western european quaternary lions: new working hypotheses
TRANSCRIPT

Western European Quaternary lions:new working hypotheses
MARCO MASSETI1 and PAUL P. A. MAZZA2*
1Department of Evolutionary Biology ‘Leo Pardi’, University of Florence, via del Proconsolo 12, 50122Florence, Italy2Department of Earth Sciences, University of Florence, via La Pira 4, 50121 Florence, Italy
Received 10 October 2012; revised 1 December 2012; accepted for publication 1 December 2012
Cave lions (Panthera spelaea), which spread throughout Western Europe for several thousand years, disappearedapproximately 14 000–14 500 years ago. They were supposedly replaced by modern lions (Panthera leo) approxi-mately 8000 years ago. Modern lions reached the steppes of Ukraine and Hungary, without penetrating the forestsof Central Europe. The present study focuses on Italian and Spanish findings that possibly bridge the allegedabsence of these big cats from Europe for 6000 years. Fossil lion remains from reliably radiocarbon-dated levelshave been plotted against the d18O curve and mapped. The accumulated evidence indicates that lions inhabitedWestern Europe uninterruptedly from the early Middle Pleistocene up to the Early Holocene. Moreover, all of thelatest Pleistocene/early Holocene lion-bearing localities do not range farther than the 44th parallel north and arelocated at relatively high altitudes. Two working hypotheses are formulated: one, which is less likely because it isnot supported by palaeontological evidence indicating earlier migrations of lions from Africa, suggests that modernlions entered Western Europe prior to 8000 years ago; the second, which is more probable, suggests that P. spelaea(or an advanced offspring of the species) survived up until the latest Pleistocene. Panthera leo accessed EasternEurope between 6000–6500 and 8000 years ago but was prevented from penetrating further west, probably becauseof the intrusive presence of their indigenous European relatives, and/or the increasing encroachment of modernhuman populations. © 2013 The Linnean Society of London, Biological Journal of the Linnean Society, 2013, 109,66–77.
ADDITIONAL KEYWORDS: dispersal – Panthera spp. – Pleistocene/Holocene – stratigraphic distribution –Western Eurasia.
INTRODUCTION
Africa is the homeland of both past and modern lionlineages. Modern sub-Saharan African lions derivefrom a number of in East and South African Pleis-tocene refugia in a time period spanning fromapproximately 169 to 324 kyr BP (Antunes et al.,2008). During the Late Pleistocene (i.e., approxi-mately 100 kyr BP), they spread into Central andNorth Africa and into Asia. Long before this date,however, at approximately 700 kyr, lion clades dis-persed in Europe (García & Arsuaga, 2001; Bischoffet al., 2007), when landscapes opened-up, hostingsavannah-like open woodlands (García & Arsuaga,
2001). Early in their history, they spread as far northas the British Isles, as attested by the remains clas-sified as Panthera fossilis or Panthera leo fossilis fromthe type Cromerian of West Runton, Norfolk (Lewis,Pacher & Turner, 2010; Stuart & Lister, 2011). Thisearly European Middle Pleistocene stock is ancestralto the Late Pleistocene Panthera spelaea or Pantheraleo spelaea of Eurasia. The latter spread out fromSpain to the easternmost reaches of Russia. A Ber-ingian population of cave lion apparently colonizedNorth America approximately 337 kyr BP, where itremained isolated and originated Panthera atrox(Barnett et al., 2009).
Rivers of ink have been spilled on whether the LatePleistocene lions of Western Europe are to be classi-fied Panthera leo, or not. Leaving aside Argant (1988),who named them Panthera mosbachensis, those who*Corresponding author. E-mail: [email protected]
bs_bs_banner
Biological Journal of the Linnean Society, 2013, 109, 66–77. With 3 figures
© 2013 The Linnean Society of London, Biological Journal of the Linnean Society, 2013, 109, 66–7766

consider P. spelaea a fully distinct species find it mor-phologically (Friant, 1940; Dietrich, 1968; Janossy,1969; Verescagin, 1969; Vereshchagin, 1971; Ballesio,1980; Spassov & Iliev, 1994; Baryshnikov & Boesko-rov, 2001; Nagel et al., 2003; Sotnikova & Nikolskiy,2006; Baryshnikov, 2011) distinct from modernlions. Cave lions are known to be generally largerthan modern lions (Ballesio, 1980). Sotnikova &Nikolskiy (2006) list a number of cranial and dentalcharacters distinguishing P. spelaea from P. leo. In thepostcranial skeleton, cave lions have relatively stur-dier, more robust limb bones, especially the metapo-dials (Ballesio, 1980). Panthera spelaea is alsomarkedly distinct genetically from P. leo (Burgeret al., 2004; Barnett et al., 2006), and closer to modernsub-Saharan lions than to North African and Asianrepresentatives (Barnett et al., 2006; Stuart & Lister,2011). Because no genetic interchange has been iden-tified between cave and modern lions (Burger et al.,2004; Barnett et al., 2009), Stuart & Lister (2011)assumed that the European spelaea lineage developedin complete genetic isolation from its African rela-tives. These conclusions are in stark contrast to thoseof Hemmer (1967, 1974), Kurtén (1968, 1985), Turner(1984), García & Arsuaga (1998), Turner & Antón(1997), Anconetani et al. (1995), Yamaguchi et al.(2004), Álvarez-Lao & García-García (2006), GarcíaGarcía et al. (2009), Croitor & Brugal (2010), Schnit-zler (2011), Molara (2012), who considered P. spelaeaand P. leo to be conspecific, at most distinct at thesubspecies level.
The European or Eurasian lion first appeared in theearly Middle Pleistocene at Isernia La Pineta, south-eastern Italy (Sala, 1990). According to Stuart & Lister(2011), P. spelaea became extinct across Eurasiaapproximately 14–14.5 cal kyr BP. Stuart & Lister(2011) also state that, in the time period span-ning approximately 7–14 kyr BP (i.e. during thePleistocene/Holocene transition), there was an expan-sion of modern lions from southern refugia northwardstowards East Africa (Antunes et al., 2008) and towardsEurope. Therefore, P. leo would have entered Europeduring the Holocene, more exactly between 6.0–6.5and 8.0 cal kyr BP, presumably from Asia Minor,passing through the Bosphorus. The reconstruction byStuart & Lister (2011) is in line with those proposed bySpassov & Bozkov (1982) and Spassov & Iliev (1994),who assumed that several Asian elements enteredEurope not earlier than the last glacial episode, prob-ably via a terrestrial land-bridge across the Bosphorus(Por, 1987). Schnitzler (2011) suggested an alternativeway through the Caucasus.
Largely because of human hunting and a loss ofhabitats, modern lions lasted a few thousand years.Historians of the Fifth Century BC to the SecondCentury AD (Herodotus, Xenophon, Aristotle, Pliny
the Elder, Dion Chrysostom, Pausanias, and Heli-anus) warned about the treacherous presence of lionsin Eastern Europe, as well as in the southern Balkanregions between the Fifth and First Centuries BC(Ninov, 1999; Yannouli, 2003; Salari, 2006). Harper(1945) first noted that the lions disappeared fromEurope (the Balkans) in approximately the FirstCentury AD. Between the Fourth and Third centuryBC, lions still persisted in Bulgaria (Spassov & Popov,2007). The latest European reports are from Greece(AD 80–100: Bartosiewicz, 2009); more precisely, fromKastanas (Becker, 1986) and Delphi (Lefèvre et al.,1992). Nonetheless, when lions actually becameextinct in the southern Balkan peninsula remainsuncertain. It is generally agreed that this occurred inthe second half of the First Millennium BC, or at thebeginning of the Christian era (Meyer, 1903; Yannouli,2003). In recent historical times P. leo was distributedin the South-western Palearctic from Western India toalmost all the Near East, from Persia and Asia Minorto North Africa. Until 1942, the Barbary lion, Panth-era leo leo L., 1758, lived on the Western Atlas, inMorocco (Aulagnier & Thevenot, 1986; Cuzin, 2003).
