vessel generator noise as a settlement cue for marine biofouling species
TRANSCRIPT

� � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �� � ! " # " ! $ % � � � � $ & � ' () � � � � � � � � � � � � � * � � � � � �+ � � � � � � � � � , � � � � � � - , � � � � � � � � � � , � � � � � . � � � � $ ! & " ( / % � , � � � � � � � � � � � � � � � � � � 0 � � �' & 1 % $ � � � � � � � 2 � � � � � � � � � $ � ' # 0 � 3 4
5 6 7 8 9 : ; < = > ? @ A B CBiofouling: The Journal of Bioadhesion and Biofilm
ResearchPublication details, including instructions for authors and subscription information:
http://www.tandfonline.com/loi/gbif20
Vessel generator noise as a settlement cue for marine
biofouling speciesJ.I. McDonald
a, S.L. Wilkens
b, J.A. Stanley
c & A.G. Jeffs
c
a Western Australian Department of Fisheries, WA Fisheries and Marine Research
Laboratories, Perth, Australiab National Institute of Water and Atmospheric Research, Marine Biodiversity and Biosecurity,
Kilbirnie, Wellington, New Zealandc University of Auckland, Auckland, New Zealand
Published online: 28 May 2014.
To cite this article: J.I. McDonald, S.L. Wilkens, J.A. Stanley & A.G. Jeffs (2014): Vessel generator noise as a
settlement cue for marine biofouling species, Biofouling: The Journal of Bioadhesion and Biofilm Research, DOI:
10.1080/08927014.2014.919630
To link to this article: http://dx.doi.org/10.1080/08927014.2014.919630) � - � 2 - 2 D � � � � � � � . � � � � � � + D � -� � � � � � * � � � � � � � � E � F � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � G � � H D � � � I J � � � � � �� � � � � � � � � � � � � � � � � � � � � � � � K 0 � F � � � � � � � � * � � � � � � � � � � , � � � � � � � � � � � � � � � � E �� � � � � � � � � � � � � � � � � � � � � � � � F � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � D � � � K � � � � � � � � � � F � � L � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � F � � � � � � � � � � � � � � �� � � � � � F � � � � � � � � � � � � � � � � � � � * � � � � � � K � � � � � � � � � � � � � D � � � � � � � � � � � � � � � � � � �� � � � � � � � � � � � � F � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � K � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �� � � � � � � � � � � � � � � � � � � � � � � � � � � � , � � � � � � � � � � � � � � L � � � � � � � � , � � � � � � � � � � � � � � � � � � � � � � � F �� � � � � � F � � � � � � � � � � , � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � , � � � � � � � � �� � D � � � K� � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � , � � � � � � F � � � � � � � � � � � � K � � � � � � � � � � � � � � � � � � � � � �� � � � � � � � � � � � � � � � � � � � � � � � � � � , � � � � � � � 1 � � � � � , � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �� � � � � � � � � � � L � � � � � � � � � � � � � K � � � � * D � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � M M K � � � � � � � K � � � M � � , M � � � � 1 � � 1 � � � � � � � �

Vessel generator noise as a settlement cue for marine biofouling species
J.I. McDonalda, S.L. Wilkensb*, J.A. Stanleyc and A.G. Jeffsc
aWestern Australian Department of Fisheries, WA Fisheries and Marine Research Laboratories, Perth, Australia;bNational Institute of Water and Atmospheric Research, Marine Biodiversity and Biosecurity, Kilbirnie, Wellington, New Zealand;cUniversity of Auckland, Auckland, New Zealand
(Received 17 December 2013; accepted 25 April 2014)
Underwater noise is increasing globally, largely due to increased vessel numbers and international ocean trade. Vesselsare also a major vector for translocation of non-indigenous marine species which can have serious implications for biose-curity. The possibility that underwater noise from fishing vessels may promote settlement of biofouling on hulls wasinvestigated for the ascidian Ciona intestinalis. Spatial differences in biofouling appear to be correlated with spatial dif-ferences in the intensity and frequency of the noise emitted by the vessel’s generator. This correlation was confirmed inlaboratory experiments where C. intestinalis larvae showed significantly faster settlement and metamorphosis whenexposed to the underwater noise produced by the vessel generator. Larval survival rates were also significantly higher intreatments exposed to vessel generator noise. Enhanced settlement attributable to vessel generator noise may indicate thatvessels not only provide a suitable fouling substratum, but vessels running generators may be attracting larvae andenhancing their survival and growth.
Keywords: underwater sound; ascidian; larval settlement; metamorphosis; vessel noise; biofouling
Introduction
Ambient underwater sound has long been recognised as
one of the many cues used by marine organisms for ori-
entation, settlement and/or metamorphosis (Montgomery
et al. 2006; Stanley et al. 2012). Given this, it seems
equally likely that anthropogenic generated noise from
vessels in port environments may also have an influence
on the settlement of biofouling species, over and above
any natural background ambient sound. However, little
research has been conducted to verify if this is the case.
Anthropogenic noise generated by vessels typically falls
within the 6 to ~ 30,000 Hz frequency range, with the
highest intensities usually between 10 and 1,000 Hz. This
frequency range has been shown to be important for the
attraction, settlement and metamorphosis of many marine
invertebrate larvae (Montgomery et al. 2006; Götz et al.
2009; Jeffs et al. 2011). Several experimental studies
have shown that ambient underwater sound emanating
from coastal habitats can act as long-distance orientation
cues for settlement-stage crustacea (Jeffs et al. 2003,
2005; Radford et al. 2007; Stanley et al. 2010) and fish
(Tolimieri et al. 2000; Montgomery et al. 2006; Simpson
et al. 2008) attempting to locate suitable habitats. It is
also known that the planktonic larvae of many benthic
invertebrates can settle as soon as they find an appropri-
ate substratum or when conditions are favourable for col-
onisation (Glasby & Connell 1999). This facilitation may
be influenced by underwater sound.
The underwater noise produced by marine vessels
comes primarily from mechanical vibrations produced
by the engines, power transmission units and genera-
tors, from the hull interacting with the water whilst
underway and from cavitation on the rotating propeller
blades. Over the past decades ship traffic has increased,
and noise emissions account for > 90% of the acoustic
energy humans emit into the sea (Green et al. 1994).
Recent research has implicated underwater noise from
the generators of vessels as a possible cause of
increased settlement of fouling organisms through trig-
gering natural larval settlement cues (Wilkens et al.
