two forms of sustained xanthophyll cycle-dependent energy dissipation in overwintering euonymus...
TRANSCRIPT

ABSTRACT
Seasonal differences in PSII efficiency (Fv/Fm), the conver-sion state of the xanthophyll cycle (Z + A)/(V + A + Z), andleaf adenylate status were investigated in Euonymuskiautschovicus. On very cold days in winter, Fv/Fm assesseddirectly in the field remained low and Z + A high throughoutday and night in both sun and shade leaves. Pre-dawn trans-fer of leaves from subfreezing temperatures in the field toroom temperature revealed that recovery (increases in Fv/Fm
and conversion of Z + A to violaxanthin) consisted of one,rapid phase in shade leaves, whereas in sun leaves a rapidphase was followed by a slow phase requiring days. The pre-dawn ATP/ADP ratio, as well as that determined at midday,was similar when comparing overwintering leaves with thosesampled in the summer, although pre-dawn levels ofATP + ADP were elevated in all leaves during winter relativeto summer. After a natural transition to warmer days duringthe winter, pre-dawn Fv/Fm and Z + A in shade leaves hadreturned to values typical for summer, whereas in sun leavesFv/Fm and Z + A levels remained intermediate between thecold day in winter and the summer day. Thus two distinctforms of sustained (Z +A)-dependent energy dissipation wereidentified based upon their differing characteristics. Theform that was sustained on cold days but relaxed rapidlyupon warming occurred in all leaves and may result frommaintenance of a low lumenal pH responsible for the noctur-nal engagement of (Z + A)-dependent thermal dissipationexclusively on very cold days in the winter. The form that wassustained even upon warming and correlated with slow Z +Ato violaxanthin conversion occurred only in sun leaves andmay represent a sustained engagement of (Z + A)-dependentenergy dissipation associated with an altered PSII proteincomposition. In the latter, warm-sustained form, uncoupleror cycloheximide infiltration had no effect on the slow phaseof recovery, but lincomycin infiltration inhibited the slowincrease in Fv/Fm and the conversion of Z +Ato violaxanthin.
Key-words: Euonymus kiautschovicus; adenylate status;chlorophyll fluorescence; low temperature stress; photoinhi-bition; photoprotection; xanthophyll cycle; zeaxanthin.
INTRODUCTION
Evergreen leaves from seasonally cold environments face adifficult challenge during winter months from exposure tothe combined stresses of both high light and cold tempera-tures. While low temperatures diminish maximal rates ofphotosynthetic carbon gain they have little effect on lightabsorption (Baker 1994). An increased portion of theabsorbed energy is dissipated thermally via xanthophyllcycle-dependent energy dissipation at low temperatures(Bilger & Björkman 1991; Adams & Barker 1998). Inaddition, the conversion of the deepoxidized componentsof the xanthophyll cycle, zeaxanthin (Z) and antheraxan-thin (A), to violaxanthin (V) can be slow in overwinteringleaves, and it has been postulated that high levels of(Z + A)-dependent energy dissipation can persist through-out the night in overwintering leaves (Adams & Demmig-Adams 1994, 1995; Adams et al. 1995; Ottander, Campbell& Öquist 1995; Verhoeven, Adams & Demmig-Adams1996; Adams & Barker 1998; Barker et al. 1998). To date,the investigation of sustained energy dissipation relied pre-dominantly on characterization of leaves removed from thefield and focused on a process that persisted at warmertemperatures. In the present study, direct characterizationin the field was combined with removal of leaves from thefield and further characterization upon warming, leading tothe discovery that a large component of sustained energydissipation activity present on very cold mornings relaxesrapidly upon warming of the leaves. Thus one of the goalsof this study was to characterize the properties of thisnewly identified component of sustained xanthophyllcycle-dependent energy dissipation versus that of the pre-viously reported component that is sustained upon warm-ing of the leaves. The latter had been reported to beaccompanied by changes in PSII protein composition(Ottander et al. 1995).
Furthermore, xanthophyll cycle-dependent thermalenergy dissipation that persisted in darkness at low temper-ature was hypothesized to be due to a sustained protonationof the intrathylakoid spaces. Gilmore and Björkman(Gilmore & Björkman 1994b, 1995; Gilmore 1997) havepresented strong evidence that sustained xanthophyllcycle-dependent energy dissipation in leaves of warm-grown plants exposed to low temperatures involved themaintenance of a low intrathylakoid pH that was rapidlyreversible upon warming, and postulated that its mainte-nance might be via an ATP-dependent reverse transport of
Plant, Cell and Environment (1998) 21, 893–903
© 1998 Blackwell Science Ltd
Two forms of sustained xanthophyll cycle-dependent energy
dissipation in overwintering Euonymus kiautschovicus
A. S. VERHOEVEN,* W. W. ADAMS III & B. DEMMIG-ADAMS
Department of Environmental, Population, and Organismic Biology, University of Colorado, Boulder, CO 80309–0334, USA
ORIGINAL ARTICLE OA 220 EN
Correspondence: William W. Adams III. Fax: 303 492 8699; e-mail: [email protected]
*Present address: Department of Plant Molecular Physiology,3190 Maile Way, St. John 503, University of Hawaii at Manoa,Honolulu, Hawaii 96822, USA.
893

protons through the thylakoid ATPase. Previous characteri-zation of Euonymus kiautschovicus(Adams & Demmig-Adams 1992, 1995; Adams et al. 1992, 1995;Demmig-Adams & Adams 1996a,b; Verhoeven et al.1996), which exhibits sustained xanthophyll cycle-depen-dent energy dissipation in the winter, suggested that thisspecies would be an ideal candidate for a further character-ization of the persistent nocturnal engagement of Z + A forenergy dissipation. In addition to summer and winter fieldcharacterization of sun and shade leaves (energy dissipa-tion activity and foliar carotenoid and adenylate composi-tion) of E. kiautschovicus, the relaxation kinetics andsensitivity of the slowly reversible component of winter-induced sustained xanthophyll cycle-dependent energydissipation to uncouplers and inhibitors of protein synthe-sis were also investigated.
MATERIALS AND METHODS
Site description and plant material
Euonymus kiautschovicusLoessner was studied on thecampus of the University of Colorado at Boulder (lati-tude 40°00·8' N, longitude 105°16·8' W, elevation1730 m) from the summer of 1995 until the winter of1997. Two populations were examined, one exposed tofull sunlight throughout the day and a shade populationthat received no direct sunlight. For cold transfer experi-ments, warm-grown E. kiautschovicusplants were pur-chased at a local nursery and maintained in a greenhousein full sunlight.
Chlorophyll fluorescence
Chlorophyll fluorescence was ascertained from intactleaves under the respective growth PFD conditions with aPAM-2000 portable fluorometer (Walz, Effeltrich,Germany) or, for the uncoupler experiments, on leaf discswith the laboratory model PAM 101 and 103 (Schreiber,Schliwa & Bilger 1986). Fluorescence measurements andparameters derived therefrom were calculated as describedpreviously (Demmig-Adams & Adams 1996a; Demmig-Adams et al. 1996).
The efficiency of PSII at the actual degree of centre clo-sure (Fm' – F)/Fm' reflects the fraction of light absorbed byPSII antennae that is utilized in PSII photochemistry(Genty, Briantais & Baker 1989) while Fv'/Fm' indicatesthe efficiency of open PSII units, or the ‘excitation captureefficiency’ of PSII units (Schreiber, Schliwa & Neubauer1994). Decreases in Fv'/Fm' are apparently caused predom-inantly by increases in energy dissipation in the antennae(Demmig-Adams et al. 1996) and were therefore used asan indicator of energy dissipation in this study when it wasnot possible to compute NPQ (non-photochemical fluores-cence quenching, Fm/Fm' – 1, Bilger & Björkman 1990)under conditions of cold temperature stress when the dark-adapted Fm was already quenched (Adams et al. 1995;Verhoeven et al. 1996).
