to cross the sea or to follow the coast? flight directions and behaviour of migrating raptors...
TRANSCRIPT

TO CROSS THE SEA OR TO FOLLOW THE COASTFLIGHT DIRECTIONS AND BEHAVIOUR
OF MIGRATING RAPTORS APPROACHINGTHE MEDITERRANEAN SEA IN AUTUMN
by
SUSANNA K MEYER1) RETO SPAAR2) and BRUNO BRUDERER34)
(Swiss Ornithological Institute 6204 Sempach Switzerland)
(Acc 2-XII-1999)
Summary
We studied the ight behaviour of migrating raptors confronted with the Mediterraneansea at an average coast site near Malaga (Spain) in autumn by means of a trackingradar Behavioural reactions to the water barrier were species-specic but modi ed byenvironmental conditions Honey buzzards Pernis apivorus and black kites Milvus migranswere reluctant to crossing the water and followed the east-west leading coast the fewhoney buzzards crossing the sea at the end of the migratory period were probably juvenilesConsiderablenumbers of harriers Circus sp falcons Falco sp and ospreys Pandion haliaetuscrossed the Mediterranean sea Falcons crossing the sea had higher ight altitudes thanthose following the coast or crossing the bay Sea crossings occurred preferably in followingwinds and also in sidewinds whereas no birds were observed to cross the sea in strongopposing winds However tailwind-support only partly explained for different migratoryroutes Raptors crossing the sea in apping-gliding ight increased airspeeds with sidewindsto reduce drift but different from theory they did not decrease airspeed with increasingtailwind-support indicating that they minimised ight time above sea Time and energyrelated consequences of different ight routes are discussed
1) E-mail address MeyerSorninstch2) Corresponding author e-mail address SpaarRorninstch3) E-mail address BrudererBorninstch4) This study was part of the project lsquoTemporal and spatial pattern of bird migration inthe western Mediterranean mdash ight strategies in an ecological bottleneck arearsquo of theSwiss Ornithological Institute We thank all colleagues at the radar stations for their eldassistance especially F Liechti and H Stark T Steuri provided the radar software forthe recording and analysis of the radar tracks We thank L Jenni F Liechti B Naef-Denzer G Pasinelli L Schifferli and H Schmid for their comments on the manuscriptWe gratefully acknowledge the support by the Swiss National Science Foundation (ProjectNr 31-4324295) and the Silva-Casa Foundation
creg Koninklijke Brill NV Leiden 2000 Behaviour 137 379-399
380 MEYER SPAAR amp BRUDERER
Introduction
Due to food shortage in winter more than half of the Palearctic raptor speciesmigrate to southern Europe or even to Africa (Geacutensboslashl 1997) Large speciesmigrate predominantly by soaring-gliding as they need thermal updraftsthey y mainly across land Most raptors seem reluctant to cross large waterbodies (Richardson 1978) However smaller species such as sparrowhawksAccipiter sp falcons Falco sp and some medium sized raptors such as har-riers Circus sp honey buzzards Pernis apivorus black kite Milvus migransand osprey Pandion haliaetus regularly cross the Mediterranean in consid-erable numbers using apping ight records exist from most Mediterraneanislands (Bannermann amp Bannermann 1971 Beaman amp Galea 1974 Thi-ollay 1977 Galea amp Massa 1985 Bijlsma 1987 Agostini et al 1994Agostini amp Logozzo 1995) Since thermals are almost absent over the watersurface these sea crossings imply long powered ights with considerableenergy expenditure (Kerlinger 1989)
Most studies dealing with water crossing behaviour were carried out atconcentration points where the distance to the visible opposite coastline isshort (Evans amp Lathbury 1972 Kerlinger 1984 1985) At our study site theopposite African coast is out of sight (distance about 150 km) and migratingraptors approaching the south coast of Spain in autumn had to decide whetherto keep southerly ight directions and cross the Mediterranean sea or tochange ight direction and follow the coast
The decision whether to cross the water body or to follow the coasthoping or knowing that there will be a narrow strait or land bridge (in ourcase the Strait of Gibraltar) is in uenced by several factors such as geneticslife experience and weather conditions On their rst migration juvenileraptors in many species cannot take pro t from the adultrsquos experiencesince adults migrate earlier (Agostini et al 1994 Gustin amp Pizarri 1998)Weather wind and thermal conditions strongly in uence the migratory ightbehaviour (Spaar 1999) If sea crossings are supported by good followingwinds and soaring-gliding is hampered by bad thermal conditions along thecoast the short cut across the sea may minimise time and energy comparedto ying large detours along the land bridge
In our study we analyse the migratory ight directions and behaviour ofdifferent raptors approaching the Mediterranean sea in autumn We revealinter-speci c differences in the tendency to cross the water body andcompare ight characteristics for different migratory routes
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 381
Methods
Study site and data recording
Raptor migrationwas studied at the south coast of the Iberian Peninsula 25 km east of Malagaand 140 km north-east of Gibraltar (Fig 1) The site was 100 m inland from the shoreline and30 m above sea level The topography is not expected to have a concentration effect exceptfor birds approaching from the east and following the coast afterwards towards GibraltarTherefore the study site is considered as an average site at the east-west leading coast Thedistance to the African coast is approximately 150 km and the opposite coast is out of sightto a migratory bird
Observations lasted from 10 August to 24 October 1996 A tracking radar of the typelsquoSuper edermausrsquo was used to record the ight paths of migrating birds This radar allowsto track a single medium-sized raptor at distances up to 8 km (for details see Bruderer et al1995) While trackinga bird its exact position (x y and z co-ordinates)was digitally recordedevery second and the ight path visualised on a computer screen The species was identi edusing a 125 times telescope mounted parallel to the radar beam Only raptors identi edat least to the genus were included in the analysis Emphasis was on long ight tracksincluding entire phases of soaring and gliding The birds were visually observed duringthe entire ight path and their ight behaviour (direction ight style) was recorded Birdsnoted as locals or not showing clearly directed tracks were excluded from the analysis Visualobservation lasted from 800 h to 2000 h occasionally longer A daily break of two hoursin the afternoon was necessary for operational reasons (data backup routine evaluations) Itwas shifted from day to day to obtain records from the whole day In total 386 tracks ofmigrating raptors were recorded including honey buzzard (N = 81) black kite (N = 40)harriers (N = 102) including marsh harrier Circus aeruginosus Montagursquos harrier Circuspygargus and unidenti ed harriers osprey (N = 17) and falcons (N = 146) includingperegrine falcon Falco peregrinus hobby Falco subbuteo lesser kestrel Falco naumannikestrel Falco tinnunculus and unidenti ed falcons Unidenti ed falcons might include a fewsparrowhawks Accipiter nisus because at large distance or under poor visibility it was verydif cult to distinguish these genera
Wind speed and direction at all ight levels were obtained from pilot balloons which werereleased and tracked every four hours up to 3000 m above ground
Migratory ight routes
Directions in gliding or apping-gliding ight were considered to be the preferred migratorydirection To compare ight directions we categorised the raptor tracks by ight directionand position above ground (Fig 1) raptors lsquo ying along the coastrsquo had ight directions of240deg -300deg and ew above land whereas birds lsquo ying across the bayrsquo (240deg -270deg ) at leastpartly ew above sea Raptors lsquocrossing the searsquo ew out to the sea with directions of 95deg -240deg Birds ying north- or eastwards (300deg -95deg ) were not included in the further analysisTo separate raptors lsquocrossing the searsquo and lsquo ying across the bayrsquo we took the angle of thecoastal tip near Fuengirola (direction 240deg distance about 50 km) which is visible undergood atmospheric conditions and therefore is a possible leading point for birds ying acrossthe bay of Malaga
382 MEYER SPAAR amp BRUDERER
Fig 1 Location of the radar and sectors associated with the following migratory routeslsquocrossing the searsquo (directions of 95deg -240deg ) lsquo ying across the bayrsquo (birds at least partiallyabove the water body but keeping an angle closer to the coast ie 240deg -270deg ) lsquo ying alongthe coastrsquo (directions of 240deg -300deg above land) lsquoreversed migrationrsquo (directions of 300deg -
95deg ) The circle indicates the range of the radar for tracking medium-sized raptors
Analysis of tracks
Similarly to Spaar (1996) for raptor tracks in Israel the tracks were subdivided into intervalsof 10 s and different ight phases marked interactively on the computer screen We de nedthe following ight styles (1) lsquosoaring-gliding ightrsquo consisted of thermal circling and inter-thermal gliding phases (vertical speed lt shy 05 ms) (2) lsquosoaring combined with apping ightrsquo included birds in thermal circling and inter-thermal apping-gliding ight (verticalspeed 0 plusmn 05 ms) and (3) lsquohorizontal apping-gliding ightrsquo (vertical speed 0 plusmn 05 ms)
Pilot balloons released and tracked every four hours informed about wind direction andwind speed at all ight levels Airspeed and heading while gliding were calculated bysubtractingthe wind vector at the ight altitude from the track vector (Liechti 1993) Air- andgroundspeed are the velocities of the bird relative to the air and ground respectively Cross-country speed relative to the ground in soaring-gliding ight was calculated as the averagespeed over a distance of at least 2 km Cross-country speed relative to the air was derived bysubtracting the wind vector from the vector of cross-country speed relative to the ground
In the diagrams the time of day is expressed as a percentage of the time between sunrise(0) and sunset (100) in order to compensate for the time shift of dawn and dusk duringthe observation period Circular statistics are based on Batschelet (1981) Tailwind- and
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 383
sidewind-componentare the wind speeds in track direction and rectangular to track directionrespectivelyTo analyse seasonal trends in the proportion of sea crossing 5-day periods withat least two ight paths of a species group were included in the analysis
Results
Migratory directions and ight styles
Track directions of all raptors were south-westerly (238deg plusmn 34deg mean plusmn SD)and coincided with the direction towards the visible tip of the coastline nearFuengirola corresponding headings were 246deg plusmn 36deg (Fig 1) Tracks andheadings differed signi cantly between the species groups (Watson-Williamsmultisample test tracks F = 1884 df = 4 N = 382 p lt 00001headings F = 2148 df = 4 N = 382 p lt 00001) Black kites(track 269deg heading 276deg ) and honey buzzards (252deg 263deg ) kept westerly ight directions mainly following the east-west leading coastline whereasharriers (229deg 231deg ) falcons (228deg 240deg ) and ospreys (225deg 221deg ) hadsouth-westerly ight directions (Fig 2) Thus regarding the water-crossingbehaviour two main groups occurred no black kites and only few honeybuzzards (17 of tracks) but large proportions of harriers (65) falcons(57) and ospreys (73) crossed the sea (Fig 3A) The proportion of birds ying across the bay was below 15 of tracks in all groups For all groupswind directions were predominantly from north-west or from east (Fig 2)
The proportion of sea crossings (5-day periods) increased towards theend of the migratory season in honey buzzards and falcons (honey buzzardR 2 = 082 N = 6 p lt 001 falcons R 2 = 068 N = 12 p lt 001) Noseasonal trend was found in marsh and Montagursquos harriers (marsh harrierR 2 = shy 025 N = 5 NS Montagursquos harrier R 2 = shy 015 N = 5 NS)
According to the different proportions of migratory directions propor-tions of migratory ight styles differed between species groups soaring-gliding was the main ight style of honey buzzards and black kites (50and 71 of total tracking time respectively Fig 3B) Flapping-gliding ight was extensively used by harriers (62) falcons (66) and ospreys(86) Proportion of apping-gliding ight was lower in marsh harriers(50) than in Montagursquos harriers (76) Raptors combining soaring with apping-gliding ight had lower climbing rates than birds in pure soaring-gliding ight indicating that apping might be a compensation for weakthermal conditions (122 ms N = 39 and 158 ms N = 77 respectivelyt = shy 221 p lt 005)
384 MEYER SPAAR amp BRUDERER
Fig 2 Distribution of track (line) and heading (shaded) directions for the different raptorgroups Corresponding distribution of wind directions at the levels of the tracked birds given
at the right
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 385
Fig 3 Proportion of different migratory routes (A) and of time spent in different ightstyles (B without reversed migrants) in the species groups For de nitions see methods Totaltracking time honey buzzard (512 min) black kite (174 min) harriers (369 min) falcons
(497 min) and osprey (107 min)
In uence of environmental conditions on migratory direction
In weak winds (ie wind speed below 4 ms) numbers of falcons harriersand ospreys crossing the sea or following the coast were similar for allwind directions (df = 3 x 2 = 28 NS and x 2 = 64 NS respectivelyTable 1) In strong winds sea crossings occurred in westerly northerlyand easterly winds (ie side and following winds) but not in southerly
386 MEYER SPAAR amp BRUDERER
TABLE 1 Comparison of wind situations and migratory routes
Crossing the sea Flying along the coast
weak wind strong wind weak wind strong wind
West 13 19 5 9North 16 40 9 15East 17 22 7 9South 9 0 1 1
x 2 28 397 64 116p 04 lt 0001 01 lt 001
Number of tracks given Wind direction West (225deg -315deg ) North (315deg -45deg ) East (45deg -135deg ) South (135deg -225deg ) Weak winds are below 4 ms Tracks of harriers falcons andospreys are included
winds (ie opposing winds df = 3 x 2 = 397 p lt 0001 Table 1)Strong northerly winds providing tailwind-support were preferred comparedto strong westerly and easterly winds (df = 2 x 2 = 96 p lt 001) In birdsfollowing the coast under strong winds numbers of observed birds showeda similar pattern as in birds crossing the sea they were higher in northerlywesterly and easterly compared to southerly winds (df = 3 x 2 = 116p lt 001 Table 1) The lower number of birds following the coast instrong southerly winds might be an artefact migratory birds were driftedtowards the mountain chains northerly of the radar station and were out ofthe tracking range of the radar
The tailwind-support achieved by the birds did only partly explain fordifferent migratory routes tailwind-component (t-c) of raptors crossingthe sea was higher than a theoretical t-c for the same birds assuming to y along the coast (270deg paired t-test N = 149 t = 78 p lt 0001Fig 4) On the other hand t-c of raptors ying along the coast was lowerthan the theoretical t-c if crossing the sea (210deg N = 138 t = shy 44p lt 0001) Thus many birds ew along the coast with a lower tailwind-support than if they would have crossed the sea There is an indicationthat good thermal conditions might stimulate birds to migrate along thecoast with a lower tailwind-support compared to sea crossing birds yingalong the coast instead of crossing the sea with a higher tailwind-supporthad higher climbing rates in thermal circling than birds ying along coastwith a higher tailwind-support compared to crossing the sea (mean plusmn SD
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 387
Fig 4 The observed tailwind-components in raptors crossing the sea (A left) and yingalong the coast (B left) versus the theoretical tailwind-componentsfor the alternativestrategyfor the same birds assumed to y along the coast (A right) and crossing the sea (B right)Paired t -test (A) N = 149 t = 78 p lt 0001 () (B) N = 138 t = shy 44 p lt 0001
18 plusmn 09 ms N = 29 and 10 plusmn 05 ms N = 17 respectively t-testt = 31 p lt 0005)
Flight altitudes
For all raptors the median of ight altitudes was below 1000 m above groundlevel (agl) and the 90 altitude limit was below 1800 m agl (Fig 5)Median altitudes differed signi cantly between the species groups (blackkite median 636 m N = 40 honey buzzard 862 m N = 81 harriers910 m N = 102 falcons 949 m N = 146 osprey 951 m N = 17 Kruskal-Wallis ANOVA H = 1835 df = 4 N = 386 p lt 0005) Harriers andfalcons had signi cantly higher ight altitudes than black kites (GT2-testp lt 001) Maximum observed ight levels were at about 1400 m agl inblack kites at 1800 m in ospreys at 2000 m in harriers at 2200 m in honeybuzzards and at 2600 m in falcons (Fig 5) No black kites were tracked in theearly afternoon when ight altitudes of soaring migrants are usually highpartly due to long operational breaks in the rst week of our observationperiod Thus maximum ight levels of black kites were probably higher
388 MEYER SPAAR amp BRUDERER
Fig 5 Flight altitudes above ground (mean per track) in the course of the day (0 =sunrise 100 = sunset) Median 636 m for black kite 862 m for honey buzzard 910 m
for harriers 949 m for falcons and 951 m for osprey
Flight altitudes of honey buzzards black kites and harriers increased fromsunrise towards noon and were more or less stable thereafter (Fig 5) Falconsachieved high ight altitudes already soon after sunrise by apping-gliding ight In falcons ight altitudes differed between migratory routes birdscrossing the sea had higher ight altitudes than birds ying across the bayand along the coast (Fig 6 Kruskal-Wallis ANOVA H = 181 df =2 p = 00001 GT2-test crossing the sea versus ying across the bay(p lt 001) and versus along the coast (p lt 005)) In honey buzzards andharriers differences in ight altitudes between migratory routes showed thesame tendency but did not differ statistically (Fig 6)
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 389
Fig 6 Flight altitudes above ground (mean per track) for different migratory routes inhoney buzzard harriers and falcons Boxes are medians with 25-75 quartiles whiskersshow 10-90 range dots are outliers (ie outside the 15 box length from the upper and
lower value) GT2-test p lt 005 p lt 001
Fig 7 Mean climbing rates in thermal circling in the course of the day (0 = sunrise100 = sunset) Climbing rate did not differ between species groups (ANOVA F = 116
df = 4 N = 116 NS)
Flight characteristics in soaring-gliding
Thermal circling phases were recorded in 116 of 386 tracks and averageclimbing rate was 146 plusmn 084 ms (N = 116) Climbing rates increasedduring the morning hours (Fig 7 time of day lt 50 Spearman rankcorrelation N = 61 r = 034 p lt 001) Strong thermal convection aroundnoon allowed climbing rates of up to 45 ms From noon to sunset climbing
390 MEYER SPAAR amp BRUDERER
TA
BL
E2
Flig
htch
arac
teri
stic
sof
diffe
rent
spec
ies
grou
ps(m
s)
hone
ybu
zzar
dbl
ack
kite
harr
iers
falc
ons
ospr
eyF
Soar
ing
glid
ing
Cli
mbi
ngra
te1
35plusmn
077
(48)
171
plusmn0
97(2
2)1
54plusmn
087
(21)
134
plusmn0
84(2
4)1
86(1
)1
16N
SC
ross
-cou
ntry
grou
ndsp
eed
86
plusmn0
4(2
8)7
6plusmn
07
(7)
93
plusmn0
9(9
)9
3plusmn
10
(6)
89
(1)
079
NS
Cro
ss-c
ount
ryai
rspe
ed10
4plusmn
42
(28)
99
plusmn3
6(7
)9
3plusmn
25
(9)
95
plusmn3
1(1
0)11
3(1
)0
27N
SIn
ter-
ther
mal
grou
ndsp
eed
116
plusmn3
2(4
7)10
8plusmn
25
(21)
121
plusmn2
5(2
1)11
2plusmn
32
(23)
114
(1)
075
NS
Inte
r-th
erm
alai
rspe
ed12
8plusmn
37
(47)
134
plusmn3
0(2
1)12
0plusmn
30
(21)
127
plusmn3
6(2
3)17
0(1
)0
57N
S
Hor
izon
tal
appi
ng-g
lidi
ngi
ght
Gro
unds
peed
138
plusmn2
5(1
6)10
9plusmn
26
(9)
116
plusmn1
9(6
1)12
2plusmn
36
(68)
146
plusmn1
9(1
0)2
47
Air
spee
d11
1plusmn
23
(16)
103
plusmn2
3(9
)11
2plusmn
18
(58)
113
plusmn2
4(6
8)13
2plusmn
30
(10)
387
Mea
nplusmn
SD
(N)
Dif
fere
nces
wer
ete
sted
byA
NO
VA
S
igni
ca
nce
leve
l
plt
005
plt
001
H
oriz
onta
l
appi
ng-g
lidi
ng
ight
was
incl
uded
only
ifve
rtic
alsp
eeds
did
note
xcee
d0
plusmn0
5m
san
dif
noth
erm
alci
rcli
ngw
asob
serv
ed
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 391
rates did not decrease signi cantly (Spearman rank correlation N = 55r = shy 015 NS) Climbing rates in thermal circling below 1 ms occurredduring the whole day They did not differ between species groups (Table 2ANOVA F = 116 df = 3 N = 115 NS) Flight altitudes increased withincreasing climbing rates in birds lsquo ying along the coastrsquo (r = 049 N = 64p lt 00001) Average circling duration in completed soaring phases was106 plusmn 67 s (N = 80) and it was not related to climbing rate (r = 002N = 80 NS)
Mean airspeed and groundspeed in inter-thermal gliding did not differbetween the species groups (Table 2 ANOVA F = 057 and F = 075respectively df = 3 N = 112 NS) and means per species ranged between120-134 ms and 108-121 ms respectively Raptors increased airspeedsin inter-thermal gliding with increasing climbing rates (r = 052 N =113 p lt 00001) indicating that they maximised cross-country speeds insoaring-gliding ight (Pennycuick 1989 Spaar amp Bruderer 1997a) Meancross-country speeds in soaring-gliding did not differ between species groups(ANOVA cross-country airspeed F = 079 df = 3 N = 54 NS range 93-113 ms cross-country groundspeed F = 027 df = 3 N = 54 NS range76-93 ms)
Flight characteristics in horizontal apping-gliding
This ight style was considered if vertical speeds did not exceed 0 plusmn 05 msand no thermal circling was observed within a track Air- and groundspeedsdiffered between species groups (ANOVA airspeed F = 247 df = 4N = 161 p lt 005 groundspeed F = 387 df = 4 N = 164p lt 0005 Table 2) Ospreys had higher groundspeeds than harriers (GT2-test p lt 005) and higher ground- and airspeeds than black kites (p lt 005)Mean airspeeds per species ranged between 103-132 ms correspondinggroundspeeds between 109-146 ms
Flapping-gliding migrants reached signi cantly higher cross-country air-speeds (113 plusmn 22 ms N = 161) and groundspeeds (122 plusmn 31 ms N =164) than birds in soaring-gliding ight (airspeed 100 plusmn 36 ms N = 55Welch-test t = 249 df = 68 p lt 001 groundspeed 87 plusmn 25 msN = 55 t = 846 df = 114 p lt 00005)
In horizontal apping-gliding ight reactions to different wind con-ditions depended on migratory route raptors ying along the coast de-
392 MEYER SPAAR amp BRUDERER
Fig 8 Airspeed in horizontal apping-gliding ight versus tailwind- and sidewind-components for different migratory routes Linear regression (A) excluding one value inbrackets y = shy 0256x + 1128 R 2 = 0215 N = 56 p lt 0001 (B) r = shy 010 N = 57NS (C) r = shy 007 N = 85 NS (D) excluding two values in brackets y = 0191x+ 1072
R2 = 005 N = 84 p lt 005
creased airspeed with increasing tailwind-component (Fig 8A) but air-speed was not related to sidewind-component (Fig 8B) In contrast in rap-tors crossing the sea no relation was found between airspeed and tailwind-component (Fig 8C) but airspeed increased with increasing sidewind-component (Fig 8D)
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 393
Discussion
There are only few studies on the ight behaviour of migrating raptors cross-ing the sea mainly carried out at concentration points such as narrow wa-ter straits or peninsulas (Evans amp Lathbury 1972 Bernis 1975 Kerlinger1984) and none by tracking radar As these studies were limited to visualobservation mainly high altitudinal migration was often not detected (Ker-linger et al 1985) Our approach is different because (i) we observed raptormigration at an average coast site (ii) we used a tracking radar and (iii) wemeasured wind conditions at all ight levels
Migratory routes and in uence of environmental conditions
Although black kites and in larger numbers honey buzzards undertakelong sea-crossings in the Central Mediterranean (Beaman amp Galea 1974Agostini et al 1994) in our study no black kites and few honey buzzardswere observed attempting the sea crossing The majority of harriers falconsand ospreys crossed the sea Sea crossing honey buzzards occurred towardsthe end of the migratory season and were probably juveniles as in honeybuzzards adults migrate two weeks earlier than juveniles (Kjelleacuten 1992Agostini et al 1997) In contrast to the adults which often migrate in large ocks juveniles mainly migrate singly and have no experience about themigratory ight route This may lead to a higher proportion of juvenilescrossing the sea
Wind and thermal conditions affected migratory directions many harri-ers falcons and ospreys crossed the sea in favourable following and sidewinds but no sea crossings were observed in strong opposing winds This isreasonable because energy and time consumption for the sea crossing andtherefore the risk of exhaustion drastically increase in opposing winds How-ever tailwind-support did not fully explain the decisions on migratory routesbecause regularly birds followed the coast with a lower tailwind-supportcompared to crossing the sea These birds following the coast reached highclimbing rates in thermal circling and this might be compensating for thelower tailwind-support Under good thermal conditions birds were able tomigrate in soaring-gliding ight which consumes much less energy com-pared to apping-gliding ight when crossing the sea (see below Energyand time consumption) Anyway birds in bad shape for example those with
394 MEYER SPAAR amp BRUDERER
low fat reserves should follow the coast in order to minimise the risk ofstarvation to feed when ever it is possible and to y in soaring-gliding ight
At the same observation site migratory directions of nocturnal migrants ying below 2000 m above ground shift from 190deg to 250deg within the nightthe majority of the birds cross the sea when approaching the coast in the rsthalf of the night whereas they hesitate later on (Bruderer amp Liechti 1998)Thus the motivation to cross large barriers seems to decrease after somehours of migratory ight
Energy and time consumption
Energy consumption in apping ight disproportionately increases withbody mass while in soaring-gliding ight it is 2-3 times the basal metabolicrate (BMR) only (for herring gull Larus argentatus Baudinette amp Schmidt-Nielson 1974 Lustick et al 1978) In honey buzzards the average observedairspeed in apping-gliding ight was 111 ms and fat consumption perdistance (gkm) for an average honey buzzard (body mass in Table 3) atthis speed is about 6 times higher than in soaring-gliding ight (calculatedafter Pennycuick (1989 version 11) energy consumption in soaring-gliding ight = 2 times BMR body drag coef cient = 04) If the body dragcoef cient is set to 01 as recently proposed by Pennycuick (1997) energyconsumption in apping ight per km is reduced and energetic differencebetween soaring-gliding and apping ight become smaller but calculatedmaximum range speeds in apping ight are very high compared to ourobservations
We calculated energy and time consumption of different migratory routesbased on observed ight speeds for an average honey buzzard (Table 3) Notethat we did not take into account the time needed for refuelling consumedenergy (ie burned fat) and that we assumed zero wind Crossing the sea in apping-gliding ight (150 km distance) took on average about 4 hours andit was the time-minimising strategy for a honey buzzard (Table 3) Flyingalong the coast (250 km) required about 7 hours in soaring-gliding ightand 6 hours in apping-gliding ight Flying along the coast by soaring-gliding is the energy-minimising strategy for a honey buzzard and consumesabout 4 g fat (Table 3) Flapping-gliding ight along the coast is energeticallythe most expensive but never the fastest strategy Thus black kites andhoney buzzards migrated predominantly according to the energy minimising
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 395
TABLE 3 Observed airspeeds and corresponding estimated ight durationand fat consumption for an average honey buzzard when ying along the
coast or across the sea in different ight styles
Flight style Soaring-gliding Horizontal Horizontal apping-gliding apping-gliding
Flight route Flying along the coast Flying along the coast Crossing the seaDistance 250 km 250 km 150 km
Airspeed ms 101 (81-123) 108 (88-133) 108 (88-133)Flight duration hmin 652 (538-834) 626 (513-753) 351 (308-444)Fat consumption g 41 (34-55) 266 (230-298) 158 (138-179)for ight
Average honey buzzard = body mass 08 kg wing span 13 m wing area 025 m2 Medianand 25-75 range are given in brackets Calculations on fat consumption are based onPennycuickrsquos (1989) program of bird ight performance version 11 and birds are assumedto maximize cross-country speed in soaring-gliding ight (Spaar amp Bruderer 1997a)
strategy because a large proportion of the birds followed the coast in soaring-gliding ight
For species such as falcons or small harriers the difference in energyconsumption between apping and soaring-gliding ight is smaller Asthese species regularly use apping-gliding ight also over land or combinesoaring with apping-gliding ight differences in energy consumptionbetween land and sea route become even smaller and only short detours areenergetically favourable By combining soaring and apping-gliding ightsmaller raptors such as falcons or harriers are able to y under varyingenvironmental conditions to extend their migratory activity to times whenthermal updrafts are absent and to migrate even at night (Stark amp Liechti1993 Spaar amp Bruderer 1997b Spaar et al 1998)
Flight altitudes
Flight altitudes differed between the species groups partly due to differencesin migratory routes Birds crossing the sea ew considerably higher thanthose following the coast Kerlinger (1984) made similar observations atthe Delaware Bay birds crossing the bay ew at higher altitudes thanthose keeping on land He explained the higher ight altitudes by the bettervisibility of the other shore However the African coast is not visible fromour study site Some falcons even started crossing the sea after sunset
396 MEYER SPAAR amp BRUDERER
indicating that they continued migration into the night these birds had to y at least three hours until they reached the African coast The higher ightaltitudes of birds crossing the sea at our study site was mainly caused bytopographical features birds ying with southerly directions were forced toclimb higher up when crossing the mountains chains (1500 m height) about10-15 km northward of the coast and they afterwards maintained these ightlevels when approaching the sea
Flight altitudes of birds migrating along the coast were similar to altitudesobserved in North America and southern Israel with most raptors yingbelow 1000 m above ground and lt 5 above 1500 m (Kerlinger 19841989 Bruderer et al 1994 Spaar 1996)
Flight characteristics in soaring-gliding
Flight characteristics in soaring-gliding ight did not differ between thespecies groups (Table 2) As shown for raptor migration in Israel climb-ing rates do not depend primarily on morphological features but on environ-mental conditions (Spaar 1997) In general mean climbing rates in southernSpain were lower than in the semi-arid desert in Israel (Spaar 1997) Meanduration of the completed circling phases was shorter in Israel (88 plusmn 52 sSpaar 1995 1999) than in Spain In contrast to Israel (Spaar 1995 1997)circling time did not increase with increasing climbing rate Compared tothe semi-desert the development of thermal convection in southern Spainwas probably less stable and birds circled as long as possible in updraftsbecause they could not rely to encounter a next strong thermal within glid-ing distance This might also explain why inter-thermal gliding was oftensupported by apping it reduces sinking rate and prolongs gliding distance
