isolation of antigens from normal tissue by affinity chromatography with IgG-conjugated Sepharose ^B.

81
effects and was not attributed to the elution of protein.The normal antigens were released from the antibody-gel matrix with a low pH buffer. Similar results were obtained with a column packed with absorbed rabbit anti tumor immuno- adsorbant (Fig. 19). Fraction I and II were assessed for the presence of normal or tumor antigens. No activity could be detected by the C'H50 assay in either sample (Table 15)* Since the affinity chromatography was performed at 23°C over a 24 hr period, it was possible that denaturation or proteolysis of the antigens had occurred. The immunoadsorbant was used in a batch procedure at 4°C to reduce the possibility of this deleterious action. Some antigenic activity was found in Fraction I, but this was greatly reduced in comparison to the soluble tumor antigen (Table 15)• Fraction II still displayed no antigenic activity in the C'H50 assay.The lack of activity was not due to the presence of detergent because the soluble protein was treated with ethanol and ethylene glycol before application to the column. The (NH^)2S0^ ppt was applied to the column but none of the sample would permeate the gel bed possibly due to the formation of large ppt. It is possible that the low pH treatment destroyed activity in Fraction II; whereas, proteolytic or denaturation may have caused loss of activity in Fraction I. Although the immunoadsorbant fractions did not display antigenic activity in the complement fixation assay, positive

2.0 “
GO4020
Volume (ml)
Figure 19. The isolation of antigens from tumor tissue by affinity chromatography with IgG-conjugated Sepharose 4B.
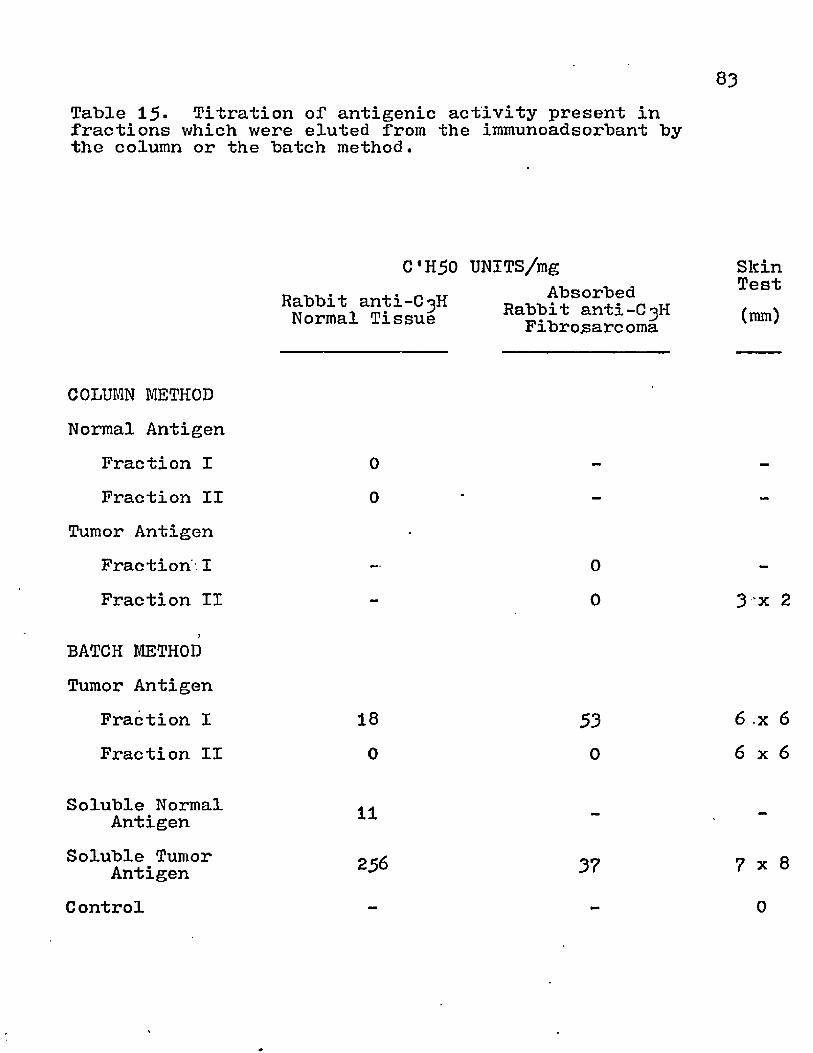
Table 15* Titration of antigenic activity present in fractions which were eluted from the immunoadsorbant by the column or the batch method.
Rabbi t ant i -C 3H Normal Tissue
C'H50 UNITS/mgAbsorbed
Rabbit anti-C3H Fibro^arc oma
COLUMN METHOD Normal Antigen
Fraction I Fraction II
Tumor Antigen Fraction.I Fraction II
00
00
BATCH METHOD Tumor Antigen
Fraction I Fraction II
180
53o
Soluble Normal Antigen
Soluble Tumor Antigen
11
256 37Control
83
SkinTest(mm)
6.x 6 6 x 6
7 x 8
0

results were obtained by skin testing immunized guinea pigs with both fractions (Table 15)■ The activity was comparable to that obtained with tumor HGF pellet and soluble protein. Since no induration was obtained with the control, the skin test reaction was the result of an immunological response.

