variation in neighbourhood context shapes frugivore-mediated facilitation and competition among...
TRANSCRIPT

Variation in neighbourhood context shapesfrugivore-mediated facilitation and competition amongco-dispersed plant speciesJ€org Albrecht1*†, Victoria Bohle1, Dana G. Berens1, Bogdan Jaroszewicz2, Nuria Selva3 andNina Farwig1
1Department of Ecology – Conservation Ecology, Faculty of Biology, Philipps-University Marburg, Karl-von-Frisch Str.8, D-35043 Marburg, Germany; 2Białowie _za Geobotanical Station, Faculty of Biology, University of Warsaw, Sportowa19, PL-17-230 Białowie _za, Poland; and 3Institute of Nature Conservation, Polish Academy of Sciences, Mickiewicza33, PL-31-120 Krak�ow, Poland
Summary
1. Co-occurring and simultaneously fruiting plant species may either compete for dispersal byshared frugivores or enhance each other’s dispersal through joint attraction of frugivores. Whilecompetitive plant–plant interactions are expected to cause the evolutionary divergence of fruit phe-nologies, facilitative interactions are assumed to promote their convergence. To which extent com-petitive and facilitative interactions among plant species with similar phenological niches arecontrolled by spatial variation in their local abundance and co-occurrence is poorly understood.2. Here, we test the hypotheses that when a plant species fruits in high densities, large phenologicaloverlap with other plant species causes competition for seed dispersers owing to frugivore satiation.Conversely, we expect large phenological overlap to enhance the dispersal of a plant species fruitingin low densities through attraction of frugivores by other species in its local neighbourhood.3. We test these predictions on plant–frugivore networks based on seed removal from 15 woody,fleshy-fruited plant species by 30 avian and 4 mammalian frugivore species across 13 study sites inBiałowie _za Forest, Poland.4. A null model indicated that fruit phenologies of the regional plant assemblage were more differ-entiated than expected by chance. In the local networks, the tendency of plants to share frugivoresincreased with phenological overlap. High phenological overlap reduced the seed removal rates,interaction strength (proportion of interactions) and the number of partners of plant species fruitingin high densities. Conversely, plant species fruiting in low densities mainly profited from high phe-nological overlap with other species. Importantly, the sharing of mutualistic partners among co-fruiting plant species was also reflected in their co-occurrence.5. Synthesis. Our study highlights that, in spite of the overall signal of competition, frugivore-medi-ated interactions among cofruiting plant species may consistently promote the establishment and per-sistence of rare species through facilitation. In addition, our results suggest that, among otherfactors, indirect coupling of species through shared mutualistic partners might be an important deter-minant of plant community assembly. The coupling through shared mutualists may cause the forma-tion of associations among co-dispersed plant species and might contribute to the coexistence ofspecies in plant–animal mutualistic communities.
Key-words: Białowie _za Forest, interaction networks, limiting similarity, niche differentiation, phe-nological overlap, plant–animal mutualism, plant–plant interactions, seed dispersal
Introduction
Dispersal is one of the key factors that sustains diversity inplant communities (Janzen 1970; Howe & Smallwood 1982;Nathan & Muller-Landau 2000; Webb & Peart 2001).
*Correspondence author: E-mail: [email protected]†Present address: Institute of Nature Conservation, Polish Academy ofSciences, Mickiewicza 33, PL-31-120 Krak�ow, Poland.
© 2015 The Authors. Journal of Ecology © 2015 British Ecological Society
Journal of Ecology doi: 10.1111/1365-2745.12375

Dispersal of seeds by frugivorous animals is prevalent in vari-ous terrestrial ecosystems and forms the basis for regenerationof fleshy-fruited plants (Herrera 2002). By dispersing theseeds of their food plants, frugivores essentially shape therates of recruitment (Janzen 1970; Webb & Peart 2001),the spatial mosaic of regeneration (Jordano et al. 2007), thegene flow between populations (Kamm et al. 2009), and thecolonization of habitats (Nathan & Muller-Landau 2000).Central to all plant–frugivore associations is that co-occur-
ring and simultaneously fruiting plant species share frugivores(Herrera 1984, 1998; Jordano 1987; Fuentes 1995; Carlo,Collazo & Groom 2003; Schupp, Jordano & Gomez 2010),because frugivores show a high plasticity in their foragingbehaviour and in their fruit preferences (Carnicer, Jordano &Melian 2009). Owing to this lack of strong reciprocal speci-ficity, plant–frugivore relationships are assumed to be diffuseand driven by indirect frugivore-mediated plant–plant interac-tions (Herrera 1985, 1995, 1998; Carlo, Aukema & Morales2007). Therefore, at the local scale of up to a few hectares,rates of seed removal from individual plants strongly dependon their con- and heterospecific fruiting neighbourhood(Carlo, Aukema & Morales 2007; Carlo & Morales 2008).The conspecific fruiting neighbourhood of a plant individualis characterized by the local occurrence of other individualsof the same species. The heterospecific fruiting neighbour-hood of a plant individual is defined by the local co-occur-rence and by the phenological overlap with individuals ofother species.Co-occurring plant species that fruit simultaneously may
either compete for dispersal by shared frugivores (Snow1965; Thompson & Willson 1979; Burns 2005) or enhanceeach other’s dispersal through joint attraction of frugivores(Wheelwright 1985; Poulin et al. 1999; Burns 2002). Accord-ing to the theory of ‘limiting similarity’, competitive plant–plant interactions are supposed to favour the evolutionarydivergence of fruit phenologies, while facilitative interactionsare expected to promote their convergence (Snow 1965; Mac-Arthur & Levins 1967; Thompson & Willson 1979). Theseselective regimes should result either in the differentiation orin the synchronization of fruit phenologies. Yet, the literaturedoes not provide conclusive evidence for these hypotheses,because temporal patterns of fruit production are stronglyconstrained by abiotic, physiological and evolutionary factorsas well as by other types of biotic interactions (e.g. pollina-tion or seed predation; Herrera 2002).Despite these constraints, the extent to which the fruit phe-
nology of a plant individual overlaps with the phenologies ofothers can vary depending on its local neighbourhood context.This variation in neighbourhood context emerges naturallydue to spatial variation in the local abundance and co-occur-rence of plant species (Thompson 2005). The concomitantspatial variation in the phenological differentiation amonglocally co-occurring plant species, in turn, may determine thecompetitive or facilitative character of indirect frugivore-medi-ated plant–plant interactions. For instance, owing to frugivoresatiation, large phenological overlap with other co-occurringplant species may cause competition for seed dispersers when
the density of con- and heterospecific fruits is high (Thomp-son & Willson 1979; Rathcke & Lacey 1985; Herrera et al.1994; Hampe 2008). Conversely, plant species that fruit in lowdensities may profit from large phenological overlap with otherco-occurring plant species that attract frugivores (Herrera 1984;Whelan et al. 1998; Carlo 2005; Carlo, Aukema & Morales2007). Therefore, considering the spatial variability in thelocal neighbourhood context seems essential to gain insightsinto the mechanisms that drive competitive and facilitativeinteractions among plant species with overlapping fruit phe-nologies.In a 2-year field study, we quantified seed removal by fru-
givorous birds and mammals from 15 woody, fleshy-fruitedplant species on 17 study sites located in the last relict of old-growth forest of the European lowland (Białowie _za, Poland).We combined these interaction data with estimates of localpopulation densities from 13 of these study sites and with theregional fruit phenologies of the species. First, we used a nullmodel to test for non-random temporal patterns in the fruitproduction of the regional plant assemblage; that is, we testedwhether fruit phenologies were more, or less, differentiatedthan expected by chance (Snow 1965; Burns 2005). Then, weassessed whether both the variation in the local co-occurrence,abundance and phenological overlap of plant species affectedtheir interactions with frugivores. To do so, we applied net-work analyses to quantify the specialization of plants (i.e. thetendency to share mutualistic partners), rates of seed removalfrom plants, their interaction strength and the number of theirmutualistic partners in the local plant–frugivore associations(Bascompte, Jordano & Olesen 2006; Bl€uthgen, Menzel &Bl€uthgen 2006; Bl€uthgen et al. 2008). We hypothesized that(i) high phenological overlap among plant species results inlow specialization of plants on frugivores and, thus, in a largeoverlap of disperser assemblages. This is a precondition forindirect frugivore-mediated competitive or facilitative interac-tions among plant species with similar phenological niches.We expected that (ii) high phenological overlap would causecompetition among plants when species fruit in high densities(i.e. phenological overlap should reduce seed removal rates,interaction strength and number of partners). Alternatively,(iii) high phenological overlap should have facilitative effectson interactions of plant species fruiting in low densities (i.e.phenological overlap should increase seed removal rates,interaction strength and number of partners).