From the accounts provided by Stuart & Lister(2011), lions apparently lacked from Europe for atleast 6000 years (i.e. from approximately 14 to 8 kyrBP), which is the gap separating P. spelaea fromP. leo. Moreover, according to Stuart & Lister (2011),modern lions would have dispersed in the steppes ofUkraine and Hungary (Krakhmalnaya, 1999) butwithout reaching the forests of Central Europe. Inthis scenario, modern lions would have reached onlyEastern Europe and only for a limited time period.Was this actually the case? There is evidence thatappears to reduce or even fill the temporal (andpartially also the spatial) gaps in the European dis-tribution of modern lions. Yet they have been scarcelyconsidered by scholars. The present study focuses onthis information, which includes both osteological andiconographical data and, in particular on the differentscenarios that might result from it. The present studyis not only based on direct observation, but also oninformation available in the literature, which wasobtained from Cardini (1972), Castelletti, Maspero &Tozzi (1994), Cilli (1993–1994), Boschian, Mallegni& Tozzi (1995), Masseti et al. (1995), Cilli, Malerba &Tozzi (1998), Berton et al. (2003), Fiore, Pino Urìa &Tagliacozzo (2004). Late Pleistocene lions are alsoreported from Cantabria (Northern Spain); moreexactly, from layers 27 (late Magdalenian-Azilian)and 29 (basal Mesolithic?) of Cueva La Riera (Altuna,1986; Sommer & Benecke, 2006), and from levels Eand F (Early–Middle Magdalenian) of Cueva deUrtiaga (García Morales, 1984–1985).
All the radiocarbon dates reported are calibrated.For conversion from noncalibrated to calibrated ages,
EUROPEAN QUATERNARY LIONS 67
© 2013 The Linnean Society of London, Biological Journal of the Linnean Society, 2013, 109, 66–77

reference was made to Kitagawa & van der Plicht(1996), Jöris & Weninger (1998), Reimer et al. (2009)and Stuiver et al. (1998).
EASTERN EUROPE (BUT NOT ONLYEASTERN EUROPE)
Finds from Italy, and perhaps also from Spain, appar-ently build a bridge across the supposed chasm of6000 years separating P. spelaea from P. leo (Figs 1,2). The remains are sparse, although they areunquestionably lion remains. Unfortunately, giventheir fragmentary nature, the specimens lack taxo-nomically diagnostic elements that would supporttheir attribution to either P. spelaea or P. leo. Thismight be resolved with recourse to ancient DNAmethods, although DNA extraction is a destructiveprocess and we were not allowed by the curators toalter such scarce lion material it in any way. This alsoimpedes the direct dating of the remains, which couldtherefore only be context-dated. The possibility thatthe specimens might have been reworked from olderlayers cannot be thoroughly ruled out. Münzel et al.(2007, 2008, 2011) have shown that this is not such aremote possibility. Although the chance that the speci-mens from all the fossiliferous levels examined in thepresent study are reworked is quite improbable, theimpossibility of analyzing the remains chemically ledto the formulation of two, most intriguing working
hypotheses: (1) P. leo entered Europe earlier thanassumed or (2) Panthera spelaea survived longer thanassumed.
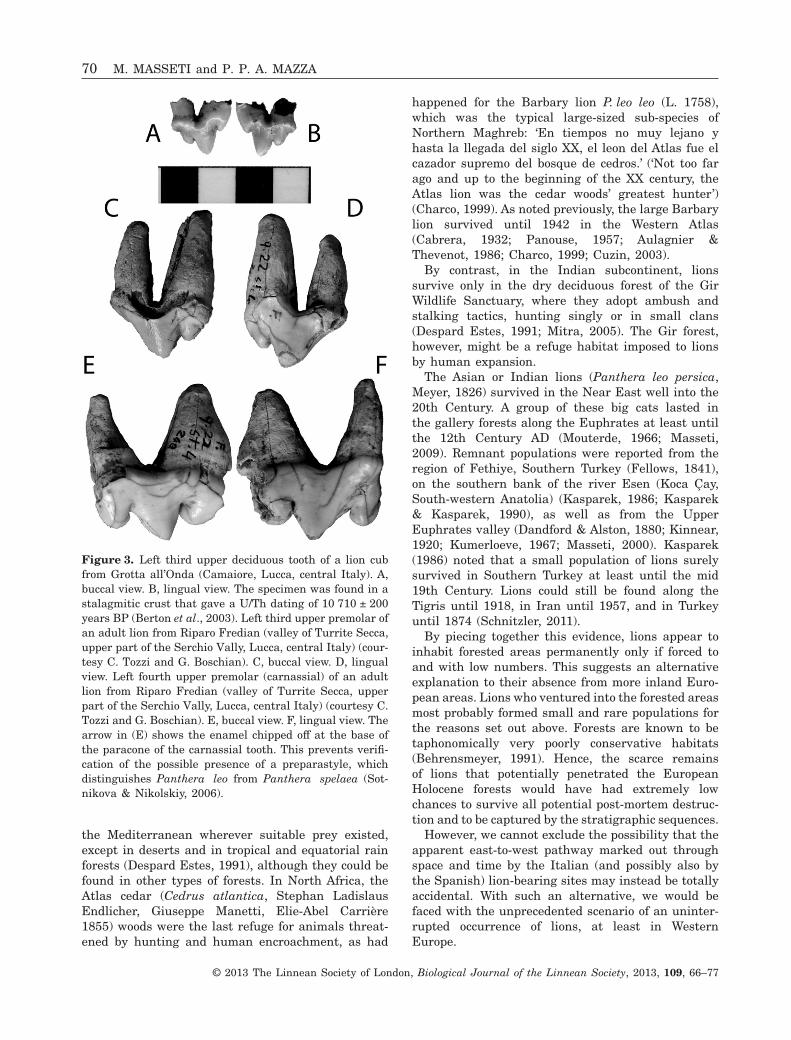
The oldest of the remains chronologically includedin the gap of 6000 years come from layer L of Grottadella Madonna, (Praia a Mare, at the feet of MountVingiolo on the Tyrrhenian coast of Calabria; latitude39°53′43″N, longitude: 15°47′11″E, approximately52 m a.s.l.), and are dated to 12 100 ± 150 (cal yearsBP 14 038–14 291)/9020 ± 125 (cal years BP 9014–10 243) years BP on charcoal and burnt bones (Alessioet al., 1966, 1967; Cardini, 1972; Fiore et al., 2004).Other early evidence of lions comes from RiparoFredian (valley of Turrite Secca, upper part of theSerchio Vally, Lucca, central Italy): these remainsgave a radiocarbon date of 10 870 ± 119 (cal years BP12 875–12 993) years BP and are therefore referred tothe late Epigravettian (Cilli, 1993–1994; Cilli et al.,1998). Another quite old Italian attestation is that ofGrotta all’Onda (Camaiore, Lucca, central Italy),where a third upper deciduous tooth of a lion cub wasfound in a stalagmitic crust that gave a U/Th datingof 10 710 ± 200 years BP (Berton et al., 2003) (Fig. 3).Somewhat younger lion finds are the remains ofan adult and of two juveniles with deciduous denti-tion, found in Riparo Fredian’s layer 4, which gave aradiocarbon date on charcoal of 9458 ± 91 years BP(cal years BP 10 738–10 951) (Sauveterrian, ItalianPreboreal Mesolithic facies) (Castelletti et al., 1994;
Figure 1. Location of the sites where osteological remains of latest Pleistocene/earliest Holocene lions were found. Thelow latitudinal position of the localities suggests either that lions were a component of the ‘warm’ interglacialcommunities, or that they were prevented from expanding more diffusedly throughout Europe, possibly because of theinvasive presence of modern humans.
