\u003ctitle\u003emicrobial remains in middle proterozoic rocks of northern...
TRANSCRIPT

Microbial remains in Middle Proterozoic rocks of Northern Australia
Marina Astafievaa, Richard B. Hooverb, Alexei Yu. Rozanova, P. Vickers-Richc, A. Wildec
aPaleontological Institute of Russian Academy of Sciences, 117647 Profsoyusnaya 123, Moscow, Russia; bAstrobiology Laboratory, NASA/NSSTC, Huntsville, Alabama, USA 35802;
cSchool of Geosciences, Monash University Clayton, Victoria 3800 Australia
ABSTRACT
Investigation of the samples of the McArthur River complex ore deposit, one of the most zinc-lead mineral province in the world, brings us to conclusion about possibility of biogenic origin of sulfides in McArthur River ore deposit and to make supposition about formation of studied rocks in the photic zone of the sea.
Key words: cocci, sulphate reduction, cyanobacteria, purple bacteria, sulfides, photic zone.
The McArthur River ore deposit, one of the most significant zinc-lead deposits in the world, is situated in the southeastern part of Northern territory of Australia. McArthur River ore deposit is one of the most well known examples of stratiform Zn-Pb-A ore deposit. The fine-grained sphalerite-galena bands are intricately inter-layered both with pelagic organic-rich beds and, as it is considered, with thin turbidite beds. The previous investigations in regional sedimentology and geochemistry of Barney Creek Formation gave us opportunity to suppose that the host rocks to the ore deposits, HYC Pyritic Shale, represent a period of water deepening. Water depth was probably in excess of several hundred meters, with the deepest water euxinic facies corresponding to the ore position at he base of the HYC Pyritic Shale.
It is supposed that mineral brines periodically introduced to the basin in connection with microseismic activity (that follows every significant seismic event in the basin) were most probably sulfate-rich (SO4
2- > H2S) and capable of transporting at moderate temperatures (150oC - 250oC), and near-neutral pH (Large et al. 1998).
But the data obtained as a result of preliminary investigation of these rocks from the bacterial paleontology point of view give opportunity to suppose a little bit different conditions of formation of the rocks under investigation – under much lower temperatures and much less depth (within the limits of photic zone).
There were 3 samples at our disposal that were studied under the scanning electron microscope (CamScan-4) with microanalyser.
In chemical composition of rocks under investigation iron and zinc predominate together with silicon and aluminum. Sulfur also are met rather often, barium and potash – sometimes, lead is rare.
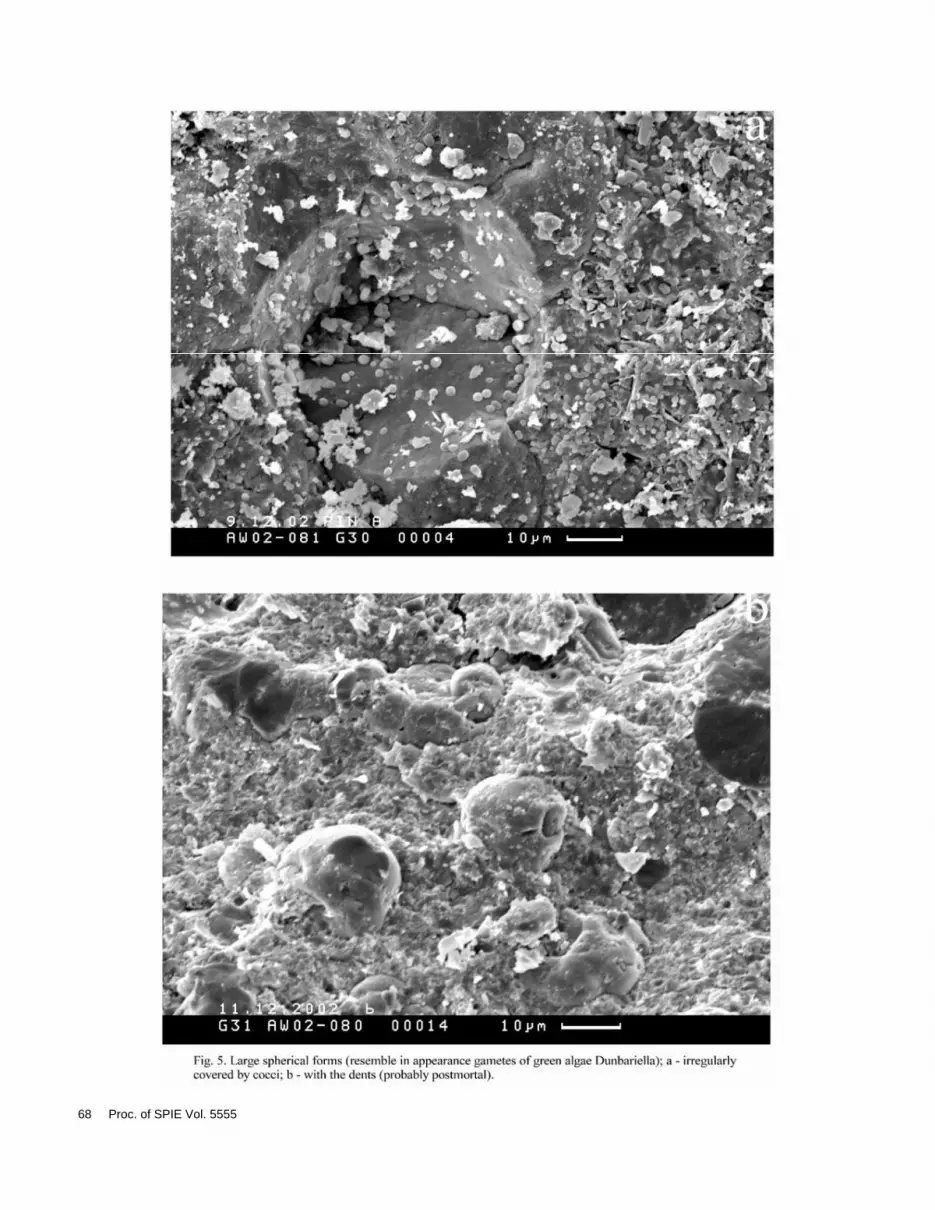
All biomorphic microstructures that we have met could be divided into several groups. At first there are coccoidal structures (Figs. 1, 2). Cocci are met both in accumulations and single. Besides peculiar structures represented probably combination of coccoidal structures with acicular crystals are met (Fig. 3). Secondly there are filament structures (thread-like) and rod-shaped structures (Fig. 4). Large spherical forms (Fig. 5), biofilms, glycocalyx etc. and scattering crystallites of ancient framboids (Fig. 6) are present.
The distribution of the discovered biomorphic structures through the samples could be represented by following way:
Instruments, Methods, and Missions for Astrobiology VIII, edited byRichard B. Hoover, Gilbert V. Levin, Alexei Y. Rozanov, Proc. of SPIE Vol. 5555(SPIE, Bellingham, WA, 2004) · 0277-786X/04/$15 · doi: 10.1117/12.558919
59