Materials and methods
STUDY AREA
We conducted this study in the Białowie_za Forest, the last primaryold-growth forest of the European lowland, extending across the bor-der between Poland and Belarus. On Polish territory, the forest coversan area of about 625 km2. Within the Białowie_za National Park(c. 100 km2), an area of about 60 km2 is strictly protected and anarea of about 45 km2 has potentially never been commercially logged(Sokołowski 2004). Commercial logging has shaped over 80% of theremaining forest stands since the First World War (Bobiec et al.
© 2015 The Authors. Journal of Ecology © 2015 British Ecological Society, Journal of Ecology
2 J. Albrecht et al.
2000; Bobiec 2002, 2012). Most riverine areas of the forest had beencleared during the 16th and 17th centuries for the purpose of hay pro-duction (Sokołowski 2004).
STUDY SPECIES AND DESIGN
We focussed our study on woody, fleshy-fruited plant species thatdepend on seed dispersal by frugivorous birds and mammals. As thefocal plant species of our study are mainly associated with ash-alderflood plain forests (Fraxino-Alnetum community; Matuszkiewicz2001), we established our study sites within this plant association.We used maps of forest communities (Fali�nski 1994) to select studysites and verified their suitability in the field. In 2011 and 2012, weestablished a total of 17 study sites that were scattered over about400 km2 covering two-thirds of the Polish part of the Białowie_za For-est. The study sites were situated in old-growth and logged foreststands and covered various landscape contexts from forest interior toforest edges to forest fragments (Table S1, Supporting information).The pairwise distance between study sites ranged from 1.4 to 23 km(11 � 5.8 km, mean � SD).
On the study sites, we identified 15 woody, fleshy-fruited plant spe-cies from 12 genera and 5 families (6 tree and 9 shrub species;Table S2). In ash–alder floodplain forests, these species occur in highdensities and often form the dominant aspect of the mid- and understo-rey (Albrecht et al. 2013, 2014). Across study sites, the number of spe-cies varied between 7 and 14 (10 � 1.7 species per site; mean � SD).Plant densities varied between 10 and 103 individuals per 100 m2
across study sites (36 � 27 individuals per 100 m2; mean � SD). Thethree most common tree species of the midstorey were Frangula alnus(5.0 � 8.3 individuals per 100 m2; mean � SD), Prunus padus(4.2 � 3.5) and Sorbus aucuparia (2.2 � 3.4). The three most com-mon shrub species of the understorey were Ribes spicatum (5.9 � 5.6),R. nigrum (4.3 � 7.4) and Viburnum opulus (1.6 � 2.0). In somestudy sites, Rubus ideaus occurred in very high densities (10 � 11). Inthe study region, the fruiting period of the focal plant species rangesfrom the end of June until mid of October (Fig. 1a). During this period,the focal plant species represent critical resources for a diverse array offrugivorous birds and mammals (Table S3). Most fruits are removedfrom plants until the end of October, while undispersed fruits oftendesiccate and can remain on pedicels until budding in the next spring(J. Albrecht, pers. observation).
FRUGIVORE OBSERVATIONS
From June to October in 2011 and 2012, we monitored fruit ripening ofthe plant species on the study sites weekly to select fruiting individualsfor the frugivore observations (Table S4). We observed each plant spe-cies on each study site and year three times for 6 h starting at sunrise(18 h 9 plant species�1 9 study site�1 9 year�1). When less thanthree fruiting individuals of a plant species were available on a studysite, we repeated observations on the same one or two individuals,respectively, on different days. In total, we were able to select threeindividuals in 93 cases, two individuals in 27 cases and one individualin 31 cases for the frugivore observations per species on each study siteper year (Table S4). Observation sessions were conducted during theearly, mid and late fruiting period of each plant species. We observedplant–frugivore interactions with binoculars from camouflaged tentssimultaneously on different study sites (7 observers in 2011 and 14observers in 2012). We recorded all frugivore species visiting the indi-vidual plants, as well as the number of frugivore individuals, durationof visits and their fruit-handling behaviour (for details see Albrechtet al. 2013). In total, we accumulated 2718 h of seed disperser observa-tions (774 h in 2011 and 1944 h in 2012), distributed over 158 days(67 days in 2011 and 91 days in 2012). After each observation session,we counted the fruits of the observed plant individuals. For trees, wecounted the fruits on representative parts of the tree crown and thenextrapolated over the whole tree crown. Later, we used these crop sizeestimates for compiling the fruit phenologies of the 15 plant species andfor estimating the conspecific fruit densities of the species on the studysites (see sections: ‘Compilation of fruit phenologies’ and ‘Fruit densi-ties and phenological differentiation in local plant communities’).
CENSUS OF PLANT POPULATIONS
We quantified plant population densities on 13 of the 17 study sites (4study sites were not surveyed due to logistic constraints). To estimateplant population densities, we established 20 alternating squares of20 9 20 m2 size on 10 study sites and 10 squares on 3 study sites,which were situated in small forest fragments. Thus, we sampled plantdensities on an area of 0.8 ha on 10 sites and on an area of 0.4 ha on 3sites. Because all of the focal plant species were perennials, we assumedthat their population density would not change significantly betweenyears. Thus, we studied interactions with seed dispersers in two consec-utive seasons (2011 and 2012), but estimated plant abundances only in
(a) (b)
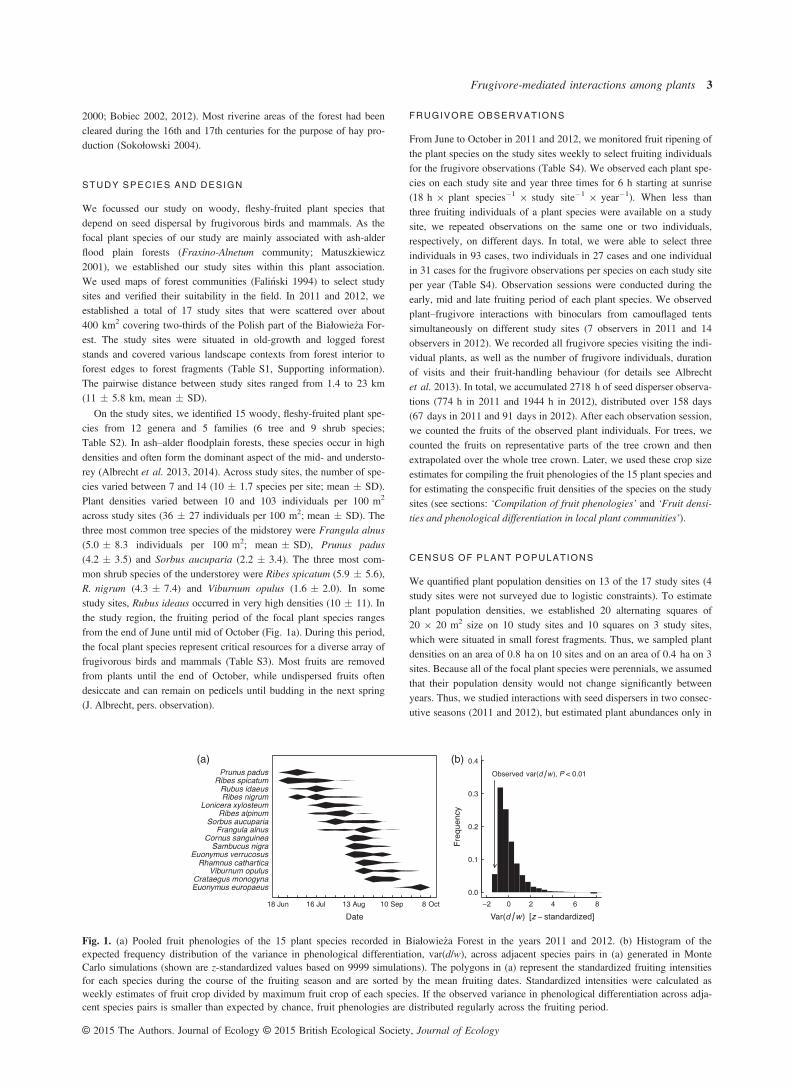
Fig. 1. (a) Pooled fruit phenologies of the 15 plant species recorded in Białowie_za Forest in the years 2011 and 2012. (b) Histogram of theexpected frequency distribution of the variance in phenological differentiation, var(d/w), across adjacent species pairs in (a) generated in MonteCarlo simulations (shown are z-standardized values based on 9999 simulations). The polygons in (a) represent the standardized fruiting intensitiesfor each species during the course of the fruiting season and are sorted by the mean fruiting dates. Standardized intensities were calculated asweekly estimates of fruit crop divided by maximum fruit crop of each species. If the observed variance in phenological differentiation across adja-cent species pairs is smaller than expected by chance, fruit phenologies are distributed regularly across the fruiting period.
