trophic ecology of the dominant fishes in elkhorn slough, california, 1974�1980
TRANSCRIPT

Estuaries VoL’19, No. 1, p. 115-138 March 1996
Trophic Ecology of the Dominant Fishes in
Elkhorn Slough, California, 1974-l 980
&MES P. BARRY’,*
-kw M. YOKLAVICH3 GREGOR M. CAILLIET DAVID A. AMBROSE~
BROORE S. ANTRIM
Moss Landing Marine Laboratories 110. Box 450 Moss Landing, Calijbrnia 95039
ABSTRACT: Food habits of the dominant fishes collected from 1974 to 1980 at eight locations in Elkhorn Slough, California, and the adjacent ocean were investigated. Epifaunal crustacea was the major prey group identified from stomach contents of more than 2,000 fishes, followed by epifaunal and infaunal worms, and mollusca. Overall, 18 &h species consumed 263 different prey taxa, ranging from 10 taxa to 125 taxa per fish species and including 99 crustacean, 56 polychaete, and 39 molluscan taxa. Mean prey richness was greatest at stations near the ocean and lowest at inshore stations. Detailed dietary data for all prey taxa were summarized as trophic spectra for each fish species. Trophic spectra represented functional groups of prey and were used for comparisons of dietary similarity. Cluster analyses, based on tropbic spectra, resulted in four feeding guilds of fishes. Of 18 fish species, seven (Amphistidw argenteus, Leptocottus armatws, Embiotoca jacksoni, Cleuelandia ios, Gillidtthys mirabilis, Cymatogaster aggregata, and Cithurichthys stigmaeus) fed principally on epifaunal crustacea. Four species (Pleuronectes vet&s, Platichthys steli%us, Phanerodon @catus, and Mylio- batus caZ@rnicu) consumed mostly molluscs and tiaunal worms. Two species (Psettichthys melanostictus and Triahis semi- fasciata) fed on mobile crustacea, and five species (H~erprosopon anale, Engraulis ntodux, Uupea pallasi, Atherinopsis uzZa$n-niensis, and Atherinops af$nis) fed largely on zooplankton and plant material. Our results suggest that high food availability enhances the nursery function of inshore habitats, and emphasize the importance of invertebrate prey pop ulations and the indirect linkage of plant production to the ichthyofaunal assemblage, particularly marine immigrant species that are likely ‘estuarine dependent.’
Introduction
Estuaries, coastal embayments, and shallow in- shore habitats are recognized as productive systems used by fishes for reproduction, feeding, and shel- ter from pre’dators (Gunter 1938; McErlean et al. 1973; Pearcy and Myers 1974; Allen and Horn 1975; Carr and Geisel 1975; Odum and Heald 1975; Darnell and Soniat 1979; Dando 1984). These systems function as important nursery areas for fishes, particularly during spring and summer when rates of occupancy are highest (Allen and Horn 1975; Allen 1982; Onuf and Quammen 1983; Yoklavich et al. 1991) and primary and secondary production are greatest (McGowan 1986). Factors enhancing the suitability of inshore habitats as nurseries are postulated to be 1) low rates of pre-
1 Corresponding author. 2 Present address: Monterey Bay Aquarium Research Institute,
160 Central Avenue, Pacific Grove, California 93950. 3 Present address: NOAA-Pacific Fisheries Environmental
Group, 1352 Lighthouse Avenue, Pacific Grove, California 93950-2097.
4 Present address: NOAA-Southwest Fisheries Science Center, P.O. Box 271, La Jolla, California 92038.
dation, 2) high levels of food availability, 3) warm- er waters conducive to rapid growth, and 4) poten- tially greater suitability as spawning habitat for some species (Nixon 1980; Toole 1980; Krygier and Pearcy 1986; Yoklavich 1982; Gunderson et al. 1990). The seasoinal coincidence of peak fish abun- dance and secondary production suggests that food availability may be a principal factor influenc- ing the nursery function of shallow inshore habi- tats. High rates of secondary production by detri- tivores, mainly infaunal and epifaunal worms (Poly- chaeta, Oligochaeta, Nemertea) and crustaceans, increase the availability of food for fish predators.
Use of estuaries and coastal embayments by fish- es has been examined most closely along the East Coast of the United States (Shenker and Dean 1979; Weinstein 1979; Bozeman and Dean 1980), where estuaries account for nearly 85% of the coastline. Although estuaries and shallow coastal bays constitute only about 20% of the western United States coast (Day and Yanez-Arancibia 1985), several West Coast sites function as impor- tant nursery and spawning grounds for marine fishes (Pearcy and Myers 1974; McGowan 1986; Gunderson et al. 1990).
0 1996 Estuarine Research Federation 115

116 J. P. Barry et al.
Several aspects of the fauna1 composition and patterns of distribution within various habitats in Elkhorn Slough, a maritime wetland embayment along the central coast of California, have been studied since the early 1970s by faculty and grad- uate students of Moss Landing Marine Laborato- ries (California State University). These studies provide information on seasonal and spatial varia- tion in the distribution of larval, juvenile, and adult fishes (Yoklavich et al. 1991; 1992), avifauna (Ramer et al. 1991), invertebrate communities (Kvitek et al. unpublished data), and on the im- pacts of marine mammal predation on local fish assemblages (Harvey et al. in press).
Considering the dominant fishes in Elkhorn Slough and adjacent benthic habitats of Monte- rey Bay, the objectives of this study were to: 1) characterize their feeding habits and trophic re- lationships; 2) assess the relative importance of general prey groups as a forage base for fishes; and 3) estimate predator selectivity for benthic invertebrate prey. Characterization of trophic re- lationships among fish and invertebrate assem- blages is critical to estimates of energy flow and productivity, predator and prey regulation, and dependence of fishes on inshore resources. Moreover, this study provides a baseline for eval- uation of anthropogenic and other impacts on the Elkhorn Slough system.
Methods
STUDY SITE Elkhorn Slough, California (36”48’N, 121”47’W)
is a shallow coastal embayment in central Monterey Bay (Fig. 1). Prior to 1946, the slough was a coastal lagoon and seasonal estuary separated from the ocean by a sand berm that was breached infre- quently during winter storms. Geologic (Schwartz 1983; Schwartz et al. 1986; Hornberger 1991) and archaeological studies (Gobalet 1990, 1992) indi- cate dominance by freshwater flora and fauna throughout much of the slough’s history (ca. 15,000 yr) . Construction of jetties at Moss Landing Harbor in 1946 exposed Elkhorn Slough to daily tidal exchange with the ocean and increased rates of erosion in tidal channels throughout the slough (Williams et al. 1992; Crampton 1994). The main channel and tidal creeks, salt evaporation ponds, mudflats, and salt marsh compose the major hab- itats in the slough. The main channel covers ap- proximately 350 acres and extends inland 7 km, with an axial length of 11 km (Browning 1972). Its depth and width vary considerably with tidal height, ranging from nearly 100 m wide and 5 m deep at the mouth to 15 m across and 1.5 m deep at the head of the slough near Hudson’s Landing
(Crampton 1994). A network of tidal creeks me- anders through the salt marshes and, together with the main channel and Moss Landing Harbor, cov- ers nearly 1,000 ha. Salt evaporation ponds (470 ha), created early in this century, were separated from tidal flow until 1982 when dikes were partially breached by erosion, allowing limited exchange of water with the main channel. Extensive mudflats, covering over 1,000 ha, lie about 1.3 m above mean low low water (mllw) and are exposed according to height of local mixed, semidiurnal tides. Tidal salt marshes cover nearly 3,500 ha and are domi- nated by Salicornia virginica (pickleweed).
The hydrography of Elkhorn Slough is typical of coastal embayments and seasonal estuaries with strong tidal influence and freshwater input during winter. Temperature and salinity are most variable in the upper slough, where salinity ranges from about 17.7%0 after episodic winter runoff to a sum- mer high of 37.7%0 (Broenkow 1977). Tempera- ture in the upper slough ranges from 21-27°C in June to 14-16°C in winter. The lower slough is much less variable, and is similar in temperature (14-18°C) and salinity (ca. 33%0) to surface waters of Monterey Bay. Tidal flow controls circulation in Elkhorn Slough, leading to daily exchange of more than 75% of the slough’s high water volume dur- ing spring tides (Smith 1973). Even during periods of high volumetric exchange, however, waters in- shore of the tidal prism, generally located about 4.8 km inland, have residence times of months (Smith 1973).
SAMPLING DESIGN ‘Fishes from three stations in the main channel
of Elkhorn Slough and two ocean stations on the sandy shelf in Monterey Bay adjacent to the slough (Fig. 1) were sampled monthly from August 1974 to June 1976. Collections were taken at stations in the upper slough (Hudson’s Landing) and tidal creeks (Rubis Creek and Long Canyon), hereafter termed tidal creeks, at least monthly from January 1979 to June 1980.
Trawling was the principal sampling technique used at primary stations. An otter trawl, with a 4.8 m head-rope, 3.8 cm square mesh body, 2.5 cm square mesh codend, 1.3 cm square codend liner, and 183 m bridles, was towed behind a 5 m Boston Whaler powered by a 55 hp outboard engine. Tows were made between fixed markers during daylight at a speed of either 3-4 knots (main channel) or 1.5-2.5 knots (tidal creeks). Additional samples were collected using other techniques (beach seine, gill net, and channel nets) at primary sta- tions and scattered locations throughout Elkhorn Slough. Fishes were sorted by species immediately upon collection, and measured to the nearest mm

Trophic Ecology of Fishes in Elkhorn Slough 117
CALIFORNIA
121“45’N
Fig. 1. Fish (dots) and benthic j invel -tebrate (circles) sampling stations in Elkhorn Slough, California, and the ne; lore ocean.