AW 02-079 In this sample biomorphic structures are the richest and most diverse, practically all morphological modifications found in samples under investigation are met in this sample: cocci (0.5-6.0 ), combination of cocci and microcrystals, “acritarchs”, biofilms.
AW 02-080 Large spherical (30 ) forms, rod-shaped and thread-like (filament) structures, biofilms, cocci.
AW 02-081 Cocci, large spherical (50-60 ) structures, filament and rod-shaped structures, combination of cocci and microcrystals, biofilms.
As a rule, coccoidal forms with diameter about 0.5-6.0 predominate in samples (Figs. 1, 2). Cocci could be met in a single position, in a dispersed form (single cocci as if ingrained in the rock) and in accumulations (colonies). Often investigated cocci morphologically are practically indistinguishable from purple sulfur bacteria of genus Thiocapsa (consultation of L. M. Gerasimenko). In rare cases there is an impression that the rock practically consists of coccoidal forms.
Peculiar structures composed of cocci surrounded or covered by microcrystals represented interest (Fig. 3). These structures are represented by biomorphic coccoidal microstructure in the center and as if covered by microneedles (microcrystals) and barium dominates the chemical composition. Barium is a component of witherite (BaCO3) and barite (Ba(SO4)). Both minerals are companions of galena (PbS) and ores containing silver. As McArthur River deposit is represented by Pb-Zn-Ag containing ores, the findings of these minerals of barium in our samples cannot arise astonishment.
It is seen on the figures both coccoidal forms surrounded by needles-extensions and a more complicated combination of biomorphic microstructures – the combination of coccoidal and rod-shaped forms with microcrystals could be seen.
Coccoidal biomorphic microstructures (diameter about 6 having openings with uneven edges are also encountered. Such structures are possible to be considered to be acritarchs on the recent stage of investigation.
In samples AW 02-080 and AW 02-081 filament structures with diameter about 1 are found. There are no such structures in sample AW 02-079. In this sample the mark of larger filament form (diameter about 8 ) is noted (fig. 4).
In samples AW 02-080 and AW 02-081 large spherical forms with diameter 30-70 , covered by coccoidal structures, with diameter about 1 were discovered (Fig. 5). These biomorphic structures resemble in appearance gametes of green algae genus Dunaliella (D. salinae) from Sivash (Gerasimenko, Zavarzin, 1993), which is in our opinion these structures could be considered as eukaryotes. As a rule these forms are irregularly covered by cocci (about 1-2 in size). It cannot be excluded that algae gametes were used by cocci as a source of carbon. Sometimes there are dents (about 5 in diameter) on gametes, but in this case the possibility of postmortal deformation of gamete is great. It is obvious that gamete inhabited in the water mass and sank to the bottom only after their death (postmortal).
Besides described above biomorphic structures represented by iron and zinc sulfides crystals of “pyrite”, more precisely iron sulfide, are met. Unfortunately there is no opportunity to speak about mineral composition of rocks. Such crystals often resemble scattering framboidal crystallites (Fig. 6), though the framboids by themselves were not noted. But sizes of founded crystallites a little bit differ from classic ones. Studied earlier framboids had typical sizes up to 10 with crystallites less than 1 (Astafieva, 2004). The crystallites encountered in Australian rocks are larger, there size is about 1.5-2.0 in general but can reach 3 , that is the crystallite sizes in the studied samples can exceed typical sizes by up to order of magnitude. Unfortunately preservation of all studied objects (pyrite crystals included) could be better. As a rule, the crystallites studied in this work are octahedral in form. In some cases, it could be possible to suggest cubic form of crystallites but
60 Proc. of SPIE Vol. 5555

