torices a, barroso-barcenilla f, cambra-moo o, pérez-garcía a, segura m. 2012. palaeontological...
TRANSCRIPT

at SciVerse ScienceDirect
Cretaceous Research 37 (2012) 231e239
Contents lists available
Cretaceous Research
journal homepage: www.elsevier .com/locate/CretRes
Palaeontological and palaeobiogeographical implications of the new Cenomanianvertebrate site of Algora, Guadalajara, Spain
Angélica Torices a,b,*, Fernando Barroso-Barcenilla b,c, Oscar Cambra-Moo d, Adán Pérez-García b,Manuel Segura c
aDepartment of Biological Sciences, Faculty of Science, CW405 Biological Sciences Building, University of Alberta, Edmonton, Alberta, Canada T6G 2E9bDepartamento de Paleontología, Facultad de Ciencias Geológicas, Universidad Complutense de Madrid, 28040 Madrid, SpaincGrupo de Investigación IBERCRETA, Universidad de Alcalá de Henares, 28871 Alcalá de Henares, SpaindDepartamento de Biología, Facultad de Ciencias, Universidad Autónoma de Madrid, 28049 Madrid, Spain
a r t i c l e i n f o
Article history:Received 6 October 2011Accepted in revised form 5 April 2012Available online 6 May 2012
Keywords:Late CretaceousCenomanianVertebrate palaeontologyPalaeobiogeographySpain
* Corresponding author. Department of BiologicalCW405 Biological Sciences Building, University ofCanada T6G 2E9.
E-mail addresses: [email protected], atorices@ge
0195-6671/$ e see front matter � 2012 Elsevier Ltd.doi:10.1016/j.cretres.2012.04.004
a b s t r a c t
A new Cenomanian fossil site has been located in Algora, in the Castilian Branch of the Iberian Ranges,central Spain. The outcrop is in the upper part of the Utrillas Formation in sediments that can beinterpreted as sandy coastal deposits (bars and channels) with subtidal and intertidal events. Thevertebrate fauna associated with this site is composed of mixed remains of fishes, turtles, crocodyliformsand dinosaurs, with Laurasian and Gondwanan affinities. The discovery of this site provides newpalaeobiogeographical information and enriches the poorly known vertebrate faunas of the EuropeanCenomanian.
� 2012 Elsevier Ltd. All rights reserved.
1. Introduction
Vertebrate remains from the Cenomanian are very scarce in theIberian Peninsula compared to the record for BerriasianeAlbianand CampanianeMaastrichtian intervals (López-Martínez et al.,2001; Barroso-Barcenilla et al., 2009). Previous works on theIberian Peninsula have reported coastal assemblages in Estrema-dura, Portugal (Jonet, 1981) and mixed assemblages in Asturias,Spain (Ruiz-Omeñaca et al., 2009; Vullo et al., 2009). In the rest ofEurope, the best documented vertebrate assemblage from this timeinterval comes from the localities of Charentes, France (Vullo et al.,2005; Vullo, 2007; Vullo and Néraudeau, 2008), but there are otherisolated findings as well (Buffetaut, 1989; Averianov et al., 2005;Fejfar et al., 2005). In this paper, we report the first results of ananalysis of a new vertebrate fauna from the Cenomanian in Algora,Spain. These remains are the first Cenomanian vertebrate fossils tobe found in the Iberian Ranges and they increase the limitedknowledge of these faunas, revealing new information aboutpalaeogeographical relationships between Laurasian and Gond-wanan assemblages.
Sciences, Faculty of Science,Alberta, Edmonton, Alberta,
o.ucm.es (A. Torices).
All rights reserved.
2. General setting
2. 1. Stratigraphy
This new Cenomanian vertebrate site, as noted by Segura et al.(2010) and Torices et al. (2010), is located in the district of Algora(Castilian Branch of the Iberian Ranges, province of Guadalajara,central Spain; Fig. 1) and, stratigraphically, is situated in the upperpart of the “Arenas de Utrillas” Formation (Aguilar et al., 1971;sensu Gil et al., 2004; Fig. 2). In Algora and the surrounding area,this terrigenous lithostratigraphic unit is represented by an intervalof sandstones 80e100 m thick (Segura et al., 1985; García et al.,1993). It overlies unconformably Jurassic and, to a lesser extent,Lower Cretaceous rocks. The Utrillas Formation changes laterallysoutheastwards, by means of a gradational boundary, to 75e85 mof limestones and dolostones with benthic foraminifera of thelower upper Cenomanian “Dolomías de Villa de Vés” Formation(Vilas et al., 1982). At the top, above a sharp boundary (reflectinga transgression), it is overlain by 10e15 m of limestones anddolostones with ammonoids of the uppermost CenomanianelowerTuronian “Calizas y Margas de Picofrentes” Formation (Floquetet al., 1982). Eastwards and southeastwards, the Utrillas Forma-tion comprises interbedded sandstones with chalky cement, dolo-stones and limestones with benthic foraminifera belonging to the
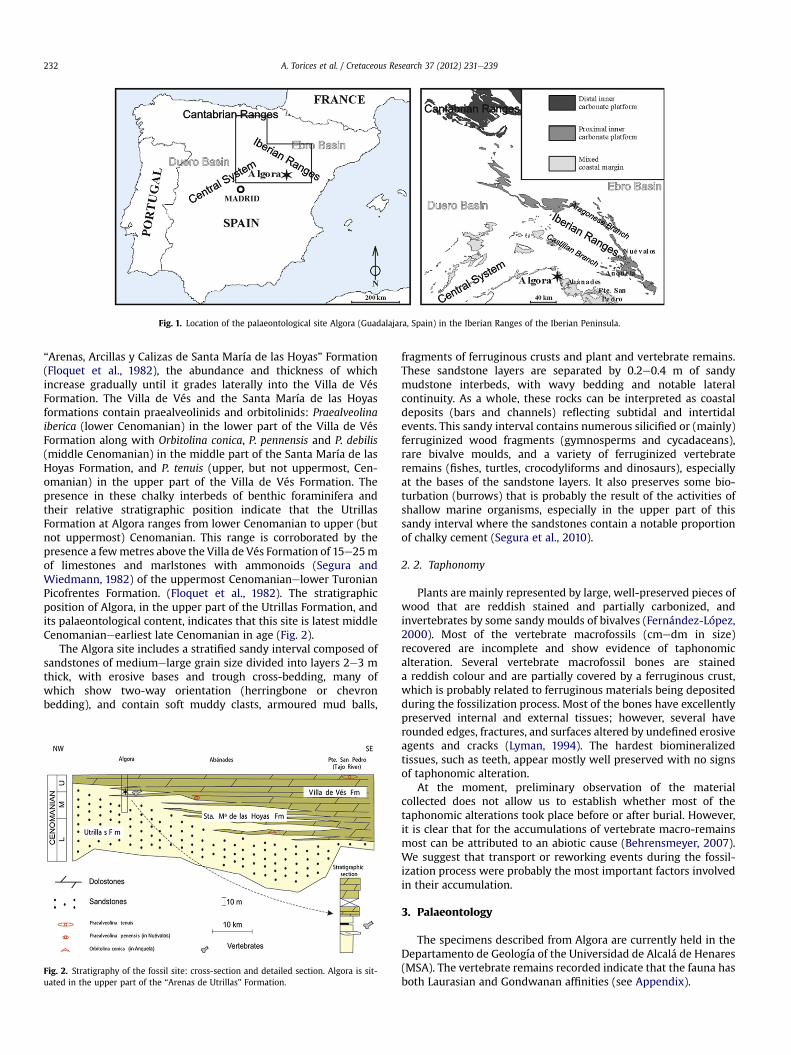
Fig. 1. Location of the palaeontological site Algora (Guadalajara, Spain) in the Iberian Ranges of the Iberian Peninsula.