2012). For example, settlement of larvae of the mussel,
Perna canaliculus, was significantly faster when
exposed to the underwater noise produced by a 125-m
long steel-hulled ferry (Wilkens et al. 2012). The
researchers suggested that reducing the noise emitted by
a vessel whilst at berth may help to reduce biofouling
on its hull.
Domestic fishing vessels were used as a model vector
in this study as they spend significant periods of time in
port running on generator power, and may be one of the
vectors in the spread of invasive species between ports.
Although best-practice management biofouling guidelines
are being developed for vessels in both New Zealand
and Australia, the risk of domestic spread of pest species
is still high. As vessel traffic is higher in human-
transport hubs, marine areas such as ports are regarded
*Corresponding author. Email: [email protected]
© 2014 Taylor & Francis
Biofouling, 2014
http://dx.doi.org/10.1080/08927014.2014.919630
N OPQR OST UTV WXN UYSZ[ \UQ[]̂_̀ ab UZ` Ua cde_fghi cXj ka[` Q\lT OQSRTi S[mn opqrst kQUsrmu

as important foci for the spread of non-indigenous
species (Floerl & Inglis 2005).
The biosecurity risks from hull fouling arise when
competent pest organisms are released into a suitable
recipient region in the form of adult life stages or
planktonic propagules and encounter a suitable settle-
ment habitat, such as a vessel hull (Floerl et al. 2004).
Due to the negative impacts of introduced marine pest
species into new non-native locations there are extensive
efforts to reduce the translocation of marine pest species
around the world (Wonham et al. 2000; Floerl et al.
2004). Vessel hull fouling may be responsible for at least
75% of the ship-mediated non-indigenous species in Port
Philip Bay, Australia (Hewitt 2004). A sampling of biota
scraped from hulls of commercial vessels in Germany
showed the presence of non-indigenous species on 96%
of the 131 ships examined (Gollasch 2002). A survey of
eight vessels in Hawaii found non-indigenous species on
the hulls of the majority of vessels, despite low levels of
overall biofouling (Godwin 2003). A single inspection of
a relatively clean passenger vessel in Australia found
several non-indigenous species in protected (niche) areas
of the ship, including the European green crab, Carcinus
maenus (Coutts et al. 2003). The fouling present on
vessels often reflects the fouling biota of the port/marina
where they spend most time, therefore movement of
fouled vessels from one location to another presents a
species translocation risk. Understanding biological and
physical variables that may influence the settlement of
hull-fouling species may be one tool in the biosecurity
management toolbox. Therefore the aim of this study
was to investigate whether spatial differences in noise
emission existed on a vessel, and what the influence
might be on the settlement of fouling species and the
overall levels of fouling present on a vessel. Evidence
that larvae respond to vessel noise was published by
Wilkens et al. (2012). This study only documented one
species (Perna canaliculus) in a controlled laboratory
environment. The present research aimed to demonstrate
that this trend is occurring in another common biofouling
species, such as ascidians (Lambert 2001; see also the
review on ascidian fouling by Aldred & Clare 2014) and
highlights spatial differences in noise intensity and
observed levels of fouling on vessels.
Materials and methods
It was not logistically possible to ‘tie up’ a number of
commercial fishing vessels to experimentally test the
effects of noise intensity on biofouling. Therefore this
study both examined in situ levels of hull fouling on
four fishing vessels (from four locations on each vessel)
and performed laboratory experiments to determine the
settlement response of a common fouling ascidian spe-
cies (Ciona intestinalis) to the different noise intensities
sampled at different locations around a vessel hull. This
approach provides a robust approach to assess the effects
of variable intensity of vessel noise on biofouling species
in a marina environment, and confirms the results by
testing in a controlled laboratory environment.
Vessel noise recording and processing
Vessel generator noises were recorded from a 25-m long
steel-hulled fishing vessel berthed in the Port of Freman-
tle, Western Australia, during February 2012. Only one
vessel was used for the recordings as the generators were
identical in the four vessels used in the fouling observa-
tions. Based on the location of the generator in this fish-
ing vessel design (Figure 1), four spatially separated hull
locations were selected for noise recordings. These were
(1) adjacent to the generator, (2) the stern, (3) the bow,
and (4) opposite the generator. At the time of recording
the vessel was operating on a ship-based generator
power supply and no other machinery was operational
during the recordings. No other vessels were operating
within the vicinity of this vessel when recordings were
taking place.
A calibrated hydrophone (HTI-96-Min, High Tech,
Inc., Long Beach, MS, USA) was used to continuously
record 5 min of underwater noise emitted by the vessel
generator at four locations around the hull (Figure 1).
The hydrophone was placed ~50 cm from the hull at
each of the four locations and lowered 2 m into the
water. During the recording phase the output was cap-
tured on a calibrated digital recorder (Edirol R09HR,
Roland Corporation, Japan).
Digital recordings were downloaded and spectral plots
were generated using the methods described in Wilkens
et al. (2012). An ANOVA was then performed to deter-
mine if there was a significant difference (p < 0.001) in
noise intensity among the four locations. The sub-samples
were then band-pass filtered into four frequency bins:
30–100, 101–500, 501–2,000, and 2,001–20,000 Hz, and
the overall mean proportion of total noise intensity was
calculated for each frequency bin. For each hull location
the proportion of total noise intensity was arcsine
Figure 1. Locations of underwater noise recordings from a25 m fishing vessel. ! denotes the location of the generator onthe vessel.
2 J.I. McDonald et al.
N OPQR OST UTV WXN UYSZ[ \UQ[]̂_̀ ab UZ` Ua cde_fghi cXj ka[` Q\lT OQSRTi S[mn opqrst kQUsrmu

transformed and analysed using a two-way ANOVA, with
Location and Frequency Bin as factors. Significant differ-
ences between proportions of total noise intensity were
determined using the Holm–Sidak test once the ANOVA
had determined an overall significant difference among
proportions (p < 0.001).
Two-minute sequences of the recordings were
transferred onto an MP3 player (A8695, Dick Smith
Electronics, Chullora, NSW, Australia) for playback
during the ascidian larval settlement experiment.
In situ observations of level of fouling
Four 25-m fishing vessels of comparable hull design and
antifouling treatment regime were berthed together at the
time of this study. The location of the vessels, and their
generator types, were identical. The level of biofouling
present on each of the four vessel hulls was estimated
using in situ diver observations and from examination of
underwater video (Snake-Eye IIITM, Aqua Communica-
tions Inc., Waltham, MA, USA). Observations were made
at the same four locations as used for the noise recordings
(ie adjacent to the generator, the stern, the bow and oppo-
site the generator). All visual estimates of hull fouling
present on all four vessels were made by both the
co-authors McDonald and Wilkens independently using
the level of fouling scale developed by Floerl et al.