DISCUSSION
The cell surface contains a variety of macromolecular components. This mosaic contains an array of antigens which may be important for the recognition of self and foreign determinants. Neoplastic cells have been shown to possess distinctive antigens which are not a part of the normal surface mosaic. Since lymphocytic recognition is due to contactual phenomena with the cell surface, the isolation of surface membranes from malignant cells would contain these tumor-specific antigens. By disrupting the cells, various subcellular constituents can be separated on the basis of size and density. When the murine fibrosarcoma homogenate was centrifuged at the higher centrifugal speeds, the pellet and interfacial material contained increasing amounts of mitochondria, endoplasmic reticulum, nuclei, and plasma membrane. However, the supernatant was mostly devoid of mitochondria, endoplasmic reticulum, and nuclei. In the 16,000 xg supernatant, the surface membrane marker activity was still present and greatly purified. These microsomal vescicles were pelleted at 113,000 xg for 60 min and contained surface membrane. The relative activity of
85

86
5'-mononucleotidase activity was increased eight-fold and a yield of 1-2 mg HGF protein per wet wt tissue could he obtained. The surface membrane marker was also found in other subcellular fractions but in much smaller quantities. Thus, the homogenization of the tumor tissue by nitrogen cavitation appears to have disrupted the plasma membrane into a variety of sizes which were sedimented at low, intermediate, and high centrifugal forces. Furthermore, most of the intracellular organelles and whole cells were sedimented at low or intermediate speeds. Similar subcellular fractionation results have been obtained with Ehrlich ascites carcinoma (110) and a murine lymphoma (7)• Therefore, the surface membrane from a murine fibrosarcoma could be procured in highly purified form by differential centrifugation.
Tumor specific antigens have been detected in subcellular fractions of neoplastic tissue (11, 3**-» 89» 111)and on viable tumor cell lines. The interfacial material and HGF pellet from the fibrosarcoma were related for animal immunizations because they contained the greater amounts of surface membrane marker activity (30.3 and 78#, respectively) and the least contamination by other organelles. So, every mg of protein injected would contain a high proportion of surface antigens. The IGF pellet was not chosen because of the presence of enzyme markers that correlate with the

87
presence of lysosomes. These organelles contain proteases which might destroy the antigenic activity of the subcellular fraction.
Rabbits which were immunized with interfacial material from the fibrosarcoma demonstrated a higher titer of antibody (1:l6o) in the microcytotoxicity assay. Sera from tumor-bearing and immunized C^H mice were quantitively less active (titer Ii40). Furthermore, the heterologous sera were more effective in producing cell cytolysis than the isologous antisera. Thus, the isologous sera may have been less effective in target cell destruction and had a lower titer due to the recognition of only a few epitopes. The rabbit antitumor sera were titrated with normal and tumor tissue and demonstrated the ability to fix complement with both antigens. The absorption of these antisera with the interfacial material from normal tissue removed most or all antibodies to normal mouse tissue antigens; whereas, the reactivity to the fibrosarcoma subcellular fractions remained. The immunoglobulins that were produced in syngeneic mice to the fibrosarcoma demonstrated no or little ability to fix complement in the C*H50 assay. Since the fibrosarcoma was derived from a C^H cell line, these syngeneic mice could respond only to the tumor specific antigens. However, the heterologous antisera may have been produced not only to these unique specificities but also species and tissue antigens. The

88
C'H50 assay may be less sensitive in the ability to detect antibody activity than the microcytotoxicity assay. The reason for this may reside in the nature of the antigen. In the microcytotoxicity assay, viable cells are used. The surface antigens may move in the fluid mosaic so that eventually antigen-antibody complexes are formed which permit complement activation and cell lysis. Only one site is needed for cytotoxic activity. With the subcellular fractions, the membrane constituents may be static and hence prevent complement fixation.
An attempt was made to elute antibody from fibrosarcoma tissue with 2.5 M NaCl; however, no activity could be detected. This may be attributed to the inefficiency of this method to elute or.of the assay to detect immunoglobulins.The antigen activity of the subcellular fraction was not drastically affected by this treatment. Treatment with low pH or denaturants may be necessary in order to achieve the elution of antibody from the tumor tissue. The interfacial material and the HGF pellet were not equivalent as immunogens. When the unabsorbed antisera for the interfacial material from the fibrosarcoma were titrated, the number of C*H50 units/ml obtained was greater with the homogenate than with HGF pellet antigen. The interfacial material contained DNA (kO.6%), plasma membrane (30.370* endoplasmic reticulum (18.8^), mitochondria (10.370 • and probably other intracellular

89
organelles that were not assayed. Since the homogenate contains all the various cellular organelles and their antigens, there would be more activity than with the purified HGF pellet which contained mostly surface membrane and some endoplasmic reticulum. When the rabbit anti-tumor from the interfacial material was absorbed with normal tissue, this difference was removed. The reactivity to tumor specific antigens in the homogenate and HGF pellet was similar because the number of these unique determinants were the same for both subcellular fractions. The titration of antisera to this purified material did not vary whether the homogenate or HGF material was used because the HGF pellet offered fewer antigens for an immune response than the interfacial material. The antigenic activity of this HGF pellet was preserved best in Tris-buffered sucrose containing 0.2?S sodium azide at 4°C . Thus, the titers of antisera for subcellular fraction can vary with the immunogen that is used for the injections and the antigen that is employed for the titrations of the antisera.
The heterologous antisera from the fibrosarcoma fractions demonstrated reactivity to tumor specific antigens. Initially, it was found that the unabsorbed antisera had a higher titer with the tumor tissue than the normal tissue. The absorption of this antiserum with normal tissue removed all activity to the liver, kidney, and muscle tissue but the