A. Torices et al. / Cretaceous Research 37 (2012) 231e239232
“Arenas, Arcillas y Calizas de Santa María de las Hoyas” Formation(Floquet et al., 1982), the abundance and thickness of whichincrease gradually until it grades laterally into the Villa de VésFormation. The Villa de Vés and the Santa María de las Hoyasformations contain praealveolinids and orbitolinids: Praealveolinaiberica (lower Cenomanian) in the lower part of the Villa de VésFormation along with Orbitolina conica, P. pennensis and P. debilis(middle Cenomanian) in the middle part of the Santa María de lasHoyas Formation, and P. tenuis (upper, but not uppermost, Cen-omanian) in the upper part of the Villa de Vés Formation. Thepresence in these chalky interbeds of benthic foraminifera andtheir relative stratigraphic position indicate that the UtrillasFormation at Algora ranges from lower Cenomanian to upper (butnot uppermost) Cenomanian. This range is corroborated by thepresence a fewmetres above the Villa de Vés Formation of 15e25mof limestones and marlstones with ammonoids (Segura andWiedmann, 1982) of the uppermost Cenomanianelower TuronianPicofrentes Formation. (Floquet et al., 1982). The stratigraphicposition of Algora, in the upper part of the Utrillas Formation, andits palaeontological content, indicates that this site is latest middleCenomanianeearliest late Cenomanian in age (Fig. 2).
The Algora site includes a stratified sandy interval composed ofsandstones of mediumelarge grain size divided into layers 2e3 mthick, with erosive bases and trough cross-bedding, many ofwhich show two-way orientation (herringbone or chevronbedding), and contain soft muddy clasts, armoured mud balls,
Fig. 2. Stratigraphy of the fossil site: cross-section and detailed section. Algora is sit-uated in the upper part of the “Arenas de Utrillas” Formation.
fragments of ferruginous crusts and plant and vertebrate remains.These sandstone layers are separated by 0.2e0.4 m of sandymudstone interbeds, with wavy bedding and notable lateralcontinuity. As a whole, these rocks can be interpreted as coastaldeposits (bars and channels) reflecting subtidal and intertidalevents. This sandy interval contains numerous silicified or (mainly)ferruginized wood fragments (gymnosperms and cycadaceans),rare bivalve moulds, and a variety of ferruginized vertebrateremains (fishes, turtles, crocodyliforms and dinosaurs), especiallyat the bases of the sandstone layers. It also preserves some bio-turbation (burrows) that is probably the result of the activities ofshallow marine organisms, especially in the upper part of thissandy interval where the sandstones contain a notable proportionof chalky cement (Segura et al., 2010).
2. 2. Taphonomy
Plants are mainly represented by large, well-preserved pieces ofwood that are reddish stained and partially carbonized, andinvertebrates by some sandy moulds of bivalves (Fernández-López,2000). Most of the vertebrate macrofossils (cmedm in size)recovered are incomplete and show evidence of taphonomicalteration. Several vertebrate macrofossil bones are staineda reddish colour and are partially covered by a ferruginous crust,which is probably related to ferruginous materials being depositedduring the fossilization process. Most of the bones have excellentlypreserved internal and external tissues; however, several haverounded edges, fractures, and surfaces altered by undefined erosiveagents and cracks (Lyman, 1994). The hardest biomineralizedtissues, such as teeth, appear mostly well preserved with no signsof taphonomic alteration.
At the moment, preliminary observation of the materialcollected does not allow us to establish whether most of thetaphonomic alterations took place before or after burial. However,it is clear that for the accumulations of vertebrate macro-remainsmost can be attributed to an abiotic cause (Behrensmeyer, 2007).We suggest that transport or reworking events during the fossil-ization process were probably the most important factors involvedin their accumulation.
3. Palaeontology
The specimens described from Algora are currently held in theDepartamento de Geología of the Universidad de Alcalá de Henares(MSA). The vertebrate remains recorded indicate that the fauna hasboth Laurasian and Gondwanan affinities (see Appendix).

A. Torices et al. / Cretaceous Research 37 (2012) 231e239 233
3.1. Plants
Plant remains consist mainly of numerous fragments of wood. Insome parts of the site, many sections of large trunks are present andcan be assigned, on the basis of their overall appearance, togymnosperms and cycadaceans (Fig. 3A).
3.2. Invertebrates
Few remains of invertebrates have been collected. These areconfined to moulds of undetermined bivalves (Fig. 3B).
3.3. Osteichthyans
Fish remains are represented by ganoid scales. One group ofthese scales is coarsely folded (Fig. 3C) and can be attributed toStromerichthys sp., a taxon of uncertain taxonomic affinities,although it is commonly referred to as an amiid (Dutheil, 1999;Vullo and Néraudeau, 2008) even though ganoid ridges can befound in the scales of some lepisosteid gars (Buffetaut et al., 1996;Cavin et al., 1996; Gottfried and Krause, 1998; Vullo and Néraudeau,2008). The other group of ganoid scales has four parallel ridges onan enamelled external surface. This ornamentation suggests thatthey could belong to semionotids or lepisosteids, or might alsobelong to Stromerichthys sp. (Vullo et al., 2009) (Fig. 3D).