(2005). This estimate is an ordinal rank scale of the rela-
tive abundance (approximate percentage cover) of fouling
assemblage present on the hull. Both observers visually
assessed a 2 m2 area of the vertical side of each vessel at
each of the locations used for noise recordings, from the
waterline to the top of the bilge keel.
Source of ascidian larvae
C. intestinalis, like many other solitary ascidian species,
is a prolific biofouler of vessel hulls and marine infra-
structures around the world (see Fitridge et al. 2012;
Aldred & Clare 2014). Adult specimens were collected
from Lyttelton Harbour, New Zealand in January 2012
and transported to the laboratory. Individual specimens
were dissected in the laboratory. The eggs and sperm
were removed using glass Pasteur pipettes. The repro-
ductive status could be assessed visually prior to dissec-
tion to ensure only sexually mature individuals were
used (protocol adapted from Cirino et al. 2002). Differ-
ent donor specimens were used for cross fertilisation.
The eggs were placed into a Petri dish containing 25 ml
of sterile seawater and ~ 300 μl of concentrated sperm
and gently agitated to ensure mixing of gametes. The
dilution of 300 μl of sperm into 25 ml of seawater pre-
vented an excess of sperm sticking to the eggs, poten-
tially compromising insemination. One hour after
insemination, the seawater was changed to remove
surplus sperm and the Petri dishes placed at 18–20°C for
15–18 h to allow embryo development. Immediately
prior to hatching (which was confirmed using light
microscopy), embryos were randomly selected and trans-
ferred into a sterile, flat bottomed 12-well tissue culture
plate (Sarstedt, Ingle Farm, Australia). Each well con-
tained 10 ml of sterile seawater at 18°C and an individ-
ual C. intestinalis larva.
Larval settlement experiment
The larval settlement experiment consisted of five treat-
ments. The first four treatments were exposed to noise
recordings from the four different locations on the vessel:
Location 1 (adjacent to generator, starboard side), Loca-
tion 2 (opposite generator, portside), Location 3 (stern),
and Location 4 (bow). The fifth treatment was a control
(ie no vessel noise) (Figure 1). For each treatment, three
replicate water baths were used to maintain a constant
water temperature at 18°C (± 1°C) throughout the experi-
ment. Each water bath contained a single 12-well tissue
culture plate which was visually and acoustically transpar-
ent. The water baths were covered with shade cloth, pro-
viding a constant low light level, thereby reducing
interference from external light cues. Foam rubber mats
were placed under the water baths to prevent any transfer
of acoustic energy from the surrounding environment into
the experimental treatments. Prior to the commencement
of the experiment, the absence of acoustic interference in
the treatment baths was confirmed by recording from each
of the replicates within each treatment using a calibrated
hydrophone (HTI-96-MIN).
Noise in each water bath was emitted from a speaker
(SBA1500, 4Ω, 100–18,000 Hz, Koninklijke Philips
Electronics, Noord-Brabant, The Netherlands) which was
sealed within a waterproof plastic bag, placed in the bot-
tom of the water bath, and held down by a lead weight.
The speakers were connected to a MP3 player which
continuously replayed a 2 min sequence of the vessel
noise recording from the corresponding location on the
vessel. Three different 2 min sequences from each loca-
tion were used to avoid pseudo-replication by using the
same vessel recording for each replicate within the treat-
ment (Kroodsma et al. 2001). The control treatment con-
tained an underwater speaker, weighted to the bottom,
but not connected to an MP3 player. For the treatments
the experimental vessel noise replayed in the water baths
was confirmed at 140.6 dB for Location 1 (adjacent to
generator, starboard side), and at 138.8 dB for Location
2 (opposite generator, portside), 135.2 dB for Location 3
(stern), and at 127.5 dB for Location 4 (bow). Examina-
tion of the recordings of the replayed generator noise
were analysed and verified to have a similar spectral
composition to the original recording of the noise from
the vessel in port.
Biofouling 3
N OPQR OST UTV WXN UYSZ[ \UQ[]̂_̀ ab UZ` Ua cde_fghi cXj ka[` Q\lT OQSRTi S[mn opqrst kQUsrmu
At two-hourly intervals, each tissue culture plate from
each treatment was removed from the water bath and
examined under a binocular microscope (× 40) to observe
the development of each larva. Larvae were classified
according to their progressive stages in the settlement
process, as described in Cirino et al. (2002): (1) swim-
ming; (2) immobile – larvae motionless when stimulated
by gentle suction from the tip of a 200 μm pipette, larvae
still coloured/opaque and body still intact; (3) attached –
larvae attached to the surface of the well or the meniscus
of the water by head, larvae remains attached when
gently stimulated by water movement; (4) metamorphic
stage 1 (M1) – tail at right angles to head, tail beginning
to turn transparent and starting to reabsorb, head darken-
ing/starting to turn pink, firmly attached to surface of well
or meniscus; (5) metamorphic stage 2 (M2) – tail reab-
sorption complete, pink coloration in head, larvae lobed
shaped, stalk starting to appear; or (6) dead – larvae
transparent or emaciated, head and tail starting to frag-
ment and shrink, no movement. The experiment was ter-
minated when all experimental larva had either attached
and/or metamorphosed or were dead. Data were exam-
ined to determine if there were any significant differences
between replicates within treatments. No significant dif-
ferences were found within treatments (controls or noise
treatments), therefore all replicates within a treatment
were pooled for ease of analysis and presentation of
results. Differences in larval, settlement, metamorphosis
and survival were tested using χ2 analyses.
