vip and pacap display different vasodilatory effects in rabbit coronary and cerebral arteries
TRANSCRIPT

VIP and PACAP display different vasodilatory effects in rabbit
coronary and cerebral arteries
Torur Dalsgaarda,*, Jens Hannibalb, Jan Fahrenkrugb, Christian Rifbjerg Larsena, Bent Ottesena
aDepartment of Obstetrics and Gynaecology 537, Hvidovre University Hospital, Kettegard Alle 30, DK-2650 Hvidovre, Copenhagen, DenmarkbDepartment of Clinical Biochemistry, Bispebjerg University Hospital, Copenhagen, Denmark
Received 7 February 2002; received in revised form 23 August 2002; accepted 26 August 2002
Abstract
Vasoactive intestinal polypeptide (VIP) and pituitary adenylate cyclase activating polypeptide (PACAP) are closely related peptides with
wide distribution in the nervous system. The aim of the present study was to investigate functional characteristics and the influence of sex
steroids on the vasodilatory effects of these two peptides in cerebral and coronary vessels from female New Zealand White (NZW) rabbits.
The localization and concentration of VIP and PACAP in cardiovascular tissue was evaluated using immunohistochemistry and
radioimmunoassays. The vasodilatory effects of VIP and PACAP were investigated using myographs, allowing isometric tension recordings.
In order to evaluate the influence of steroid hormones, the rabbits were ovariectomized and randomized to treatment for 4 weeks with 17h-estradiol (E2), Norethindrone Acetate (NETA), E2 +NETA or placebo. Ring segments of the posterior cerebral artery, the right proximal
coronary artery and the distal left coronary artery were examined.
The highest concentrations of VIP/PACAP were observed in cerebral and coronary arteries: 5.0/5.7 and 2.8/3.5 pmol/g, respectively. The
peptides were localized in nerve fibres innervating the arteries. Both peptides produced dose-dependent vasodilatory responses in all vessels
investigated. While the effects of PACAP were identical in cerebral and coronary arterial segments, the effects of VIP displayed significant
differences (Emax, pI2, Hill-slope). Treatment with sex steroids induced no changes in the vascular effects of the two peptides.
These results indicate different mechanisms of action for the vasodilating effects of the two closely related peptides VIP and PACAP in
different areas of the coronary and cerebrovascular tree. Treatment with female sex steroids does not seem to change these mechanisms.
D 2002 Elsevier Science B.V. All rights reserved.
Keywords: Cardiovascular diseases; Blood flow; Hormones; Peptides
1. Introduction
Vasoactive intestinal polypeptide (VIP) and pituitary
adenylate cyclase activating polypeptide (PACAP) are two
structurally closely related neuropeptides with widespread
distribution in the central and peripheral nervous system and
have a broad spectrum of biological actions [1–3]. PACAP
demonstrates 68% sequence homology with VIP in the N-
terminal 28 amino acids [4]. PACAP is present in two
amidated forms, the 38-amino-acid residues PACAP-38
and the C-terminally truncated PACAP-27, both derived
from a 175-amino-acid precursor [5].
Both VIP and PACAP-27/PACAP-38 have been shown
to be potent vasodilators in several species. PACAP acts via
three types of receptors, of which one is specific for PACAP
(PAC1) and two are shared with VIP (VPAC1 and VPAC2)
[6]. In humans, VIP dilates cerebral [7,8] and coronary
arteries [9,10]. Correspondingly, VIP has been shown to
dilate rabbit cerebral arteries [11] and rabbit coronary
arteries [12]. VIP-immunoreactive neuronal cell bodies have
been located to the interatrial septa of the heart [12]. Both
PACAP-27 and PACAP-38 dilate human coronary arteries
[13,14], but PACAP-27 has been found not to affect human
middle cerebral arteries [15]. However, our knowledge of
the effect of VIP and PACAP on the arteries in the heart and
brain is still limited.
Previously, it has been observed in animals that treatment
with estradiol increases the expression of all pre-pro-VIP-
derived peptides except PHI/PHV [16], and in postmeno-
pausal women, it has been shown that the vasodilatory effect
of VIP on vaginal blood vessels is dependent on the
presence of oestrogen [17]. Furthermore, the vasomotor
0167-0115/02/$ - see front matter D 2002 Elsevier Science B.V. All rights reserved.
PII: S0167 -0115 (02 )00205 -7
* Corresponding author. Tel.: +45-36-32-33-76; fax: +45-36-32-37-23.
E-mail address: [email protected] (T. Dalsgaard).
www.elsevier.com/locate/regpep
Regulatory Peptides 110 (2003) 179–188

responses of isolated rabbit coronary arteries to VIP have
been demonstrated to change with age and the development
of sexual maturity, indicating a relationship with sex ste-
roids [18].
Due to the presence and significant cardiovascular effects
of VIP in the heart, the peptide has recently been suggested
to be important in the regulation of coronary blood flow
[19]. The mechanism of the putative beneficial cardiovas-
cular effects produced by estradiol in postmenopausal
women might therefore involve VIP and/or PACAP.
In non-hysterectomized women, postmenopausal hor-
mone replacement therapy (HRT) consists of a combination
of oestrogen (as 17h-estradiol) and progestogen (as the
synthetic compound Norethindrone Acetate). These are
female sex steroids, inducing their action mainly by chan-
ging the transcription of DNA in the cell nucleus via
specific intracellular steroid receptors. Phytoestrogens are
plant-derived molecules possessing oestrogen-like activity,
most of them with the ability of inducing different estro-
genic effect in different cells and therefore also termed
selective estrogen receptor modulators (SERMs).
Based on this, the aim of the present study was to
determine the presence of VIP and PACAP in cardiovascu-
lar tissue and to examine the vascular effects of the two
peptides on cerebral and coronary arteries from normal and
sex steroid-treated rabbits [20].
2. Materials and methods
2.1. Animals
Material and methods are described earlier in detail for a
similar study of vascular function in atherosclerotic rabbits
[21]. In short, the rabbits used in the present study were
New Zealand White (NZW) rabbits (SSI:CPH), which
during the study were housed individually in stainless steel
cages under regulated environmental conditions. Protocols
met the Danish Regulations for Animal Research and all
experiments were carried out as investigator blinded studies.