3.4. Chelonians
Remains of turtles are very abundant and are represented byboth carapace and plastron plates. Although these elements aredisjointed and generally fragmentary, representatives of twogroups of turtles are recognized. One of these (Fig. 3EeI), the mostabundant, can be assigned to Pan-Pleurodira because it sharesseveral characters with the diagnosis of this node (sensu Gaffneyet al., 2006): it has a sutural articulation of the ilium with thecostal plates, and a sutural articulation of the pubis and ischiumwith the xiphiplastron; and the morphology of the ilium scarsuggests that the dorsal part of this bone is broad and robust, beingclose to the midline. The material analyzed has a reduced first rib;a long triangular ilion scar; hexagonal neural plates (from thesecond to the last), with anterolateral contacts shorter thanposterolateral contacts; and an ornament pattern consisting ofweakly granulated polygons. These characters are shared with theeupleurodiran groups Chelidae and Pelomedusoides (de Lapparentde Broin and Murelaga, 1999; de Lapparent de Broin and Molnar,2001; Gaffney et al., 2006). These plates differ from Dortokidaebecause they do not share several characters of their diagnosis (deLapparent de Broin et al., 2004) such as the presence of a strongmedial ornamentation of tubercles and pits, crests and ridges atleast on the neural plates, or longitudinal-oblique crests on someregions of the plastron; the first pleural scute being at least partlyexcluded from the second costal plate; and the presence of longischiatic scars. However, owing to the limited information availableon the Cretaceous pan-pleurodiran groups present in Europe beforethe Santonian, and to the lack of sufficient characters in the spec-imens analyzed, these representatives of Pan-Pleurodira areprovisionally allocated to Eupleurodira. The problems resultingfrom the analysis of scattered and fragmentary specimens do notallow confirmation of their assignment to a single taxon.
The other turtles recognized from Algora are referred toSolemydidae (Stem Testudines sensu Joyce et al., 2011) (Fig. 3J). Theirornamentation, with granulation, allows their assignment to thisgroup (de Lapparent de Broin andMurelaga,1999), the earliest recordof which is from Late Jurassic deposits in Europe. As with Dortokidaeor Pelomedusoides, this group is very common in European
uppermost Cretaceous sites. Solemydidae are also identified in theCretaceous of North America, between the Aptian and Campanian(Joyce et al., 2011). In theCenomanianof Europe, it has been identifiedin several sites in Charentes, southwestern France (Vullo et al., 2005,2010; Vullo, 2007; Vullo and Néraudeau, 2008). The relative distancebetween each granulation or group of granulations appears to begreater in the Algora plates than in morphotype A identified in theCharentes fauna (Vullo et al., 2010). This distance between thegranulations observed in thematerial fromAlgora is similar to that ofthe second morphotype from Charentes but differs from it becausesome tubercles are fused. However, our limited knowledge of repre-sentatives of European Cenomanian solemydids, which are onlyrepresented by fragmentary elements of the shell, has prevented anytaxon from being identified at the generic level (Vullo et al., 2010).Therefore, the Algora material is referred to Solemydidae indet.
3.5. Crocodyliforms
Late Cretaceous crocodyliformsmainly consist of representativesof the Eusuchia. However, more primitive taxa are known fromsome late Late Cretaceous sites (Martin et al., 2010; Martin andDelfino, 2010). Currently, the early Late Cretaceous crocodyliformsare extremely poorly known; they are represented by non-eusuchian Neosuchia and Eusuchia from southwestern France andSpain (Vullo et al., 2005, 2009). The currently available specimensfromAlgora, represented by osteoderms, scarce fragments of bones,and teeth (Fig. 4AeE), show no particular diagnostic features. Theirknown combination of characters is compatible with that ofadvanced neosuchians: conical teeth, slightly ornamented bya pattern of smooth ridges, with mesial and distal carinae lackingdenticles or crenulations. The presence of a pitted ornamentalpattern has been considered a synapomorphy of Neosuchia þAraripesuchus (Ortega et al., 2000). Within Neosuchia, the presenceof sub-square dorsal osteoderms suggests that the Algora specimenmay be referable to Eusuchia or Neosuchia. Taking into account thescarce evidence for European crocodyliforms from the late Barre-mian until the beginning of the Campanian (Buscalioni et al., 2001;Martin and Delfino, 2010), the process of the replacement of morebasal Neosuchia by Eusuchia representatives in the Europeanecosystems through the Cretaceous is still poorly understood(Martin and Delfino, 2010), and in this context, more remains fromthe Algora site could be especially noteworthy.
3.6. Dinosaurs
Three theropod teeth have been found and assigned to Carch-arodontosauridae indet. (Fig. 4I, J). They are laterally compressedand the apex of the crown is slightly curved distally. Their height isbetween 2.5 and 4 cm and their fore-aft-basal-length (FABL) isbetween 1 and 1.7 cm. They have denticles on both carinae. Thedenticles on the mesial carina extend to the base of the crown.Denticle density is 2.5e3 denticles per mm on the distal carina and3 denticles per mm on the mesial carina. The denticles are chisel-like, and they have slightly oblique blood grooves between theirbases. The enamel is rough and exhibits arcuate wrinkles. InEurope, theropod teeth from the Cenomanian have been found inAsturias, northern Spain, and Charentes, southwestern France(Vullo, 2007; Ruiz-Omeñaca et al., 2009). Themorphometric data ofthe Algora teeth are compatible with those described by Vullo(2007) for the teeth classified as Carcharodontosauridae indet. inthe Charentes sites. They have an ornament of arcuate wrinkles incommon with the teeth of Carcharodontosaurus saharicus (Serenoet al., 1996). Although enamel wrinkles have been found in othertheropod groups besides carcharodontosaurids (Brusatte et al.,2007), it seems that there are morphological differences that may

Fig. 3. A, MSA-001, fragment of wood. B, MSA-002, internal mould of bivalve. C, MSA-003, osteichthyan scale (Stromerichthys sp.). D, MSA-004, osteichthyan scale. E, MSA-005, cf.Eupleurodira, first costal plate in ventral view. F, MSA-006, cf. Eupleurodira, neural plate in dorsal view. G, MSA-007, cf. Eupleurodira, last costal plate in ventral view. H, MSA-008, cf.Eupleurodira, peripheral plate in ventral view. I, MSA-009, cf. Eupleurodira, anterior region of the xiphiplastron in dorsal view, showing the partial scars of the ischium (posterior)and pubis (anterior). J, MSA-010, fragment of a plate of a turtle identified as Solemydidae indet.