Results
Vessel noise
The average noise intensity recorded from each location
around the vessel was measured at 140.6 dB re 1 μPa
RMS at Location 1, 138.8 at Location 2, 135.2 at Loca-
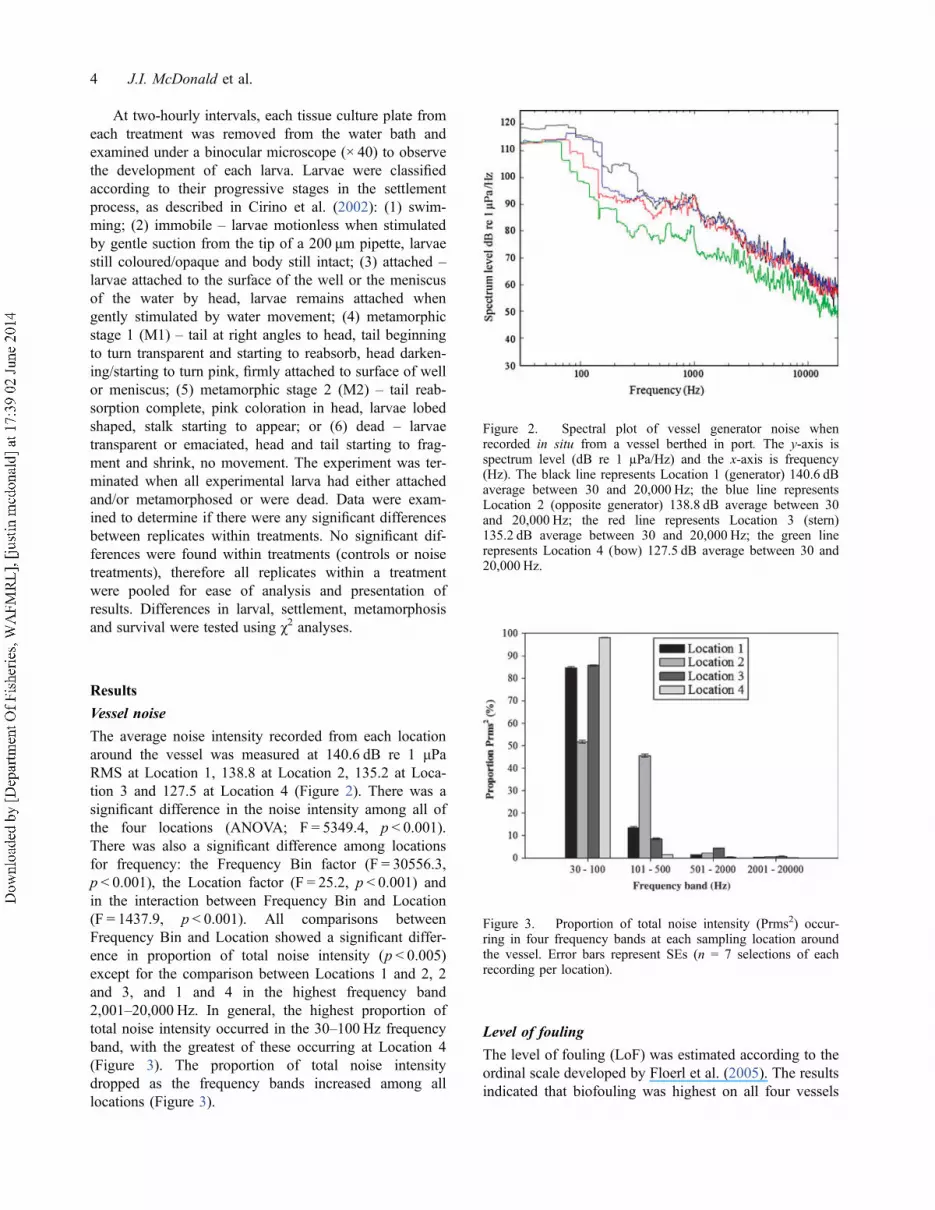
tion 3 and 127.5 at Location 4 (Figure 2). There was a
significant difference in the noise intensity among all of
the four locations (ANOVA; F = 5349.4, p < 0.001).
There was also a significant difference among locations
for frequency: the Frequency Bin factor (F = 30556.3,
p < 0.001), the Location factor (F = 25.2, p < 0.001) and
in the interaction between Frequency Bin and Location
(F = 1437.9, p < 0.001). All comparisons between
Frequency Bin and Location showed a significant differ-
ence in proportion of total noise intensity (p < 0.005)
except for the comparison between Locations 1 and 2, 2
and 3, and 1 and 4 in the highest frequency band
2,001–20,000 Hz. In general, the highest proportion of
total noise intensity occurred in the 30–100 Hz frequency
band, with the greatest of these occurring at Location 4
(Figure 3). The proportion of total noise intensity
dropped as the frequency bands increased among all
locations (Figure 3).
Level of fouling
The level of fouling (LoF) was estimated according to the
ordinal scale developed by Floerl et al. (2005). The results
indicated that biofouling was highest on all four vessels
Figure 3. Proportion of total noise intensity (Prms2) occur-ring in four frequency bands at each sampling location aroundthe vessel. Error bars represent SEs (n = 7 selections of eachrecording per location).
Figure 2. Spectral plot of vessel generator noise whenrecorded in situ from a vessel berthed in port. The y-axis isspectrum level (dB re 1 µPa/Hz) and the x-axis is frequency(Hz). The black line represents Location 1 (generator) 140.6 dBaverage between 30 and 20,000 Hz; the blue line representsLocation 2 (opposite generator) 138.8 dB average between 30and 20,000 Hz; the red line represents Location 3 (stern)135.2 dB average between 30 and 20,000 Hz; the green linerepresents Location 4 (bow) 127.5 dB average between 30 and20,000 Hz.
4 J.I. McDonald et al.
N OPQR OST UTV WXN UYSZ[ \UQ[]̂_̀ ab UZ` Ua cde_fghi cXj ka[` Q\lT OQSRTi S[mn opqrst kQUsrmu
examined, at the location closest to the generator
(Figure 4). This site was also confirmed to have the high-
est intensity of noise when analysed. The biofouling level
(relative abundance of fouling present on the vessel sur-
face) decreased with increasing distance away from the
noise source (ie the generator), with the bow showing the
lowest level of biofouling. All four vessels examined
showed a similar trend, with the highest overall LoF at
the generator and lowest at the bow. Intermediate LoF
ranks were determined for the sites opposite the generator
and at the stern. Both observers consistently scored the
same fouling estimate for every vessel and location sam-
pled. Biofouling consisted predominantly of colonial and
solitary ascidians (Polycitor sp., Sigillina sp., Botrylloides
sp., Styela sp.) bryozoans (Bugula sp., Zoobotryon
verticillatum, Watersipora subtorquata, W. arcuata), ser-
pulid polychaetes (unidentified) and porifera (unidenti-
fied). Although specific species data are not presented
here, a total of 24 morphotypes were identified from the
biofouling samples, of which four were confirmed to be
non-indigenous species and three were cryptogenic.
Larval settlement experiment
At the commencement of the experiment, all C. intestinalis
larvae were swimming and viable when introduced into the
experimental chambers.