118 J. P. Barry et al.
(standard length, SL) . Individuals selected for food habit analyses were preserved in 10% formalin and seawater for 1-2 d and transferred to 70% isopro- pyl alcohol.
DIETARY ANALYSES
We analyzed diets of numerically dominant fish- es collected in trawls from Elkhorn Slough and two nearshore ocean stations representing as many months or seasons as possible. Some species that were sampled inefficiently in trawls but that ac- counted for a high percentage of the catch using other methods were included in dietary analyses. Within months of collection, individuals of each species were selected by chance for dietary analy- ses. Contents of the upper digestive tract were identified to the lowest feasible taxa (see Appendix 1) using dissecting and compound microscopes, with the aid of experts on local floral and inver- tebrate fauna1 taxa (Nybakken and Jong 1977). All items were counted and the percentage contribu- tion of each prey type to the total prey volume was estimated visually. Individuals with empty stomachs were excluded from analyses.
Three indices (prey richness [number of prey taxa], prey diversity [Shannon-Weaver H; Shannon and Weaver 19631, and percent dominance [Simp- son 19491) were used to evaluate sufficiency of sampling effort. Cumulative indices were calculat- ed from values of prey importance (%IRI; see be- low) for all prey taxa, combined in groups of sam- ples (stomachs) ranging from one to N (all stom- achs examined), and plotted versus the number of stomachs combined. Sample size was considered sufficient when prey dominance stabilized (slope approached zero) for each fish species (usually from 10 to 15 stomachs examined), indicating that additional samples had little effect on prey domi- nance and contributed little new information on major dietary components. Because prey richness, and to a lesser degree prey diversity, are sensitive to rare taxa, these indices often continued to rise with higher sample sizes. Thus, this method max- imizes the likelihood that samples characterize the major dietary components for each fish species, but may be inadequate to identify total dietary breadth (see Ambrose 1976; Ruagh 1976; Antrim 1981; Barry 1983).
Dietary data were summarized at each station us- ing an index of relative importance (IRI; Pinkas et al. 1971), calculated from the mean percentage contribution by number (%N) , volume (%V), and frequency of occurrence (%FO) of each prey tax- on within each stomach, over the number of stom- achs that contained food items:
IRI = %FO(%V + %N)
IRIS were calculated after excluding unidentifiable digested material, which for some species account- ed for a large proportion (>40%) of the gut con- tents. IRIS were standardized to 100% for each fish species by calculating the percentage of the total IRI contributed by each prey type (%IRI), and ranged from 0 (absent from diet) to 100 (the only prey consumed).
Trophic Spectrum Analyses Four major trophic categories (flora, epifauna,
infauna, and mobile fauna) were defined from tax- onomic and natural history information, including availability of prey species as a consequence of for- aging behavior and morphological constraints of the predators (Nybakken and Jong 1977; Kvitek et al. unpublished data; see Appendix 1 for list of all prey taxa within each trophic category). These tro- phic categories represent functional groups of prey that have greater ecological meaning than those based solely on taxonomic divisions (Hay 1994). Within the flora category, prey were assigned to the subcategories: algae, vascular plants, and plant detritus, and comprised mainly of macroalgae. Blue-green algae (Cyanobacteria) and diatoms (Chyrsophyta) also were included under the algae subcategory. Epifauna was the most diverse cate- gory, comprising surface-dwelling Crustacea, Mol- lusca, Echinodermata, vermiform taxa (worms, nearly always dominated by polychaetes) , and mis- cellaneous lesser taxa. Species living deeper than the top centimeter of sedimentary habitats, includ- ing Crustacea, Mollusca, and various worms, were assigned to Infauna. Epifauna and infauna were defined primarily by their estimated availability to foraging fishes, though the distinction between these groups frequently is obscure for prey that move through sediments to varying depths, or are partially exposed (e.g., bivalve siphons). Crustacea included several taxa, but gammaridean amphi- pods and isopods were by far the most abundant groups. Bivalves were often the most common mol- luscan prey. Mobile fauna included subcategories of zooplankton, other Crustacea and Insecta, and fish and Cephalopoda. Zooplankton largely were passive drifters or weak swimmers (e.g., copepods). Other subcategories of mobile fauna were divided taxonomically but included species capable of es- cape behavior requiring somewhat greater pursuit than smaller and more sedentary epifauna and in- fauna.
A trophic spectrum (TS) was compiled for each species of fish predator at each permanent sam- pling station in order to characterize the feeding habits of ichthyofauna in Elkhorn Slough and ad- jacent ocean stations (Darnell 1961; Cailliet et al. 1978). For each station (s) , percentage IRIS for all

Trophic Ecology of Fishes in Elkhorn Slough 119
prey taxa of a fish predator (f) were summed into the trophic subcategories (defined above) :
TS, = 2 %IRI, j=l
where TS,, is the ith trophic subcategory, including all (n) prey taxa within the subcategory. Similarly, trophic spectra were calculated for each station by averaging trophic spectra for one station over all fish species:
i (TS,,. %N,J TS, = 100 i=lf
f:c i=l j=1 (TS,,. %Nsij) ’
Thus, each subcategory of a trophic spectrum for any station (TS,,) is the average TS,, over all fish species, weighted by each fish’s relative abundance at the station, and standardized over the total TS for all trophic categories (p).
Finally, an adjusted trophic spectrum (Darnell and Soniat 1979; Langton and Wading 1990) char- acterizing the feeding ecology of fishes in Elkhorn Slough as a whole was calculated by averaging tro- phic spectra from each station (TS,), weighted by the relative abundance of all fishes at each station (%C), and standardized to 100%:
TS,,, = 100 $ (TSs,* %Cj) ’ s
AC i-1 j=1 (TS,ij* %Cj)
Similar analyses of trophic spectra were completed for each of three size classes of Platichthys stellatus because this species exhibited the greatest varia- tion in size from juveniles (<99 mm) to large adults (>200 mm), which could influence prey se- lection.
Dietary Similarity Comparisons of diets between fish species and
stations were made using the pairwise percentage similarity index (PSI; Krebs 1989). PSIs were cal- culated for each species pair, based on dietary data for all prey taxa %IRI, and for trophic spectra (TS) :
PSI = 2 minimum(P,,, PZi) i=l
where PSI is the sum of the smallest %IRl or TS (PIi and PZi) of each pair of n prey categories (i) of both fish species of stations.
Similarity among diets of fishes in Elkhorn Slough also was evaluated using cluster analyses for
both the generalized diets based on trophic spectra data and detailed data including all prey taxa. Clus- tering was performed using the Ward Minimum Variance method and Pearson correlation coeffi- cient as a measure of similarity (Systat 1992). Dis- similarity among clusters of 40% of the maximum overall distance was considered a major division, and was used to define a distinct consumer guild.
Prey Availability and Predator Selectivity
The only data on distribution and abundance of any invertebrate prey of fishes in Elkhorn Slough are from sediment cores (volume = 1.0 X 1O-3 m to 3.1 X 10m3 m”) collected at several intertidal sites in the slough and Moss Landing Harbor (see Ny- bakken and Jong 1977). Overlap was estimated from pairwise comparisons (PSI indices) between abundance of benthic invertebrate prey in the cores and fish dietary data. These analyses includ- ed comparisons using both detailed prey taxa and trophic subcategories of fish diets.
Results
SAMPLE COLLECTION Otter trawl tows (n = 355) were made from Au-
gust 1974 to June 1980 at six stations in the main channel and tidal creeks of Elkhorn Slough and two inshore ocean stations; 22,334 fishes were col- lected from 65 species and 27 families. Over 3,300 additional fishes were collected from seines and gill nets deployed in channels, marsh, and ocean beach habitats. Details of the survey are discussed by Cailliet et al. (1977)) Barry and Cailliet (1981), Barry (1983), and Yoklavich et al. (1991).
Eighteen species of fish from 10 families were selected for analyses of food habits and trophic re- lationships. Ten species represented at least 1% of the total abundance in otter trawl samples (Table 1). Three species (Amphistichus argenteus, Hype$ro- sopon anale, and Psettichthys melanostictus) were in- cluded because of their relatively high abundance (>5%) in otter trawls at the ocean stations, and six additional species (Athen’nops af$nis, Atherinojxis californiensis, Cleuelandia ios, Gillichthys mirabilis, My- liobatus cal@rnica, and Triakis semifasciata) were added owing to their high abundance in non-trawl collections (beach seine, gill net, and channel nets). Overall, 11-14 species accounted for 77- 97% of the total abundance of fish captured in trawls at each station, representing the dominant fishes of Elkhorn Slough.
DIETARY ANAL~ES Overall, 263 prey taxa were identified from stem-
ach contents of 2,063 fishes, representing from 10 to 125 prey taxa per species of fish (Table 1). Crus-
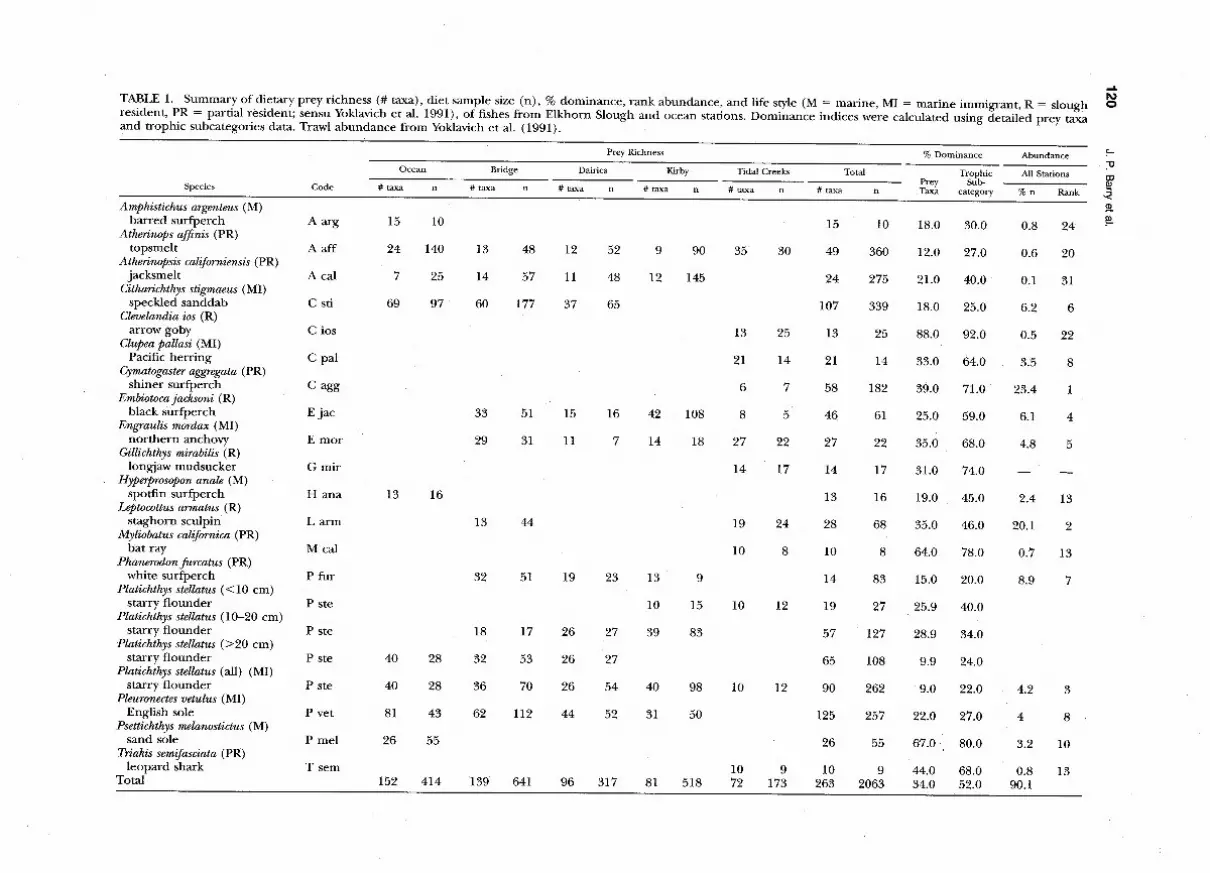
TABLE 1. Summary of dietary prey richness (# taxa), diet sample size (n), % dominance, rank abundance, and life style (M = marine, MI = marine immigrant, R = slough 6
resident, PR = partial resident; sensu Yoklavich et al. 1991), of fishes from Elkhorn Slough and ocean stations. Dominance indices were calculated using detailed prey taxa 0
and trophic subcategories data. Trawl abundance from Yoklavich et al. (1991).
Species Code
Amfihistichus argenteus (M) barred surfp&ch
Athti’nops affinis (PR) topsmelt
Atherinqtxis calij&rniensis (PR) jacksmelt
Citharichthys stigmaeus (MI) speckled sanddab
Clevelandia ios (R) arrow goby
C&pea pallasi (MI) Pacific herring
Cymatogaster aggregata (PR) shiner surfperch
Emb-iotoca jacksoni (R) black surfperch
Engraulis mordax (MI) northern anchovy
Gillichthys mirabilis (R) longjaw mudsucker
Hyperjwosopon anale (M) spotfm surfperch
Leptocottus armatus (R) staghorn sculpin
Myliobatus californica (PR) bat ray
Phanerodon furcatus (PR) white surfperch
Platichthys stellatus (< 10 cm) starry flounder
PZatichthys stellatus (lo-20 cm) starry flounder
Platichthys stelkztus (>20 cm) starry flounder
Platichthys stellatus (all) (MI) starry flounder
Plewonectes vet&us (MI) English sole
Psettichthys melanostictus (M) sand sole
Triakis semqasciata (PR) leopard shark
Total
Prey Richness % Dominance Abundance +
OCGI” Bridge Dairies Kirby Tidal Creeks TOti Tropbic P
All Stations
#taxa n #taxa n #taxa n #taxa n #taxa ” #taxa n Prey sub- I T?iXa category %n Rank 3
m
*=-g
A aff
A cal
c sti
C ios
C pal
C agg
E jac
E mor
G mir
H ana
L arm
M cal
P fur
P ste
P ste
P ste
P ste
P vet
P me1
T sem
15 10
24 140 13
7 25 14
69 97 60
24 c
48 12 52
57 11 48
177 37 65
9
12
35 30
33
29
51 15 16 42
31 11 7 14
13 25
21 14
6 7
8 5
27 22
14 17
0.8
0.6
0.1
6.2
0.5
3.5
23.4
6.1
4.8
20
31
6
22
8
1
4
5
13 16
13 44 24
8
32 51 19 23 13
10
39
- -
2.4 13
20.1 2
0.7 13
8.9 7
18
40 28 32
40 28 36
81 43 62
26 55
17 26 27
53 26 27
70 26 54
112 44 52
40
31
152 414 139 641 96 317 81
90
145
108
18
9
15
83
98
50
518
19
10
10
10
10
12
12
9
15
49
24
107
13
21
58
46
27
14
13
28
10
14
19
57
65
90
125
26
10
10
360
275
339
25
14
182
61
22
17
16
68
8
83
27
127
108
262
257
55
9
18.0 30.0
12.0 27.0
21.0 40.0
18.0 25.0
88.0 92.0
33.0 64.0
39.0 71.0
25.0 59.0
35.0 68.0
31.0 74.0
19.0 45.0
35.0 46.0
64.0 78.0
15.0 20.0
25.9 40.0
28.9 34.0
9.9 24.0
9.0 22.0
22.0 27.0
67.0 80.0
44.0 68.0 34.0 52.0
4.2
4
3.2
0.8 90.1
3
8
10
13 72 173 263 2063