In raptors migrating along the coast ight altitude and gliding airspeedswere related to thermal strength raptors circling in strong thermals hadhigher ight altitudes and higher inter-thermal gliding airspeeds than thosein weak thermals This is consistent with optimal ight theory for birds max-imising cross-country speed in soaring-gliding ight (Pennycuick 1989) asdemonstrated for steppe buzzards Buteo buteo vulpinus (Spaar amp Bruderer1997a)
Flight characteristics in horizontal apping-gliding
According to optimal ight theory maximum range speed decreases withfollowing winds and increases with head- and sidewinds (Pennycuick 1975
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 397
Liechti et al 1994) In raptors ying along the coast we observed decreas-ing airspeeds with increasing tailwind-support whereas birds crossing thesea did not reduce airspeeds Why did the birds react differently Sea cross-ings imply higher risks there is no possibility to land and weather (wind)conditions may change rapidly Higher airspeeds in following winds lead toshorter crossing times and lower the risk of unpredictable weather changesFurthermore drift by sidewinds may considerably extent crossing distancesand in order to reduce drift raptors increased airspeeds with increasingsindwind-components In contrast to sharp-shinned hawks Accipiter stria-tus avoiding crossings in strong lateral winds (Kerlinger 1985) we observedmany raptors to cross the sea in strong north-westerly (ie lateral) winds ifthey provided at least partial tailwind-support
Conclusions
Our observations at an average site at the southern coast of Spain showthat differences in water crossing behaviour are mainly species-speci cpresumably due to the costs for different ight strategies additionallyindividuals reacted to environmental factors such as wind and thermalconditions In medium sized raptors such as honey buzzard soaring-gliding ight along the coast is energy-minimising whereas crossing the sea istime-minimising Sea crossings occurred preferably with a good tailwind-support but also under strong sidewinds Raptors crossing the sea did notreduce airspeeds in following winds but increased airspeeds with increasingsidewinds They seemed to minimise the water crossing duration and theeffect of drift by sidewinds Under good thermal conditions birds regularlyfollowed the coast despite a lower tailwind-support compared to crossing thesea
References
Agostini N amp Logozzo D (1995) Autumn migration of honey buzzards in Southern Italymdash J Raptor Res 29 p 275-277
mdash mdash Logozzo D amp Colero C (1997) Migration of ocks of honey buzzards in SouthernItaly and Malta mdash J Raptor Res 31 p 84-86
mdash mdash Malara G Neri F Mollicone D amp Melotto S (1994) Flight strategies of honeybuzzards during spring migration across the Central Mediterranean mdash Avocetta 18p 73-76
398 MEYER SPAAR amp BRUDERER
Bannermann D-A amp Bannermann WM (1971) Handbook of the birds of Cyprus andmigrants of the Middle East mdash Oliver amp Boyd Edinburgh
Batschelet E (1981) Circular statistics in biology mdash Academic Press LondonBaudinette RV amp Schmidt-Nielsen K (1974) Energy costs of gliding ight in herring
gulls mdash Nature 248 p 83-84Beaman M amp Galea C (1974) The visible migration of raptors over the Maltese Islands
mdash Ibis 116 p 419-431Bernis F (1975) Migracion de Falconiformes y Ciconea spp por Gibraltar mdash IV Avance
sobre recuentos en el verano-otontildeo 1974 mdash Ardeola 21 p 581-592Bijlsma RG (1987) Bottleneck areas for migratory birds in the Mediterranean region mdash
ICBP Study report No 18 CambridgeBruderer B amp Liechti F (1998) Flight behaviour of nocturnally migrating birds in coastal
areas mdash crossing or coasting mdash J Avian Biol 29 p 499-510mdash mdash Blitzblau S amp Peter D (1994) Migration and ight behaviour of honey buzzards
Pernis apivorus in southern Israel observed by radar mdash Ardea 82 p 111-122mdash mdash Steuri T amp Baumgartner M (1995) Short-range high-precision surveillance of
nocturnal migration and tracking of single targets mdash Israel J Zool 41 p 207-220Evans PR amp Lathbury GW (1972) Raptor migration across the Strait of Gibraltar mdash Ibis
115 p 572-585Galea C amp Massa B (1985) Notes on the raptor migration across the central Mediter-
ranean mdash In Conservation studies on raptors (I Newton amp RD Chancellor eds)ICBP Technical Publication No 5 p 257-261
Gensboslashl B (1997) Greifvoumlgel mdash BLV VerlagsgesellschaftMuumlnchenGustin M amp Pizzari T (1998) Migratory pattern in the genus Circus sex and age
differential migration in Italy mdash Ornis Svecica 8 p 23-26Kerlinger P (1984) Flight behaviour of sharp-shinned hawks during migration 2 Over
water mdash Anim Beh 32 p 1029-1034mdash mdash (1985) Water-crossing behaviour of raptors during migration mdash Wilson Bull 97
p 109-113mdash mdash (1989) Flight strategies of migrating hawks mdash University of Chicago Press Chicagomdash mdash Bingman VP amp Able KP (1985) Comparative ight behaviour of migrating hawks
studied with tracking radar during autumn in central New York mdash Can J Zool 63p 755-761
Kjelleacuten N (1992) Differential timing of autumn migration between sex and age groups inraptors at Falsterbo Sweden mdash Orn Scan 23 p 420-434
Liechti F (1993) Naumlchtlicher Vogelzug im Herbst uumlber Suumlddeutschland Winddrift undKompensation mdash J Ornithol 134 p 373-404
mdash mdash Hedenstroumlm A amp Alerstam T (1994) Effects of sidewinds on optimal ight speedsof birds mdash J theor Biol 170 p 219-225
Lustick S Batterby B amp Kelty M (1978) Behavioural thermoregulation orientationtoward the sun in herring gulls mdash Sience 200 p 81-83
Pennycuick C (1975) Mechanics of ight mdash In Avian biology Vol 5 (DS Farner ampRJ King eds) Academic Press New York p 1-75
mdash mdash (1989) Bird ight performance a practical calculation manual mdash Oxford UniversityPress Oxford
mdash mdash (1997) Actual and lsquooptimumrsquo ight speeds eld data reassessedmdash J Exp Biol 200p 2355-2361
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 399
Richardson WJ (1978) Timing and amount of bird migration in relation to weather areview mdash Oikos 30 p 224-272
Spaar R (1995) Flight behaviour of steppe buzzard (Buteo buteo vulpinus) during springmigration in southern Israel a tracking radar study mdash Israel J Zool 41 p 489-500
mdash mdash (1996) Flight behaviour of migrating raptors in southern Israel mdash PhD thesisUniversity of Basel Verlag Schweizerische Vogelwarte Sempach
mdash mdash (1997) Flight strategies of migrating raptors a comparative study of interspecicvariation in ight characteristicsmdash Ibis 139 p 523-535
mdash mdash (1999) Flight behaviour of migrating raptors under varying environmental conditionsmdash In Proc 22 Int Ornithol Congr (N Adams amp R Slotow eds) University of NatalDurban p 1844-1862
mdash mdash amp Bruderer B (1997a) Optimal ight behaviour of soaring migrants A case studyof migrating steppe buzzards Buteo buteo vulpinus mdash Behav Ecol 8 p 288-297)
mdash mdash amp Bruderer B (1997b) Migration by apping or soaring ight strategies of marshMontagursquos and pallid harriers in southern Israel mdash Condor 99 p 458-469
mdash mdash Stark H amp Liechti F (1998) Migratory ight strategies of Levant sparrowhawkstime or energy minimization mdash Anim Behav 56 p 1185-1197
Stark H amp Liechti F (1993) Do Levant sparrowhawks Accipiter brevipes also migrate atnight mdash Ibis 135 p 233-236
Thiollay J-M (1977) Importance des populations de rapaces migrateurs en mediterraneeoccidentale mdash Alauda 45 p 115-121

380 MEYER SPAAR amp BRUDERER
Introduction
Due to food shortage in winter more than half of the Palearctic raptor speciesmigrate to southern Europe or even to Africa (Geacutensboslashl 1997) Large speciesmigrate predominantly by soaring-gliding as they need thermal updraftsthey y mainly across land Most raptors seem reluctant to cross large waterbodies (Richardson 1978) However smaller species such as sparrowhawksAccipiter sp falcons Falco sp and some medium sized raptors such as har-riers Circus sp honey buzzards Pernis apivorus black kite Milvus migransand osprey Pandion haliaetus regularly cross the Mediterranean in consid-erable numbers using apping ight records exist from most Mediterraneanislands (Bannermann amp Bannermann 1971 Beaman amp Galea 1974 Thi-ollay 1977 Galea amp Massa 1985 Bijlsma 1987 Agostini et al 1994Agostini amp Logozzo 1995) Since thermals are almost absent over the watersurface these sea crossings imply long powered ights with considerableenergy expenditure (Kerlinger 1989)
Most studies dealing with water crossing behaviour were carried out atconcentration points where the distance to the visible opposite coastline isshort (Evans amp Lathbury 1972 Kerlinger 1984 1985) At our study site theopposite African coast is out of sight (distance about 150 km) and migratingraptors approaching the south coast of Spain in autumn had to decide whetherto keep southerly ight directions and cross the Mediterranean sea or tochange ight direction and follow the coast
The decision whether to cross the water body or to follow the coasthoping or knowing that there will be a narrow strait or land bridge (in ourcase the Strait of Gibraltar) is in uenced by several factors such as geneticslife experience and weather conditions On their rst migration juvenileraptors in many species cannot take pro t from the adultrsquos experiencesince adults migrate earlier (Agostini et al 1994 Gustin amp Pizarri 1998)Weather wind and thermal conditions strongly in uence the migratory ightbehaviour (Spaar 1999) If sea crossings are supported by good followingwinds and soaring-gliding is hampered by bad thermal conditions along thecoast the short cut across the sea may minimise time and energy comparedto ying large detours along the land bridge
In our study we analyse the migratory ight directions and behaviour ofdifferent raptors approaching the Mediterranean sea in autumn We revealinter-speci c differences in the tendency to cross the water body andcompare ight characteristics for different migratory routes
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 381
Methods
Study site and data recording
Raptor migrationwas studied at the south coast of the Iberian Peninsula 25 km east of Malagaand 140 km north-east of Gibraltar (Fig 1) The site was 100 m inland from the shoreline and30 m above sea level The topography is not expected to have a concentration effect exceptfor birds approaching from the east and following the coast afterwards towards GibraltarTherefore the study site is considered as an average site at the east-west leading coast Thedistance to the African coast is approximately 150 km and the opposite coast is out of sightto a migratory bird
Observations lasted from 10 August to 24 October 1996 A tracking radar of the typelsquoSuper edermausrsquo was used to record the ight paths of migrating birds This radar allowsto track a single medium-sized raptor at distances up to 8 km (for details see Bruderer et al1995) While trackinga bird its exact position (x y and z co-ordinates)was digitally recordedevery second and the ight path visualised on a computer screen The species was identi edusing a 125 times telescope mounted parallel to the radar beam Only raptors identi edat least to the genus were included in the analysis Emphasis was on long ight tracksincluding entire phases of soaring and gliding The birds were visually observed duringthe entire ight path and their ight behaviour (direction ight style) was recorded Birdsnoted as locals or not showing clearly directed tracks were excluded from the analysis Visualobservation lasted from 800 h to 2000 h occasionally longer A daily break of two hoursin the afternoon was necessary for operational reasons (data backup routine evaluations) Itwas shifted from day to day to obtain records from the whole day In total 386 tracks ofmigrating raptors were recorded including honey buzzard (N = 81) black kite (N = 40)harriers (N = 102) including marsh harrier Circus aeruginosus Montagursquos harrier Circuspygargus and unidenti ed harriers osprey (N = 17) and falcons (N = 146) includingperegrine falcon Falco peregrinus hobby Falco subbuteo lesser kestrel Falco naumannikestrel Falco tinnunculus and unidenti ed falcons Unidenti ed falcons might include a fewsparrowhawks Accipiter nisus because at large distance or under poor visibility it was verydif cult to distinguish these genera
Wind speed and direction at all ight levels were obtained from pilot balloons which werereleased and tracked every four hours up to 3000 m above ground
Migratory ight routes
Directions in gliding or apping-gliding ight were considered to be the preferred migratorydirection To compare ight directions we categorised the raptor tracks by ight directionand position above ground (Fig 1) raptors lsquo ying along the coastrsquo had ight directions of240deg -300deg and ew above land whereas birds lsquo ying across the bayrsquo (240deg -270deg ) at leastpartly ew above sea Raptors lsquocrossing the searsquo ew out to the sea with directions of 95deg -240deg Birds ying north- or eastwards (300deg -95deg ) were not included in the further analysisTo separate raptors lsquocrossing the searsquo and lsquo ying across the bayrsquo we took the angle of thecoastal tip near Fuengirola (direction 240deg distance about 50 km) which is visible undergood atmospheric conditions and therefore is a possible leading point for birds ying acrossthe bay of Malaga
382 MEYER SPAAR amp BRUDERER
Fig 1 Location of the radar and sectors associated with the following migratory routeslsquocrossing the searsquo (directions of 95deg -240deg ) lsquo ying across the bayrsquo (birds at least partiallyabove the water body but keeping an angle closer to the coast ie 240deg -270deg ) lsquo ying alongthe coastrsquo (directions of 240deg -300deg above land) lsquoreversed migrationrsquo (directions of 300deg -
95deg ) The circle indicates the range of the radar for tracking medium-sized raptors
Analysis of tracks
Similarly to Spaar (1996) for raptor tracks in Israel the tracks were subdivided into intervalsof 10 s and different ight phases marked interactively on the computer screen We de nedthe following ight styles (1) lsquosoaring-gliding ightrsquo consisted of thermal circling and inter-thermal gliding phases (vertical speed lt shy 05 ms) (2) lsquosoaring combined with apping ightrsquo included birds in thermal circling and inter-thermal apping-gliding ight (verticalspeed 0 plusmn 05 ms) and (3) lsquohorizontal apping-gliding ightrsquo (vertical speed 0 plusmn 05 ms)
Pilot balloons released and tracked every four hours informed about wind direction andwind speed at all ight levels Airspeed and heading while gliding were calculated bysubtractingthe wind vector at the ight altitude from the track vector (Liechti 1993) Air- andgroundspeed are the velocities of the bird relative to the air and ground respectively Cross-country speed relative to the ground in soaring-gliding ight was calculated as the averagespeed over a distance of at least 2 km Cross-country speed relative to the air was derived bysubtracting the wind vector from the vector of cross-country speed relative to the ground
In the diagrams the time of day is expressed as a percentage of the time between sunrise(0) and sunset (100) in order to compensate for the time shift of dawn and dusk duringthe observation period Circular statistics are based on Batschelet (1981) Tailwind- and
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 383
sidewind-componentare the wind speeds in track direction and rectangular to track directionrespectivelyTo analyse seasonal trends in the proportion of sea crossing 5-day periods withat least two ight paths of a species group were included in the analysis
Results
Migratory directions and ight styles
Track directions of all raptors were south-westerly (238deg plusmn 34deg mean plusmn SD)and coincided with the direction towards the visible tip of the coastline nearFuengirola corresponding headings were 246deg plusmn 36deg (Fig 1) Tracks andheadings differed signi cantly between the species groups (Watson-Williamsmultisample test tracks F = 1884 df = 4 N = 382 p lt 00001headings F = 2148 df = 4 N = 382 p lt 00001) Black kites(track 269deg heading 276deg ) and honey buzzards (252deg 263deg ) kept westerly ight directions mainly following the east-west leading coastline whereasharriers (229deg 231deg ) falcons (228deg 240deg ) and ospreys (225deg 221deg ) hadsouth-westerly ight directions (Fig 2) Thus regarding the water-crossingbehaviour two main groups occurred no black kites and only few honeybuzzards (17 of tracks) but large proportions of harriers (65) falcons(57) and ospreys (73) crossed the sea (Fig 3A) The proportion of birds ying across the bay was below 15 of tracks in all groups For all groupswind directions were predominantly from north-west or from east (Fig 2)
The proportion of sea crossings (5-day periods) increased towards theend of the migratory season in honey buzzards and falcons (honey buzzardR 2 = 082 N = 6 p lt 001 falcons R 2 = 068 N = 12 p lt 001) Noseasonal trend was found in marsh and Montagursquos harriers (marsh harrierR 2 = shy 025 N = 5 NS Montagursquos harrier R 2 = shy 015 N = 5 NS)
According to the different proportions of migratory directions propor-tions of migratory ight styles differed between species groups soaring-gliding was the main ight style of honey buzzards and black kites (50and 71 of total tracking time respectively Fig 3B) Flapping-gliding ight was extensively used by harriers (62) falcons (66) and ospreys(86) Proportion of apping-gliding ight was lower in marsh harriers(50) than in Montagursquos harriers (76) Raptors combining soaring with apping-gliding ight had lower climbing rates than birds in pure soaring-gliding ight indicating that apping might be a compensation for weakthermal conditions (122 ms N = 39 and 158 ms N = 77 respectivelyt = shy 221 p lt 005)
384 MEYER SPAAR amp BRUDERER
Fig 2 Distribution of track (line) and heading (shaded) directions for the different raptorgroups Corresponding distribution of wind directions at the levels of the tracked birds given
at the right
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 385
Fig 3 Proportion of different migratory routes (A) and of time spent in different ightstyles (B without reversed migrants) in the species groups For de nitions see methods Totaltracking time honey buzzard (512 min) black kite (174 min) harriers (369 min) falcons
(497 min) and osprey (107 min)
In uence of environmental conditions on migratory direction
In weak winds (ie wind speed below 4 ms) numbers of falcons harriersand ospreys crossing the sea or following the coast were similar for allwind directions (df = 3 x 2 = 28 NS and x 2 = 64 NS respectivelyTable 1) In strong winds sea crossings occurred in westerly northerlyand easterly winds (ie side and following winds) but not in southerly
386 MEYER SPAAR amp BRUDERER
TABLE 1 Comparison of wind situations and migratory routes
Crossing the sea Flying along the coast
weak wind strong wind weak wind strong wind
West 13 19 5 9North 16 40 9 15East 17 22 7 9South 9 0 1 1
x 2 28 397 64 116p 04 lt 0001 01 lt 001
Number of tracks given Wind direction West (225deg -315deg ) North (315deg -45deg ) East (45deg -135deg ) South (135deg -225deg ) Weak winds are below 4 ms Tracks of harriers falcons andospreys are included
winds (ie opposing winds df = 3 x 2 = 397 p lt 0001 Table 1)Strong northerly winds providing tailwind-support were preferred comparedto strong westerly and easterly winds (df = 2 x 2 = 96 p lt 001) In birdsfollowing the coast under strong winds numbers of observed birds showeda similar pattern as in birds crossing the sea they were higher in northerlywesterly and easterly compared to southerly winds (df = 3 x 2 = 116p lt 001 Table 1) The lower number of birds following the coast instrong southerly winds might be an artefact migratory birds were driftedtowards the mountain chains northerly of the radar station and were out ofthe tracking range of the radar
The tailwind-support achieved by the birds did only partly explain fordifferent migratory routes tailwind-component (t-c) of raptors crossingthe sea was higher than a theoretical t-c for the same birds assuming to y along the coast (270deg paired t-test N = 149 t = 78 p lt 0001Fig 4) On the other hand t-c of raptors ying along the coast was lowerthan the theoretical t-c if crossing the sea (210deg N = 138 t = shy 44p lt 0001) Thus many birds ew along the coast with a lower tailwind-support than if they would have crossed the sea There is an indicationthat good thermal conditions might stimulate birds to migrate along thecoast with a lower tailwind-support compared to sea crossing birds yingalong the coast instead of crossing the sea with a higher tailwind-supporthad higher climbing rates in thermal circling than birds ying along coastwith a higher tailwind-support compared to crossing the sea (mean plusmn SD
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 387
Fig 4 The observed tailwind-components in raptors crossing the sea (A left) and yingalong the coast (B left) versus the theoretical tailwind-componentsfor the alternativestrategyfor the same birds assumed to y along the coast (A right) and crossing the sea (B right)Paired t -test (A) N = 149 t = 78 p lt 0001 () (B) N = 138 t = shy 44 p lt 0001
18 plusmn 09 ms N = 29 and 10 plusmn 05 ms N = 17 respectively t-testt = 31 p lt 0005)
Flight altitudes
For all raptors the median of ight altitudes was below 1000 m above groundlevel (agl) and the 90 altitude limit was below 1800 m agl (Fig 5)Median altitudes differed signi cantly between the species groups (blackkite median 636 m N = 40 honey buzzard 862 m N = 81 harriers910 m N = 102 falcons 949 m N = 146 osprey 951 m N = 17 Kruskal-Wallis ANOVA H = 1835 df = 4 N = 386 p lt 0005) Harriers andfalcons had signi cantly higher ight altitudes than black kites (GT2-testp lt 001) Maximum observed ight levels were at about 1400 m agl inblack kites at 1800 m in ospreys at 2000 m in harriers at 2200 m in honeybuzzards and at 2600 m in falcons (Fig 5) No black kites were tracked in theearly afternoon when ight altitudes of soaring migrants are usually highpartly due to long operational breaks in the rst week of our observationperiod Thus maximum ight levels of black kites were probably higher
388 MEYER SPAAR amp BRUDERER
Fig 5 Flight altitudes above ground (mean per track) in the course of the day (0 =sunrise 100 = sunset) Median 636 m for black kite 862 m for honey buzzard 910 m
for harriers 949 m for falcons and 951 m for osprey
Flight altitudes of honey buzzards black kites and harriers increased fromsunrise towards noon and were more or less stable thereafter (Fig 5) Falconsachieved high ight altitudes already soon after sunrise by apping-gliding ight In falcons ight altitudes differed between migratory routes birdscrossing the sea had higher ight altitudes than birds ying across the bayand along the coast (Fig 6 Kruskal-Wallis ANOVA H = 181 df =2 p = 00001 GT2-test crossing the sea versus ying across the bay(p lt 001) and versus along the coast (p lt 005)) In honey buzzards andharriers differences in ight altitudes between migratory routes showed thesame tendency but did not differ statistically (Fig 6)
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 389
Fig 6 Flight altitudes above ground (mean per track) for different migratory routes inhoney buzzard harriers and falcons Boxes are medians with 25-75 quartiles whiskersshow 10-90 range dots are outliers (ie outside the 15 box length from the upper and
lower value) GT2-test p lt 005 p lt 001
Fig 7 Mean climbing rates in thermal circling in the course of the day (0 = sunrise100 = sunset) Climbing rate did not differ between species groups (ANOVA F = 116
df = 4 N = 116 NS)
Flight characteristics in soaring-gliding
Thermal circling phases were recorded in 116 of 386 tracks and averageclimbing rate was 146 plusmn 084 ms (N = 116) Climbing rates increasedduring the morning hours (Fig 7 time of day lt 50 Spearman rankcorrelation N = 61 r = 034 p lt 001) Strong thermal convection aroundnoon allowed climbing rates of up to 45 ms From noon to sunset climbing
390 MEYER SPAAR amp BRUDERER
TA
BL
E2
Flig
htch
arac
teri
stic
sof
diffe
rent
spec
ies
grou
ps(m
s)
hone
ybu
zzar
dbl
ack
kite
harr
iers
falc
ons
ospr
eyF
Soar
ing
glid
ing
Cli
mbi
ngra
te1
35plusmn
077
(48)
171
plusmn0
97(2
2)1
54plusmn
087
(21)
134
plusmn0
84(2
4)1
86(1
)1
16N
SC
ross
-cou
ntry
grou
ndsp
eed
86
plusmn0
4(2
8)7
6plusmn
07
(7)
93
plusmn0
9(9
)9
3plusmn
10
(6)
89
(1)
079
NS
Cro
ss-c
ount
ryai
rspe
ed10
4plusmn
42
(28)
99
plusmn3
6(7
)9
3plusmn
25
(9)
95
plusmn3
1(1
0)11
3(1
)0
27N
SIn
ter-
ther
mal
grou
ndsp
eed
116
plusmn3
2(4
7)10
8plusmn
25
(21)
121
plusmn2
5(2
1)11
2plusmn
32
(23)
114
(1)
075
NS
Inte
r-th
erm
alai
rspe
ed12
8plusmn
37
(47)
134
plusmn3
0(2
1)12
0plusmn
30
(21)
127
plusmn3
6(2
3)17
0(1
)0
57N
S
Hor
izon
tal
appi
ng-g
lidi
ngi
ght
Gro
unds
peed
138
plusmn2
5(1
6)10
9plusmn
26
(9)
116
plusmn1
9(6
1)12
2plusmn
36
(68)
146
plusmn1
9(1
0)2
47
Air
spee
d11
1plusmn
23
(16)
103
plusmn2
3(9
)11
2plusmn
18
(58)
113
plusmn2
4(6
8)13
2plusmn
30
(10)
387
Mea
nplusmn
SD
(N)
Dif
fere
nces
wer
ete
sted
byA
NO
VA
S
igni
ca
nce
leve
l
plt
005
plt
001
H
oriz
onta
l
appi
ng-g
lidi
ng
ight
was
incl
uded
only
ifve
rtic
alsp
eeds
did
note
xcee
d0
plusmn0
5m
san
dif
noth
erm
alci
rcli
ngw
asob
serv
ed
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 391
rates did not decrease signi cantly (Spearman rank correlation N = 55r = shy 015 NS) Climbing rates in thermal circling below 1 ms occurredduring the whole day They did not differ between species groups (Table 2ANOVA F = 116 df = 3 N = 115 NS) Flight altitudes increased withincreasing climbing rates in birds lsquo ying along the coastrsquo (r = 049 N = 64p lt 00001) Average circling duration in completed soaring phases was106 plusmn 67 s (N = 80) and it was not related to climbing rate (r = 002N = 80 NS)
Mean airspeed and groundspeed in inter-thermal gliding did not differbetween the species groups (Table 2 ANOVA F = 057 and F = 075respectively df = 3 N = 112 NS) and means per species ranged between120-134 ms and 108-121 ms respectively Raptors increased airspeedsin inter-thermal gliding with increasing climbing rates (r = 052 N =113 p lt 00001) indicating that they maximised cross-country speeds insoaring-gliding ight (Pennycuick 1989 Spaar amp Bruderer 1997a) Meancross-country speeds in soaring-gliding did not differ between species groups(ANOVA cross-country airspeed F = 079 df = 3 N = 54 NS range 93-113 ms cross-country groundspeed F = 027 df = 3 N = 54 NS range76-93 ms)
Flight characteristics in horizontal apping-gliding
This ight style was considered if vertical speeds did not exceed 0 plusmn 05 msand no thermal circling was observed within a track Air- and groundspeedsdiffered between species groups (ANOVA airspeed F = 247 df = 4N = 161 p lt 005 groundspeed F = 387 df = 4 N = 164p lt 0005 Table 2) Ospreys had higher groundspeeds than harriers (GT2-test p lt 005) and higher ground- and airspeeds than black kites (p lt 005)Mean airspeeds per species ranged between 103-132 ms correspondinggroundspeeds between 109-146 ms
Flapping-gliding migrants reached signi cantly higher cross-country air-speeds (113 plusmn 22 ms N = 161) and groundspeeds (122 plusmn 31 ms N =164) than birds in soaring-gliding ight (airspeed 100 plusmn 36 ms N = 55Welch-test t = 249 df = 68 p lt 001 groundspeed 87 plusmn 25 msN = 55 t = 846 df = 114 p lt 00005)
In horizontal apping-gliding ight reactions to different wind con-ditions depended on migratory route raptors ying along the coast de-
392 MEYER SPAAR amp BRUDERER
Fig 8 Airspeed in horizontal apping-gliding ight versus tailwind- and sidewind-components for different migratory routes Linear regression (A) excluding one value inbrackets y = shy 0256x + 1128 R 2 = 0215 N = 56 p lt 0001 (B) r = shy 010 N = 57NS (C) r = shy 007 N = 85 NS (D) excluding two values in brackets y = 0191x+ 1072
R2 = 005 N = 84 p lt 005
creased airspeed with increasing tailwind-component (Fig 8A) but air-speed was not related to sidewind-component (Fig 8B) In contrast in rap-tors crossing the sea no relation was found between airspeed and tailwind-component (Fig 8C) but airspeed increased with increasing sidewind-component (Fig 8D)
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 393
Discussion
There are only few studies on the ight behaviour of migrating raptors cross-ing the sea mainly carried out at concentration points such as narrow wa-ter straits or peninsulas (Evans amp Lathbury 1972 Bernis 1975 Kerlinger1984) and none by tracking radar As these studies were limited to visualobservation mainly high altitudinal migration was often not detected (Ker-linger et al 1985) Our approach is different because (i) we observed raptormigration at an average coast site (ii) we used a tracking radar and (iii) wemeasured wind conditions at all ight levels
Migratory routes and in uence of environmental conditions
Although black kites and in larger numbers honey buzzards undertakelong sea-crossings in the Central Mediterranean (Beaman amp Galea 1974Agostini et al 1994) in our study no black kites and few honey buzzardswere observed attempting the sea crossing The majority of harriers falconsand ospreys crossed the sea Sea crossing honey buzzards occurred towardsthe end of the migratory season and were probably juveniles as in honeybuzzards adults migrate two weeks earlier than juveniles (Kjelleacuten 1992Agostini et al 1997) In contrast to the adults which often migrate in large ocks juveniles mainly migrate singly and have no experience about themigratory ight route This may lead to a higher proportion of juvenilescrossing the sea
Wind and thermal conditions affected migratory directions many harri-ers falcons and ospreys crossed the sea in favourable following and sidewinds but no sea crossings were observed in strong opposing winds This isreasonable because energy and time consumption for the sea crossing andtherefore the risk of exhaustion drastically increase in opposing winds How-ever tailwind-support did not fully explain the decisions on migratory routesbecause regularly birds followed the coast with a lower tailwind-supportcompared to crossing the sea These birds following the coast reached highclimbing rates in thermal circling and this might be compensating for thelower tailwind-support Under good thermal conditions birds were able tomigrate in soaring-gliding ight which consumes much less energy com-pared to apping-gliding ight when crossing the sea (see below Energyand time consumption) Anyway birds in bad shape for example those with
394 MEYER SPAAR amp BRUDERER
low fat reserves should follow the coast in order to minimise the risk ofstarvation to feed when ever it is possible and to y in soaring-gliding ight
At the same observation site migratory directions of nocturnal migrants ying below 2000 m above ground shift from 190deg to 250deg within the nightthe majority of the birds cross the sea when approaching the coast in the rsthalf of the night whereas they hesitate later on (Bruderer amp Liechti 1998)Thus the motivation to cross large barriers seems to decrease after somehours of migratory ight
Energy and time consumption