68 M. MASSETI and P. P. A. MAZZA
© 2013 The Linnean Society of London, Biological Journal of the Linnean Society, 2013, 109, 66–77
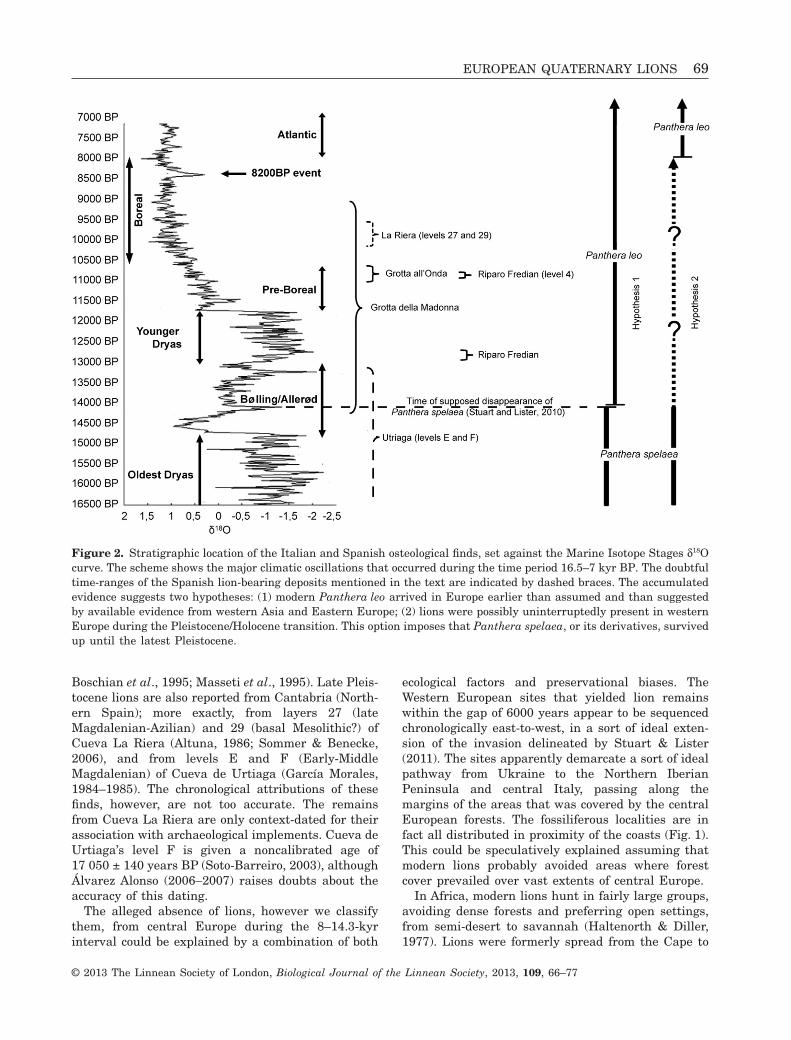
Boschian et al., 1995; Masseti et al., 1995). Late Pleis-tocene lions are also reported from Cantabria (North-ern Spain); more exactly, from layers 27 (lateMagdalenian-Azilian) and 29 (basal Mesolithic?) ofCueva La Riera (Altuna, 1986; Sommer & Benecke,2006), and from levels E and F (Early-MiddleMagdalenian) of Cueva de Urtiaga (García Morales,1984–1985). The chronological attributions of thesefinds, however, are not too accurate. The remainsfrom Cueva La Riera are only context-dated for theirassociation with archaeological implements. Cueva deUrtiaga’s level F is given a noncalibrated age of17 050 ± 140 years BP (Soto-Barreiro, 2003), althoughÁlvarez Alonso (2006–2007) raises doubts about theaccuracy of this dating.
The alleged absence of lions, however we classifythem, from central Europe during the 8–14.3-kyrinterval could be explained by a combination of both
ecological factors and preservational biases. TheWestern European sites that yielded lion remainswithin the gap of 6000 years appear to be sequencedchronologically east-to-west, in a sort of ideal exten-sion of the invasion delineated by Stuart & Lister(2011). The sites apparently demarcate a sort of idealpathway from Ukraine to the Northern IberianPeninsula and central Italy, passing along themargins of the areas that was covered by the centralEuropean forests. The fossiliferous localities are infact all distributed in proximity of the coasts (Fig. 1).This could be speculatively explained assuming thatmodern lions probably avoided areas where forestcover prevailed over vast extents of central Europe.
In Africa, modern lions hunt in fairly large groups,avoiding dense forests and preferring open settings,from semi-desert to savannah (Haltenorth & Diller,1977). Lions were formerly spread from the Cape to
Figure 2. Stratigraphic location of the Italian and Spanish osteological finds, set against the Marine Isotope Stages d18Ocurve. The scheme shows the major climatic oscillations that occurred during the time period 16.5–7 kyr BP. The doubtfultime-ranges of the Spanish lion-bearing deposits mentioned in the text are indicated by dashed braces. The accumulatedevidence suggests two hypotheses: (1) modern Panthera leo arrived in Europe earlier than assumed and than suggestedby available evidence from western Asia and Eastern Europe; (2) lions were possibly uninterruptedly present in westernEurope during the Pleistocene/Holocene transition. This option imposes that Panthera spelaea, or its derivatives, survivedup until the latest Pleistocene.
EUROPEAN QUATERNARY LIONS 69
© 2013 The Linnean Society of London, Biological Journal of the Linnean Society, 2013, 109, 66–77
the Mediterranean wherever suitable prey existed,except in deserts and in tropical and equatorial rainforests (Despard Estes, 1991), although they could befound in other types of forests. In North Africa, theAtlas cedar (Cedrus atlantica, Stephan LadislausEndlicher, Giuseppe Manetti, Elie-Abel Carrière1855) woods were the last refuge for animals threat-ened by hunting and human encroachment, as had
happened for the Barbary lion P. leo leo (L. 1758),which was the typical large-sized sub-species ofNorthern Maghreb: ‘En tiempos no muy lejano yhasta la llegada del siglo XX, el leon del Atlas fue elcazador supremo del bosque de cedros.’ (‘Not too farago and up to the beginning of the XX century, theAtlas lion was the cedar woods’ greatest hunter’)(Charco, 1999). As noted previously, the large Barbarylion survived until 1942 in the Western Atlas(Cabrera, 1932; Panouse, 1957; Aulagnier &Thevenot, 1986; Charco, 1999; Cuzin, 2003).
By contrast, in the Indian subcontinent, lionssurvive only in the dry deciduous forest of the GirWildlife Sanctuary, where they adopt ambush andstalking tactics, hunting singly or in small clans(Despard Estes, 1991; Mitra, 2005). The Gir forest,however, might be a refuge habitat imposed to lionsby human expansion.
The Asian or Indian lions (Panthera leo persica,Meyer, 1826) survived in the Near East well into the20th Century. A group of these big cats lasted inthe gallery forests along the Euphrates at least untilthe 12th Century AD (Mouterde, 1966; Masseti,2009). Remnant populations were reported from theregion of Fethiye, Southern Turkey (Fellows, 1841),on the southern bank of the river Esen (Koca Çay,South-western Anatolia) (Kasparek, 1986; Kasparek& Kasparek, 1990), as well as from the UpperEuphrates valley (Dandford & Alston, 1880; Kinnear,1920; Kumerloeve, 1967; Masseti, 2000). Kasparek(1986) noted that a small population of lions surelysurvived in Southern Turkey at least until the mid19th Century. Lions could still be found along theTigris until 1918, in Iran until 1957, and in Turkeyuntil 1874 (Schnitzler, 2011).
By piecing together this evidence, lions appear toinhabit forested areas permanently only if forced toand with low numbers. This suggests an alternativeexplanation to their absence from more inland Euro-pean areas. Lions who ventured into the forested areasmost probably formed small and rare populations forthe reasons set out above. Forests are known to betaphonomically very poorly conservative habitats(Behrensmeyer, 1991). Hence, the scarce remainsof lions that potentially penetrated the EuropeanHolocene forests would have had extremely lowchances to survive all potential post-mortem destruc-tion and to be captured by the stratigraphic sequences.
However, we cannot exclude the possibility that theapparent east-to-west pathway marked out throughspace and time by the Italian (and possibly also bythe Spanish) lion-bearing sites may instead be totallyaccidental. With such an alternative, we would befaced with the unprecedented scenario of an uninter-rupted occurrence of lions, at least in WesternEurope.
Figure 3. Left third upper deciduous tooth of a lion cubfrom Grotta all’Onda (Camaiore, Lucca, central Italy). A,buccal view. B, lingual view. The specimen was found in astalagmitic crust that gave a U/Th dating of 10 710 ± 200years BP (Berton et al., 2003). Left third upper premolar ofan adult lion from Riparo Fredian (valley of Turrite Secca,upper part of the Serchio Vally, Lucca, central Italy) (cour-tesy C. Tozzi and G. Boschian). C, buccal view. D, lingualview. Left fourth upper premolar (carnassial) of an adultlion from Riparo Fredian (valley of Turrite Secca, upperpart of the Serchio Vally, Lucca, central Italy) (courtesy C.Tozzi and G. Boschian). E, buccal view. F, lingual view. Thearrow in (E) shows the enamel chipped off at the base ofthe paracone of the carnassial tooth. This prevents verifi-cation of the possible presence of a preparastyle, whichdistinguishes Panthera leo from Panthera spelaea (Sot-nikova & Nikolskiy, 2006).
70 M. MASSETI and P. P. A. MAZZA
© 2013 The Linnean Society of London, Biological Journal of the Linnean Society, 2013, 109, 66–77

NOT TOO FAR NORTH
The latest Pleistocene lions were not only preferen-tially distributed in more open, marginal habitats ofWestern Europe. Unlike their earlier counterparts,which dispersed as far north as Britain (Stuart &Lister, 2011), the latest Pleistocene finds are all con-fined to fairly low, temperate latitudes, possibly inresponse to particular ecological constraints. TheWestern European localities that yielded lion remainsare located between the latitudes of 39°N and 44°N(Fig. 1). These carnivores are therefore a componentof the typical ‘warm’ interglacial fauna (Antuneset al., 2008).
The low latitudinal, marginal coastal fringes of theperi-Mediterranean region inhabited by the latestPleistocene Western European lions perhaps offeredrelatively more varied resources compared to moreinland areas. We can speculate that the number ofpotential prey species for lion prides, when consideredon a large scale, was probably higher towards theareas at the margins of the forest canopy, in accord-ance with latitudinal and areal factors.