Pigment analysis
Collection, extraction, and analysis of pigments byHPLC were as described previously (Adams & Demmig-Adams 1992), using a column described by Gilmore &Yamamoto (1991).
Adenylate analysis
The collection of leaf material and extraction of adenylateswere performed as previously described by Gilmore &Björkman (1994a). Adenylates were analysed via HPLCusing a Rainin Hydropore-AX column module (5µmspherical silica, 4·6 mm ID× 25 cm). Solvent A was 5 mMNH4H2PO4, pH 3·9 and solvent B was 1M NH4H2PO4,pH 3·9. The flow rate was 1 mL min–1 with detection at254 nm. The gradient was 30–60% B in 15 min, then to100% B in 5 min, holding at 100% B for 5 min with a 2 minlinear gradient back to initial conditions and a 15 min re-equilibration time. It was not possible to analyse AMP dueto interference from UV-absorbing background at the lowsalt concentrations.
Field transfer experiments
Leaves were initially measured in the field prior to sunrisefor fluorescence and samples were taken for pigment andadenylate analysis. Leaves were then transferred to the lab-oratory and maintained at a continuous low photon fluxdensity (PFD) of 10–15µmol m–2 s–1 and room tempera-ture (20–22 °C) on moist filter paper in Petri dishes.Leaves were monitored for fluorescence and leaf discswere collected for pigment and adenylate analysis periodi-cally over the course of 100 h of recovery.
For the experiments incorporating protein synthesisinhibitors or uncouplers into winter-stressed leaves, leaveswere either characterized in the field prior to sunrise, andthen collected and transferred to the laboratory (inhibitorexperiments), or collected and transferred to the laboratoryprior to measurement (uncoupler experiments). Leaveswere kept in darkness on moist paper towels on ice. Theabaxial surface of the leaves was lightly abraded, andleaves were floated in a solution containing 0·4M sucrose,0·1% TWEEN, and either 5 mM lincomycin, 7µM cyclo-heximide or no inhibitor, or 133µM nigericin, 314µM
FCCP, both uncouplers together, or no uncouplers. Theconcentration of lincomycin used was 10 times the concen-tration reported to prevent protein synthesis upon uptake oflincomycin through the petiole (Aro et al. 1994), and waschosen to ensure inhibition of protein synthesis upon infil-tration through the cell walls, whereas the concentration ofcycloheximide was that used by Greer, Berry & Björkman(1986) for similarly treated leaves. Leaves were vacuum-infiltrated and maintained on the solution for 15 min at lowlight (≈ 10 µmol photons m–2 s–1). In the inhibitor experi-ments leaves were then transferred to moist filter paper inPetri dishes and assayed for fluorescence and pigments at 6and 26 h. In the uncoupler experiments, leaves were
894 A. S. Verhoeven et al.
© 1998 Blackwell Science Ltd, Plant, Cell and Environment,21, 893–903
remeasured for Fv/Fm after 15 min of uptake in the solutioncontaining uncouplers, and then illuminated with200 µmol photons m–2 s–1 while monitoring fluorescenceto ascertain successful incorporation of the uncoupler.
Cold room transfer experiments
Plants were collected from the greenhouse prior to sunriseand pre-light fluorescence, pigments, and adenylate sam-ples were obtained. Plants were then placed in a cold roomand exposed to between 700 and 1000µmol photons m–2