Energy consumption in apping ight disproportionately increases withbody mass while in soaring-gliding ight it is 2-3 times the basal metabolicrate (BMR) only (for herring gull Larus argentatus Baudinette amp Schmidt-Nielson 1974 Lustick et al 1978) In honey buzzards the average observedairspeed in apping-gliding ight was 111 ms and fat consumption perdistance (gkm) for an average honey buzzard (body mass in Table 3) atthis speed is about 6 times higher than in soaring-gliding ight (calculatedafter Pennycuick (1989 version 11) energy consumption in soaring-gliding ight = 2 times BMR body drag coef cient = 04) If the body dragcoef cient is set to 01 as recently proposed by Pennycuick (1997) energyconsumption in apping ight per km is reduced and energetic differencebetween soaring-gliding and apping ight become smaller but calculatedmaximum range speeds in apping ight are very high compared to ourobservations
We calculated energy and time consumption of different migratory routesbased on observed ight speeds for an average honey buzzard (Table 3) Notethat we did not take into account the time needed for refuelling consumedenergy (ie burned fat) and that we assumed zero wind Crossing the sea in apping-gliding ight (150 km distance) took on average about 4 hours andit was the time-minimising strategy for a honey buzzard (Table 3) Flyingalong the coast (250 km) required about 7 hours in soaring-gliding ightand 6 hours in apping-gliding ight Flying along the coast by soaring-gliding is the energy-minimising strategy for a honey buzzard and consumesabout 4 g fat (Table 3) Flapping-gliding ight along the coast is energeticallythe most expensive but never the fastest strategy Thus black kites andhoney buzzards migrated predominantly according to the energy minimising
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 395
TABLE 3 Observed airspeeds and corresponding estimated ight durationand fat consumption for an average honey buzzard when ying along the
coast or across the sea in different ight styles
Flight style Soaring-gliding Horizontal Horizontal apping-gliding apping-gliding
Flight route Flying along the coast Flying along the coast Crossing the seaDistance 250 km 250 km 150 km
Airspeed ms 101 (81-123) 108 (88-133) 108 (88-133)Flight duration hmin 652 (538-834) 626 (513-753) 351 (308-444)Fat consumption g 41 (34-55) 266 (230-298) 158 (138-179)for ight
Average honey buzzard = body mass 08 kg wing span 13 m wing area 025 m2 Medianand 25-75 range are given in brackets Calculations on fat consumption are based onPennycuickrsquos (1989) program of bird ight performance version 11 and birds are assumedto maximize cross-country speed in soaring-gliding ight (Spaar amp Bruderer 1997a)
strategy because a large proportion of the birds followed the coast in soaring-gliding ight
For species such as falcons or small harriers the difference in energyconsumption between apping and soaring-gliding ight is smaller Asthese species regularly use apping-gliding ight also over land or combinesoaring with apping-gliding ight differences in energy consumptionbetween land and sea route become even smaller and only short detours areenergetically favourable By combining soaring and apping-gliding ightsmaller raptors such as falcons or harriers are able to y under varyingenvironmental conditions to extend their migratory activity to times whenthermal updrafts are absent and to migrate even at night (Stark amp Liechti1993 Spaar amp Bruderer 1997b Spaar et al 1998)
Flight altitudes
Flight altitudes differed between the species groups partly due to differencesin migratory routes Birds crossing the sea ew considerably higher thanthose following the coast Kerlinger (1984) made similar observations atthe Delaware Bay birds crossing the bay ew at higher altitudes thanthose keeping on land He explained the higher ight altitudes by the bettervisibility of the other shore However the African coast is not visible fromour study site Some falcons even started crossing the sea after sunset
396 MEYER SPAAR amp BRUDERER
indicating that they continued migration into the night these birds had to y at least three hours until they reached the African coast The higher ightaltitudes of birds crossing the sea at our study site was mainly caused bytopographical features birds ying with southerly directions were forced toclimb higher up when crossing the mountains chains (1500 m height) about10-15 km northward of the coast and they afterwards maintained these ightlevels when approaching the sea
Flight altitudes of birds migrating along the coast were similar to altitudesobserved in North America and southern Israel with most raptors yingbelow 1000 m above ground and lt 5 above 1500 m (Kerlinger 19841989 Bruderer et al 1994 Spaar 1996)
Flight characteristics in soaring-gliding
Flight characteristics in soaring-gliding ight did not differ between thespecies groups (Table 2) As shown for raptor migration in Israel climb-ing rates do not depend primarily on morphological features but on environ-mental conditions (Spaar 1997) In general mean climbing rates in southernSpain were lower than in the semi-arid desert in Israel (Spaar 1997) Meanduration of the completed circling phases was shorter in Israel (88 plusmn 52 sSpaar 1995 1999) than in Spain In contrast to Israel (Spaar 1995 1997)circling time did not increase with increasing climbing rate Compared tothe semi-desert the development of thermal convection in southern Spainwas probably less stable and birds circled as long as possible in updraftsbecause they could not rely to encounter a next strong thermal within glid-ing distance This might also explain why inter-thermal gliding was oftensupported by apping it reduces sinking rate and prolongs gliding distance
In raptors migrating along the coast ight altitude and gliding airspeedswere related to thermal strength raptors circling in strong thermals hadhigher ight altitudes and higher inter-thermal gliding airspeeds than thosein weak thermals This is consistent with optimal ight theory for birds max-imising cross-country speed in soaring-gliding ight (Pennycuick 1989) asdemonstrated for steppe buzzards Buteo buteo vulpinus (Spaar amp Bruderer1997a)
Flight characteristics in horizontal apping-gliding
According to optimal ight theory maximum range speed decreases withfollowing winds and increases with head- and sidewinds (Pennycuick 1975
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 397
Liechti et al 1994) In raptors ying along the coast we observed decreas-ing airspeeds with increasing tailwind-support whereas birds crossing thesea did not reduce airspeeds Why did the birds react differently Sea cross-ings imply higher risks there is no possibility to land and weather (wind)conditions may change rapidly Higher airspeeds in following winds lead toshorter crossing times and lower the risk of unpredictable weather changesFurthermore drift by sidewinds may considerably extent crossing distancesand in order to reduce drift raptors increased airspeeds with increasingsindwind-components In contrast to sharp-shinned hawks Accipiter stria-tus avoiding crossings in strong lateral winds (Kerlinger 1985) we observedmany raptors to cross the sea in strong north-westerly (ie lateral) winds ifthey provided at least partial tailwind-support
Conclusions
Our observations at an average site at the southern coast of Spain showthat differences in water crossing behaviour are mainly species-speci cpresumably due to the costs for different ight strategies additionallyindividuals reacted to environmental factors such as wind and thermalconditions In medium sized raptors such as honey buzzard soaring-gliding ight along the coast is energy-minimising whereas crossing the sea istime-minimising Sea crossings occurred preferably with a good tailwind-support but also under strong sidewinds Raptors crossing the sea did notreduce airspeeds in following winds but increased airspeeds with increasingsidewinds They seemed to minimise the water crossing duration and theeffect of drift by sidewinds Under good thermal conditions birds regularlyfollowed the coast despite a lower tailwind-support compared to crossing thesea
References
Agostini N amp Logozzo D (1995) Autumn migration of honey buzzards in Southern Italymdash J Raptor Res 29 p 275-277
mdash mdash Logozzo D amp Colero C (1997) Migration of ocks of honey buzzards in SouthernItaly and Malta mdash J Raptor Res 31 p 84-86
mdash mdash Malara G Neri F Mollicone D amp Melotto S (1994) Flight strategies of honeybuzzards during spring migration across the Central Mediterranean mdash Avocetta 18p 73-76
398 MEYER SPAAR amp BRUDERER
Bannermann D-A amp Bannermann WM (1971) Handbook of the birds of Cyprus andmigrants of the Middle East mdash Oliver amp Boyd Edinburgh
Batschelet E (1981) Circular statistics in biology mdash Academic Press LondonBaudinette RV amp Schmidt-Nielsen K (1974) Energy costs of gliding ight in herring
gulls mdash Nature 248 p 83-84Beaman M amp Galea C (1974) The visible migration of raptors over the Maltese Islands
mdash Ibis 116 p 419-431Bernis F (1975) Migracion de Falconiformes y Ciconea spp por Gibraltar mdash IV Avance
sobre recuentos en el verano-otontildeo 1974 mdash Ardeola 21 p 581-592Bijlsma RG (1987) Bottleneck areas for migratory birds in the Mediterranean region mdash
ICBP Study report No 18 CambridgeBruderer B amp Liechti F (1998) Flight behaviour of nocturnally migrating birds in coastal
areas mdash crossing or coasting mdash J Avian Biol 29 p 499-510mdash mdash Blitzblau S amp Peter D (1994) Migration and ight behaviour of honey buzzards
Pernis apivorus in southern Israel observed by radar mdash Ardea 82 p 111-122mdash mdash Steuri T amp Baumgartner M (1995) Short-range high-precision surveillance of
nocturnal migration and tracking of single targets mdash Israel J Zool 41 p 207-220Evans PR amp Lathbury GW (1972) Raptor migration across the Strait of Gibraltar mdash Ibis
115 p 572-585Galea C amp Massa B (1985) Notes on the raptor migration across the central Mediter-
ranean mdash In Conservation studies on raptors (I Newton amp RD Chancellor eds)ICBP Technical Publication No 5 p 257-261
Gensboslashl B (1997) Greifvoumlgel mdash BLV VerlagsgesellschaftMuumlnchenGustin M amp Pizzari T (1998) Migratory pattern in the genus Circus sex and age
differential migration in Italy mdash Ornis Svecica 8 p 23-26Kerlinger P (1984) Flight behaviour of sharp-shinned hawks during migration 2 Over
water mdash Anim Beh 32 p 1029-1034mdash mdash (1985) Water-crossing behaviour of raptors during migration mdash Wilson Bull 97
p 109-113mdash mdash (1989) Flight strategies of migrating hawks mdash University of Chicago Press Chicagomdash mdash Bingman VP amp Able KP (1985) Comparative ight behaviour of migrating hawks
studied with tracking radar during autumn in central New York mdash Can J Zool 63p 755-761
Kjelleacuten N (1992) Differential timing of autumn migration between sex and age groups inraptors at Falsterbo Sweden mdash Orn Scan 23 p 420-434
Liechti F (1993) Naumlchtlicher Vogelzug im Herbst uumlber Suumlddeutschland Winddrift undKompensation mdash J Ornithol 134 p 373-404
mdash mdash Hedenstroumlm A amp Alerstam T (1994) Effects of sidewinds on optimal ight speedsof birds mdash J theor Biol 170 p 219-225
Lustick S Batterby B amp Kelty M (1978) Behavioural thermoregulation orientationtoward the sun in herring gulls mdash Sience 200 p 81-83
Pennycuick C (1975) Mechanics of ight mdash In Avian biology Vol 5 (DS Farner ampRJ King eds) Academic Press New York p 1-75
mdash mdash (1989) Bird ight performance a practical calculation manual mdash Oxford UniversityPress Oxford
mdash mdash (1997) Actual and lsquooptimumrsquo ight speeds eld data reassessedmdash J Exp Biol 200p 2355-2361
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 399
Richardson WJ (1978) Timing and amount of bird migration in relation to weather areview mdash Oikos 30 p 224-272
Spaar R (1995) Flight behaviour of steppe buzzard (Buteo buteo vulpinus) during springmigration in southern Israel a tracking radar study mdash Israel J Zool 41 p 489-500
mdash mdash (1996) Flight behaviour of migrating raptors in southern Israel mdash PhD thesisUniversity of Basel Verlag Schweizerische Vogelwarte Sempach
mdash mdash (1997) Flight strategies of migrating raptors a comparative study of interspecicvariation in ight characteristicsmdash Ibis 139 p 523-535
mdash mdash (1999) Flight behaviour of migrating raptors under varying environmental conditionsmdash In Proc 22 Int Ornithol Congr (N Adams amp R Slotow eds) University of NatalDurban p 1844-1862
mdash mdash amp Bruderer B (1997a) Optimal ight behaviour of soaring migrants A case studyof migrating steppe buzzards Buteo buteo vulpinus mdash Behav Ecol 8 p 288-297)
mdash mdash amp Bruderer B (1997b) Migration by apping or soaring ight strategies of marshMontagursquos and pallid harriers in southern Israel mdash Condor 99 p 458-469
mdash mdash Stark H amp Liechti F (1998) Migratory ight strategies of Levant sparrowhawkstime or energy minimization mdash Anim Behav 56 p 1185-1197
Stark H amp Liechti F (1993) Do Levant sparrowhawks Accipiter brevipes also migrate atnight mdash Ibis 135 p 233-236
Thiollay J-M (1977) Importance des populations de rapaces migrateurs en mediterraneeoccidentale mdash Alauda 45 p 115-121

MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 381
Methods
Study site and data recording
Raptor migrationwas studied at the south coast of the Iberian Peninsula 25 km east of Malagaand 140 km north-east of Gibraltar (Fig 1) The site was 100 m inland from the shoreline and30 m above sea level The topography is not expected to have a concentration effect exceptfor birds approaching from the east and following the coast afterwards towards GibraltarTherefore the study site is considered as an average site at the east-west leading coast Thedistance to the African coast is approximately 150 km and the opposite coast is out of sightto a migratory bird
Observations lasted from 10 August to 24 October 1996 A tracking radar of the typelsquoSuper edermausrsquo was used to record the ight paths of migrating birds This radar allowsto track a single medium-sized raptor at distances up to 8 km (for details see Bruderer et al1995) While trackinga bird its exact position (x y and z co-ordinates)was digitally recordedevery second and the ight path visualised on a computer screen The species was identi edusing a 125 times telescope mounted parallel to the radar beam Only raptors identi edat least to the genus were included in the analysis Emphasis was on long ight tracksincluding entire phases of soaring and gliding The birds were visually observed duringthe entire ight path and their ight behaviour (direction ight style) was recorded Birdsnoted as locals or not showing clearly directed tracks were excluded from the analysis Visualobservation lasted from 800 h to 2000 h occasionally longer A daily break of two hoursin the afternoon was necessary for operational reasons (data backup routine evaluations) Itwas shifted from day to day to obtain records from the whole day In total 386 tracks ofmigrating raptors were recorded including honey buzzard (N = 81) black kite (N = 40)harriers (N = 102) including marsh harrier Circus aeruginosus Montagursquos harrier Circuspygargus and unidenti ed harriers osprey (N = 17) and falcons (N = 146) includingperegrine falcon Falco peregrinus hobby Falco subbuteo lesser kestrel Falco naumannikestrel Falco tinnunculus and unidenti ed falcons Unidenti ed falcons might include a fewsparrowhawks Accipiter nisus because at large distance or under poor visibility it was verydif cult to distinguish these genera
Wind speed and direction at all ight levels were obtained from pilot balloons which werereleased and tracked every four hours up to 3000 m above ground
Migratory ight routes
Directions in gliding or apping-gliding ight were considered to be the preferred migratorydirection To compare ight directions we categorised the raptor tracks by ight directionand position above ground (Fig 1) raptors lsquo ying along the coastrsquo had ight directions of240deg -300deg and ew above land whereas birds lsquo ying across the bayrsquo (240deg -270deg ) at leastpartly ew above sea Raptors lsquocrossing the searsquo ew out to the sea with directions of 95deg -240deg Birds ying north- or eastwards (300deg -95deg ) were not included in the further analysisTo separate raptors lsquocrossing the searsquo and lsquo ying across the bayrsquo we took the angle of thecoastal tip near Fuengirola (direction 240deg distance about 50 km) which is visible undergood atmospheric conditions and therefore is a possible leading point for birds ying acrossthe bay of Malaga
382 MEYER SPAAR amp BRUDERER
Fig 1 Location of the radar and sectors associated with the following migratory routeslsquocrossing the searsquo (directions of 95deg -240deg ) lsquo ying across the bayrsquo (birds at least partiallyabove the water body but keeping an angle closer to the coast ie 240deg -270deg ) lsquo ying alongthe coastrsquo (directions of 240deg -300deg above land) lsquoreversed migrationrsquo (directions of 300deg -
95deg ) The circle indicates the range of the radar for tracking medium-sized raptors
Analysis of tracks
Similarly to Spaar (1996) for raptor tracks in Israel the tracks were subdivided into intervalsof 10 s and different ight phases marked interactively on the computer screen We de nedthe following ight styles (1) lsquosoaring-gliding ightrsquo consisted of thermal circling and inter-thermal gliding phases (vertical speed lt shy 05 ms) (2) lsquosoaring combined with apping ightrsquo included birds in thermal circling and inter-thermal apping-gliding ight (verticalspeed 0 plusmn 05 ms) and (3) lsquohorizontal apping-gliding ightrsquo (vertical speed 0 plusmn 05 ms)
Pilot balloons released and tracked every four hours informed about wind direction andwind speed at all ight levels Airspeed and heading while gliding were calculated bysubtractingthe wind vector at the ight altitude from the track vector (Liechti 1993) Air- andgroundspeed are the velocities of the bird relative to the air and ground respectively Cross-country speed relative to the ground in soaring-gliding ight was calculated as the averagespeed over a distance of at least 2 km Cross-country speed relative to the air was derived bysubtracting the wind vector from the vector of cross-country speed relative to the ground
In the diagrams the time of day is expressed as a percentage of the time between sunrise(0) and sunset (100) in order to compensate for the time shift of dawn and dusk duringthe observation period Circular statistics are based on Batschelet (1981) Tailwind- and
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 383
sidewind-componentare the wind speeds in track direction and rectangular to track directionrespectivelyTo analyse seasonal trends in the proportion of sea crossing 5-day periods withat least two ight paths of a species group were included in the analysis
Results
Migratory directions and ight styles
Track directions of all raptors were south-westerly (238deg plusmn 34deg mean plusmn SD)and coincided with the direction towards the visible tip of the coastline nearFuengirola corresponding headings were 246deg plusmn 36deg (Fig 1) Tracks andheadings differed signi cantly between the species groups (Watson-Williamsmultisample test tracks F = 1884 df = 4 N = 382 p lt 00001headings F = 2148 df = 4 N = 382 p lt 00001) Black kites(track 269deg heading 276deg ) and honey buzzards (252deg 263deg ) kept westerly ight directions mainly following the east-west leading coastline whereasharriers (229deg 231deg ) falcons (228deg 240deg ) and ospreys (225deg 221deg ) hadsouth-westerly ight directions (Fig 2) Thus regarding the water-crossingbehaviour two main groups occurred no black kites and only few honeybuzzards (17 of tracks) but large proportions of harriers (65) falcons(57) and ospreys (73) crossed the sea (Fig 3A) The proportion of birds ying across the bay was below 15 of tracks in all groups For all groupswind directions were predominantly from north-west or from east (Fig 2)
The proportion of sea crossings (5-day periods) increased towards theend of the migratory season in honey buzzards and falcons (honey buzzardR 2 = 082 N = 6 p lt 001 falcons R 2 = 068 N = 12 p lt 001) Noseasonal trend was found in marsh and Montagursquos harriers (marsh harrierR 2 = shy 025 N = 5 NS Montagursquos harrier R 2 = shy 015 N = 5 NS)
According to the different proportions of migratory directions propor-tions of migratory ight styles differed between species groups soaring-gliding was the main ight style of honey buzzards and black kites (50and 71 of total tracking time respectively Fig 3B) Flapping-gliding ight was extensively used by harriers (62) falcons (66) and ospreys(86) Proportion of apping-gliding ight was lower in marsh harriers(50) than in Montagursquos harriers (76) Raptors combining soaring with apping-gliding ight had lower climbing rates than birds in pure soaring-gliding ight indicating that apping might be a compensation for weakthermal conditions (122 ms N = 39 and 158 ms N = 77 respectivelyt = shy 221 p lt 005)
384 MEYER SPAAR amp BRUDERER
Fig 2 Distribution of track (line) and heading (shaded) directions for the different raptorgroups Corresponding distribution of wind directions at the levels of the tracked birds given
at the right
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 385
Fig 3 Proportion of different migratory routes (A) and of time spent in different ightstyles (B without reversed migrants) in the species groups For de nitions see methods Totaltracking time honey buzzard (512 min) black kite (174 min) harriers (369 min) falcons
(497 min) and osprey (107 min)
In uence of environmental conditions on migratory direction
In weak winds (ie wind speed below 4 ms) numbers of falcons harriersand ospreys crossing the sea or following the coast were similar for allwind directions (df = 3 x 2 = 28 NS and x 2 = 64 NS respectivelyTable 1) In strong winds sea crossings occurred in westerly northerlyand easterly winds (ie side and following winds) but not in southerly
386 MEYER SPAAR amp BRUDERER
TABLE 1 Comparison of wind situations and migratory routes
Crossing the sea Flying along the coast
weak wind strong wind weak wind strong wind
West 13 19 5 9North 16 40 9 15East 17 22 7 9South 9 0 1 1
x 2 28 397 64 116p 04 lt 0001 01 lt 001
Number of tracks given Wind direction West (225deg -315deg ) North (315deg -45deg ) East (45deg -135deg ) South (135deg -225deg ) Weak winds are below 4 ms Tracks of harriers falcons andospreys are included
winds (ie opposing winds df = 3 x 2 = 397 p lt 0001 Table 1)Strong northerly winds providing tailwind-support were preferred comparedto strong westerly and easterly winds (df = 2 x 2 = 96 p lt 001) In birdsfollowing the coast under strong winds numbers of observed birds showeda similar pattern as in birds crossing the sea they were higher in northerlywesterly and easterly compared to southerly winds (df = 3 x 2 = 116p lt 001 Table 1) The lower number of birds following the coast instrong southerly winds might be an artefact migratory birds were driftedtowards the mountain chains northerly of the radar station and were out ofthe tracking range of the radar
The tailwind-support achieved by the birds did only partly explain fordifferent migratory routes tailwind-component (t-c) of raptors crossingthe sea was higher than a theoretical t-c for the same birds assuming to y along the coast (270deg paired t-test N = 149 t = 78 p lt 0001Fig 4) On the other hand t-c of raptors ying along the coast was lowerthan the theoretical t-c if crossing the sea (210deg N = 138 t = shy 44p lt 0001) Thus many birds ew along the coast with a lower tailwind-support than if they would have crossed the sea There is an indicationthat good thermal conditions might stimulate birds to migrate along thecoast with a lower tailwind-support compared to sea crossing birds yingalong the coast instead of crossing the sea with a higher tailwind-supporthad higher climbing rates in thermal circling than birds ying along coastwith a higher tailwind-support compared to crossing the sea (mean plusmn SD
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 387
Fig 4 The observed tailwind-components in raptors crossing the sea (A left) and yingalong the coast (B left) versus the theoretical tailwind-componentsfor the alternativestrategyfor the same birds assumed to y along the coast (A right) and crossing the sea (B right)Paired t -test (A) N = 149 t = 78 p lt 0001 () (B) N = 138 t = shy 44 p lt 0001
18 plusmn 09 ms N = 29 and 10 plusmn 05 ms N = 17 respectively t-testt = 31 p lt 0005)
Flight altitudes
For all raptors the median of ight altitudes was below 1000 m above groundlevel (agl) and the 90 altitude limit was below 1800 m agl (Fig 5)Median altitudes differed signi cantly between the species groups (blackkite median 636 m N = 40 honey buzzard 862 m N = 81 harriers910 m N = 102 falcons 949 m N = 146 osprey 951 m N = 17 Kruskal-Wallis ANOVA H = 1835 df = 4 N = 386 p lt 0005) Harriers andfalcons had signi cantly higher ight altitudes than black kites (GT2-testp lt 001) Maximum observed ight levels were at about 1400 m agl inblack kites at 1800 m in ospreys at 2000 m in harriers at 2200 m in honeybuzzards and at 2600 m in falcons (Fig 5) No black kites were tracked in theearly afternoon when ight altitudes of soaring migrants are usually highpartly due to long operational breaks in the rst week of our observationperiod Thus maximum ight levels of black kites were probably higher
388 MEYER SPAAR amp BRUDERER
Fig 5 Flight altitudes above ground (mean per track) in the course of the day (0 =sunrise 100 = sunset) Median 636 m for black kite 862 m for honey buzzard 910 m
for harriers 949 m for falcons and 951 m for osprey
Flight altitudes of honey buzzards black kites and harriers increased fromsunrise towards noon and were more or less stable thereafter (Fig 5) Falconsachieved high ight altitudes already soon after sunrise by apping-gliding ight In falcons ight altitudes differed between migratory routes birdscrossing the sea had higher ight altitudes than birds ying across the bayand along the coast (Fig 6 Kruskal-Wallis ANOVA H = 181 df =2 p = 00001 GT2-test crossing the sea versus ying across the bay(p lt 001) and versus along the coast (p lt 005)) In honey buzzards andharriers differences in ight altitudes between migratory routes showed thesame tendency but did not differ statistically (Fig 6)
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 389
Fig 6 Flight altitudes above ground (mean per track) for different migratory routes inhoney buzzard harriers and falcons Boxes are medians with 25-75 quartiles whiskersshow 10-90 range dots are outliers (ie outside the 15 box length from the upper and
lower value) GT2-test p lt 005 p lt 001
Fig 7 Mean climbing rates in thermal circling in the course of the day (0 = sunrise100 = sunset) Climbing rate did not differ between species groups (ANOVA F = 116
df = 4 N = 116 NS)
Flight characteristics in soaring-gliding
Thermal circling phases were recorded in 116 of 386 tracks and averageclimbing rate was 146 plusmn 084 ms (N = 116) Climbing rates increasedduring the morning hours (Fig 7 time of day lt 50 Spearman rankcorrelation N = 61 r = 034 p lt 001) Strong thermal convection aroundnoon allowed climbing rates of up to 45 ms From noon to sunset climbing
390 MEYER SPAAR amp BRUDERER
TA
BL
E2
Flig
htch
arac
teri
stic
sof
diffe
rent
spec
ies
grou
ps(m
s)
hone
ybu
zzar
dbl
ack
kite
harr
iers
falc
ons
ospr
eyF
Soar
ing
glid
ing
Cli
mbi
ngra
te1
35plusmn
077
(48)
171
plusmn0
97(2
2)1
54plusmn
087
(21)
134
plusmn0
84(2
4)1
86(1
)1
16N
SC
ross
-cou
ntry
grou
ndsp
eed
86
plusmn0
4(2
8)7
6plusmn
07
(7)
93
plusmn0
9(9
)9
3plusmn
10
(6)
89
(1)
079
NS
Cro
ss-c
ount
ryai
rspe
ed10
4plusmn
42
(28)
99
plusmn3
6(7
)9
3plusmn
25
(9)
95
plusmn3
1(1
0)11
3(1
)0
27N
SIn
ter-
ther
mal
grou
ndsp
eed
116
plusmn3
2(4
7)10
8plusmn
25
(21)
121
plusmn2
5(2
1)11
2plusmn
32
(23)
114
(1)
075
NS
Inte
r-th
erm
alai
rspe
ed12
8plusmn
37
(47)
134
plusmn3
0(2
1)12
0plusmn
30
(21)
127
plusmn3
6(2
3)17
0(1
)0
57N
S
Hor
izon
tal
appi
ng-g
lidi
ngi
ght
Gro
unds
peed
138
plusmn2
5(1
6)10
9plusmn
26
(9)
116
plusmn1
9(6
1)12
2plusmn
36
(68)
146
plusmn1
9(1
0)2
47
Air
spee
d11
1plusmn
23
(16)
103
plusmn2
3(9
)11
2plusmn
18
(58)
113
plusmn2
4(6
8)13
2plusmn
30
(10)
387
Mea
nplusmn
SD
(N)
Dif
fere
nces
wer
ete
sted
byA
NO
VA
S
igni
ca
nce
leve
l
plt
005
plt
001
H
oriz
onta
l
appi
ng-g
lidi
ng
ight
was
incl
uded
only
ifve
rtic
alsp
eeds
did
note
xcee
d0
plusmn0
5m
san
dif
noth
erm
alci
rcli
ngw
asob
serv
ed
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 391
rates did not decrease signi cantly (Spearman rank correlation N = 55r = shy 015 NS) Climbing rates in thermal circling below 1 ms occurredduring the whole day They did not differ between species groups (Table 2ANOVA F = 116 df = 3 N = 115 NS) Flight altitudes increased withincreasing climbing rates in birds lsquo ying along the coastrsquo (r = 049 N = 64p lt 00001) Average circling duration in completed soaring phases was106 plusmn 67 s (N = 80) and it was not related to climbing rate (r = 002N = 80 NS)
Mean airspeed and groundspeed in inter-thermal gliding did not differbetween the species groups (Table 2 ANOVA F = 057 and F = 075respectively df = 3 N = 112 NS) and means per species ranged between120-134 ms and 108-121 ms respectively Raptors increased airspeedsin inter-thermal gliding with increasing climbing rates (r = 052 N =113 p lt 00001) indicating that they maximised cross-country speeds insoaring-gliding ight (Pennycuick 1989 Spaar amp Bruderer 1997a) Meancross-country speeds in soaring-gliding did not differ between species groups(ANOVA cross-country airspeed F = 079 df = 3 N = 54 NS range 93-113 ms cross-country groundspeed F = 027 df = 3 N = 54 NS range76-93 ms)
Flight characteristics in horizontal apping-gliding
This ight style was considered if vertical speeds did not exceed 0 plusmn 05 msand no thermal circling was observed within a track Air- and groundspeedsdiffered between species groups (ANOVA airspeed F = 247 df = 4N = 161 p lt 005 groundspeed F = 387 df = 4 N = 164p lt 0005 Table 2) Ospreys had higher groundspeeds than harriers (GT2-test p lt 005) and higher ground- and airspeeds than black kites (p lt 005)Mean airspeeds per species ranged between 103-132 ms correspondinggroundspeeds between 109-146 ms
Flapping-gliding migrants reached signi cantly higher cross-country air-speeds (113 plusmn 22 ms N = 161) and groundspeeds (122 plusmn 31 ms N =164) than birds in soaring-gliding ight (airspeed 100 plusmn 36 ms N = 55Welch-test t = 249 df = 68 p lt 001 groundspeed 87 plusmn 25 msN = 55 t = 846 df = 114 p lt 00005)
In horizontal apping-gliding ight reactions to different wind con-ditions depended on migratory route raptors ying along the coast de-
392 MEYER SPAAR amp BRUDERER
Fig 8 Airspeed in horizontal apping-gliding ight versus tailwind- and sidewind-components for different migratory routes Linear regression (A) excluding one value inbrackets y = shy 0256x + 1128 R 2 = 0215 N = 56 p lt 0001 (B) r = shy 010 N = 57NS (C) r = shy 007 N = 85 NS (D) excluding two values in brackets y = 0191x+ 1072
R2 = 005 N = 84 p lt 005
creased airspeed with increasing tailwind-component (Fig 8A) but air-speed was not related to sidewind-component (Fig 8B) In contrast in rap-tors crossing the sea no relation was found between airspeed and tailwind-component (Fig 8C) but airspeed increased with increasing sidewind-component (Fig 8D)
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 393
Discussion
There are only few studies on the ight behaviour of migrating raptors cross-ing the sea mainly carried out at concentration points such as narrow wa-ter straits or peninsulas (Evans amp Lathbury 1972 Bernis 1975 Kerlinger1984) and none by tracking radar As these studies were limited to visualobservation mainly high altitudinal migration was often not detected (Ker-linger et al 1985) Our approach is different because (i) we observed raptormigration at an average coast site (ii) we used a tracking radar and (iii) wemeasured wind conditions at all ight levels