Trophic Ecology of Fishes in Elkhorn Slough 121
tacea were the most diverse prey group, with 101 taxa consumed by 18 species of fish. Fifty-six poly- chaete taxa were identified, as well as 39 molluscan taxa, of which 29 were bivalves. Algae and other plant material accounted for 26 prey taxa.
Diets of flatfishes (Cithatichthys stigmaeus, Pleura- nectes vetulus, Platichthys stellatus, and Psettichthys me- lanostictus) were more diverse (i.e., higher prey taxa richness; mean = 87 taxa) than those of surf- perches (Amphistichus argenteus, Cymatogaster aggre- gata, Embiotoca jacksoni, Hyperprosopon anale, and Phanerodon furcatus, mean = 35 taxa), yet both groups fed on a greater number of prey types than the nine other fish predators (mean = 22 taxa). Pleuronectes vetulus had the most diverse diet (125 prey taxa) and Myliobatus californica and Triakis sem- ifasciata fed on the fewest prey taxa (10). Sample size partially explains the low prey richness for My- liobatus calijornica (n = 8) and Ttiakis semifaciata (n = 9), as indicated by the positive correlation between prey richness and number of stomachs ex- amined (r = 0.74, p < 0.91).
Epifaunal Crustacea was clearly the dominant prey group, accounting for >50% of the total %IRI for six species of fish. Mobile Crustacea contrib- uted to >80% of the diets of Psettichthys melanostic- tus and Triakis semifasciata. Gammarid amphipods (18 species total), particularly Coro@hium sp. and Allorchestes angusta, were a major component of fish diets. Infaunal subcategories (worms, crustacea, molluscs) ranked third in importance overall, av- eraging 16.5% of the total %IRl, but were major dietary components for Myliobatus calijornia, Pha- nerodon furcatus, and Platichthys stellatus, represent- ing 88%, 47%, and 44% of the diet, respectively. Algae and marine plants generally were unimpor- tant in diets of Elkhorn Slough ichthyofauna, with the exception of the two atherinid species Atheri- no@ af$nis and Atherinopsis californiensis. Mobile zooplankton were dominant only in the diets of the pelagic fish species Clupea pallasi and Engraulis mordax, and to a lesser extent Hyperposopon anale.
In general, diets were more limited for fishes in upper slough and tidal creek habitats. Prey rich- ness was highest at the ocean and bridge stations, where >152 prey taxa were identified over all fish species, and declined to 72 taxa at the tidal creeks stations (Table 1). Similarly, mean number of prey taxa per predator declined with distance from the ocean, due primarily to changes in prey richness in diets of marine immigrant species (sensu Yok- lavich et al. 1991; Fig. 2a). Benthic-feeding fishes (principally those species in Epifaunal Crustacea and Mollusc/infaunal worm feeding guilds; see be- low) had diets of fewer taxa when collected further inshore compared to fishes feeding in the water column (Fig. 2b).
Marine Marine Immigrant Partial Resident
T Resident
$ 75 - b= Epifautial Crustacea
f Mollusc / lnfaunal Worm
.- Mobile Crustacea I Fish
y 50- Zooplankton I Flora
S 2 # 25-
E
= 0 I I I I I I
C. $ 0.8
s .; 0.6
2 0.4 8
0.0 .I,
Fig. 2. Mean number of prey taxa per fish species by (a) life style (see Table 1 for definitions) and (b) feeding guild (see text), and (c) mean percent dominance of prey taxa by feeding guild (symbols as in 2b) at Elkhorn Slough and nearshore ocean stations (standard error bars are indicated).
Prey dominance reflected patterns of prey rich- ness, with highest dominance in the upper slough for most species (Fig. 2~). Dominance indices cal- culated from all prey taxa and from trophic sub- categories, coupled with prey richness (Table l), indicated the most specialized diets for Clevelandia

122 J. P. Barry et al.
ios, Myliobatus califwnica, and Psettichthys melanostic- tus. Flatfish, surfperch, and atherinids generally fed more equitably on a wide variety of prey. Dom- inance indices, like total prey richness, were relat- ed significantly to sample size (r = 0.56; p < 0.05), decreasing as sample size increased.
Sufficient data were collected to examine sea- sonal variation in the diets of three surfperch spe- cies and three flatfish species. Although Cymatogas- ter awegata primarily consumed polychaete worms and pericaridean crustacea, the relative propor- tions and dominant species varied among seasons. During fall 1974, gammarid amphipods were taken nearly exclusively (%IRI = 91.0), while in winter polychaetes were the major prey taxon (49%). By spring 1975, various crustacea (primarily isopods) made up 60% of the diet, and copepods contrib- uted 60% in summer. However, the heavy reliance on gammarid amphipods during fall 1974 did not recur during fall 1975, when polychaetes (72%) were the major prey type. Phanerodon furcatus and Embiotoca jacksoni were most abundant at the bridge, and showed similar seasonal shifts in diet. Through fall and spring, polychaete worms were a large portion (48% to 84%) of their diets, while in summer, the importance of crustacea, especially gammarid amphipods, increased to 42%, as did the importance of other pericaridean prey. Diets of flatfish species (Citham’chthys stigmaeus, Pleuronec- tes vet&s, Platichthys stellatus) varied less with sea- son than did the diets of surfperch species (see Ambrose 1976 and Antrim 1981 for greater detail).
TROPHIC PATTERNS OF ELKHORN SLOUGH FEEDING GUILDS
Epzfaunal Crustacea Guild
Seven species of fish specialized on epifaunal crustacea, especially gammarid amphipods and harpacticoid copepods (Fig. 3). Two, Amphistichus argenteus and Citharichthys stigrnaeus, are principally marine species that enter lower regions of the slough seasonally. Five core species-Clevelandia ios, Cymatogaster aggregata, Embiotoca j’acksoni, Gil- lichthys mirabilis, and Leptocottus armatus, reside throughout the slough most or all of the year. Diets were least diverse (i.e., lowest species richness) in the upper slough, where most species ingested two gammarid amphipods, Coqbhium sp. and to a less- er degree Orchestia traskiana, and harpacticoid co- pepods (Fig. 2b; Appendix 1).
The cottid Leptocottus armatus varied consider- ably in total length within the slough, with larger individuals found in the lower slough and juveniles abundant near tidal creek habitats. Large adults consumed small shore crabs (Hemigrapsus sp.) and gammarid amphipods, macroalgae (Enteromqha
sp.) , and other minor prey items. Small individuals fed on smaller, less mobile prey, especially gam- marid amphipods (Corobhium sp.), harpacticoid co- pepods, and polychaete worms.
Embiotica jacksoni was common throughout the slough but most abundant near the bridge station, where it averaged 30% of the total fish catch. This species forages for epifaunal and infaunal prey consisting of polychaete worms, caprellid amphi- pods (Caprella sp.), and other amphipods at the bridge station. Bivalves and mobile decapods were taken most frequently at the dairy station, while epifaunal amphipods (Corophium sp. and Allorches- tes angusta) and isopods (Ianiropsis sp.) dominated diets at upper slough stations.
Resident fishes restricted to shallow mud flats and tidal creeks (Clevelandia ios and Gillichthys mi- rabilis) fed mainly on epifaunal crustacea (Fig. 3; Appendix 1). Cleuelandia ios is small (total length -2-3 cm) and incapable of taking large prey. Its diet consisted largely of harpacticoid copepods. Gillichthys mirabilis is larger (total length -15 cm) and fed predominantly on gammarid amphipods, especially Orchestia traskianna, and to a lesser ex- tent on polychaete worms. Cymatogaster aggregata ranked first in overall abundance in the main channel of Elkhorn Slough, where it fed mainly on epifaunal worms (Armandia breuis and other poly- chaetes) and crustacea (harpacticoid copepods, gammarid amphipods) . The relative importance of gammarid amphipods (primarily Corophium sp.) , and to a lesser degree isopods (Ianiropsis sp.), was greater in the upper main channel (Kirby Park) than at lower slough stations.
Amphistichus argenteus and Citharichthys stigmaeus had the most distinct diets within this guild (Fig. 3; Appendix 1) and fed on several gammarid am- phipod species (mainly Aoroides columbiae, Atlus tri- dens, and Monoculoides spinipes), as well as other prey taxa (bivalve siphons, polychaete worms). Am- phistichus argenteus ingested prey more typical of its coastal ocean habitat (i.e., sand dollars [Dendraster excentricus] and sand crabs [Emerita analoga] ) . Cith- arichthys stigmaeus is a marine immigrant that fed heavily on mysids at the ocean stations, and to a lesser degree on gammarid amphipods and deca- pods. Epifaunal worms, (primarily Armandia brevis) ,
crustacea (Aoroides columbiae) and bivalve siphons were ingested most commonly at the bridge sta- tion. Near the dairy station, gammarid amphipods (Aoroides sp.) and bivalve siphons dominated the diet.
Mollusc and Infaunal Worm Guild Molluscs and infaunal worms dominated the di-
ets (>65% overall) of Pleuronectes vetulus, Platichthys stellatus, Phanerodon furcatus, and Myliobatus califor-