s–1 for 12 h, with leaf temperatures of 8–10 °C. The lightsource was a 1000 W metal-halide lamp. A circulatingwater-filled heat filter was placed between the light sourceand the plant to absorb most of the infrared radiation. Plantpots were covered with heating pads and insulated to main-tain soil temperature at 20–22 °C. After 12 h exposure thelights were extinguished and leaves were monitored for24 h in darkness at either room temperature (22 °C) or inthe cold (4–5 °C). Measurements of fluorescence, pig-ments, and adenylates were taken after 12 h high light andafter 15 min, 1 h and 24 h in darkness (recovery).
Statistical analysis
Calculations of correlation coefficients and Student’s t-tests were performed using Microsoft Excel 5·0. The statis-tical software package SAS was used to perform analysisof variance of the means of the three sets of the adenylatedata. When a significant difference occurred Scheffé’spairwise analysis was performed to elucidate which meanswere different from each other.
RESULTS
Short-term treatment at low temperature followedby recovery at cold versus warm temperatures
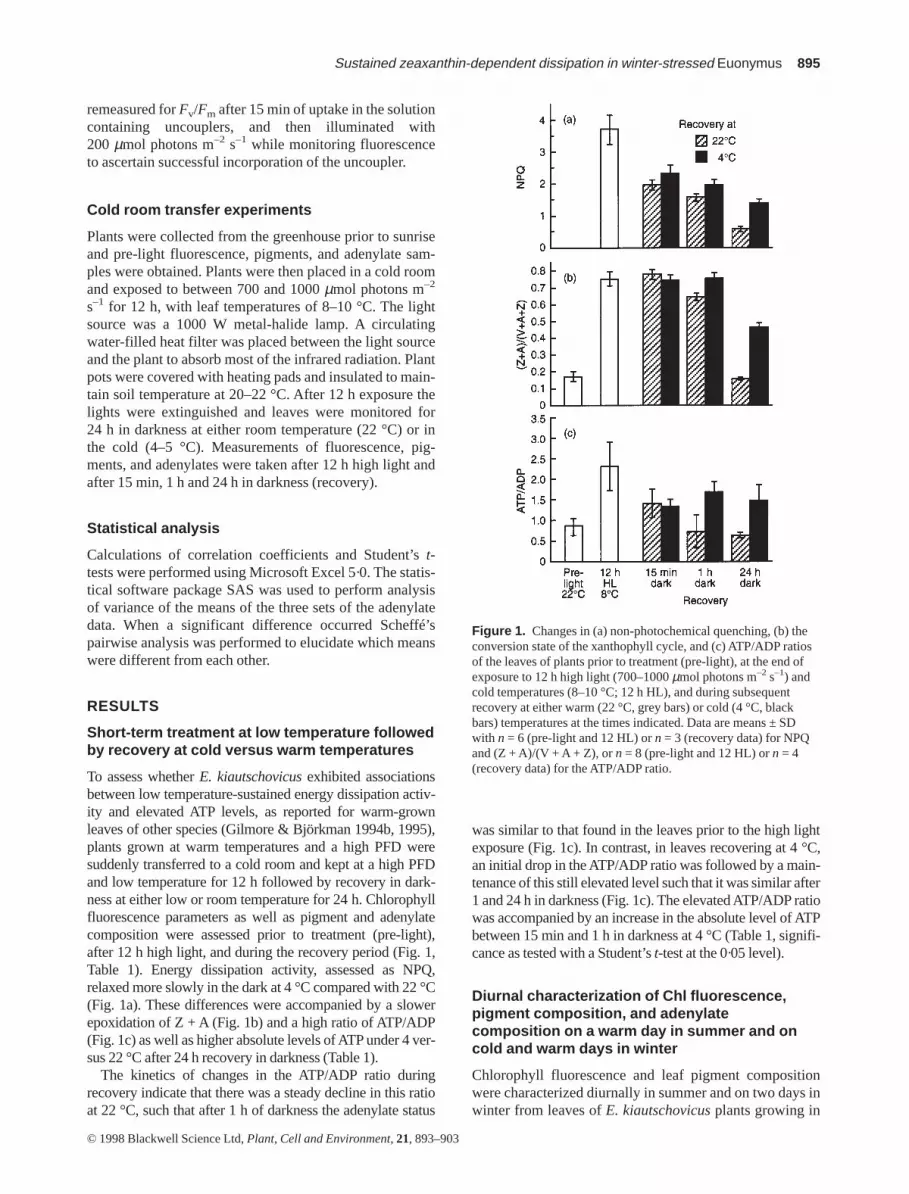
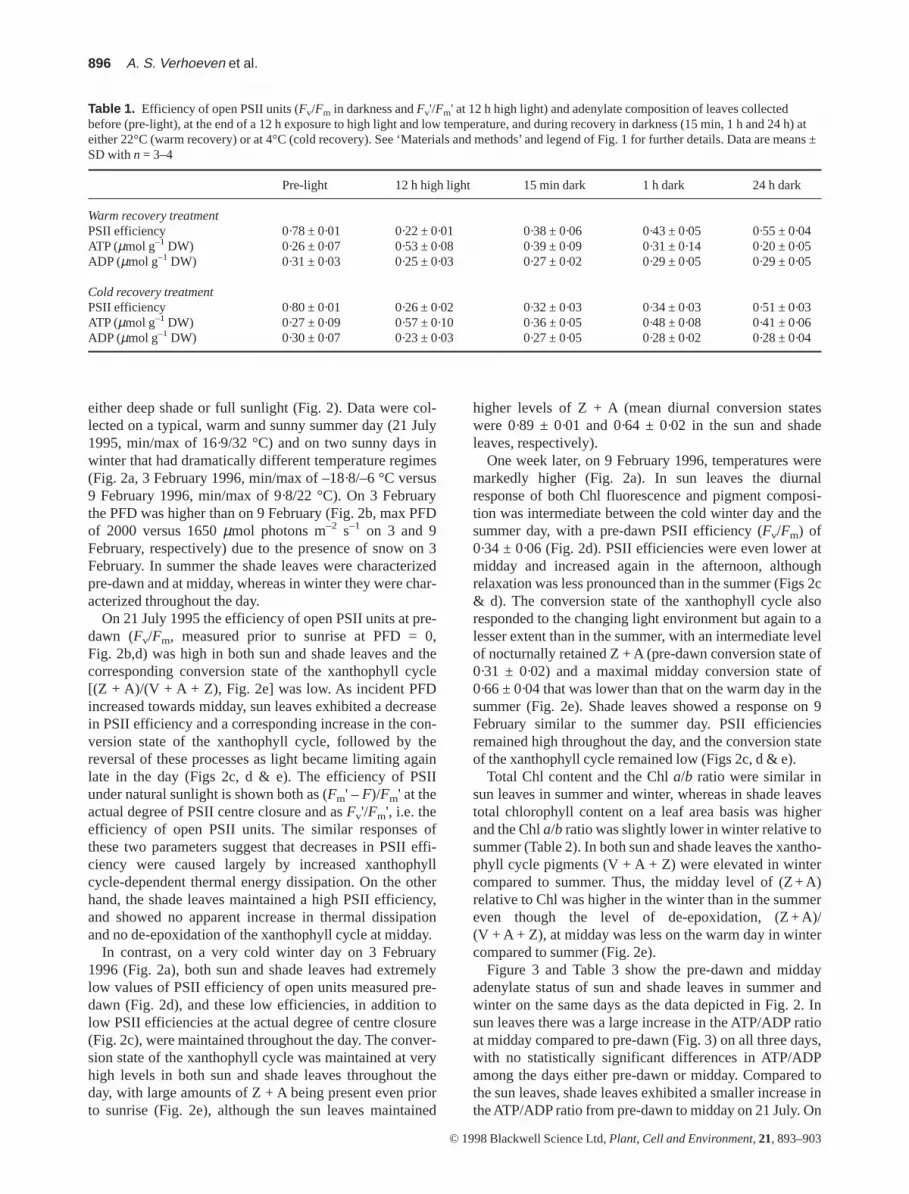
To assess whether E. kiautschovicusexhibited associationsbetween low temperature-sustained energy dissipation activ-ity and elevated ATP levels, as reported for warm-grownleaves of other species (Gilmore & Björkman 1994b, 1995),plants grown at warm temperatures and a high PFD weresuddenly transferred to a cold room and kept at a high PFDand low temperature for 12 h followed by recovery in dark-ness at either low or room temperature for 24 h. Chlorophyllfluorescence parameters as well as pigment and adenylatecomposition were assessed prior to treatment (pre-light),after 12 h high light, and during the recovery period (Fig. 1,Table 1). Energy dissipation activity, assessed as NPQ,relaxed more slowly in the dark at 4 °C compared with 22 °C(Fig. 1a). These differences were accompanied by a slowerepoxidation of Z + A (Fig. 1b) and a high ratio of ATP/ADP(Fig. 1c) as well as higher absolute levels of ATP under 4 ver-sus 22 °C after 24 h recovery in darkness (Table 1).
The kinetics of changes in the ATP/ADP ratio duringrecovery indicate that there was a steady decline in this ratioat 22 °C, such that after 1 h of darkness the adenylate status
was similar to that found in the leaves prior to the high lightexposure (Fig. 1c). In contrast, in leaves recovering at 4 °C,an initial drop in the ATP/ADP ratio was followed by a main-tenance of this still elevated level such that it was similar after1 and 24 h in darkness (Fig. 1c). The elevated ATP/ADP ratiowas accompanied by an increase in the absolute level of ATPbetween 15 min and 1 h in darkness at 4 °C (Table 1, signifi-cance as tested with a Student’s t-test at the 0·05 level).
Diurnal characterization of Chl fluorescence,pigment composition, and adenylatecomposition on a warm day in summer and oncold and warm days in winter
Chlorophyll fluorescence and leaf pigment compositionwere characterized diurnally in summer and on two days inwinter from leaves of E. kiautschovicusplants growing in
Sustained zeaxanthin-dependent dissipation in winter-stressed Euonymus 895
© 1998 Blackwell Science Ltd, Plant, Cell and Environment,21, 893–903
Figure 1. Changes in (a) non-photochemical quenching, (b) theconversion state of the xanthophyll cycle, and (c) ATP/ADP ratiosof the leaves of plants prior to treatment (pre-light), at the end ofexposure to 12 h high light (700–1000 µmol photons m–2 s–1) andcold temperatures (8–10 °C; 12 h HL), and during subsequentrecovery at either warm (22 °C, grey bars) or cold (4 °C, blackbars) temperatures at the times indicated. Data are means ± SDwith n = 6 (pre-light and 12 HL) or n = 3 (recovery data) for NPQand (Z + A)/(V + A + Z), or n = 8 (pre-light and 12 HL) or n = 4(recovery data) for the ATP/ADP ratio.