Settlement
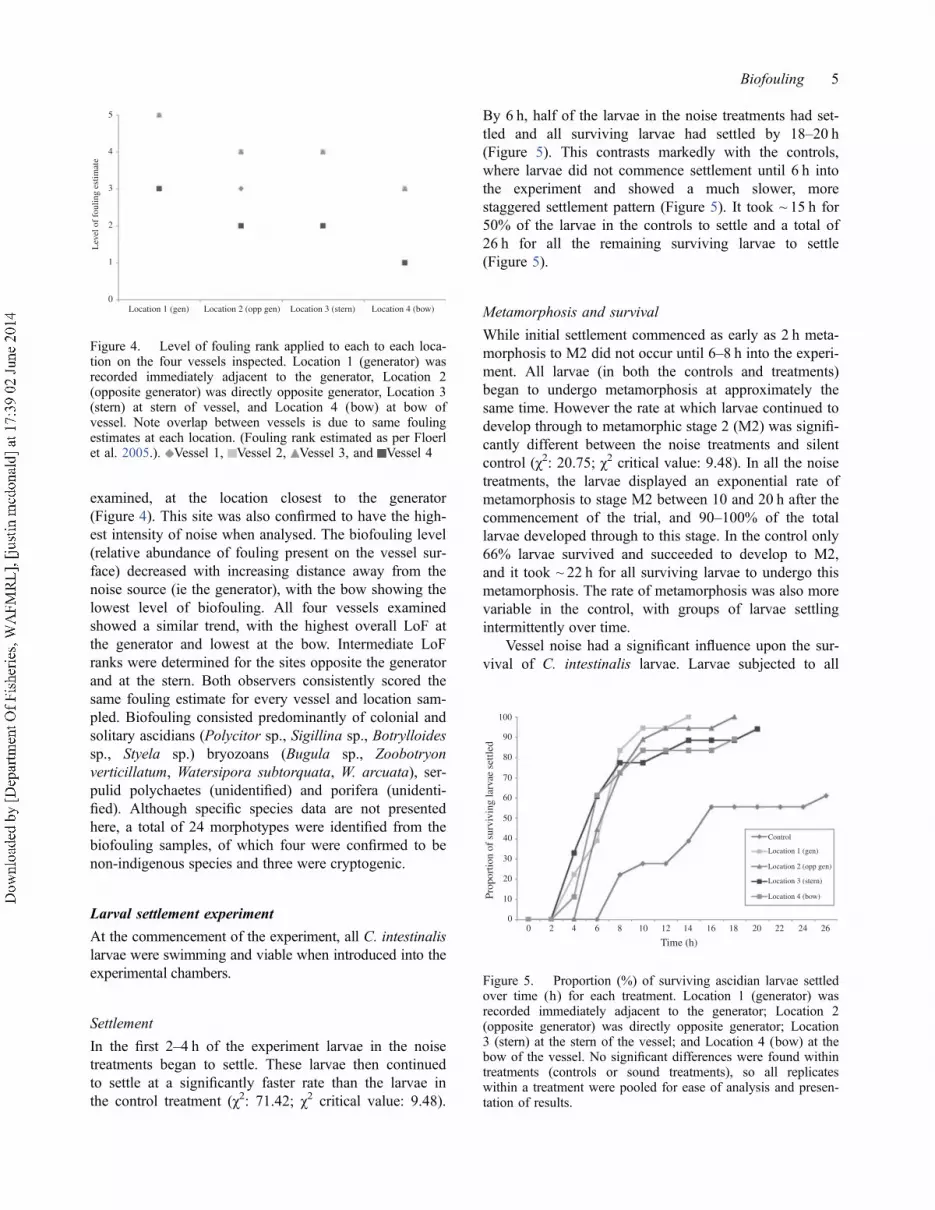
In the first 2–4 h of the experiment larvae in the noise
treatments began to settle. These larvae then continued
to settle at a significantly faster rate than the larvae in
the control treatment (χ2: 71.42; χ2 critical value: 9.48).
By 6 h, half of the larvae in the noise treatments had set-
tled and all surviving larvae had settled by 18–20 h
(Figure 5). This contrasts markedly with the controls,
where larvae did not commence settlement until 6 h into
the experiment and showed a much slower, more
staggered settlement pattern (Figure 5). It took ~ 15 h for
50% of the larvae in the controls to settle and a total of
26 h for all the remaining surviving larvae to settle
(Figure 5).
Metamorphosis and survival
While initial settlement commenced as early as 2 h meta-
morphosis to M2 did not occur until 6–8 h into the experi-
ment. All larvae (in both the controls and treatments)
began to undergo metamorphosis at approximately the
same time. However the rate at which larvae continued to
develop through to metamorphic stage 2 (M2) was signifi-
cantly different between the noise treatments and silent
control (χ2: 20.75; χ2 critical value: 9.48). In all the noise
treatments, the larvae displayed an exponential rate of
metamorphosis to stage M2 between 10 and 20 h after the
commencement of the trial, and 90–100% of the total
larvae developed through to this stage. In the control only
66% larvae survived and succeeded to develop to M2,
and it took ~ 22 h for all surviving larvae to undergo this
metamorphosis. The rate of metamorphosis was also more
variable in the control, with groups of larvae settling
intermittently over time.
Vessel noise had a significant influence upon the sur-
vival of C. intestinalis larvae. Larvae subjected to all
5
4
3
2
1
0
Lev
el o
f fo
uli
ng e
stim
ate
Location 1 (gen) Location 2 (opp gen) Location 3 (stern) Location 4 (bow)
Figure 4. Level of fouling rank applied to each to each loca-tion on the four vessels inspected. Location 1 (generator) wasrecorded immediately adjacent to the generator, Location 2(opposite generator) was directly opposite generator, Location 3(stern) at stern of vessel, and Location 4 (bow) at bow ofvessel. Note overlap between vessels is due to same foulingestimates at each location. (Fouling rank estimated as per Floerlet al. 2005.). Vessel 1, Vessel 2, Vessel 3, and Vessel 4
Pro
port
ion o
f su
rviv
ing l
arvae
set
tled
100
90
80
70
60
50
40
30
20
10
00 2 4 6 8 10 12 14
Time (h)
Control
Location 1 (gen)
Location 2 (opp gen)
Location 3 (stern)
Location 4 (bow)
16 18 20 22 24 26
Figure 5. Proportion (%) of surviving ascidian larvae settledover time (h) for each treatment. Location 1 (generator) wasrecorded immediately adjacent to the generator; Location 2(opposite generator) was directly opposite generator; Location3 (stern) at the stern of the vessel; and Location 4 (bow) at thebow of the vessel. No significant differences were found withintreatments (controls or sound treatments), so all replicateswithin a treatment were pooled for ease of analysis and presen-tation of results.