Trophic Ecology of Fishes in Elkhorn Slough 123
Fig. 3. Dietary summary of trophic subcategories, grouped by station, for seven fish species in ;he Epifaunal Crustacea feeding guild. Fill pattern as in Fig. 6.
&a (Fig. 4; Appendix 1). The pleuronectid flat- fishes, Pleuronectes vetulus and Platichthys stellatus, are marine immigrants that use the slough season- ally for reproduction and as a nursery (Yoklavich et al. 1991). Pleuronectes vetulus had a somewhat broader diet at the ocean stations, where 81 prey taxa were dominated by epifaunal and infaunal polychaetes, ostracods, amphipods, and bivalve si- phons. Further inland, prey richness decreased, and bivalve siphons, epifaunal and infaunal poly- chaetes, and epifaunal amphipods were most im- portant (Fig. 3).
The diet of Platichthys stellatus also varied from the ocean to upper slough, perhaps related to changes in prey availability and to fish size distri- bution. The number of prey taxa was fairly con-
stant in the main channel and decreased strongly only at tidal creek stations (10 taxa; Table 1). Greatest variation in fish size (ca. 50-300 mm SL) occurred at the bridge and dairy stations. Most Pla- tichthys stellatus from upper slough and shallow marsh habitats were less than 150 mm, while size at the ocean stations was mainly >150 mm (Am- brose 1976). Larger individuals ingested a wider l
variety of prey (Fig. 5), relying on polychaete worms and bivalve siphons, in addition to a higher relative importance of prey potentially unavailable to small fishes, such as mobile crustacea (decapod crabs) and infaunal molluscs.
Polychaete worms were a lesser component in diets of Phanerodon furcatus and Myliobatus calafor- nicu. Phanerodon furcatus was captured at all stations

124 J. P. Barry et al.
80
Fig. 4. Dietary summary of trophic subcategories, grouped by station, for four fish species in the Mollusc/Infaunal Worm feeding guild. Fill pattern as in Fig. 6.
except the tidal creeks but was abundant only in the lower slough (bridge and dairy stations), where it accounted for 18-34% of the total fish abun- dance (Table 1). Infaunal polychaete worms and caprellid amphipods were its most common prey near the bridge and dairy stations, with a shift to decapod crabs and bivalves in the upper slough (Fig. 4; Appendix 1). Myliobatus calijbrnica fed pri- marily on infaunal bivalves (95 %IRI) and epifaun- al gammarid amphipods. and gastropods.
MOBILE CRUSTACEA AND FISH GUILD
Although mobile crustacea were the main prey of Psettichthys melanostictus and Tm’akis semifasciata (Fig. 6), they consumed different species (Appen- dix 1). Psettichthys melanostictus fed on mysids and
Crangon sp. as juveniles (Cl50 mm), then switched to larger more mobile prey (mysidacea [mainly Ex- acanthomysis da&ii]) as adults, and even ate juve- nile conspecifics. Triakis semifasciata ingested mo- bile grapsid crabs (principally Hemigrapsus oregonen- sis) and a green alga (Enteromorpha sp.), which was probably taken incidentally in pursuit of animal prey.
Zooplankton and Flora Guild
Hype$wosopon anale, Engraulis mordax, Clupea pal- lasi, Atherinopsis californiensis, and Athem’nops affinis ate zooplankton and marine plants (Fig. 7). The atherinids (Atherinopsis californiensis and Atherinops af$nis) are partial residents (Yoklavich et al. 1991) distributed throughout the slough. Atherinops af

80
Fig. 5. Dietary summary of trophic subcategories for Pla- tichthys stellatus, grouped by size. Fill pattern as in Fig. 6.
finis had a generalized diet at the ocean stations, where mobile fauna, such as cyclopoid and cal- anoid copepods, euphausids, and diatoms were the dominant prey taxa. Zooplankton was less im- portant further inshore, where diatoms and mac- roalgae (Enteromorpha sp. and Ulna sp.) increased in the diet. Infaunal worms (mainly nematodes) represented a large proportion (29.2 %IRI) of the diet of Atherinops afJinis at the Kirby Park station; however, nematodes likely were gastrointestinal parasites rather than prey captured from benthic sediments. The most common food ingested by Atherino$sis californiensis was green algae (Ulna sp. and Enteromo$ha sp.), and diatoms (mainly Melo- sira moniliformis) . Eggs of Atherino@sis californiensis were found in stomachs of several adults of this species at the bridge and Kirby Park stations. Zoo- plankton represented >97% of the trophic spec- trum at the ocean stations, owing to a statistical artifact whereby one euphausid was the only countable item in the stomach (other items were too small or amorphous to identify) and thus con- tributed 100% to the numerical component of the RI.
Trophic Ecology of Fishes in Elkhorn Slough 125
Diets of clupeiform fishes ( Clupea pallasi and En- graulis mordax) were available only for the com- bined data from Hudson’s Landing and tidal creeks stations. Both fed on zooplankton, consist- ing mainly of harpacticoid and calanoid copepods (Fig. 7). Because they were not identified to lower taxonomic levels, contributions to diets by harpac- ticoid taxa were indistinguishable, even though this group has members common in benthic sedi- ments and the water column. Thus, harpacticoids were designated as benthic crustacea, except for fishes known to forage in the water column (Ath- erinops af$nis, Clupea pallasi, and Engraulis mordax) ,
for which harpacticoids were assigned to zooplank- ton. Unlike other surfperches, Hyperprosopon anale fed mainly on planktonic invertebrate larvae (me- galopae and zoea larvae), mysids, and harpacticoid copepods.
DIETARY SIMILARITY
Dietary similarity (PSI), calculated from detailed data on all prey taxa, indicated high overlap (PSI>SO) for several species of fishes in Elkhorn Slough (Table 2). Clupea pallasi and Engraulis mor- dax both forage in the water column and fed on very similar prey (PSI = 67.8). Engraulis mordax’s diet also was similar to Atherinops afJinis (PSI = 36.9), due to harpacticoid and calanoid copepods and the chrysophyte, Gyrosigma sp. Both atherinids consumed similar chrysophyte taxa, calanoid co- pepods, and the green alga, Enteromorpha, resulting in a high mean overlap (PSI = 31.5). Three spe- ties-Leptocottus armatus, Cymatogaster aggregata, and Embiotoca jacksoni, fed heavily on the benthic amphipod, Corophium sp., and had high overlap (PSI = 52-67). Phanerodon furcatus had a very broad diet, which was similar to Embiotoca jacksoni (PSI = 34.4) because both fed on polychaetes, Cor- ophium sp., amphipods, decapods, and caprellid amphipods. Platichthys stellatus, Pleuronectes vetulus, and Citharichthys stigmaeus held the 4th to 6th high- est ranks in overlap (PSI = 38-50) by ingesting bivalve siphons and polychaetes (mainly Armandia brevis) .
Dietary similarity, calculated from trophic spec- tra data, resulted in much higher resource overlap (Table 2). However, PSI based on TS values were significantly correlated with PSI values calculated from %IRI values for detailed prey taxa (r = 0.62; n = 153; p < 0.01). Cymatogaster aggregata, Clevelan- dia ios, Citharichthys stigmaeus, Embiotoca jacksoni, and Gillichthys mirabilis, relying largely on epifaunal crustacea, had a mean PSI of 72.0. High dietary overlap between Engraulis mordax and Clupea pallasi (PSI = 85.1) resulted from foraging on harpacti- coid copepods, classified here as zooplankton be- cause of the filter-feeding behavior of these fish.