90
ability to react with the fibrosarcoma still remained.This remaining activity may represent* (i) tumor specific antigens which are not found on untransformed cells, (ii) antigen amplification in which there is a quantitative increase on malignant cells of a normal antigen (i.e. embryonic or fetal antigens). Also, there appeared to be a difference in the quantity of normal tissue antigens that were present in the transformed cells. When the malignant and normal tissue were assayed for the normal tissue antigens, normal tissue contained a higher quantity of these antigens than the transformed tissue. This may be due to: (i) antigenic deletion in which some normal specificities are no longer presnet, (ii) antigenic displacement in which the tumor specific antigens occupy space normally available to other antigens. Thus, a qualitative and a quantitative change in antigens may occur during oncogenic transformation.
The rabbit anti-HGF pellet was used to quantitate the amounts of normal and tumor specific antigen present in the various cell fractions. There was a significant correlation between the surface membrane marker activity and the amount of C*H50 units/mg present in any fraction for both the normal and tumor-defined antigens. The HGF pellet yielded the highest activities for surface membrane and antigen content. Thus, this complement-fixing activity may represent surface membrane antigens and not those of the internal mileu.

91
The content of tumor specific transplantation antigen in the various subcellular fractions was demonstrated by their ability to protect C^H mice against challenge with a tumor- forming dose of fibrosarcoma cells. Only the HGF pellet provided immunoprotection. The resistance demonstrated a dose-dependent response since high doses of the HGF may not induce a tumoricidal immune response. A dose-related immunity has been noted with soluble tumor antigens and has been attributed to the weak antigenicity of the tumor specific transplantation antigens (90). The dose response character of such antigens may be explained by the following provisions. Injection of low amounts of antigen may not produce immunostimulation because suboptimal amounts of antigen do not result in detectable lymphocytic stimulation. The amount of antigen necessary to induce a reaction appears at a threshold. The response occurs over a narrow range of doses. The immune response may then be diverted with high concentrations of antigen. This nullification in the high-dose range may reflect the induction tolerance or enhancement.
The antigenic configurations of the cell surface are intercalated in the lipid bilayer. Treatments with enzymes (97)i chaotropic salts (66, 90), hypotonic solutions (116), and detergents (2, 5*0 have been used to release antigens from the membrane. The effects of pH, osmolality, detergent concentration, time, and temperature on solubilization

92
were studied with the interfacial material. This subcellular fraction was used because it provided an adequate amount of protein for studying these variables (5-10 mg protein per g wet wt tissue). The solubilization of protein from these membranes was pH dependent. This was also noticed with another experimental tumor in which soluble material from Sarcoma I cells enhanced tumor growth in C57 B1 mice (60). The amount of protein released was approximately 90$ at standard conditions. This phenomenon may be a result of* (i) ionization of moieties associated with the membrane components (phospholipids, cholesterol, proteins, carbohydrates) which are unchanged at low or physiological pH. The carboxyl residues have low pKa values but amino, phosphatidyl , and sulfydryl substituents have pKa values in the alkaline region. This ionization process may result in charge repulsion of various macromolecules. (ii) various noncovalent bonds may also be broken by changes in pH such as van der Waals forces and hydrogen bonds.
The molar concentration of the Tris buffer also significantly effected protein solubilization. By increasing the osmolality of Tris, the percent solubilization decreased from 90$ to 44$. This has been also demonstrated with the extraction of proteins from human erythrocyte ghosts (80) and may be attributed to the stabilizing effect of high molar Tris buffers on membranes.

93
Proteins are oriented with the hydrophilic domain exposed to the aqueous environment; whereas, the hydrophilic proteins are inserted in the nonpolar environment of the lipid bilayer. In order to achieve the extraction of proteins from this matrix, an amphipathic molecule may be used to convert this hydrophilic domain to one that has an affinity for a polar environment. Solubilization with Triton X-114 was optimal at ImM concentrations and was rapidly diminished with higher quantities of this nonionic detergent. The solubilization process requires the detergent to be monomer in form. With ImM Triton X-114, approximately 90# of the amphipathic molecules are present in a monomeric form (41). As the quantity of detergent increases above the critical micellar concentration (3xl0%I), more monomers are complexed in micelles (?4). These monomers become integrated into micelles and are no longer available for the solubilization process. Also, various components may be released from the membrane with varying concentrations of detergent by the process of differential solubilization (62). Proteins have been selectively extracted from erythrocytes by using a nonionic detergent in the 0.2-lmM range.The use of 0.5-10mM detergent solubilizes the phospholipids and 5-20mM of the cholesterol (63). The binding of Triton X-114 to the hydrophobic surface is weak, k=2.2 x 10^ 1/fA (74) . Extraction of protein with nonpolar solvents or