A. Torices et al. / Cretaceous Research 37 (2012) 231e239234

Fig. 4. A, MSA-011, Neosuchia indet., probably a fragment of the fronto-parietal area of the skull table in dorsal view. B, MSA-011, Neosuchia indet., probably a fragment of thefronto-parietal area of the skull table in ventral view. C, MSA-014, Neosuchia indet., tooth. D, E, MSA-012 and MSA-013, Neosuchia indet., osteoderms. F, MSA-019, coprolite. G, H,MSA-015 and MSA-016, unidentified vertebrate bones. I, J, MSA-017 and MSA-018, Carcharodontosauridae indet., teeth with arcuate wrinkles.
A. Torices et al. / Cretaceous Research 37 (2012) 231e239 235
be diagnostic of different theropod groups. In fact, enamel wrinklesin carcharodontosaurids seem to be more pronounced near theserrations and can continue across the crown as shallow bands(Brusatte et al., 2007). The Asturian tooth is fragmentary, but thedenticles are slightly larger than those present in Algora andCharentes, and the tooth lacks enamel wrinkles (Ruiz-Omeñacaet al., 2009). Although there is a difference in the number ofdenticles, this is minimal and could be considered to fall within thenormal range of intraspecific variability (Torices, 2007). However,owing to its fragmentary condition and lack of ornamentation,Ruiz-Omeñaca et al. (2009) classified the Asturian tooth as Ther-opoda indet. The teeth from Algora and Charentes seem to belong
to the same type of theropod dinosaur, and it is hoped that newfinds of more complete material in Asturias will also confirm thepresence of the Carcharodontosauridae at that site in the future.
Large isolated, altered and incomplete fossil bones have beenrecovered from the site. Although their size could suggest that theybelong to dinosaurs they are currently regarded as unidentifiedvertebrate bones (Fig. 4G, H).
3.7. Trace fossils
Several coprolites have been found at Algora (Fig. 4F). Some-times, coprolites can be difficult to distinguish from nodules, but
A. Torices et al. / Cretaceous Research 37 (2012) 231e239236
a distinctive helicoid shape has been an important characteristic fortheir determination. Helically-coiled coprolites suggest an intestineanatomy that is fairly common in both chondrichthyan andosteichthyan fishes (McAllister, 1985; Coy, 1995). The coprolites arearound 3 cm in length. Some have an elliptical outline, while othershave a more rounded shape. Their surface is reddish in colourowing to iron oxide staining and is unornamented.
4. Palaeobiogeography
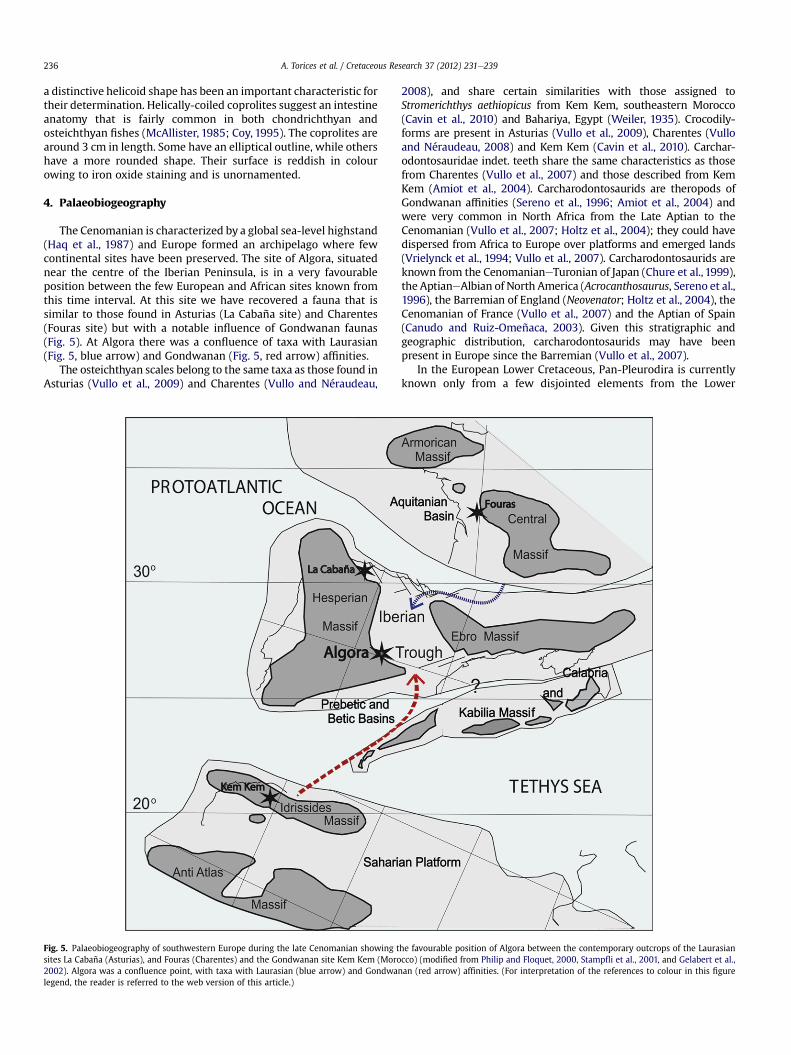
The Cenomanian is characterized by a global sea-level highstand(Haq et al., 1987) and Europe formed an archipelago where fewcontinental sites have been preserved. The site of Algora, situatednear the centre of the Iberian Peninsula, is in a very favourableposition between the few European and African sites known fromthis time interval. At this site we have recovered a fauna that issimilar to those found in Asturias (La Cabaña site) and Charentes(Fouras site) but with a notable influence of Gondwanan faunas(Fig. 5). At Algora there was a confluence of taxa with Laurasian(Fig. 5, blue arrow) and Gondwanan (Fig. 5, red arrow) affinities.
The osteichthyan scales belong to the same taxa as those found inAsturias (Vullo et al., 2009) and Charentes (Vullo and Néraudeau,
Fig. 5. Palaeobiogeography of southwestern Europe during the late Cenomanian showing tsites La Cabaña (Asturias), and Fouras (Charentes) and the Gondwanan site Kem Kem (Moro2002). Algora was a confluence point, with taxa with Laurasian (blue arrow) and Gondwalegend, the reader is referred to the web version of this article.)