The modern Western European vegetation startedappearing since the Tertiary (Magri et al., 2007). Theclimatic and vegetational evolution during the Pleis-tocene promoted habitat fragmentation. Remnants ofPleistocene vegetation lingered on until 9000 yearsBP (Huntley, 1988, 1990, 1992; Starkel, 1991; Grove& Rackham, 2001) and these patches of Pleistocenevegetation acted as refugia for Pleistocene and EarlyHolocene fauna (Stewart & Lister, 2001; Reumer,2007). Patchiness increased relative diversity byaccelerating race and species formation amongisolated populations (Finch & Western, 1977). Thenarrower species-ranges produced ungulates of com-paratively smaller body sizes. A trend towardssmaller body size has been observed in several largemammals (mammoth, bison, horse, mastodon, etc.;Reumer, 2007) in the course of the Late Pleistocene.Populations enclosed in these fragments reacted tothe environmental stresses by selectively evolvinginto smaller sizes. This was the result of inbreedingand genetic drift phenomena, which replicated islandevolution on oceanic islands (Reumer, 2007). By con-trast, the habitats at the borders of the great forestedregions were probably not affected by fragmentationand offered a relatively higher variety of prey items.
Modern lions preferably hunt ungulates of40–1200 kg, and especially between 40–60 kg (femalelions) and > 400 kg (male lions) (Wright, 1960; Bour-lière, 1963; Mitchell, Shenton & Uys, 1965; Foster &Kearney, 1967; Kruuk & Turner, 1967; de Pienaar,1969; Schaller, 1972; Elliot & Cowan, 1978; Mills,1984; Van Orsdol, 1984; Ruggiero, 1991; Johnsingh,1992; Mills & Shenk, 1992; Mills & Biggs, 1993;
Palmqvist, Martinez-Navarro & Arribas, 1996;Radloff & Du Toit, 2004). In the Italian and Spanishlocalities noted above, lions are associated with wildboar, roe deer, red deer, aurochs, ibex, and chamois. Inthe Early–Middle Magdalenian levels E and F ofCueva de Urtiaga, lions are also accompanied byhorse, chamois, and reindeer (García Morales, 1984–1985; García & Arsuaga, 2003). Although investiga-tions have not yet been extended to the possibleoccurrence of lions ravaging on ungulate bones, mostof these species (i.e. horse, red deer, reindeer, aurochs,ibex) fall in the range of lions’ potential prey. Usingisotopic methodologies, Bocherens et al. (2011) haveshown that cave lions were predators of reindeer, upuntil the Bölling. Hence, with respect to speculationsabout an interaction between the two species, thecoexistence with reindeer adds credit to the hypoth-esis that these Late Glacial lions might be survivorsof the cave lion lineage.
NOT TOO HIGH
The Italian and Spanish lion-bearing sites prove thatmodern lions patrolled territories that could extend inaltitude up to over 700 m a.s.l. The Grotta all’Ondacave opens on the slopes of Mount Matanna, at thebottom of a morphological crag at an altitude of708 m. Riparo Fredian opens at 400–500 m. Wecannot exclude the possibility that these rupestral,rather than mountainous areas were refugia for lionsthreatened by human encroachment, analogous to theBarbary lion, nor that humans may have selectivelytransported particularly durable joints of felid car-casses from far afield.
LIONS IN WESTERN EUROPEANPREHISTORIC ART
Big cats have been the recurrent subject matters ofmany prehistoric artists. Leroi-Gourhan (1965, 1967)analyzed the characters of the Palaeolithic EuropeanArt’s so-called Classical Style IV (group B). This is aquite realistic art expression, in which animals aredepicted with great anatomical accuracy, and atten-tion is focused on the portrayal of movement. Stylisticcomparisons led Leroi-Gourhan (1965, 1967) to referit to the Magdalenian and advanced Epigravettianand, consequently, to date it between 12 000 and 9000years BP. There are six (or seven) portrayed atLascaux, at least four at Les Trois-Fréres; one in eachof the last set of pictures of Les Combarelles’ secondand third tunnels. All the known pictures of felids arein inconspicuous locations, in secluded rooms of thecaves (Les Trois-Fréres), in narrow, deep tunnels(Fond-de-Gaume), or at the margins of a set of
EUROPEAN QUATERNARY LIONS 71
© 2013 The Linnean Society of London, Biological Journal of the Linnean Society, 2013, 109, 66–77

figures, such as at Labastide and Gabillou (Leroi-Gourhan, 1980). Spassov & Stoychev (2005) considerthat females and subadults are the primary subjectsof cave paintings, whereas adult males are rare.According to Spassov & Stoychev (2005), males areshown at Combarelle 1 and Gabilou, where theyappear with short manes, as if they were hybridsbetween lions and tigers. At Grotte Chauvet (Ardeche,Southern France), which was discovered in the firsthalf of the 1990s, next to the numerous lions, there isalso the image of a leopard, Panthera pardus (L.1758). Although the stylistic comparison is an impor-tant means of dating these works, it can be danger-ously arbitrary. The drawings in the Grotte Chauvethave been dated by the NotCal04 calibrationto approximately 36 kyr BP (cal years BP 41 310)(Chauvet, Brunel Deschamps & Hillaire, 1996;Mellars, 2006).
A common trait of all these lion paintings is thatthey are completely maneless. A mane is barelydrawn on the left side of a lion’s head, in a sort of‘cameo’, on a rock surface at Les Trois-Fréres. Thehead faces the observer, whereas the body is turnedsideways (Ucko & Rosenfeld, 1967).
A presumably adult, maneless lion (Masseti, 2003,2010) was found carved on a limestone block in levelT 10 of Riparo Tagliente (Northwestern Italy)(Fontana, Guerreschi & Liagre, 2002). The specimenwas associated with an inhumated human skeleton,together with final Epigravettian lithic implements(Guerreschi & Squintani, 1995). A sample (OxA –1067) collected from the skeleton’s right femur gave anoncalibrated radiocarbon date of 13 190 ± 90 yearsBP (cal years BP 15 760–15 950) (Bartolomei et al.,1974, 1982; Rocci Ris et al., 2006), which was consist-ent with previous noncalibrated radiocarbon dates of12 650 ± 160 to 13 000 ± 170 kyr BP (cal years BP15 070–15 468 to 15 470–15 820) from the same level(Broglio & Improta, 1994–1995).
The absence of the mane in all cave paintings oflions had been emphasized in numerous studies(Capitan, Breuil & Peyrony, 1924; Friant, 1940;Spassov & Stoychev, 2005) and was regarded as adistinctive character of the cave lion, although thetail, when figured, shows a tuft of hair like that ofmodern lions (Agustí & Antón, 1997; Spassov &Stoychev, 2005). Noteworthy is that none of the por-trayals of modern male lions analyzed by Hemmer(1967) in a comprehensive study of artistic figurationsof the Classical Period (which can be referred toeither P. leo leo or P. leo persica) show the absence ofthe mane. The presence or lack of mane, however,might not have been so diagnostic for the EuropeanPleistocene lions. Nagel et al. (2003) proposed,inspired by West & Packer (2002), a thermoregulatoryexplanation for the lack of mane in European Pleis-
tocene lions, claiming that felids would developmanes only under warm conditions. Nagel et al.(2003) assumed that Pleistocene lions trapped theirinternal heat with a thick fur cover, and thus did notneed the additional isolation of manes. If so, animportant diagnostic discriminant between P. spelaeaand P. leo would be lost. Spassov (1992), however,questioned this opinion, suggesting that the maneplays a role in regulating intraspecific relationships.
DISCUSSION
The migration of modern lions from Africa intoEurope 8000 years ago is largely documented in bothWestern Asia and Eastern Europe. By contrast, thereis no sign of any earlier immigration. Althoughmigrating populations might have left no evidence oftheir passage, the hypothesis that P. leo reachedEurope at the very end of the Pleistocene appears tobe quite improbable. The fact that all the calibratedages of Italian and Spanish specimens fall within thelatest Pleistocene, as well as the association of lionsand reindeer at Cueva de Urtiaga, appears to supportthe second option, which entails that lions were unin-terruptedly present in Western Europe from the timeof Isernia La Pineta up to approximately 8000–7500years ago. This is possible provided that it can bedemonstrated that Western Europe preserved condi-tions favourable to lions compared to anywhere elsethroughout the immense ranges of Eurasia. Solidevidence shows that the species became extinct notonly across Eurasia in the interval approximately14–14.5 cal kyr BP, but also in Alaska/Yukon approxi-mately 1000 years later (Stuart & Lister, 2011). Iflions actually survived in Western Europe, furtherissues need to be addressed: what were the endmembers of this lineage? Did P. spelaea originateindigenous ecological competitors of P. leo? Did theselate, indigenous representatives exhibit gradualmorphological differences over time, without clear,unquestionable taxonomic demarcations? Further-more, were the later, allochthonous groups of Africanlions possibly limited in their penetration by theobtrusive presence of their autochthonous, Europeanrelatives?