Migratory routes and in uence of environmental conditions
Although black kites and in larger numbers honey buzzards undertakelong sea-crossings in the Central Mediterranean (Beaman amp Galea 1974Agostini et al 1994) in our study no black kites and few honey buzzardswere observed attempting the sea crossing The majority of harriers falconsand ospreys crossed the sea Sea crossing honey buzzards occurred towardsthe end of the migratory season and were probably juveniles as in honeybuzzards adults migrate two weeks earlier than juveniles (Kjelleacuten 1992Agostini et al 1997) In contrast to the adults which often migrate in large ocks juveniles mainly migrate singly and have no experience about themigratory ight route This may lead to a higher proportion of juvenilescrossing the sea
Wind and thermal conditions affected migratory directions many harri-ers falcons and ospreys crossed the sea in favourable following and sidewinds but no sea crossings were observed in strong opposing winds This isreasonable because energy and time consumption for the sea crossing andtherefore the risk of exhaustion drastically increase in opposing winds How-ever tailwind-support did not fully explain the decisions on migratory routesbecause regularly birds followed the coast with a lower tailwind-supportcompared to crossing the sea These birds following the coast reached highclimbing rates in thermal circling and this might be compensating for thelower tailwind-support Under good thermal conditions birds were able tomigrate in soaring-gliding ight which consumes much less energy com-pared to apping-gliding ight when crossing the sea (see below Energyand time consumption) Anyway birds in bad shape for example those with
394 MEYER SPAAR amp BRUDERER
low fat reserves should follow the coast in order to minimise the risk ofstarvation to feed when ever it is possible and to y in soaring-gliding ight
At the same observation site migratory directions of nocturnal migrants ying below 2000 m above ground shift from 190deg to 250deg within the nightthe majority of the birds cross the sea when approaching the coast in the rsthalf of the night whereas they hesitate later on (Bruderer amp Liechti 1998)Thus the motivation to cross large barriers seems to decrease after somehours of migratory ight
Energy and time consumption
Energy consumption in apping ight disproportionately increases withbody mass while in soaring-gliding ight it is 2-3 times the basal metabolicrate (BMR) only (for herring gull Larus argentatus Baudinette amp Schmidt-Nielson 1974 Lustick et al 1978) In honey buzzards the average observedairspeed in apping-gliding ight was 111 ms and fat consumption perdistance (gkm) for an average honey buzzard (body mass in Table 3) atthis speed is about 6 times higher than in soaring-gliding ight (calculatedafter Pennycuick (1989 version 11) energy consumption in soaring-gliding ight = 2 times BMR body drag coef cient = 04) If the body dragcoef cient is set to 01 as recently proposed by Pennycuick (1997) energyconsumption in apping ight per km is reduced and energetic differencebetween soaring-gliding and apping ight become smaller but calculatedmaximum range speeds in apping ight are very high compared to ourobservations
We calculated energy and time consumption of different migratory routesbased on observed ight speeds for an average honey buzzard (Table 3) Notethat we did not take into account the time needed for refuelling consumedenergy (ie burned fat) and that we assumed zero wind Crossing the sea in apping-gliding ight (150 km distance) took on average about 4 hours andit was the time-minimising strategy for a honey buzzard (Table 3) Flyingalong the coast (250 km) required about 7 hours in soaring-gliding ightand 6 hours in apping-gliding ight Flying along the coast by soaring-gliding is the energy-minimising strategy for a honey buzzard and consumesabout 4 g fat (Table 3) Flapping-gliding ight along the coast is energeticallythe most expensive but never the fastest strategy Thus black kites andhoney buzzards migrated predominantly according to the energy minimising
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 395
TABLE 3 Observed airspeeds and corresponding estimated ight durationand fat consumption for an average honey buzzard when ying along the
coast or across the sea in different ight styles
Flight style Soaring-gliding Horizontal Horizontal apping-gliding apping-gliding
Flight route Flying along the coast Flying along the coast Crossing the seaDistance 250 km 250 km 150 km
Airspeed ms 101 (81-123) 108 (88-133) 108 (88-133)Flight duration hmin 652 (538-834) 626 (513-753) 351 (308-444)Fat consumption g 41 (34-55) 266 (230-298) 158 (138-179)for ight
Average honey buzzard = body mass 08 kg wing span 13 m wing area 025 m2 Medianand 25-75 range are given in brackets Calculations on fat consumption are based onPennycuickrsquos (1989) program of bird ight performance version 11 and birds are assumedto maximize cross-country speed in soaring-gliding ight (Spaar amp Bruderer 1997a)
strategy because a large proportion of the birds followed the coast in soaring-gliding ight
For species such as falcons or small harriers the difference in energyconsumption between apping and soaring-gliding ight is smaller Asthese species regularly use apping-gliding ight also over land or combinesoaring with apping-gliding ight differences in energy consumptionbetween land and sea route become even smaller and only short detours areenergetically favourable By combining soaring and apping-gliding ightsmaller raptors such as falcons or harriers are able to y under varyingenvironmental conditions to extend their migratory activity to times whenthermal updrafts are absent and to migrate even at night (Stark amp Liechti1993 Spaar amp Bruderer 1997b Spaar et al 1998)
Flight altitudes
Flight altitudes differed between the species groups partly due to differencesin migratory routes Birds crossing the sea ew considerably higher thanthose following the coast Kerlinger (1984) made similar observations atthe Delaware Bay birds crossing the bay ew at higher altitudes thanthose keeping on land He explained the higher ight altitudes by the bettervisibility of the other shore However the African coast is not visible fromour study site Some falcons even started crossing the sea after sunset
396 MEYER SPAAR amp BRUDERER
indicating that they continued migration into the night these birds had to y at least three hours until they reached the African coast The higher ightaltitudes of birds crossing the sea at our study site was mainly caused bytopographical features birds ying with southerly directions were forced toclimb higher up when crossing the mountains chains (1500 m height) about10-15 km northward of the coast and they afterwards maintained these ightlevels when approaching the sea
Flight altitudes of birds migrating along the coast were similar to altitudesobserved in North America and southern Israel with most raptors yingbelow 1000 m above ground and lt 5 above 1500 m (Kerlinger 19841989 Bruderer et al 1994 Spaar 1996)
Flight characteristics in soaring-gliding
Flight characteristics in soaring-gliding ight did not differ between thespecies groups (Table 2) As shown for raptor migration in Israel climb-ing rates do not depend primarily on morphological features but on environ-mental conditions (Spaar 1997) In general mean climbing rates in southernSpain were lower than in the semi-arid desert in Israel (Spaar 1997) Meanduration of the completed circling phases was shorter in Israel (88 plusmn 52 sSpaar 1995 1999) than in Spain In contrast to Israel (Spaar 1995 1997)circling time did not increase with increasing climbing rate Compared tothe semi-desert the development of thermal convection in southern Spainwas probably less stable and birds circled as long as possible in updraftsbecause they could not rely to encounter a next strong thermal within glid-ing distance This might also explain why inter-thermal gliding was oftensupported by apping it reduces sinking rate and prolongs gliding distance
In raptors migrating along the coast ight altitude and gliding airspeedswere related to thermal strength raptors circling in strong thermals hadhigher ight altitudes and higher inter-thermal gliding airspeeds than thosein weak thermals This is consistent with optimal ight theory for birds max-imising cross-country speed in soaring-gliding ight (Pennycuick 1989) asdemonstrated for steppe buzzards Buteo buteo vulpinus (Spaar amp Bruderer1997a)
Flight characteristics in horizontal apping-gliding
According to optimal ight theory maximum range speed decreases withfollowing winds and increases with head- and sidewinds (Pennycuick 1975
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 397
Liechti et al 1994) In raptors ying along the coast we observed decreas-ing airspeeds with increasing tailwind-support whereas birds crossing thesea did not reduce airspeeds Why did the birds react differently Sea cross-ings imply higher risks there is no possibility to land and weather (wind)conditions may change rapidly Higher airspeeds in following winds lead toshorter crossing times and lower the risk of unpredictable weather changesFurthermore drift by sidewinds may considerably extent crossing distancesand in order to reduce drift raptors increased airspeeds with increasingsindwind-components In contrast to sharp-shinned hawks Accipiter stria-tus avoiding crossings in strong lateral winds (Kerlinger 1985) we observedmany raptors to cross the sea in strong north-westerly (ie lateral) winds ifthey provided at least partial tailwind-support
Conclusions
Our observations at an average site at the southern coast of Spain showthat differences in water crossing behaviour are mainly species-speci cpresumably due to the costs for different ight strategies additionallyindividuals reacted to environmental factors such as wind and thermalconditions In medium sized raptors such as honey buzzard soaring-gliding ight along the coast is energy-minimising whereas crossing the sea istime-minimising Sea crossings occurred preferably with a good tailwind-support but also under strong sidewinds Raptors crossing the sea did notreduce airspeeds in following winds but increased airspeeds with increasingsidewinds They seemed to minimise the water crossing duration and theeffect of drift by sidewinds Under good thermal conditions birds regularlyfollowed the coast despite a lower tailwind-support compared to crossing thesea
References
Agostini N amp Logozzo D (1995) Autumn migration of honey buzzards in Southern Italymdash J Raptor Res 29 p 275-277
mdash mdash Logozzo D amp Colero C (1997) Migration of ocks of honey buzzards in SouthernItaly and Malta mdash J Raptor Res 31 p 84-86
mdash mdash Malara G Neri F Mollicone D amp Melotto S (1994) Flight strategies of honeybuzzards during spring migration across the Central Mediterranean mdash Avocetta 18p 73-76
398 MEYER SPAAR amp BRUDERER
Bannermann D-A amp Bannermann WM (1971) Handbook of the birds of Cyprus andmigrants of the Middle East mdash Oliver amp Boyd Edinburgh
Batschelet E (1981) Circular statistics in biology mdash Academic Press LondonBaudinette RV amp Schmidt-Nielsen K (1974) Energy costs of gliding ight in herring
gulls mdash Nature 248 p 83-84Beaman M amp Galea C (1974) The visible migration of raptors over the Maltese Islands
mdash Ibis 116 p 419-431Bernis F (1975) Migracion de Falconiformes y Ciconea spp por Gibraltar mdash IV Avance
sobre recuentos en el verano-otontildeo 1974 mdash Ardeola 21 p 581-592Bijlsma RG (1987) Bottleneck areas for migratory birds in the Mediterranean region mdash
ICBP Study report No 18 CambridgeBruderer B amp Liechti F (1998) Flight behaviour of nocturnally migrating birds in coastal
areas mdash crossing or coasting mdash J Avian Biol 29 p 499-510mdash mdash Blitzblau S amp Peter D (1994) Migration and ight behaviour of honey buzzards
Pernis apivorus in southern Israel observed by radar mdash Ardea 82 p 111-122mdash mdash Steuri T amp Baumgartner M (1995) Short-range high-precision surveillance of
nocturnal migration and tracking of single targets mdash Israel J Zool 41 p 207-220Evans PR amp Lathbury GW (1972) Raptor migration across the Strait of Gibraltar mdash Ibis
115 p 572-585Galea C amp Massa B (1985) Notes on the raptor migration across the central Mediter-
ranean mdash In Conservation studies on raptors (I Newton amp RD Chancellor eds)ICBP Technical Publication No 5 p 257-261
Gensboslashl B (1997) Greifvoumlgel mdash BLV VerlagsgesellschaftMuumlnchenGustin M amp Pizzari T (1998) Migratory pattern in the genus Circus sex and age
differential migration in Italy mdash Ornis Svecica 8 p 23-26Kerlinger P (1984) Flight behaviour of sharp-shinned hawks during migration 2 Over
water mdash Anim Beh 32 p 1029-1034mdash mdash (1985) Water-crossing behaviour of raptors during migration mdash Wilson Bull 97
p 109-113mdash mdash (1989) Flight strategies of migrating hawks mdash University of Chicago Press Chicagomdash mdash Bingman VP amp Able KP (1985) Comparative ight behaviour of migrating hawks
studied with tracking radar during autumn in central New York mdash Can J Zool 63p 755-761
Kjelleacuten N (1992) Differential timing of autumn migration between sex and age groups inraptors at Falsterbo Sweden mdash Orn Scan 23 p 420-434
Liechti F (1993) Naumlchtlicher Vogelzug im Herbst uumlber Suumlddeutschland Winddrift undKompensation mdash J Ornithol 134 p 373-404
mdash mdash Hedenstroumlm A amp Alerstam T (1994) Effects of sidewinds on optimal ight speedsof birds mdash J theor Biol 170 p 219-225
Lustick S Batterby B amp Kelty M (1978) Behavioural thermoregulation orientationtoward the sun in herring gulls mdash Sience 200 p 81-83
Pennycuick C (1975) Mechanics of ight mdash In Avian biology Vol 5 (DS Farner ampRJ King eds) Academic Press New York p 1-75
mdash mdash (1989) Bird ight performance a practical calculation manual mdash Oxford UniversityPress Oxford
mdash mdash (1997) Actual and lsquooptimumrsquo ight speeds eld data reassessedmdash J Exp Biol 200p 2355-2361
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 399
Richardson WJ (1978) Timing and amount of bird migration in relation to weather areview mdash Oikos 30 p 224-272
Spaar R (1995) Flight behaviour of steppe buzzard (Buteo buteo vulpinus) during springmigration in southern Israel a tracking radar study mdash Israel J Zool 41 p 489-500
mdash mdash (1996) Flight behaviour of migrating raptors in southern Israel mdash PhD thesisUniversity of Basel Verlag Schweizerische Vogelwarte Sempach
mdash mdash (1997) Flight strategies of migrating raptors a comparative study of interspecicvariation in ight characteristicsmdash Ibis 139 p 523-535
mdash mdash (1999) Flight behaviour of migrating raptors under varying environmental conditionsmdash In Proc 22 Int Ornithol Congr (N Adams amp R Slotow eds) University of NatalDurban p 1844-1862
mdash mdash amp Bruderer B (1997a) Optimal ight behaviour of soaring migrants A case studyof migrating steppe buzzards Buteo buteo vulpinus mdash Behav Ecol 8 p 288-297)
mdash mdash amp Bruderer B (1997b) Migration by apping or soaring ight strategies of marshMontagursquos and pallid harriers in southern Israel mdash Condor 99 p 458-469
mdash mdash Stark H amp Liechti F (1998) Migratory ight strategies of Levant sparrowhawkstime or energy minimization mdash Anim Behav 56 p 1185-1197
Stark H amp Liechti F (1993) Do Levant sparrowhawks Accipiter brevipes also migrate atnight mdash Ibis 135 p 233-236
Thiollay J-M (1977) Importance des populations de rapaces migrateurs en mediterraneeoccidentale mdash Alauda 45 p 115-121

382 MEYER SPAAR amp BRUDERER
Fig 1 Location of the radar and sectors associated with the following migratory routeslsquocrossing the searsquo (directions of 95deg -240deg ) lsquo ying across the bayrsquo (birds at least partiallyabove the water body but keeping an angle closer to the coast ie 240deg -270deg ) lsquo ying alongthe coastrsquo (directions of 240deg -300deg above land) lsquoreversed migrationrsquo (directions of 300deg -
95deg ) The circle indicates the range of the radar for tracking medium-sized raptors
Analysis of tracks
Similarly to Spaar (1996) for raptor tracks in Israel the tracks were subdivided into intervalsof 10 s and different ight phases marked interactively on the computer screen We de nedthe following ight styles (1) lsquosoaring-gliding ightrsquo consisted of thermal circling and inter-thermal gliding phases (vertical speed lt shy 05 ms) (2) lsquosoaring combined with apping ightrsquo included birds in thermal circling and inter-thermal apping-gliding ight (verticalspeed 0 plusmn 05 ms) and (3) lsquohorizontal apping-gliding ightrsquo (vertical speed 0 plusmn 05 ms)
Pilot balloons released and tracked every four hours informed about wind direction andwind speed at all ight levels Airspeed and heading while gliding were calculated bysubtractingthe wind vector at the ight altitude from the track vector (Liechti 1993) Air- andgroundspeed are the velocities of the bird relative to the air and ground respectively Cross-country speed relative to the ground in soaring-gliding ight was calculated as the averagespeed over a distance of at least 2 km Cross-country speed relative to the air was derived bysubtracting the wind vector from the vector of cross-country speed relative to the ground
In the diagrams the time of day is expressed as a percentage of the time between sunrise(0) and sunset (100) in order to compensate for the time shift of dawn and dusk duringthe observation period Circular statistics are based on Batschelet (1981) Tailwind- and
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 383
sidewind-componentare the wind speeds in track direction and rectangular to track directionrespectivelyTo analyse seasonal trends in the proportion of sea crossing 5-day periods withat least two ight paths of a species group were included in the analysis
Results
Migratory directions and ight styles
Track directions of all raptors were south-westerly (238deg plusmn 34deg mean plusmn SD)and coincided with the direction towards the visible tip of the coastline nearFuengirola corresponding headings were 246deg plusmn 36deg (Fig 1) Tracks andheadings differed signi cantly between the species groups (Watson-Williamsmultisample test tracks F = 1884 df = 4 N = 382 p lt 00001headings F = 2148 df = 4 N = 382 p lt 00001) Black kites(track 269deg heading 276deg ) and honey buzzards (252deg 263deg ) kept westerly ight directions mainly following the east-west leading coastline whereasharriers (229deg 231deg ) falcons (228deg 240deg ) and ospreys (225deg 221deg ) hadsouth-westerly ight directions (Fig 2) Thus regarding the water-crossingbehaviour two main groups occurred no black kites and only few honeybuzzards (17 of tracks) but large proportions of harriers (65) falcons(57) and ospreys (73) crossed the sea (Fig 3A) The proportion of birds ying across the bay was below 15 of tracks in all groups For all groupswind directions were predominantly from north-west or from east (Fig 2)
The proportion of sea crossings (5-day periods) increased towards theend of the migratory season in honey buzzards and falcons (honey buzzardR 2 = 082 N = 6 p lt 001 falcons R 2 = 068 N = 12 p lt 001) Noseasonal trend was found in marsh and Montagursquos harriers (marsh harrierR 2 = shy 025 N = 5 NS Montagursquos harrier R 2 = shy 015 N = 5 NS)
According to the different proportions of migratory directions propor-tions of migratory ight styles differed between species groups soaring-gliding was the main ight style of honey buzzards and black kites (50and 71 of total tracking time respectively Fig 3B) Flapping-gliding ight was extensively used by harriers (62) falcons (66) and ospreys(86) Proportion of apping-gliding ight was lower in marsh harriers(50) than in Montagursquos harriers (76) Raptors combining soaring with apping-gliding ight had lower climbing rates than birds in pure soaring-gliding ight indicating that apping might be a compensation for weakthermal conditions (122 ms N = 39 and 158 ms N = 77 respectivelyt = shy 221 p lt 005)
384 MEYER SPAAR amp BRUDERER
Fig 2 Distribution of track (line) and heading (shaded) directions for the different raptorgroups Corresponding distribution of wind directions at the levels of the tracked birds given
at the right
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 385
Fig 3 Proportion of different migratory routes (A) and of time spent in different ightstyles (B without reversed migrants) in the species groups For de nitions see methods Totaltracking time honey buzzard (512 min) black kite (174 min) harriers (369 min) falcons
(497 min) and osprey (107 min)
In uence of environmental conditions on migratory direction
In weak winds (ie wind speed below 4 ms) numbers of falcons harriersand ospreys crossing the sea or following the coast were similar for allwind directions (df = 3 x 2 = 28 NS and x 2 = 64 NS respectivelyTable 1) In strong winds sea crossings occurred in westerly northerlyand easterly winds (ie side and following winds) but not in southerly
386 MEYER SPAAR amp BRUDERER
TABLE 1 Comparison of wind situations and migratory routes
Crossing the sea Flying along the coast
weak wind strong wind weak wind strong wind
West 13 19 5 9North 16 40 9 15East 17 22 7 9South 9 0 1 1
x 2 28 397 64 116p 04 lt 0001 01 lt 001
Number of tracks given Wind direction West (225deg -315deg ) North (315deg -45deg ) East (45deg -135deg ) South (135deg -225deg ) Weak winds are below 4 ms Tracks of harriers falcons andospreys are included
winds (ie opposing winds df = 3 x 2 = 397 p lt 0001 Table 1)Strong northerly winds providing tailwind-support were preferred comparedto strong westerly and easterly winds (df = 2 x 2 = 96 p lt 001) In birdsfollowing the coast under strong winds numbers of observed birds showeda similar pattern as in birds crossing the sea they were higher in northerlywesterly and easterly compared to southerly winds (df = 3 x 2 = 116p lt 001 Table 1) The lower number of birds following the coast instrong southerly winds might be an artefact migratory birds were driftedtowards the mountain chains northerly of the radar station and were out ofthe tracking range of the radar
The tailwind-support achieved by the birds did only partly explain fordifferent migratory routes tailwind-component (t-c) of raptors crossingthe sea was higher than a theoretical t-c for the same birds assuming to y along the coast (270deg paired t-test N = 149 t = 78 p lt 0001Fig 4) On the other hand t-c of raptors ying along the coast was lowerthan the theoretical t-c if crossing the sea (210deg N = 138 t = shy 44p lt 0001) Thus many birds ew along the coast with a lower tailwind-support than if they would have crossed the sea There is an indicationthat good thermal conditions might stimulate birds to migrate along thecoast with a lower tailwind-support compared to sea crossing birds yingalong the coast instead of crossing the sea with a higher tailwind-supporthad higher climbing rates in thermal circling than birds ying along coastwith a higher tailwind-support compared to crossing the sea (mean plusmn SD
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 387
Fig 4 The observed tailwind-components in raptors crossing the sea (A left) and yingalong the coast (B left) versus the theoretical tailwind-componentsfor the alternativestrategyfor the same birds assumed to y along the coast (A right) and crossing the sea (B right)Paired t -test (A) N = 149 t = 78 p lt 0001 () (B) N = 138 t = shy 44 p lt 0001
18 plusmn 09 ms N = 29 and 10 plusmn 05 ms N = 17 respectively t-testt = 31 p lt 0005)
Flight altitudes
For all raptors the median of ight altitudes was below 1000 m above groundlevel (agl) and the 90 altitude limit was below 1800 m agl (Fig 5)Median altitudes differed signi cantly between the species groups (blackkite median 636 m N = 40 honey buzzard 862 m N = 81 harriers910 m N = 102 falcons 949 m N = 146 osprey 951 m N = 17 Kruskal-Wallis ANOVA H = 1835 df = 4 N = 386 p lt 0005) Harriers andfalcons had signi cantly higher ight altitudes than black kites (GT2-testp lt 001) Maximum observed ight levels were at about 1400 m agl inblack kites at 1800 m in ospreys at 2000 m in harriers at 2200 m in honeybuzzards and at 2600 m in falcons (Fig 5) No black kites were tracked in theearly afternoon when ight altitudes of soaring migrants are usually highpartly due to long operational breaks in the rst week of our observationperiod Thus maximum ight levels of black kites were probably higher
388 MEYER SPAAR amp BRUDERER
Fig 5 Flight altitudes above ground (mean per track) in the course of the day (0 =sunrise 100 = sunset) Median 636 m for black kite 862 m for honey buzzard 910 m
for harriers 949 m for falcons and 951 m for osprey
Flight altitudes of honey buzzards black kites and harriers increased fromsunrise towards noon and were more or less stable thereafter (Fig 5) Falconsachieved high ight altitudes already soon after sunrise by apping-gliding ight In falcons ight altitudes differed between migratory routes birdscrossing the sea had higher ight altitudes than birds ying across the bayand along the coast (Fig 6 Kruskal-Wallis ANOVA H = 181 df =2 p = 00001 GT2-test crossing the sea versus ying across the bay(p lt 001) and versus along the coast (p lt 005)) In honey buzzards andharriers differences in ight altitudes between migratory routes showed thesame tendency but did not differ statistically (Fig 6)
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 389
Fig 6 Flight altitudes above ground (mean per track) for different migratory routes inhoney buzzard harriers and falcons Boxes are medians with 25-75 quartiles whiskersshow 10-90 range dots are outliers (ie outside the 15 box length from the upper and
lower value) GT2-test p lt 005 p lt 001
Fig 7 Mean climbing rates in thermal circling in the course of the day (0 = sunrise100 = sunset) Climbing rate did not differ between species groups (ANOVA F = 116
df = 4 N = 116 NS)
Flight characteristics in soaring-gliding
Thermal circling phases were recorded in 116 of 386 tracks and averageclimbing rate was 146 plusmn 084 ms (N = 116) Climbing rates increasedduring the morning hours (Fig 7 time of day lt 50 Spearman rankcorrelation N = 61 r = 034 p lt 001) Strong thermal convection aroundnoon allowed climbing rates of up to 45 ms From noon to sunset climbing
390 MEYER SPAAR amp BRUDERER
TA
BL
E2
Flig
htch
arac
teri
stic
sof
diffe
rent
spec
ies
grou
ps(m
s)
hone
ybu
zzar
dbl
ack
kite
harr
iers
falc
ons
ospr
eyF
Soar
ing
glid
ing
Cli
mbi
ngra
te1
35plusmn
077
(48)
171
plusmn0
97(2
2)1
54plusmn
087
(21)
134
plusmn0
84(2
4)1
86(1
)1
16N
SC
ross
-cou
ntry
grou
ndsp
eed
86
plusmn0
4(2
8)7
6plusmn
07
(7)
93
plusmn0
9(9
)9
3plusmn
10
(6)
89
(1)
079
NS
Cro
ss-c
ount
ryai
rspe
ed10
4plusmn
42
(28)
99
plusmn3
6(7
)9
3plusmn
25
(9)
95
plusmn3
1(1
0)11
3(1
)0
27N
SIn
ter-
ther
mal
grou
ndsp
eed
116
plusmn3
2(4
7)10
8plusmn
25
(21)
121
plusmn2
5(2
1)11
2plusmn
32
(23)
114
(1)
075
NS
Inte
r-th
erm
alai
rspe
ed12
8plusmn
37
(47)
134
plusmn3
0(2
1)12
0plusmn
30
(21)
127
plusmn3
6(2
3)17
0(1
)0
57N
S
Hor
izon
tal
appi
ng-g
lidi
ngi
ght
Gro
unds
peed
138
plusmn2
5(1
6)10
9plusmn
26
(9)
116
plusmn1
9(6
1)12
2plusmn
36
(68)
146
plusmn1
9(1
0)2
47
Air
spee
d11
1plusmn
23
(16)
103
plusmn2
3(9
)11
2plusmn
18
(58)
113
plusmn2
4(6
8)13
2plusmn
30
(10)
387
Mea
nplusmn
SD
(N)
Dif
fere
nces
wer
ete
sted
byA
NO
VA
S
igni
ca
nce
leve
l
plt
005
plt
001
H
oriz
onta
l
appi
ng-g
lidi
ng
ight
was
incl
uded
only
ifve
rtic
alsp
eeds
did
note
xcee
d0
plusmn0
5m
san
dif
noth
erm
alci
rcli
ngw
asob
serv
ed
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 391
rates did not decrease signi cantly (Spearman rank correlation N = 55r = shy 015 NS) Climbing rates in thermal circling below 1 ms occurredduring the whole day They did not differ between species groups (Table 2ANOVA F = 116 df = 3 N = 115 NS) Flight altitudes increased withincreasing climbing rates in birds lsquo ying along the coastrsquo (r = 049 N = 64p lt 00001) Average circling duration in completed soaring phases was106 plusmn 67 s (N = 80) and it was not related to climbing rate (r = 002N = 80 NS)
Mean airspeed and groundspeed in inter-thermal gliding did not differbetween the species groups (Table 2 ANOVA F = 057 and F = 075respectively df = 3 N = 112 NS) and means per species ranged between120-134 ms and 108-121 ms respectively Raptors increased airspeedsin inter-thermal gliding with increasing climbing rates (r = 052 N =113 p lt 00001) indicating that they maximised cross-country speeds insoaring-gliding ight (Pennycuick 1989 Spaar amp Bruderer 1997a) Meancross-country speeds in soaring-gliding did not differ between species groups(ANOVA cross-country airspeed F = 079 df = 3 N = 54 NS range 93-113 ms cross-country groundspeed F = 027 df = 3 N = 54 NS range76-93 ms)
Flight characteristics in horizontal apping-gliding
This ight style was considered if vertical speeds did not exceed 0 plusmn 05 msand no thermal circling was observed within a track Air- and groundspeedsdiffered between species groups (ANOVA airspeed F = 247 df = 4N = 161 p lt 005 groundspeed F = 387 df = 4 N = 164p lt 0005 Table 2) Ospreys had higher groundspeeds than harriers (GT2-test p lt 005) and higher ground- and airspeeds than black kites (p lt 005)Mean airspeeds per species ranged between 103-132 ms correspondinggroundspeeds between 109-146 ms
Flapping-gliding migrants reached signi cantly higher cross-country air-speeds (113 plusmn 22 ms N = 161) and groundspeeds (122 plusmn 31 ms N =164) than birds in soaring-gliding ight (airspeed 100 plusmn 36 ms N = 55Welch-test t = 249 df = 68 p lt 001 groundspeed 87 plusmn 25 msN = 55 t = 846 df = 114 p lt 00005)
In horizontal apping-gliding ight reactions to different wind con-ditions depended on migratory route raptors ying along the coast de-
392 MEYER SPAAR amp BRUDERER
Fig 8 Airspeed in horizontal apping-gliding ight versus tailwind- and sidewind-components for different migratory routes Linear regression (A) excluding one value inbrackets y = shy 0256x + 1128 R 2 = 0215 N = 56 p lt 0001 (B) r = shy 010 N = 57NS (C) r = shy 007 N = 85 NS (D) excluding two values in brackets y = 0191x+ 1072
R2 = 005 N = 84 p lt 005
creased airspeed with increasing tailwind-component (Fig 8A) but air-speed was not related to sidewind-component (Fig 8B) In contrast in rap-tors crossing the sea no relation was found between airspeed and tailwind-component (Fig 8C) but airspeed increased with increasing sidewind-component (Fig 8D)
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 393
Discussion
There are only few studies on the ight behaviour of migrating raptors cross-ing the sea mainly carried out at concentration points such as narrow wa-ter straits or peninsulas (Evans amp Lathbury 1972 Bernis 1975 Kerlinger1984) and none by tracking radar As these studies were limited to visualobservation mainly high altitudinal migration was often not detected (Ker-linger et al 1985) Our approach is different because (i) we observed raptormigration at an average coast site (ii) we used a tracking radar and (iii) wemeasured wind conditions at all ight levels
Migratory routes and in uence of environmental conditions