896 A. S. Verhoeven et al.
© 1998 Blackwell Science Ltd, Plant, Cell and Environment,21, 893–903
Table 1. Efficiency of open PSII units (Fv/Fm in darkness and Fv'/Fm' at 12 h high light) and adenylate composition of leaves collectedbefore (pre-light), at the end of a 12 h exposure to high light and low temperature, and during recovery in darkness (15 min, 1 h and 24 h) ateither 22°C (warm recovery) or at 4°C (cold recovery). See ‘Materials and methods’ and legend of Fig. 1 for further details. Data are means ±SD with n = 3–4
Pre-light 12 h high light 15 min dark 1 h dark 24 h dark
Warm recovery treatmentPSII efficiency 0·78 ± 0·01 0·22 ± 0·01 0·38 ± 0·06 0·43 ± 0·05 0·55 ± 0·04ATP (µmol g–1 DW) 0·26 ± 0·07 0·53 ± 0·08 0·39 ± 0·09 0·31 ± 0·14 0·20 ± 0·05ADP (µmol g–1 DW) 0·31 ± 0·03 0·25 ± 0·03 0·27 ± 0·02 0·29 ± 0·05 0·29 ± 0·05
Cold recovery treatmentPSII efficiency 0·80 ± 0·01 0·26 ± 0·02 0·32 ± 0·03 0·34 ± 0·03 0·51 ± 0·03ATP (µmol g–1 DW) 0·27 ± 0·09 0·57 ± 0·10 0·36 ± 0·05 0·48 ± 0·08 0·41 ± 0·06ADP (µmol g–1 DW) 0·30 ± 0·07 0·23 ± 0·03 0·27 ± 0·05 0·28 ± 0·02 0·28 ± 0·04
either deep shade or full sunlight (Fig. 2). Data were col-lected on a typical, warm and sunny summer day (21 July1995, min/max of 16·9/32 °C) and on two sunny days inwinter that had dramatically different temperature regimes(Fig. 2a, 3 February 1996, min/max of –18·8/–6 °C versus9 February 1996, min/max of 9·8/22 °C). On 3 Februarythe PFD was higher than on 9 February (Fig. 2b, max PFDof 2000 versus 1650µmol photons m–2 s–1 on 3 and 9February, respectively) due to the presence of snow on 3February. In summer the shade leaves were characterizedpre-dawn and at midday, whereas in winter they were char-acterized throughout the day.
On 21 July 1995 the efficiency of open PSII units at pre-dawn (Fv/Fm, measured prior to sunrise at PFD = 0,Fig. 2b,d) was high in both sun and shade leaves and thecorresponding conversion state of the xanthophyll cycle[(Z + A)/(V + A + Z), Fig. 2e] was low. As incident PFDincreased towards midday, sun leaves exhibited a decreasein PSII efficiency and a corresponding increase in the con-version state of the xanthophyll cycle, followed by thereversal of these processes as light became limiting againlate in the day (Figs 2c, d & e). The efficiency of PSIIunder natural sunlight is shown both as (Fm' – F)/Fm' at theactual degree of PSII centre closure and as Fv'/Fm', i.e. theefficiency of open PSII units. The similar responses ofthese two parameters suggest that decreases in PSII effi-ciency were caused largely by increased xanthophyllcycle-dependent thermal energy dissipation. On the otherhand, the shade leaves maintained a high PSII efficiency,and showed no apparent increase in thermal dissipationand no de-epoxidation of the xanthophyll cycle at midday.
In contrast, on a very cold winter day on 3 February1996 (Fig. 2a), both sun and shade leaves had extremelylow values of PSII efficiency of open units measured pre-dawn (Fig. 2d), and these low efficiencies, in addition tolow PSII efficiencies at the actual degree of centre closure(Fig. 2c), were maintained throughout the day. The conver-sion state of the xanthophyll cycle was maintained at veryhigh levels in both sun and shade leaves throughout theday, with large amounts of Z + A being present even priorto sunrise (Fig. 2e), although the sun leaves maintained
higher levels of Z + A (mean diurnal conversion stateswere 0·89 ± 0·01 and 0·64 ± 0·02 in the sun and shadeleaves, respectively).
One week later, on 9 February 1996, temperatures weremarkedly higher (Fig. 2a). In sun leaves the diurnalresponse of both Chl fluorescence and pigment composi-tion was intermediate between the cold winter day and thesummer day, with a pre-dawn PSII efficiency (Fv/Fm) of0·34 ± 0·06 (Fig. 2d). PSII efficiencies were even lower atmidday and increased again in the afternoon, althoughrelaxation was less pronounced than in the summer (Figs 2c& d). The conversion state of the xanthophyll cycle alsoresponded to the changing light environment but again to alesser extent than in the summer, with an intermediate levelof nocturnally retained Z + A (pre-dawn conversion state of0·31 ± 0·02) and a maximal midday conversion state of0·66 ± 0·04 that was lower than that on the warm day in thesummer (Fig. 2e). Shade leaves showed a response on 9February similar to the summer day. PSII efficienciesremained high throughout the day, and the conversion stateof the xanthophyll cycle remained low (Figs 2c, d & e).
Total Chl content and the Chl a/b ratio were similar insun leaves in summer and winter, whereas in shade leavestotal chlorophyll content on a leaf area basis was higherand the Chl a/b ratio was slightly lower in winter relative tosummer (Table 2). In both sun and shade leaves the xantho-phyll cycle pigments (V + A + Z) were elevated in wintercompared to summer. Thus, the midday level of (Z + A)relative to Chl was higher in the winter than in the summereven though the level of de-epoxidation, (Z + A)/(V + A + Z), at midday was less on the warm day in wintercompared to summer (Fig. 2e).
Figure 3 and Table 3 show the pre-dawn and middayadenylate status of sun and shade leaves in summer andwinter on the same days as the data depicted in Fig. 2. Insun leaves there was a large increase in the ATP/ADP ratioat midday compared to pre-dawn (Fig. 3) on all three days,with no statistically significant differences in ATP/ADPamong the days either pre-dawn or midday. Compared tothe sun leaves, shade leaves exhibited a smaller increase inthe ATP/ADP ratio from pre-dawn to midday on 21 July. On

the extremely cold day in winter (3 February), the shadeleaves showed no increase in ATP/ADP from pre-dawn tomidday; the ratio remained at the pre-dawn level. A weeklater (9 February) when temperatures were relatively high(Fig. 2a) there was a large increase in ATP/ADP from pre-dawn to midday, with the midday ratio reaching levels com-parable to the midday levels observed in sun leaves.
The pre-dawn levels of both ATP and ADP were higherin sun and shade leaves in winter relative to summer(Table 3). In addition, it appears that, for both sun andshade leaves, the pre-dawn retention of ADP was relatively
more pronounced than that of ATP on the cold winter day(3 February) and that ATP retention was relatively morepronounced on the warmer winter day (9 February).
Changes in the conversion state of thexanthophyll cycle and PSII efficiency ( Fv/Fm)during recovery from winter stress in leavescollected pre-dawn during winter
Leaves from sun and shade populations of Euonymuskiautschovicuswere collected from the field prior to sunrise
Sustained zeaxanthin-dependent dissipation in winter-stressed Euonymus 897
© 1998 Blackwell Science Ltd, Plant, Cell and Environment,21, 893–903
Figure 2. Diurnal characterization of (a) leaf temperature, (b) PFD, (c & d) PSII efficiency, and (e) the conversion state of the xanthophyllcycle on leaves measured in the summer (21 July 1995) and on two days in the winter (3 and 9 February 1996). Changes in PFD indicate PFDincident on the leaf. Changes in PSII efficiency are depicted as both (c) (Fm′ – F)/Fm′ (PSII efficiency at closed units) and as (d) Fv/Fm
determined pre-dawn and Fv′/Fm′ (PSII efficiency at open units). Changes in the conversion state of the xanthophyll cycle are expressed as(Z + A)/(V + A + Z). Error bars represent the SD with n = 3.

on 26 January 1996 after an overnight minimum temperatureof –14°C. Chlorophyll fluorescence characteristics, pigmentcomposition, and leaf adenylate status were determined fromleaves both in the field (time = 0) and periodically for 100 hfollowing the rapid transfer (between t = 0 and t = 2 min)from the field to low light and room temperature.