Biofouling 5
N OPQR OST UTV WXN UYSZ[ \UQ[]̂_̀ ab UZ` Ua cde_fghi cXj ka[` Q\lT OQSRTi S[mn opqrst kQUsrmu

vessel noise treatments had significantly greater survival
rates than those in the silent treatment (χ2: 56.55; χ2 criti-
cal value: 9.48). In noise treatment 1 (immediately adja-
cent to the generator) and 2 (directly opposite the
generator), survival was 100%. In noise treatments 3 (at
the stern) and 4 (at the bow), survival was 94 and 88%
respectively. However in the control (which was silent),
survival was only 66%.
Discussion
The research presented here provides evidence that
spatial differences in noise emission exist at different
locations on a vessel, and demonstrates a relationship
between these noise emissions and in situ biofouling.
The study experimentally demonstrated that when
exposed to increased levels of vessel noise, larval settle-
ment, metamorphosis and survival increases, compared
with a silent control.
Spatial distribution of sound on the vessel
The intensity of the vessel generator noise was con-
firmed to be highest at the site closest to the generator
(Location 1) and decreased away from the noise source
(with Location 4 with the lowest intensity). The propor-
tion of total noise intensity in each frequency bin dif-
fered significantly around the vessel. However, at all
sites on the vessel low frequency (30–100 Hz frequency
bin) dominated the noise spectra. Therefore it appears
that the low frequency noise may be the important factor
which is driving increased larval settlement and survival.
One possible explanation for increased settlement on
vessel hulls is that low frequency vessel noise may
reduce the boundary layer near the surface of the hull
and therefore make the settlement surface more accessi-
ble to larvae. However more research is required to
ascertain whether this is a behavioural or physical mech-
anism which is influencing larval settlement. Noise is
certainly not the only larval settlement or metamorphosis
cue and a major function of the settlement stage is in
response to a range of environmental inputs (Rittschof
et al. 1998). Chemical cues (Alfaro et al. 2006), surface
rugosity (Alfaro et al. 2004), light and temperature
(Bayne 1964; Carl et al. 2011) are also important. How-
ever, noise is likely to be an important factor in ports
and marinas due to its ability to traverse relatively large
distances and through turbid, turbulent waters which
may restrict or alter chemical or visual cues.
Level of fouling
The influence of the spatial distribution of sound inten-
sity was reflected in the observed LoF ranks at each of
the four locations on the vessel. Observed levels of
biofouling (applied according to the LoF rank; Floerl
et al. 2005) were higher on the fishing vessel in the sites
closest to the generator. The observed LoF data coupled
with the spatial distribution of sound around the vessel
suggest that low frequency noise is influencing a range
of biofouling species and the trend is more widespread
than initially thought.
Laboratory experiments
Results from this research clearly demonstrated that set-
tlement, metamorphosis and survival in C. intestinalis
larvae are influenced by vessel generator noise in labora-
tory experiments. C. intestinalis larvae exposed to vessel
generator noise settled and metamorphosed significantly
faster than larvae which were not exposed to any genera-
tor noise. Approximately 50% of the surviving larvae
exposed to vessel generator noise had settled 6 h after
the commencement of the experiment, with the remain-
ing larvae all settling by 18 h. This contrasts markedly
with the control where it took 15 h for 50% of surviving
larvae to settle and a total of 26 h for all surviving larvae
to settle. The enhanced settlement attributable to vessel
generator noise may indicate that stationary vessels not
only provide a suitable substratum for the settlement of
fouling species (Godwin 2003) but the noise from vessel
generators may be attracting larvae and enhancing their
survival. Substratum vibration resulting from vessel
sound may also contribute to increased settlement in that
the physical vibration influences the behaviour of the
larvae. Ascidian larvae attach to surfaces when there is
contact with the apical papillae so it is possible that
vibration of the settlement surface is sufficient to stimu-
late adhesive release (Rittschof et al. 1998; Aldred &
Clare 2014) (a physical rather than behavioural mecha-
nism, driven by low frequency sound vibration). Many
larvae have mechanosensory ability and can detect vibra-
tion, possibly signalling permanence to the larvae, that
is, the likelihood that a surface will last sufficiently long
for growth, maturation and reproduction (Rittschof et al.
1998). Clearly, further investigation into the mechanism
by which vessel sound influences larvae settlement
behaviour is required.
In its native range, natural populations of C. intestinalis
are found attached to rocky substrata (Petersen & Riisgard
1992), suggesting that ambient underwater sound produced
by rocky coastal habitats may be one important cue for the
settlement of C. intestinalis larvae. C. intestinalis also has
a tendency to attach to the hulls of ships and foul pylons
and other marine infrastructure (McDonald 2004) of noisy
ports and marinas around the globe. Although
C. intestinalis was not present on the vessels sampled, this
is an excellent candidate for laboratory trials as it is such a
prolific biofouler. In this experiment, not only did noise
have a significant influence on settlement, but once settled,
6 J.I. McDonald et al.
N OPQR OST UTV WXN UYSZ[ \UQ[]̂_̀ ab UZ` Ua cde_fghi cXj ka[` Q\lT OQSRTi S[mn opqrst kQUsrmu

larvae underwent metamorphosis at a much faster rate
when exposed to vessel generator noise. Development to
M2 stage (recognised as when a larvae has settled,
reabsorbed the tail and developed into an adult form) was
achieved in 60% of the larvae exposed to the noise
treatments (over a 12 h period), compared with only 20%
in the control treatment over the same period. Larvae
subjected to the two highest intensity noise treatments
(immediately adjacent to the generator and opposite the
generator) had a 100% survival rate compared to a maxi-
mum survival of only 66% in the silent control. The
enhanced survival (89–100% across all noise treatments)
and development of larvae when influenced by vessel
noise represents a significant risk in terms of vessel medi-
ated spread of invasive species. While sound reception has
been reported in some cephalopod molluscs (Packard et al.
1990; Williamson 1995; Mooney et al. 2010), to the
authors’ knowledge, the discovery that ascidian larvae can
detect and respond to vessel noise is the first description of
an auditory response in ascidians, which is a major group
in terms of biomass in shallow aquatic communities and
also biofouling assemblages (Lambert & Lambert 2003).
This response to vessel noise has been previously
documented in mussel larvae (Wilkens et al. 2012),
suggesting the response is active in other fouling taxa, but
until now this theory has not been documented in fouling
species such as ascidians.