under increasing magnification the crystallites that we consider to be cubic turned to be microscopic section of octahedron crystallites.
It is necessary to say that the absence of clear framboidal structures typical for biogenic iron sulfides in studied samples put us a little on our guard. Probably framboidal structures formed simultaneously with sedimentation have been destroyed with time. The larger sizes of our crystallites could be due to specific conditions of their formation. So in some authors opinion (Kizilshtein, Minaeva, 1972), crystallization of less saturated brine leads to formation of comparatively larger crystal structures.
It should be mentioned that the framboidal pyrite formation usually takes place in the sediment on the stage of early diagenesis (Berner, 1984; Berner, Westrich, 1985). But its formation is possible also in water mass, for example in the Black sea, fjord Framvaren (Norway) and estuary of Pettaquamscutt river. Some authors considered that framboidal pyrite formation in water mass is connected with the region adjacent to (or below) the boundary between aerobic and anaerobic zones, with a high degree of supersaturation with respect to pyrite and a lesser degree of supersaturation with respect to iron monosulfides (Wilkin, Barnes, 1997b).
Biogenic formation of metal sulfides, including iron sulfides, (hosted rocks of ore deposits of the McArthur River deposit are represented exactly by pyretic shales), as a rule is limited by concentration of organic carbon available for metabolism, because in marine sediments iron and sulfate of marine waters usually are rather rich (Gorlenko et al., 1977).
It should be said that necessary for pyrite formation soluble sulfates represent organic matter, “destroyed” by sulfate-reduction, and soluble iron – by pore waters of sediment (Raiswell et al., 1993). Besides in the opinion of these authors the connection between pyrite formations with pore waters rather close to boarder sediment/water is clear (so as to free diffusive connection with superincumbent water be supported) (Raiswell et al., 1993).
As it is known, the activity of sulfate-reducing bacteria (SRB) is more active in the dark time of day, and microbial anaerobic sulfide oxidation is more intensive in light (Visscher et al., 1998). Consequently pyrite formation is connected with daily variation of illumination that is deposition of iron sulfides is possible only in dark time of day, when the consumption of H2S by purple bacteria is stopped. During time when light is present, hydrogen sulfide is used by the phototrophic purple bacteria.
It is necessary to mention that biogenic framboids not only pyrite but also black jack (sphalerite) (ZnS) are known (Bawden et al., 2003).
Crystallites of pyritohedron (pentagon dodecahedron) were found, but their sizes are so large – 5-6 and more that it is most likely in this case it was said about real pyrite crystals (not about component of framboids crystallites).
We consider all of the biomorphic structures described above to be formed in situ, that is simultaneously with sedimentation. But along with “rock-forming” biomorphic structures we found microstructures, which we consider to be subsequent contamination, probably actinomycetes and algae.
CONCLUSIONS
The deposition of heavy metal (primarily iron) sulfides is due to sulfate-reduction. In silts and dark anaerobic zone hydrogen sulfide is connected in insoluble iron sulfides, deposits and goes away from sulfur circulation (Gorlenko et al., 1977). In the case of sufficient illumination the main consumers of hydrogen sulfide – phototrophic purple bacteria. As it was mentioned, the coccoidal structures found by us morphologically look like purple bacteria of genus Thiocapsa. The specifics of our material do not allow us to speak with confidence about their identity, but the possibility of it is rather high. Under deficit of hydrogen, sulfide the main product of oxidation of hydrogen sulfide are sulfates, sulfur does not accumulate. The formation of intermediate sulfur inside cells of purple bacteria happens if the rate of hydrogen sulfide formation or its coming in exceeds the rate
Proc. of SPIE Vol. 5555 61