2008), and share certain similarities with those assigned toStromerichthys aethiopicus from Kem Kem, southeastern Morocco(Cavin et al., 2010) and Bahariya, Egypt (Weiler, 1935). Crocodily-forms are present in Asturias (Vullo et al., 2009), Charentes (Vulloand Néraudeau, 2008) and Kem Kem (Cavin et al., 2010). Carchar-odontosauridae indet. teeth share the same characteristics as thosefrom Charentes (Vullo et al., 2007) and those described from KemKem (Amiot et al., 2004). Carcharodontosaurids are theropods ofGondwanan affinities (Sereno et al., 1996; Amiot et al., 2004) andwere very common in North Africa from the Late Aptian to theCenomanian (Vullo et al., 2007; Holtz et al., 2004); they could havedispersed from Africa to Europe over platforms and emerged lands(Vrielynck et al., 1994; Vullo et al., 2007). Carcharodontosaurids areknown from the CenomanianeTuronian of Japan (Chure et al., 1999),the AptianeAlbian of North America (Acrocanthosaurus, Sereno et al.,1996), the Barremian of England (Neovenator; Holtz et al., 2004), theCenomanian of France (Vullo et al., 2007) and the Aptian of Spain(Canudo and Ruiz-Omeñaca, 2003). Given this stratigraphic andgeographic distribution, carcharodontosaurids may have beenpresent in Europe since the Barremian (Vullo et al., 2007).
In the European Lower Cretaceous, Pan-Pleurodira is currentlyknown only from a few disjointed elements from the Lower
he favourable position of Algora between the contemporary outcrops of the Laurasiancco) (modified from Philip and Floquet, 2000, Stampfli et al., 2001, and Gelabert et al.,nan (red arrow) affinities. (For interpretation of the references to colour in this figure

A. Torices et al. / Cretaceous Research 37 (2012) 231e239 237
Barremian of Vallipón (province of Teruel, Spain). These have beenassigned to an undetermined representative of Dortokidae, beingthe oldest known record of this group of turtles endemic to Europe(Murelaga Bereikua, 1998). The presence of this family has beenrecognized in the Cenomanian of Charentes (Vullo et al., 2010). It isknown, intermittently, up to the upper Paleocene (de Lapparent deBroin et al., 2004; Pérez-García et al., 2012). In the Late Cretaceousthis taxon coexisted with another group of Pan-Pleurodira,Bothremydidae. These representatives of Eupleurodira are presentin European fossil sites from the Santonian, with a very abundantand diverse record (de Lapparent de Broin, 2001; Gaffney et al.,2006; Rabi et al., in press).
The specimens assigned to Pan-Pleurodira in Algora are,together with the representatives of Dortokidae from the Barre-mian of Vallipón, and the Cenomanian of Charentes, the onlyspecimens of this clade known from European Cretaceous fossilsites prior to the Santonian. The Pan-Pleurodira probably originatedin the European region of Pangea in the Triassic (de Lapparent deBroin, 2001). Subsequently, European endemic forms evolved, asis the case of Dortokidae and other groups in Gondwana, such asEupleurodira (de Lapparent de Broin, 2001). These two lineages hadbeen separate at least from the Late Jurassic (de Lapparent de Broinet al., 2004). The known record has led to the interpretation thattwo clades of Eupleurodira evolved in Gondwana, and occurred indifferent areas during the Early Cretaceous. The chelid turtlesdeveloped in the south of Gondwana, and the pelomedusoides inthe north (de la Fuente, 2003). In fact, representatives of fourgroups of Pelomedusoides have been recognized in the Cen-omanian of Kem Kem, Morocco (Tong and Buffetaut, 1996; Gaffneyet al., 2002, 2006; Cavin et al., 2010): Dirqadim schaefferi (Eurax-emydidae); Galianemys emringeri and G. whitei (Bothremydidae);Hamadachelys escuilliei (Podocnemididae); and Araripemys sp.(Araripemydidae). All of these taxa have been diagnosed only bycranial material. Only some elements of the shell have beenassigned, tentatively, to Galianemys (Gaffney et al., 2006; Karl,2010). It is believed that some representatives of Eupleurodiracame to Europe during the Late Cretaceous, and then several timesduring the Cenozoic (de Lapparent de Broin, 2001). The likelypresence of representatives of Eupleurodira in the Cenomanian ofAlgora, a group not otherwise represented in Europe before theSantonian (Rabi et al., in press) could indicate that it arrived inEurope earlier than previously thought.
At the Algora site another group of turtles has also been found,the Solemydidae, which are Laurasian turtles unknown in Asia(Danilov, 2005). This group is now identified in the Cenomanian ofcentral Spain, having been previously recognized in levels ofsimilar age in southwestern France (Vullo et al., 2005, 2010; Vullo,2007; Vullo and Néraudeau, 2008). Although several taxa areprobably represented in these sites, the known material in bothregions is too fragmentary to determine whether it belongs to oneor more taxa.
Fragmentary material of ?Carettochelyidae has been identified inthe Cenomanian of Charentes and La Cabaña Formation (Asturias),and remains of another indeterminate group of turtles have alsobeen recognized. These latter turtles have an ornamentation that ischelydroid in aspect, formed of small, irregular, and more or lessdichotomous grooves (Vullo and Néraudeau, 2008; Vullo et al., 2009,2010). To date, these two groups are not known in Algora.
It seems clear from the finding at the Algora site and all of theprevious taxawith Gondwanan affinities in Albian and Cenomaniansites that the faunas of Europe from that period of time wereinfluenced by Gondwanan dispersals though a palaeogeographicalbridge between Europe and Africa (Rage, 1997; Canudo and Ruiz-Omeñaca, 2003; Vullo et al., 2005; Vullo et al., 2007; Weishampelet al., 2010; Zarcone et al., 2010).
5. Conclusions
A newmiddleeupper Cenomanian fossil site has been located inAlgora, central Spain. The fossil record for this interval in Europe isscarce and knowledge of it is limited, so this find is particularlynoteworthy. The faunal diversity at the site is high, with fishes andreptiles being the groups best represented. Fishes are representedby two types of ganoid scales. Onemorphotype of ganoid scales canbe attributed to the problematic taxon Stromerichthys, and theother could belong to semionotids or lepisosteids. Turtles are rep-resented by plates with granular decorations that are assigned toSolemydidae. However, most specimens are attributed to a group ofPan-Pleurodira that is not Dortokidae, the group previouslyrecognized in Cretaceous European outcrops before the Senonian.Crocodyliforms are represented by osteoderms, bones and teethattributed to Neosuchia. Theropod remains include teeth that haverough enamel and arcuate wrinkles, and have been assigned toCarcharodontosauridae indet.