Both sun and shade leaves exhibited extremely low val-ues of Fv/Fm in the field prior to sunrise (sun, 0·12 ± 0·02;shade, 0·04 ± 0·03; Fig. 4). Leaves cut in the field werekept on ice in darkness during rapid transfer to the labora-tory. Upon transfer to room temperature Fv/Fm increasedquickly in the shade leaves, reaching 0·38 ± 0·03 after1 min and 0·65 after 1 h. Sun leaves also showed an initial,rapid increase in Fv/Fm (e.g. to 0·45 ± 0·01 after 30 min)followed by continual but slow increases over the next100 h. Thus, while recovery appeared to consist of onlyone, rapid phase in shade leaves, it consisted of a rapid anda second, slow phase in sun leaves. The increase in Fv/Fm
was due to a simultaneous increase of both Fm and Fo inthe sun leaves during recovery, while in the shade leavesthere was an initial drop in Fo upon warming (occurring inthe first 2 min), followed by a slower increase in Fo thatwas proportional to increases in Fm throughout theremainder of the recovery of Fv/Fm (data not shown; seeAdams & Demmig-Adams 1994; Verhoeven et al. 1996).Proportional increases in Fo and Fm during recovery arethought to be indicative of changes in the rate constantkD
for energy dissipation in PSII antenna complexes(Kitajima & Butler 1975), suggesting that there was sig-nificant relaxation of energy dissipation in the PSII light-harvesting antennae during recovery in both sun and shadeleaves (see Verhoeven et al. 1996).
Both sun and shade leaves started with a large fractionof the xanthophyll cycle pool converted to Z + A (sun,
898 A. S. Verhoeven et al.
© 1998 Blackwell Science Ltd, Plant, Cell and Environment,21, 893–903
Figure 3. ATP/ADP ratios in leaves collected in the field eitherpre-dawn (P) or midday (M) during exposure to maximal incidentPFD on 21 July 1995, 3 February 1996, and 9 February 1996 (seeFig. 2 for environmental conditions). The shaded columns are pre-dawn data and the open columns are midday data. The error barsrepresent the SD with n = 3–5.
Table 2. Chlorophyll and xanthophyll cycle composition of Euonymus kiautschovicusleaves sampled from summer versus winter. Summerdata are means from leaf discs collected on 21 July 1995 and winter data are means from three separate sampling days (26 January, 3February, and 9 February 1996). See Fig. 2 for environmental conditions. Data are mean ± SD, n = 3 for the summer leaves, and n = 9 for thewinter leaves. NS, no significant difference in summer vs winter (Student's t-test). (**) Significance at the 0.01 level
Sun Shade
summer winter % change summer winter % change
Chl a+b (µmol m–2) 389 ± 23 421 ± 63 NS 611 ± 61 748 ± 73 +22%**Chl a/b 3·90 ± 0·15 3·91 ± 0·28 NS 3·37 ± 0·06 3·21 ± 0·07 –5%**V+A+Z (µmol m–2) 47 ± 7 88 ± 10 +87%** 22 ± 1 44 ± 5 +100%**V+A+Z (mmol mol–1) Chl a+b 119 ± 13 217 ± 36 +82%** 38 ± 1 57 ± 5 +50%**
ATP (µmol g–1 DW) ADP (µmol g–1 DW)
Pre-dawn Midday Pre-dawn Midday
Sun samples21 July 1995 0·19 ± 0·05a 0·59 ± 0·06a 0·26 ± 0·04a 0·24 ± 0·06a3 February 1996 0·26 ± 0·04ab 0·54 ± 0·09a 0·40 ± 0·06b 0·19 ± 0·02a9 February 1996 0·33 ± 0·08b 0·57 ± 0·07a 0·36 ± 0·02b 0·18 ± 0·03a
Shade samples21 July 1995 0·10 ± 0·03a 0·26 ± 0·03a 0·15 ± 0·04a 0·23 ± 0·05a3 February 1996 0·19 ± 0·06ab 0·22 ± 0·02a 0·48 ± 0·05b 0·48 ± 0·04b9 February 1996 0·22 ± 0·06b 0·47 ± 0·03b 0·29 ± 0·04c 0·19 ± 0·02a
Table 3. Adenylate composition ofEuonymus kiautschovicusleaves collected ona summer day (21 July 1995) and on both acold (3 February 1996) and a warm day (9February 1996) in winter. See Fig. 2 forenvironmental conditions. Data are means ±SD with n = 3–4. Each group of 3 wasanalysed together using analysis of variancefollowed by Scheffe's pairwise testing; meanswithin each group with different letters aresignificantly different from each other

0·69 ± 0·02; shade, 0·59 ± 0·04). Changes in the conversionstate of the xanthophyll cycle mirrored the response ofPSII efficiency very closely, with epoxidation occurringmore rapidly in shade leaves relative to sun leaves (Fig. 4).
Shade leaves showed no significant changes in thecontents of Chl a, Chl b, neoxanthin, lutein, VAZ, α-carotene and β-carotene, all per unit leaf area, or the Chla/b ratio between sampling in the field (time = 0) and74 h following transfer to room temperature (not shown).Sun leaves showed a 35% decrease in the content of thexanthophyll cycle pigments (V + A + Z), a 10% increasein Chl a, and an 8% increase in β-carotene, all per unit
leaf area, but no significant changes in any of the otherpigments (not shown).
The adenylate composition of sun and shade leaves wasassayed in the field at time 0 and after 30 min and 100 hrecovery in the shade leaves, or after 7 and 100 h recov-ery in the sun leaves (Table 4). No marked differenceswere observed in overall adenylate status between thesesampling times.
Effects of protein synthesis inhibitors on therecovery of E. kiautschovicus leaves transferredto room temperature from the field during thewinter
Leaves collected pre-dawn, with reduced Fv/Fm valuesand elevated Z + A content, were infiltrated with either asolution containing 5 mM lincomycin (an inhibitor ofchloroplast-encoded protein synthesis), 7µM cyclohex-imide (an inhibitor of nuclear-encoded protein synthesis),or a control solution, and allowed to recover at room tem-perature in low light. The results from two separateexperiments are shown in Table 5. Differences in initialvalues (time 0) on the two days are probably due to differ-ences in minimum nocturnal temperature (–14·5 °C ver-sus –5 °C on 31 January 1996 and 12 February 1997,respectively). After 26 h recovery, leaves infiltrated withlincomycin had lower Fv/Fm levels and higher(Z + A)/(V + A + Z) ratios than the control or cyclohex-imide-infiltrated leaves.
Effects of infiltration with uncouplers on the PSIIefficiency of E. kiautschovicus leaves collectedfrom the field in winter
Leaves with reduced Fv/Fm values were collected pre-dawn on cold winter days, transferred to warm tempera-tures in the laboratory, and infiltrated with variousuncouplers at warm temperatures. Figure 5 depicts fluo-rescence traces from three infiltration experiments eachincluding a control and an uncoupler-infiltrated leaf.There was no significant increase in Fo or Fm in any of theexperiments after infiltration with either nigericin orFCCP or both. Evidence for the effectiveness of the
Sustained zeaxanthin-dependent dissipation in winter-stressed Euonymus 899
© 1998 Blackwell Science Ltd, Plant, Cell and Environment,21, 893–903
Figure 4. Changes in (a) the intrinsic PSII efficiency (Fv/Fm) and(b) the conversion state of the xanthophyll cycle in sun and shadeleaves of Euonymus kiautschovicusupon transfer to low PFD androom temperature after collection prior to sunrise on 26 January1996 (nocturnal minimum of – 14 °C). The error bars representthe SD with n = 3.