94
treatment with polystyrene beads (56) way remove the detergent from the hydrophobic region so that antigenic activity and water solubility are diminished.
Lipids in cellular membranes demonstrate phase transitions when exposed to various temperatures (^2). The role of this solid gel conversion on solubilization was investigated but no dramatic effects were noticed with variations in temperature.
The solubilization of antigens from the interfacial material may be proportional to the amount of time which it is exposed to the surfactant. The results showed that the extraction process was complete after 60 min . Thus, pH, osmolality, and detergent concentration played a major role in the release of protein from the subcellular fraction membranes. The detergent could be removed from the medium by various methods and the antigen activity remained.
The ability to solubilize proteins makes possible the purification or isolation of antigenic specificities. Isoelectric focusing (10), gel filtration (66), salt precipitation (90), ion exchange chromatography (28), and affinity chromatography (15) have been used for this purpose. Affinity chromatography was used to attempt the isolation of a tumor specific antigen because it would permit the selection of only the desired antigenic specificities on a preparative scale. Absorbed rabbit anti-fibrosarcoma

95
was conjugated to the CNBr-activated Sepharose and soluble tumor antigen was adsorbed to the column. This antigen could be dissociated from the immobilized antibody by using a low pH buffer but not 2.5 M NaCl. The inability of the high ionic strength solution to desorb the protein indicates that the attachment to the column was due to a specific antigen-antibody interaction and not nonspecific adsorption effects. Although the low pH buffer was effective in eluting the antigen from the column, it may have also resulted in denaturation of the antigen because no activity was seen in the C'H50 assay. However, intradermal injection of the eluted protein into an immunized guinea pig resulted in a cutaneous anaphylaxis-type reaction which disappeared after 2k hrs. The adsorbed rabbit IgG to tumor tissue demonstrated specificity for only antigens from tumor and not normal tissue. To the column, this material may represent tumor specific antigens.
In summary, a subcellular fraction was obtained by the differential centrifugation of a tumor homogenate which was a purified preparation of surface membrane.This high centrifugal force pellet had a high surface antigen content and was able to confer immunity against the fibrosarcoma cells. More than 90% of the membrane antigens could be solubilized from the subcellular fraction with a nonionic detergent. The soluble antigens were separated by affinity chromatography.

SUMMARY
Tumors were produced in C^H/COX mice by injecting syngeneic polyoma-induced fibrosarcoma cells. Palpable tumors were harvested in 1^-16 days, disaggregated by pushing the tissue through a nylon screen, and homogenized by nitrogen cavitation. Various subcellular fractions were obtained by using differential centrifugation and an aqueous two-phase polymer system. The polymer system consisted of dextran and polyethylene glycol in a phosphate buffer and has been shown to separate biological substances on the basis of size, density, and surface properties. All fractions were checked for antigen content by a quantitative complement fixation assay, for an enzyme marker that is associated with the cell surface (5'-mononucleotidase), and for other enzyme markers located in other organelles (cytochrome oxidase, DPNH diaphorase, and diphenylamine assays). Results demonstrated that a 113,000 xg pellet (HGF) contained a nine-fold increase in the surface membrane marker activity and minimal contamination by organelles. This HGF pellet has also been injected into rabbits and guinea pigs. After
96

97
absorbing the antibody to normal antigen specificities, activity was still expressed toward tumor HGF pellet in complement fixation assays. This HGF pellet was used in a tumor challenge assay. The injection of 0.5 mg protein delayed significantly the appearance of a palpable tumor in comparison to control animals? however, a higher dose did not display such protection. Thus, a tumor specific transplantation antigen was present in the HGF membrane preparation and evoked a protective response which was dose dependent. Hence, a response to the tumor specific antigen could be elicited not only in syngeneic animals but also in xenogeneic species.
Various parameters for solubilizing protein from the surface membrane with Triton X-114 were studied. The effects of pH, osmolality, detergent concentration, temperature, and time were examined. The most dramatic changes were produced by pH, osmolality, and detergent concentration; whereas, temperature and time did not have profound effects beyond a certain minimum. Data from a quantitative complement fixation assay revealed that greater than 9k% of the antigens were released and 85-90^ of the protein remained in solution after a 6x10^ g-min centrifugation. Gel filtration, extraction with organic solvents, salt fractionation with ammonium sulfate, and micelle dispersion were investigated as methods for removing the detergent from the soluble

98
antigen. Salt fractionation was found to be the best method. This soluble antigenic material also gave positive results in complement fixation, skin test, and passive hemagglutination assays.
Using soluble antigens from the HGF pellet, various antigenic specificities from the murine fibrosarcoma were separated by affinity chromatography. Rabbit antisera that contained antibodies only to tumor specific antigens was conjugated to CNBr-activated Sepharose. Sera from tumor- bearing mice was not used because only minimal activity could be detected by complement fixation. By monitoring the effluent from the column, material could be eluted from the immunoadsorbant by using a low pH buffer. This eluted antigen displayed no activity in the C'H50 assay but did give a positive response in skin tests. Therefore, there were antigens present in the soluble protein which, were capable of combining in a specific manner with the immobilized antibody.