Although black kites and in larger numbers honey buzzards undertakelong sea-crossings in the Central Mediterranean (Beaman amp Galea 1974Agostini et al 1994) in our study no black kites and few honey buzzardswere observed attempting the sea crossing The majority of harriers falconsand ospreys crossed the sea Sea crossing honey buzzards occurred towardsthe end of the migratory season and were probably juveniles as in honeybuzzards adults migrate two weeks earlier than juveniles (Kjelleacuten 1992Agostini et al 1997) In contrast to the adults which often migrate in large ocks juveniles mainly migrate singly and have no experience about themigratory ight route This may lead to a higher proportion of juvenilescrossing the sea
Wind and thermal conditions affected migratory directions many harri-ers falcons and ospreys crossed the sea in favourable following and sidewinds but no sea crossings were observed in strong opposing winds This isreasonable because energy and time consumption for the sea crossing andtherefore the risk of exhaustion drastically increase in opposing winds How-ever tailwind-support did not fully explain the decisions on migratory routesbecause regularly birds followed the coast with a lower tailwind-supportcompared to crossing the sea These birds following the coast reached highclimbing rates in thermal circling and this might be compensating for thelower tailwind-support Under good thermal conditions birds were able tomigrate in soaring-gliding ight which consumes much less energy com-pared to apping-gliding ight when crossing the sea (see below Energyand time consumption) Anyway birds in bad shape for example those with
394 MEYER SPAAR amp BRUDERER
low fat reserves should follow the coast in order to minimise the risk ofstarvation to feed when ever it is possible and to y in soaring-gliding ight
At the same observation site migratory directions of nocturnal migrants ying below 2000 m above ground shift from 190deg to 250deg within the nightthe majority of the birds cross the sea when approaching the coast in the rsthalf of the night whereas they hesitate later on (Bruderer amp Liechti 1998)Thus the motivation to cross large barriers seems to decrease after somehours of migratory ight
Energy and time consumption
Energy consumption in apping ight disproportionately increases withbody mass while in soaring-gliding ight it is 2-3 times the basal metabolicrate (BMR) only (for herring gull Larus argentatus Baudinette amp Schmidt-Nielson 1974 Lustick et al 1978) In honey buzzards the average observedairspeed in apping-gliding ight was 111 ms and fat consumption perdistance (gkm) for an average honey buzzard (body mass in Table 3) atthis speed is about 6 times higher than in soaring-gliding ight (calculatedafter Pennycuick (1989 version 11) energy consumption in soaring-gliding ight = 2 times BMR body drag coef cient = 04) If the body dragcoef cient is set to 01 as recently proposed by Pennycuick (1997) energyconsumption in apping ight per km is reduced and energetic differencebetween soaring-gliding and apping ight become smaller but calculatedmaximum range speeds in apping ight are very high compared to ourobservations
We calculated energy and time consumption of different migratory routesbased on observed ight speeds for an average honey buzzard (Table 3) Notethat we did not take into account the time needed for refuelling consumedenergy (ie burned fat) and that we assumed zero wind Crossing the sea in apping-gliding ight (150 km distance) took on average about 4 hours andit was the time-minimising strategy for a honey buzzard (Table 3) Flyingalong the coast (250 km) required about 7 hours in soaring-gliding ightand 6 hours in apping-gliding ight Flying along the coast by soaring-gliding is the energy-minimising strategy for a honey buzzard and consumesabout 4 g fat (Table 3) Flapping-gliding ight along the coast is energeticallythe most expensive but never the fastest strategy Thus black kites andhoney buzzards migrated predominantly according to the energy minimising
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 395
TABLE 3 Observed airspeeds and corresponding estimated ight durationand fat consumption for an average honey buzzard when ying along the
coast or across the sea in different ight styles
Flight style Soaring-gliding Horizontal Horizontal apping-gliding apping-gliding
Flight route Flying along the coast Flying along the coast Crossing the seaDistance 250 km 250 km 150 km
Airspeed ms 101 (81-123) 108 (88-133) 108 (88-133)Flight duration hmin 652 (538-834) 626 (513-753) 351 (308-444)Fat consumption g 41 (34-55) 266 (230-298) 158 (138-179)for ight
Average honey buzzard = body mass 08 kg wing span 13 m wing area 025 m2 Medianand 25-75 range are given in brackets Calculations on fat consumption are based onPennycuickrsquos (1989) program of bird ight performance version 11 and birds are assumedto maximize cross-country speed in soaring-gliding ight (Spaar amp Bruderer 1997a)
strategy because a large proportion of the birds followed the coast in soaring-gliding ight
For species such as falcons or small harriers the difference in energyconsumption between apping and soaring-gliding ight is smaller Asthese species regularly use apping-gliding ight also over land or combinesoaring with apping-gliding ight differences in energy consumptionbetween land and sea route become even smaller and only short detours areenergetically favourable By combining soaring and apping-gliding ightsmaller raptors such as falcons or harriers are able to y under varyingenvironmental conditions to extend their migratory activity to times whenthermal updrafts are absent and to migrate even at night (Stark amp Liechti1993 Spaar amp Bruderer 1997b Spaar et al 1998)
Flight altitudes
Flight altitudes differed between the species groups partly due to differencesin migratory routes Birds crossing the sea ew considerably higher thanthose following the coast Kerlinger (1984) made similar observations atthe Delaware Bay birds crossing the bay ew at higher altitudes thanthose keeping on land He explained the higher ight altitudes by the bettervisibility of the other shore However the African coast is not visible fromour study site Some falcons even started crossing the sea after sunset
396 MEYER SPAAR amp BRUDERER
indicating that they continued migration into the night these birds had to y at least three hours until they reached the African coast The higher ightaltitudes of birds crossing the sea at our study site was mainly caused bytopographical features birds ying with southerly directions were forced toclimb higher up when crossing the mountains chains (1500 m height) about10-15 km northward of the coast and they afterwards maintained these ightlevels when approaching the sea
Flight altitudes of birds migrating along the coast were similar to altitudesobserved in North America and southern Israel with most raptors yingbelow 1000 m above ground and lt 5 above 1500 m (Kerlinger 19841989 Bruderer et al 1994 Spaar 1996)
Flight characteristics in soaring-gliding
Flight characteristics in soaring-gliding ight did not differ between thespecies groups (Table 2) As shown for raptor migration in Israel climb-ing rates do not depend primarily on morphological features but on environ-mental conditions (Spaar 1997) In general mean climbing rates in southernSpain were lower than in the semi-arid desert in Israel (Spaar 1997) Meanduration of the completed circling phases was shorter in Israel (88 plusmn 52 sSpaar 1995 1999) than in Spain In contrast to Israel (Spaar 1995 1997)circling time did not increase with increasing climbing rate Compared tothe semi-desert the development of thermal convection in southern Spainwas probably less stable and birds circled as long as possible in updraftsbecause they could not rely to encounter a next strong thermal within glid-ing distance This might also explain why inter-thermal gliding was oftensupported by apping it reduces sinking rate and prolongs gliding distance
In raptors migrating along the coast ight altitude and gliding airspeedswere related to thermal strength raptors circling in strong thermals hadhigher ight altitudes and higher inter-thermal gliding airspeeds than thosein weak thermals This is consistent with optimal ight theory for birds max-imising cross-country speed in soaring-gliding ight (Pennycuick 1989) asdemonstrated for steppe buzzards Buteo buteo vulpinus (Spaar amp Bruderer1997a)
Flight characteristics in horizontal apping-gliding
According to optimal ight theory maximum range speed decreases withfollowing winds and increases with head- and sidewinds (Pennycuick 1975
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 397
Liechti et al 1994) In raptors ying along the coast we observed decreas-ing airspeeds with increasing tailwind-support whereas birds crossing thesea did not reduce airspeeds Why did the birds react differently Sea cross-ings imply higher risks there is no possibility to land and weather (wind)conditions may change rapidly Higher airspeeds in following winds lead toshorter crossing times and lower the risk of unpredictable weather changesFurthermore drift by sidewinds may considerably extent crossing distancesand in order to reduce drift raptors increased airspeeds with increasingsindwind-components In contrast to sharp-shinned hawks Accipiter stria-tus avoiding crossings in strong lateral winds (Kerlinger 1985) we observedmany raptors to cross the sea in strong north-westerly (ie lateral) winds ifthey provided at least partial tailwind-support
Conclusions
Our observations at an average site at the southern coast of Spain showthat differences in water crossing behaviour are mainly species-speci cpresumably due to the costs for different ight strategies additionallyindividuals reacted to environmental factors such as wind and thermalconditions In medium sized raptors such as honey buzzard soaring-gliding ight along the coast is energy-minimising whereas crossing the sea istime-minimising Sea crossings occurred preferably with a good tailwind-support but also under strong sidewinds Raptors crossing the sea did notreduce airspeeds in following winds but increased airspeeds with increasingsidewinds They seemed to minimise the water crossing duration and theeffect of drift by sidewinds Under good thermal conditions birds regularlyfollowed the coast despite a lower tailwind-support compared to crossing thesea
References
Agostini N amp Logozzo D (1995) Autumn migration of honey buzzards in Southern Italymdash J Raptor Res 29 p 275-277
mdash mdash Logozzo D amp Colero C (1997) Migration of ocks of honey buzzards in SouthernItaly and Malta mdash J Raptor Res 31 p 84-86
mdash mdash Malara G Neri F Mollicone D amp Melotto S (1994) Flight strategies of honeybuzzards during spring migration across the Central Mediterranean mdash Avocetta 18p 73-76
398 MEYER SPAAR amp BRUDERER
Bannermann D-A amp Bannermann WM (1971) Handbook of the birds of Cyprus andmigrants of the Middle East mdash Oliver amp Boyd Edinburgh
Batschelet E (1981) Circular statistics in biology mdash Academic Press LondonBaudinette RV amp Schmidt-Nielsen K (1974) Energy costs of gliding ight in herring
gulls mdash Nature 248 p 83-84Beaman M amp Galea C (1974) The visible migration of raptors over the Maltese Islands
mdash Ibis 116 p 419-431Bernis F (1975) Migracion de Falconiformes y Ciconea spp por Gibraltar mdash IV Avance
sobre recuentos en el verano-otontildeo 1974 mdash Ardeola 21 p 581-592Bijlsma RG (1987) Bottleneck areas for migratory birds in the Mediterranean region mdash
ICBP Study report No 18 CambridgeBruderer B amp Liechti F (1998) Flight behaviour of nocturnally migrating birds in coastal
areas mdash crossing or coasting mdash J Avian Biol 29 p 499-510mdash mdash Blitzblau S amp Peter D (1994) Migration and ight behaviour of honey buzzards
Pernis apivorus in southern Israel observed by radar mdash Ardea 82 p 111-122mdash mdash Steuri T amp Baumgartner M (1995) Short-range high-precision surveillance of
nocturnal migration and tracking of single targets mdash Israel J Zool 41 p 207-220Evans PR amp Lathbury GW (1972) Raptor migration across the Strait of Gibraltar mdash Ibis
115 p 572-585Galea C amp Massa B (1985) Notes on the raptor migration across the central Mediter-
ranean mdash In Conservation studies on raptors (I Newton amp RD Chancellor eds)ICBP Technical Publication No 5 p 257-261
Gensboslashl B (1997) Greifvoumlgel mdash BLV VerlagsgesellschaftMuumlnchenGustin M amp Pizzari T (1998) Migratory pattern in the genus Circus sex and age
differential migration in Italy mdash Ornis Svecica 8 p 23-26Kerlinger P (1984) Flight behaviour of sharp-shinned hawks during migration 2 Over
water mdash Anim Beh 32 p 1029-1034mdash mdash (1985) Water-crossing behaviour of raptors during migration mdash Wilson Bull 97
p 109-113mdash mdash (1989) Flight strategies of migrating hawks mdash University of Chicago Press Chicagomdash mdash Bingman VP amp Able KP (1985) Comparative ight behaviour of migrating hawks
studied with tracking radar during autumn in central New York mdash Can J Zool 63p 755-761
Kjelleacuten N (1992) Differential timing of autumn migration between sex and age groups inraptors at Falsterbo Sweden mdash Orn Scan 23 p 420-434
Liechti F (1993) Naumlchtlicher Vogelzug im Herbst uumlber Suumlddeutschland Winddrift undKompensation mdash J Ornithol 134 p 373-404
mdash mdash Hedenstroumlm A amp Alerstam T (1994) Effects of sidewinds on optimal ight speedsof birds mdash J theor Biol 170 p 219-225
Lustick S Batterby B amp Kelty M (1978) Behavioural thermoregulation orientationtoward the sun in herring gulls mdash Sience 200 p 81-83
Pennycuick C (1975) Mechanics of ight mdash In Avian biology Vol 5 (DS Farner ampRJ King eds) Academic Press New York p 1-75
mdash mdash (1989) Bird ight performance a practical calculation manual mdash Oxford UniversityPress Oxford
mdash mdash (1997) Actual and lsquooptimumrsquo ight speeds eld data reassessedmdash J Exp Biol 200p 2355-2361
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 399
Richardson WJ (1978) Timing and amount of bird migration in relation to weather areview mdash Oikos 30 p 224-272
Spaar R (1995) Flight behaviour of steppe buzzard (Buteo buteo vulpinus) during springmigration in southern Israel a tracking radar study mdash Israel J Zool 41 p 489-500
mdash mdash (1996) Flight behaviour of migrating raptors in southern Israel mdash PhD thesisUniversity of Basel Verlag Schweizerische Vogelwarte Sempach
mdash mdash (1997) Flight strategies of migrating raptors a comparative study of interspecicvariation in ight characteristicsmdash Ibis 139 p 523-535
mdash mdash (1999) Flight behaviour of migrating raptors under varying environmental conditionsmdash In Proc 22 Int Ornithol Congr (N Adams amp R Slotow eds) University of NatalDurban p 1844-1862
mdash mdash amp Bruderer B (1997a) Optimal ight behaviour of soaring migrants A case studyof migrating steppe buzzards Buteo buteo vulpinus mdash Behav Ecol 8 p 288-297)
mdash mdash amp Bruderer B (1997b) Migration by apping or soaring ight strategies of marshMontagursquos and pallid harriers in southern Israel mdash Condor 99 p 458-469
mdash mdash Stark H amp Liechti F (1998) Migratory ight strategies of Levant sparrowhawkstime or energy minimization mdash Anim Behav 56 p 1185-1197
Stark H amp Liechti F (1993) Do Levant sparrowhawks Accipiter brevipes also migrate atnight mdash Ibis 135 p 233-236
Thiollay J-M (1977) Importance des populations de rapaces migrateurs en mediterraneeoccidentale mdash Alauda 45 p 115-121

MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 383
sidewind-componentare the wind speeds in track direction and rectangular to track directionrespectivelyTo analyse seasonal trends in the proportion of sea crossing 5-day periods withat least two ight paths of a species group were included in the analysis
Results
Migratory directions and ight styles
Track directions of all raptors were south-westerly (238deg plusmn 34deg mean plusmn SD)and coincided with the direction towards the visible tip of the coastline nearFuengirola corresponding headings were 246deg plusmn 36deg (Fig 1) Tracks andheadings differed signi cantly between the species groups (Watson-Williamsmultisample test tracks F = 1884 df = 4 N = 382 p lt 00001headings F = 2148 df = 4 N = 382 p lt 00001) Black kites(track 269deg heading 276deg ) and honey buzzards (252deg 263deg ) kept westerly ight directions mainly following the east-west leading coastline whereasharriers (229deg 231deg ) falcons (228deg 240deg ) and ospreys (225deg 221deg ) hadsouth-westerly ight directions (Fig 2) Thus regarding the water-crossingbehaviour two main groups occurred no black kites and only few honeybuzzards (17 of tracks) but large proportions of harriers (65) falcons(57) and ospreys (73) crossed the sea (Fig 3A) The proportion of birds ying across the bay was below 15 of tracks in all groups For all groupswind directions were predominantly from north-west or from east (Fig 2)
The proportion of sea crossings (5-day periods) increased towards theend of the migratory season in honey buzzards and falcons (honey buzzardR 2 = 082 N = 6 p lt 001 falcons R 2 = 068 N = 12 p lt 001) Noseasonal trend was found in marsh and Montagursquos harriers (marsh harrierR 2 = shy 025 N = 5 NS Montagursquos harrier R 2 = shy 015 N = 5 NS)
According to the different proportions of migratory directions propor-tions of migratory ight styles differed between species groups soaring-gliding was the main ight style of honey buzzards and black kites (50and 71 of total tracking time respectively Fig 3B) Flapping-gliding ight was extensively used by harriers (62) falcons (66) and ospreys(86) Proportion of apping-gliding ight was lower in marsh harriers(50) than in Montagursquos harriers (76) Raptors combining soaring with apping-gliding ight had lower climbing rates than birds in pure soaring-gliding ight indicating that apping might be a compensation for weakthermal conditions (122 ms N = 39 and 158 ms N = 77 respectivelyt = shy 221 p lt 005)
384 MEYER SPAAR amp BRUDERER
Fig 2 Distribution of track (line) and heading (shaded) directions for the different raptorgroups Corresponding distribution of wind directions at the levels of the tracked birds given
at the right
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 385
Fig 3 Proportion of different migratory routes (A) and of time spent in different ightstyles (B without reversed migrants) in the species groups For de nitions see methods Totaltracking time honey buzzard (512 min) black kite (174 min) harriers (369 min) falcons
(497 min) and osprey (107 min)
In uence of environmental conditions on migratory direction
In weak winds (ie wind speed below 4 ms) numbers of falcons harriersand ospreys crossing the sea or following the coast were similar for allwind directions (df = 3 x 2 = 28 NS and x 2 = 64 NS respectivelyTable 1) In strong winds sea crossings occurred in westerly northerlyand easterly winds (ie side and following winds) but not in southerly
386 MEYER SPAAR amp BRUDERER
TABLE 1 Comparison of wind situations and migratory routes
Crossing the sea Flying along the coast
weak wind strong wind weak wind strong wind
West 13 19 5 9North 16 40 9 15East 17 22 7 9South 9 0 1 1
x 2 28 397 64 116p 04 lt 0001 01 lt 001
Number of tracks given Wind direction West (225deg -315deg ) North (315deg -45deg ) East (45deg -135deg ) South (135deg -225deg ) Weak winds are below 4 ms Tracks of harriers falcons andospreys are included
winds (ie opposing winds df = 3 x 2 = 397 p lt 0001 Table 1)Strong northerly winds providing tailwind-support were preferred comparedto strong westerly and easterly winds (df = 2 x 2 = 96 p lt 001) In birdsfollowing the coast under strong winds numbers of observed birds showeda similar pattern as in birds crossing the sea they were higher in northerlywesterly and easterly compared to southerly winds (df = 3 x 2 = 116p lt 001 Table 1) The lower number of birds following the coast instrong southerly winds might be an artefact migratory birds were driftedtowards the mountain chains northerly of the radar station and were out ofthe tracking range of the radar
The tailwind-support achieved by the birds did only partly explain fordifferent migratory routes tailwind-component (t-c) of raptors crossingthe sea was higher than a theoretical t-c for the same birds assuming to y along the coast (270deg paired t-test N = 149 t = 78 p lt 0001Fig 4) On the other hand t-c of raptors ying along the coast was lowerthan the theoretical t-c if crossing the sea (210deg N = 138 t = shy 44p lt 0001) Thus many birds ew along the coast with a lower tailwind-support than if they would have crossed the sea There is an indicationthat good thermal conditions might stimulate birds to migrate along thecoast with a lower tailwind-support compared to sea crossing birds yingalong the coast instead of crossing the sea with a higher tailwind-supporthad higher climbing rates in thermal circling than birds ying along coastwith a higher tailwind-support compared to crossing the sea (mean plusmn SD
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 387
Fig 4 The observed tailwind-components in raptors crossing the sea (A left) and yingalong the coast (B left) versus the theoretical tailwind-componentsfor the alternativestrategyfor the same birds assumed to y along the coast (A right) and crossing the sea (B right)Paired t -test (A) N = 149 t = 78 p lt 0001 () (B) N = 138 t = shy 44 p lt 0001
18 plusmn 09 ms N = 29 and 10 plusmn 05 ms N = 17 respectively t-testt = 31 p lt 0005)
Flight altitudes
For all raptors the median of ight altitudes was below 1000 m above groundlevel (agl) and the 90 altitude limit was below 1800 m agl (Fig 5)Median altitudes differed signi cantly between the species groups (blackkite median 636 m N = 40 honey buzzard 862 m N = 81 harriers910 m N = 102 falcons 949 m N = 146 osprey 951 m N = 17 Kruskal-Wallis ANOVA H = 1835 df = 4 N = 386 p lt 0005) Harriers andfalcons had signi cantly higher ight altitudes than black kites (GT2-testp lt 001) Maximum observed ight levels were at about 1400 m agl inblack kites at 1800 m in ospreys at 2000 m in harriers at 2200 m in honeybuzzards and at 2600 m in falcons (Fig 5) No black kites were tracked in theearly afternoon when ight altitudes of soaring migrants are usually highpartly due to long operational breaks in the rst week of our observationperiod Thus maximum ight levels of black kites were probably higher
388 MEYER SPAAR amp BRUDERER
Fig 5 Flight altitudes above ground (mean per track) in the course of the day (0 =sunrise 100 = sunset) Median 636 m for black kite 862 m for honey buzzard 910 m
for harriers 949 m for falcons and 951 m for osprey
Flight altitudes of honey buzzards black kites and harriers increased fromsunrise towards noon and were more or less stable thereafter (Fig 5) Falconsachieved high ight altitudes already soon after sunrise by apping-gliding ight In falcons ight altitudes differed between migratory routes birdscrossing the sea had higher ight altitudes than birds ying across the bayand along the coast (Fig 6 Kruskal-Wallis ANOVA H = 181 df =2 p = 00001 GT2-test crossing the sea versus ying across the bay(p lt 001) and versus along the coast (p lt 005)) In honey buzzards andharriers differences in ight altitudes between migratory routes showed thesame tendency but did not differ statistically (Fig 6)
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 389
Fig 6 Flight altitudes above ground (mean per track) for different migratory routes inhoney buzzard harriers and falcons Boxes are medians with 25-75 quartiles whiskersshow 10-90 range dots are outliers (ie outside the 15 box length from the upper and
lower value) GT2-test p lt 005 p lt 001
Fig 7 Mean climbing rates in thermal circling in the course of the day (0 = sunrise100 = sunset) Climbing rate did not differ between species groups (ANOVA F = 116
df = 4 N = 116 NS)
Flight characteristics in soaring-gliding
Thermal circling phases were recorded in 116 of 386 tracks and averageclimbing rate was 146 plusmn 084 ms (N = 116) Climbing rates increasedduring the morning hours (Fig 7 time of day lt 50 Spearman rankcorrelation N = 61 r = 034 p lt 001) Strong thermal convection aroundnoon allowed climbing rates of up to 45 ms From noon to sunset climbing
390 MEYER SPAAR amp BRUDERER
TA
BL
E2
Flig
htch
arac
teri
stic
sof
diffe
rent
spec
ies
grou
ps(m
s)
hone
ybu
zzar
dbl
ack
kite
harr
iers
falc
ons
ospr
eyF
Soar
ing
glid
ing
Cli
mbi
ngra
te1
35plusmn
077
(48)
171
plusmn0
97(2
2)1
54plusmn
087
(21)
134
plusmn0
84(2
4)1
86(1
)1
16N
SC
ross
-cou
ntry
grou
ndsp
eed
86
plusmn0
4(2
8)7
6plusmn
07
(7)
93
plusmn0
9(9
)9
3plusmn
10
(6)
89
(1)
079
NS
Cro
ss-c
ount
ryai
rspe
ed10
4plusmn
42
(28)
99
plusmn3
6(7
)9
3plusmn
25
(9)
95
plusmn3
1(1
0)11
3(1
)0
27N
SIn
ter-
ther
mal
grou
ndsp
eed
116
plusmn3
2(4
7)10
8plusmn
25
(21)
121
plusmn2
5(2
1)11
2plusmn
32
(23)
114
(1)
075
NS
Inte
r-th
erm
alai
rspe
ed12
8plusmn
37
(47)
134
plusmn3
0(2
1)12
0plusmn
30
(21)
127
plusmn3
6(2
3)17
0(1
)0
57N
S
Hor
izon
tal
appi
ng-g
lidi
ngi
ght
Gro
unds
peed
138
plusmn2
5(1
6)10
9plusmn
26
(9)
116
plusmn1
9(6
1)12
2plusmn
36
(68)
146
plusmn1
9(1
0)2
47
Air
spee
d11
1plusmn
23
(16)
103
plusmn2
3(9
)11
2plusmn
18
(58)
113
plusmn2
4(6
8)13
2plusmn
30
(10)
387
Mea
nplusmn
SD
(N)
Dif
fere
nces
wer
ete
sted
byA
NO
VA
S
igni
ca
nce
leve
l
plt
005
plt
001
H
oriz
onta
l
appi
ng-g
lidi
ng
ight
was
incl
uded
only
ifve
rtic
alsp
eeds
did
note
xcee
d0
plusmn0
5m
san
dif
noth
erm
alci
rcli
ngw
asob
serv
ed
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 391
rates did not decrease signi cantly (Spearman rank correlation N = 55r = shy 015 NS) Climbing rates in thermal circling below 1 ms occurredduring the whole day They did not differ between species groups (Table 2ANOVA F = 116 df = 3 N = 115 NS) Flight altitudes increased withincreasing climbing rates in birds lsquo ying along the coastrsquo (r = 049 N = 64p lt 00001) Average circling duration in completed soaring phases was106 plusmn 67 s (N = 80) and it was not related to climbing rate (r = 002N = 80 NS)
Mean airspeed and groundspeed in inter-thermal gliding did not differbetween the species groups (Table 2 ANOVA F = 057 and F = 075respectively df = 3 N = 112 NS) and means per species ranged between120-134 ms and 108-121 ms respectively Raptors increased airspeedsin inter-thermal gliding with increasing climbing rates (r = 052 N =113 p lt 00001) indicating that they maximised cross-country speeds insoaring-gliding ight (Pennycuick 1989 Spaar amp Bruderer 1997a) Meancross-country speeds in soaring-gliding did not differ between species groups(ANOVA cross-country airspeed F = 079 df = 3 N = 54 NS range 93-113 ms cross-country groundspeed F = 027 df = 3 N = 54 NS range76-93 ms)
Flight characteristics in horizontal apping-gliding
This ight style was considered if vertical speeds did not exceed 0 plusmn 05 msand no thermal circling was observed within a track Air- and groundspeedsdiffered between species groups (ANOVA airspeed F = 247 df = 4N = 161 p lt 005 groundspeed F = 387 df = 4 N = 164p lt 0005 Table 2) Ospreys had higher groundspeeds than harriers (GT2-test p lt 005) and higher ground- and airspeeds than black kites (p lt 005)Mean airspeeds per species ranged between 103-132 ms correspondinggroundspeeds between 109-146 ms
Flapping-gliding migrants reached signi cantly higher cross-country air-speeds (113 plusmn 22 ms N = 161) and groundspeeds (122 plusmn 31 ms N =164) than birds in soaring-gliding ight (airspeed 100 plusmn 36 ms N = 55Welch-test t = 249 df = 68 p lt 001 groundspeed 87 plusmn 25 msN = 55 t = 846 df = 114 p lt 00005)
In horizontal apping-gliding ight reactions to different wind con-ditions depended on migratory route raptors ying along the coast de-
392 MEYER SPAAR amp BRUDERER
Fig 8 Airspeed in horizontal apping-gliding ight versus tailwind- and sidewind-components for different migratory routes Linear regression (A) excluding one value inbrackets y = shy 0256x + 1128 R 2 = 0215 N = 56 p lt 0001 (B) r = shy 010 N = 57NS (C) r = shy 007 N = 85 NS (D) excluding two values in brackets y = 0191x+ 1072
R2 = 005 N = 84 p lt 005
creased airspeed with increasing tailwind-component (Fig 8A) but air-speed was not related to sidewind-component (Fig 8B) In contrast in rap-tors crossing the sea no relation was found between airspeed and tailwind-component (Fig 8C) but airspeed increased with increasing sidewind-component (Fig 8D)
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 393
Discussion
There are only few studies on the ight behaviour of migrating raptors cross-ing the sea mainly carried out at concentration points such as narrow wa-ter straits or peninsulas (Evans amp Lathbury 1972 Bernis 1975 Kerlinger1984) and none by tracking radar As these studies were limited to visualobservation mainly high altitudinal migration was often not detected (Ker-linger et al 1985) Our approach is different because (i) we observed raptormigration at an average coast site (ii) we used a tracking radar and (iii) wemeasured wind conditions at all ight levels
Migratory routes and in uence of environmental conditions
Although black kites and in larger numbers honey buzzards undertakelong sea-crossings in the Central Mediterranean (Beaman amp Galea 1974Agostini et al 1994) in our study no black kites and few honey buzzardswere observed attempting the sea crossing The majority of harriers falconsand ospreys crossed the sea Sea crossing honey buzzards occurred towardsthe end of the migratory season and were probably juveniles as in honeybuzzards adults migrate two weeks earlier than juveniles (Kjelleacuten 1992Agostini et al 1997) In contrast to the adults which often migrate in large ocks juveniles mainly migrate singly and have no experience about themigratory ight route This may lead to a higher proportion of juvenilescrossing the sea
Wind and thermal conditions affected migratory directions many harri-ers falcons and ospreys crossed the sea in favourable following and sidewinds but no sea crossings were observed in strong opposing winds This isreasonable because energy and time consumption for the sea crossing andtherefore the risk of exhaustion drastically increase in opposing winds How-ever tailwind-support did not fully explain the decisions on migratory routesbecause regularly birds followed the coast with a lower tailwind-supportcompared to crossing the sea These birds following the coast reached highclimbing rates in thermal circling and this might be compensating for thelower tailwind-support Under good thermal conditions birds were able tomigrate in soaring-gliding ight which consumes much less energy com-pared to apping-gliding ight when crossing the sea (see below Energyand time consumption) Anyway birds in bad shape for example those with
394 MEYER SPAAR amp BRUDERER
low fat reserves should follow the coast in order to minimise the risk ofstarvation to feed when ever it is possible and to y in soaring-gliding ight
At the same observation site migratory directions of nocturnal migrants ying below 2000 m above ground shift from 190deg to 250deg within the nightthe majority of the birds cross the sea when approaching the coast in the rsthalf of the night whereas they hesitate later on (Bruderer amp Liechti 1998)Thus the motivation to cross large barriers seems to decrease after somehours of migratory ight
Energy and time consumption
Energy consumption in apping ight disproportionately increases withbody mass while in soaring-gliding ight it is 2-3 times the basal metabolicrate (BMR) only (for herring gull Larus argentatus Baudinette amp Schmidt-Nielson 1974 Lustick et al 1978) In honey buzzards the average observedairspeed in apping-gliding ight was 111 ms and fat consumption perdistance (gkm) for an average honey buzzard (body mass in Table 3) atthis speed is about 6 times higher than in soaring-gliding ight (calculatedafter Pennycuick (1989 version 11) energy consumption in soaring-gliding ight = 2 times BMR body drag coef cient = 04) If the body dragcoef cient is set to 01 as recently proposed by Pennycuick (1997) energyconsumption in apping ight per km is reduced and energetic differencebetween soaring-gliding and apping ight become smaller but calculatedmaximum range speeds in apping ight are very high compared to ourobservations