126 J. P. Barry et al.
80
E 60
m Algae Vascular Plant Detritus Epifaunal worm
m Epifaunal Crustacea
m Epifaunal Mollusc
~ Epifaunal Echinoderm
m Miscellaneous Epifauna
Infaunal Worm Infaunal Crustacea lnfaunal Mollusc
I1 Mobile Fish/Cephalopod l777I Mobile Crustacea 6Sl Zooplan kton
Fig. 6. Dietary summary of trophic subcategories, grouped by station, for two fish species in the Mobile Crustacea/Fish feeding guild.
Cymatogaster aggregata, Embiotoca jacksoni, Gillichthys mirabilis, Leptocottus armatus, Amphistichus urger&us, and Citham’chthys stigmaeus) fed principally on epi- fauna1 crustacea, with lesser ingestion of infaunal and epifaunal worms. A second cluster (Phanerodon furcatus, Platichthys stellatus, Pleuronectes vetulus, and Myliobatus calfornica) generally had broad diets of . epifaunal and infaunal molluscs, and infaunal worms. The only predators (Psettichthys melanostic- tus and Ttiakis semifasciata) to include mobile crus- tacea as their primary food source comprised a third cluster. The fourth cluster (Atherinop af$nis, Atha’nopsis californiensis, Clupea pallasi, Engraulis mordax, and Hyperprosopon anale) fed primarily in the water column, with zooplankton and diatoms as principal prey.
Hyper$wos@on anale fed similarly to clupeiform fish- es (mean PSI = 65.9) owing to consumption of zooplankton. High overlap between Platichthys stel- latus and Pleuronectes vet&us (PSI = 71.6) resulted mainly from eating infaunal and epifaunal worms. Triakis semvasciata and Psettichthys melanostictus fed on different species of mobile crustacea, as indi- cated by a PSI of 0.0 based on detailed data; using importance values from trophic spectrum catego- ries, a much higher dietary overlap (PSI = 80.9) was demonstrated.
Cluster analysis of predator diets, based on %IRIs from trophic subcategories, produced four feeding clusters for fishes in Elkhorn Slough and nearby ocean stations (Fig. 8). Not surprisingly, the most well-defined cluster (including Cleuelandia ios,

Trophic Ecology of Fishes in Elkhorn Slough 127
80
Fig. 7. Dietary summary of trophic subcategories, grouped by station, for five fish species in the Zooplankton/Flora feeding guild. Fill pattern as in Fig. 6.
PREY AVAILABILITY AND PREDATOR SELECTMTY
Infaunal and epifaunal worms (primarily the polychaetes Armandia breois, Capitella capitata, Exo- gene lourea, and Streblospio benedicti) were the most abundant invertebrates in the sediment cores (Ta- ble 3). Epifaunal crustacea (tanaids, Leptochelia du- bia; cumaceans, Cyclaspis sp.; gammarid amphi- pods, Corophium sp.) were second in abundance. Together these species accounted for SO-90% of total invertebrate abundance.
Similarity between fish diet and prey availability was highest for benthic foragers, especially those that fed on both epifaunal crustacea and epifaunal and infaunal worms (Table 4). The diet of Cyma-
togaster aggregata was most similar to invertebrate abundance (mean PSI = 68.5), with highest simi- larity in the lower slough (PSI = 81.3) where this species was most abundant. Diet of juvenile Pla- tichthys stellatus (<lo cm) also was very similar to prey abundance, with a mean PSI of 66.4 over all stations. Similarity between diet and invertebrate abundance was marginally higher at Kirby Park (PSI = 34.1) than at Moss Landing Harbor (PSI = 31.9). Diets of marine species were least similar to invertebrate abundance (mean PSI = 14.1), while resident species diets were most similar (mean PSI = 43.0).
Estimates of prey availability based on core sam- ples were not adequate to represent the range of

128 J. P. Barry et al
prey available for several species of fishes in Elk- horn Slough. Most flora, zooplankton, and mobile fauna were not sampled effectively, or at all, by sed- iment cores.
SPATIAL VARIATION IN ICHTHYOFAUNAL FORAGING PRESSURE
The overall trophic spectrum for Elkhorn Slough, calculated by weighting trophic spectra by total fish abundance at each station, indicates sev- eral prey groups contributed to the total energy budget of the ichthyofaunal assemblage (Fig. 9). Epifauna accounted for >65% of the total prey consumed in the slough. Epifaunal crustacea and both epifaunal and infaunal worms (principally Polychaeta) represented the majority of the food base for the fish assemblage. Mobile fauna were ingested heavily by a few species, accounting for nearly 12% of the total prey. Flora were a minor component of the forage base.
The increase in relative importance of epifaunal crustacea, and decrease in nearly all other groups, from the bridge to tidal creek stations, is evident from the adjusted trophic spectra for each station (Fig. 9). Comparison of average fish foraging pat- terns among stations was possible by calculating simi!arity indices (PSI) for station pairs, using ad- justed trophic spectra for each station. As expect- ed, overlap in foraging pressure was least similar between the ocean and the three Elkhorn Slough stations (Table 5). Overlap increased between sta- tions further inshore, notably in response to nar- rower prey richness at inshore stations.
Overlap between prey availability and overall tro- phic spectra for each station corroborates the pat- tern of greater dietary specialization with increased distance inshore (Table 6). The overall trophic spectrum for Kirby Park was less similar to prey availability at other stations (mean PSI = 47.7) than were trophic spectra of the dairy and bridge to other stations (mean PSI = 61.7). Furthermore, prey abundance at Kirby Park was, on average, more similar to trophic spectra of all stations (mean PSI = 70.5) than were prey abundances from other stations (mean PSI = 57.8).
Discussion LINKAGE OF ICHTHYOFAUNA TO
PRIMARY PRODUCTION Estuarine food webs are complex, and include
important linkages to primary production via ben- thic microalgae and macroalgae, vascular plants, and phytoplankton (Haines and Montague 1979; Hackney and Haines 1980). Marine plants and plant detritus are thought to be a primary food source for fishes in some systems (Darnell 1958, 1961; Odum and Heald 1975; Naiman and Sibert

Trophic Ecology of Fishes in Elkhorn Slough 129
Trophic Guilds
Epifaunal Crustacea
Epifaunal Molluscs, lnfaunal Worms and Molluscs
Mobile Crustacea, Fish
Zooplankton, Flora
Predators Amphistichus argenteus
Leptocottus armatus
Embiotoca jacksoni
Clevelandia ios
Gillichthys mirabilis
Cymatogaster aggregata
Citharichthys stigmaeus
Pleuronectes vetulus Platichthys stellatus
Phanerodon furcatus Myliobatus californica
Psettichthys melanostictus
Triakis semifasciata Hyperprosopon anale
Engraulis mordax
Clupea pallasi
Atherinopsis californiensis
Atherinops affinis
Cluster Analysis
1 t
0.0 Dissimilarity 5.0
Fig. 8. Cluster analysis of ichthyofauna in Elkhorn Slough and nearshore ocean stations, analysis based on dietary information summed to trophic spectrum categories (see text).
1979; Barry and Ehret 1993). Epiphytic microbial communities and microfauna associated with de- tritus are, however, thought to be more important nutritionally than associated detrital plant material (Tenore and Rice 1980; Marinucci 1982), and vas- cular plants are less productive in some estuaries and coastal embayments than benthic microalgae and macroalgae (Zedler et al. 1978; Haines 1979). Moreover, organic aggregates of microalgae, mac- roalgae, microbes, vascular plants, and other
TABLE 3. Summary of invertebrate abundance (% number) estimated from sediment cores summed by trophic subcatego- ries (raw data from Nybakken et al. 1977 and Kvitek et al. un- published data).
Trophic Category M.L. Kirby Subcategory Harbor Bridge Dairy Park Average
Epifauna ‘Worms’ 26.3 45.7 19.6 22.9 Crustacea 38.9 10.6 36.3 59.8 36.4 Mollusca 6.5 1.6 Echinodermata Miscellaneous Groups
Infauna ‘Worms’ 31.6 39.4 63.7 14.1 37.2 Crustacea Mollusca 3.2 4.3 1.9
100.0 100.0 100.0 100.0 100.0
sources comprise >98% of the detrital export of a Gulf Coast salt marsh (Ribelin and Collier 1979), and are a primary food source for small inverte- brates.
These sources may require processing by micro- bial communities and herbivorous invertebrates and fishes before they are used by the majority of fishes. Although direct consumption of plant de- tritus was relatively unimportant to fishes in Elk- horn Slough, detritus-consuming prey, such as har- pacticoid copepods, gammarid amphipods, and a few species of polychaete worms, dominated the diets of several fish species. Thus, small inverte- brates, the primary prey of the slough’s fish assem- blage, represent secondary, tertiary, or higher lev- els of consumption via complex pathways poten- tially based on plant detritus. In contrast to many tropical and subtemperate maritime wetlands worldwide, mullet (Nlugil ce@zZus) and other de- tritivorous fishes are uncommon in Elkhorn Slough. Nonetheless, the indirect importance of plant material to higher consumers does not de- crease its potential role in regulating patterns of production, or vice versa, via cascading trophic in- teractions (Vanni et al. 1990; Carpenter and Kitch- ell 1992). Indeed, a suite of invertebrate prey may link food web compartments comprising primary