2.2. Tissue extraction and radioimmunoassays (RIA)
Six female rabbits at the age of 22 weeks were anae-
sthetized (pentobarbital 100 mg/kg intravenously). The
abdomen and thorax were cut open and the entire heart
with aorta, the iliac arteries and the inferior caval vein were
removed. The head was separated from the body, the skull
was carefully opened with an oscillating saw and the entire
brain with the brainstem was removed. The tissue was
rapidly placed in ice-cold Krebs buffer (NaCl 119 mM,
KCl 4.6 mM, NaHCO3 15 mM, CaCl2 1.5 mM, MgCl2 1.2
mM, NaH2PO4 1.2 mM and glucose 11 mM) to reduce
metabolism and anoxia. All Krebs buffer solutions were
constantly bubbled with 5% CO2 + 95% O2 to maintain pH
at 7.4. Throughout the subsequent dissection, the tissue was
bathed in cold Krebs buffer at 4 jC. The cerebral arteries ofthe posterior part of the circle of Willis (the posterior
communicating arteries, the posterior cerebral arteries and
approximately 15 mm of the basilar artery) were isolated
from the ventral surface of the brain by careful micro-
dissection under a stereomicroscope and dissected free of
connective tissue. The intramural part of the left anterior
descending coronary artery (f 20 mm) was isolated and
freed of fat tissue, ventricular muscle and connective tissue.
A square of the myocardium (f 5� 5 mm, without visible
coronary arteries) was cut from the left ventricle near apex.
Furthermore, ring segments (f 20 mm in length) were cut
from the thoracic aorta, the caval vein and the proximal iliac
arteries (bilaterally, f 10 mm from each side). All samples
were immediately stored at � 80 jC. Before peptide ana-
lysis, the frozen tissue specimens were weighed and extrac-
ted in boiling water/acetic acid [22]. The extracted samples
were reconstituted and analyzed by radioimmunoassays
specific for PACAP-38 (antiserum code no. 733C) [22]
and VIP (antiserum code no. 5603) [23,24], respectively.
The radioimmunoassays were specific for the respective
peptides and did not show any cross-reactivity with structur-
ally related peptides. The within- and between-assay coef-
ficients of variation were below 10%. The concentrations
were expressed as pmol neuropeptide per gram wet weight
tissue (n = 6).
2.3. Immunohistochemistry
Four adult male rabbits were used for the immunhisto-
chemical analysis. On the day of tissue preparation, the
rabbits were killed by an overdose of pentobarbital intra-
cardially (500 mg/animal), which does not seem to affect
cardiovascular tissue. The brain and heart were removed,
and the major extracerebral arteries on the ventral surface of
the brain were identified using a stereomicroscope and
carefully dissected from the brain. The vessels and the heart
were placed in ice-cold Stefanini fixative for 16–24 h at 4
jC. The heart tissue was cryoprotected for 24–48 h in 30%
sucrose at 4 jC, then frozen at � 20 jC before it was cut in
12-Am-thick sections on a cryostat. The blood vessels were
placed in cryoprotectant at � 20 jC and reacted as whole
mount as described below. Double immunohistochemistry
was performed as previously described [25] using a well-
characterized mouse monoclonal PACAP antibody (code
no. JHH1, supernatant diluted 1:5) [22] and a guinea pig
anti-VIP antiserum (code no. B-GP340-1, obtained from
Euro-Diagnostica, Malmo, Sweden, diluted 1:500). PACAP
and VIP immunoreactivity were visualized using a biotiny-
lated donkey anti-mouse antiserum (code no. 115-065-151,
diluted 1:800, Jackson Immunoresearch Laboratories, West
Grove, PA, USA) and FITC-conjugated donkey anti-guinea
pig antiserum (code no. 706-045-148, diluted 1:50) in
combination with biotinylated tyramide (Tyramide System
amplification, DuPont NENR, Boston, MA, USA) and
streptavidin–Texas RedR. As controls, sections and/or
T. Dalsgaard et al. / Regulatory Peptides 110 (2003) 179–188180

whole mounts were routinely incubated with antibodies
preabsorbed with the respective antigen (20 Ag/ml), which
abolished all staining. Images were grabbed via a Leica
DC200 camera using Leica DC200 software (Leica, Cam-
bridge, UK). Image editing software (Adobe Photoshop/
Adobe Illustrator) was used to combine the obtained images
into plates, and figures were printed on a Tektronix (Wilson-
ville, OR) Phaser 450 dye sublimation printer.
2.4. Functional studies
2.4.1. Surgery and experimental design
Following 1 week of acclimatisation, at the age of 16
weeks, 32 female rabbits were bilaterally ovariectomised
during anaesthesia with 0.4 ml/kg body weight of a mixture
of Hypnorm (fentanyl citrate 0.315 mg/ml + fluanisone 10
mg/ml, Janssen-Cilag, Sauderton, High Wycombe, Buck-
inghamshire HP14 4HJ, UK), Dormicum (5 mg/ml, F.
Hoffman-La Roche, Basel, Switzerland) and sterile water
(1:1:2). After 2 weeks of recovery, from the age of 18
weeks, the rabbits were randomized to four different groups
of treatment (each group n = 8): placebo control, 17h-estra-diol (E2), Norethindrone Acetate (NETA) or E2 +NETA. To
avoid the influence of phytoestrogens, the rabbits were fed
100 g/animal/day of semisynthetic pelleted diet C2000
(Altromin International, D-4937 Lage, Germany) during
the recovery and treatment periods. Furthermore, the batch
of semisynthetic chow was tested for the content of the
major isoflavonoids commonly found in soy, using extrac-
tion with water/methanol/formic acid followed by analysis
with high-pressure liquid chromatography (HPLC) and
liquid chromatography/mass spectrometry (LCMS). No
traces of these phytoestrogens were found in the semi-
synthetic chow. During the 4-week treatment period, the
concentrations of E2 and NETA (both Novo Nordisk Far-
maka, Lyngby, Denmark) added to the chow were 0% and
0% for the placebo group, 0.004% and 0% for the E2 group,
0% and 0.003% for the NETA group and 0.004% and
0.003% for the E2 +NETA group, respectively. One rabbit
from the E2 +NETA group suffered from diarrhoea for the
last 2 weeks of the treatment period and displayed oedema
of the gut and the brain at dissection. Consequently, this
rabbit was excluded from the study.