We calculated energy and time consumption of different migratory routesbased on observed ight speeds for an average honey buzzard (Table 3) Notethat we did not take into account the time needed for refuelling consumedenergy (ie burned fat) and that we assumed zero wind Crossing the sea in apping-gliding ight (150 km distance) took on average about 4 hours andit was the time-minimising strategy for a honey buzzard (Table 3) Flyingalong the coast (250 km) required about 7 hours in soaring-gliding ightand 6 hours in apping-gliding ight Flying along the coast by soaring-gliding is the energy-minimising strategy for a honey buzzard and consumesabout 4 g fat (Table 3) Flapping-gliding ight along the coast is energeticallythe most expensive but never the fastest strategy Thus black kites andhoney buzzards migrated predominantly according to the energy minimising
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 395
TABLE 3 Observed airspeeds and corresponding estimated ight durationand fat consumption for an average honey buzzard when ying along the
coast or across the sea in different ight styles
Flight style Soaring-gliding Horizontal Horizontal apping-gliding apping-gliding
Flight route Flying along the coast Flying along the coast Crossing the seaDistance 250 km 250 km 150 km
Airspeed ms 101 (81-123) 108 (88-133) 108 (88-133)Flight duration hmin 652 (538-834) 626 (513-753) 351 (308-444)Fat consumption g 41 (34-55) 266 (230-298) 158 (138-179)for ight
Average honey buzzard = body mass 08 kg wing span 13 m wing area 025 m2 Medianand 25-75 range are given in brackets Calculations on fat consumption are based onPennycuickrsquos (1989) program of bird ight performance version 11 and birds are assumedto maximize cross-country speed in soaring-gliding ight (Spaar amp Bruderer 1997a)
strategy because a large proportion of the birds followed the coast in soaring-gliding ight
For species such as falcons or small harriers the difference in energyconsumption between apping and soaring-gliding ight is smaller Asthese species regularly use apping-gliding ight also over land or combinesoaring with apping-gliding ight differences in energy consumptionbetween land and sea route become even smaller and only short detours areenergetically favourable By combining soaring and apping-gliding ightsmaller raptors such as falcons or harriers are able to y under varyingenvironmental conditions to extend their migratory activity to times whenthermal updrafts are absent and to migrate even at night (Stark amp Liechti1993 Spaar amp Bruderer 1997b Spaar et al 1998)
Flight altitudes
Flight altitudes differed between the species groups partly due to differencesin migratory routes Birds crossing the sea ew considerably higher thanthose following the coast Kerlinger (1984) made similar observations atthe Delaware Bay birds crossing the bay ew at higher altitudes thanthose keeping on land He explained the higher ight altitudes by the bettervisibility of the other shore However the African coast is not visible fromour study site Some falcons even started crossing the sea after sunset
396 MEYER SPAAR amp BRUDERER
indicating that they continued migration into the night these birds had to y at least three hours until they reached the African coast The higher ightaltitudes of birds crossing the sea at our study site was mainly caused bytopographical features birds ying with southerly directions were forced toclimb higher up when crossing the mountains chains (1500 m height) about10-15 km northward of the coast and they afterwards maintained these ightlevels when approaching the sea
Flight altitudes of birds migrating along the coast were similar to altitudesobserved in North America and southern Israel with most raptors yingbelow 1000 m above ground and lt 5 above 1500 m (Kerlinger 19841989 Bruderer et al 1994 Spaar 1996)
Flight characteristics in soaring-gliding
Flight characteristics in soaring-gliding ight did not differ between thespecies groups (Table 2) As shown for raptor migration in Israel climb-ing rates do not depend primarily on morphological features but on environ-mental conditions (Spaar 1997) In general mean climbing rates in southernSpain were lower than in the semi-arid desert in Israel (Spaar 1997) Meanduration of the completed circling phases was shorter in Israel (88 plusmn 52 sSpaar 1995 1999) than in Spain In contrast to Israel (Spaar 1995 1997)circling time did not increase with increasing climbing rate Compared tothe semi-desert the development of thermal convection in southern Spainwas probably less stable and birds circled as long as possible in updraftsbecause they could not rely to encounter a next strong thermal within glid-ing distance This might also explain why inter-thermal gliding was oftensupported by apping it reduces sinking rate and prolongs gliding distance
In raptors migrating along the coast ight altitude and gliding airspeedswere related to thermal strength raptors circling in strong thermals hadhigher ight altitudes and higher inter-thermal gliding airspeeds than thosein weak thermals This is consistent with optimal ight theory for birds max-imising cross-country speed in soaring-gliding ight (Pennycuick 1989) asdemonstrated for steppe buzzards Buteo buteo vulpinus (Spaar amp Bruderer1997a)
Flight characteristics in horizontal apping-gliding
According to optimal ight theory maximum range speed decreases withfollowing winds and increases with head- and sidewinds (Pennycuick 1975
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 397
Liechti et al 1994) In raptors ying along the coast we observed decreas-ing airspeeds with increasing tailwind-support whereas birds crossing thesea did not reduce airspeeds Why did the birds react differently Sea cross-ings imply higher risks there is no possibility to land and weather (wind)conditions may change rapidly Higher airspeeds in following winds lead toshorter crossing times and lower the risk of unpredictable weather changesFurthermore drift by sidewinds may considerably extent crossing distancesand in order to reduce drift raptors increased airspeeds with increasingsindwind-components In contrast to sharp-shinned hawks Accipiter stria-tus avoiding crossings in strong lateral winds (Kerlinger 1985) we observedmany raptors to cross the sea in strong north-westerly (ie lateral) winds ifthey provided at least partial tailwind-support
Conclusions
Our observations at an average site at the southern coast of Spain showthat differences in water crossing behaviour are mainly species-speci cpresumably due to the costs for different ight strategies additionallyindividuals reacted to environmental factors such as wind and thermalconditions In medium sized raptors such as honey buzzard soaring-gliding ight along the coast is energy-minimising whereas crossing the sea istime-minimising Sea crossings occurred preferably with a good tailwind-support but also under strong sidewinds Raptors crossing the sea did notreduce airspeeds in following winds but increased airspeeds with increasingsidewinds They seemed to minimise the water crossing duration and theeffect of drift by sidewinds Under good thermal conditions birds regularlyfollowed the coast despite a lower tailwind-support compared to crossing thesea
References
Agostini N amp Logozzo D (1995) Autumn migration of honey buzzards in Southern Italymdash J Raptor Res 29 p 275-277
mdash mdash Logozzo D amp Colero C (1997) Migration of ocks of honey buzzards in SouthernItaly and Malta mdash J Raptor Res 31 p 84-86
mdash mdash Malara G Neri F Mollicone D amp Melotto S (1994) Flight strategies of honeybuzzards during spring migration across the Central Mediterranean mdash Avocetta 18p 73-76
398 MEYER SPAAR amp BRUDERER
Bannermann D-A amp Bannermann WM (1971) Handbook of the birds of Cyprus andmigrants of the Middle East mdash Oliver amp Boyd Edinburgh
Batschelet E (1981) Circular statistics in biology mdash Academic Press LondonBaudinette RV amp Schmidt-Nielsen K (1974) Energy costs of gliding ight in herring
gulls mdash Nature 248 p 83-84Beaman M amp Galea C (1974) The visible migration of raptors over the Maltese Islands
mdash Ibis 116 p 419-431Bernis F (1975) Migracion de Falconiformes y Ciconea spp por Gibraltar mdash IV Avance
sobre recuentos en el verano-otontildeo 1974 mdash Ardeola 21 p 581-592Bijlsma RG (1987) Bottleneck areas for migratory birds in the Mediterranean region mdash
ICBP Study report No 18 CambridgeBruderer B amp Liechti F (1998) Flight behaviour of nocturnally migrating birds in coastal
areas mdash crossing or coasting mdash J Avian Biol 29 p 499-510mdash mdash Blitzblau S amp Peter D (1994) Migration and ight behaviour of honey buzzards
Pernis apivorus in southern Israel observed by radar mdash Ardea 82 p 111-122mdash mdash Steuri T amp Baumgartner M (1995) Short-range high-precision surveillance of
nocturnal migration and tracking of single targets mdash Israel J Zool 41 p 207-220Evans PR amp Lathbury GW (1972) Raptor migration across the Strait of Gibraltar mdash Ibis
115 p 572-585Galea C amp Massa B (1985) Notes on the raptor migration across the central Mediter-
ranean mdash In Conservation studies on raptors (I Newton amp RD Chancellor eds)ICBP Technical Publication No 5 p 257-261
Gensboslashl B (1997) Greifvoumlgel mdash BLV VerlagsgesellschaftMuumlnchenGustin M amp Pizzari T (1998) Migratory pattern in the genus Circus sex and age
differential migration in Italy mdash Ornis Svecica 8 p 23-26Kerlinger P (1984) Flight behaviour of sharp-shinned hawks during migration 2 Over
water mdash Anim Beh 32 p 1029-1034mdash mdash (1985) Water-crossing behaviour of raptors during migration mdash Wilson Bull 97
p 109-113mdash mdash (1989) Flight strategies of migrating hawks mdash University of Chicago Press Chicagomdash mdash Bingman VP amp Able KP (1985) Comparative ight behaviour of migrating hawks
studied with tracking radar during autumn in central New York mdash Can J Zool 63p 755-761
Kjelleacuten N (1992) Differential timing of autumn migration between sex and age groups inraptors at Falsterbo Sweden mdash Orn Scan 23 p 420-434
Liechti F (1993) Naumlchtlicher Vogelzug im Herbst uumlber Suumlddeutschland Winddrift undKompensation mdash J Ornithol 134 p 373-404
mdash mdash Hedenstroumlm A amp Alerstam T (1994) Effects of sidewinds on optimal ight speedsof birds mdash J theor Biol 170 p 219-225
Lustick S Batterby B amp Kelty M (1978) Behavioural thermoregulation orientationtoward the sun in herring gulls mdash Sience 200 p 81-83
Pennycuick C (1975) Mechanics of ight mdash In Avian biology Vol 5 (DS Farner ampRJ King eds) Academic Press New York p 1-75
mdash mdash (1989) Bird ight performance a practical calculation manual mdash Oxford UniversityPress Oxford
mdash mdash (1997) Actual and lsquooptimumrsquo ight speeds eld data reassessedmdash J Exp Biol 200p 2355-2361
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 399
Richardson WJ (1978) Timing and amount of bird migration in relation to weather areview mdash Oikos 30 p 224-272
Spaar R (1995) Flight behaviour of steppe buzzard (Buteo buteo vulpinus) during springmigration in southern Israel a tracking radar study mdash Israel J Zool 41 p 489-500
mdash mdash (1996) Flight behaviour of migrating raptors in southern Israel mdash PhD thesisUniversity of Basel Verlag Schweizerische Vogelwarte Sempach
mdash mdash (1997) Flight strategies of migrating raptors a comparative study of interspecicvariation in ight characteristicsmdash Ibis 139 p 523-535
mdash mdash (1999) Flight behaviour of migrating raptors under varying environmental conditionsmdash In Proc 22 Int Ornithol Congr (N Adams amp R Slotow eds) University of NatalDurban p 1844-1862
mdash mdash amp Bruderer B (1997a) Optimal ight behaviour of soaring migrants A case studyof migrating steppe buzzards Buteo buteo vulpinus mdash Behav Ecol 8 p 288-297)
mdash mdash amp Bruderer B (1997b) Migration by apping or soaring ight strategies of marshMontagursquos and pallid harriers in southern Israel mdash Condor 99 p 458-469
mdash mdash Stark H amp Liechti F (1998) Migratory ight strategies of Levant sparrowhawkstime or energy minimization mdash Anim Behav 56 p 1185-1197
Stark H amp Liechti F (1993) Do Levant sparrowhawks Accipiter brevipes also migrate atnight mdash Ibis 135 p 233-236
Thiollay J-M (1977) Importance des populations de rapaces migrateurs en mediterraneeoccidentale mdash Alauda 45 p 115-121
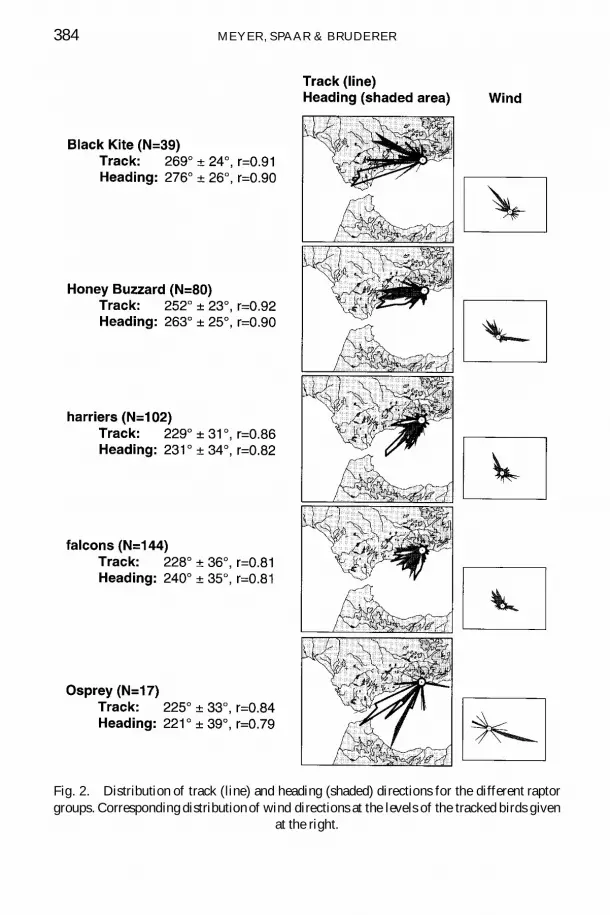
384 MEYER SPAAR amp BRUDERER
Fig 2 Distribution of track (line) and heading (shaded) directions for the different raptorgroups Corresponding distribution of wind directions at the levels of the tracked birds given
at the right
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 385
Fig 3 Proportion of different migratory routes (A) and of time spent in different ightstyles (B without reversed migrants) in the species groups For de nitions see methods Totaltracking time honey buzzard (512 min) black kite (174 min) harriers (369 min) falcons
(497 min) and osprey (107 min)
In uence of environmental conditions on migratory direction
In weak winds (ie wind speed below 4 ms) numbers of falcons harriersand ospreys crossing the sea or following the coast were similar for allwind directions (df = 3 x 2 = 28 NS and x 2 = 64 NS respectivelyTable 1) In strong winds sea crossings occurred in westerly northerlyand easterly winds (ie side and following winds) but not in southerly
386 MEYER SPAAR amp BRUDERER
TABLE 1 Comparison of wind situations and migratory routes
Crossing the sea Flying along the coast
weak wind strong wind weak wind strong wind
West 13 19 5 9North 16 40 9 15East 17 22 7 9South 9 0 1 1
x 2 28 397 64 116p 04 lt 0001 01 lt 001
Number of tracks given Wind direction West (225deg -315deg ) North (315deg -45deg ) East (45deg -135deg ) South (135deg -225deg ) Weak winds are below 4 ms Tracks of harriers falcons andospreys are included
winds (ie opposing winds df = 3 x 2 = 397 p lt 0001 Table 1)Strong northerly winds providing tailwind-support were preferred comparedto strong westerly and easterly winds (df = 2 x 2 = 96 p lt 001) In birdsfollowing the coast under strong winds numbers of observed birds showeda similar pattern as in birds crossing the sea they were higher in northerlywesterly and easterly compared to southerly winds (df = 3 x 2 = 116p lt 001 Table 1) The lower number of birds following the coast instrong southerly winds might be an artefact migratory birds were driftedtowards the mountain chains northerly of the radar station and were out ofthe tracking range of the radar
The tailwind-support achieved by the birds did only partly explain fordifferent migratory routes tailwind-component (t-c) of raptors crossingthe sea was higher than a theoretical t-c for the same birds assuming to y along the coast (270deg paired t-test N = 149 t = 78 p lt 0001Fig 4) On the other hand t-c of raptors ying along the coast was lowerthan the theoretical t-c if crossing the sea (210deg N = 138 t = shy 44p lt 0001) Thus many birds ew along the coast with a lower tailwind-support than if they would have crossed the sea There is an indicationthat good thermal conditions might stimulate birds to migrate along thecoast with a lower tailwind-support compared to sea crossing birds yingalong the coast instead of crossing the sea with a higher tailwind-supporthad higher climbing rates in thermal circling than birds ying along coastwith a higher tailwind-support compared to crossing the sea (mean plusmn SD
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 387
Fig 4 The observed tailwind-components in raptors crossing the sea (A left) and yingalong the coast (B left) versus the theoretical tailwind-componentsfor the alternativestrategyfor the same birds assumed to y along the coast (A right) and crossing the sea (B right)Paired t -test (A) N = 149 t = 78 p lt 0001 () (B) N = 138 t = shy 44 p lt 0001
18 plusmn 09 ms N = 29 and 10 plusmn 05 ms N = 17 respectively t-testt = 31 p lt 0005)
Flight altitudes
For all raptors the median of ight altitudes was below 1000 m above groundlevel (agl) and the 90 altitude limit was below 1800 m agl (Fig 5)Median altitudes differed signi cantly between the species groups (blackkite median 636 m N = 40 honey buzzard 862 m N = 81 harriers910 m N = 102 falcons 949 m N = 146 osprey 951 m N = 17 Kruskal-Wallis ANOVA H = 1835 df = 4 N = 386 p lt 0005) Harriers andfalcons had signi cantly higher ight altitudes than black kites (GT2-testp lt 001) Maximum observed ight levels were at about 1400 m agl inblack kites at 1800 m in ospreys at 2000 m in harriers at 2200 m in honeybuzzards and at 2600 m in falcons (Fig 5) No black kites were tracked in theearly afternoon when ight altitudes of soaring migrants are usually highpartly due to long operational breaks in the rst week of our observationperiod Thus maximum ight levels of black kites were probably higher
388 MEYER SPAAR amp BRUDERER
Fig 5 Flight altitudes above ground (mean per track) in the course of the day (0 =sunrise 100 = sunset) Median 636 m for black kite 862 m for honey buzzard 910 m
for harriers 949 m for falcons and 951 m for osprey
Flight altitudes of honey buzzards black kites and harriers increased fromsunrise towards noon and were more or less stable thereafter (Fig 5) Falconsachieved high ight altitudes already soon after sunrise by apping-gliding ight In falcons ight altitudes differed between migratory routes birdscrossing the sea had higher ight altitudes than birds ying across the bayand along the coast (Fig 6 Kruskal-Wallis ANOVA H = 181 df =2 p = 00001 GT2-test crossing the sea versus ying across the bay(p lt 001) and versus along the coast (p lt 005)) In honey buzzards andharriers differences in ight altitudes between migratory routes showed thesame tendency but did not differ statistically (Fig 6)
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 389
Fig 6 Flight altitudes above ground (mean per track) for different migratory routes inhoney buzzard harriers and falcons Boxes are medians with 25-75 quartiles whiskersshow 10-90 range dots are outliers (ie outside the 15 box length from the upper and
lower value) GT2-test p lt 005 p lt 001
Fig 7 Mean climbing rates in thermal circling in the course of the day (0 = sunrise100 = sunset) Climbing rate did not differ between species groups (ANOVA F = 116
df = 4 N = 116 NS)
Flight characteristics in soaring-gliding
Thermal circling phases were recorded in 116 of 386 tracks and averageclimbing rate was 146 plusmn 084 ms (N = 116) Climbing rates increasedduring the morning hours (Fig 7 time of day lt 50 Spearman rankcorrelation N = 61 r = 034 p lt 001) Strong thermal convection aroundnoon allowed climbing rates of up to 45 ms From noon to sunset climbing
390 MEYER SPAAR amp BRUDERER
TA
BL
E2
Flig
htch
arac
teri
stic
sof
diffe
rent
spec
ies
grou
ps(m
s)
hone
ybu
zzar
dbl
ack
kite
harr
iers
falc
ons
ospr
eyF
Soar
ing
glid
ing
Cli
mbi
ngra
te1
35plusmn
077
(48)
171
plusmn0
97(2
2)1
54plusmn
087
(21)
134
plusmn0
84(2
4)1
86(1
)1
16N
SC
ross
-cou
ntry
grou
ndsp
eed
86
plusmn0
4(2
8)7
6plusmn
07
(7)
93
plusmn0
9(9
)9
3plusmn
10
(6)
89
(1)
079
NS
Cro
ss-c
ount
ryai
rspe
ed10
4plusmn
42
(28)
99
plusmn3
6(7
)9
3plusmn
25
(9)
95
plusmn3
1(1
0)11
3(1
)0
27N
SIn
ter-
ther
mal
grou
ndsp
eed
116
plusmn3
2(4
7)10
8plusmn
25
(21)
121
plusmn2
5(2
1)11
2plusmn
32
(23)
114
(1)
075
NS
Inte
r-th
erm
alai
rspe
ed12
8plusmn
37
(47)
134
plusmn3
0(2
1)12
0plusmn
30
(21)
127
plusmn3
6(2
3)17
0(1
)0
57N
S
Hor
izon
tal
appi
ng-g
lidi
ngi
ght
Gro
unds
peed
138
plusmn2
5(1
6)10
9plusmn
26
(9)
116
plusmn1
9(6
1)12
2plusmn
36
(68)
146
plusmn1
9(1
0)2
47
Air
spee
d11
1plusmn
23
(16)
103
plusmn2
3(9
)11
2plusmn
18
(58)
113
plusmn2
4(6
8)13
2plusmn
30
(10)
387
Mea
nplusmn
SD
(N)
Dif
fere
nces
wer
ete
sted
byA
NO
VA
S
igni
ca
nce
leve
l
plt
005
plt
001
H
oriz
onta
l
appi
ng-g
lidi
ng
ight
was
incl
uded
only
ifve
rtic
alsp
eeds
did
note
xcee
d0
plusmn0
5m
san
dif
noth
erm
alci
rcli
ngw
asob
serv
ed
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 391
rates did not decrease signi cantly (Spearman rank correlation N = 55r = shy 015 NS) Climbing rates in thermal circling below 1 ms occurredduring the whole day They did not differ between species groups (Table 2ANOVA F = 116 df = 3 N = 115 NS) Flight altitudes increased withincreasing climbing rates in birds lsquo ying along the coastrsquo (r = 049 N = 64p lt 00001) Average circling duration in completed soaring phases was106 plusmn 67 s (N = 80) and it was not related to climbing rate (r = 002N = 80 NS)
Mean airspeed and groundspeed in inter-thermal gliding did not differbetween the species groups (Table 2 ANOVA F = 057 and F = 075respectively df = 3 N = 112 NS) and means per species ranged between120-134 ms and 108-121 ms respectively Raptors increased airspeedsin inter-thermal gliding with increasing climbing rates (r = 052 N =113 p lt 00001) indicating that they maximised cross-country speeds insoaring-gliding ight (Pennycuick 1989 Spaar amp Bruderer 1997a) Meancross-country speeds in soaring-gliding did not differ between species groups(ANOVA cross-country airspeed F = 079 df = 3 N = 54 NS range 93-113 ms cross-country groundspeed F = 027 df = 3 N = 54 NS range76-93 ms)
Flight characteristics in horizontal apping-gliding
This ight style was considered if vertical speeds did not exceed 0 plusmn 05 msand no thermal circling was observed within a track Air- and groundspeedsdiffered between species groups (ANOVA airspeed F = 247 df = 4N = 161 p lt 005 groundspeed F = 387 df = 4 N = 164p lt 0005 Table 2) Ospreys had higher groundspeeds than harriers (GT2-test p lt 005) and higher ground- and airspeeds than black kites (p lt 005)Mean airspeeds per species ranged between 103-132 ms correspondinggroundspeeds between 109-146 ms
Flapping-gliding migrants reached signi cantly higher cross-country air-speeds (113 plusmn 22 ms N = 161) and groundspeeds (122 plusmn 31 ms N =164) than birds in soaring-gliding ight (airspeed 100 plusmn 36 ms N = 55Welch-test t = 249 df = 68 p lt 001 groundspeed 87 plusmn 25 msN = 55 t = 846 df = 114 p lt 00005)
In horizontal apping-gliding ight reactions to different wind con-ditions depended on migratory route raptors ying along the coast de-
392 MEYER SPAAR amp BRUDERER
Fig 8 Airspeed in horizontal apping-gliding ight versus tailwind- and sidewind-components for different migratory routes Linear regression (A) excluding one value inbrackets y = shy 0256x + 1128 R 2 = 0215 N = 56 p lt 0001 (B) r = shy 010 N = 57NS (C) r = shy 007 N = 85 NS (D) excluding two values in brackets y = 0191x+ 1072
R2 = 005 N = 84 p lt 005
creased airspeed with increasing tailwind-component (Fig 8A) but air-speed was not related to sidewind-component (Fig 8B) In contrast in rap-tors crossing the sea no relation was found between airspeed and tailwind-component (Fig 8C) but airspeed increased with increasing sidewind-component (Fig 8D)
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 393
Discussion
There are only few studies on the ight behaviour of migrating raptors cross-ing the sea mainly carried out at concentration points such as narrow wa-ter straits or peninsulas (Evans amp Lathbury 1972 Bernis 1975 Kerlinger1984) and none by tracking radar As these studies were limited to visualobservation mainly high altitudinal migration was often not detected (Ker-linger et al 1985) Our approach is different because (i) we observed raptormigration at an average coast site (ii) we used a tracking radar and (iii) wemeasured wind conditions at all ight levels
Migratory routes and in uence of environmental conditions
Although black kites and in larger numbers honey buzzards undertakelong sea-crossings in the Central Mediterranean (Beaman amp Galea 1974Agostini et al 1994) in our study no black kites and few honey buzzardswere observed attempting the sea crossing The majority of harriers falconsand ospreys crossed the sea Sea crossing honey buzzards occurred towardsthe end of the migratory season and were probably juveniles as in honeybuzzards adults migrate two weeks earlier than juveniles (Kjelleacuten 1992Agostini et al 1997) In contrast to the adults which often migrate in large ocks juveniles mainly migrate singly and have no experience about themigratory ight route This may lead to a higher proportion of juvenilescrossing the sea
Wind and thermal conditions affected migratory directions many harri-ers falcons and ospreys crossed the sea in favourable following and sidewinds but no sea crossings were observed in strong opposing winds This isreasonable because energy and time consumption for the sea crossing andtherefore the risk of exhaustion drastically increase in opposing winds How-ever tailwind-support did not fully explain the decisions on migratory routesbecause regularly birds followed the coast with a lower tailwind-supportcompared to crossing the sea These birds following the coast reached highclimbing rates in thermal circling and this might be compensating for thelower tailwind-support Under good thermal conditions birds were able tomigrate in soaring-gliding ight which consumes much less energy com-pared to apping-gliding ight when crossing the sea (see below Energyand time consumption) Anyway birds in bad shape for example those with
394 MEYER SPAAR amp BRUDERER
low fat reserves should follow the coast in order to minimise the risk ofstarvation to feed when ever it is possible and to y in soaring-gliding ight
At the same observation site migratory directions of nocturnal migrants ying below 2000 m above ground shift from 190deg to 250deg within the nightthe majority of the birds cross the sea when approaching the coast in the rsthalf of the night whereas they hesitate later on (Bruderer amp Liechti 1998)Thus the motivation to cross large barriers seems to decrease after somehours of migratory ight
Energy and time consumption
Energy consumption in apping ight disproportionately increases withbody mass while in soaring-gliding ight it is 2-3 times the basal metabolicrate (BMR) only (for herring gull Larus argentatus Baudinette amp Schmidt-Nielson 1974 Lustick et al 1978) In honey buzzards the average observedairspeed in apping-gliding ight was 111 ms and fat consumption perdistance (gkm) for an average honey buzzard (body mass in Table 3) atthis speed is about 6 times higher than in soaring-gliding ight (calculatedafter Pennycuick (1989 version 11) energy consumption in soaring-gliding ight = 2 times BMR body drag coef cient = 04) If the body dragcoef cient is set to 01 as recently proposed by Pennycuick (1997) energyconsumption in apping ight per km is reduced and energetic differencebetween soaring-gliding and apping ight become smaller but calculatedmaximum range speeds in apping ight are very high compared to ourobservations
We calculated energy and time consumption of different migratory routesbased on observed ight speeds for an average honey buzzard (Table 3) Notethat we did not take into account the time needed for refuelling consumedenergy (ie burned fat) and that we assumed zero wind Crossing the sea in apping-gliding ight (150 km distance) took on average about 4 hours andit was the time-minimising strategy for a honey buzzard (Table 3) Flyingalong the coast (250 km) required about 7 hours in soaring-gliding ightand 6 hours in apping-gliding ight Flying along the coast by soaring-gliding is the energy-minimising strategy for a honey buzzard and consumesabout 4 g fat (Table 3) Flapping-gliding ight along the coast is energeticallythe most expensive but never the fastest strategy Thus black kites andhoney buzzards migrated predominantly according to the energy minimising
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 395
TABLE 3 Observed airspeeds and corresponding estimated ight durationand fat consumption for an average honey buzzard when ying along the
coast or across the sea in different ight styles
Flight style Soaring-gliding Horizontal Horizontal apping-gliding apping-gliding
Flight route Flying along the coast Flying along the coast Crossing the seaDistance 250 km 250 km 150 km
Airspeed ms 101 (81-123) 108 (88-133) 108 (88-133)Flight duration hmin 652 (538-834) 626 (513-753) 351 (308-444)Fat consumption g 41 (34-55) 266 (230-298) 158 (138-179)for ight
Average honey buzzard = body mass 08 kg wing span 13 m wing area 025 m2 Medianand 25-75 range are given in brackets Calculations on fat consumption are based onPennycuickrsquos (1989) program of bird ight performance version 11 and birds are assumedto maximize cross-country speed in soaring-gliding ight (Spaar amp Bruderer 1997a)
strategy because a large proportion of the birds followed the coast in soaring-gliding ight
For species such as falcons or small harriers the difference in energyconsumption between apping and soaring-gliding ight is smaller Asthese species regularly use apping-gliding ight also over land or combinesoaring with apping-gliding ight differences in energy consumptionbetween land and sea route become even smaller and only short detours areenergetically favourable By combining soaring and apping-gliding ightsmaller raptors such as falcons or harriers are able to y under varyingenvironmental conditions to extend their migratory activity to times whenthermal updrafts are absent and to migrate even at night (Stark amp Liechti1993 Spaar amp Bruderer 1997b Spaar et al 1998)
Flight altitudes
Flight altitudes differed between the species groups partly due to differencesin migratory routes Birds crossing the sea ew considerably higher thanthose following the coast Kerlinger (1984) made similar observations atthe Delaware Bay birds crossing the bay ew at higher altitudes thanthose keeping on land He explained the higher ight altitudes by the bettervisibility of the other shore However the African coast is not visible fromour study site Some falcons even started crossing the sea after sunset
396 MEYER SPAAR amp BRUDERER
indicating that they continued migration into the night these birds had to y at least three hours until they reached the African coast The higher ightaltitudes of birds crossing the sea at our study site was mainly caused bytopographical features birds ying with southerly directions were forced toclimb higher up when crossing the mountains chains (1500 m height) about10-15 km northward of the coast and they afterwards maintained these ightlevels when approaching the sea
Flight altitudes of birds migrating along the coast were similar to altitudesobserved in North America and southern Israel with most raptors yingbelow 1000 m above ground and lt 5 above 1500 m (Kerlinger 19841989 Bruderer et al 1994 Spaar 1996)
Flight characteristics in soaring-gliding
Flight characteristics in soaring-gliding ight did not differ between thespecies groups (Table 2) As shown for raptor migration in Israel climb-ing rates do not depend primarily on morphological features but on environ-mental conditions (Spaar 1997) In general mean climbing rates in southernSpain were lower than in the semi-arid desert in Israel (Spaar 1997) Meanduration of the completed circling phases was shorter in Israel (88 plusmn 52 sSpaar 1995 1999) than in Spain In contrast to Israel (Spaar 1995 1997)circling time did not increase with increasing climbing rate Compared tothe semi-desert the development of thermal convection in southern Spainwas probably less stable and birds circled as long as possible in updraftsbecause they could not rely to encounter a next strong thermal within glid-ing distance This might also explain why inter-thermal gliding was oftensupported by apping it reduces sinking rate and prolongs gliding distance