130 J. P. Barry et al.
TABLE 4. Percent Similarity (PSI) between diet of fishes (av- eraged by species over all stations) and prey availability in sed- iment cores collected at several locations in Moss Landing Har- bor and Elkhorn Slough.
Species
Amphistichus argenteus (MI) Atherinops afinis (PR) Atkerinopsis californiensis (PR) Citharichthys s&ma,, (MI) Clevelundia ios (R) Clupea palhi (Mi) Cymatogaster agp-egata (PR) Embiotoca jacksoni (R) Engraulis mwdax (Ml) Gillichthys mirabilis (R) Hyp~rosopon an& (M) Leptocottus armatus (R) Myliobatus califmnica (PR) Phanerodon furcatus (PR) Platichthys stellatus (<lo cm) Platichthys stellatus (IO-20 cm) Platichthys stellatus (>20 cm) Platichthys stellatus (all) (MI) Pbzuronectes vet&us (MI) Psettichthys melanostictus (M) Triakis semifasciata (PR) Average
Mean Marine Mean Marine Immigrant Mean Partial Resident Mean Resident
Benthic Sampling Stations
M.L. Harbor Bridee Dairv
Kirby Park MGUl
40.9 12.5 36.3 44.8 33.6 15.0 15.0 14.9 15.0 14.9
0.1 0.1 0.1 0.1 0.1 59.5 31.2 41.3 66.3 49.6 43.1 14.8 37.9 64.0 39.9 17.4 17.4 17.3 17.3 17.3 81.3 80.7 54.2 57.6 68.5 49.8 21.8 43.9 67.6 45.8
3.4 3.4 3.4 3.4 3.4 48.6 20.3 44.5 69.5 45.7
6.2 6.2 6.2 6.2 6.2 46.7 18.4 40.7 56.9 40.7
6.3 7.4 3.1 9.7 6.6 52.1 38.1 47.2 45.0 45.6 70.6 50.1 84.4 60.7 66.4 38.2 39.3 20.7 41.5 34.9 37.2 38.3 31.6 24.4 32.8 52.8 51.8 43.4 39.7 46.9 48.0 46.5 30.4 48.1 43.2
2.5 2.5 2.5 2.5 2.5 0.7 0.7 0.6 0.6 0.6
31.9 21.6 26.0 34.1 28.4
16.5 7.1 15.0 17.8 14.1 36.2 30.1 27.2 34.9 32.1 25.9 23.7 20.0 21.3 22.7 47.0 18.8 41.7 64.5 43.0
producers and detritivores with higher level pred- ators (sensu Raffaelli and Hall 1992). Temporal and spatial variation in fish abundance and prey productivity may be regulated largely by patterns of primary production and plant detritus (e.g., Vet- ter 1994), especially for fishes that are ‘estuarine dependent.’ Our results suggest enhancement of ichthyofaunal productivity in inshore habitats, ow- ing to high abundance of invertebrate prey and coupled with high rates of primary production by diatoms and marine plants (Odum 1971; Nybak- ken 1987; Langton and Wading 1990).
The ichthyofauna of Elkhorn Slough, like other temperate bays and estuaries, consists of a few spe- cies that reside all or most of the year within the system, and several dominant species that occupy the slough seasonally as juveniles or reproductively active adults (Allen and Horn 1975; Hoff and Ibara 1977; Allen 1982; Onuf and Quammen 1983; Yok- lavich et al. 1991). The nursery function of shallow inshore habitats has been attributed to increased growth potential owing to higher food abundance and elevated water temperature, as well as reduced predation (Nixon 1980; Boesch and Turner 1984). Analyses of diets of the dominant fishes in Elkhorn
Slough indicate high prey abundance, but do not discount alternative or complementary hypotheses.
Consumption of epifaunal crustacea as the pri- mary food resource for surfperch and flatfishes in Elkhorn Slough during spring and summer em- phasizes the importance of a plant detritusbased system, particularly considering the nursery func- tion of the slough for juveniles of several species during these seasons (e.g., Ambrose 1976; Cailliet et al. 1977; Antrim 1981; Barry and Cailliet 1981; Barry 1983; Yoklavich et al. 1991, 1992). Small her- bivorous and omnivorous crustacea and polychaete worms respond rapidly to increases in microalgal and macroalgal blooms during spring and summer, resulting in increased prey abundance and avail- ability to fishes,
High population abundance of a variety of small benthic crustacea and worms may be a critical fea- ture of the nursery function, due to limitations on the maximum prey size that juvenile fishes are ca- pable of handling. Diets of fish species often vary with size (Orcutt 1950; Bregnballe 1961; Haertel and Osterberg 1967; Miller 1967; Levings 1974). Changes in diet with size, documented for Pla- tichthys stellatus (Fig. 5) and Psettichthys melanostictus in Elkhorn Slough (Ambrose 1976), suggest that these fishes consume the largest prey possible un- less small prey are abundant and densely concen- trated (Werner and Gilliam 1984). In this way, the distribution of juvenile fishes may be regulated partially by the distribution of their prey.
AFWNDANCE AND DISTRIBUTION OF PREY RESOURCES
‘Diets of fishes in Elkhorn Slough generally were similar to patterns of prey abundance, and the lat- ter likely influenced the local distribution of fishes. Ichthyofauna of estuaries and shallow marsh hab- itats often are categorized as trophic generalists that use abundant, but highly variable, prey re- sources. Densities of intertidal invertebrates at sta- tions adjacent to fish sampling sites in Elkhorn Slough varied considerably from 1974 to 1976, with episodic peaks for several species and near fivefold changes in total invertebrate abundance (Nybak- ken and Jong 1977).
Patterns of invertebrate abundance were not clearly seasonal or coherent among taxa. Density of polychaetes tended to increase during winter, and crustacean density was most often highest in summer. Polychaetes, especially capitellids (Capitel- la capitata and Notomastus ten&), spionids (Streblos- pie benedict+, and opheliids (Armandia brewis), gen- erally were present in greater densities than other taxa. Crustacea were occasionally most abundant (particularly in the upper slough) but usually ranked second in density. Gammarid amphipods,

Trophic Ecology of Fishes in Elkhorn Slough 131
80
E 6o
9 0
40
*20
0
Fig. 9. Integrated trophic spectrum for the entire fish assemblage at each Elkhorn Slough and nearshore ocean station, and at all stations combined.
Corophium sp., the cumacean Cyclaspis sp., and the tanadacean Leptochelia dubia occurred in high den- sities episodically at all stations.
The most abundant polychaetes and crustaceans generally were prevalent in fish diets, particularly at stations and during periods of high densities, indicating an opportunistic shift in spatial distri- bution by either the fish assemblage, the principal prey, or both. For example, where Corofhium sp. was most abundant in core samples (Kirby Park), it also increased in relative importance in fish di- ets. Moreover, similarity between adjusted trophic spectra of fish diets at each station and the abun-
TABLE 5. Percentage Similarity Indices comparing adjusted trophic spectra among stations in Elkhorn Slough and ocean stations.
Bridge Dairy Kirby Creeks
OG%Ul
34.6 45.9 44.6 39.5
Bridge
67.2 67.9 52.6
Dairy
76.1 48.8
Kirby
72.5
dance of invertebrate prey was higher within sta- tions rather than between stations (Table 6; ANO- VA, F = 4.1, p < 0.10)) indicating predator forag- ing patterns were most similar to prey distributions at the location of capture.
ENVIRONMENTAL CONSTRAINTS ON FISH AND INVERTEBRATE DISTRIBUTION
Shifts in predator and prey abundance and dis- tribution from the ocean to upper slough were as-
TABLE 6. Similarity between adjusted trophic spectra for ichthyofaunal diets (left column) and invertebrate abundances estimated from core samples (top row), for stations where both datasets are available. Similarity between stations was signifi- candy lower (mean PSI = 57.0) than within stations (mean PSI = 72.0; ANOVA F = 4.1, p < 0.10).
Invertebrate Core Samples
Diet
+ion Bridge Dairy Kirby Park
Bridge 75.6 54.0 78.0 Dairy 58.0 63.5 56.6 Kirby Park 43.6 51.8 76.9

132 J. P. Barry et al.
sociated with and probably controlled by physical and biological factors. Variation in temperature, sa- linity, turbidity, and oxygen increases with distance inshore from the mouth of Elkhorn Slough (Broenkow 1977), and likely is important in regu- lating the diversity and abundance of food re- sources for both invertebrate and fish assemblages. Fluctuations in temperature and salinity may re- strict stenothermal and stenohaline species to low- er slough or ocean habitats, thereby decreasing di- versity of fishes and invertebrate prey. Thus, nar- row dietary breadth for fishes in the upper slough could be related to high abundance of preferred prey and reduced prey diversity.
The general decrease in invertebrate species richness with distance inshore (Nybakken and Jong 1977) is similar to patterns reported for other marine wetlands (Dahlberg 1972; Boesch 1977; Gainey and Greenberg 197; Weinstein et al. 1980; Smith et al. 1984). The average number of inver- tebrate species decreased from Moss Landing Har- bor to Kirby Park, due mainly to reduction in poly- chaete diversity. Crustacea diversity increased slightly. Thus, overall prey density generally was similar throughout Elkhorn Slough, but species composition and relative abundance were spatially and temporally variable. The fish assemblage likely tracks prey abundance by migration and feeding specialization, particularly when preferred prey are highly abundant, as observed at some upper slough sites.
Density of benthic invertebrates alone may be inadequate to define food availability, and rates of prey production may better indicate levels of en- ergy available for consumption by fishes. Temporal changes in invertebrate density include population and individual growth, in addition to mortality due to predation and other sources. Predation by fishes in Elkhorn Slough was undoubtedly highest during summer when total fish abundance was 10 to 25 times greater than winter (Yoklavich et al. 1991). In contrast, fish abundance at shallow inshore ocean stations increased only fivefold in summer.
Although predation pressure as related to vari- ation in fish abundance may affect seasonal pat- terns of prey abundance, further study is required to evaluate this hypothesis. Experiments by Quam- men (1984) in southern California indicated that predation by fishes had little effect on infaunal densities in intertidal habitats, with some impact by shorebird foraging. Subtidal prey populations likely experience higher rates of fish predation ow- ing to greater immersion time, but the magnitude of these effects may vary (Robinson and Wellborn 1988). Seasonal predation by fishes in Elkhorn Slough is balanced partially, at least in intertidal habitats, by migrating shorebird populations that
increase in number and biomass nearly tenfold during fall and winter (Ramer et al. 1991). Nev- ertheless, minor temporal changes in invertebrate density in the slough from 1974 to 1976 may un- derestimate seasonal patterns in prey production. Prey productivity and availability to predators are likely coupled to seasonal increases in primary pro- duction by marine plants, with peak growth rates during spring and summer.
The trophic patterns described herein for dom- inant fishes surveyed during the 1970s may be af- fected by recent changes occurring in Elkhorn Slough. Historically, the slough has experienced three episodes of significant hydrologic perturba- tion. Diversion of the Salinas River in 1910 de- creased sedimentation and freshwater input; the slough was a shallow embayment with minimal tid- al current and no intertidal mudflats (Oliver et al. unpublished data). Exposure to strong tidal flush- ing and erosion was initiated in 1947 with the con- struction of Moss Landing Harbor, which directly connected the slough to Monterey Bay. Intertidal mudflats emerged, salinity increased, and maxi- mum depth of the slough’s main channel in- creased from 1 m to 7 m (Oliver et al. unpublished data). Rates of tidal scour, erosion, and loss of salt marshes have increased since 1982 due to failure of dikes surrounding wetland pastures and salt ponds.
The main channel and tidal creeks of the slough continue to broaden and deepen, potentially alter- ing the species composition of fish predators and invertebrate prey. Additionally, the number of har- bor seals (Phoca vitulina richardsi), a voracious pis- civore, has increased threefold in Elkhorn Slough over the last decade (Oxman 1995), with peak abundance of seals coincident with that of fishes in summer months. A comparison of fish assem- blages in the main channel of Elkhorn Slough col- lected in the 1970s and in 1991 indicates a reduc- tion in species richness, dominance, and abun- dance (Oxman 1995). Dominant species of fishes largely remained the same over the ll-yr period, and major changes in rank order of abundance remained within feeding guilds (e.g., in the Epi- fauna1 Crustacea guild, relative abundance of Cith- arichthys stigmaeus and Leptocottous armatus in- creased while that of Cymatogaster aggregata and Em- biotoca jacksoni decreased; similar changes occurred in other guilds). Mean number of fishes per tow, however, has decreased twofold to twelvefold (Ox- man 1995), with potential consequences to overall prey densities.
RESOURCE PARTITIONING AMONG ELKHORN SLOUGH FISHES
The potential for interspecific competition for prey resources seems greatest during summer