The location of this site in the Iberian Peninsula is very significantbecause it preserves a fauna with both Laurasian and Gondwananaffinities. Laurasian forms are represented by solemydid turtles.Further searching of the site may lead to the recovery of moreLaurasian forms like those that occur in southwestern France andAsturias. The rest of the Algoran fauna have Gondwanan affinities.Gondwanan influences in Laurasia during the Cenomanian weresignificant, with groups such as Carcharodontosauridae beingpresent in both North African and European sites. The appearance offossils which could be assigned to Eupleurodira is remarkablebecause this group was not previously known in Europe before theSantonian. The Algoran fossils also confirm the establishment ofpalaeogeographical connections between Africa and Europe duringthe Cenomanian (Vullo et al., 2007; Zarcone et al., 2010) that fav-oured the dispersal of Gondwanan fauna to Laurasia.
Acknowledgements
Part of this research has been financed by the projects PEII11-0237-7926 of the Junta de Comunidades de Castilla-La Mancha,and CGL2008-03112, CGL2009-09000 and CGL2009-12008 of theMinisterio de Ciencia e Innovación (Spain). We thank AlisonMurray, Mark Wilson, Barbara Grandstaff, David Batten, FranciscoOrtega and two anonymous reviewers for comments and sugges-tions and Victoria Arbour for her help with our English. AngelicaTorices is funded by FECYT and the program “Ayudas para lamovilidad postdoctoral en centros extrajeros”.
References
Aguilar, M.J., Ramírez del Pozo, J., Riba, O., 1971. Algunas precisiones sobre la sed-imentación y paleoecología del Cretácico inferior en la Zona de Utrillas-Villarroya de los Pinares (Teruel). Estudios Geológicos 27, 497e512.
Amiot, R., Buffetaut, E., Tong, H., Boudad, L., Kabiri, L., 2004. Isolated theropod teethfrom the Cenomanian of Morocco and their palaeobiogeographical significance.Revue de Paléobiologie Volume Spécial 9, 143e149.
Averianov, A.O., Kurochkin, E.N., Pervushov, E.M., Ivanov, A.V., 2005. Two bonefragments of ornithocheiroid pterosaurs from the Cenomanian of VolgogradRegion, southern Russia. Acta Palaeontologica Polonica 50, 289e294.
Barroso-Barcenilla, F., Cambra-Moo,O., Escaso, F., Ortega, F., Pascual, A., Pérez-García, A.,Rodríguez-Lázaro, J., Sanz, J.L., Segura, M., Torices, A., 2009. New and exceptionaldiscovery in the Upper Cretaceous of the Iberian Peninsula: the palaeontologicalsite of “Lo Hueco”, Cuenca, Spain. Cretaceous Research 30, 1268e1278.
Behrensmeyer, A.K., 2007. Bonebeds through time. In: Rogers, R.R., Eberth, D.A.,Fiorillo, A.R. (Eds.), Bonebeds; Genesis, Analysis and Paleobiological Signifi-cance. The University of Chicago Press, Chicago, IL, pp. 65e102.
Brusatte, S.L., Benson, R.B.J., Carr, T.D., Williamson, T.E., Sereno, P.C., 2007.The systematic utility of theropod enamel wrinkles. Journal of VertebratePaleontology 27, 1052e1056.
Buffetaut, E., 1989. Une vertèbre de Dinosaure Titanosauridé dans le Cénomaniendu Mans et ses implications paléobiogéographiques. Comptes Rendus del’Académie des Sciences, Paris, Série IIa 309, 437e443.

A. Torices et al. / Cretaceous Research 37 (2012) 231e239238
Buffetaut, E., Costa, G., Le Lœuff, J., Martin, M., Cavin, L., Rage, J.-C., Valentin, X.,Tong, H., 1996. An early Campanian vertebrate fauna from the Villeveyrac Basin(Hérault, southern France). Neues Jahrbuch für Geologie und Paläontologie,Monatschefte 1996, 1e16.
Buscalioni, A.D., Ortega, F., Weishampel, D.B., Jianu, C.M., 2001. A revision of thecrocodyliform Allodaposuchus precedens from the Upper Cretaceous of theHateg Basin, Romania. Its relevance in the phylogeny of Eusuchia. Journal ofVertebrate Paleontology 21, 74e86.
Canudo, J.I., Ruiz-Omeñaca, J.I., 2003. Los restos directos de dinosaurios terópodos(excluyendo Aves) en España. In: Pérez-Lorente, F. (Ed.), Dinosaurios y OtrosReptiles Mesozoicos en España. Fundación Patrimonio Paleontológico de LaRioja, Instituto de Estudios Riojanos, Universidad de La Rioja, Logroño, Colec-ción Ciencias de la Tierra 26, pp. 269e312.
Cavin, L., Martin, M., Valentin, X., 1996. Découverte d’Atractosteus africanus (Actino-pterygii, Lepisosteidae) dans le Campanien inférieur de Ventabren (Boûches-du-Rhône, France). Implications paléobiogéographiques. Revue de Paléobiologie 15,1e7.
Cavin, L., Tong, H., Boudad, L., Meister, C., Piuz, A., Tabouelle, J., Aarab, M., Amiot, R.,Buffetaut, E., Dyke, G., Hua, S., Le Loeuff, J., 2010. Vertebrate assemblages fromthe early Late Cretaceous of southeastern Morocco: an overview. Journal ofAfrican Earth Sciences 57, 391e412.
Chure, D.J., Manabe, M., Tanimoto, M., Tomidam, Y., 1999. An unusual theropodtooth from the Mifune Group (Late Cenomanian to Early Turonian), Kumamoto,Japan. In: Tomida, Y., Rich, T.H., Vickers-Rich, P. (Eds.), Proceedings of theSecond Gondwanan Dinosaur Symposium. National Science Museum Mono-graphs 15, pp. 291e296.
Coy, C.E., 1995. The first record of spiral coprolites from the Dinosaur Park Forma-tion (Judith River Group, Upper Cretaceous), southern Alberta, Canada. Journalof Paleontology 69, 1191e1194.
Danilov, I.G., 2005. Die fossilen Schildkröten Europas. In: Fritz, U. (Ed.), Handbuchder Reptilien und Amphibien Europas. Band 3/IIIB: Schildkroten II. Wiebelsheim(Aula), pp. 329e441.
Dutheil, D.B., 1999. Freshwater fish fauna from the Upper Cretaceous of Morocco. In:Arratia, G., Schultze, H.P. (Eds.), Mesozoic Fishes 2. Systematics and FossilRecord. Verlag Dr. F. Pfeil, München, pp. 553e564.
Fejfar, O., Ko�s�ták, M., Kva�cek, J., Mazuch, M., Mouèka, M., 2005. First Cenomaniandinosaur from Central Europe (Czech Republic). Acta Palaeontologica Polonica50, 295e300.