2.4.2. Tissue preparation and myograph experiments
At the age of 22 weeks, two rabbits from different groups
of treatment were anaesthetized (pentobarbital 100 mg/kg
intravenously). The entire heart and brain were removed,
and the cerebral and coronary arteries were microdissected
as described above. Ring preparations (f 1 mm in length)
of three artery segments were prepared for the myograph
experiments: from the middle part of the posterior cerebral
artery (PCA), from the proximal epicardial part of the right
coronary artery as close to its origination from aorta as
possible (PROX) and from the intramural part of the left
anterior descending coronary artery (LAD), halfway
between aorta and apex of the heart. One LAD segment
from the placebo group and two PROX segments (from the
placebo and E2 group, respectively) could not be dissected.
Blood samples (nz 7 for all groups) were analyzed for E2
and estrone [26]. Two 40-Am wires were carefully inserted
through the lumen of each artery ring preparation for
mounting in double myographs [27] (Myo-Interface 500A,
JP Trading, Aarhus, Denmark), allowing measurement of
the isometric tension relations. All the experiments were
performed with two corresponding artery segments from
two differently treated rabbits, which were assigned ran-
domly and mounted on the near or far transducer in the
same double myograph, thus allowing the two arteries to be
tested and compared simultaneously and exposed to the
same solutions. The relation between resting wall tension
and internal circumference was determined, and from this,
the internal circumference, L100, corresponding to a trans-
mural pressure of 13.3 kPa ( = 100 mm Hg) for a relaxed
vessel in situ was calculated [28]. The vessels were set to the
internal circumference Ll = 0.9� L100. Previously, it has
Table 1
Characteristics of the rabbits used in this investigation
Treatment Placebo E2 NETA E2 +NETA ANOVA
Number 8 8 8 7a –
Body weight, start of treatment (kg) 3.3 (0.1) 3.3 (0.1) 3.2 (0.2) 3.3 (0.1) NS
Body weight, end of treatment (kg) 3.7 (0.1) 3.7 (0.1) 3.4 (0.2) 3.6 (0.1) NS
Avg. weight gain during treatment (g/week) 102 (6) 98 (15) 71 (13) 88 (16) NS
Avg. intake, E2 [mg (kg bw)� 1 day� 2] 0 (0) 1.1 (0)*,** 0 (0) 1.0 (0.1)*,** p< 0.001
Avg. intake, NETA [mg (kg bw)� 1 day� 2] 0 (0) 0 (0) 0.8 (0)*,*** 0.8 (0)*,*** p< 0.001
Serum–E2 at endpoint (pmol l� 1) 39 (0) 101 (10)*,** 39 (0) 106 (8)*,** p< 0.001
Serum–Estrone at endpoint (pmol l� 1) 53 (3) 123 (25)b 84 (11)c 122 (10)* p< 0.01
Values are mean (S.E.M.). E2 = 17h-estradiol, NETA=Norethindrone Acetate. Avg. = average, bw= body weight.a One rabbit from the E2 +NETA group was excluded because of diarrhoea and pathology at dissection.b Indicates borderline significance ( p= 0.14) compared to placebo (Student’s t-test [Bonferroni correction]).c Indicates borderline significance ( p= 0.10) compared to placebo (Student’s t-test [Bonferroni correction]).
* Indicates p< 0.001 compared to placebo.
** Indicates p< 0.001 compared to NETA.
***Indicates p< 0.001 compared to E2.
T. Dalsgaard et al. / Regulatory Peptides 110 (2003) 179–188 181

been shown that at Ll, the force production in vascular
smooth muscle is close to maximum, also in rabbit coronary
and cerebral arteries [29]. By changing the normal Krebs
buffer with 124 mM K+–Krebs buffer, which is normal
Krebs buffer with KCl exchanged for NaCl on an equimolar
basis, repeated contractions were induced until reproducible
responses were obtained, accepting a variation of V 10%. In
vessels precontracted with 30 mM K+–Krebs buffer (com-
position as normal Krebs buffer, except KCl, 30 mM, and
NaCl, 93.7 mM) and allowed to equilibrate for 20 min to
reach a plateau, the cumulative dose–response curves to
VIP and PACAP-27 (Peninsula Laboratories, St. Helens,
UK, both 10� 10–10� 6 M) were established (half-logarith-
mic steps, 3 min between each addition). With an estimated
intracellular [K+] of 150 mM, the change in extracellular
[K+] from 4.6 to 30 mM would change the membrane
potential (Vm) from approximately � 93 to approximately
� 43 mV, calculated using the Nernst equation, Vm =
RT(zF)� 1ln(Co/Ci), where R is the gas constant ( = 8.314 J
K� 1 mol� 1), T the absolute temperature (310 K= 37 jC in
the myograph bath), z the number of charges for the ion
(= + 1), F the Faraday constant ( = 9.65 104 A s mol� 1), Co
and Ci the molal [K+] outside and inside the cell membrane,
respectively. In pilot studies, we observed that the vaso-
dilating effect of PACAP-27 was more pronounced than for
PACAP-38. Earlier studies have indicated that concentration
and effect of neuropeptides are independent parameters; for
instance, it has been demonstrated that sex steroids can
influence the motor effect of VIP in smooth muscle without
changing the VIP concentration [30]. Therefore, it was
decided to use PACAP-27 for the functional studies, but
PACAP-38 for the RIA studies, since the tissue concen-
trations of PACAP-38 are considerably higher compared to
PACAP-27. Artery segments with a maximum force deve-
lopment of less than 1 mN/mm following depolarization
with 124 or 30 mM potassium were discarded. Following all
the mentioned criterions of exclusion, nz 6 for all groups
and for all types of vessels in all experiments.