of its oxidation by microorganisms (Gorlenko et al., 1977). The vital activity of cyanobacteria (including those found by us to be possible analogs of Microcoleus) also is connected with photic zone.
So it is possible to suppose periodical variation of conditions of sedimentation (photic zone – dark zone), because the deposition of sulfides during dark time of the day, it seems, is not enough for formation of complex metal deposits of industrial value. In our opinion it is bacterial sulfate-reduction that mainly provides the formation of hydrogen sulfide, which in its turn is connected in metal sulfides.
It could be proposed seasonal variations of illumination of water masses connected with the presence or absence of ice cover. The presence of dark zone could be connected not only with the depth of basin but also with the turbidity of the water, the presence of great amount of suspended particles and so on (and even to less extent with the climatic peculiarities – fewer sunny days, etc).
Besides under laboratory conditions the total dependence of form and sizes of framboids from temperature was revealed – with the increasing of temperature the dimensions and circularity of aggregates are increasing, aggregates formed under lower temperatures have smaller sizes (Sunagawa et al., 1971). But these conclusions based upon investigation of formation of framboidal pyrite under temperatures from 200o to 300oC and under pressure from 15 to 85 atmospheres, which is under conditions significantly different from the conditions under which framboidal pyrite forms in nature.
Therefore our investigation of these tells us about the possibility of the biogenic formation of sulfides of the McArthur River deposit and although it is a little bit too early to speak confidently about the mechanisms of the formation of the sulfides of this deposit, it is possible to suggest that investigated rocks formed in the photic zone of the sea (that is depth most likely was not more than 200 m) with alternation of dark and light conditions.
ACKNOWLEDGMENTS
The authors are grateful to everybody who has promoted the carrying out of this work, especially to L. M. Gerasimenko for her contribution in discussion of the manuscript and definition of bacterial structures; the authors are also grateful to G. T.Ushatinskaja for the discussion of material; A. V. Kravtsev and L. T. Protasevich for their assistance in electron microscope investigation; and to everybody who has given the material at our disposal.
This study was fulfilled under Program of RAS Presidium “Origin and evolution of biosphere”, subprogram II and supported by the Russian Foundation for Basic Research (SS – 974.2003.5; projects 02-04-48094 and 03-05-64499).
REFERENCES
1. Bawden Th.M., Einaudi M.T., Bostick B.C., Meibom A., Wooden J., Norby J.W., Orobona M.J.T., Chamberlain C.P. “Extreme 34S depletions in ZnS at the Mike gold deposit, Carlin Trend, Nevada: Evidence for bacteriogenic supergene sphalerite”, Geology, 31, 913-916, 2003.
2. Berner R.A., «Sedimentary pyrite formation: An update», Geochim. et Cosmochim. Acta, 48, 605-615, 1984. 3. Berner R.A., Westrich J.T., «Bioturbation and the Early Diagenesis of Carbon and Sulfur», Amer. Journ.,
Science, 285, 193-206, 1985. 4. Fortin D., Ferris F.G., Beveridge F.G. “Surface-mediated Mineral Development by Bacteria, Reviews in
Mineralogy”, Geomicrobiology: Interactions between Microbes and Minerals, Eds.:Banfield J.F., Nealson K.H.P., 35, 161-180, 1997.
5. Gerasimenko L.M., Zavarzin G.A., “The relict cyanobacterial communities”, The problems of pre-anthropogenous evolution of the biosphere, 222-253, Nauka, Moscow, 1993.
6. Gorlenko V.M., Dubinina G.A., Kusnezov S.I., Ecology of water microorganisms, 288 p., Nauka, Moscow, 1977.
7. Gromov B.V., Pavlenko G.V., Ecology of bacteria, 248 p, Leningrad University Press, Leningrad, 1989. 8. Kizilshtein L.J., Minaeva L.G., “Origin of framboidal forms of pyrite”, Reports of Ac. Sci. of the USSR, 206, no
5, 1187-1189.
62 Proc. of SPIE Vol. 5555