CONCLUSIONS
The Middle and early Late Pleistocene big cats werepart of the fauna of Western Europe for over 700 000years and were widespread. They endured the rigorsof at least nine glacial events, some of which (MarineIsotope Stages 10, 6, 2) were particularly intense. Yetthey survived. In the light of the data reported in thepresent study, Western European lions, regardlessof whether they were allochthonous, newly-arrived
72 M. MASSETI and P. P. A. MAZZA
© 2013 The Linnean Society of London, Biological Journal of the Linnean Society, 2013, 109, 66–77

modern lions, or the indigenous derivatives of P. spe-laea, also outlived the cold snaps of the Allerød andYounger Dryas, only to rapidly demise thereafter, asindicated not only by the drastic contraction of theirarea of distribution, but also by the sudden scarcity offinds. This is quite puzzling, especially at a time whenthe general conditions in Europe were improving.Those cold episodes evidently marked a watershed forlions, between a favourable past and a more criticalfuture. Climate likely played a minor role in thischange; however, the ever-growing presence ofhumans must have been more intrusive. Early intheir history, European lions witnessed the dawn,successful spread, and ultimate sunset of the Nean-derthal lineage, and the arrival of Homo sapiens, whosteadily spread thereafter. The expansion of modernhumans most probably impacted on the latest Pleis-tocene and early Holocene lions, and we cannot ruleout the possibility that the latest lions could notparallel the success of their Middle and early LatePleistocene counterparts exactly as a result of theintrusive presence of H. sapiens. Modern humans hadco-evolved alongside P. leo in Africa and had mostprobably developed effective countering strategies(Brantingham, 1998; Stiner, 2002).
In Western Europe, lions were probably mainlyconfined to marginal open habitats at relatively lowlatitudes. The Grotta della Madonna finds show thatthey maintained at southern latitudes, perhapsbecause of the hostile Younger Dryas conditionsfarther north (Starkel, 1991). Lions formed fairlystably settled populations, as the cub teeth from thechronologically coeval Riparo Fredian (level 4) andGrotta all’Onda clearly prove. Nonetheless, the rela-tively high altitudes at which these and all the otherlatest Pleistocene/early Holocene lion-bearing sitesare located suggests that lions were probably gradu-ally pressed towards ever more restricted refugia,possibly under pressure from the growing encroach-ment of humans. Lions lasted a little longer, untiltheir eventual regional extinction from WesternEurope in the early Holocene.
ACKNOWLEDGEMENTS
We wish to thank Dr Fabrice Cuzin of the Laboratoirede Biogéographie et Ecologie des Vertébrès de EcolePratique des hautes Etudes de la Faculté de Sciences,Montellier; Professor Nuria García García of theDepartamento de Paleontología, Facultad de CienciasGeológicas, Universidad Complutense de Madrid andof the Centro de Investigación (UCM-ISCIII) deEvolución y Comportamiento Humanos, Madrid; Pro-fessor Adrian M. Lister of the Department of Palae-ontology of the Natural History Museum, London;Professor Anthony Stuart of the Department of Pal-
aeontology, Natural History Museum, London; andProfessor N. Spassov of the National Museum ofNatural History, Sofia, for their comments and mosthelpful suggestions. The authors would also like tothank Dr Leonardo Salari of the DipartimentoScienze della Terra dell’Università ‘Sapienza’ diRoma, for enjoyable and stimulating discussions onthe earliest Western Palaearctic modern lions. We aremost grateful to Dr Stefania Campetti and to theMuseo Archeologico di Camaiore for granting usaccess to the Grotta all’Onda fossil material. Cordialthanks go to the anonymous reviewers for their mosthelpful comments on the manuscript. The study wasfunded by ex 60%MIUR (Ministry of Education, Uni-versity and Research) grants.
REFERENCES
Agustí J, Antón M. 1997. Memoria de la tierra. Vertebradosfósiles de la Peninsula Ibérica. Barcelona: Ediciones delSerbal.
Alessio M, Bella F, Bachechi F, Cortesi C. 1966. Univer-sity of Rome, Carbon-14 Dates IV. Radiocarbon 8: 401–404.
Alessio M, Bella F, Bachechi F, Cortesi C. 1967. Univer-sity of Rome, Carbon-14 Dates V. Radiocarbon 9: 354–357.
Altuna J. 1986. The mammalian fauna from the prehistoricsite of La Riera. In: Straus LG, Clark G, eds. La Riera Cave.Stone age hunter gatherer adaptations in northern Spain.Tempe, AR: Anthropological Research Paper 36, ArizonaState University, 237–274, 421–479, 481–497.
Álvarez Alonso D. 2006–2007. The Cantabrian Magdale-nian. Lateglacial Chronology in Northern Iberian Penin-sula. Portugalia, Nova Série 27–28: 5–18.
Álvarez-Lao D, García-García N. 2006. A new site from theSpanish Middle Pleistocene with cold-resistant faunal ele-ments: La Parte (Asturias, Spain). Quaternary International142–143: 107–118.
Anconetani P, Ferrari M, Giusberti G, Peretto C. 1995.Analisi dei resti ossei fratturati intenzionalmente del giaci-mento di Isernia La Pineta (Molise, Italia): reperti dellaporzione più profonda dell’archeosuperficie T 3A del Isettore di scavo. In: Peretto C, De Curtis O, eds. Atto del 1°Convegno Nazionale di Archeozoologia (Rovigo, 5–7 marzo1993). Rovigo: Padusa Quaderni, 1: 55–59.
Antunes A, Troyer JL, Roelke ME, Pecon-Slattery J,Packer C, Winterbach C, Winterbach H, Hemson G,Frank L, Stander P, Siefert L, Driciru M, Funston PJ,Alexander KA, Prager KC, Mills G, Wildt D, Bush M,O’Brien SJ, Johnson WE. 2008. The evolutionary dynam-ics of the lion Panthera leo revealed by host and viralpopulation genomics. PLoS Genetics 4: e1000251.
Argant A. 1988. Etude de l’exemplaire de Panthera spelaea(Goldfuss, 1810) (Mammalia, Carnivora, Felidae) du gise-ment Pleistocene Moyen recent de la Grotte d’Aze (Saone etLoire). Revue de Paleobiologie 7: 449–466.
Aulagnier S, Thevenot M. 1986. Catalogue des mammiferessauvages du Maroc. Rabat: Royame du Maroc, Institut
EUROPEAN QUATERNARY LIONS 73
© 2013 The Linnean Society of London, Biological Journal of the Linnean Society, 2013, 109, 66–77

Scientifique, Universite Mohammed V/Ministere del’Education Nationale.
Ballesio R. 1980. Le gisement pléistocene supérieure lagrotte de Jaurens à Nespouls, Corrèze, France.; les Carni-vores (Mammalia, Carnivora), 2 – Felidae. NouvellesArchives du Museum d’Histoire Naturelle de Lyon 18:61–102.
Barnett R, Shapiro B, Barnes I, Ho SY, Burger J,Yamaguchi N, Higham TFG, Wheeler HT, RosendahlW, Sher AV, Sotnikova M, Kuznetsova T, BaryshnikovGF, Martin LD, Harington CR, Burns JA, Cooper A.2009. Phylogeography of lions (Panthera leo ssp.) revealsthree distinct taxa and a Late Pleistocene reduction indiversity. Molecular Ecology 18: 1668–1677.
Barnett R, Yamaguchi N, Barnes I, Cooper A. 2006.The origin, current diversity and future conservation ofthe modern lion (Panthera leo). Proceedings of the RoyalSociecty of London Series B, Biological Sciences 273: 2119–2125.
Bartolomei G, Broglio A, Cattani L, Cremaschi M, Guer-reschi A, Mantovani E, Peretto C, Sala B. 1982. Idepositi würmiani del Riparo Tagliente. 4. Ferrara, Italy:Annali dell’Università di Ferrara, ns, 15 (3).
Bartolomei G, Broglio A, Guerreschi A, Leonardi P,Peretto C, Sala B. 1974. Una sepoltura epigravettiana neldeposito pleistocenico del Riparo tagliente in Valpantena(Verona). Rivista di Scienze Preistoriche 29: 1–52.
Bartosiewicz L. 2009. A lion’s share of attention: archaeol-ogy and the historical record. Acta Archaeologica AcademiaeScientarium Hungaricae 60: 275–289.