© 2015 The Authors. Journal of Ecology © 2015 British Ecological Society, Journal of Ecology
Frugivore-mediated interactions among plants 3

the second season (2012). In each square, we counted the number ofindividuals of each focal plant species with a stem diameter of at least1 cm at 50 cm height. We counted all individuals of R. idaeus, becausethe species is a climber and the rule was not applicable. In addition, weclassified individuals into juveniles and adults based on their stemdiameter at 50 cm height. In R. idaeus, lignification of stems was usedfor classification. Plant individuals were considered as adults, if theirstem diameter was larger than 2 cm. Total plant density and the densityof adult plants were highly correlated (mean � SD of Spearman’s rankcorrelation across the 15 plant species: q = 0.91 � 0.11, in all casesn = 13 and P < 0.01). The results of subsequent analyses were similarfor both measures. Since in our study system total population densitiesof plants are strong predictors of interactions with pollinators and seeddispersers (Albrecht et al. 2014), we used the mean total density perspecies per square as an estimate of the population density on the studysites.
COMPILAT ION OF FRUIT PHENOLOGIES
We used the data on the crop size of the plant species from the frugi-vore observations to obtain estimates of the mean crop size of eachplant species in the course of the fruiting season in each of the 2study years. From the frugivore observations, we obtained an averageof 33 crop size estimates for each plant species throughout the fruit-ing season (Table S5). The crop size estimates had a temporal resolu-tion of approximately 2 days in both years (1.5 � 1.4 days;mean � SD; range: 1―6). Based on these data, we constructed amatrix with I rows representing the plant species and J columns rep-resenting periods of 1 week, in which each cell aij gave the meanindividual fruit crop of plant species i in week j. In each cell aij, theseestimates of fruit crop encompassed on average seven samples(6.5 � 3.8 samples 9 week�1 across plant species; mean � SD;range: 1.8–14). To increase the resolution of the fruit phenologies, weaveraged the weekly crop size estimates across the 2 years for the 10species that produced fruits in both years. This procedure seems justi-fiable, because the fruiting sequence of the species was consistentacross years (Spearman’s rank correlation, q = 0.76, P = 0.016), andthe between-year variation in the timing of fruit production relative tothe length of the fruiting periods was low (ratio: 0.06 � 0.32;mean � SD across ten plant species that produced fruits in bothyears). We standardized the phenology of each species to rangebetween 0 and 1, by dividing the fruit crop of plant species i in eachweek j by the maximum fruit crop of the respective species duringthe fruiting period (Figs 1a and 2b).
COMMUNITY-WIDE PHENOLOGICAL DIFFERENTIATION
IN THE REGIONAL PLANT ASSEMBLAGE
In the next step, we used the fruit phenologies of the 15 focal plant spe-cies to test for community-wide phenological differentiation of the fruitphenologies in the regional plant assemblage. To characterize phenolog-ical niche differentiation, we used the metric d/w (Taper & Chase1985). This metric is defined as the difference d between the averagephenotypes of two species divided by the sum of their standard devia-tions w. In the first step, species were ranked according to their averagefruiting times (Fig. 1a), which we calculated as the average week offruit production, weighted by the relative number of fruits produced ineach week. The length of the fruiting periods was defined as one stan-dard deviation around the average fruiting time, weighted by the rela-tive number of fruits produced in each week (Burns 2005). In thesecond step, we calculated the index d/w for all adjacent species pairs in
the temporal sequence and used the variance of these values to quantifythe community-level differentiation of fruit phenologies (Poole & Rath-cke 1979; Taper & Chase 1985; Burns 2005).
We used a null model to test whether the fruit phenologies weremore or less differentiated than expected by chance (Ashton, Givnish& Appanah 1988; Burns 2005). To do so, we compared the observedvariance in the index d/w to variance values expected from MonteCarlo simulations (Fig. 1b). To reduce type I error rates, we preservedthe length of the original fruiting season by keeping the positions ofthe fruiting periods of the first and last species fixed (Ashton, Givnish& Appanah 1988; Burns 2005). Simulations started by choosing ran-dom numbers from within the observed range of average fruitingtimes for the 13 remaining species. The fruiting periods of the plantspecies were then assigned to the random numbers according to theobserved species sequence. Preserving the original species sequenceaccounts for potential phylogenetic constraints on the length of thefruiting periods of the plant species (Ashton, Givnish & Appanah1988). Then, the index d/w was calculated for all adjacent speciespairs in the sequence, and the variance of these values was calculated.This procedure was iterated 9999 times. We multiplied the fraction ofiterations generating variances that were smaller than the observedvariance by two to obtain two-tailed P-values.
FRUIT DENSIT IES AND PHENOLOGICAL
DIFFERENTIAT ION IN LOCAL PLANT COMMUNIT IES
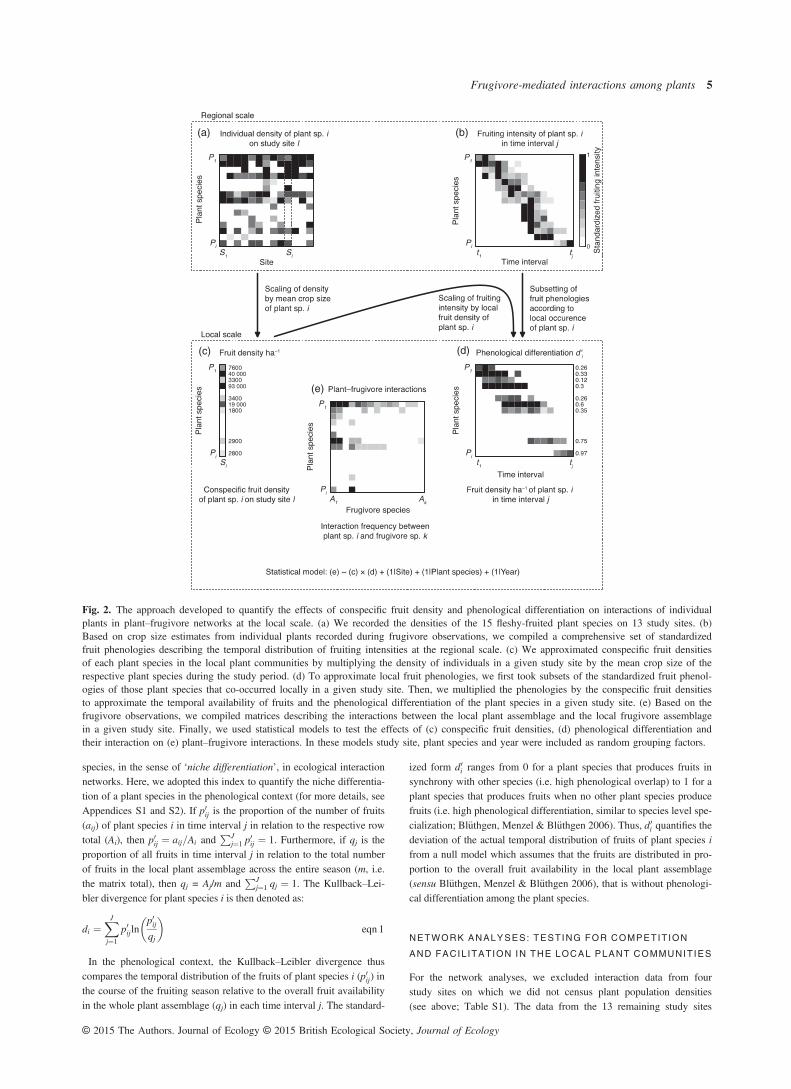
The main prediction of our study was that variation in the abundanceand co-occurrence of plant species between study sites determines theeffect of phenological differentiation on interactions between frugi-vores and individual plants. To capture this variability, we quantifiedthe conspecific fruit density of a plant species (Fig. 2c), as well as itsphenological differentiation from other plant species in the local com-munity (Fig. 2d).
To quantify the fruit density of plant species i on each study site(mean density of fruits per hectare on each study site in each year),we multiplied the number of individuals of plant species i on thestudy sites by the mean crop size of the respective plant species in agiven year (Fig. 2a,c). Therefore, the fruit densities of a plant speciescould vary between both study years depending on the average fruitcrop of a species in a given year. Thus, we assumed that all plantindividuals in a population produced fruits in a given year. Althoughthis approach may overestimate the total fruit density within popula-tions, our conclusions should not be affected, since this bias shouldbe consistent across plant species.
To quantify the phenological differentiation of each plant species iin the local plant communities, we constructed local ‘realized’ phe-nologies of the plants based on the regional set of fruit phenologies(Fig. 2b,d). For each study site in each year, we created a subset ofphenologies for those plant species that produced fruits on the respec-tive study site in the respective year. The regional set of fruit phenol-ogies was based on the standardized fruiting intensities of each plantspecies (see above; range: 0–1). Thus, we multiplied these standard-ized phenologies with the conspecific fruit density of the plant specieson a given study site in the respective study year to get an estimateof the temporal distribution of the conspecific fruit density aij for eachplant species i in each week j and on each study site in each year.