As the density of the water transmits acoustic energy
very efficiently and over greater distances than in air, the
effects of underwater noise in a port environment may
extend over vast distances. The transmission of vessel
generator noise may also be increasing the area over
which larvae are attracted to a vessel. The vessel noises
recorded in this study had a noise intensity of between
127.5 dB and 140.6 dB re 1 μPa RMS (and between
50–24,000 Hz) which is within the range of sound inten-
sities recorded for natural reefs (Radford et al. 2008a,
2008b, 2010; Simpson et al. 2011). Assuming cylindrical
spreading of noise from the ship hull, which is appropri-
ate in the shallow water where vessels are berthed, then
the noise would occur at ~ 500 m out from the hull. Thus
if a clean vessel enters an infected port and runs on gen-
erator power it could be attracting fouling pest species
from a ~ 500 m radius. It has been demonstrated that
vessels acquire the majority of hull fouling while moored
in coastal ports, primarily because these environments
act as hubs for domestic and international shipping
movements (Carlton 1987). In 2002, Callow and Callow
reported that biofouling was a large and continuing prob-
lem, one that costs the US Navy up to $1 billion per
annum to manage. Shultz et al. (2011) also document
massive economic costs and highlight the fact that for a
single vessel class within the US Navy fleet biofouling
costs US$56 million.
Interestingly, there was no significant difference in
larval settlement, metamorphosis or survival rates between
the different noise treatments. This contrasts with the
observed level of fouling present on the vessels, where
significantly greater fouling cover was associated with
higher noise intensity (immediately adjacent to the genera-
tor and decreasing with distance from the noise source). A
possible explanation for this may be that C. intestinalis
larvae are not able to discriminate between the different
intensities associated with the four noise treatments. Many
other species such as crabs, fish and coral have been
documented to respond to ambient noise within the
100–1,000 Hz range (Montgomery et al. 2006; Radford
et al. 2007; Vermeij et al. 2010), which is certainly within
the range tested in these settlement experiments, and pos-
sibly other species are more capable of discriminating
between noise intensity, thereby explaining the observed
differences in biofouling on the vessel hulls.
Conclusion
This research suggests that vessels running generators in
ports may be promoting the settlement and survival of
marine pest species on their submerged surfaces. In the
absence of light, temperature or chemical cues from con-
specifics, the rates of settlement, metamorphosis and sur-
vival are significantly increased in C. intestinalis larvae
when exposed to vessel noise. It is, of course, more cost-
effective to prevent invasive species from establishing
themselves than it is to try to eradicate them once they
have become established. The goal therefore should be to
minimise the arrival of new species. As ports are apparent
havens for introduced marine species, vessels running
generators in ports seem to be at higher risk of attracting
biofouling based on the results presented here. Mecha-
nisms to ameliorate this biofouling potential should be
further investigated and tested. The dampening of genera-
tor noise may reduce the rate at which fouling accrues, or
vessels may be able to run on land-based power supplies
as opposed to ship-based generators. Alternatively there
may be a frequency at which vessel noise either ceases to
be a concern or can be ‘tuned’ to repel fouling. These
areas need to be further investigated.
Acknowledgements
The authors thank Kailis Marine (Terry Hewitt) for access to vesselsin Fremantle, Caroline Williams (NIWA) for collecting Cionaintestinalis adults, the University of Auckland Leigh MarineLaboratory for provision of laboratory space and equipment,Samantha Bridgwood for assistance with fieldwork and internalreviewers for constructive feedback and review of the manuscript.This research was funded by NIWA under the Innovative Seed Fund(2011/2012) and Coasts and Oceans Programme 4 – MarineBiosecurity (2014/15 SCI), the Glenn Family Foundation andWestern Australian Department of Fisheries. The authors wish tothank Wayne Young and Dan Pederson (Dampier Port Authority)for their support and enthusiasm.
Biofouling 7
N OPQR OST UTV WXN UYSZ[ \UQ[]̂_̀ ab UZ` Ua cde_fghi cXj ka[` Q\lT OQSRTi S[mn opqrst kQUsrmu

References
Aldred N, Clare AS. 2014. Mini-review: impact and dynamicsof surface fouling by solitary and compound ascidians.Biofouling. 30:259–270.
Alfaro AC, Jeffs AG, Creese RG. 2004. Bottom-drifting algal/mussel spat association along sandy coastal region innorthern New Zealand. Aquaculture. 241:269–290.
Alfaro AC, Copp BR, Appleton DR, Kelly S, Jeffs AG. 2006.Chemical cues promote settlement in larvae of the green-lipped mussel, Perna canaliculus. Aquacult Int. 14:405–412.
Bayne BL. 1964. The responses of the larvae of Mytilus edulisL. to light and to gravity. Oikos. 15:162–174.
Callow ME, Callow JA. 2002. Marine biofouling: a stickyproblem. Biologist. 49:1–5.
Carl C, Poole AJ, Vucko MJ, Williams MR, Whalan S, de Nys R.2011. Optimising settlement assays of pediveligers andplantigrades of Mytilus galloprovincialis. Biofouling. 27:859–868.
Carlton JT. 1987. Patterns of transoceanic marine biologicalinvasions in the Pacific Ocean. Bull Mar Sci. 41:452–465.
Cirino P, Toscano A, Caramiello D, Macina A, Miraglia V,Monte A 2002. Laboratory culture of the ascidian Cionaintestinalis (L.): a model system for molecular developmen-tal biology research. Mar Mod Elec Rec. [cited 2012 Oct10]. Available from: http://www.mbl.edu/html/BB/MMER/CIR/CirTit.html
Coutts ADM, Moore KM, Hewitt CL. 2003. Ships’ sea-chests:an overlooked transfer mechanism for non-indigenous mar-ine species? Mar Poll Bull. 46:1504–1515.
Fitridge I, Dempster T, Guenther J, DeNys R. 2012. Theimpact and control of biofouling in marine aquaculture: areview. Biofouling. 28:649–669.
Floerl O, Pool TK, Inglis GJ. 2004. Positive interactionsbetween nonindigenous species facilitate transport byhuman vectors. Ecol Appl. 14:1724–1736.
Floerl O, Inglis GJ, Hayden BJ. 2005. A risk-based predictivetool to prevent accidental introductions of non-indigenousmarine species. Environ Manage. 35:765–778.
Glasby TM, Connell SD. 1999. Urban structures as marinehabitats. Ambio. 28:595–598.
Godwin LS. 2003. Hull fouling of maritime vessels as a path-way for marine species invasions to Hawaiian islands. Bio-fouling. 19:123–131.