2.5. Data, calculations and statistical analysis
Responses of the vessels were measured as force (F)
and expressed as active wall tension (T), which is F
divided by twice the segment length [28]. Internal lumen
diameter was calculated as l = L/p. The effective active
pressure (yPK+, max) corresponding to the maximum active
tension response to 124 mM potassium (TK+, max) was
calculated as yPK+, max = TK+, max/(Ll/2p). By computerized
iteration (GraphPad Prism Software, version 2.00, San
Diego, CA), the responses to the cumulative addition of
the peptides were fitted to sigmoidal dose–response curves
with variable Hill-slopes, used to determine the maximum
relaxant effect (Emax) and the concentration required to
produce half the maximum response (EC50), the latter
expressed as the negative logarithm to the molar concen-
tration, pI2 =� log(EC50). Results are expressed as mean
F S.E.M. Significance of difference between the treatment
groups was assessed by one-way ANOVA. If this test
indicated statistical significance, Student’s two-tailed t-test
was used to compare groups two by two, using the
Bonferroni correction for multiple comparisons. Statistical
significance was set at p < 0.05.
3. Results
3.1. Characteristics of the rabbits
Baseline values, body weight at arrival, before and after
the treatment period, average weight gain per week during
treatment and relative feed intake during the study demon-
strated comparability of the four groups of treatment (Table
1). The rabbits receiving active treatment with E2 and
E2 +NETA displayed E2 levels three times that of the placebo
and NETA group (Table 1). An average relative intake of 1.1
mg E2/kg body weight/day for the E2 group and of 1.0 mg E2/
kg body weight/day for the E2 +NETA group resulted in
physiological levels of E2 in the blood of these two groups.
The placebo and NETA group had no intake of E2, and
serum–E2 levels at euthanization for these two groups were
at the detection limit of 40 pM. Consequently, serum–E2
levels for both the E2 and E2 +NETA group were signifi-
cantly higher ( p < 0.001, Student’s t-test with Bonferroni
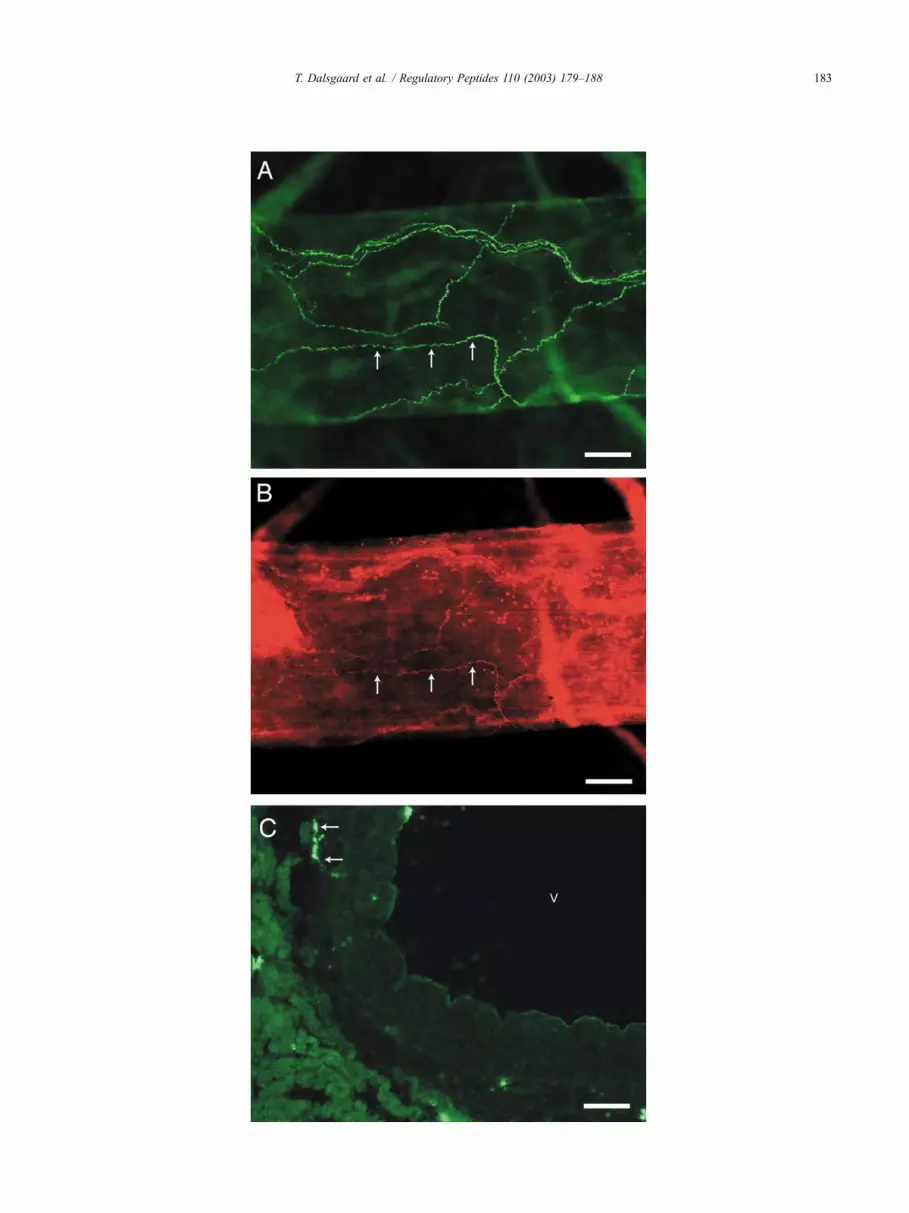
Fig. 2. Fluorescence photomicrographs of PACAP (A and C) and VIP immunoreactivity (B) in whole mount preparation of cerebral arteries isolated from the
basal surface of the rabbit brain (A and B) and in coronary arteries (C). Note that VIP is located in a subpopulation of PACAP containing nerve fibres
(exemplified by arrows in A and B). PACAP immunoreactive nerve fibres were found in coronary arteries located in the adventitia and media (arrows in C). V
indicates lumen of a coronary artery. Scale bars: (A) and (B) = 100 Am; (C) = 50 Am.
Fig. 1. Concentration of immunoreactive VIP (single hatched columns) and
PACAP-38 (double hatched columns) in rabbit cerebral arteries (CER), left
anterior descending coronary arteries (COR), myocardium (MYO), thoracic
aorta (AO), iliac arteries (IA) and caval vein (CV). Values (pmol/g wet
weight) are given as mean, horizontal bars indicate S.E.M. (n= 6).