To quantify the phenological differentiation of plant species i in thelocal plant communities, we used the Kullback–Leibler divergence di asa measure of relative entropy in its standardized form (Bl€uthgen, Men-zel & Bl€uthgen 2006). This index was originally suggested byBl€uthgen, Menzel & Bl€uthgen (2006) to measure the specialization of
© 2015 The Authors. Journal of Ecology © 2015 British Ecological Society, Journal of Ecology
4 J. Albrecht et al.
species, in the sense of ‘niche differentiation’, in ecological interactionnetworks. Here, we adopted this index to quantify the niche differentia-tion of a plant species in the phenological context (for more details, seeAppendices S1 and S2). If p0ij is the proportion of the number of fruits(aij) of plant species i in time interval j in relation to the respective rowtotal (Ai), then p0ij ¼ aij=Ai and
PJj¼1 p
0ij ¼ 1. Furthermore, if qj is the
proportion of all fruits in time interval j in relation to the total numberof fruits in the local plant assemblage across the entire season (m, i.e.the matrix total), then qj = Aj/m and
PJj¼1 qj ¼ 1. The Kullback–Lei-
bler divergence for plant species i is then denoted as:
di ¼XJj¼1
p0ijlnp0ijqj
� �eqn 1
In the phenological context, the Kullback–Leibler divergence thuscompares the temporal distribution of the fruits of plant species i (p0ij) inthe course of the fruiting season relative to the overall fruit availabilityin the whole plant assemblage (qj) in each time interval j. The standard-
ized form d0i ranges from 0 for a plant species that produces fruits insynchrony with other species (i.e. high phenological overlap) to 1 for aplant species that produces fruits when no other plant species producefruits (i.e. high phenological differentiation, similar to species level spe-cialization; Bl€uthgen, Menzel & Bl€uthgen 2006). Thus, d0i quantifies thedeviation of the actual temporal distribution of fruits of plant species ifrom a null model which assumes that the fruits are distributed in pro-portion to the overall fruit availability in the local plant assemblage(sensu Bl€uthgen, Menzel & Bl€uthgen 2006), that is without phenologi-cal differentiation among the plant species.
NETWORK ANALYSES: TESTING FOR COMPETIT ION
AND FACIL ITAT ION IN THE LOCAL PLANT COMMUNIT IES
For the network analyses, we excluded interaction data from fourstudy sites on which we did not census plant population densities(see above; Table S1). The data from the 13 remaining study sites
(a)
(c) (d)
(e)
(b)
Fig. 2. The approach developed to quantify the effects of conspecific fruit density and phenological differentiation on interactions of individualplants in plant–frugivore networks at the local scale. (a) We recorded the densities of the 15 fleshy-fruited plant species on 13 study sites. (b)Based on crop size estimates from individual plants recorded during frugivore observations, we compiled a comprehensive set of standardizedfruit phenologies describing the temporal distribution of fruiting intensities at the regional scale. (c) We approximated conspecific fruit densitiesof each plant species in the local plant communities by multiplying the density of individuals in a given study site by the mean crop size of therespective plant species during the study period. (d) To approximate local fruit phenologies, we first took subsets of the standardized fruit phenol-ogies of those plant species that co-occurred locally in a given study site. Then, we multiplied the phenologies by the conspecific fruit densitiesto approximate the temporal availability of fruits and the phenological differentiation of the plant species in a given study site. (e) Based on thefrugivore observations, we compiled matrices describing the interactions between the local plant assemblage and the local frugivore assemblagein a given study site. Finally, we used statistical models to test the effects of (c) conspecific fruit densities, (d) phenological differentiation andtheir interaction on (e) plant–frugivore interactions. In these models study site, plant species and year were included as random grouping factors.
© 2015 The Authors. Journal of Ecology © 2015 British Ecological Society, Journal of Ecology
Frugivore-mediated interactions among plants 5

comprised 2304 h of seed disperser observations. For these studysites, we constructed quantitative interaction matrices based on thefrequency of interactions between plants and frugivores (Fig. 2e). Ifwe had data from 2 years for a study site, we constructed separateinteraction matrices for each year. Across the 13 study sites and bothyears, we quantified a total of 20 interaction matrices. In contrast toprevious network studies (Schleuning et al. 2011; Menke, B€ohning-Gaese & Schleuning 2012; Albrecht et al. 2013), here we defined theinteraction frequency as the number of seeds that frugivores removedfrom plants. Thus, our estimates of interaction frequency reflect thequantity component of seed dispersal (Schupp, Jordano & Gomez2010). To do so, we first estimated the mean number of fruits thatfrugivore species k removed from plant species i per visit based onthe frugivore observations (Farwig, B€ohning-Gaese & Bleher 2006;Albrecht, Neuschulz & Farwig 2012). Then, we multiplied the num-ber of visits of frugivore species k on plant species i by the meannumber of fruits that frugivore species k removed from the plant spe-cies i per visit. Finally, we multiplied the number of fruits removedfrom plant species i by the mean number of seeds per fruit(Table S2). The total interaction frequency of a frugivore species wasdefined as the number of seeds removed from all plant species withina network, whereas the interaction frequency from the plants’ per-spective was defined as the total number of seeds removed from aplant species in a given network. The standardized sampling effortper plant species across the study sites allowed us to draw specificconclusions from the plants’ perspective in terms of seed removalrates and potential consequences for seed dispersal.
In order to test our first hypothesis that the tendency of plant spe-cies to share frugivores should increase with phenological overlap, wemeasured the specialization of plants on frugivores. In analogy tophenological differentiation above, we quantified the degree of com-plementary specialization among plants in the networks by calculatingthe standardized Kullback–Leibler divergence (d0i ) for each plant spe-cies i in each network (Bl€uthgen, Menzel & Bl€uthgen 2006). In thecontext of interaction networks, a value of 0 indicates highest possiblegeneralization and 1 indicates highest possible specialization of plantson frugivores. Therefore, d0 quantifies the tendency of plants to sharefrugivores with other plants, that is the degree of ‘niche exclusive-ness’ in mutualistic interactions (Bl€uthgen 2010).
We used a combination of three measures to test our second hypothe-sis that plant species with overlapping fruit phenologies should competefor frugivores if they occur in high densities, but should profit from phe-nological overlap if they occur in low densities. First, we used absoluteseed removal rates from the plants in the interaction networks as a sur-rogate of their potential dispersal capacity (Schupp, Jordano & Gomez2010). Secondly, we used the interaction strength of the plant species tomeasure their ‘relative impact’ in the interaction networks (Bascompte,Jordano & Olesen 2006; V�azquez et al. 2007). Interaction strength dif-fers conceptually from the niche property d0 because it directly reflectsthe interaction counts (Bl€uthgen 2010). We defined the interactionstrength of a plant species in a given network as the interaction fre-quency of plant species i divided by the total number of interactions inthe network (proportion of interactions that plant species i has in thenetwork). Thirdly, we used the number of partners (plant degree) toquantify the number of frugivore species that visited plant species i(Bascompte, Jordano & Olesen 2006).
STATIST ICAL ANALYSES
We used linear mixed-effects models to analyse the variation in thespecialization of plants, rates of seed removal from plants, their inter-
action strength and the number of mutualistic partners. In these analy-ses, we treated conspecific fruit density, phenological differentiationand their interaction as fixed continuous predictors. Conspecific fruitdensity and phenological differentiation were only weakly correlated(r = 0.18), which suggested no collinearity between these predictors.As not all plant species occurred on every study site and in eachstudy year (i.e. site, year and plant species were not full factorial), wefitted site (13 sites), year (2 years) and plant species (15 plant spe-cies) as separate random grouping factors. The model for plant spe-cialization was fitted with a Gaussian error distribution and anidentity link function. For the number of seeds Ai removed from plantspecies i, we fitted a model with Poisson error distribution and a loglink function. Since the residuals of this model were overdispersed(v2 = 21883.74, ratio = 180.9, d.f. = 121, P < 0.001), we fitted anadditional observation-level random effect to account for this extravariation (Hinde 1982; Williams 1982; Breslow 1984; Bates et al.2013). For the interaction strength of plants, we fitted a model withbinomial error distribution and a logit link function. In this model, weanalysed the interaction strength (response variable) as the interactionfrequency Ai of plant species i versus the summed interaction fre-quency of the remaining plant species in a network (m � Ai). Similarto the model for seed removal rates, we fitted an additional observa-tion-level random effect, because model residuals were overdispersed(v2 = 21152.10, ratio = 174.8, d.f. = 121, P < 0.001). The numberof partners of plants represented a count variable, but was notstrongly skewed. Thus, we fitted a model with Poisson error distribu-tion and a square-root link function for the number of partners. Westandardized the predictor variables to zero mean and unit variance toallow for a comparison of effect sizes. We used the estimates of thethree models to examine under which conditions the plant species ina local community tended to compete for frugivores or tended toenhance each other’s interactions with frugivores. All statistical analy-ses were performed in R (R Development Core Team 2013) using thepackage lme4 for analyses of linear mixed-effects models (Bates et al.2013) and the package bipartite for calculation of species level spe-cialization d0 (Dormann et al. 2009).