Trophic Ecology of Fishes in Elkhorn Slough 133
when ichthyofaunal abundance peaks, yet little in- formation is available to evaluate its occurrence or effects. Four resident fishes (Clevelandia ios, Embi- otoca jacksoni, Gillichthys mirabilis, and Lqbtocottus ar- matus) within the Epifaunal Crustacea feeding guild (mean PSI for TS subcategories = 80.2; Table 2) fed on different prey taxa (mean PSI for de- tailed prey data = 17.0; Table 2). Thus, resource partitioning is possible, but high gut fullness (Am- brose 1976) suggests resource availability is high and competition low.
Overall, marine immigrants that enter the slough during summer had lower dietary similarity using trophic spectra, and slightly higher similarity using detailed prey data than did resident species. Dietary similarity for marine immigrant species within feeding guilds, however, was high (n = 2; mean = 71.1). This suggests that while resource partitioning may be important for resident species, species immigrating into Elkhorn Slough during summer use similar prey resources owing to high prey availability and low competition. If diets are shaped by competition, resource partitioning may be most apparent among species present during ‘competitive crunch’ periods (sensu Weins 1977) of lower food abundance (i.e., winter). Thus, tro- phic opportunism (Darnell 1961) is a more likely foraging mode, particularly for seasonal immi- grants occupying Elkhorn Slough as juveniles. Sim- ilar foraging patterns have been reported for sea- sonal populations in other estuaries and bays (Smith et al. 1984).
ACKNOWLEDGMENTS
We acknowledge many students at Moss Landing Marine Lab- oratories for help with field collections. This study was funded primarily by a grant from the National Sea Grant College Pro- gram, National Oceanic and Atmospheric Administration (NOAA), United States Department of Commerce, under grant number 048-MOl-189, project number R/CZ45B to G. Cailliet through the California Sea Grant College, and Pacific Gas and Electric Company to J. Nybakken, G. Cailliet, and W. Broenkow. The views expressed herein are those of the authors and do not necessarily reflect the views of NOAA or any of its subagencies. The United States Government is authorized to reproduce and distribute for governmental purposes. Additional funding was provided by NOAA, Sanctuary and Reserve Division (Contract No. NA89AA-D-CZ117) to J. Oliver, G. Cailliet, and M. Silber- stein, and by Elkhorn Slough Foundation for manuscript prep aration and publication. Comments by two anonymous review- ers improved the organization and content of our manuscript.
LITERATURE CITED
ALLEN, L. G. 1982. Seasonal abundance, composition, and pro- ductivity of the littoral fish assemblage in upper Newport Bay, California. Fishery Bulletin United States 80:769-790.
ALLEN, L. G. AND M. H. HORN. 1975. Abundance, diversity and seasonality of fishes in Colorado Lagoon, Alamitos Bay, Cali- fornia. Estuarine and Coastal Marine Science 3:371-380.
AMBROSE, D. A. 1976. The distribution, abundance, and feed- ing ecology of four species of flatfish in the vicinity of Elkhorn
Slough, California. M.A. Thesis, San Jose State University, San Jose, California.
ANTRIM, B. S. 1981. Habitat and food resource utilization of three species of embiotocids in Elkhorn Slough, Monterey Bay, California. M.A. Thesis, California State University, Fres- no, California.
BARRY, J. P. 1983. Utilization of shallow marsh habitats by fishes in Elkhorn Slough, California. M.A. Thesis, San Jose State University, San Jose, California.
BARRY, J. P. AND G. M. CAILLIET. 1981. The utilization of shallow marsh habitats by commercially important fishes in Elkhorn Slough, California. Cal-Neva Wild@ Transactions 1981:38-47.
BARRY, J. P. AND M. J. EHRET. 1993. Diet, food preference, and algal availability for fishes and crabs on intertidal reef com- munities in southern California. Environmental Biology ofFishes 37:75-95.
BOESCH, D. F. 1977. A new look at the zonation of benthos along the estuarine gradient, p. 245-266. In B. C. Coull (ed.), Ecology of Marine Benthos. University South Carolina Press, Columbia, South Carolina.
BOESCH, D. F. AND R. E. TURNER. 1984. Dependence of fishery species on salt marshes: The role of food and refuge. Estuaries 7:460-468.
BOZEMAN, E. L., JR. AND J. M. DEAN. 1980. The abundance of estuarine larval and juvenile fish in a South Carolina intertid- al creek. Estuaries 3:89-97.
BREGNBALLE, F. 1961. Plaice and flounder as consumers of the microscopic bottom fauna. Meddelelser pa Danmarks Fiskwi-og Havundersogels~ 3:133-182.
BROENKOW, W. 1977. Water chemistry of Elkhorn Slough and Moss Landing Harbor, p. 387-465. In J. Nybakken, G. Cailliet, and W. Broenkow (eds.), Ecologic and Hydrographic Studies of Elkhorn Slough Moss Landing Harbor and Nearshore Coastal Waters. Moss Landing Marine Laboratories Technical Publication, Moss Landing, California.
BROWNING, B. M. 1972. The natural resources of Elkhorn Slough: Their present and future use. Cal@rnia Department of Fish and Game, Coastal Wetlands Series 4.
CAILLIET, G. M., B. S. ANTRIM, AND D. A. AMBROSE. 1978. Tro- phic spectrum analysis of fishes in Elkhorn Slough and near- by waters, p. 118-128. In S. J. Lipovsky and C. A. Simenstad (eds.), Fish Food Habits Studies, Proceedings of the Second Pacific Northwest Technical Workshop. University of Wash- ington, Washington Sea Grant WSGWO-77-2. Seattle, Wash- ington.
CAILLIET, G. M., B. ANTRIM, D. AMBROSE, S. PACE, AND M. STE- VENSON. 1977. Species composition, abundance and ecolog- ical studies of fishes, larval fishes, and zooplankton in Elk- horn Slough, p. 216-386. In J. Nybakken, G. Cailliet, and W. Broenkow (eds.), Ecologic and Hydrographic Studies of Elk- horn Slough Moss Landing Harbor and Nearshore Coastal Waters. Moss Landing Marine Laboratories Technical Publi- cation, Moss Landing, California.
CAILLIET, G. M. AND J. P. BARRY. 1978. Comparison of food array overlap measures useful in fish feeding habit analysis, p. 67- 79. In S. J. Lipovsky and C. A. Simenstad (eds.), Fish Food Habits Studies, Proceedings of the Second Pacific Northwest Technical Workshop. University of Washington, Washington Sea Grant WSGWO-77-2. Seattle, Washington.
CARPENTER, S. R. PD J. F. KITCHELL. 1992. Trophic cascade and biomanipulation: Interface of research and management-A reply to the comment by DeMelo et al. Limnology and Ocean- ography 37:208-213.
CARR, W. E. S. AND J. T. GEISEL. 1975. Impact of thermal efflu- ent from a steam-electric station on a marshland nursery area during the hot season. Fishery Bulletin United States 73:67-80.
CRAMPTON, T. A. 1994. Long-term effects of Moss Landing Har- bor on the wetlands of Elkhorn Slough. M.S. Thesis, Univer- sity of California, Santa Cruz, California.