LITERATURE CITED
1 .
2.
3.
4.
5.
6.
7*
8.
9*
10.
Alexander, P. 1967* Immunotherapy of leukemiai the use of different classes of immune lymphocytes.Cancer Res., 27*2521-2526.________ . 197^. Escape from immune destruction by thehost through shedding of surface antigens* is this a characteristic sharec by malignant and embryonic cells? Cancer Res., 3^*2077-2082.Alexander, P. and Fairley, G. H. 1967* Cellular resistance to tumours. Brit. Med. Bull., 23*86-92.Andrews, A., Congdon, C. C., Edwards, C. L., Gengozian, N., and Vodopick, N. H. 19^7■ Preliminary trials of clinical immunotherapy. Cancer Res.. 27*2535“25*H •Ariyan, S. and Gershon, R. K. 1973* Augmentation of the adoptive transfer of specific tumor immunity by nonspecifically immunized macrophages. J. Natl. Cancer Inst., 51! 114-5-11^8*Avis, P. and Lewis, M. G. 1973* Tumor-associated fetal antigens in human tumors. J. Natl. Cancer Inst., 51*1063-1066.Avis, P. J. G, 1972. In Subcellular components* preparation and fractionation. London*Butterworth..Baldwin, R. W. and Barker, C. R. 1967. Antigenic composition of transplanted rat hepatomas originally induced by dimethylaminoazobenzene. Brit. J. Cancer.21*338-3^5*Bashford, E. F. 1909* Cancer in man and animals.The Lancet. 2*691-701.Bernier, I., Dautigny, A., Colombani, J., and Jolles, P. 1973* Detergent solubilized HL-A antigens from human platelets* a comparative study of various purification techniques. Biochem Biophys Acta, 356*82-90.
99

100
11. Bhattachaya, M. and Barlow, J. J. 1973* Immunologic studies of human serous cystadenocarcinoma of ovary.Cancer. 31*588-595*
12. Blair, P. B. 19^9* Immunodiffusion of mouse mammarytumor virus in mild samples and correlation with‘tumor development. Cancer Res., 29:745-748.
13* Blarney, R. W. 1989* The effect of isogenic lymphoidcells on primary sarcoma in the rat. Cancer Res.,29*333-334-.
14. Bonavida, B. 1974. Studies on the induction andexpression of T cells-mediated cytolysis by membrane antigens. J. Iran., 112:926-934.
15* Bowen, J. G. and Balswin, R. W. 1976. Isolation and characterization of tumor-specific antigen from the serum of rats bearing transplanted aminoazo dye-induced hepatomas. Transnl.. 21:213-219.
16. Boyse, E. A., Old, L. J., Stockert, E., and Shegino, N.1908. Genetic origin of tumor antigens. Cancer Res.,28:1280-1287*
17* Brooks, P. and Lawley, P. D. i960. Aldylating agents.Brit. Med. Bull., 20:91-95*
18. Bubenik, J., Perlman, P., Helmstein, K., and Moberger, G.1970. Cellular and humoral responses to human urinarybladder carcinomas. Int. J. Cancer.. 5*310-319*
19* Burk, K. H., Drewinko, B., Lichtiger, B., and Trijilli, J.1976. Cell cycle dependency of human sarcoma-associated tumor antigen expression. Cancer Res.. 36*1278-1283.
20. Burtin, P., Chavanel, G., and von Kleist, S. 1972. Demonstration in normal human plasma of an antigen that cross reacts with the carcinoembryonic antigen of digestive tract tumors. J. Natl. Cancer Inst., 49:1727-1728.
21. Burtin, P., von Kleist, S., and Savine, M. C. 1971*Loss of a normal colonic membrane antigen in human cancers of the colon. Cancer Res., 31*1038-1041.
22. Burton, K. 1956. A study of the conditions and mechanisms of the dephenylamine reaction for the colorimetric estimation of deoxyribonucleic acid. Biochem. J., 62:315-323*

Clark, D. A. and Nathanson, L. 197^* Patterns of cellular immunity in malignant melanoma. Recent Results in Cancer Research, 47*67-78.’Claugh, S. C.t Parikh, I., and Cuatrecasas, P. 197^.A somplified method for cyanogen bromide activation of agarose for affinity chromatography. Anal. Biochem.,60i149-152.Coggins, J. H. and Anderson, N. G. 197^* In Developmental aspects of carcinogenesis and immunity. New York*Academic Press.Cooperstein, S. J. and Lazarow, A. 1951* A micro- spectrophotometric method for determination of cytochrome oxidase. J. Biol. Chera., 189*665-670.Cryan, V/. S., Hyde, R. M., and Garb, S. 1966.Demonstration by gel diffusion of antigen in spontaneous mouse tumor. Cancer Res., 26t1458-1465.Davies, D. A. L., Boyse, E, A., Old, L. J., and Stockert,E. 1967* Separation of soluble TL antigen from soluble H-2 histocompatibility antigen by column chromatography.J* Sxp. Med., 125*5^9-558.Day, E. D. and Bigner, D. D. 1973* Specificity, cross reactivity, and affinity of iodine-labelled antiglioma antibodies for monolayers of cultured glioma celis.Cancer Res., 3312362-2372.Deckers, P. J., Edgerton B. W,, Thomas, B. J., and Pilch, Y. H. 1971* The adoptive transfer of concomittant immunity to murine tumor isografts with spleen cells from tumor-bearing animals. Cancer Res.,31*73^-7^2.Diller, I. C. and Watson, G. F. 19^9■ The effect of aqueous spleen extract on growth of tumor cells in mice. Growth, 13*27-44.Eilber, F. R. and Morton, D. L. 1970. Immunologic studies of human sorcomas* additional evidence suggesting an associated sarcoma virus. Cancer. 261588-596.Eisen, M. J. and Woglom, W. H. 1941. The nonspecific nature of imduced resistance to tumors. Cancer Res., It629-631.