Sun samples t = 0 h t = 7 h t = 100 h
ATP (µmol g–1 DW) 0·26 ± 0·06 0·25 ± 0·09 0·19 ± 0·02ADP (µmol g–1 DW) 0·37 ± 0·05 0·46 ± 0·07 0·36 ± 0·05ε (µmol g–1 DW) 0·88 ± 0·14 0·96 ± 0·14 0·75 ± 0·08ATP/ADP 0·70 ± 0·13 0·56 ± 0·26 0·53 ± 0·04
Shade samples t = 0 h t = 0·5 h t = 100 h
ATP (µmol g–1 DW) 0·23 ± 0·04 0·22 ± 0·09 0·25 ± 0·07ADP (µmol g–1 DW) 0·37 ± 0·04 0·32 ± 0·06 0·41 ± 0·05ε (µmol g–1 DW) 0·83 ± 0·09 0·76 ± 0·23 0·91 ± 0·18ATP/ADP 0·62 ± 0·11 0·67 ± 0·17 0·60 ± 0·13
Table 4. Adenylate composition, adenylatephosphate charge ε, and ATP/ADP ratios ofsun and shade Euonymus kiautschovicusleaves collected prior to sunrise on 26January 1996 (nocturnal minimum of–14°C), and after either 7 and 100 h (sun), or0.5 and 100 h (shade) at low PFD and roomtemperature, respectively. The data aremeans ± SD, n = 4

uncoupler in uncoupling thylakoid membranes is providedby the absence of, or a reduction in, a light-induced addi-tional increase in NPQ (decrease in Fm').
DISCUSSION
In the present study, characterization of Euonymuskiautschovicusin the field revealed that all leaves, fromsun and shade populations alike, showed a persistent anddramatic lowering of PSII efficiency at the prevailing lowtemperatures during winter. In all instances the noctur-nally persistent low PSII efficiency was associated withthe nocturnal retention of Z + A, even in the most deeplyshaded leaves. Furthermore, additional characterizationof sun and shade leaves suggested that two forms of noc-turnally sustained Z + A engagement for energy dissipa-tion occurred, with one persisting exclusively atsubfreezing temperatures (and first reported for the fieldin the present study) and the other being stable even atwarm temperatures.
Persistent low PSII efficiency that relaxed rapidlyupon warming
At subfreezing temperatures in the field, nocturnally sus-tained xanthophyll cycle-dependent thermal energy dissipa-tion was observed in both shade and sun leaves of E.kiautschovicus. In shade leaves, all of the nocturnally, lowtemperature-sustained lowering of PSII efficiency reversedrapidly upon removal of leaves from the field (Fig. 4) or uponwarming in the field (Fig. 2). In sun leaves only a portion ofthe low PSII efficiency that persisted nocturnally on verycold days had the same characteristics; this portion of thelow temperature-sustained thermal energy dissipation was
reversed on a warm day in the winter (Fig. 2) and was rapidlyreversible upon transfer to warm temperatures (Fig. 4). Thischaracteristic of rapid reversibility upon warming would beconsistent with the hypothesis of Gilmore & Björkman(1995) that a sustained engagement of Z + A in energy dissi-pation in darkness results from the maintenance of a lowintrathylakoid pH at low temperatures. Upon warming, thepH gradient across the thylakoid membrane is presumablyrapidly dissipated and Z + A rapidly disengaged from energydissipation activity (Gilmore & Björkman 1994b, 1995).
As had been observed in other species (Gilmore &Björkman 1994b, 1995), leaves of warm-grown E.kiautschovicusexhibited an elevated ATP/ADP ratio whensubjected to low temperatures in the dark following expo-sure to high light (Fig. 1 & Table 1). In contrast, the pre-dawn foliar ATP/ADP ratio did not differ between winterand summer from plants characterized in the field (Fig. 3).Although elevated total levels of ATP + ADP were presentpre-dawn in overwintering E. kiautschovicus leaves(Table 3; see also Hurry et al. 1995; Strand et al. 1997),there is no supporting evidence for reverse proton pumpingduring the winter since, from a thermodynamic considera-tion, substrate/product ratios have to change in order toreverse the direction of the ATPase-catalysed reactions. Itis, however, possible that other factors might have alloweda nocturnal retention of lumenal acidity at subfreezing tem-peratures, including low temperature-induced membranepermeability changes affecting proton leakage.
Ecologically this appears to be an elegant approach tothe employment of (Z + A)-dependent photoprotectiveenergy dissipation. It is present when it is needed, remain-ing engaged for energy dissipation on very cold morningsand days, and it becomes disengaged almost instanta-neously upon the cessation of the low temperature stress.
900 A. S. Verhoeven et al.
© 1998 Blackwell Science Ltd, Plant, Cell and Environment,21, 893–903
Hours of recovery
0 6 26
31 January 1996Fv/Fm control 0·11 ± 0·06 0·48 ± 0·09 0·58 ± 0·10
+ lincomycin 0·31 ± 0·02 0·33 ± 0·06*+ cycloheximide 0·42 ± 0·04 0·54 ± 0·02
(Z+A)/(V+A+Z) control 0·86 ± 0·02 0·71 ± 0·17 0·27 ± 0·07+ lincomycin 0·87 ± 0·04 0·93 and 0·81+ cycloheximide 0·85 ± 0·09 0·49 and 0·33
12 February 1997Fv/Fm control 0·40 ± 0·06 0·59 ± 0·06 0·66 ± 0·04
+ lincomycin 0·47 ± 0·07 0·55 ± 0·04*+ cycloheximide 0·60 ± 0·04 0·65 ± 0·03
(Z+A)/(V+A+Z) control 0·73 ± 0·06 0·61 ± 0·14 0·19 ± 0·02+ lincomycin 0·74 ± 0·04 0·50 ± 0·11+ cycloheximide 0·55 ± 0·20 0·23 ± 0·05
Table 5. The effects of the protein synthesisinhibitors lincomycin and cycloheximide onthe recovery of Fv/Fm and the conversionstate of the xanthophyll cycle upon transfer ofE. kiautschovicusleaves to room temperatureand low PFD after collection prior to sunriseon a cold winter morning. Data depicted arefrom two experiments, one in 1996 and one in1997. Time 0 measurements were taken in thefield prior to collection of leaves forinfiltration. Both Fv/Fm and the conversionstate of the xanthophyll cycle(Z+A)/(V+A+Z) are depicted after both 6 and26 h of recovery. Data are means ± SD for 3leaves for each treatment. *Significantlydifferent from the control at the 0·05 level.The pigment data from 31 January 1996(nocturnal minimum of –14·5°C) at the 26 htime point for the inhibitor treatmentsconsisted of only two data points, so both aredepicted. On 12 February 1997 (nocturnalminimum of –5°C) the Studentst-testrevealed that the lincomycin-treated leaveshad a higher value for (Z+A)/(V+A+Z) with aP value of 0·056 at the 26 h time point

Interestingly, it appears that a lower temperature thresholdwas present for the persistent engagement of this energydissipation process in E. kiautschovicusacclimated to thewinter field conditions, i.e. below 0 °C (unpublished obser-vations), versus 5–15 °C for several plant species grown atwarmer temperatures (Gilmore & Björkman 1994b, 1995).
Persistent low PSII efficiency maintained uponwarming
Sustained xanthophyll cycle-dependent energy dissipationthat reversed only slowly upon warming of the leaves[either on a warmer day in the winter (Fig. 2, see alsoAdams & Demmig-Adams 1995) or upon transfer of theleaves into the laboratory (Fig. 4, see also Verhoeven et al.1996)] was observed only in leaves of E. kiautschovicusplants growing in fully exposed sites. A considerable por-tion of the nocturnally sustained energy dissipation activityobserved on the cold day (3 February) was still present onthe warm day (9 February) in the winter (Fig. 2), and a sim-ilar portion of that present on another cold day (26 January)in the winter required days to relax fully upon transfer ofthe sun leaves to the laboratory (Fig. 4). This portion thatrelaxed slowly upon warming was highly correlated (cor-relation coefficient of 0·975) with the reconversion ofZ + A to V (Fig. 4; see also Adams et al. 1995; Verhoevenet al. 1996).