T. Dalsgaard et al. / Regulatory Peptides 110 (2003) 179–188182
T. Dalsgaard et al. / Regulatory Peptides 110 (2003) 179–188 183

correction) compared to both the placebo and NETA group.
The average relative intake of NETA was 0.8 mg/kg body-
weight/day for both the E2 +NETA and NETA group. The
placebo and E2 group had no intake of NETA. No laboratory
kit was available for measuring the concentration of NETA,
but serum–estrone can be regarded as an indicator of the
NETA uptake. NETA and other progestogens induce an
increased activity of P450-aromatase [31] and 17h-hydrox-ysteroid dehydrogenase [32,33] in extra-ovarian tissue,
thereby increasing the conversion to estrone. Accordingly,
serum–estrone was significantly higher in the E2 +NETA
group compared to the placebo group ( p < 0.001, Student’s t-
test with Bonferroni correction). Estrone levels for the NETA
and E2 group were also higher compared to the placebo
group, thus confirming the intake of NETA from the chow
in the NETA group, but with a larger variation. Therefore,
the p-values were only borderline significant following
Bonferroni correction of the results from Student’s t-test
( p = 0.10 and 0.14, respectively).
3.2. Radioimmunoassays
The concentration of immunoreactive VIP and PACAP-
38 was significantly higher in coronary and cerebral
arteries compared to both myocardium, thoracic aorta, iliac
arteries and caval vein, where the concentrations were
negligible. Although the concentrations of VIP tended to
be lower than the concentrations of PACAP in the coronary
and cerebral arteries, they were not significantly different
(Fig. 1).
3.3. Immunohistochemistry
3.3.1. Basal arteries of the brain
Whole mount preparation of the basal arteries showed a
distinct pattern of PACAP immunostaining consisting of a
well-developed plexus of perivascular nerve fibres observed
Table 2
Functional characteristics for the vessels used for myograph investigations
in this study
Treatment Placebo E2 NETA E2 +NETA
PCA, ll (Am) 444 (9) 475 (30) 439 (21) 488 (26)
PCA, yTK+, max
(mN mm� 1)
2.5 (0.3) 3.2 (0.4) 2.8 (0.3) 2.8 (0.4)
PCA, yPK+, max
(mm Hg)
85 (10) 98 (12) 95 (11) 86 (10)
PCA, first
K(30/124)+ ratio (%)
113 (9) 95 (3) 92 (5) 104 (11)
PCA, second
K(30/124)+ ratio (%)
91 (11) 81 (6) 86 (7) 95 (7)
LAD, ll (Am) 790 (61) 744 (63) 596 (87) 848 (49)
LAD, yTK+, max
(mN mm� 1)
6.2 (1.2) 5.5 (0.7) 3.7 (0.9) 5.3 (0.6)
LAD, yPK+, max
(mm Hg)
121 (23) 114 (14) 87 (12) 96 (11)
LAD, first
K(30/124)+ ratio (%)
97 (6) 97 (9) 109 (9) 93 (9)
LAD, second
K(30/124)+ ratio (%)
100 (9) 101 (8) 121 (9) 96 (14)
PROX, ll (Am) 884 (64) 890 (78) 899 (102) 916 (33)
PROX, yTK+, max
(mN mm� 1)
4.1 (0.7) 5.3 (0.6) 4.2 (0.3) 4.2 (0.7)
PROX, yPK+, max
(mm Hg)
69 (9) 94 (6) 76 (8) 69 (11)
PROX, first
K(30/124)+ ratio (%)
94 (10) 96 (10) 93 (10) 107 (5)
PROX, second
K(30/124)+ ratio (%)
99 (9) 96 (12) 100 (12) 112 (7)
Values are mean (S.E.M.). PCA=posterior cerebral artery, LAD= left
anterior descending coronary artery, PROX= proximal right coronary
artery. ll = normalized effective lumen diameter; yTK+, max =maximum
active wall tension at ll for 124 mM K+ response; yPK+, max =maximum
effective active pressure at ll for 124 mM K+ response; K+(30/124)
ratio=(maximum active wall tension for 30 mM K+ response)/(yTK+, max)
ratio in percent, first ratio before the VIP experiment, second ratio before
the PACAP-27 experiment. ANOVA, all NS.
Fig. 3. Maximum vasodilatory effect (Emax, percentage of precontraction
induced by 30 mM potassium–Krebs buffer, top panel) and pI2[� log(EC50), lower panel] for VIP (panel to the left) and PACAP-27
(panel to the right) in rabbit posterior cerebral artery (PCA), left anterior
descending coronary artery (LAD) and proximal right coronary artery
(PROX). White columns = placebo group, single hatched columns = 17h-estradiol (E2) group, double hatched columns =Norethindrone Acetate
(NETA) group and black columns =E2 +NETA group. Values are mean
with S.E.M. shown as vertical bars. ANOVA, all NS (nz 6).
T. Dalsgaard et al. / Regulatory Peptides 110 (2003) 179–188184

in the wall of all major cerebral arteries and their branches
(Fig. 2A). VIP immunostaining was less intense compared
to the PACAP staining (Fig. 2B) and seemed to be located in
a subpopulation of the PACAP-immunoreactive nerve fibres
(Fig. 2A and B).
3.3.2. Coronary arteries
Few densely stained PACAP-immunoreactive nerve
fibres were found in coronary arteries located in the adven-
titia and media (Fig. 2C). VIP could not be demonstrated in
rabbit coronary arteries using immunohistochemistry with
the VIP guinea pig antiserum.
3.4. Functional studies
3.4.1. Functional characteristics of the vessels
There were no differences between the treatment groups,
for any of the vessels, in the number of steps at the
normalization procedure. The functional characteristics of
the vessels used in this investigation are summarized in
Table 2. For all vessels, activation by 124 mM potassium
induced an initial fast increase in tension, most pronounced
in the coronary arteries, which declined within 10 s to a
tonic contraction. The normalized effective lumen diameter
(ll) and the amplitude of the maximum potassium-induced
contraction (yTK+, max and yPK+, max) at ll displayed no
differences between the four groups of treatment. In all
vessels, activation by 30 mM potassium induced a slowly
developing contraction, which reached a plateau with a tonic
contraction in approximately 10–15 min. No differences
were observed between the treatment groups for any of the
vessels for the first or second 30 mM potassium response
(before the VIP and PACAP experiment, respectively) and
neither for the first and second K30/120+ ratio (Table 2).