Embleton, M. J. and Heidelberger, C. 1975* Neoantigens on chemically transformed clone CoH mouse embryo cells. Cancer Res.. 35*2047-2055. *
Fischinger, P. J. and Haapala, D. K. 1974. Oncoduction. A unifying hypothesis of viral carcinogenesis.Prog. Expl. Tumor Res., 19*1-22.Fisher, B.f Saffer, E. A., and Fisher, E. R. 1971* Experiences with lymphocyet immunotherapy in experimental tumor systems. Cancer, 27*771-781.Fisher, B.t Soliman, 0., and Fisher, E. R. 1970.Further observations concerning effects of antilymphocyte serum on tumro growth* with special references to allogeneic inhibition. Cancer Res., 30:66-68.Flexner, S. and Jobling, J. W. 1907* Restraint and promotion of tumor growth. Proc. Son. Expl. Biol. Med., 5 *16-18.Foley, E. J. 1953* Antigenic properties of methyl-cholanthacene-induced tumors in mice of the strain of origin. Cancer Res.. 13*835-837*Fossati, G. Canevari, S., and DellaPorta, G. 1973* Cellular immunity in cancer patients against embryonal cells. J. Natl. Cancer Inst., 51*867-671*Frasch, C. E. 1975* Removal of detergent by ultrafiltration. Dialog. 7*1-3*Frye, L. D. and Edinin, M. 1970. The rapid intermixing of cell surface antigens after formation of mouse-human heterokaryons . J. Cell Sci.. .'7* 319-336.Gorer, P. A. 1937* The genetic and antigenic basis oftumor transplantation. J. Path. Bact., 44:691-697.Gerfo, P. L., Krupey, J., and Hansen, H. J. 1971*Demonstration of an antigen common to several varietiesof neoplasia. New Eng. J. Med., 285*138-141.Gold, P. and Freedman, S. 0. 1965* Specific carcino-embryonic antigens on the human digestive system.J. Ex£. Med., 121*439-462.

103
46. Gold, P. and Freedman, S. 0. 1965* Demonstration oftumor-specific antigens in human colonic carcinomata by immunological tolerance and absorption techniques.J. E x p . Med., 122*467-481.
4?. Graham, J. B. and Graham, R. M. 1955* Antibodieselicited by cancer in patients. Cancer, 8*409-416.48. Gross, L. 19 *3* Intradermal immunization of C3Hmice against a sarcoma that originated in an animal of
the same line. Cancer Res., 3*326-333*49* . 19^5* The specificity of acquired tumor
immunity. J. Imm., 50*91-99*50. Hayami, M., Hellstrom, I., Hellstrom, K., and Lammin, D. R.
197^* Further studies on the ability of regressorsera to block cell-mediated destruction of Rous sarcomas. Int. J. Cancer. 13*^3-53*
51. Hellstrom, I. and Sjogren, H. 0. 19^5* Demonstrationof H-2 isoantigens and polyoma specific tumor antigens by measuring colony formation in vitro. Expl. Cell Res., 40*212-215.
52. Hellstrom, I. and Sjogren, H. 0. 1966. Demonstrationof common specific antigens in mouse and hamster polyoma tumors. Int. J. Cancer, 1*481-489.
53* Hellstrom, I., Hellstrom, K. E., Sjogren, H. 0., andWarner, G. A. 1971. Demonstration of cell-mediated immunity to human neoplasms of various histological types. Int. J. Cancer, 7*1-16.
54. Hilgert, I., Kandutsch, A. A., Cherry, M., and Snell, G. D. 1969. Fractionation of murine H-2 antigens with the use of detergents. Transpl.. 8*251-262.55* Habel, K. and Atanasiu, P. 1959* Tranplantation ofpolyoma-indeced tumor in the hamster. Proc. Soc.. Expl. Biol. Med., 102*99-102.56. Halloway, P. W. 1973* A simple procedure for removalof Triton X-100 from protein samples. Anal. Biochem., 53*304-308.