134 J. P. Barry et al.
DAHLBERG, M. D. 1972. An ecological study of Georgia coastal fishes. Fishery Bulletin United States 70:323-353.
DANDO, P. R. 1984. Reproduction in estuarine fish, p. 155-170. In G. W. Potts and R. J. Wootton (eds.), Fish Reproduction Strategies and Tactics. Academic Press, London.
DARNELL, R. M. 1958. Food habits of fishes and larger inver- tebrates of Lake Pontchartrain, Louisiana, an estuarine com- munity. Publications of the Zwtitute of Marine Science, University of Tocas 5:353-416.
DARNELL, R. M. 1961. Trophic spectrum of an estuarine com- munity, based on studies of Lake Pontchartrain, Louisiana. Ecology 42:553-568.
DARNELL, R. M. AND T. M. SONIAT. 1979. The Estuary/Conti- nental Shelf as an Interactive System. In R. J. Livingston (ed.), Ecological Processes in Coastal and Marine Systems. Plenum Press, New York.
DAY, J. W., JR. AND A. YANEZ-ARANCIBIA. 1985. Coastal lagoons and estuaries as an environment for nekton, Chapter 3, p. 17-34. In A. Yanez-Arancibia (ed.), Fish Community Ecology in Estuaries and Coastal Lagoons: Toward an Ecosystem In- tegration. DR(R) UNAM Press Mexico, Mexico City.
GAINEY, L. F., JR. AND M. J. GREENBERG. 1977. Physiological basis of the species abundance-salinity relationship in molluscs: A speculation. Matine Biology (Berl.) 40:41-49.
GOBALET, K. W. 1990. Prehistoric status of freshwater fishes of the Pajaro-Salinas River system of California. Cop&a 1990:680- 685.
GOBALET, K. W. 1992. Inland utilization of marine fishes by Native Americans along the central California coast. Jownal of Calzjknia and Great Basin Anthropology 14:72-84.
GUNDERSON, D. R., D. A. ARMSTRONG, Y-B. SHI, AND R. A. MC~ONNAUGHEY. 1990. Patterns of estuarine use by juvenile English sole (Parophrys vetulus) and Dungeness crab (Cancer mug&m). Estuaries 13:59-71.
GUNTER, G. 1938. Seasonal variations in abundance of certain estuarine and marine fishes in Louisiana, with particular ref- erence to life histories. Ecological Monographs 8:313-345.
HACKNEY, C. T. AND E. B. ONES. 1980. Stable carbon isotope composition of fauna and organic matter collected in a Mis- sissippi estuary. Estuarine and Coastal Marine Science 10:703- 708.
HAERTEL, L. AND C. OSTE.RBERG. 1967. Ecology of zooplankton, benthos and fishes in the Columbia River estuary. Ecoloa 48: 459-472.
HAINES, E. B. 1979. Interactions between Georgia salt marshes and coastal waters: A changing paradigm, p. 35-46. In R. J. Livingston (ed.), Ecological Processes in Coastal and Marine systems. Plenum Press, New York.
HAINES, E. B. AND C. L. MONTAGUE. 1979. Food sources of es- tuarine invertebrates analyzed using ‘3C/‘2C ratios. Ecology 60: 48-56.
HARVEY, J. T., R. HELM, AND G. V. MOREJOHN. In press. Food habits of harbor seals inhabiting Elkhorn Slough, California. Cal~omia Department of Fish and Game Fish Bulletin
HAY, M. E. 1994. Species as ‘noise’ in community ecology: Do seaweeds block our view of the kelp forest? Trends In Ecology and Evolution 9:414-416.
HOFF, J. G. AND R. M. IBARA. 1977. Factors affecting the sea- sonal abundance, composition and diversity of fishes in a southeastern New England estuary. Estuutine and Coastal Ma- rine Science 5:665-678.
HORNBERGER, M. I. 1991. Paleoenvironment of Elkhorn Slough and surrounding wetland habitats: A geological study using an ecological approach. M.S. Thesis, Moss Landing Marine Laboratories and San Jose State University, California.
RREBS, C. J. 1989. Ecological Methodology. Harper and Row, Publishers, New York.
KRYGIER, E. E. AND W. G. PEARCY. 1986. The role of estuarine and offshore nursery areas for young English sole, Parophrys
vet&us Girard, off Oregon. Fishery Bulletin United States 84:119- 132.
LANGTON, R. W. AND L. WATLING. 1990. The fish-benthos con- nection: A definition of prey groups in the Gulf of Maine, p. 424-438. In M. Barnes and R. N. Gibson (eds.), Trophic Re- lationships in the Marine Environment, Proceedings 24th Eu- ropean Marine Biology Symposium, Aberdeen University Press, Scotland.
LEVINGS, C. D. 1974. Seasonal changes in feeding and particle selection by winter flounder (Pseudopleuronectes am&anus). Transactions of the American Fisheries Society 103:828-832.
kfARINUCC1, A. C. 1982. Trophic importance of Spurtina alter- nzflora production and decomposition to the marsh-estuarine ecosystem. Biological Conservation 22:35-58.
MCERLEAN, A. J., S. G. O’CONNOR, J. A. MIHURSKY, AND C. I. GIBSON. 1973. Abundance, diversity and seasonal patterns of estuarine fish populations. Estuatilze and Coastal Marine Science 1:19-36.
MCGOWAN, M. F. 1986. Northern anchovy, Engmulis mordax, spawning in San Francisco Bay, California, 1978-79, relative to hydrography and zooplankton prey of adults and larvae. Fish- e~y Bulletin United States 84879-894.
MILLER, B. S. 1967. Stomach contents of adult starry flounder and sand sole in East Sound, Orcas Island, Washington. Jour- nal of the Fisheries Research Board of Canada 242515-2526.
NAIMAN, R. J. AND J. R. SIBERT. 1979. Detritus and juvenile salm- on production in the Nanaimo Estuary: III. Importance of detrital carbon to ‘the estuarine ecosystem. Journal of the Fish- eries Research Board of Canada 36:504-520.
NIXON, S. W. 1980. Between coastal marshes and coastal wa- ters-A review of twenty years of speculation and research on the role of salt marshes in estuarine productivity and water chemistry, p. 437-525 In P. Hamilton and K. B. MacDonald (eds.), Estuarine and Wetland Processes. Plenum Press, New York.
NYBAKKEN, J. W. 1987. Marine Biology: An Ecological Approach, second ed. Harper and Row, New York.
NYB~N, J. AND C. JONG. 1977. Benthic invertebrate baseline studies of the Moss Landing-Elkhorn Slough environment, p. 1-215. In J. Nybakken, G. Cailliet, and W. Broenkow (eds.), Ecologic and Hydrographic Studies of Elkhorn Slough Moss Landing Harbor and Nearshore Coastal Waters. Moss Land- ing Marine Laboratories Technical Publication, Moss Land- ing, California.
ODUM, E. P. 1971. Fundamentals of Ecology, third ed. W. B. Saunders, Co., Philadelphia.
ODUM, W. E. AND E. J. HF.ALD. 1975. The detritus-based food web of an estuarine mangrove community, p. 265-286. In L. E. Cronin (ed.), Estuarine Research, Vol. 1. Academic Press, New York.
ONUF, C. P. AND M. L. QUAMMEN. 1983. Fishes in a California coastal lagoon: Effects of major storms on distribution and abundance. Marine Ecology Progress Series 12:1-14.
ORCUTT, H. G. 1950. The life history of the starry flounder, Platichthys stellatus (Pallas). Calij&aia Department of Fish and Game Fish Bulletin 78:1-64.
OXMAN, D. S. 1995. Seasonal abunda%ce, movements, and food habits of harbor seals (Phoca vitulina richardsi) in Elkhorn Slough, California. MS. Thesis, California State University, Stanislaus, California.
PEARCY, W. G. AND S. S. MYERS. 1974. Larval fishes of Yaquina Bay, Oregon: A nursery ground for marine fishes? Fishery Bul- letin United States 72:201-213.
PINKAS, L., M. S. OLIPHANT, AND I. L. K. IVERSON. 1971. Food habits of albacore, bluefin tuna, and bonito in California wa- ters. Calijbrnia Department of Fish and Game Fish Bulletin 152.
QUAMMEN, M. L. 1984. Predation by shorebirds, fish, and crabs on invertebrates in intertidal mudflats: An experimental test. Ecobgy 65:529-537.

RAFFAELLI, D. AND S. J. HALL. 1992. Compartments and pre- dation in an estuarine food web. Journal of Animal Ecology 61: 551-560.
RAMER, B.A., G. W. PAGE, AND M. M. YOKLAVICH. 1991. Seasonal abundance, habitat use, and diet of shorebirds in Elkhorn Slough, California. Western Birds 22:157-174.
RIBELIN, B. W. AND A. W. COLLIER. 1979. Ecological considera- tion of detrital aggregates in the salt marsh, p. 47-68. In R. J. Livingston (ed.), Ecological Processes in Coastal and Marine Systems. Plenum Press, New York.
ROBINSON, J. V. AND G. A. WELLBORN. 1988. Ecological resis- tance to the invasion of a freshwater clam, Corbicula$uminea: Fish predation effects. Oecologia 77:445-452.
RUAGH, A. A. 1976. Feeding habits of silversides (family Ath- erinidae) in Elkhorn Slough, Monterey Bay, California. M.A.
‘Thesis, California State University, Fresno, California. SCHWARTZ, D. L. 1983. Geologic history of Elkhorn Slough,
Monterey County, California. M.S. Thesis, Moss Landing Ma- rine Laboratories and San Jose State University, San Jose, Cal- ifornia.
SCHWARTZ, D. L., H. T. MULLINS, AND D. F. BELKNAP. 1986. Ho- locene geologic history of a transform margin estuary: Elk- horn Slough, central California. Estuarine, Coastal and Shelf Science 22:285-302.
SHANNON, C. E. AND W. WEAVER. 1963. The Mathematical The- ory of Communication. University of Illinois Press, Urbana, Illinois.
SHENKER, J. M. AND J. M. DEAN. 1979. The utilization of an intertidal salt marsh creek by larval and juvenile fishes; abun- dance, diversity, and temporal variation. Estuaries 2:154-163.
SIMPSON, E. H. 1949. Measurement of diversity. Nature 163:688. SMITH, R. E. 1973. The hydrography of Elkhorn Slough, a shal-
low California coastal embayment. Moss Landing Marine Lab- oratories Technical Publication 73-2, Moss Landing, Califor- nia.
SMITH, S. M., J. G. HOFF, S. P. O’NEIL, AND M. P. WEINSTEIN. 1984. Community and trophic organization of nekton utiliz- ing shallow marsh habitats, York River, Virginia. Fishery Bulletin Uked States 82:455-467.
SESTAT. 1992. SYSTAT for Windows: Statistics, Version 5 Edi- tion. Systat, Inc. Evanston, Illinois.
TENORE, K R. AND D. L. RICE. 1980. A review of trophic factors affecting secondary production of deposit feeders, p. 325- 340. In Marine Benthic Dynamics. University of South Caro- lina Press, Columbia, South Carolina.
Trophic Ecology of Fishes in Elkhorn Slough 135
TOOLE, C. L. 1980. Intertidal recruitment and feeding in rela- tion to optimal utilization of nursery areas by juvenile English sole (Parophrys vetulus). Environmental Biolqy of Fishes 5:383- 390.
VANNI, M. J., C. LUEC&, J. F. KITCHELL, Y. ALLEN, J. TEMTE, AND J. J. MAGNUSON. 1990. Effects on lower trophic levels of mas- sive fish mortality. Nature 344:333-335.
VETTER, E. W. 1994. Hotspots of benthic production. Nature 372:47.
WEINS, J. A. 1977. On competition ip variable environments. American Scientist 65:590-597.
WEINSTEIN, M. P. 1979. Shallow marsh habitats as primary nurs- eries for fishes and shellfish, Cape Fear River, North Carolina. Fishery Bulletin United States 77:339-357.
WEINSTEIN, M. P., S. L. WEISS, AND M. F. WALTERS. 1980. Mul- tiple determinants of community structure in shallow marsh habitats, Cape Fear River estuary, North Carolina. Marine Bi- o&y (Berl.) 58:227-243.
WERNER, E. E. AND J. F. GILLIAM. 1984. The ontogenetic niche and species interactions in size-structured populations. An- nual Review Ecology and Systemmatics 15:393-425.
WILLIAMS, P. AND ASSOCIATES AND MOFFATT AND NICHOL ENGI- NEERS. 1992. Elkhorn Slough tidal hydraulics erosion study. Prepared for United States Army Corps of Engineers, San Francisco District, San Francisco, California.
YOKLAVICH, M. M. 1982. Growth, food consumption, and con- version efficiency of juvenile English sole (Parophrys vetulus), p. 97-105. In G. M. Cailliet and C. A. Simenstad (eds.), Fish Food Habit Studies, Prqceedings Third Pacific Workshop. University of Washington, Washington Sea Grant. Seattle, Washington.
YOKLAVICH, M. M., G. M. CAILLIET, J. P. BARRY, D. A. AMBROSE, AND B. S. ANTFUM. 1991. Temporal and spatial patterns in abundance and diversity of fish assemblages in Elkhorn Slough, California. Estuaries 14465-480.
YOKLAVICH, M. M., M. STEVENSON, AND G. M. CAILLIET. 1992. Seasonal and spatial patterns of ichthyoplankton abundance in Elkhorn Slough, California. Estuatine, Coastal and Shelf Sci- ence 34:109-126.
ZEDLER, J., T. WINFIELD, AND D. MAURIELLO. 1978. Primary pro- ductivity in a southern California estuary. Coastal Zone 3:649- 662.
Received for consideration, July 5, Accepted for publication, Apn’l 17,
1994 1995

136 J. P. Barry et al.
“9 9 wo 0
091o-!G? 2000
9 0 qy0?- MOdA i
” 0
r( 0 d 6
9 0

Trophic Ecology of Fishes in Elkhorn Slough 137
9 0
9 0
0 d 9 9 0 0
9 0
0 6
I- ” 0
68 ei6
99 moo 00 c66d
‘? M)
9 0999 0 000
r: 9 T! zoo
9 0 9 0
9 00 0 dd
0 0 d 6
” 0
9 0
h t.
6 cri 9 0

138 J. P. Barry et al.
9 0 9 0
0 6