Baryshnikov G. 2011. Pleistocene Felidae (Mammalia, Car-nivora) from the Kudaro Paleolithic cave sites in the Cau-casus. Proceedings of the Zoological Institute RAS 315:197–226.
Baryshnikov G, Boeskorov G. 2001. The Pleistocene cavelion Panthera spelaea (Carnivora, Felidae) from Yakutia,Russia. Cranium 18: 7–24.
Becker C. 1986. Kastanas. Die Tierknochenfunde. Prähistor-ische Archäeologie in Südesteuropa. 5. Berlin: Wissen-schaftsverlag Volker Spiess.
Behrensmeyer AK. 1991. Terrestrial vertebrate accumula-tions. In: Allison PA, Briggs DEG, eds. Releasing the datalocked in the fossil record. Taphonomy. New York, NY:Plenum Press, 291–335.
Berton A, Bonato M, Borsato A, Campetti S, Fabbri PF,Mallegni F, Perrini L, Piccini L. 2003. Nuove datazioniradiometriche con il metodo U/Th sulle formazioni stalag-mitiche di Grotta all’Onda. Rivista di Scienze Preistoriche53: 241–256.
Bischoff JL, Williams RW, Rosenbauer RJ, Aranburu A,Arsuaga JL, García N, Cuenca-Bescós G. 2007. High-resolution U-series dates from the Sima de los Huesos yields600 + 8 - 66 kyrs: implications for the evolution of the Nean-derthal lineage. Journal of Archaeological Science 34: 763–770.
Bocherens H, Drucker DG, Bonjean D, Bridault A,Conard NJ, Cupillard C, Germonpré M, Höneisen M,Münzel SC, Napierrala H, Patou-Mathis M, Stephan E,
Uerpmann H-P, Ziegler R. 2011. Isotopic evidence fordietary ecology of cave lion (Panthera spelaea) in North-western Europe: prey choice, competition and implicationsfor extinction. Quaternary International 245: 249–261.
Boschian G, Mallegni F, Tozzi C. 1995. The Epigravet-tian and Mesolithic site of Fredian Shelter (in Tuscany).Quaternaria Nova 5: 45–80.
Bourlière F. 1963. Specific feeding habits of Africancarnivores. African Wildlife Journal 17: 21–27.
Brantingham JP. 1998. Hominid–carnivore coevolution andinvasion of the predatory guild. Journal of AnthropologicalArchaeology 17: 327–353.
Broglio A, Improta S. 1994–1995. Nuovi dati di cronologiaassoluta del Paleolitico superiore e del Mesolitico delVeneto, del Trentino e del Friuli. Atti dell’Istituto Veneto diScienze, Lettere ed Arti 153: 9–28.
Burger J, Rosendahl W, Loreille O, Hemmer H, Eriks-son T, Götherström A, Hiller J, Collins MJ, Wess T,Alta KW. 2004. Molecular phylogeny of the extinct cavelion Panthera leo spelaea. Molecular Phylogenetics andEvolution 30: 841–849.
Cabrera A. 1932. Los mamíferos de Marruecos. Trabajos delMuseo Nacional de Ciencias Naturales, Serie Zoologica 57:1–362.
Capitan L, Breuil H, Peyrony D. 1924. Les Combarellesaux Eyzies (Dordogne). Paris: Masson.
Cardini L. 1972. Praia a Mare. Relazione degli scavi1957–1970 dell’Istituto Italiano di Paleontologia Umana.Bullettino di Paletnologia Italiana 79: 31–59.
Castelletti L, Maspero A, Tozzi C. 1994. Il popolamentodella Valle del Serchio (Toscana settentrionale) duranteil Tardiglaciale würmiano e l’Olocene antico. In: Biagi P,Nandris J, eds. Highland Zone Exploitation in SouthernEurope. Brescia: Monografie di Natura Bresciana, 20: 189–204.
Charco J. 1999. El bosque mediterráneo en el norte de África.Biodiversidad y lucha contra la desertification. Madrid:Agenzia Española de Cooperación Internacional.
Chauvet J-M, Brunel Deschamps E, Hillaire C. 1996.Chaivet Cave. The discovery of the word’s oldest paintings.London: Thames and Hudson.
Cilli C. 1993–1994. Analisi archeozoologica e tafonomica deiresti faunistici del Riparo Fredian (Molazzana, Lucca).DPhil Thesis, University of Turin, Italy.
Cilli C, Malerba G, Tozzi C. 1998. Analyse archeozoologiqueet modifications de surface des restes fauniques de deuxsites du Paléolithique supérieur de Toscane (Italie). ActesUnion Internationale des Sciences Préhistoriques et Proto-historiques 13: 675–679.
Croitor R, Brugal J. 2010. Ecological and evolutionarydynamics of the carnivore community in Europe during thelast 3 million years. Quaternary International 212: 98–108.
Cuzin F. 2003. La régression des grands Mammifères terres-tres du Maroc méridional au cours du XXe siècle. NaturaliaMaroccana 1: 93–99.
Dandford CG, Alston ER. 1880. On the mammals of AsiaMinor. Part II. Proceedings of the Zoological Society ofLondon 1880: 50–64.
74 M. MASSETI and P. P. A. MAZZA
© 2013 The Linnean Society of London, Biological Journal of the Linnean Society, 2013, 109, 66–77

Despard Estes R. 1991. The behavior guide to Africanmammals. Including hoofed mammals, carnivora, primates.Berkley, CA: The University of California Press.
Dietrich WD. 1968. Fossile Löwen in europäischen and afri-kanischen Pleistozän. Paläontologische AbhandlungenBerlin (A) 3: 323–366.
Elliot JP, Cowan IM. 1978. Territoriality, density, and preyof the lion in Ngorongoro Crater, Tanzania. CanadianJournal of Zoology 56: 1726–1734.
Fellows C. 1841. An account of discoveries in Lycia, being aJournal kept during a second Excursion in Asia Minor(1840). London.
Finch VA, Western D. 1977. Cattle colours in pastoral herds:natural selection or social preference. Ecology 58: 1384–1392.
Fiore I, Pino Urìa B, Tagliacozzo A. 2004. L’exploita-tion des petits animaux au Paléolithique supérieur –Mésolithique en Italie: l’exemple de la Grotta del Santuario
della Madonna a Praia a Mare (Cosenza, Italie). In: BrugalJ-P, Desse J, eds. Petits animaux et sociétés humaines. Ducomplément alimentaire aux ressources utilitaires. Antibes:Rencontres Internationales Archéologie Histoire d’Antibes,XXIV, APDCA, 417–430.
Fontana F, Guerreschi A, Liagre J. 2002. RiparoTagliente. La serie epigravettiana. In: Aspes A, ed. Preisto-ria Veronese. Contributi e aggiornamenti. Verona: Memoriedel Museo Civico di Storia Naturale di Verona, II serie, Sez.Scienze dell’Uomo, 5: 42–47.
Foster J, Kearney D. 1967. Nairobi National Park gamecensus, 1966. East African Wildlife Journal 5: 112–120.
Friant M. 1940. Sur la question des affinités du Grand Chatdes Cavernes. Mammalia 4: 87–93.
García N, Arsuaga JL. 1998. The carnivore remainsfrom the hominid-bearing Trinchera-Galería, Sierra deAtapuerca, Middle Pleistocene site (Spain). Geobios 31: 659–674.
García N, Arsuaga JL. 2001. Les carnivores (Mammalia)des sites du Pleistocene ancien et moyen d’Atapuerca(Espagne). L’Anthropologie 105: 83–93.
García N, Arsuaga JL. 2003. Last Glaciation cold-adaptedfaunas in the Iberian Peninsula. In: Reumer JWF, de Vos J,Mol D, eds. Advances in mammoth research (Proceedings ofthe Second International Mammoth Conference, Rotterdam,16–20 May 1999). Rotterdam: Deinsea, 9: 159–169.
García García N, Feranec RS, Arsuaga JL, Bermúdez deCastro JM, Carbonell E. 2009. Isotopic analysis of theecology of herbivores and carnivores from the Middle Pleis-tocene deposits of the Sierra De Atapuerca, northern Spain.Journal of Archaeological Science 36: 1142–1151.
García Morales M. 1984-1985. Sobre el valor cronológico dela presencia del ciervo y el reno en el arte PaleolíticoCantábrico. Zephyrus 37–38: 125–136.
Grove AT, Rackham O. 2001. The nature of MediterraneanEurope. An ecological history. New Haven, CT: Yale Univer-sity Press.
Guerreschi A, Squintani MB. 1995. La prima officina liticaepigravettiana (T. 10A) di Riparo Tagliente (Grezzana,Verona). Bullettino di Paletnologia Italiana 86: 19–58.
Haltenorth T, Diller H. 1977. Säugetiere Afrikas und Mada-gaskars. München: BLV Verlagsgesellschaft mbH.