Results
The null model analysis indicated that the fruit phenologies ofthe regional plant assemblage were more differentiated thanexpected by chance [var(d/w)observed = 0.23; var(d/w)expected= 0.35 � 0.095, mean � SD; P < 0.01; Fig. 1].Overall, the 20 analysed plant–frugivore networks describe
a total of 5748 visits of 34 frugivore species (30 bird and 4mammal species) on the 15 plant species. During these visits,the frugivores removed an estimated number of 15 766 fruitsand 30 805 seeds, respectively, from the focal plants. Threebird species were the most frequent visitors: Sylvia atricapilla(2352 visits), Turdus merula (1140) and Erithacus rubecula(877). The plant species that attracted most frugivores per18 h were Prunus padus (127 � 115 visits; mean � SD),Sambucus nigra (115 � 97) and Rhamnus cathartica(86 � 143). The fruit density of the plant species in the localnetworks had no effect on the specialization of plants onfrugivores (Table 1). However, as expected, those plant spe-cies with a high phenological overlap shared most of theirfrugivores with other plant species, whereas frugivore assem-blages became more exclusive as phenological differentiationincreased (Table 1; Fig. 3a).
© 2015 The Authors. Journal of Ecology © 2015 British Ecological Society, Journal of Ecology
6 J. Albrecht et al.
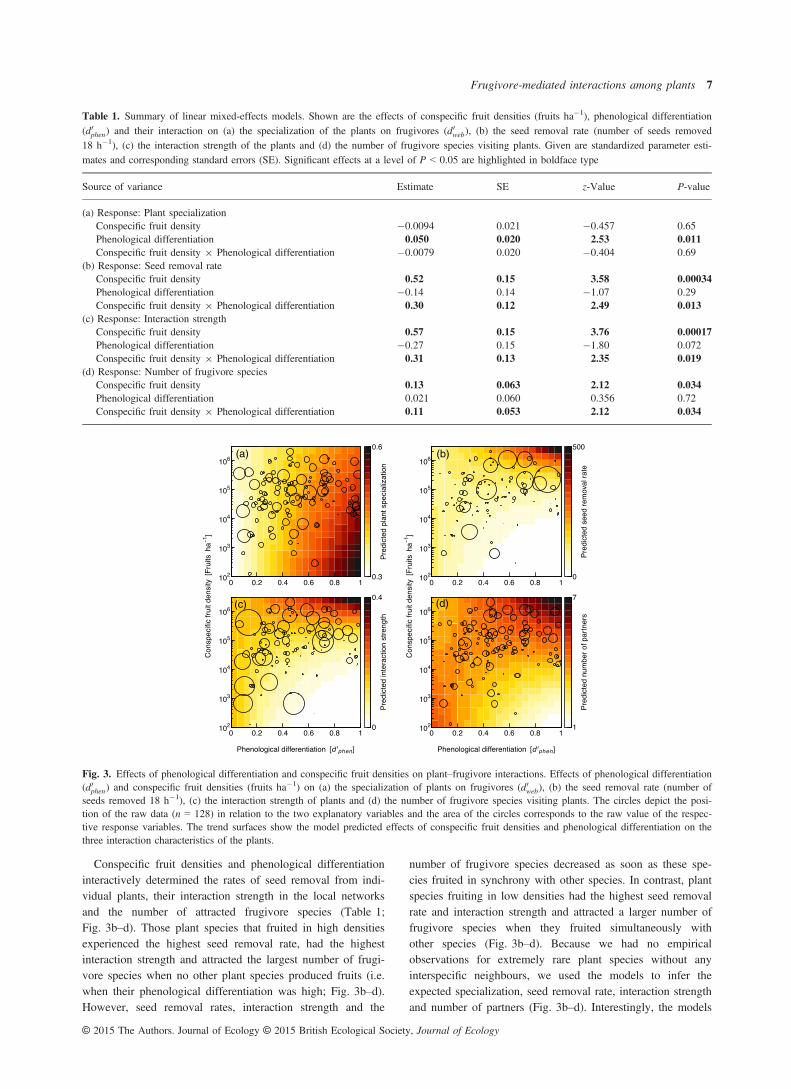
Conspecific fruit densities and phenological differentiationinteractively determined the rates of seed removal from indi-vidual plants, their interaction strength in the local networksand the number of attracted frugivore species (Table 1;Fig. 3b–d). Those plant species that fruited in high densitiesexperienced the highest seed removal rate, had the highestinteraction strength and attracted the largest number of frugi-vore species when no other plant species produced fruits (i.e.when their phenological differentiation was high; Fig. 3b–d).However, seed removal rates, interaction strength and the
number of frugivore species decreased as soon as these spe-cies fruited in synchrony with other species. In contrast, plantspecies fruiting in low densities had the highest seed removalrate and interaction strength and attracted a larger number offrugivore species when they fruited simultaneously withother species (Fig. 3b–d). Because we had no empiricalobservations for extremely rare plant species without anyinterspecific neighbours, we used the models to infer theexpected specialization, seed removal rate, interaction strengthand number of partners (Fig. 3b–d). Interestingly, the models
Table 1. Summary of linear mixed-effects models. Shown are the effects of conspecific fruit densities (fruits ha�1), phenological differentiation(d0phen) and their interaction on (a) the specialization of the plants on frugivores (d0web), (b) the seed removal rate (number of seeds removed18 h�1), (c) the interaction strength of the plants and (d) the number of frugivore species visiting plants. Given are standardized parameter esti-mates and corresponding standard errors (SE). Significant effects at a level of P < 0.05 are highlighted in boldface type
Source of variance Estimate SE z-Value P-value
(a) Response: Plant specializationConspecific fruit density �0.0094 0.021 �0.457 0.65Phenological differentiation 0.050 0.020 2.53 0.011Conspecific fruit density 9 Phenological differentiation �0.0079 0.020 �0.404 0.69
(b) Response: Seed removal rateConspecific fruit density 0.52 0.15 3.58 0.00034Phenological differentiation �0.14 0.14 �1.07 0.29Conspecific fruit density 9 Phenological differentiation 0.30 0.12 2.49 0.013
(c) Response: Interaction strengthConspecific fruit density 0.57 0.15 3.76 0.00017Phenological differentiation �0.27 0.15 �1.80 0.072Conspecific fruit density 9 Phenological differentiation 0.31 0.13 2.35 0.019
(d) Response: Number of frugivore speciesConspecific fruit density 0.13 0.063 2.12 0.034Phenological differentiation 0.021 0.060 0.356 0.72Conspecific fruit density 9 Phenological differentiation 0.11 0.053 2.12 0.034
(a)
(c) (d)
(b)
Fig. 3. Effects of phenological differentiation and conspecific fruit densities on plant–frugivore interactions. Effects of phenological differentiation(d0phen) and conspecific fruit densities (fruits ha�1) on (a) the specialization of plants on frugivores (d0web), (b) the seed removal rate (number ofseeds removed 18 h�1), (c) the interaction strength of plants and (d) the number of frugivore species visiting plants. The circles depict the posi-tion of the raw data (n = 128) in relation to the two explanatory variables and the area of the circles corresponds to the raw value of the respec-tive response variables. The trend surfaces show the model predicted effects of conspecific fruit densities and phenological differentiation on thethree interaction characteristics of the plants.
© 2015 The Authors. Journal of Ecology © 2015 British Ecological Society, Journal of Ecology
Frugivore-mediated interactions among plants 7

indicated that these locally rare species – with a conspecificfruit density of 100 to 1000 fruits per hectare – were unlikelyto attract any frugivores at times when no other plant speciesproduce fruits (Fig. 3b–d).
Discussion
PHENOLOGICAL DIFFERENTIAT ION AND PLANT
SPECIAL IZAT ION ON FRUGIVORES
We found evidence that fruit phenologies of the 15 plant spe-cies in the regional assemblage were more differentiated thanexpected by chance. These results contrast with earlier studiesthat reported random patterns of fruit production (Wheel-wright 1985; Smith-Ramirez, Armesto & Figueroa 1998;Burns 2005) and add to those studies that reported non-ran-dom patterns in fruit production (Poulin et al. 1999; Thies &Kalko 2004). Non-random temporal patterns in the fruit pro-duction of plants can result from various factors, includingabiotic, physiological and phylogenetic constraints, as well asbiotic interactions (MacArthur & Levins 1967; Herrera 2002).Although competition for frugivores might explain the differ-entiation of fruit phenologies among the plant species at theregional scale, we cannot rule out the roles of other factors.Instead, we aimed at going one step further and asked to
which extent competitive and facilitative interactions amongplant species with overlapping phenologies are driven by spatialvariation in their local abundance and co-occurrence. The shar-ing of frugivores is a prerequisite for frugivore-mediated com-petition or facilitation between locally co-occurring plantindividuals (Snow 1965; Carlo & Aukema 2005). We foundthat, at the local scale, plant species whose fruit phenologiesoverlapped were indirectly coupled through shared frugivores.The overlap of frugivore assemblages among co-occurring andsimultaneously fruiting plant species is in line with theoreticalexpectations (V�azquez et al. 2009) and can be attributed to thelow degree of reciprocal specialization in plant–frugivore asso-ciations (Jordano 1987; Fuentes 1995). In the majority of thelocal plant–frugivore assemblages, cofruiting plant species werecoupled through highly generalized frugivore species, such asS. atricapilla, T. merula and E. rubecula (Table S3). A recentstudy suggests that such generalized animal mutualists have amajor impact on community-level dynamics in mutualisticassemblages (Guimar~aes, Jordano & Thompson 2011). Ourresults add to these findings, as they suggest that generalizedfrugivores are probably the main drivers of indirect interactionsbetween co-occurring and cofruiting plant species.