Fig. 4. Concentration– response curves for relaxation induced by VIP
(graph A) and PACAP-27 (graph B), both 10� 10–10� 6 M (half-
logarithmic steps), in rabbit posterior cerebral arteries (closed, black
circles), distal left coronary arteries (closed, black triangles) and proximal
right coronary arteries (open circles) for pooled data (26V nV 30). Data are
expressed as percentage of the precontraction induced by Krebs buffer with
potassium, 30 mM, at the beginning of each experiment. Symbols indicate
mean with S.E.M. shown by vertical bars. *** indicates p< 0.0001 for Emax
(ANOVA).
Fig. 5. Maximum vasodilatory effect (Emax, percentage of precontraction
induced by 30 mM potassium–Krebs buffer, top panel), pI2 [� log(EC50),
middle panel] and Hill-slopes (lower panel) for VIP (panel to the left) and
PACAP-27 (panel to the right) in rabbit posterior cerebral artery (PCA), left
anterior descending coronary artery (LAD) and proximal right coronary
artery (PROX) for pooled data (26V nV 30). Values are mean with S.E.M.
shown as vertical bars. *** indicates p< 0.001, ** indicates p< 0.01 and *
indicates p< 0.05 (Student’s t-test [Bonferroni correction]).
T. Dalsgaard et al. / Regulatory Peptides 110 (2003) 179–188 185

3.4.2. Myograph dose–response experiments, VIP and
PACAP-27
Both VIP and PACAP-27 induced a dose-dependent
vasodilatory response in cerebral as well as coronary arteries.
The computerised iterations showed that treatment with sex
steroids induced no changes in Emax, pI2 or Hill-slopes in any
of the vessels for any of the two examined peptides (Fig. 3).
In general, there was a similar pattern for each treatment
group: that the VIP response was different for each vessel
type, with the smallest response in PROX, larger in LAD and
largest in PCA, while for PACAP-27, the response was
practically similar in all three artery segments. To further
examine these variations, and since treatment induced no
changes, the results for all four treatment groups were pooled
(Fig. 4). The differences between the vessel segments in
Emax, pI2 and Hill-slopes for the VIP response were all
statistically significant (ANOVA, p < 0.0001, p < 0.0001
and p < 0.05, respectively, Fig. 5). For the PACAP response,
on the contrary, Emax, pI2 and Hill-slopes were similar in all
three examined artery segments.
4. Discussion
The present study demonstrates that rabbit cerebral
arteries are supplied by VIP- and PACAP-containing nerve
fibres, and in contrast to previous findings in rat [34], the
two peptides were partly co-localized and VIP immuno-
staining was less intense compared to the PACAP staining.
Only PACAP-immunoreactive fibres were demonstrated in
the coronary arteries. Due to the species, we had to use a
guinea pig antiserum rather than our highly sensitive rabbit
anti-VIP antiserum, which could explain the inability to
detect VIP in the coronary arteries at the immunohistoche-
mical level. Both VIP and PACAP were able to produce a
vasodilatory response, but while the vasodilatory effect of
VIP differed from one vascular bed to another, PACAP
displayed identical vascular responses in all vessel seg-
ments. This observation is in accordance with a previous
study in atherosclerotic cerebral and coronary arteries from
Watanabe Heritable Hyperlipidemic (WHHL) rabbits (Dals-
gaard et al., unpublished data). The finding is unexpected,
since only three receptors for VIP and PACAP are known
today [6]: VPAC1 and VPAC2 that respond to both VIP and
PACAP and PAC1 that only responds to PACAP. We
observed that the maximum effect of VIP in PCA was
significantly higher compared to both the maximum VIP
response in the coronary arteries and to the maximum
PACAP response in PCA ( p < 0.001, Student’s t-test). At
the same time, there was a trend towards a decreased
maximum response to VIP in PROX compared to LAD.
This was supported by a significantly smaller maximum
response to VIP compared to PACAP in PROX ( p < 0.05,
Student’s t-test). Therefore, our results indicate that VIP
induces different mechanisms of action in the three exa-
mined artery segments. In PCA, VIP may either act through
a new unknown receptor or directly on a different mecha-
nism in the cell membrane, for instance, the potassium
channels. In porcine coronary arteries, it has been demon-
strated that VIP induces relaxation via three mechanisms: a
decrease in [Ca2 +]i by inhibiting the Ca2 + influx (presu-
mably through membrane hyperpolarization mediated by
activation of K+ channels); a decrease in [Ca2 +]i by seques-
tering cytosolic Ca2 + into store sites and a decrease in the
Ca2 + sensitivity of the contractile apparatus through the
activation of G-protein [35]. In an earlier study examining
the vasodilatory effect of VIP on coronary vessels in the
isolated perfused rabbit heart, the slope of the dose–
response curve of VIP suggested that the effect was induced
through one receptor subtype [12]. This is not supported by
the present study, where the Hill-slopes of both proximal
and distal coronary arteries suggest the presence of more
than one receptor subtype.
Alternatively, the regional differences in vascular re-
sponses to the two peptides may be explained by the
interaction of various substances produced and present in
the tissue, or that the vessels display different populations of
the receptor subtypes, some of them being able to counteract
the others or to cluster with each other and G-proteins, and
thereby induce configurational changes. These hypotheses
would explain the observed significant differences for the
Hill coefficients. Earlier, it has been demonstrated in guinea
pig cerebral arteries that the vasoactive mechanism for
neuropeptides may be complex. For instance, NPY, which
is co-localized not only with norepinephrine in sympathetic
perivascular fibres but also with VIP in some parasympa-
thetic neurons, can greatly reduce the vasodilatory effect of
VIP [36]. Recently, it has been demonstrated in rat cerebral
vessels that VPAC1 immunoreactivity is localized to the
plasmalemma of smooth muscle cells on superficial cerebral
arteries and arterioles from the basal surface of the brain
[34]. VIP- and PACAP-containing nerve fibres were found
in separate nerve populations, with different distribution
pattern and density, but both with an intimate contact to the
receptor protein, PACAP to a lesser extent than VIP. In the
present study in rabbits, the number of PACAP-immuno-
reactive nerve fibres in the basal cerebral arteries was higher
than those containing VIP, and VIP seemed to present in a
subpopulation of the PACAP-immunoreactive fibres.