104
5?. Itoh, T. and Southam, C. M. 1963* Isoantibodies to human cancer cells in healthy recipients of cancer hmomtransplants. J. Imm.. 91*469-483.
58. Jacobs, B. B. 1972. Altered host-allograft relationships for mouse tumors modified by prior passage in vitro and in vivo. II Reactivity of lymphoid cells.J* Natl. Cancer Inst.. 4-9* 1085-1091.
59* Jeejeebhoy, H. P. / 1975* Tbe role of the immune response in oncogenesis. In Vitro. 11:166-172.
60. Kandutsch, A. A. i960. Intracellular distribution and extraction of tumor homograft-enhancing antigens.Cancer Res., 20*264-273-
6l,. Kidd, J. J. 1930. A complement-binding antigen inextracts of the Brown-Pearce carcinoma of rabbits.Proc. Soc. Expl. Biol. Med., 38*292-295*
62. Kirkpatrick, P. H. and Sandberg, H. E. 1973* Effectof anionic surfactants, nonionic surfactants, and neutral salts on the conformation of spin-labelled erythrocyte membrane proteins. Biochem Biophys Acta. 298:209-216.
63. Kirkpatrick, F. H., Garkesky, S. E., and Marinetti, 0. V. 1974. Differential solubilization of proteins, phospholipids, and cholesterol of erythrocyte membranes by detergents. Biochem Biophys Acta, 345*154-162.
64. Klein, G. 1966. Tumor antigens. Ann. Rev. Micro.. 20:223-252.65. Koch, M. A. and Sabin, A. A. 1963* Specificity of virus-induced resistance to transplantation of polyoma
and SV40 tumors in adult hamsters. Proc. Soc. Expl.Biol. Med., 113*4-12.
66. Leonard, E. J., Meltzer, M. S., Borsos, T., and Rapp, H. J. 1972. Properties of tumor^-specific antigens solubilizedby hypotonic potassium chloride. Natl. Inst. Cancer Monogr.. 35*129-134.
67. Lesko, L. , Donlon, M., Mainetti, M. , and Hare, J. D.1973* A rapid method for the isolation of rat liver plasma membrane using an aqueous two-phase polymer system. Biochem Biophys Acta, 311*173-179*

105
68. Levin, I. and Sittenfield, M. J. 1911* Studiesonimmunity in cancers of the white rat: the significance of athrepsia. J. Exp. Med., 13*511-520.
69* Levine, L. 1973* In Handbook of experimental immunology. London1Blackwell.
70. Lewis, M. G., Ikonopisov, R. L., Nairn, R. C., Phillips,T. M., Fairley, G. 11., Bodenham, D. C., and Alexander, P.1969* Tumor-specific antibodies in human malignantmelanoma and their relationship to the extent of the disease. Brit. Med. J., 3*547-552.
71. Lowry, 0. H., Rosebrough, N. J., Farr, A. J., and Randall,R. J. 1951* Protein measurement with the Folin phenolreagent. J. Biol. Chem., 193*265-275*
72. Lowry, 0. H. and Lopez, J. A. 19^6* The determination of inorganic phosphate in the presence of labile phosphate esters. J. Biol. Chem., 162*421-428.
73* Lumsden, T. 1937* On cytotoxins lethal to nucleated mammalian cells norma and malignant. Amer. J. Cancer, 31*430-439.
74. Makino, S., Reynolds, A., and Tanford, C. 1973* The binding of deoxycholate and Triton X-100 to proteins.J. Biol. Chem. 248:4926-4932.
75* Martin, F. and Devant, J. 1973* Carcinoembryonicantigen in normal human saliva. J. Natl. Cancer Inst. 50:1375-1379*
76. Mavligit, G.M., Gutterman, J.U., McBride, C.M., and Hersh, E.M. 1973* Cell-mediated immunity to human solid tumors: in vitro detection by lymphocyte blast-ogenic responses to cell-associated and solubilized tumor antigens. Recent Results in Cancer Res. 47:84-96.
77. Mavligit, G.M., Ambus, U.t Gutterman, J.U., Hersh, E.M., and McBride, C.M. 1973* Antigen solubilized from human solid tumors: lymphocyte stimulation and cutaneousdelayed hypersensitivity. Nature (New Biology), 243* 188-190.
78. McCarthy, R.E. 1964. Modification of the immune response of mice to skin homografts and deterografts by Ehrlich ascites carcinoma. Cancer Res. 24*915-919*

10 6
79* McPeak, C.J. 1971* Intralymphatic therapy with immunelymphocytes. Cancer. 28*1126-1127•
80. Miller, D.M. 1970. Total solubilization of erythrocyte membranes by nonionic detergents. Biochem.Biophys. Res. Comm. 40*716-722.
81. Miller, L.S., Evans, S.B., Rossio, J.L., and Dodd, M.C.197^. A rapid method for the isolation of plasma membranes from a murine fibrosarcoma using an aqueous two-phase polymer system. Prep. Biochem. ^*489-^98.
82. Mitchison, N.A. 1955* Studies on the immunological response to foreign tumor transplants in the mouse.I. The role of lymph node cells in conferring immunity by adoptive transfer. J. Ex p . Med. 102*157-177.
83. Moller, E. and Moller, G. 19&7* Inhibition of tumor grov/th by confrontation with imcompatible cells. Possible surveillance mechanism against neoplasma. Cancer. 20*871-879.
8^. . 1955* Quantitative studies on the sensitivityof normal and neoplastic mouse cells to the cytotoxic action of isoantibodies. J. Exp. Med. 102*527-553■
85. Muller, H.K, and Flannery, G.R. 1973* Epidermal antigens in experimental keratoacanthoma and squamous cell carcinoma. Cancer Res. 33*2181-2186.
86. Murphy, J.B, and Morton, J.J. 1915* The lymphocytein natural and induced resistance to transplanted cancer.II. Studies in lymphoid activity. J. Exp. Med. 22* 20^-211. “
87. Nordquist, R.E. 1973* Specific antigens in humanalveolar cell carcinoma. Cancer Res. 33*1790-1795*
88. Old, L.J. and Boyse, E.A. 1965. Antigens of tumorsand leukemias induced by virus. Fed. Proc. 2^*1009-1017.
89. Paranjpe, M.S. and Boone, C.W. 1975* Specific depression of the antitumor cellular immune response with autologous tumor homogenate. Cancer Res. 35*1205-1209•
90. Pellis, N.R. and Kahan, B.D. 1975* Specific tumor immunity induced with soluble material* restricted range of antigen dose and of challenge tumor load for immunoprotection. J. Imm. 115*1717-1722.