EFFECTS OF CONSPECIF IC FRUIT DENSIT IES AND
PHENOLOGICAL DIFFERENTIAT ION ON PLANT–
FRUGIVORE INTERACTIONS
Under which conditions do co-occurring and cofruiting plantspecies compete for dispersal by shared frugivores or facilitateeach other’s dispersal? Our results show that the outcome ofthese frugivore-mediated plant–plant interactions depended onboth: the conspecific fruiting neighbourhood and on the
degree of phenological differentiation from other fruiting spe-cies. Plant species that occurred in high fruit densities had alow seed removal rate, a low interaction strength and lownumbers of frugivore species when their phenology over-lapped with other fruiting species. These results suggest thathighly abundant cofruiting plant species are likely to competefor frugivores. Conversely, we found positive effects of phe-nological overlap on seed removal rates, interaction strengthand the number of partners of plant species occurring in lowdensities. These results indicate facilitative neighbourhoodeffects on interactions of these rare species.The negative effects of high phenological overlap on inter-
actions of plant species fruiting in high conspecific densitiesmay be explained by frugivore satiation (Herrera et al. 1994;Hampe 2008). Frugivore satiation has already been suggestedas a main driver of mast fruiting strategies in animal-dis-persed plant species, because mast fruiting should minimizeseed loss due to predation by specialized seed eaters (Herreraet al. 1998). At the same time, high conspecific fruit densitiesmay reduce fruit removal by legitimate seed dispersersthrough a dispersion of frugivores into the local fruitingneighbourhood. This would limit the availability of frugivoresto individual plants (Thompson & Willson 1979).The high seed removal rate and interaction strength
observed in plant species fruiting in high densities but lackingother cofruiting species in their neighbourhood may beexplained by the spatial distribution of fleshy-fruited plants inthe Białowie _za forest. In this region, fleshy-fruited plants aremainly associated with ash-alder floodplain forests, whichshow a patchy distribution in the landscape since they requirespecial edaphic conditions (Fali�nski 1994). Thus, fruitresources are highly aggregated, and single species fruitpatches of high density may act as foci for frugivores (Carlo& Morales 2008; Morales et al. 2012). Previous studies haveshown that frugivores track the availability of fruit resourcesover large spatial and temporal scales (Telleria, Ramirez &P�erez-Tris 2008). The interaction strength of plant species fru-iting in high densities may thus increase with phenologicaldifferentiation, because frugivores are likely to concentrate onthese plant species as soon as other species in the local fruit-ing neighbourhood become scarce (Prasad & Sukumar 2010).The most captivating aspect of our study is the finding that,
despite the overall signal of competition among the dominantspecies in the local plant assemblages, plant species fruitingin low densities may actually benefit from interspecific facili-tation (Fig. 3b–d). These results lend support to the hypothe-sis that an attractive fruiting neighbourhood may facilitate thedispersal of less attractive plant species (Carlo, Aukema &Morales 2007). The increase in absolute seed removal mayenhance the quantity of seed dispersal and recruitment, iffrugivores move seeds to suitable microhabitats. Moreover, adiverse frugivore assemblage may increase the quality of dis-persal, if frugivores differ in their movement behaviour or intheir use of microhabitats (Carlo 2005; Jordano et al. 2007;Carlo et al. 2013; Carlo & Tewksbury 2014). Importantly,species that occur in low densities are likely to be dispersal-limited (Carlo & Tewksbury 2014). Thus, our results suggest
© 2015 The Authors. Journal of Ecology © 2015 British Ecological Society, Journal of Ecology
8 J. Albrecht et al.

that the low dispersal capacity of rare plants is enhanced bythe local co-occurrence of other plant species that attractfrugivores.
POTENTIAL CONSEQUENCES FOR NETWORK
STRUCTURE AND COMMUNITY ASSEMBLY PROCESSES
Besides the implications for animal-mediated seed dispersal,our study has important implications for the structure ofplant–animal mutualistic networks and the potential effects offrugivores on community assembly processes.First, our results provide an alternative explanation to the
question why some links in ecological networks remain unob-served even with intense sampling effort (Olesen et al. 2011).To date, these missing links have mainly been attributed tomismatches in species’ phenotypic traits and to spatiotemporalmismatches in the co-occurrence of plants and animal mutual-ists (Jordano, Bascompte & Olesen 2003; Olesen et al. 2011).Our study adds another dimension to these explanations, as itsuggests that unobserved links might not be realized in somesituations because of the specific neighbourhood context inwhich a species occurs.Secondly, our findings offer an ecological mechanism to
explain the nested structure of mutualistic networks, that iswhy specialists tend to interact with generalists (Jordano, Bas-compte & Olesen 2003). Generally, nestedness is supposed toreduce interspecific competition and to enhance the number ofcoexisting species (Bascompte, Jordano & Olesen 2006;Bastolla et al. 2009; but see James, Pitchford & Plank 2012).Simulations predict that a nested network emerges naturally,if new species enter a community where they have minimalcompetitive load (Bastolla et al. 2009; Encinas-Viso, Revilla& Etienne 2012). Along these lines, Olesen et al. (2008)found for arctic plant–pollinator networks that plant speciesentering a community during the course of the season tend tointeract with the most connected pollinator species. Our studyadds to these findings by suggesting that phenological cou-pling of rare and common plant species through shared inter-actions with generalized animal mutualists may facilitate theestablishment and persistence of rare species. This may alsoexplain why rare animal-dispersed plant species typically co-occur with widespread species (L�azaro, Mark & Olesen2005). Literally speaking, rare species may be ‘passengers’that profit from the presence of abundant species, which drivethe movement of frugivores through the landscape (Peterson,Allen & Holling 1998; Carlo, Aukema & Morales 2007).Thirdly, our results imply that the fruiting neighbourhood
may, besides the seed dispersal quantity and quality of rareplant species, also affect the directionality of seed dispersaland community assembly (Carlo, Aukema & Morales 2007).Previous studies suggested that simultaneous fruit productioncan result in the co-dispersal of different plant species and indirectional seed dispersal between these species (Wenny2001; Clark et al. 2004; Carlo & Aukema 2005). Ultimately,this form of directed seed dispersal might contribute to theformation of species associations. We found support for thishypothesis in our data, because the co-occurrence of plant
species across study sites was correlated with the similarity oftheir phenological niches (Procrustes analysis: r = 0.75,P < 0.01; Appendix S3). Overall, the results of our studyindicate that the co-dispersal of plant species with similarphenological niches might, in addition to other abiotic andbiotic factors, influence plant community assembly processes.
Conclusions
Our study highlights that variation in neighbourhood contextis an important determinant of the outcome of indirect frugi-vore-mediated interactions among co-occurring plant species.While abundant cofruiting plant species competed for interac-tions with frugivores, the interactions of rare plants wereenhanced by phenological overlap with other fruiting species.These results indicate that indirect interactions among plantsthat share seed dispersers may consistently promote the estab-lishment and persistence of rare species through facilitation.Importantly, the sharing of seed dispersers among cofruitingplant species was also reflected in their co-occurrence. Thissuggests that, besides other factors, indirect coupling of spe-cies through shared mutualistic partners might be an impor-tant determinant of plant community assembly processes. Theindirect coupling through shared mutualists may drive the for-mation of associations among co-dispersed plant species andmight contribute to species coexistence in plant–animal mutu-alistic communities.
Acknowledgements
T. A. Carlo and an anonymous referees provided insightful comments, whichimproved the manuscript. We greatly thank the administration of theBiałowie _za National Park, the forestry administrations of Białowie _za, Hajn�owkaand Browsk, and Polish authorities (Ministry of Environment, GDOS [PolishGeneral Directorate of Environment Conservation, Warsaw] and RDOS [Regio-nal Directorate of Environment Conservation, Białystok]) for the permissions towork in Białowie _za Forest. We are indebted to the following volunteer ornithol-ogists who assisted with frugivore observations and without whom realizationof fieldwork would not have been possible: F. Bunsen, A. Fritzsch, C. Gayer,I. Grass, N. Gr€on, J. Hagge, J. Hennlein, C. Herche, M. Hoffmann, C. H€ofs, L.Jeske, M. Jung, S. Kaack, E. Kubitz, I. M€oller and M. P€onichen. N.F. andD.G.B. are supported by the Robert Bosch Foundation. This study is part ofthe PhD thesis of J.A. at the University of Marburg. The project is funded bythe German Federal Foundation for Environment (DBU) in the framework of aPhD scholarship. N.S. is supported by funds of the Institute of Nature Conser-vation. The authors have no conflict of interest to declare.