Gollasch S. 2002. The importance of ship hull fouling as a vec-tor of species introductions into the North Sea. Biofouling.18:105–121.
Götz T, Hastie G, Hatch LT, Raustein O, Southall BL, Tasker M,Thomsen F. 2009. Overview of the impacts of anthropogenicunderwater sound in the marine environment. London:OSPAR Commission; p. 134.
Green DM, DeFerrari HA, McFadden D, Pearse JS, PopperAN, Richardson WJ, Ridgway SH, Tyack PL. 1994.Low-frequency sound and marine mammals: currentknowledge and research needs. Washington (DC): NationalAcademy Press NRC Report.
Hewitt CL. 2004. Invasive species – vectors and managementstrategies. Book Review. J Exp Mar Biol Ecol. 311:186–187.
Jeffs AG, Radford C, Stanley JA. 2011. Ecological effects ofacoustic pollution in the marine environment. In: Allodi S,Nazari E, editors. Pollution in the marine environment.New Delhi: Research Signpost Publishing; p. 176–198.
Jeffs A, Tolimieri N, Montgomery JC. 2003. Crabs on cue forthe coast: the use of underwater sound for orientation bypelagic crab stages. Mar Freshwater Res. 54:841–845.
Jeffs AG, Montgomery JC, Tindle CT. 2005. How do spinylobster post-larvae find the coast? New Zeal J Mar Fresh.39:605–617.
Kroodsma DE, Byers BE, Goodale E, Johnson S, Liu WC.2001. Pseudoreplication in playback experiments, revisiteda decade later. Anim Behav. 61:1029–1033.
Lambert G. 2001. A global review of ascidian introduction andtheir possible impact on the endemic fauna. In: Sawada R,Yokosawa H, Lambert CC, editors. The biology of ascidians.Heidelberg, New York, Tokyo: Springer-Verlag; p.249–257.
Lambert CG, Lambert G. 2003. Persistence and differentialdistribution of nonindigenous ascidians in harbours of theSouthern California Bight. Mar Ecol Prog Ser. 259:145–161.
McDonald JI. 2004. The invasive pest species Ciona intestinalis(Linnaeus, 1767) reported in a harbour in southern WesternAustralia. Mar Poll Bull. 49:854–874.
Montgomery JC, Jeffs AG, Simpson S, Meekan M, Tindle C.2006. Sound as an orientation cue for the pelagic larvae ofreef fish and decapod crustaceans. Adv Mar Biol. 51:143–196.
Mooney TA, Hanlon RT, Christensen-Dalsgaard J, Madsen PT,Ketten DR, Nachtigall PE. 2010. Sounds detection by thelongfin squid (Loligo pealeii) studied with auditory evokedpotentials: sensitivity to low-frequency particle motion andnot pressure. J Exp Biol. 213:3748–3759.
Packard A, Karlsen HE, Sand O. 1990. Low frequency hearingin cephalopods. J Comp Physiol. 166:501–505.
Peterson JK, Riisgard HU. 1992. Filtration capacity of theascidian Ciona intestinalis and its grazing impact in ashallow fjord. Mar Ecol Prog Ser. 88:9–17.
Radford CA, Jeffs AG, Montgomery JC. 2007. Directionalswimming response of five species of crab postlarvae inresponse to reef sound. Bull Mar Sci. 80:69–378.
Radford CA, Jeffs AG, Tindle CT, Montgomery JC. 2008a.Resonating sea urchin skeletons create coastal choruses.Mar Ecol Prog Ser. 362:37–43.
Radford CA, Jeffs AG, Tindle CT, Montgomery JC. 2008b.Temporal patterns in ambient noise of biological originfrom a shallow water temperate reef. Oecologia. 156:921–929.
Radford CA, Stanley JA, Tindle CT, Montgomery JC, JeffsAG. 2010. Localised coastal habitats have distinct under-water sound signatures. Mar Ecol Prog Ser. 401:21–29.
Rittschof D, Forward RB, Cannon G, Welch JM, McClary Jr. M,Holm ER, Clare AS, Conova S, McKelvey LM, Bryan P,Van Dover CL. 1998. Cues and context: larval responses tophysical and chemical cues. Biofouling. 12:31–44.
Schultz MP, Bendick JA, Holm ER, Hertel WM. 2011. Eco-nomic impact of biofouling on a naval surface ship. Bio-fouling. 27:87–98.
Simpson SD, Meekan MG, Jeffs AG, Montgomery JC,McCauley RD. 2008. Settlement-stage coral reef fishes pre-fer the higher frequency invertebrate-generated audiblecomponent of reef noise. Anim Behav. 75:1861–1868.
Simpson SD, Radford AN, Tickle EJ, Meekan MG, Jeffs AG.2011. Adaptive avoidance of reef noise. PLoS ONE. 6:e16625.
Stanley J, Radford C, Jeffs AG. 2010. Induction of settlementin crab megalopae by ambient underwater reef sound.Behav Ecol. 21:113–120.
Stanley J, Radford C, Jeffs AG. 2012. Location, location, loca-tion: finding a suitable home among the noise. Proc R SocBiol Sci Ser B. 279:3622–3631.
8 J.I. McDonald et al.
N OPQR OST UTV WXN UYSZ[ \UQ[]̂_̀ ab UZ` Ua cde_fghi cXj ka[` Q\lT OQSRTi S[mn opqrst kQUsrmu

Tolimieri N, Jeffs AG, Montgomery JC. 2000. Ambient soundas a cue for navigation by the pelagic larvae of reef fishes.Mar Ecol Prog Ser. 207:219–224.
Vermeij MJA, Marhaver KL, Huijbera CM, Nagelkerken I,Simpson SD. 2010. Coral larvae move toward reef sounds.PLoS ONE. 5:e10660.
Wilkens SL, Stanley JA, Jeffs AG. 2012. Induction of settle-ment in mussel (Perna canaliculus) larvae by vessel noise.Biofouling. 28:65–72.
Williamson R. 1995. A sensory basis for orientation incephalopods. J Mar Biol Assoc UK. 75:83–92.
Wonham MJ, Carlton JT, Ruiz GM, Smith LD. 2000. Fish andships: relating dispersal frequency to success in biologicalinvasions. Mar Biol. 136:1111–1121.
Biofouling 9
N OPQR OST UTV WXN UYSZ[ \UQ[]̂_̀ ab UZ` Ua cde_fghi cXj ka[` Q\lT OQSRTi S[mn opqrst kQUsrmu