10?
91* Phelps, H.J. 1937* A note on the non-specific actionof so-called "anti-cancer serum." Amer. J. Cancer. 31*441-445.92. Pitot, H.C, 1965* Some biochemical aspects of malignancy. Ann. Rev. Biochem. 35*335-368.93* Potter, V.R, 1958. The biochemical approach to thecancer problem. Fed. Proc. 17*691-697.9*K Prehn, R.T. and Main, J.M. 1957* Immunity to methyl-cholanthrene-induced sarcomas. J. Natl. C.ancer Inst.
18*769-778.95* Priori, E.S., Semon, G. , Demochowski, L., Gallager, H.S.,
and Anderson, D.E. 1971. Immunofluorescence studies on sera of patients with breast carcinoma. Cancer. 28:1462-1471.96. Rapp, F. 1973* Do herpesviruses cause cancer? J.Natl. Caneer Inst. 50*825-831.97* Rosenberg, E.B., Mann, D.L., Hill, J.J., and Fahey, J.L.
1971. Prolonged skin allograft survival in mice pretreated with soluble transplantation antigens. Transol. 12:402-405.
99. Rous, P. 1910. An experimental comparison of transplanted tumor and a transplanted normal tissue sapable of growth. J. Exo. Med. 12*344-365.
100. Segerling, M., Chanian, S.H., and Borsos, T. 1975* Effect of metabolic inhibitors on the ability of tumor cells to express antigen and bind complement components C4 and C3* Cancer Res. 35*3204-3208.
101. Sjogren, H.0, and Hellstrom, I. 1965* Induction of polyoma specific transplantation antigenicity in moloney leukemia cells. E x t >. Cell Res. 40*208-212.
102. Sjogren, H.0. and Jonsson, N. 1957- Resistance against isotransplantation of mouse tumors induced by Rous sarcoma virus. Ex p . Cell Res. 32*618-621.
103. Sjogren, H.0., Hellstrom, I., and Klein, G. I96I. Resistance of polyoma virus immunized mice to transplantation of established polyoma tumors. Exp. Cell Res. 23*204-208.

108
104.
105.
106.
107.
108.
109.
110.
111.
112.
113.
114.
115.
116.
Steele, G., Sjogren, H.O., and Price, M.R. 1975* Tumor-associated and embryonic antigens in soluble fractions of a chemically-induced rat colon carcinoma. Int. J. Cancer, 16:33-51.Tuffrey, M.A. and Batchelor, J.R. 1964, Tumour specific immunity against murine epitheliomas induced with 9,10- dimethyl-1,2-benzanthracene. Nature. 204:349-351*Twomey, P.L., Catalona, W.J., and Chretien, P.B. 1974. Cellular immunity in cured cancer patients. Cancer, 33:435-440.Vaughan, J.W. 1912. Blood changes caused by the hypodermic administration of the cancer proteid. J. Amer.Med. Assoc. 59*1784-1769*________ . 1914. Cancer vaccine and anticancer globulins as an aid in the surgical treatment of malignancy.J* Amer. Med. Assoc. 53*1258-1265.Wahren, B. 1966. Demonstration of a tumor-specific antigen in spontaneously developing AKR lymphomas. Int. J. Cancer, 1:41-50.Wallach, D.P.H. and Kamat, V.B. 1966. In Methods in Bnzymology. Academic Press, New York.Weiler, E. 1956. Antigenic differences between normal hamster kidney and stilbesterol-induced kidney carcinoma: complement fixation reactions with cytoplasmicparticles. Brit. J. Cancer. 10*553-559*Wells, H.G. 1909* The resistance of the human body to cancer. J. Amer. Med. Assoc. 52*1731-1740.Woglom, V/.H. 1922. A critique of tumor resistance.J. Cancer Res. 7*283-301.Yoshida, T.0. 1965* Further evidence of immunologicreaction against methylcholanthrene-induced autochthonous tumors. Japan J. Exp. Med. 35*115-124.Youn, J.K., LeFrancois, D., and Barski, G. 1973* In vitro studies on the mechanism of the eclipse of cell- mediated immunity in mice bearing advanced tumors. J. Natl. Cancer Inst. 50*921-926.Young, J.M. and Gyenes, L. 1973* The induction of cellular immunity with soluble murine histocompatibility antigens. Cell Imm. 6:231-242.