Data accessibility
Data from this study are available from the Dryad Digital Repository:http://dx.doi.org/10.5061/dryad.s1b7q (Albrecht et al. 2015).
References
Albrecht, J., Neuschulz, E.L. & Farwig, N. (2012) Impact of habitat structureand fruit abundance on avian seed dispersal and fruit predation. Basic andApplied Ecology, 13, 347–354.
Albrecht, J., Berens, D.G., Bl€uthgen, N., Jaroszewicz, B., Selva, N. & Farwig, N.(2013) Logging and forest edges reduce redundancy in plant–frugivore net-works in an old-growth European forest. Journal of Ecology, 101, 990–999.
Albrecht, J., Berens, D.G., Jaroszewicz, B., Selva, N., Brandl, R. & Farwig, N.(2014) Correlated loss of ecosystem services in coupled mutualistic networks.Nature Communications, 5, 3810.
© 2015 The Authors. Journal of Ecology © 2015 British Ecological Society, Journal of Ecology
Frugivore-mediated interactions among plants 9

Albrecht, J., Bohle, V., Berens, D.G., Jaroszewicz, B., Selva, N. & Farwig, N.(2015) Data from: variation in neighbourhood context shapes frugivore-medi-ated facilitation and competition among co-dispersed plant species. DryadDigital Repository, doi:10.5061/dryad.s1b7q.
Ashton, P.S., Givnish, T.J. & Appanah, S. (1988) Staggered flowering in theDipterocarpaceae: new insights into floral induction and the evolution ofmast fruiting in the aseasonal tropics. The American Naturalist, 132, 44–66.
Bascompte, J., Jordano, P. & Olesen, J.M.M. (2006) Asymmetric coevolution-ary networks facilitate biodiversity maintenance. Science, 312, 431–433.
Bastolla, U., Fortuna, M.A., Pascual-Garcia, A., Ferrera, A., Luque, B. & Bas-compte, J. (2009) The architecture of mutualistic networks minimizes compe-tition and increases biodiversity. Nature, 458, 1018–1021.
Bates, D., Maechler, M., Bolker, B. & Walker, S. (2013) lme4: Linear Mixed-Effects Models Using Eigen and S4. Available at http://CRAN.R-project.org/package=lme4.
Bl€uthgen, N. (2010) Why network analysis is often disconnected from commu-nity ecology: a critique and an ecologist’s guide. Basic and Applied Ecology,11, 185–195.
Bl€uthgen, N., Menzel, F. & Bl€uthgen, N. (2006) Measuring specialization inspecies interaction networks. BMC Ecology, 6, 9–20.
Bl€uthgen, N., Fr€und, J., V�azquez, D.P. & Menzel, F. (2008) What do interac-tion network metrics tell us about specialization and biological traits? Ecol-ogy, 89, 3387–3399.
Bobiec, A. (2002) Living stands and dead wood in the Białowieza forest: sug-gestions for restoration management. Forest Ecology and Management, 165,125–140.
Bobiec, A. (2012) Białowieza Primeval Forest as a remnant of culturally modi-fied ancient forest. European Journal of Forest Research, 131, 1269–1285.
Bobiec, A., van der Burgt, H., Meijer, K., Zuyderduyn, C., Haga, J. & Vlaand-eren, B. (2000) Rich deciduous forests in Białowieza as a dynamic mosaic ofdevelopmental phases: premises for nature conservation and restoration man-agement. Forest Ecology and Management, 130, 159–175.
Breslow, N.E. (1984) Extra-poisson variation in log-linear models. Applied Sta-tistics, 33, 38–44.
Burns, K.C. (2002) Seed dispersal facilitation and geographic consistency inbird-fruit abundance patterns. Global Ecology and Biogeography, 11, 253–259.
Burns, K.C. (2005) Is there limiting similarity in the phenology of fleshy fruits?Journal of Vegetation Science, 16, 617–624.
Carlo, T.A. (2005) Interspecific neighbors change seed dispersal pattern of anavian-dispersed plant. Ecology, 86, 2440–2449.
Carlo, T.A. & Aukema, J.E. (2005) Female-directed dispersal and facilitationbetween a tropical mistletoe and a dioecious host. Ecology, 86, 3245–3251.
Carlo, T.A., Aukema, J.E. & Morales, J.M. (2007) Plant–frugivore interactionsas spatially explicit networks: Integrating frugivore foraging with fruitingplant spatial patterns. Seed Dispersal: Theory and its Application in aChanging World (eds A.J. Dennis, E.W. Schupp, R.J. Green & D.A. West-cott), pp. 369–390. CAB International, Oxfordshire, UK.
Carlo, T.A., Collazo, J.A. & Groom, M.J. (2003) Avian fruit preferences acrossa Puerto Rican forested landscape: pattern consistency and implications forseed removal. Oecologia, 134, 119–131.
Carlo, T.A. & Morales, J.M. (2008) Inequalities in fruit-removal and seed dis-persal: consequences of bird behaviour, neighbourhood density and landscapeaggregation. Journal of Ecology, 96, 609–618.
Carlo, T.A. & Tewksbury, J.J. (2014) Directness and tempo of avian seed dis-persal increases emergence of wild chiltepins in desert grasslands. Journal ofEcology, 102, 248–255.
Carlo, T.A., Garc�ıa, D., Mart�ınez, D., Gleditsch, J.M. & Morales, J.M. (2013)Where do seeds go when they go far? Distance and directionality of avianseed dispersal in heterogeneous landscapes. Ecology, 94, 301–307.
Carnicer, J., Jordano, P. & Melian, C.J. (2009) The temporal dynamics ofresource use by frugivorous birds: a network approach. Ecology, 90, 1958–1970.
Clark, C.J., Poulsen, J.R., Connor, E.F. & Parker, V.T. (2004) Fruiting trees asdispersal foci in a semi-deciduous tropical forest. Oecologia, 139, 66–75.
Dormann, C.F., Fr€und, J., Bl€uthgen, N. & Gruber, B. (2009) Indices, graphsand null models: analyzing bipartite ecological networks. The Open EcologyJournal, 2, 7–24.
Encinas-Viso, F., Revilla, T.A. & Etienne, R.S. (2012) Phenology drives mutu-alistic network structure and diversity. Ecology Letters, 15, 198–208.
Fali�nski, J.B. (1994) Concise geobotanical atlas of Białowieza forest. Phytocoe-nosis, 6, 3–34.
Farwig, N., B€ohning-Gaese, K. & Bleher, B. (2006) Enhanced seed dispersal ofPrunus africana in fragmented and disturbed forests? Oecologia, 147, 238–252.
Fuentes, M. (1995) How specialized are fruit–bird interactions? Overlap of fru-givore assemblages within and between plant species. Oikos, 74, 324–330.
Guimar~aes, P.R., Jordano, P. & Thompson, J.N. (2011) Evolution and coevolu-tion in mutualistic networks. Ecology Letters, 14, 877–885.
Hampe, A. (2008) Fruit tracking, frugivore satiation, and their consequencesfor seed dispersal. Oecologia, 156, 137–145.
Herrera, C.M. (1984) A study of avian frugivores, bird-dispersed plants, and theirinteraction in Mediterranean scrublands. Ecological Monographs, 54, 1–23.
Herrera, C.M. (1985) Determinants of plant–animal coevolution – the case ofmutualistic dispersal of seeds by vertebrates. Oikos, 44, 132–141.
Herrera, C.M. (1995) Plant-vertebrate seed dispersal systems in the Mediterra-nean – ecological, evolutionary, and historical determinants. Annual Reviewof Ecology and Systematics, 26, 705–727.
Herrera, C.M. (1998) Long-term dynamics of Mediterranean frugivorous birdsand fleshy fruits: a 12-year study. Ecological Monographs, 68, 511–538.
Herrera, C.M. (2002) Seed dispersal by vertebrates. Plant–Animal Interactions:An Evolutionary Approach (eds C.M. Herrera & O. Pellmyr), pp. 185–208.Blackwell, Oxford, UK.
Herrera, C.M., Jordano, P., Lopezsoria, L. & Amat, J.A. (1994) Recruitment ofa mast-fruiting, bird-dispersed tree – bridging frugivore activity and seedlingestablishment. Ecological Monographs, 64, 315–344.
Herrera, C.M., Jordano, P., Guitian, J. & Traveset, A. (1998) Annual variabilityin seed production by woody plants and the masting concept: reassessmentof principles and relationship to pollination and seed dispersal. The AmericanNaturalist, 152, 576–594.