Harper F. 1945. Extinct and Vanishing Mammals of the OldWorld. Special Publication No.12. American Committee forInternational Wild Life Protection. New York, NY: Zoologi-cal Park.
Hemmer H. 1967. Uber das Aiussehen der klein-bzw. vorder-asiatischen Löwen (Panthera leo ssp. persica). Säuge-tierkundliche Mitteilungen 15: 50–53.
Hemmer H. 1974. Untersuchungen zur Stammesgeschichteder Pantherkatzen (Pantherinae) Teil 3. Zur Artgeschichtede Löwen Panthera (Panthera) leo (Linnaeus, 1758). Verof-fentlichungen der Zoologischen Staatssammlung München17: 167–280.
Huntley B. 1988. Europe. In: Huntley B, Webb T, eds. Vegeta-tion history. London: Kluwer Academic Publishers, 341–384.
Huntley B. 1990. European post-glacial forests: composi-tional changes in response to climatic change. Journal ofVegetation Science 1: 507–518.
Huntley B. 1992. Pollen-climate response surfaces and thestudy of climate change. In: Gray JM, ed. Applications ofquaternary research. Quaternary Proceedings 2. Cambridge:Quaternary Research Association, 91–99.
Janossy D. 1969. Stratigraphische Auswertung dedseuropäiscehn mittelpleistozän Wirbelfauna. Teil II. Berichteder Gesellschaft für Geologische Wissenschaften, Reihe A,Geologie und Paläontologie 14: 573–643.
Johnsingh AJT. 1992. Prey selection in three large sympat-ric carnivores in Bandipur. Mammalia 56: 517–526.
Jöris O, Weninger B. 1998. Extension of the 14C calibrationcurve to ca. 40,000 cal BC by synchronizing Greenland180/160 ice core records and North Atlantic Foraminiferaprofiles: a comparison with U/Th coral data. Radiocarbon40: 495–504.
Kasparek A, Kasparek M. 1990. Reiseführer Natur Türkei.München: BLV Verlagsgesellschaft mbH.
Kasparek M. 1986. On a historical occurrence of the lion,Panthera leo, in Turkey. Zoology in the Middle East 1: 9–10.
Kinnear NB. 1920. The past and present distribution of thelion in south eastern Asia. Journal of Bombay NaturalHistory Society 27: 33–39.
Kitagawa H, van der Plicht J. 1996. 40,000-year varvechronology from lake Suigetsu, Japan: extension of the 14Ccalibration curve. Radiocarbon 40: 505–515.
Krakhmalnaya TV. 1999. Carnivore remains from LatePleistocene and Holocene deposits in the Ukraine. Archäolo-gie in Eurasien 6: 223–235.
Kruuk H, Turner M. 1967. Comparative notes on predationby lion, leopard, cheetah and wild dog in the Serengeti area,East Africa. Mammalia 31: 1–27.
Kumerloeve H. 1967. Zur Verbreitung kleinasiatischerRaub-und Huftiere sowie einer Grossnager. Säugetierkund-liche Mitteilungen 15: 337–409.
Kurtén B. 1968. Pleistocene mammals of Europe. London:Weidenfeld and Nicholson.
Kurtén B. 1985. The Pleistocene lion of Beringia. AnnalesZoologici Fennici 22: 117–121.
Lefèvre F, Mulliez D, Oulhen J, Rousset D, Luce J-M,
EUROPEAN QUATERNARY LIONS 75
© 2013 The Linnean Society of London, Biological Journal of the Linnean Society, 2013, 109, 66–77

Moretti J-C, Déroche V, Pétridis P. 1992. Etudes, chro-niques et rapports: Delphes. Bulletin de correspondancehellénique 116: 686–711.
Leroi-Gourhan A. 1965. Préhistoire de l’art occidental.Mazenod, Paris: L’art et les grandes civilisations.
Leroi-Gourhan A. 1967. Treasures of prehistoric art. NewYork, NY: Abrams.
Leroi-Gourhan A. 1980. Les Débuts de l’Art. Colloquesinternationaux du C.N.R.S. 599. Les Processus del’Hominisation 599: 131–132.
Lewis M, Pacher M, Turner A. 2010. The larger Carnivoraof the West Runton Freshwater Bed. Quaternary Interna-tional 228: 116–135.
Magri D, Fineschi S, Bellarosa R, Buonamici A, Sebas-tiani F, Schirone B, Simeone MC, Vendramin GG.2007. The distribution of Quercus suber chloroplast haplo-types matches the palaeogeographical history of the westernMediterranean. Molecular Ecology 16: 5259–5266.
Masseti M. 2000. Wild cats (Mammalia, Carnivora) of Ana-tolia. With some observations on the former and presentoccurrence of leopards in south-eastern Turkey and on theGreek island of Samos. Biogeographia 20: 607–618.
Masseti M. 2003. Estinzione dei carnivori e degli ungulatiitaliani nell’Olocene. In: Boitani L, Lovari S, Vigna TagliantiA, eds. Fauna d’Italia. Mammalia III. Carnivora – Artio-dactyla. Bologna: Edizioni Calderini de Il Sole 24 OreEdagricole, 1–16.
Masseti M. 2009. Carnivores of Syria. In: Neubert E, Amr Z,Taiti S, Gümüs B, eds. Animal biodiversity in the MiddleEast. Proceedings of the First International Congress: Docu-menting, Analysing and Managing Biodiversity in theMiddle East. Aqaba, Jordan: ZooKeys, 31: 229–252.
Masseti M. 2010. Zoologia storica e archeologica dei felidiitaliani. In: Randi E, Ragni B, Bizzarrini L, Agostini N,Tedaldi G, eds. Biologia e conservazione dei Felidi in Italia.Atti del convegno – Santa Sofia (FC), 7–8 Novembre 2008.Arezzo, Italy: Ente Parco Nazionale Foreste Casentinesi,9–28.
Masseti M, Mazza P, Rustioni M, Sala B. 1995. Large-sized Italian ungulates at the Late Pleistocene–Holocenetransition: an overview. In: Peretto C, De Curtis O, eds. Attodel 1° Convegno Nazionale di Archeozoologia (Rovigo, 5–7marzo 1993). Rovigo: Padusa Quaderni, 1: 89–96.
Mellars P. 2006. Going East: new genetic and archaeologicalperspectives on the modern human colonization of Eurasia.Science 313: 796–800.
Meyer AB. 1903. The antiquity of the lion in Greece.Zoologischer Garten Berlin 44: 65–73.
Mills MGL. 1984. Prey selection and feeding habits ofthe large carnivores in the southern Kalahari. Koedoe 27(Suppl.): 281–294.
Mills MGL, Biggs HC. 1993. Prey apportionment andrelated ecological relationships between large carnivores inKruger National Park. Symposium of the Zoological Societyof London 65: 253–268.
Mills MGL, Shenk TM. 1992. Predator–prey relationships:the impact of lion predation on wildbeest and zebra popu-lations. Journal of Animal Ecology 61: 693–702.
Mitchell BL, Shenton JB, Uys JCM. 1965. Predation onlarge mammals in the Kafue National Park, Zambia. Zoo-logica Africana 1: 297–318.
Mitra S. 2005. Gir forest and the saga of the Asiatic Lion.Delhi: Indus Publishing Company.
Molara G. 2012. Resti faunistici provenienti dai livelli delPleistocene superiore di Grotta all’Onda (Camaiore, Lucca).In: De Grossi Mazzorin J, Saccà D, Tozzi C, eds. Atti del 6°Convegno Nazionale di Archeozoologia. Pisa: AssociazioneItaliana di Acheozoologia/Dipartimento di Scienze Archeo-logiche, Università di Pisa, 57–62.
Mouterde P. 1966. Nouvelle flore du Liban et de la Syria.Lyban I: Editions de l’imprimerie catholique Beyrouth T I,II, III.
Münzel SC, Hofreiter M, Rabeder G, Bocherens H, Uer-pmann HP, Conard NJ. 2007. Correlating genetic resultsfor the replacement of cave bears at 28,000 B.P. from theAch Valley (Swabian Jura) with metrical and morphologicaldata. Scripta Facultatis Scientiarium Naturalium Univer-sitatis Masarykianae Brunensis 35: 123–128.
Münzel SC, Hofreiter M, Stiller M, Conard NJ, Bocher-ens H. 2008. Neue Ergebnisse zur Alb (Chronologie, Iso-topie und Paläogenetik). Stalactite 58: 27–30.
Münzel SC, Hofreiter M, Stiller M, Mittnik A, ConardNJ, Bocherens H. 2011. Pleistocene bears in the SwabianJura (Germany): genetic replacement, ecological displace-ment, extinctions and survival. Quaternary International245: 225–237.