The vasodilatory effect of PACAP in rabbit cerebral
arteries is demonstrated in this study for the first time, but
has previously been reported in other species [37–39].
Earlier studies have demonstrated regional differences in
the vasodilating effect of VIP and PACAP. In canine
cerebral vessels, the maximum vasodilating effect of
PACAP-27 has been demonstrated to be higher, the more
rostrally the vessel is situated [37], with the most potent
vasodilating effect of PACAP-27 in the anterior cerebral
arteries. The vasodilatory potency of VIP and PACAP is
also different in isolated human coronary arteries [14].
Interestingly, it has earlier been demonstrated that the
relative frequency of immunohistochemically visible VIP
T. Dalsgaard et al. / Regulatory Peptides 110 (2003) 179–188186

nerve fibres in cerebral arteries from rabbits was lower than
in several other species, and that the concentration of VIP in
the same arteries was lowest of the examined species,
including cow, pig, cat, guinea pig, rat and mouse [40].
This is the first study examining the effect of clinically
used HRT regimens on the function of non-atherosclerotic
coronary and cerebral arteries, at the same time excluding
the influence of phytoestrogens. Earlier, we have observed
that feeding with a standard plant-component-based diet is
able to change the effect of VIP on atherosclerotic cerebral
arteries (Dalsgaard et al., unpublished data). In the present
study, pretreatment with sex steroids, leading to physiolo-
gical plasma levels of estradiol, and plasma levels of estrone
that confirmed the uptake of NETA, did not influence the
peptide-induced vasodilation. Similar treatment regimens in
female rabbits have previously been reported to induce
significant changes in the functional and mechanical cha-
racteristics of cerebral and coronary arteries [29]. In non-
vascular uterine rabbit smooth muscle, treatment with estra-
diol and progesterone for 4 weeks was able to induce
significant changes in the receptor expression and effect of
VIP [30]. Thus, the cardioprotective effects produced by
estradiol in non-atherosclerotic arteries seem not to involve
the actions of VIP/PACAP directly.
Acknowledgements
This study was supported by grants from the Danish
Medical Research Council (SSVF), the Novo Nordisk
Foundation, the Danish Heart Foundation and the Research
Council of the Copenhagen Hospital Corporation. The
authors also wish to thank Novo Nordisk Farmaka, Lyngby,
Denmark, for supplying 17h-estradiol and Norethindrone
Acetate for the study.
References
[1] Fahrenkrug J. Transmitter role of vasoactive intestinal peptide. Phar-
macol Toxicol 1993;72:354–63.
[2] Ottesen B, Fahrenkrug J. Vasoactive intestinal polypeptide and other
preprovasoactive intestinal polypeptide-derived peptides in the female
and male genital tract: localization, biosynthesis, and functional and
clinical significance. Am J Obstet Gynecol 1995;172:1615–31.
[3] Vaudry D, Gonzalez BJ, Basille M, Yon L, Fournier A, Vaudry H.
Pituitary adenylate cyclase-activating polypeptide and its receptors:
from structure to functions. Pharmacol Rev 2000;52:269–324.
[4] Arimura A. Pituitary adenylate cyclase activating polypeptide (PA-
CAP): discovery and current status of research. Regul Pept 1992;37:
287–303.
[5] Kimura C, Ohkubo S, Ogi K, Hosoya M, Itoh Y, Onda H, et al. A
novel peptide which stimulates adenylate cyclase: molecular cloning
and characterization of the ovine and human cDNAs. Biochem Bio-
phys Res Commun 1990;166:81–9.
[6] Harmar AJ, Arimura A, Gozes I, Journot L, Laburthe M, Pisegna JR,
et al. International Union of Pharmacology: XVIII. Nomenclature of
receptors for vasoactive intestinal peptide and pituitary adenylate cy-
clase-activating polypeptide. Pharmacol Rev 1998;50:265–70.
[7] Uddman R, Edvinsson L. Neuropeptides in the cerebral circulation.
Cerebrovasc Brain Metab Rev 1989;1:230–52.
[8] White RP. Responses of human basilar arteries to vasoactive intestinal
polypeptide. Life Sci 1987;41:1155–63.
[9] Saetrum OO, Edvinsson L. Effect of parasympathetic and sensory
transmitters on human epicardial coronary arteries and veins. Phar-
macol Toxicol 1996;78:273–9.
[10] Gulbenkian S, Saetrum OO, Ekman R, Costa AN, Wharton J, Polak
JM, et al. Peptidergic innervation of human epicardial coronary ar-
teries. Circ Res 1993;73:579–88.
[11] Tsukahara T, Hongo K, Kassell NF, Ogawa H. The influence of ex-
perimental subarachnoid hemorrhage on the relaxation induced by
vasoactive intestinal polypeptide in the cerebral arteries of the rabbit.
Neurosurgery 1989;24:731–5.
[12] Accili EA, Buchan AM, Kwok YN, Ledsome JR, Brown JC. Presence
and actions of vasoactive intestinal peptide in the isolated rabbit heart.
Can J Physiol Pharm 1995;73:134–9.
[13] Bruch L, Bychkov R, Kastner A, Bulow T, Ried C, Gollasch M, et al.
Pituitary adenylate-cyclase-activating peptides relax human coronary
arteries by activating K(ATP) and K(Ca) channels in smooth muscle
cells. J Vasc Res 1997;34:11–8.
[14] Kastner A, Bruch L, Will-Shahab L, Modersohn D, Baumann G.
Pituitary adenylate cyclase activating peptides are endothelium-inde-
pendent dilators of human and porcine coronary arteries. Agents Ac-
tions Suppl 1995;45:283–9.
[15] Dorner GT, Wolzt M, Eichler HG, Schmetterer L. Effect of pituitary
adenylate cyclase activating polypeptide 1–27 on ocular, cerebral and
skin blood flow in humans. Naunyn-Schmiedeberg’s Arch Pharmacol
1998;358:657–62.
[16] Skakkebaek ML, Georg B, Mikkelsen JD, Ottesen B, Fahrenkrug J.