Hinde, J. (1982) Compound poisson regression models. GLIM 82: Proceedingsof the International Conference on Generalised Linear Models, vol. 14 (ed.R. Gilchrist), pp. 109–121. Springer, New York, USA.
Howe, H.F. & Smallwood, J. (1982) Ecology of seed dispersal. Annual Reviewof Ecology and Systematics, 13, 201–228.
James, A., Pitchford, J.W. & Plank, M.J. (2012) Disentangling nestedness frommodels of ecological complexity. Nature, 487, 227–230.
Janzen, D.H. (1970) Herbivores and the number of tree species in tropical for-ests. The American Naturalist, 104, 501–528.
Jordano, P. (1987) Patterns of mutualistic interactions in pollination and seeddispersal: connectance, dependence asymmetries, and coevolution. The Amer-ican Naturalist, 129, 657–677.
Jordano, P., Bascompte, J. & Olesen, J.M. (2003) Invariant properties in coevo-lutionary networks of plant–animal interactions. Ecology Letters, 6, 69–81.
Jordano, P., Garcia, C., Godoy, J. & Garcia-Casta~no, J. (2007) Differential con-tribution of frugivores to complex seed dispersal patterns. Proceedings of theNational Academy of Sciences of the United States of America, 104, 3278–3282.
Kamm, U., Rotach, P., Gugerli, F. & Siroky, M. (2009) Frequent long-distancegene flow in a rare temperate forest tree (Sorbus domestica) at the landscapescale. Heredity, 103, 476–482.
L�azaro, A., Mark, S. & Olesen, J.M. (2005) Bird-made fruit orchards in north-ern Europe: nestedness and network properties. Oikos, 110, 321–329.
MacArthur, R.H. & Levins, R. (1967) The limiting similarity, convergence, anddivergence of coexisting species. The American Naturalist, 101, 377–385.
Matuszkiewicz, W. (2001) Przewodnik Do Oznaczania Zbiorowisk RoolinnychPolski [Guide for Determination of Polish Plant Associations]. Wydawnic-two Naukowe PWN, Warszawa, Poland.
Menke, S., B€ohning-Gaese, K. & Schleuning, M. (2012) Plant–frugivore net-works are less specialized and more robust at forest-farmland edges than inthe interior of a tropical forest. Oikos, 121, 1553–1566.
Morales, J.M., Rivarola, M.D., Amico, G. & Carlo, T.A. (2012) Neighborhoodeffects on seed dispersal by frugivores: testing theory with a mistletoe–mar-supial system in Patagonia. Ecology, 93, 741–748.
Nathan, R. & Muller-Landau, H.C. (2000) Spatial patterns of seed dispersal,their determinants and consequences for recruitment. Trends in Ecology &Evolution, 15, 278–285.
Olesen, J.M., Bascompte, J., Elberling, H. & Jordano, P. (2008) Temporaldynamics in a pollination network. Ecology, 89, 1573–1582.
Olesen, J.M., Bascompte, J., Dupont, Y.L., Elberling, H., Rasmussen, C. &Jordano, P. (2011) Missing and forbidden links in mutualistic networks. Pro-ceedings of the Royal Society of London, Series B: Biological Sciences, 278,725–732.
Peterson, G., Allen, C.R. & Holling, C.S. (1998) Ecological resilience, biodi-versity, and scale. Ecosystems, 1, 6–18.
Poole, R.W. & Rathcke, B.J. (1979) Regularity, randomness, and aggregationin flowering phenologies. Science (New York, N.Y.), 203, 470–471.
Poulin, B., Wright, S.J., Lefebvre, G. & Calder�on, O. (1999) Interspecific syn-chrony and asynchrony in the fruiting phenologies of congeneric bird-dis-persed plants in Panama. Journal of Tropical Ecology, 15, 213–227.
© 2015 The Authors. Journal of Ecology © 2015 British Ecological Society, Journal of Ecology
10 J. Albrecht et al.

Prasad, S. & Sukumar, R. (2010) Context-dependency of a complex fruit-frugi-vore mutualism: temporal variation in crop size and neighborhood effects.Oikos, 119, 514–523.
R Development Core Team. (2013) R: A Language and Environment for Statis-tical Computing. Available at http://www.R-project.org/.
Rathcke, B. & Lacey, E.P. (1985) Phenological patterns of terrestrial plants.Annual Review of Ecology and Systematics, 16, 179–214.
Schleuning, M., Bl€uthgen, N., Fl€orchinger, M., Braun, J., Schaefer, H.M. &B€ohning-Gaese, K. (2011) Specialization and interaction strength in a tropi-cal plant–frugivore network differ among forest strata. Ecology, 92, 26–36.
Schupp, E.W., Jordano, P. & Gomez, J.M. (2010) Seed dispersal effectivenessrevisited: a conceptual review. New Phytologist, 188, 333–353.
Smith-Ramirez, C., Armesto, J.J. & Figueroa, J. (1998) Flowering, fruiting andseed germination in Chilean rain forest myrtaceae: ecological and phyloge-netic constraints. Plant Ecology, 136, 119–131.
Snow, D.W. (1965) A possible selective factor in the evolution of fruiting sea-sons in tropical forest. Oikos, 15, 274–281.
Sokołowski, A. (2004) Lasy Puszczy Białowieskiej [The Forests of the Białowi-eza Forest]. Centrum Informacyjne Lasow Panstwowych, Warszawa, Poland.
Taper, M.L. & Chase, T.J. (1985) Quantitative genetic models for the coevolu-tion of character displacement. Ecology, 66, 355.
Telleria, J.L., Ramirez, A. & P�erez-Tris, J. (2008) Fruit tracking between sitesand years by birds in Mediterranean wintering grounds. Ecography, 31, 381–388.
Thies, W. & Kalko, E.K.V. (2004) Phenology of neotropical pepper plants(Piperaceae) and their association with their main dispersers, two short-tailedfruit bats, Carollia perspicillata and C. castanea (Phyllostomidae). Oikos, 104,362–376.
Thompson, J.N. (2005) The Geographic Mosaic of Coevolution. UniversityChicago Press, Chicago, Illinois.
Thompson, J.N. & Willson, M.F. (1979) Evolution of temperate fruit/bird inter-actions: phenological strategies. Evolution, 33, 973–982.
V�azquez, D.P., Melian, C.J., Williams, N.M., Bl€uthgen, N., Krasnov, B.R. &Poulin, R. (2007) Species abundance and asymmetric interaction strength inecological networks. Oikos, 116, 1120–1127.
V�azquez, D.P., Bl€uthgen, N., Cagnolo, L. & Chacoff, N.P. (2009) Uniting pat-tern and process in plant–animal mutualistic networks: a review. Annals ofBotany, 103, 1445–1457.
Webb, C.O. & Peart, D.R. (2001) High seed dispersal rates in faunally intacttropical rain forest: theoretical and conservation implications. Ecology Let-ters, 4, 491–499.
Wenny, D.G. (2001) Advantages of seed dispersal: a re-evaluation of directeddispersal. Evolutionary Ecology Research, 3, 51–74.
Wheelwright, N.T. (1985) Competition for dispersers, and the timing of flower-ing and fruiting in a guild of tropical trees. Oikos, 44, 465–477.
Whelan, C.J., Schmidt, K.A., Steele, B.B., Quinn, W.J. & Dilger, S. (1998)Are bird-consumed fruits complementary resources? Oikos, 83, 195–205.
Williams, D.A. (1982) Extra-binomial variation in logistic linear models.Applied Statistics, 31, 144–148.
Received 9 April 2014; accepted 15 January 2015Handling Editor: Mark Rees
Supporting Information
Additional Supporting Information may be found in the online ver-sion of this article:
Appendix S1. Properties of the phenological niche.
Appendix S2. Standardized Kullback–Leibler divergence as a mea-sure of phenological niche differentiation.
Appendix S3. Correlation between the co-occurrence and the pheno-logical niche similarity of the plant species.
Figure S1. Properties that qualify the phenological niche of speciesin the context of other species.
Figure S2. Range of the empirical data for relative fruit abundanceand relative length of the phenology of the focal plants in the dataset.
Figure S3. Results of the simulation study showing the behaviour ofthe standardized Kullback–Leibler divergence d0 as a measure of phe-nological niche differentiation.
Table S1. Summary of the geographic coordinates and the character-istics of the study sites.
Table S2. Fruit traits, growth form and mean density of the 15 plantspecies included in the study.
Table S3. List of frugivore species with their taxonomic affiliations,vernacular names and total number of recorded visits to plants duringthe study.
Table S4. Summary of sampling of seed disperser assemblages onthe 15 plant species across the study sites and the 2 study years.
Table S5. Summary of the number of crop size estimates that wereobtained in the course of the fruiting season during the 2 study years.
© 2015 The Authors. Journal of Ecology © 2015 British Ecological Society, Journal of Ecology
Frugivore-mediated interactions among plants 11