Fernández-López, S.R., 2000. Temas de Tafonomía. Departamento de Paleontología,Universidad Complutense de Madrid, Madrid, 167 pp.
Floquet, M., Alonso, A., Meléndez, A., 1982. El Cretácico Superior de Cameros-Castilla. In: García, A. (Ed.), El Cretácico de España. Universidad Complutense deMadrid, Madrid, pp. 387e456.
de la Fuente, M., 2003. Two new pleurodiran turtles from the Portezuelo Formation(Upper Cretaceous) of Northern Patagonia, Argentina. Journal of Paleontology77, 559e575.
Gaffney, E.S., Tong, H., Meylan, P.A., 2002. Galianemys, a new side-necked turtle(Pelomedusoides: Bothremydidae) from the Late Cretaceous of Morocco.American Museum Novitates 3379, 1e20.
Gaffney, E.S., Tong, H., Meylan, P.A., 2006. Evolution of the side-necked turtles: thefamilies Bothremydidae, Euraxemydidae, and Araripemydidae. Bulletin of theAmerican Museum of Natural History 300, 1e698.
García, A., Segura, M., García-Hidalgo, J.F., Carenas, B., 1993. Mixed siliciclastic andcarbonate platform of AlbianeCenomanian age from the Iberian Basin (Spain).In: Simo, T., Scott, B.W., Masse, J.P. (Eds.), Cretaceous Carbonate Platforms.American Association of Petroleum Geologists, Memoir 56, pp. 255e269.
Gelabert, B., Sàbat, F., Rodríguez-Perea, A., 2002. A new proposal for the late Cenozoicgeodynamic evolution of the western Mediterranean. Terranova 14, 93e100.
Gil, J., Carenas, B., Segura,M., García-Hidalgo, J.F., García, A., 2004. Revisióny correlaciónde las unidades litoestratigráficas del Cretácico Superior en la región central yoriental de España. Revista de la Sociedad Geológica de España 17, 249e266.
Gottfried, M.D., Krause, D., 1998. First record of gars (Lepisosteidae, Actinopterygii)on Madagascar: Late Cretaceous remains from the Mahajanga Basin. Journal ofVertebrate Paleontology 18, 275e279.
Haq, B.U., Hardenbol, J., Vail, P.R., 1987. Chronology of fluctuating sea levels since theTriassic. Science 235, 1156e1167.
Holtz, T.R., Molnar, R., Currie, P.J., 2004. Basal Tetanure. In: Weishampel, D.B.,Dodson, P., Osmólska, H. (Eds.), The Dinosauria, second ed. University ofCalifornia Press, Berkeley, CA, pp. 71e111.
Jonet, S., 1981. Contribution à l’étude des vertébrés du Crétacé portugais et spé-cialement du Cénomanien de l’Estrémadure. Comunicações dos Serviços Geo-lógicos de Portugal 67, 191e300.
Joyce, W.G., Chapman, S.D., Moody, R.T.J., Walker, C.A., 2011. The skull of the sol-emydid turtleHelochelydra nopcsai from the Early Cretaceous of the Isle ofWight(UK) and a review of Solemydidae. Special Papers in Palaeontology 86, 75e97.
Karl, H.V., 2010. Turtle shell remains (Testudines: Bothremydidae) from the Ceno-manian of Morocco. Studia Geologica Salmanticensis 46, 47e54.
de Lapparent de Broin, F., 2001. The European turtle fauna from the Triassic to thePresent. Dumerilia 4, 155e216.
de Lapparent de Broin, F., Molnar, R.E., 2001. Eocene chelid turtles from RedbankPlains, southeast Queensland, Australia. Geodiversitas 23, 41e79.
de Lapparent de Broin, F.,Murelaga, X.,1999. Turtles from theUpper Cretaceous of Laño(Iberian Peninsula). Estudios delMuseodeCienciasNaturales deÁlava 14,135e211.
de Lapparent de Broin, F., Murelaga, X., Codrea, V., 2004. Presence of Dortokidae(Chelonii, Pleurodira) in the earliest Tertiary of the Jibou Formation, Romania:paleobiogeographical implications. Acta Palaeontologica Romaniae 4, 203e215.
López-Martínez, N., Canudo, J.I., Ardévol, L., Pereda Suberbiola, X., Orue-Etxebarría, X., Cuenca-Bescós, G., Ruiz-Omeñaca, J.I., Murelaga, X., Feist, M.,2001. New dinosaur sites correlated with Upper Maastrichtian pelagic depositsin the Spanish Pyrenees: implications for the dinosaur extinction pattern inEurope. Cretaceous Research 22, 41e61.
Lyman, R.L., 1994. Vertebrate Taphonomy. In: Cambridge Manuals on Archaeology.Cambridge University Press, Cambridge, 524 pp.
Martin, J.E., Delfino, M., 2010. Recent advances in the comprehension of thebiogeography of Cretaceous European eusuchians. Palaeogeography, Palaeo-climatology, Palaeoecology 293, 406e418.
Martin, J.E., Rabi, M., Csiki, Z., 2010. Survival of Theriosuchus (Mesoeucrocodylia:Atoposauridae) in a Late Cretaceous archipelago: a new species from theMaastrichtian of Romania. Die Naturwissenschaften 97, 845e854.
McAllister, J.A., 1985. Reevaluation of the formation of spiral coprolites. Universityof Kansas Paleontological Contributions, Paper 114, 1e12.
Murelaga Bereikua, X., 1998. Primeros restos de tortugas del Cretácico Inferior (Barre-miense Superior) de Vallipón (Castellote, Teruel). Mas de las Matas 17, 189e200.
Ortega, F., Gasparini, Z., Buscalioni, A.D., Calvo, J.O., 2000. A new species ofAraripesuchus (Crocodylomorpha, Mesoeucrocodylia) from the Lower Creta-ceous of Patagonia (Argentina). Journal of Vertebrate Paleontology 20, 57e76.
Pérez-García, A, Scheyer, T.M., Murelaga, X., 2012. New interpretations of Dortokavasconica Lapparent de Broin and Murelaga, a freshwater turtle with anunusual carapace. Cretaceous Research 36, 151e161.
Philip, J., Floquet, M., 2000. Late Cenomanian; Map 14. In: Dercourt, J., Gaetani, M.,Vrielynck, B., Barrier, E., Biju-Duval, B., Brunet, M.F., Cadet, J.P., Crasquin, S.,Sandulescu, M. (Eds.), Peri-Tethys Atlas, Paleogeographical Maps, ExplanatoryNotes. CVGM/CGMV, Paris, pp. 129e136.