In raptors migrating along the coast ight altitude and gliding airspeedswere related to thermal strength raptors circling in strong thermals hadhigher ight altitudes and higher inter-thermal gliding airspeeds than thosein weak thermals This is consistent with optimal ight theory for birds max-imising cross-country speed in soaring-gliding ight (Pennycuick 1989) asdemonstrated for steppe buzzards Buteo buteo vulpinus (Spaar amp Bruderer1997a)
Flight characteristics in horizontal apping-gliding
According to optimal ight theory maximum range speed decreases withfollowing winds and increases with head- and sidewinds (Pennycuick 1975
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 397
Liechti et al 1994) In raptors ying along the coast we observed decreas-ing airspeeds with increasing tailwind-support whereas birds crossing thesea did not reduce airspeeds Why did the birds react differently Sea cross-ings imply higher risks there is no possibility to land and weather (wind)conditions may change rapidly Higher airspeeds in following winds lead toshorter crossing times and lower the risk of unpredictable weather changesFurthermore drift by sidewinds may considerably extent crossing distancesand in order to reduce drift raptors increased airspeeds with increasingsindwind-components In contrast to sharp-shinned hawks Accipiter stria-tus avoiding crossings in strong lateral winds (Kerlinger 1985) we observedmany raptors to cross the sea in strong north-westerly (ie lateral) winds ifthey provided at least partial tailwind-support
Conclusions
Our observations at an average site at the southern coast of Spain showthat differences in water crossing behaviour are mainly species-speci cpresumably due to the costs for different ight strategies additionallyindividuals reacted to environmental factors such as wind and thermalconditions In medium sized raptors such as honey buzzard soaring-gliding ight along the coast is energy-minimising whereas crossing the sea istime-minimising Sea crossings occurred preferably with a good tailwind-support but also under strong sidewinds Raptors crossing the sea did notreduce airspeeds in following winds but increased airspeeds with increasingsidewinds They seemed to minimise the water crossing duration and theeffect of drift by sidewinds Under good thermal conditions birds regularlyfollowed the coast despite a lower tailwind-support compared to crossing thesea
References
Agostini N amp Logozzo D (1995) Autumn migration of honey buzzards in Southern Italymdash J Raptor Res 29 p 275-277
mdash mdash Logozzo D amp Colero C (1997) Migration of ocks of honey buzzards in SouthernItaly and Malta mdash J Raptor Res 31 p 84-86
mdash mdash Malara G Neri F Mollicone D amp Melotto S (1994) Flight strategies of honeybuzzards during spring migration across the Central Mediterranean mdash Avocetta 18p 73-76
398 MEYER SPAAR amp BRUDERER
Bannermann D-A amp Bannermann WM (1971) Handbook of the birds of Cyprus andmigrants of the Middle East mdash Oliver amp Boyd Edinburgh
Batschelet E (1981) Circular statistics in biology mdash Academic Press LondonBaudinette RV amp Schmidt-Nielsen K (1974) Energy costs of gliding ight in herring
gulls mdash Nature 248 p 83-84Beaman M amp Galea C (1974) The visible migration of raptors over the Maltese Islands
mdash Ibis 116 p 419-431Bernis F (1975) Migracion de Falconiformes y Ciconea spp por Gibraltar mdash IV Avance
sobre recuentos en el verano-otontildeo 1974 mdash Ardeola 21 p 581-592Bijlsma RG (1987) Bottleneck areas for migratory birds in the Mediterranean region mdash
ICBP Study report No 18 CambridgeBruderer B amp Liechti F (1998) Flight behaviour of nocturnally migrating birds in coastal
areas mdash crossing or coasting mdash J Avian Biol 29 p 499-510mdash mdash Blitzblau S amp Peter D (1994) Migration and ight behaviour of honey buzzards
Pernis apivorus in southern Israel observed by radar mdash Ardea 82 p 111-122mdash mdash Steuri T amp Baumgartner M (1995) Short-range high-precision surveillance of
nocturnal migration and tracking of single targets mdash Israel J Zool 41 p 207-220Evans PR amp Lathbury GW (1972) Raptor migration across the Strait of Gibraltar mdash Ibis
115 p 572-585Galea C amp Massa B (1985) Notes on the raptor migration across the central Mediter-
ranean mdash In Conservation studies on raptors (I Newton amp RD Chancellor eds)ICBP Technical Publication No 5 p 257-261
Gensboslashl B (1997) Greifvoumlgel mdash BLV VerlagsgesellschaftMuumlnchenGustin M amp Pizzari T (1998) Migratory pattern in the genus Circus sex and age
differential migration in Italy mdash Ornis Svecica 8 p 23-26Kerlinger P (1984) Flight behaviour of sharp-shinned hawks during migration 2 Over
water mdash Anim Beh 32 p 1029-1034mdash mdash (1985) Water-crossing behaviour of raptors during migration mdash Wilson Bull 97
p 109-113mdash mdash (1989) Flight strategies of migrating hawks mdash University of Chicago Press Chicagomdash mdash Bingman VP amp Able KP (1985) Comparative ight behaviour of migrating hawks
studied with tracking radar during autumn in central New York mdash Can J Zool 63p 755-761
Kjelleacuten N (1992) Differential timing of autumn migration between sex and age groups inraptors at Falsterbo Sweden mdash Orn Scan 23 p 420-434
Liechti F (1993) Naumlchtlicher Vogelzug im Herbst uumlber Suumlddeutschland Winddrift undKompensation mdash J Ornithol 134 p 373-404
mdash mdash Hedenstroumlm A amp Alerstam T (1994) Effects of sidewinds on optimal ight speedsof birds mdash J theor Biol 170 p 219-225
Lustick S Batterby B amp Kelty M (1978) Behavioural thermoregulation orientationtoward the sun in herring gulls mdash Sience 200 p 81-83
Pennycuick C (1975) Mechanics of ight mdash In Avian biology Vol 5 (DS Farner ampRJ King eds) Academic Press New York p 1-75
mdash mdash (1989) Bird ight performance a practical calculation manual mdash Oxford UniversityPress Oxford
mdash mdash (1997) Actual and lsquooptimumrsquo ight speeds eld data reassessedmdash J Exp Biol 200p 2355-2361
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 399
Richardson WJ (1978) Timing and amount of bird migration in relation to weather areview mdash Oikos 30 p 224-272
Spaar R (1995) Flight behaviour of steppe buzzard (Buteo buteo vulpinus) during springmigration in southern Israel a tracking radar study mdash Israel J Zool 41 p 489-500
mdash mdash (1996) Flight behaviour of migrating raptors in southern Israel mdash PhD thesisUniversity of Basel Verlag Schweizerische Vogelwarte Sempach
mdash mdash (1997) Flight strategies of migrating raptors a comparative study of interspecicvariation in ight characteristicsmdash Ibis 139 p 523-535
mdash mdash (1999) Flight behaviour of migrating raptors under varying environmental conditionsmdash In Proc 22 Int Ornithol Congr (N Adams amp R Slotow eds) University of NatalDurban p 1844-1862
mdash mdash amp Bruderer B (1997a) Optimal ight behaviour of soaring migrants A case studyof migrating steppe buzzards Buteo buteo vulpinus mdash Behav Ecol 8 p 288-297)
mdash mdash amp Bruderer B (1997b) Migration by apping or soaring ight strategies of marshMontagursquos and pallid harriers in southern Israel mdash Condor 99 p 458-469
mdash mdash Stark H amp Liechti F (1998) Migratory ight strategies of Levant sparrowhawkstime or energy minimization mdash Anim Behav 56 p 1185-1197
Stark H amp Liechti F (1993) Do Levant sparrowhawks Accipiter brevipes also migrate atnight mdash Ibis 135 p 233-236
Thiollay J-M (1977) Importance des populations de rapaces migrateurs en mediterraneeoccidentale mdash Alauda 45 p 115-121
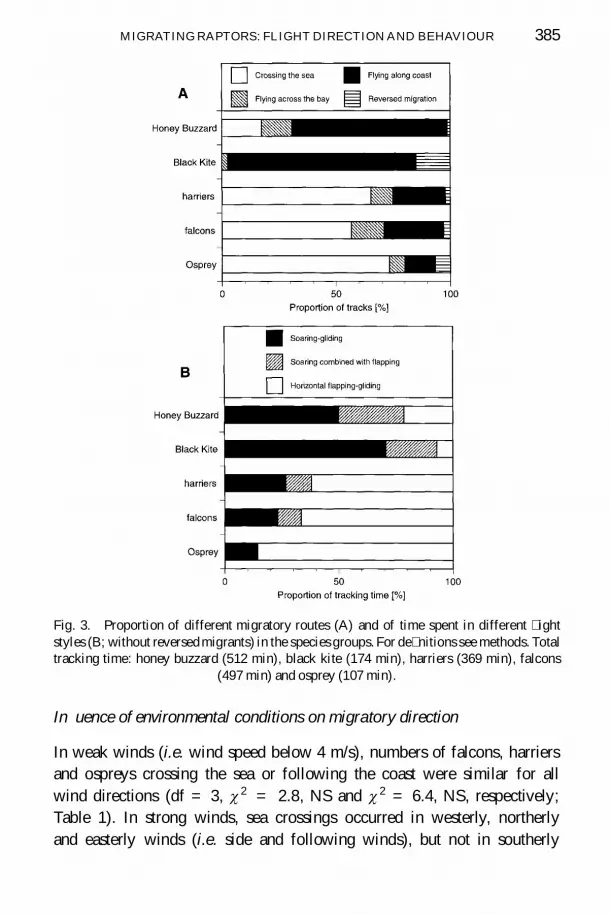
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 385
Fig 3 Proportion of different migratory routes (A) and of time spent in different ightstyles (B without reversed migrants) in the species groups For de nitions see methods Totaltracking time honey buzzard (512 min) black kite (174 min) harriers (369 min) falcons
(497 min) and osprey (107 min)
In uence of environmental conditions on migratory direction
In weak winds (ie wind speed below 4 ms) numbers of falcons harriersand ospreys crossing the sea or following the coast were similar for allwind directions (df = 3 x 2 = 28 NS and x 2 = 64 NS respectivelyTable 1) In strong winds sea crossings occurred in westerly northerlyand easterly winds (ie side and following winds) but not in southerly
386 MEYER SPAAR amp BRUDERER
TABLE 1 Comparison of wind situations and migratory routes
Crossing the sea Flying along the coast
weak wind strong wind weak wind strong wind
West 13 19 5 9North 16 40 9 15East 17 22 7 9South 9 0 1 1
x 2 28 397 64 116p 04 lt 0001 01 lt 001
Number of tracks given Wind direction West (225deg -315deg ) North (315deg -45deg ) East (45deg -135deg ) South (135deg -225deg ) Weak winds are below 4 ms Tracks of harriers falcons andospreys are included
winds (ie opposing winds df = 3 x 2 = 397 p lt 0001 Table 1)Strong northerly winds providing tailwind-support were preferred comparedto strong westerly and easterly winds (df = 2 x 2 = 96 p lt 001) In birdsfollowing the coast under strong winds numbers of observed birds showeda similar pattern as in birds crossing the sea they were higher in northerlywesterly and easterly compared to southerly winds (df = 3 x 2 = 116p lt 001 Table 1) The lower number of birds following the coast instrong southerly winds might be an artefact migratory birds were driftedtowards the mountain chains northerly of the radar station and were out ofthe tracking range of the radar
The tailwind-support achieved by the birds did only partly explain fordifferent migratory routes tailwind-component (t-c) of raptors crossingthe sea was higher than a theoretical t-c for the same birds assuming to y along the coast (270deg paired t-test N = 149 t = 78 p lt 0001Fig 4) On the other hand t-c of raptors ying along the coast was lowerthan the theoretical t-c if crossing the sea (210deg N = 138 t = shy 44p lt 0001) Thus many birds ew along the coast with a lower tailwind-support than if they would have crossed the sea There is an indicationthat good thermal conditions might stimulate birds to migrate along thecoast with a lower tailwind-support compared to sea crossing birds yingalong the coast instead of crossing the sea with a higher tailwind-supporthad higher climbing rates in thermal circling than birds ying along coastwith a higher tailwind-support compared to crossing the sea (mean plusmn SD
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 387
Fig 4 The observed tailwind-components in raptors crossing the sea (A left) and yingalong the coast (B left) versus the theoretical tailwind-componentsfor the alternativestrategyfor the same birds assumed to y along the coast (A right) and crossing the sea (B right)Paired t -test (A) N = 149 t = 78 p lt 0001 () (B) N = 138 t = shy 44 p lt 0001
18 plusmn 09 ms N = 29 and 10 plusmn 05 ms N = 17 respectively t-testt = 31 p lt 0005)
Flight altitudes
For all raptors the median of ight altitudes was below 1000 m above groundlevel (agl) and the 90 altitude limit was below 1800 m agl (Fig 5)Median altitudes differed signi cantly between the species groups (blackkite median 636 m N = 40 honey buzzard 862 m N = 81 harriers910 m N = 102 falcons 949 m N = 146 osprey 951 m N = 17 Kruskal-Wallis ANOVA H = 1835 df = 4 N = 386 p lt 0005) Harriers andfalcons had signi cantly higher ight altitudes than black kites (GT2-testp lt 001) Maximum observed ight levels were at about 1400 m agl inblack kites at 1800 m in ospreys at 2000 m in harriers at 2200 m in honeybuzzards and at 2600 m in falcons (Fig 5) No black kites were tracked in theearly afternoon when ight altitudes of soaring migrants are usually highpartly due to long operational breaks in the rst week of our observationperiod Thus maximum ight levels of black kites were probably higher
388 MEYER SPAAR amp BRUDERER
Fig 5 Flight altitudes above ground (mean per track) in the course of the day (0 =sunrise 100 = sunset) Median 636 m for black kite 862 m for honey buzzard 910 m
for harriers 949 m for falcons and 951 m for osprey
Flight altitudes of honey buzzards black kites and harriers increased fromsunrise towards noon and were more or less stable thereafter (Fig 5) Falconsachieved high ight altitudes already soon after sunrise by apping-gliding ight In falcons ight altitudes differed between migratory routes birdscrossing the sea had higher ight altitudes than birds ying across the bayand along the coast (Fig 6 Kruskal-Wallis ANOVA H = 181 df =2 p = 00001 GT2-test crossing the sea versus ying across the bay(p lt 001) and versus along the coast (p lt 005)) In honey buzzards andharriers differences in ight altitudes between migratory routes showed thesame tendency but did not differ statistically (Fig 6)
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 389
Fig 6 Flight altitudes above ground (mean per track) for different migratory routes inhoney buzzard harriers and falcons Boxes are medians with 25-75 quartiles whiskersshow 10-90 range dots are outliers (ie outside the 15 box length from the upper and
lower value) GT2-test p lt 005 p lt 001
Fig 7 Mean climbing rates in thermal circling in the course of the day (0 = sunrise100 = sunset) Climbing rate did not differ between species groups (ANOVA F = 116
df = 4 N = 116 NS)
Flight characteristics in soaring-gliding
Thermal circling phases were recorded in 116 of 386 tracks and averageclimbing rate was 146 plusmn 084 ms (N = 116) Climbing rates increasedduring the morning hours (Fig 7 time of day lt 50 Spearman rankcorrelation N = 61 r = 034 p lt 001) Strong thermal convection aroundnoon allowed climbing rates of up to 45 ms From noon to sunset climbing
390 MEYER SPAAR amp BRUDERER
TA
BL
E2
Flig
htch
arac
teri
stic
sof
diffe
rent
spec
ies
grou
ps(m
s)
hone
ybu
zzar
dbl
ack
kite
harr
iers
falc
ons
ospr
eyF
Soar
ing
glid
ing
Cli
mbi
ngra
te1
35plusmn
077
(48)
171
plusmn0
97(2
2)1
54plusmn
087
(21)
134
plusmn0
84(2
4)1
86(1
)1
16N
SC
ross
-cou
ntry
grou
ndsp
eed
86
plusmn0
4(2
8)7
6plusmn
07
(7)
93
plusmn0
9(9
)9
3plusmn
10
(6)
89
(1)
079
NS
Cro
ss-c
ount
ryai
rspe
ed10
4plusmn
42
(28)
99
plusmn3
6(7
)9
3plusmn
25
(9)
95
plusmn3
1(1
0)11
3(1
)0
27N
SIn
ter-
ther
mal
grou
ndsp
eed
116
plusmn3
2(4
7)10
8plusmn
25
(21)
121
plusmn2
5(2
1)11
2plusmn
32
(23)
114
(1)
075
NS
Inte
r-th
erm
alai
rspe
ed12
8plusmn
37
(47)
134
plusmn3
0(2
1)12
0plusmn
30
(21)
127
plusmn3
6(2
3)17
0(1
)0
57N
S
Hor
izon
tal
appi
ng-g
lidi
ngi
ght
Gro
unds
peed
138
plusmn2
5(1
6)10
9plusmn
26
(9)
116
plusmn1
9(6
1)12
2plusmn
36
(68)
146
plusmn1
9(1
0)2
47
Air
spee
d11
1plusmn
23
(16)
103
plusmn2
3(9
)11
2plusmn
18
(58)
113
plusmn2
4(6
8)13
2plusmn
30
(10)
387
Mea
nplusmn
SD
(N)
Dif
fere
nces
wer
ete
sted
byA
NO
VA
S
igni
ca
nce
leve
l
plt
005
plt
001
H
oriz
onta
l
appi
ng-g
lidi
ng
ight
was
incl
uded
only
ifve
rtic
alsp
eeds
did
note
xcee
d0
plusmn0
5m
san
dif
noth
erm
alci
rcli
ngw
asob
serv
ed
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 391
rates did not decrease signi cantly (Spearman rank correlation N = 55r = shy 015 NS) Climbing rates in thermal circling below 1 ms occurredduring the whole day They did not differ between species groups (Table 2ANOVA F = 116 df = 3 N = 115 NS) Flight altitudes increased withincreasing climbing rates in birds lsquo ying along the coastrsquo (r = 049 N = 64p lt 00001) Average circling duration in completed soaring phases was106 plusmn 67 s (N = 80) and it was not related to climbing rate (r = 002N = 80 NS)
Mean airspeed and groundspeed in inter-thermal gliding did not differbetween the species groups (Table 2 ANOVA F = 057 and F = 075respectively df = 3 N = 112 NS) and means per species ranged between120-134 ms and 108-121 ms respectively Raptors increased airspeedsin inter-thermal gliding with increasing climbing rates (r = 052 N =113 p lt 00001) indicating that they maximised cross-country speeds insoaring-gliding ight (Pennycuick 1989 Spaar amp Bruderer 1997a) Meancross-country speeds in soaring-gliding did not differ between species groups(ANOVA cross-country airspeed F = 079 df = 3 N = 54 NS range 93-113 ms cross-country groundspeed F = 027 df = 3 N = 54 NS range76-93 ms)
Flight characteristics in horizontal apping-gliding
This ight style was considered if vertical speeds did not exceed 0 plusmn 05 msand no thermal circling was observed within a track Air- and groundspeedsdiffered between species groups (ANOVA airspeed F = 247 df = 4N = 161 p lt 005 groundspeed F = 387 df = 4 N = 164p lt 0005 Table 2) Ospreys had higher groundspeeds than harriers (GT2-test p lt 005) and higher ground- and airspeeds than black kites (p lt 005)Mean airspeeds per species ranged between 103-132 ms correspondinggroundspeeds between 109-146 ms
Flapping-gliding migrants reached signi cantly higher cross-country air-speeds (113 plusmn 22 ms N = 161) and groundspeeds (122 plusmn 31 ms N =164) than birds in soaring-gliding ight (airspeed 100 plusmn 36 ms N = 55Welch-test t = 249 df = 68 p lt 001 groundspeed 87 plusmn 25 msN = 55 t = 846 df = 114 p lt 00005)
In horizontal apping-gliding ight reactions to different wind con-ditions depended on migratory route raptors ying along the coast de-
392 MEYER SPAAR amp BRUDERER
Fig 8 Airspeed in horizontal apping-gliding ight versus tailwind- and sidewind-components for different migratory routes Linear regression (A) excluding one value inbrackets y = shy 0256x + 1128 R 2 = 0215 N = 56 p lt 0001 (B) r = shy 010 N = 57NS (C) r = shy 007 N = 85 NS (D) excluding two values in brackets y = 0191x+ 1072
R2 = 005 N = 84 p lt 005
creased airspeed with increasing tailwind-component (Fig 8A) but air-speed was not related to sidewind-component (Fig 8B) In contrast in rap-tors crossing the sea no relation was found between airspeed and tailwind-component (Fig 8C) but airspeed increased with increasing sidewind-component (Fig 8D)
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 393
Discussion
There are only few studies on the ight behaviour of migrating raptors cross-ing the sea mainly carried out at concentration points such as narrow wa-ter straits or peninsulas (Evans amp Lathbury 1972 Bernis 1975 Kerlinger1984) and none by tracking radar As these studies were limited to visualobservation mainly high altitudinal migration was often not detected (Ker-linger et al 1985) Our approach is different because (i) we observed raptormigration at an average coast site (ii) we used a tracking radar and (iii) wemeasured wind conditions at all ight levels
Migratory routes and in uence of environmental conditions
Although black kites and in larger numbers honey buzzards undertakelong sea-crossings in the Central Mediterranean (Beaman amp Galea 1974Agostini et al 1994) in our study no black kites and few honey buzzardswere observed attempting the sea crossing The majority of harriers falconsand ospreys crossed the sea Sea crossing honey buzzards occurred towardsthe end of the migratory season and were probably juveniles as in honeybuzzards adults migrate two weeks earlier than juveniles (Kjelleacuten 1992Agostini et al 1997) In contrast to the adults which often migrate in large ocks juveniles mainly migrate singly and have no experience about themigratory ight route This may lead to a higher proportion of juvenilescrossing the sea
Wind and thermal conditions affected migratory directions many harri-ers falcons and ospreys crossed the sea in favourable following and sidewinds but no sea crossings were observed in strong opposing winds This isreasonable because energy and time consumption for the sea crossing andtherefore the risk of exhaustion drastically increase in opposing winds How-ever tailwind-support did not fully explain the decisions on migratory routesbecause regularly birds followed the coast with a lower tailwind-supportcompared to crossing the sea These birds following the coast reached highclimbing rates in thermal circling and this might be compensating for thelower tailwind-support Under good thermal conditions birds were able tomigrate in soaring-gliding ight which consumes much less energy com-pared to apping-gliding ight when crossing the sea (see below Energyand time consumption) Anyway birds in bad shape for example those with
394 MEYER SPAAR amp BRUDERER
low fat reserves should follow the coast in order to minimise the risk ofstarvation to feed when ever it is possible and to y in soaring-gliding ight
At the same observation site migratory directions of nocturnal migrants ying below 2000 m above ground shift from 190deg to 250deg within the nightthe majority of the birds cross the sea when approaching the coast in the rsthalf of the night whereas they hesitate later on (Bruderer amp Liechti 1998)Thus the motivation to cross large barriers seems to decrease after somehours of migratory ight
Energy and time consumption
Energy consumption in apping ight disproportionately increases withbody mass while in soaring-gliding ight it is 2-3 times the basal metabolicrate (BMR) only (for herring gull Larus argentatus Baudinette amp Schmidt-Nielson 1974 Lustick et al 1978) In honey buzzards the average observedairspeed in apping-gliding ight was 111 ms and fat consumption perdistance (gkm) for an average honey buzzard (body mass in Table 3) atthis speed is about 6 times higher than in soaring-gliding ight (calculatedafter Pennycuick (1989 version 11) energy consumption in soaring-gliding ight = 2 times BMR body drag coef cient = 04) If the body dragcoef cient is set to 01 as recently proposed by Pennycuick (1997) energyconsumption in apping ight per km is reduced and energetic differencebetween soaring-gliding and apping ight become smaller but calculatedmaximum range speeds in apping ight are very high compared to ourobservations
We calculated energy and time consumption of different migratory routesbased on observed ight speeds for an average honey buzzard (Table 3) Notethat we did not take into account the time needed for refuelling consumedenergy (ie burned fat) and that we assumed zero wind Crossing the sea in apping-gliding ight (150 km distance) took on average about 4 hours andit was the time-minimising strategy for a honey buzzard (Table 3) Flyingalong the coast (250 km) required about 7 hours in soaring-gliding ightand 6 hours in apping-gliding ight Flying along the coast by soaring-gliding is the energy-minimising strategy for a honey buzzard and consumesabout 4 g fat (Table 3) Flapping-gliding ight along the coast is energeticallythe most expensive but never the fastest strategy Thus black kites andhoney buzzards migrated predominantly according to the energy minimising
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 395
TABLE 3 Observed airspeeds and corresponding estimated ight durationand fat consumption for an average honey buzzard when ying along the
coast or across the sea in different ight styles
Flight style Soaring-gliding Horizontal Horizontal apping-gliding apping-gliding
Flight route Flying along the coast Flying along the coast Crossing the seaDistance 250 km 250 km 150 km
Airspeed ms 101 (81-123) 108 (88-133) 108 (88-133)Flight duration hmin 652 (538-834) 626 (513-753) 351 (308-444)Fat consumption g 41 (34-55) 266 (230-298) 158 (138-179)for ight
Average honey buzzard = body mass 08 kg wing span 13 m wing area 025 m2 Medianand 25-75 range are given in brackets Calculations on fat consumption are based onPennycuickrsquos (1989) program of bird ight performance version 11 and birds are assumedto maximize cross-country speed in soaring-gliding ight (Spaar amp Bruderer 1997a)
strategy because a large proportion of the birds followed the coast in soaring-gliding ight
For species such as falcons or small harriers the difference in energyconsumption between apping and soaring-gliding ight is smaller Asthese species regularly use apping-gliding ight also over land or combinesoaring with apping-gliding ight differences in energy consumptionbetween land and sea route become even smaller and only short detours areenergetically favourable By combining soaring and apping-gliding ightsmaller raptors such as falcons or harriers are able to y under varyingenvironmental conditions to extend their migratory activity to times whenthermal updrafts are absent and to migrate even at night (Stark amp Liechti1993 Spaar amp Bruderer 1997b Spaar et al 1998)
Flight altitudes
Flight altitudes differed between the species groups partly due to differencesin migratory routes Birds crossing the sea ew considerably higher thanthose following the coast Kerlinger (1984) made similar observations atthe Delaware Bay birds crossing the bay ew at higher altitudes thanthose keeping on land He explained the higher ight altitudes by the bettervisibility of the other shore However the African coast is not visible fromour study site Some falcons even started crossing the sea after sunset
396 MEYER SPAAR amp BRUDERER
indicating that they continued migration into the night these birds had to y at least three hours until they reached the African coast The higher ightaltitudes of birds crossing the sea at our study site was mainly caused bytopographical features birds ying with southerly directions were forced toclimb higher up when crossing the mountains chains (1500 m height) about10-15 km northward of the coast and they afterwards maintained these ightlevels when approaching the sea
Flight altitudes of birds migrating along the coast were similar to altitudesobserved in North America and southern Israel with most raptors yingbelow 1000 m above ground and lt 5 above 1500 m (Kerlinger 19841989 Bruderer et al 1994 Spaar 1996)
Flight characteristics in soaring-gliding
Flight characteristics in soaring-gliding ight did not differ between thespecies groups (Table 2) As shown for raptor migration in Israel climb-ing rates do not depend primarily on morphological features but on environ-mental conditions (Spaar 1997) In general mean climbing rates in southernSpain were lower than in the semi-arid desert in Israel (Spaar 1997) Meanduration of the completed circling phases was shorter in Israel (88 plusmn 52 sSpaar 1995 1999) than in Spain In contrast to Israel (Spaar 1995 1997)circling time did not increase with increasing climbing rate Compared tothe semi-desert the development of thermal convection in southern Spainwas probably less stable and birds circled as long as possible in updraftsbecause they could not rely to encounter a next strong thermal within glid-ing distance This might also explain why inter-thermal gliding was oftensupported by apping it reduces sinking rate and prolongs gliding distance
In raptors migrating along the coast ight altitude and gliding airspeedswere related to thermal strength raptors circling in strong thermals hadhigher ight altitudes and higher inter-thermal gliding airspeeds than thosein weak thermals This is consistent with optimal ight theory for birds max-imising cross-country speed in soaring-gliding ight (Pennycuick 1989) asdemonstrated for steppe buzzards Buteo buteo vulpinus (Spaar amp Bruderer1997a)
Flight characteristics in horizontal apping-gliding
According to optimal ight theory maximum range speed decreases withfollowing winds and increases with head- and sidewinds (Pennycuick 1975
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 397
Liechti et al 1994) In raptors ying along the coast we observed decreas-ing airspeeds with increasing tailwind-support whereas birds crossing thesea did not reduce airspeeds Why did the birds react differently Sea cross-ings imply higher risks there is no possibility to land and weather (wind)conditions may change rapidly Higher airspeeds in following winds lead toshorter crossing times and lower the risk of unpredictable weather changesFurthermore drift by sidewinds may considerably extent crossing distancesand in order to reduce drift raptors increased airspeeds with increasingsindwind-components In contrast to sharp-shinned hawks Accipiter stria-tus avoiding crossings in strong lateral winds (Kerlinger 1985) we observedmany raptors to cross the sea in strong north-westerly (ie lateral) winds ifthey provided at least partial tailwind-support
Conclusions
Our observations at an average site at the southern coast of Spain showthat differences in water crossing behaviour are mainly species-speci cpresumably due to the costs for different ight strategies additionallyindividuals reacted to environmental factors such as wind and thermalconditions In medium sized raptors such as honey buzzard soaring-gliding ight along the coast is energy-minimising whereas crossing the sea istime-minimising Sea crossings occurred preferably with a good tailwind-support but also under strong sidewinds Raptors crossing the sea did notreduce airspeeds in following winds but increased airspeeds with increasingsidewinds They seemed to minimise the water crossing duration and theeffect of drift by sidewinds Under good thermal conditions birds regularlyfollowed the coast despite a lower tailwind-support compared to crossing thesea
References
Agostini N amp Logozzo D (1995) Autumn migration of honey buzzards in Southern Italymdash J Raptor Res 29 p 275-277
mdash mdash Logozzo D amp Colero C (1997) Migration of ocks of honey buzzards in SouthernItaly and Malta mdash J Raptor Res 31 p 84-86
mdash mdash Malara G Neri F Mollicone D amp Melotto S (1994) Flight strategies of honeybuzzards during spring migration across the Central Mediterranean mdash Avocetta 18p 73-76
398 MEYER SPAAR amp BRUDERER
Bannermann D-A amp Bannermann WM (1971) Handbook of the birds of Cyprus andmigrants of the Middle East mdash Oliver amp Boyd Edinburgh
Batschelet E (1981) Circular statistics in biology mdash Academic Press LondonBaudinette RV amp Schmidt-Nielsen K (1974) Energy costs of gliding ight in herring
gulls mdash Nature 248 p 83-84Beaman M amp Galea C (1974) The visible migration of raptors over the Maltese Islands
mdash Ibis 116 p 419-431Bernis F (1975) Migracion de Falconiformes y Ciconea spp por Gibraltar mdash IV Avance
sobre recuentos en el verano-otontildeo 1974 mdash Ardeola 21 p 581-592Bijlsma RG (1987) Bottleneck areas for migratory birds in the Mediterranean region mdash
ICBP Study report No 18 CambridgeBruderer B amp Liechti F (1998) Flight behaviour of nocturnally migrating birds in coastal
areas mdash crossing or coasting mdash J Avian Biol 29 p 499-510mdash mdash Blitzblau S amp Peter D (1994) Migration and ight behaviour of honey buzzards
Pernis apivorus in southern Israel observed by radar mdash Ardea 82 p 111-122mdash mdash Steuri T amp Baumgartner M (1995) Short-range high-precision surveillance of
nocturnal migration and tracking of single targets mdash Israel J Zool 41 p 207-220Evans PR amp Lathbury GW (1972) Raptor migration across the Strait of Gibraltar mdash Ibis
115 p 572-585Galea C amp Massa B (1985) Notes on the raptor migration across the central Mediter-
ranean mdash In Conservation studies on raptors (I Newton amp RD Chancellor eds)ICBP Technical Publication No 5 p 257-261
Gensboslashl B (1997) Greifvoumlgel mdash BLV VerlagsgesellschaftMuumlnchenGustin M amp Pizzari T (1998) Migratory pattern in the genus Circus sex and age
differential migration in Italy mdash Ornis Svecica 8 p 23-26Kerlinger P (1984) Flight behaviour of sharp-shinned hawks during migration 2 Over
water mdash Anim Beh 32 p 1029-1034mdash mdash (1985) Water-crossing behaviour of raptors during migration mdash Wilson Bull 97
p 109-113mdash mdash (1989) Flight strategies of migrating hawks mdash University of Chicago Press Chicagomdash mdash Bingman VP amp Able KP (1985) Comparative ight behaviour of migrating hawks
studied with tracking radar during autumn in central New York mdash Can J Zool 63p 755-761
Kjelleacuten N (1992) Differential timing of autumn migration between sex and age groups inraptors at Falsterbo Sweden mdash Orn Scan 23 p 420-434