9. Large R.R., Bull S.W., Cooke D.R., Goldrick P.J., “A genetic model for the HYC deposit, Australia: based on regional sedimentology, geochemistry, and sulfide-sediment relationships”, Economic Geology, 93, 1345-1368, 1998.
10. Murowchic J.B., Barnes H.L., “Effects of temperature and degree of supersaturation on pyrite morphology”, Amer. Mineralogist, 72, 1241-1250, 1987.
11. Raiswell R., Whaler K., Dean S., Coleman M.L., Briggs D.E.G. “A simple three-dimensional model of diffusion-with-precipitation applied to localised pyrite formation in framboids, fossils and detrital iron minerals”, Marine Geology, 113, 89-100, 1993.
12. Sunagawa I., Endo Y., Nakai N., «Hydrothermal synthesis of framboidal pyrite», Soc. Mining Geol. Japan, Spec. Issue, 2, 10-14.
13. Visscher P.T., Reid R.P., Bebout B.M., et al. “Formation of lithified micritic laminae in modern marine stromatolities (Bahamas): the role of sulfur cycling”, Amer. Mineralogist, 83, 1482-1493, 1998.
Proc. of SPIE Vol. 5555 63

64 Proc. of SPIE Vol. 5555

Proc. of SPIE Vol. 5555 65

66 Proc. of SPIE Vol. 5555

Proc. of SPIE Vol. 5555 67
68 Proc. of SPIE Vol. 5555

Proc. of SPIE Vol. 5555 69