Nagel D, Hilsberg S, Benesch A, Scholz J. 2003. Func-tional morphology and fur patterns in Recent and fossilPanthera species. Scripta Geologica 126: 227–241.
Ninov L. 1999. Vergleichende Untersuchungen zur Jagd undzum Jagdwild während des Neolithikums und Äneolithi-kums in Bulgarien. Archäologie Eurasien 6: 323–338.
Palmqvist P, Martinez-Navarro B, Arribas A. 1996. Preyselection by terrestrial carnivores in a Lower Pleistocenepaleocommunity. Paleobiology 22: 514–534.
Panouse JB. 1957. Les Mammifères du Maroc. Travaux del’Institut scientifique chérifien. Série Zoologie 5: 1–206.
de Pienaar U. 1969. Predator–prey relations amongst thelarger mammals of the Kruger National Park. Koedoe 12:108–176.
Por FD. 1987. The levantine landbridge: historical andpresent patterns. In: Krupp F, Schneider W, Kinzelbach R,eds. Proceedings of the Symposium on the Fauna and Zoog-eography of the Middle East. Wiesbaden: Dr LudwigReichert Verlag, 23–28.
Radloff FGT, Du Toit JT. 2004. Large predators and theirprey in a southern African savanna: a predator’s size deter-mines its prey size range. Journal of Animal Ecology 73:410–423.
Reimer PJ, Baillie MGL, Bard E, Bayliss A, Beck JW,Blackwell PG, Bronk Ramsey C, Buck CE, Burr GS,Edwards RL, Friedrich M, Grootes PM, GuildersonTP, Hajdas I, Heaton TJ, Hogg AG, Hughen KA, KaiserKF, Kromer B, McCormac FG, Manning SW, ReimerRW, Richards DA, Southon JR, Talamo S, TurneyCSM, van der Plicht J, Weyhenmeyer CE. 2009.
76 M. MASSETI and P. P. A. MAZZA
© 2013 The Linnean Society of London, Biological Journal of the Linnean Society, 2013, 109, 66–77

INTCAL09 and MARINE09 radiocarbon age calibrationcurves, 0–50,000 years Cal BP. Radiocarbon 51: 1111–1150.
Reumer JWF. 2007. Habitat fragmentation and the extinc-tion of mammoths (Mammuthus primigenius, Proboscidea,Mammalia): arguments for a causal relationship. In: KahlkeR-D, Maul LC, Mazza PP, eds. Late Neogene and Quater-nary biodiversity and evolution: regional developments andinterregional correlations. Vol. I. Proceedings of the 18thInternational Senckenberg Conference (VI International Pal-aeontological Colloquium in Weimar). Stuttgart: CourierForschungsinstitut Senckenberg, 256: 279–286.
Rocci Ris A, Cilli C, Malerba G, Giacobini G, GuerreschiA. 2006. Archeozoologia e tafonomia dei reperti provenientida un livello epigravettiano (taglio 10) di Riparo Tagliente(Grezzana, VR). In: Malerba G, Visentini P, eds. Atti del 4°Convegno Nazionale di Archeozoologia, Pordenone, 13–15novembre 2003. Pordenone: Quaderni Museo ArcheologicoFriuli Occidentale, 6: 111–123.
Ruggiero RG. 1991. Prey selection of the lion (Panthera leoL.) in Manovo-Gounda-St Floris National Park, CentralAfrican Republic. Mammalia 55: 23–33.
Sala B. 1990. Panthera leo fossilis (v. Reich., 1906) (Felidae)de Isernia la Pineta (Pléistocène moyen inférieur d’Italie).Geobios 23: 189–194.
Salari L. 2006. Mosaico Nilotico di Palestrina: animali realio fantastici? Archeozoologia di una produzione artistica dietà ellenistica. Palestrina, Rome: Circolo Culturale Pre-nestino Simeoni.
Schaller GB. 1972. The Serengeti lion. A study of predator–prey relations. Chicago, IL: University of Chicago Press.
Schnitzler AE. 2011. Past and present distribution of theNorth African–Asian lion subgroup: a review. MammalReview 41: 220–243.
Sommer RS, Benecke N. 2006. Late Pleistocene andHolocene development of the felid fauna (Felidae) of Europe:a review. Journal of Zoology 269: 7–19.
Sotnikova M, Nikolskiy P. 2006. Systematic position of thecave lion Panthera spelaea (Goldfuss) based on cranial anddental characters. Quaternay International 142/143: 218–228.
Soto-Barreiro MJ. 2003. Cronología Radiométrica, Ecologíay Clima del Paleolítico Cantábrico. Madrid: Monografías delMuseo Nacional y Centro de Investigación de Altamira 19.In Spanish.
Spassov N. 1992. Display and hyperbolization of signalsymbols: camels and two-horn rhinoceroses. In: Proceedingsof the symposium. ‘Ungulates 91’, Toulouse 2-6 September1991. IRGM – I.N.R.A., Toulouse, 467–470.
Spassov N, Bozkov D. 1982. Rapport sur le lion de caverne etle lion de l’antiquite aux Balkans et en Bulgarie. EuropeanRegional Conference on Speleology. Proceedings I: 228–233.
Spassov N, Iliev N. 1994. Animal remains from thesubmerged Late Eneolithic – early Bronze Age settlementnear in Sozopol (the South Bulgarian Black Sea Coast).Proceedings of the International Symposium ThraciaPontica 6: 287–314.
Spassov N, Popov V. 2007. History of the formation of theBulgarian mammal fauna. In: Miteva S, Mihova B, GeorgievK, Petrov B, Vansink D, eds. The mammals, important forconservation in Bulgaria. Arnhem: Dutch Mammal SocietyVZZ, 31–46 (in Bulgarian).
Spassov N, Stoychev T. 2005. In the shadow of the cave lionand other cats: leopards and snow leopards in the prehis-toric art of Europe, Caucasus and central Asia. Annuary ofDepartment of Archaeology – NBU 6: 5–15.
Starkel L. 1991. Environmental changes at the YoungerDryas – Preboreal Transition and during the earlyHolocene: some distinctive aspects in central Europe. TheHolocene 1: 234–242.
Stewart JR, Lister AM. 2001. Cryptic northern refugia andthe origins of the modern biota. Trends in Ecology andEvolution 16: 608–613.
Stiner MC. 2002. Carnivory, coevolution, and the geographicspread of the genus Homo. Journal of ArchaeologicalResearch 10: 1–62.
Stuart AJ, Lister AM. 2011. Extinction chronology of thecave lion Panthera spelaea. Quaternary Science Review 30:17–18.
Stuiver M, Reimer PJ, Bard E, Beck JW, Burr GS,Hughen KA, Kromer B, McCormac G, van der Plicht J,Spurk M. 1998. IntCal98 radiocarbon age calibration,24,000–0 cal BP. Radiocarbon 40: 1041–1083.
Turner A. 1984. Dental sex dimorphism in European lions(Panthera leo L.) of the Upper Pleistocene: palaeoecologicalimplications. Annales Zoologici Fennici 21: 1–8.
Turner A, Antón M. 1997. The big cats and their fossilrelatives: an illustrated guide to their evolution and naturalhistory. New York, NY: Columbia University Press.
Ucko PJ, Rosenfeld A. 1967. Palaeolithic cave art. London:World University Library.
Van Orsdol KG. 1984. Foraging behavior and huntingsuccess of lions in Queen Elizabeth National Park, Uganda.African Journal of Ecology 22: 79–99.
Verescagin NK. 1969. Le lion des caverns: Panthera (Leo)spelaea Goldfuss et son histoire dans l’Holarctique. Etudessur le quaternaire dans le monde. VIII Congress INQUA,Paris 1: 463–464.
Vereshchagin NK. 1971. The cave lion and its history inHolarctic and in USSR. Trudy Zoologicheskogo institute 49:123–199.
West PM, Packer C. 2002. Sexual selection, temperature,and the lion’s mane. Science 297: 1339–1343.
Wright BS. 1960. Predation on big game in East Africa.Journal of Wildlife Management 24: 1–15.
Yamaguchi N, Cooper A, Werdelin L, MacDonald DW.2004. Evolution of the mane and group-living in the lion(Panthera leo): a review. Journal of Zoology 263: 329–342.
Yannouli E. 2003. Non-domestic carnivores in Greek prehis-tory: a review. In: Kotjabopoulou E, Hamilakis Y, HalsteadP, Gamble C, Elefanti P, eds. Zooarchaeology in Greece,recent advances, studies 9, 18. Athens: British School atAthens, 175–192.
EUROPEAN QUATERNARY LIONS 77
© 2013 The Linnean Society of London, Biological Journal of the Linnean Society, 2013, 109, 66–77