Whereas the rapid relaxation of the nocturnally sus-tained low PSII efficiency upon warming was consistentwith an involvement of a persistent low intrathylakoid pHin the sustained engagement of Z + A in energy dissipation,the low PSII efficiency in sun leaves that was slowlyreversible upon warming was not. Uncoupler infiltration ofoverwintering sun leaves brought to warm temperaturesdid not result in a rapid increase in the persistent low PSIIefficiency (Fig. 5). As a cautionary note, it should beacknowledged that uncoupling of other cellular mem-branes possibly leading to an overall low chloroplast pHcannot be excluded. On the other hand, an involvement ofchloroplast-encoded protein synthesis in the slow increasein PSII efficiency is suggested by the fact that infiltrationof sun leaves with lincomycin inhibited both this slowincrease of PSII efficiency and the removal of Z + A uponwarming of the leaves, whereas infiltration with aninhibitor of nuclear-encoded protein synthesis (cyclohex-imide) did not (Table 5). These findings are consistent withprevious suggestions that chloroplast-encoded protein syn-thesis is required for the reversal of the persistent low PSIIefficiency that results from high light and low temperature(Hurry & Huner 1992) and, particularly, the observation ofconcomitant increases in PSII efficiency, Z + A epoxida-tion, and the level of the D1 protein of PSII in needles ofScots pine during the transition from winter to summer(Ottander et al. 1995). It should also be noted that increasesin Chl a and β-carotene content in sun leaves of E.kiautschovicustransferred to room temperature and lowlight for 74 h (see ‘Results’) are consistent with de novosynthesis of PSII (and/or PSI) cores (see Yamamoto &Bassi 1996).
While these results, especially the effect of lincomycin,suggest that reversal of high levels of (Z + A)-dependentenergy dissipation in overwintering sun leaves involves denovosynthesis of PSII cores, data garnered from the use ofinhibitors alone must always be interpreted with caution.Chloramphenicol (see the Hurry & Huner 1992 study) can
Sustained zeaxanthin-dependent dissipation in winter-stressed Euonymus 901
© 1998 Blackwell Science Ltd, Plant, Cell and Environment,21, 893–903
Figure 5. Fluorescence traces of leaves collected prior to sunriseon cold winter days before and after vacuum infiltration with variousuncouplers. Leaves were collected on 22 February 1994 (+ nigericinand + FCCP treatments) and on 11 March 1994 (+ both uncouplerstreatment) when the nocturnal minima were – 7·2 and – 1·7 °C,respectively. A control leaf was assayed with each uncouplertreatment as indicated. The first trace in each series depicts the dark-adapted Fo and Fm level before infiltration of the leaf, as indicated inthe bottom panel. The post-infiltration trace depicts the Fo and Fm
after infiltration and 15 min of uptake (measured in darkness), andthen followed by illumination with 200µmol photons m–2 s–1. Lightpulses were given periodically during illumination to measure Fm′.The baseline at 1·6 kHz, which was used to ascertain the Fo level offluorescence, and the baseline at 100 kHz, which was used toascertain the Fm level of fluorescence, are both indicated in thebottom panel. The upward facing arrows indicate the point at whichthe actinic light was turned on. The Fv/Fm values for each of thetreatments before and after infiltration, respectively, are control #1(0·472, 0·508), + nigericin (0·429, 0·552), control #2 (0·543,0·542), + FCCP (0·500, 0·545), and control #3 (0·699, 0·674), bothuncouplers (0·610, 0·650).

have direct inhibitory effects on photosynthesis (Okada,Satoh & Katoh 1991; see also Schnettger et al. 1994).Moreover, both lincomycin and streptomycin (seeSchnettger et al. 1994) have been reported to have sec-ondary effects, i.e. effects on Ca2+ channels and trans-membrane ion gradients in animal systems (Fiekers et al.1979; Prior et al. 1990). Therefore it cannot be excludedthat the effects of lincomycin in preventing the increasein PSII efficiency and disengagement of Z + A fromenergy dissipation activity may involve Ca2+ and/or pro-ton gradients. The low PSII efficiency that persists uponwarming of evergreen leaves that have experienced thecombined stresses of high light and low temperatures dur-ing winter has often been referred to as ‘photoinhibition’(for a review, see Adams et al. 1995). The question arises,however, as to whether this phenomenon actually limitsthe productivity of plants. The plant species in which thisphenomenon has been observed are all likely to ceasegrowth during the winter and apparently downregulatephotosynthetic activity (see e.g. Ottander & Öquist1991). Thus the persistent lowering of PSII efficiencythrough sustained (Z + A)-dependent energy dissipation,which occurs primarily in those species maintaining con-siderable levels of chlorophyll while downstream pro-cesses are downregulated, may very well have littleimpact on plant productivity, and perhaps serves a photo-protective role in chlorophyll preservation as suggestedby Ottander et al. (1995).
In conclusion, two different forms of low temperature-sustained engagement of Z + A in energy dissipation result-ing in persistent low PSII efficiency were observed inoverwintering leaves of E. kiautschovicus. Whereas oneform (that which relaxes rapidly upon warming) mayinvolve a low temperature-induced maintenance of a lowlumenal pH, the other form is stable even upon warming ofsun leaves. A restructuring of PSII, without the involve-ment of a transthylakoid pH gradient, is likely to beinvolved in the stable engagement of Z + A in energy dissi-pation following the transition from cold to warm tempera-tures. While it is thus possible that a structural change ofanother nature replaces protein protonation (Crofts &Yerkes 1994; Gilmore 1997), inducing Z + A engagementfor energy dissipation, it should also be considered that achange in the environment of the protonated groups mayconvert reversible protonation of the affected PSII proteinsinto a more stable form.
ACKNOWLEDGMENTS
The authors are indebted to Dr O. Björkman for allowingA.S.V. to spend several weeks in his laboratory to masterthe technique of adenylate analysis. T. N. Rosenstiel andDrs B. A. Logan and S. C. Grace are gratefully acknowl-edged for assistance in the field. We also thank Drs A. M.Gilmore and R. A. Dilley for helpful discussions and K.Mueh for bringing the references on the effects of lin-comycin on calcium channels to our attention. This workwas supported by the U.S. National Science Foundation,
award # IBN-9631064, a fellowship from the David andLucile Packard Foundation to B.D.-A., and a NationalScience Foundation graduate fellowship to A.S.V.
REFERENCES
Adams W.W. III & Barker D.H. (1998) Seasonal changes in xantho-phyll cycle-dependent energy dissipation in Yucca glaucaNuttall. Plant, Cell and Environment21, 501–512.
Adams W.W. III & Demmig-Adams B. (1992) Operation of thexanthophyll cycle in higher plants in response to diurnal changesin incident sunlight.Planta186,390–398.
Adams W.W. III & Demmig-Adams B. (1994) Carotenoid composi-tion and down regulation of photosystem II in three coniferspecies during the winter.Physiologia Plantarum92,451–458.
Adams W.W. III & Demmig-Adams B. (1995) The xanthophyllcycle and sustained energy dissipation activity in Vinca minorand Euonymus kiautschovicusin winter. Plant, Cell andEnvironment18,117–127.
Adams W.W. III, Demmig-Adams B., Verhoeven A.S. & BarkerD.H. (1995) ‘Photoinhibition’ during winter stress: involvementof sustained xanthophyll cycle-dependent energy dissipation.Australian Journal of Plant Physiology22,261–276.