All prepro-VIP-derived peptides, except PHI/PHV, are expressed in
the female rat anterior pituitary and increased by estrogen. Peptides
1995;16:1287–94.
[17] Palle C, Bredkjaer HE, Fahrenkrug J, Ottesen B. Vasoactive intestinal
polypeptide loses its ability to increase vaginal blood flow after men-
opause. Am J Obstet Gynecol 1991;164:556–8.
[18] Corr LA, Poole-Wilson P, Burnstock G. Responses of coronary ar-
teries to neurotransmitters: changes with sexual maturity in the female
rabbit. J Cardiovasc Pharmacol 1991;18:144–50.
[19] Henning RJ, Sawmiller DR. Vasoactive intestinal peptide: cardiovas-
cular effects. Cardiovasc Res 2001;49:27–37.
[20] Wild RA. Estrogen: effects on the cardiovascular tree. Obstet Gynecol
1996;87:27S–35S.
[21] Dalsgaard T, Larsen CR, Mortensen A, Larsen JJ, Ottesen B. New
animal model for the study of postmenopausal coronary and cerebral
artery function: the Watanabe heritable hyperlipidemic rabbit fed on a
diet avoiding phytoestrogens. Climacteric 2002;5:178–89.
[22] Hannibal J, Mikkelsen JD, Clausen H, Holst JJ, Wulff BS, Fahrenkrug
J. Gene expression of pituitary adenylate cyclase activating polypep-
tide (PACAP) in the rat hypothalamus. Regul Pept 1995;55:133–48.
[23] Fahrenkrug J, Schaffalitzky de Muckadell OB. Radioimmunoassay of
vasoactive intestinal polypeptide (VIP) in plasma. J Lab Clin Med
1977;89:1379–88.
[24] Fahrenkrug J, Schaffalitzky de Muckadell OB. Distribution of vaso-
active intestinal polypeptide (VIP) in the porcine central nervous
system. J Neurochem 1978;31:1445–51.
[25] Fahrenkrug J, Hannibal J. Pituitary adenylate cyclase activating poly-
peptide innervation of the rat female reproductive tract and the asso-
ciated paracervical ganglia: effect of capsaicin. Neuroscience 1996;73:
1049–60.
[26] Emment Y, Collins WP, Sommerville IF. Radioimmunoassay of oes-
trone and oestradiol in human plasma. Acta Endocrinol (Copenh)
1972;69:567–82.
[27] Mulvany MJ, Nyborg N. An increased calcium sensitivity of mesen-
teric resistance vessels in young and adult spontaneously hypertensive
rats. Br J Pharmacol 1980;71:585–96.
[28] Mulvany MJ, Halpern W. Contractile properties of small arterial re-
T. Dalsgaard et al. / Regulatory Peptides 110 (2003) 179–188 187

sistance vessels in spontaneously hypertensive and normotensive rats.
Circ Res 1977;41:19–26.
[29] Hansen VB, Skajaa K, Aalkjaer C, Oxlund H, Glavind EB, Petersen
OB, et al. The effect of oophorectomy on mechanical properties of
rabbit cerebral and coronary isolated small arteries. Am J Obstet
Gynecol 1996;175:1272–80.
[30] Ottesen B, Larsen JJ, Staun-Olsen P, Gammeltoft S, Fahrenkrug J.
Influence of pregnancy and sex steroids on concentration, motor ef-
fect and receptor binding of VIP in the rabbit female genital tract.
Regul Pept 1985;11:83–92.
[31] Tseng JK, Sun B, Tseng L. The effect of progestin on rabbit endo-
metrial aromatase activity. J Steroid Biochem 1988;29:9–13.
[32] Murugesan K, Vij U, Lal B, Farooq A. Effect of progestins, estradiol,
and coenzymes NAD and NADPH on the interconversion of estradiol
and estrone in rabbit uterus in vitro. Steroids 1989;53:695–712.
[33] Coldham NG, James VH. A possible mechanism for increased breast
cell proliferation by progestins through increased reductive 17 beta-
hydroxysteroid dehydrogenase activity. Int J Cancer 1990;45:174–8.
[34] Fahrenkrug J, Hannibal J, Tams J, Georg B. Immunohistochemical
localization of the VIP1 receptor (VPAC1R) in rat cerebral blood
vessels: relation to PACAP and VIP containing nerves. J Cereb Blood
Flow Metab 2000;20:1205–14.
[35] Kawasaki J, Kobayashi S, Miyagi Y, Nishimura J, Fujishima M,
Kanaide H. The mechanisms of the relaxation induced by vasoactive
intestinal peptide in the porcine coronary artery. Br J Pharmacol
1997;121:977–85.
[36] Edvinsson L, Adamsson M. Neuropeptide Y inhibits relaxation of
guinea pig cerebral, coronary, and uterine arteries: blockade by D-
myo-inositol-1,2,6-triphosphate. J Cardiovasc Pharmacol 1992;20:
466–72.
[37] Anzai M, Suzuki Y, Takayasu M, Kajita Y, Mori Y, Seki Y, et al.
Vasorelaxant effect of PACAP-27 on canine cerebral arteries and rat
intracerebral arterioles. Eur J Pharmacol 1995;285:173–9.
[38] Tong S, Parfenova H, Shibata M, Zuckerman S, Armstead WM, Lef-
fler CW. Pituitary adenylate cyclase-activating polypeptide dilates cer-
ebral arterioles of newborn pigs. Proc Soc Exp Biol Med 1993;203:
343–7.
[39] Jansen-Olesen I, Goadsby PJ, Uddman R, Edvinsson L. Vasoactive
intestinal peptide (VIP) like peptides in the cerebral circulation of the
cat. J Auton Nerv Syst 1994;49:S97–S103 [Suppl].
[40] Edvinsson L, Fahrenkrug J, Hanko J, McCulloch J, Owman C, Udd-
man R. Vasoactive intestinal polypeptide: distribution and effects on
cerebral blood flow and metabolism. In: Cervos-Navarro J, Fritschka
E, editors. Cerebral microcirculation and metabolism, vol. 49. New
York: Raven Press; 1981. p. 147–155.
T. Dalsgaard et al. / Regulatory Peptides 110 (2003) 179–188188