Rage, J.C., 1997. Terrestrial trans-Tethyan dispersals: an overview. In: First EuropeanWorkshop on Vertebrate Palaeontology, Copenhagen. Geological Society ofDenmark. Online Series 1. www.purl.dk/net/9710-0100.
Rabi, M., Tong, H., Botfalvai, G. A new species of the side-necked turtle Foxemys(Pelomedusoides: Bothremydidae) from the Late Cretaceous of Hungaryand the historical biogeography of the Bothremydini. Geological Magazine,doi:10.1017/S0016756811000756, in press.
Ruiz-Omeñaca, J.I., Vullo, R., Bernárdez, E., Buscalioni, A., 2009. El primer restodirecto de terópodo del Cenomaniense de la Peninsula Ibérica: un diente deLimanes (Oviedo, Asturias). Geogaceta 47, 29e32.
Segura, M., Barroso-Barcenilla, F., Cambra-Moo, O., Pérez-García, A., Torices, A.,2010. Introduction to the new Cenomanian palaeontological site “Algora”(Upper Cretaceous, Guadalajara, Spain). Carnets de Géologie 7, 232e233.
Segura, M., Carenas, B., García, A., 1985. Albiense y Cenomaniense de la regiónAtienza-Sacecorbo (Cordillera Ibérica, provincia de Guadalajara). Revista deMateriales y Procesos Geológicos 3, 211e226.
Segura, M., Wiedmann, J., 1982. La transgresión del Cretácico Superior en el sectorde Atienza-Sigüenza (Guadalajara, Cordillera Ibérica) y el significado de la faunaAmmonitífera. Cuadernos Geología Ibérica 8, 293e307.
Sereno, P.C., Dutheil, D.B., Iarochene, M., Larsson, H.C.E., Lyon, G.H., Magwene, P.M.,Sidor, C.A., Varricchio, D.J., Wilson, J.A., 1996. Predatory dinosaurs from theSahara and Late Cretaceous faunal differentiation. Science 272, 986e991.
Stampfli, G., Borel, G., Cavazza, W., Mosar, J., Ziegler, P.A., 2001. The PaleotectonicAtlas of the Peritethyan Domain. In: European Geophysical Society, CD ROM,ISBN 3-9804862-6-5.
Tong, H., Buffetaut, E., 1996. A new genus and species of pleurodiran turtle from theCretaceous of southern Morocco. Neues Jahrbuch für Geologie und Paläonto-logie Abhandlungen 199, 133e150.
Torices, A., 2007. Los dinosaurios terópodos del Cretácico superior de la Cuenca Sur-Pirenaica. Unpublished PhD Thesis, Universidad Complutense de Madrid,Madrid, 409 pp.
Torices, A., Barroso-Barcenilla, F., Cambra-Moo, O., Pérez-García, A., Segura, M.,2010. The new Cenomanian vertebrate site “Algora” (Guadalajara, Spain).Journal of Vertebrate Paleontology 28 (Supplement to No. 3), 176.
Vilas, L., Mas, R., García, A., Alonso, A., Meléndez, N., Rincón, R., 1982. Ibérica Suro-ccidental. In: García, A. (Ed.), El Cretácico de España. Universidad Complutensede Madrid, Madrid, pp. 457e508.
Vrielynck, B., Dercourt, J., Cottereau, N.,1994. Des seuils lithosphériques dans la Téthys.Comptes Rendus de l’Académie des Sciences, Paris, Série IIa 318, 1677e1685.
Vullo, R., 2007. Les vertébrés du Crétacé supérieur des Charentes (Sud-Ouest de laFrance): biodiversité, taphonomie, paléoécologie et paléobiogéographie.Mémoires Géosciences Rennes 125, 1e302.
Vullo, R., Néraudeau, D., 2008. Cenomanian vertebrate assemblages from south-western France: a new insight into the European mid-Cretaceous continentalfauna. Cretaceous Research 29, 935e940.
Vullo, R., Néraudeau,D., Allain, R., Cappetta, H., 2005.Unnouveaugisement àmicrorestesde vertébrés continentaux et littoraux dans le Cénomanien inférieur de Fouras(Charente-Maritime, Sud-Ouest de la France). Comptes Rendus Palevol 4, 95e107.
Vullo, R., Néraudeau, D., Lenglet, T., 2007. Dinosaur teeth from the Cenomanian ofCharentes, western France: evidence for a mixed Laurasian-Gondwananassemblage. Journal of Vertebrate Paleontology 27, 931e943.
Vullo, R., Bernárdez, E., Buscalioni, A.D., 2009. Vertebrates from the middle?elateCenomanian La Cabaña Formation (Asturias, northern Spain): palaeo-environmental and palaeobiogeographic implications. Palaeogeography,Palaeoclimatology, Palaeoecology 276, 120e129.
Vullo, R., de Lapparent de Broin, F., Néraudeau, D., Durrieu, N., 2010. Turtles fromthe Early Cenomanian paralic deposits (Late Cretaceous) of Charentes, France.Oryctos 9, 37e48.

A. Torices et al. / Cretaceous Research 37 (2012) 231e239 239
Weiler, W., 1935. Ergebnisse der Forchungsreisen Prof. E. Stromers in den WüstenÄgyptens, II. Wirbeltierreste der Baharîje-Stufe (unterstes Cenoman). 16. NeueUntersuchungenanden Fischresten. AbhandlungenderBayerischenAkademiederWissenschaften, Mathematische-Naturwissenschaftliche Abteilung 32, 1e57.
Weishampel, D.B., Csiki, Z., Benton, M.J., Grigorescu, D., Codrea, V., 2010. Palaeo-biogeographic relationships of the Hateg biota e between isolation and inno-vation. Palaeogeography, Palaeoclimatology, Palaeoecology 293, 419e437.
Zarcone, G., Petti, F.M., Cillari, A., Di Stefano, P., Guzzetta, D., Nicosia, U., 2010.A possible bridge between Adria and Africa: new palaeobiogeographic andstratigraphic constraints on the Mesozoic palaeogeography of the CentralMediterranean area. Earth-Science Reviews 103, 154e162.
Appendix
Fossil groups from the palaeontological site of AlgoraPlantaeGymnospermsCycadaceans
MolluscaBivalvia indet.
OsteichthyesStromerichthys sp.Undetermined remains (Semionotids/Lepisosteids/Stromerichthys sp.)
CheloniaPan-Pleurodiracf. Eupleurodira indet.
Stem TestudinesSolemydidae indet.
CrocodylomorphaNeosuchia indet.
DinosauriaSaurischiaTheropoda
Carcharodontosauridae indet.Trace fossilsCoprolites