Adams W.W. III, Volk M., Hoehn A. & Demmig-Adams B. (1992)Leaf orientation and the response of the xanthophyll cycle toincident light.Oecologia90,404–410.
Aro E.M., McCaffery S. & Anderson J.M. (1994) Recovery fromphotoinhibition in peas acclimated to varying growth irradiances:Role of D1 protein-turnover.Plant Physiology104,1033–1041.
Baker N.R. (1994) Chilling stress and photosynthesis. In Causes ofPhotooxidative Stress and Ameolioration of Defense Systems inPlants(eds C.H. Foyer & P.M. Mullineaux), pp. 105–126. CRCPress, Boca Raton.
Barker D.H., Logan B.A., Adams W.W. III & Demmig-Adams B.(1998) Photochemistry and xanthophyll cycle-dependent energydissipation in differently oriented cladodes of Opuntia strictadur-ing the winter.Australian Journal of Plant Physiology25,95–104.
Bilger W. & Björkman O. (1990) Role of the xanthophyll cycle inphotoprotection elucidated by measurements of light-inducedabsorbance changes, fluorescence and photosynthesis in Hederacanariensis. Photosynthesis Research25,173–185.
Bilger W. & Björkman O. (1991) Temperature dependent of viola-xanthin de-epoxidation and non-photochemical fluorescencequenching in intact leaves of Gossypium hirsutumL. & Malvaparviflora L. Planta184,226–234.
Crofts A.R. & Yerkes C.T. (1994) A molecular mechanism for qE-quenching.FEBS Letters352,265–270.
Demmig-Adams B. & Adams W.W. III (1996a) Xanthophyll cycleand light stress in nature: uniform response to excess direct sun-light among higher plant species.Planta198,460–470.
Demmig-Adams B. & Adams W.W. III (1996b) Chlorophyll andcarotenoid composition in leaves of Euonymus kiautschovicusacclimated to different degrees of light stress in the field.Australian Journal of Plant Physiology23,649–659.
Demmig-Adams B., Adams W.W. III, Barker D.H., Logan B.A.,Bowling D.R. & Verhoeven A.S. (1996) Using chlorophyll fluo-rescence to assess the fraction of absorbed light allocated tothermal dissipation of excess excitation.Physiologia Plantarum98,254–264.
Fiekers J.F., Marshal I.B. & Parsons R.L. (1979) Clindamycin andlincomycin alter miniature endplate current decay.Nature281,680–682.
Genty B., Briantais J.-M. & Baker N.R. (1989) The relationshipbetween the quantum yield of photosynthetic electron transportand quenching of chlorophyll fluorescence.Biochimica etBiophysica Acta990,87–92.
902 A. S. Verhoeven et al.
© 1998 Blackwell Science Ltd, Plant, Cell and Environment,21, 893–903

Gilmore A.M. (1997) Mechanistic aspects of xanthophyll cycle-dependent photoprotection in higher plant chloroplasts andleaves.Physiologia Plantarum99,197–209.
Gilmore A.M. & Björkman O. (1994a) Adenine nucleotides and thexanthophyll cycle in leaves I. Effects of CO2 and temperature-limited photosynthesis on adenylate energy charge and violaxan-thin de-epoxidation.Planta192,526–536.
Gilmore A.M. & Björkman O. (1994b) Adenine nucleotides and thexanthophyll cycle in leaves II. Comparison of the effects of CO2
and temperature-limited photosynthesis on photosystem II fluo-rescence quenching, the adenylate energy charge and violaxan-thin de-epoxidation in cotton.Planta192,537–544.
Gilmore A.M. & Björkman O. (1995) Temperature-sensitive cou-pling and uncoupling of ATP-ase-mediated, nonradiative energydissipation: Similarities between chloroplasts and leaves.Planta197,646–654.
Gilmore A.M. & Yamamoto H.Y. (1991) Resolution of lutein andzeaxanthin using a nonendcapped, lightly carbon-loaded C-18high-performance liquid chromatographic column.Journal ofChromatography543,137–145.
Greer D.H., Berry J.A. & Björkman O. (1986) Photoinhibition ofphotosynthesis in intact bean leaves: role of light, temperatureand requirement for chloroplast-protein synthesis during recov-ery.Planta168,253–260.
Hurry V.M. & Huner N.P.A. (1992) Effect of cold hardening on sen-sitivity of winter and spring wheat leaves to short-term photoin-hibition and recovery of photosynthesis.Plant Physiology100,1283–1290.
Hurry V.M., Tobiæson M., Krömer S., Gardeström P. & Öquist G.(1995) Mitochondria contribute to increased photosyntheticcapacity of leaves of winter rye (Secale cerealeL.) followingcold-hardening.Plant, Cell and Environment18,69–76.
Kitajima M. & Butler W.L. (1975) Quenching of chlorophyll flu-orescence and primary photochemistry in chloroplasts bydibromothymoquinone.Biochimica et Biophysica Acta376,105–115.
Okada K., Satoh K. & Katoh S. (1991) Chloramphenicol is aninhibitor of photosynthesis.FEBS Letters295,155–158.
Ottander C., Campbell D. & Öquist G. (1995) Seasonal changes inphotosystem II organization and pigment composition in Pinussylvestris. Planta197,176–183.
Ottander C. & Öquist G. (1991) Recovery of photosynthesis in win-ter-stressed Scots pine.Plant, Cell and Environment14,345–349.
Prior C., Fiekers J.F., Henderson F., Dempster J., Marshall I.G. &Parsons R.L. (1990) End-plate ion channel block produced by lin-cosamide antiobiotics and their chemical analogs.The Journal ofPharmacology and Experimental Therapeutics255,1170–1176.
Schnettger B., Critchley C., Santore U.J., Graf M. & Krause G.H.(1994) Relationship between photoinhibition of photosynthesis,D1 protein turnover and chloroplast structure: effects of proteinsynthesis inhibitors.Plant, Cell and Environment17,55–64.
Schreiber U., Bilger W. & Neubauer C. (1994) Chlorophyll fluores-cence as a nonintrusive indicator for rapid assessment of in vivophotosynthesis. In Ecophysiology of Photosynthesis(eds E.-D.Schulze & M.M. Caldwell), pp. 49–70. Springer, Berlin.
Schreiber U., Schliwa U. & Bilger W. (1986) Continuous recordingof photochemical and non-photochemical fluorescence quench-ing with a new type of modulation fluorometer.PhotosynthesisResearch10,51–62.
Strand Å., Hurry V., Gustafsson P. & Gardeström P. (1997)Development of Arabidopsis thalianaleaves at low temperaturesreleases the suppression of photosynthesis and photosyntheticgene expression despite the accumulation of soluble carbohy-drates.The Plant Journal12,605–614.
Verhoeven A.S., Adams W.W. III & Demmig-Adams B. (1996)Close relationship between the state of the xanthophyll cycle pig-ments and photosystem II efficiency during recovery from winterstress.Physiologia Plantarum96,567–576.
Yamamoto H.Y. & Bassi R. (1996) Carotenoids: localization andfunction. In Oxygenic Photosynthesis: The Light Reactions.Advances in Photosynthesis, Vol. 4. (eds D.R. Ort & C.F.Yocum), pp. 539–563. Kluwer Academic Publishers, Dordrecht,The Netherlands.
Received 10 February 1998; received in revised form 6 May 1998;accepted for publication 18 May 1998
Sustained zeaxanthin-dependent dissipation in winter-stressed Euonymus 903
© 1998 Blackwell Science Ltd, Plant, Cell and Environment,21, 893–903