Liechti F (1993) Naumlchtlicher Vogelzug im Herbst uumlber Suumlddeutschland Winddrift undKompensation mdash J Ornithol 134 p 373-404
mdash mdash Hedenstroumlm A amp Alerstam T (1994) Effects of sidewinds on optimal ight speedsof birds mdash J theor Biol 170 p 219-225
Lustick S Batterby B amp Kelty M (1978) Behavioural thermoregulation orientationtoward the sun in herring gulls mdash Sience 200 p 81-83
Pennycuick C (1975) Mechanics of ight mdash In Avian biology Vol 5 (DS Farner ampRJ King eds) Academic Press New York p 1-75
mdash mdash (1989) Bird ight performance a practical calculation manual mdash Oxford UniversityPress Oxford
mdash mdash (1997) Actual and lsquooptimumrsquo ight speeds eld data reassessedmdash J Exp Biol 200p 2355-2361
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 399
Richardson WJ (1978) Timing and amount of bird migration in relation to weather areview mdash Oikos 30 p 224-272
Spaar R (1995) Flight behaviour of steppe buzzard (Buteo buteo vulpinus) during springmigration in southern Israel a tracking radar study mdash Israel J Zool 41 p 489-500
mdash mdash (1996) Flight behaviour of migrating raptors in southern Israel mdash PhD thesisUniversity of Basel Verlag Schweizerische Vogelwarte Sempach
mdash mdash (1997) Flight strategies of migrating raptors a comparative study of interspecicvariation in ight characteristicsmdash Ibis 139 p 523-535
mdash mdash (1999) Flight behaviour of migrating raptors under varying environmental conditionsmdash In Proc 22 Int Ornithol Congr (N Adams amp R Slotow eds) University of NatalDurban p 1844-1862
mdash mdash amp Bruderer B (1997a) Optimal ight behaviour of soaring migrants A case studyof migrating steppe buzzards Buteo buteo vulpinus mdash Behav Ecol 8 p 288-297)
mdash mdash amp Bruderer B (1997b) Migration by apping or soaring ight strategies of marshMontagursquos and pallid harriers in southern Israel mdash Condor 99 p 458-469
mdash mdash Stark H amp Liechti F (1998) Migratory ight strategies of Levant sparrowhawkstime or energy minimization mdash Anim Behav 56 p 1185-1197
Stark H amp Liechti F (1993) Do Levant sparrowhawks Accipiter brevipes also migrate atnight mdash Ibis 135 p 233-236
Thiollay J-M (1977) Importance des populations de rapaces migrateurs en mediterraneeoccidentale mdash Alauda 45 p 115-121

386 MEYER SPAAR amp BRUDERER
TABLE 1 Comparison of wind situations and migratory routes
Crossing the sea Flying along the coast
weak wind strong wind weak wind strong wind
West 13 19 5 9North 16 40 9 15East 17 22 7 9South 9 0 1 1
x 2 28 397 64 116p 04 lt 0001 01 lt 001
Number of tracks given Wind direction West (225deg -315deg ) North (315deg -45deg ) East (45deg -135deg ) South (135deg -225deg ) Weak winds are below 4 ms Tracks of harriers falcons andospreys are included
winds (ie opposing winds df = 3 x 2 = 397 p lt 0001 Table 1)Strong northerly winds providing tailwind-support were preferred comparedto strong westerly and easterly winds (df = 2 x 2 = 96 p lt 001) In birdsfollowing the coast under strong winds numbers of observed birds showeda similar pattern as in birds crossing the sea they were higher in northerlywesterly and easterly compared to southerly winds (df = 3 x 2 = 116p lt 001 Table 1) The lower number of birds following the coast instrong southerly winds might be an artefact migratory birds were driftedtowards the mountain chains northerly of the radar station and were out ofthe tracking range of the radar
The tailwind-support achieved by the birds did only partly explain fordifferent migratory routes tailwind-component (t-c) of raptors crossingthe sea was higher than a theoretical t-c for the same birds assuming to y along the coast (270deg paired t-test N = 149 t = 78 p lt 0001Fig 4) On the other hand t-c of raptors ying along the coast was lowerthan the theoretical t-c if crossing the sea (210deg N = 138 t = shy 44p lt 0001) Thus many birds ew along the coast with a lower tailwind-support than if they would have crossed the sea There is an indicationthat good thermal conditions might stimulate birds to migrate along thecoast with a lower tailwind-support compared to sea crossing birds yingalong the coast instead of crossing the sea with a higher tailwind-supporthad higher climbing rates in thermal circling than birds ying along coastwith a higher tailwind-support compared to crossing the sea (mean plusmn SD
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 387
Fig 4 The observed tailwind-components in raptors crossing the sea (A left) and yingalong the coast (B left) versus the theoretical tailwind-componentsfor the alternativestrategyfor the same birds assumed to y along the coast (A right) and crossing the sea (B right)Paired t -test (A) N = 149 t = 78 p lt 0001 () (B) N = 138 t = shy 44 p lt 0001
18 plusmn 09 ms N = 29 and 10 plusmn 05 ms N = 17 respectively t-testt = 31 p lt 0005)
Flight altitudes
For all raptors the median of ight altitudes was below 1000 m above groundlevel (agl) and the 90 altitude limit was below 1800 m agl (Fig 5)Median altitudes differed signi cantly between the species groups (blackkite median 636 m N = 40 honey buzzard 862 m N = 81 harriers910 m N = 102 falcons 949 m N = 146 osprey 951 m N = 17 Kruskal-Wallis ANOVA H = 1835 df = 4 N = 386 p lt 0005) Harriers andfalcons had signi cantly higher ight altitudes than black kites (GT2-testp lt 001) Maximum observed ight levels were at about 1400 m agl inblack kites at 1800 m in ospreys at 2000 m in harriers at 2200 m in honeybuzzards and at 2600 m in falcons (Fig 5) No black kites were tracked in theearly afternoon when ight altitudes of soaring migrants are usually highpartly due to long operational breaks in the rst week of our observationperiod Thus maximum ight levels of black kites were probably higher
388 MEYER SPAAR amp BRUDERER
Fig 5 Flight altitudes above ground (mean per track) in the course of the day (0 =sunrise 100 = sunset) Median 636 m for black kite 862 m for honey buzzard 910 m
for harriers 949 m for falcons and 951 m for osprey
Flight altitudes of honey buzzards black kites and harriers increased fromsunrise towards noon and were more or less stable thereafter (Fig 5) Falconsachieved high ight altitudes already soon after sunrise by apping-gliding ight In falcons ight altitudes differed between migratory routes birdscrossing the sea had higher ight altitudes than birds ying across the bayand along the coast (Fig 6 Kruskal-Wallis ANOVA H = 181 df =2 p = 00001 GT2-test crossing the sea versus ying across the bay(p lt 001) and versus along the coast (p lt 005)) In honey buzzards andharriers differences in ight altitudes between migratory routes showed thesame tendency but did not differ statistically (Fig 6)
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 389
Fig 6 Flight altitudes above ground (mean per track) for different migratory routes inhoney buzzard harriers and falcons Boxes are medians with 25-75 quartiles whiskersshow 10-90 range dots are outliers (ie outside the 15 box length from the upper and
lower value) GT2-test p lt 005 p lt 001
Fig 7 Mean climbing rates in thermal circling in the course of the day (0 = sunrise100 = sunset) Climbing rate did not differ between species groups (ANOVA F = 116
df = 4 N = 116 NS)
Flight characteristics in soaring-gliding
Thermal circling phases were recorded in 116 of 386 tracks and averageclimbing rate was 146 plusmn 084 ms (N = 116) Climbing rates increasedduring the morning hours (Fig 7 time of day lt 50 Spearman rankcorrelation N = 61 r = 034 p lt 001) Strong thermal convection aroundnoon allowed climbing rates of up to 45 ms From noon to sunset climbing
390 MEYER SPAAR amp BRUDERER
TA
BL
E2
Flig
htch
arac
teri
stic
sof
diffe
rent
spec
ies
grou
ps(m
s)
hone
ybu
zzar
dbl
ack
kite
harr
iers
falc
ons
ospr
eyF
Soar
ing
glid
ing
Cli
mbi
ngra
te1
35plusmn
077
(48)
171
plusmn0
97(2
2)1
54plusmn
087
(21)
134
plusmn0
84(2
4)1
86(1
)1
16N
SC
ross
-cou
ntry
grou
ndsp
eed
86
plusmn0
4(2
8)7
6plusmn
07
(7)
93
plusmn0
9(9
)9
3plusmn
10
(6)
89
(1)
079
NS
Cro
ss-c
ount
ryai
rspe
ed10
4plusmn
42
(28)
99
plusmn3
6(7
)9
3plusmn
25
(9)
95
plusmn3
1(1
0)11
3(1
)0
27N
SIn
ter-
ther
mal
grou
ndsp
eed
116
plusmn3
2(4
7)10
8plusmn
25
(21)
121
plusmn2
5(2
1)11
2plusmn
32
(23)
114
(1)
075
NS
Inte
r-th
erm
alai
rspe
ed12
8plusmn
37
(47)
134
plusmn3
0(2
1)12
0plusmn
30
(21)
127
plusmn3
6(2
3)17
0(1
)0
57N
S
Hor
izon
tal
appi
ng-g
lidi
ngi
ght
Gro
unds
peed
138
plusmn2
5(1
6)10
9plusmn
26
(9)
116
plusmn1
9(6
1)12
2plusmn
36
(68)
146
plusmn1
9(1
0)2
47
Air
spee
d11
1plusmn
23
(16)
103
plusmn2
3(9
)11
2plusmn
18
(58)
113
plusmn2
4(6
8)13
2plusmn
30
(10)
387
Mea
nplusmn
SD
(N)
Dif
fere
nces
wer
ete
sted
byA
NO
VA
S
igni
ca
nce
leve
l
plt
005
plt
001
H
oriz
onta
l
appi
ng-g
lidi
ng
ight
was
incl
uded
only
ifve
rtic
alsp
eeds
did
note
xcee
d0
plusmn0
5m
san
dif
noth
erm
alci
rcli
ngw
asob
serv
ed
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 391
rates did not decrease signi cantly (Spearman rank correlation N = 55r = shy 015 NS) Climbing rates in thermal circling below 1 ms occurredduring the whole day They did not differ between species groups (Table 2ANOVA F = 116 df = 3 N = 115 NS) Flight altitudes increased withincreasing climbing rates in birds lsquo ying along the coastrsquo (r = 049 N = 64p lt 00001) Average circling duration in completed soaring phases was106 plusmn 67 s (N = 80) and it was not related to climbing rate (r = 002N = 80 NS)
Mean airspeed and groundspeed in inter-thermal gliding did not differbetween the species groups (Table 2 ANOVA F = 057 and F = 075respectively df = 3 N = 112 NS) and means per species ranged between120-134 ms and 108-121 ms respectively Raptors increased airspeedsin inter-thermal gliding with increasing climbing rates (r = 052 N =113 p lt 00001) indicating that they maximised cross-country speeds insoaring-gliding ight (Pennycuick 1989 Spaar amp Bruderer 1997a) Meancross-country speeds in soaring-gliding did not differ between species groups(ANOVA cross-country airspeed F = 079 df = 3 N = 54 NS range 93-113 ms cross-country groundspeed F = 027 df = 3 N = 54 NS range76-93 ms)
Flight characteristics in horizontal apping-gliding
This ight style was considered if vertical speeds did not exceed 0 plusmn 05 msand no thermal circling was observed within a track Air- and groundspeedsdiffered between species groups (ANOVA airspeed F = 247 df = 4N = 161 p lt 005 groundspeed F = 387 df = 4 N = 164p lt 0005 Table 2) Ospreys had higher groundspeeds than harriers (GT2-test p lt 005) and higher ground- and airspeeds than black kites (p lt 005)Mean airspeeds per species ranged between 103-132 ms correspondinggroundspeeds between 109-146 ms
Flapping-gliding migrants reached signi cantly higher cross-country air-speeds (113 plusmn 22 ms N = 161) and groundspeeds (122 plusmn 31 ms N =164) than birds in soaring-gliding ight (airspeed 100 plusmn 36 ms N = 55Welch-test t = 249 df = 68 p lt 001 groundspeed 87 plusmn 25 msN = 55 t = 846 df = 114 p lt 00005)
In horizontal apping-gliding ight reactions to different wind con-ditions depended on migratory route raptors ying along the coast de-
392 MEYER SPAAR amp BRUDERER
Fig 8 Airspeed in horizontal apping-gliding ight versus tailwind- and sidewind-components for different migratory routes Linear regression (A) excluding one value inbrackets y = shy 0256x + 1128 R 2 = 0215 N = 56 p lt 0001 (B) r = shy 010 N = 57NS (C) r = shy 007 N = 85 NS (D) excluding two values in brackets y = 0191x+ 1072
R2 = 005 N = 84 p lt 005
creased airspeed with increasing tailwind-component (Fig 8A) but air-speed was not related to sidewind-component (Fig 8B) In contrast in rap-tors crossing the sea no relation was found between airspeed and tailwind-component (Fig 8C) but airspeed increased with increasing sidewind-component (Fig 8D)
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 393
Discussion
There are only few studies on the ight behaviour of migrating raptors cross-ing the sea mainly carried out at concentration points such as narrow wa-ter straits or peninsulas (Evans amp Lathbury 1972 Bernis 1975 Kerlinger1984) and none by tracking radar As these studies were limited to visualobservation mainly high altitudinal migration was often not detected (Ker-linger et al 1985) Our approach is different because (i) we observed raptormigration at an average coast site (ii) we used a tracking radar and (iii) wemeasured wind conditions at all ight levels
Migratory routes and in uence of environmental conditions
Although black kites and in larger numbers honey buzzards undertakelong sea-crossings in the Central Mediterranean (Beaman amp Galea 1974Agostini et al 1994) in our study no black kites and few honey buzzardswere observed attempting the sea crossing The majority of harriers falconsand ospreys crossed the sea Sea crossing honey buzzards occurred towardsthe end of the migratory season and were probably juveniles as in honeybuzzards adults migrate two weeks earlier than juveniles (Kjelleacuten 1992Agostini et al 1997) In contrast to the adults which often migrate in large ocks juveniles mainly migrate singly and have no experience about themigratory ight route This may lead to a higher proportion of juvenilescrossing the sea
Wind and thermal conditions affected migratory directions many harri-ers falcons and ospreys crossed the sea in favourable following and sidewinds but no sea crossings were observed in strong opposing winds This isreasonable because energy and time consumption for the sea crossing andtherefore the risk of exhaustion drastically increase in opposing winds How-ever tailwind-support did not fully explain the decisions on migratory routesbecause regularly birds followed the coast with a lower tailwind-supportcompared to crossing the sea These birds following the coast reached highclimbing rates in thermal circling and this might be compensating for thelower tailwind-support Under good thermal conditions birds were able tomigrate in soaring-gliding ight which consumes much less energy com-pared to apping-gliding ight when crossing the sea (see below Energyand time consumption) Anyway birds in bad shape for example those with
394 MEYER SPAAR amp BRUDERER
low fat reserves should follow the coast in order to minimise the risk ofstarvation to feed when ever it is possible and to y in soaring-gliding ight
At the same observation site migratory directions of nocturnal migrants ying below 2000 m above ground shift from 190deg to 250deg within the nightthe majority of the birds cross the sea when approaching the coast in the rsthalf of the night whereas they hesitate later on (Bruderer amp Liechti 1998)Thus the motivation to cross large barriers seems to decrease after somehours of migratory ight
Energy and time consumption
Energy consumption in apping ight disproportionately increases withbody mass while in soaring-gliding ight it is 2-3 times the basal metabolicrate (BMR) only (for herring gull Larus argentatus Baudinette amp Schmidt-Nielson 1974 Lustick et al 1978) In honey buzzards the average observedairspeed in apping-gliding ight was 111 ms and fat consumption perdistance (gkm) for an average honey buzzard (body mass in Table 3) atthis speed is about 6 times higher than in soaring-gliding ight (calculatedafter Pennycuick (1989 version 11) energy consumption in soaring-gliding ight = 2 times BMR body drag coef cient = 04) If the body dragcoef cient is set to 01 as recently proposed by Pennycuick (1997) energyconsumption in apping ight per km is reduced and energetic differencebetween soaring-gliding and apping ight become smaller but calculatedmaximum range speeds in apping ight are very high compared to ourobservations
We calculated energy and time consumption of different migratory routesbased on observed ight speeds for an average honey buzzard (Table 3) Notethat we did not take into account the time needed for refuelling consumedenergy (ie burned fat) and that we assumed zero wind Crossing the sea in apping-gliding ight (150 km distance) took on average about 4 hours andit was the time-minimising strategy for a honey buzzard (Table 3) Flyingalong the coast (250 km) required about 7 hours in soaring-gliding ightand 6 hours in apping-gliding ight Flying along the coast by soaring-gliding is the energy-minimising strategy for a honey buzzard and consumesabout 4 g fat (Table 3) Flapping-gliding ight along the coast is energeticallythe most expensive but never the fastest strategy Thus black kites andhoney buzzards migrated predominantly according to the energy minimising
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 395
TABLE 3 Observed airspeeds and corresponding estimated ight durationand fat consumption for an average honey buzzard when ying along the
coast or across the sea in different ight styles
Flight style Soaring-gliding Horizontal Horizontal apping-gliding apping-gliding
Flight route Flying along the coast Flying along the coast Crossing the seaDistance 250 km 250 km 150 km
Airspeed ms 101 (81-123) 108 (88-133) 108 (88-133)Flight duration hmin 652 (538-834) 626 (513-753) 351 (308-444)Fat consumption g 41 (34-55) 266 (230-298) 158 (138-179)for ight
Average honey buzzard = body mass 08 kg wing span 13 m wing area 025 m2 Medianand 25-75 range are given in brackets Calculations on fat consumption are based onPennycuickrsquos (1989) program of bird ight performance version 11 and birds are assumedto maximize cross-country speed in soaring-gliding ight (Spaar amp Bruderer 1997a)
strategy because a large proportion of the birds followed the coast in soaring-gliding ight
For species such as falcons or small harriers the difference in energyconsumption between apping and soaring-gliding ight is smaller Asthese species regularly use apping-gliding ight also over land or combinesoaring with apping-gliding ight differences in energy consumptionbetween land and sea route become even smaller and only short detours areenergetically favourable By combining soaring and apping-gliding ightsmaller raptors such as falcons or harriers are able to y under varyingenvironmental conditions to extend their migratory activity to times whenthermal updrafts are absent and to migrate even at night (Stark amp Liechti1993 Spaar amp Bruderer 1997b Spaar et al 1998)
Flight altitudes
Flight altitudes differed between the species groups partly due to differencesin migratory routes Birds crossing the sea ew considerably higher thanthose following the coast Kerlinger (1984) made similar observations atthe Delaware Bay birds crossing the bay ew at higher altitudes thanthose keeping on land He explained the higher ight altitudes by the bettervisibility of the other shore However the African coast is not visible fromour study site Some falcons even started crossing the sea after sunset
396 MEYER SPAAR amp BRUDERER
indicating that they continued migration into the night these birds had to y at least three hours until they reached the African coast The higher ightaltitudes of birds crossing the sea at our study site was mainly caused bytopographical features birds ying with southerly directions were forced toclimb higher up when crossing the mountains chains (1500 m height) about10-15 km northward of the coast and they afterwards maintained these ightlevels when approaching the sea
Flight altitudes of birds migrating along the coast were similar to altitudesobserved in North America and southern Israel with most raptors yingbelow 1000 m above ground and lt 5 above 1500 m (Kerlinger 19841989 Bruderer et al 1994 Spaar 1996)
Flight characteristics in soaring-gliding
Flight characteristics in soaring-gliding ight did not differ between thespecies groups (Table 2) As shown for raptor migration in Israel climb-ing rates do not depend primarily on morphological features but on environ-mental conditions (Spaar 1997) In general mean climbing rates in southernSpain were lower than in the semi-arid desert in Israel (Spaar 1997) Meanduration of the completed circling phases was shorter in Israel (88 plusmn 52 sSpaar 1995 1999) than in Spain In contrast to Israel (Spaar 1995 1997)circling time did not increase with increasing climbing rate Compared tothe semi-desert the development of thermal convection in southern Spainwas probably less stable and birds circled as long as possible in updraftsbecause they could not rely to encounter a next strong thermal within glid-ing distance This might also explain why inter-thermal gliding was oftensupported by apping it reduces sinking rate and prolongs gliding distance
In raptors migrating along the coast ight altitude and gliding airspeedswere related to thermal strength raptors circling in strong thermals hadhigher ight altitudes and higher inter-thermal gliding airspeeds than thosein weak thermals This is consistent with optimal ight theory for birds max-imising cross-country speed in soaring-gliding ight (Pennycuick 1989) asdemonstrated for steppe buzzards Buteo buteo vulpinus (Spaar amp Bruderer1997a)
Flight characteristics in horizontal apping-gliding
According to optimal ight theory maximum range speed decreases withfollowing winds and increases with head- and sidewinds (Pennycuick 1975
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 397
Liechti et al 1994) In raptors ying along the coast we observed decreas-ing airspeeds with increasing tailwind-support whereas birds crossing thesea did not reduce airspeeds Why did the birds react differently Sea cross-ings imply higher risks there is no possibility to land and weather (wind)conditions may change rapidly Higher airspeeds in following winds lead toshorter crossing times and lower the risk of unpredictable weather changesFurthermore drift by sidewinds may considerably extent crossing distancesand in order to reduce drift raptors increased airspeeds with increasingsindwind-components In contrast to sharp-shinned hawks Accipiter stria-tus avoiding crossings in strong lateral winds (Kerlinger 1985) we observedmany raptors to cross the sea in strong north-westerly (ie lateral) winds ifthey provided at least partial tailwind-support
Conclusions
Our observations at an average site at the southern coast of Spain showthat differences in water crossing behaviour are mainly species-speci cpresumably due to the costs for different ight strategies additionallyindividuals reacted to environmental factors such as wind and thermalconditions In medium sized raptors such as honey buzzard soaring-gliding ight along the coast is energy-minimising whereas crossing the sea istime-minimising Sea crossings occurred preferably with a good tailwind-support but also under strong sidewinds Raptors crossing the sea did notreduce airspeeds in following winds but increased airspeeds with increasingsidewinds They seemed to minimise the water crossing duration and theeffect of drift by sidewinds Under good thermal conditions birds regularlyfollowed the coast despite a lower tailwind-support compared to crossing thesea
References
Agostini N amp Logozzo D (1995) Autumn migration of honey buzzards in Southern Italymdash J Raptor Res 29 p 275-277
mdash mdash Logozzo D amp Colero C (1997) Migration of ocks of honey buzzards in SouthernItaly and Malta mdash J Raptor Res 31 p 84-86
mdash mdash Malara G Neri F Mollicone D amp Melotto S (1994) Flight strategies of honeybuzzards during spring migration across the Central Mediterranean mdash Avocetta 18p 73-76
398 MEYER SPAAR amp BRUDERER
Bannermann D-A amp Bannermann WM (1971) Handbook of the birds of Cyprus andmigrants of the Middle East mdash Oliver amp Boyd Edinburgh
Batschelet E (1981) Circular statistics in biology mdash Academic Press LondonBaudinette RV amp Schmidt-Nielsen K (1974) Energy costs of gliding ight in herring
gulls mdash Nature 248 p 83-84Beaman M amp Galea C (1974) The visible migration of raptors over the Maltese Islands
mdash Ibis 116 p 419-431Bernis F (1975) Migracion de Falconiformes y Ciconea spp por Gibraltar mdash IV Avance
sobre recuentos en el verano-otontildeo 1974 mdash Ardeola 21 p 581-592Bijlsma RG (1987) Bottleneck areas for migratory birds in the Mediterranean region mdash
ICBP Study report No 18 CambridgeBruderer B amp Liechti F (1998) Flight behaviour of nocturnally migrating birds in coastal
areas mdash crossing or coasting mdash J Avian Biol 29 p 499-510mdash mdash Blitzblau S amp Peter D (1994) Migration and ight behaviour of honey buzzards
Pernis apivorus in southern Israel observed by radar mdash Ardea 82 p 111-122mdash mdash Steuri T amp Baumgartner M (1995) Short-range high-precision surveillance of
nocturnal migration and tracking of single targets mdash Israel J Zool 41 p 207-220Evans PR amp Lathbury GW (1972) Raptor migration across the Strait of Gibraltar mdash Ibis
115 p 572-585Galea C amp Massa B (1985) Notes on the raptor migration across the central Mediter-
ranean mdash In Conservation studies on raptors (I Newton amp RD Chancellor eds)ICBP Technical Publication No 5 p 257-261
Gensboslashl B (1997) Greifvoumlgel mdash BLV VerlagsgesellschaftMuumlnchenGustin M amp Pizzari T (1998) Migratory pattern in the genus Circus sex and age
differential migration in Italy mdash Ornis Svecica 8 p 23-26Kerlinger P (1984) Flight behaviour of sharp-shinned hawks during migration 2 Over
water mdash Anim Beh 32 p 1029-1034mdash mdash (1985) Water-crossing behaviour of raptors during migration mdash Wilson Bull 97
p 109-113mdash mdash (1989) Flight strategies of migrating hawks mdash University of Chicago Press Chicagomdash mdash Bingman VP amp Able KP (1985) Comparative ight behaviour of migrating hawks
studied with tracking radar during autumn in central New York mdash Can J Zool 63p 755-761
Kjelleacuten N (1992) Differential timing of autumn migration between sex and age groups inraptors at Falsterbo Sweden mdash Orn Scan 23 p 420-434
Liechti F (1993) Naumlchtlicher Vogelzug im Herbst uumlber Suumlddeutschland Winddrift undKompensation mdash J Ornithol 134 p 373-404
mdash mdash Hedenstroumlm A amp Alerstam T (1994) Effects of sidewinds on optimal ight speedsof birds mdash J theor Biol 170 p 219-225
Lustick S Batterby B amp Kelty M (1978) Behavioural thermoregulation orientationtoward the sun in herring gulls mdash Sience 200 p 81-83
Pennycuick C (1975) Mechanics of ight mdash In Avian biology Vol 5 (DS Farner ampRJ King eds) Academic Press New York p 1-75
mdash mdash (1989) Bird ight performance a practical calculation manual mdash Oxford UniversityPress Oxford
mdash mdash (1997) Actual and lsquooptimumrsquo ight speeds eld data reassessedmdash J Exp Biol 200p 2355-2361
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 399
Richardson WJ (1978) Timing and amount of bird migration in relation to weather areview mdash Oikos 30 p 224-272
Spaar R (1995) Flight behaviour of steppe buzzard (Buteo buteo vulpinus) during springmigration in southern Israel a tracking radar study mdash Israel J Zool 41 p 489-500
mdash mdash (1996) Flight behaviour of migrating raptors in southern Israel mdash PhD thesisUniversity of Basel Verlag Schweizerische Vogelwarte Sempach
mdash mdash (1997) Flight strategies of migrating raptors a comparative study of interspecicvariation in ight characteristicsmdash Ibis 139 p 523-535
mdash mdash (1999) Flight behaviour of migrating raptors under varying environmental conditionsmdash In Proc 22 Int Ornithol Congr (N Adams amp R Slotow eds) University of NatalDurban p 1844-1862
mdash mdash amp Bruderer B (1997a) Optimal ight behaviour of soaring migrants A case studyof migrating steppe buzzards Buteo buteo vulpinus mdash Behav Ecol 8 p 288-297)
mdash mdash amp Bruderer B (1997b) Migration by apping or soaring ight strategies of marshMontagursquos and pallid harriers in southern Israel mdash Condor 99 p 458-469
mdash mdash Stark H amp Liechti F (1998) Migratory ight strategies of Levant sparrowhawkstime or energy minimization mdash Anim Behav 56 p 1185-1197
Stark H amp Liechti F (1993) Do Levant sparrowhawks Accipiter brevipes also migrate atnight mdash Ibis 135 p 233-236
Thiollay J-M (1977) Importance des populations de rapaces migrateurs en mediterraneeoccidentale mdash Alauda 45 p 115-121

MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 387
Fig 4 The observed tailwind-components in raptors crossing the sea (A left) and yingalong the coast (B left) versus the theoretical tailwind-componentsfor the alternativestrategyfor the same birds assumed to y along the coast (A right) and crossing the sea (B right)Paired t -test (A) N = 149 t = 78 p lt 0001 () (B) N = 138 t = shy 44 p lt 0001
18 plusmn 09 ms N = 29 and 10 plusmn 05 ms N = 17 respectively t-testt = 31 p lt 0005)
Flight altitudes
For all raptors the median of ight altitudes was below 1000 m above groundlevel (agl) and the 90 altitude limit was below 1800 m agl (Fig 5)Median altitudes differed signi cantly between the species groups (blackkite median 636 m N = 40 honey buzzard 862 m N = 81 harriers910 m N = 102 falcons 949 m N = 146 osprey 951 m N = 17 Kruskal-Wallis ANOVA H = 1835 df = 4 N = 386 p lt 0005) Harriers andfalcons had signi cantly higher ight altitudes than black kites (GT2-testp lt 001) Maximum observed ight levels were at about 1400 m agl inblack kites at 1800 m in ospreys at 2000 m in harriers at 2200 m in honeybuzzards and at 2600 m in falcons (Fig 5) No black kites were tracked in theearly afternoon when ight altitudes of soaring migrants are usually highpartly due to long operational breaks in the rst week of our observationperiod Thus maximum ight levels of black kites were probably higher
388 MEYER SPAAR amp BRUDERER
Fig 5 Flight altitudes above ground (mean per track) in the course of the day (0 =sunrise 100 = sunset) Median 636 m for black kite 862 m for honey buzzard 910 m
for harriers 949 m for falcons and 951 m for osprey
Flight altitudes of honey buzzards black kites and harriers increased fromsunrise towards noon and were more or less stable thereafter (Fig 5) Falconsachieved high ight altitudes already soon after sunrise by apping-gliding ight In falcons ight altitudes differed between migratory routes birdscrossing the sea had higher ight altitudes than birds ying across the bayand along the coast (Fig 6 Kruskal-Wallis ANOVA H = 181 df =2 p = 00001 GT2-test crossing the sea versus ying across the bay(p lt 001) and versus along the coast (p lt 005)) In honey buzzards andharriers differences in ight altitudes between migratory routes showed thesame tendency but did not differ statistically (Fig 6)
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 389
Fig 6 Flight altitudes above ground (mean per track) for different migratory routes inhoney buzzard harriers and falcons Boxes are medians with 25-75 quartiles whiskersshow 10-90 range dots are outliers (ie outside the 15 box length from the upper and
lower value) GT2-test p lt 005 p lt 001
Fig 7 Mean climbing rates in thermal circling in the course of the day (0 = sunrise100 = sunset) Climbing rate did not differ between species groups (ANOVA F = 116
df = 4 N = 116 NS)
Flight characteristics in soaring-gliding
Thermal circling phases were recorded in 116 of 386 tracks and averageclimbing rate was 146 plusmn 084 ms (N = 116) Climbing rates increasedduring the morning hours (Fig 7 time of day lt 50 Spearman rankcorrelation N = 61 r = 034 p lt 001) Strong thermal convection aroundnoon allowed climbing rates of up to 45 ms From noon to sunset climbing
390 MEYER SPAAR amp BRUDERER
TA
BL
E2
Flig
htch
arac
teri
stic
sof
diffe
rent
spec
ies
grou
ps(m
s)
hone
ybu
zzar
dbl
ack
kite
harr
iers
falc
ons
ospr
eyF
Soar
ing
glid
ing
Cli
mbi
ngra
te1
35plusmn
077
(48)
171
plusmn0
97(2
2)1
54plusmn
087
(21)
134
plusmn0
84(2
4)1
86(1
)1
16N
SC
ross
-cou
ntry
grou
ndsp
eed
86
plusmn0
4(2
8)7
6plusmn
07
(7)
93
plusmn0
9(9
)9
3plusmn
10
(6)
89
(1)
079
NS
Cro
ss-c
ount
ryai
rspe
ed10
4plusmn
42
(28)
99
plusmn3
6(7
)9
3plusmn
25
(9)
95
plusmn3
1(1
0)11
3(1
)0
27N
SIn
ter-
ther
mal
grou
ndsp
eed
116
plusmn3
2(4
7)10
8plusmn
25
(21)
121
plusmn2
5(2
1)11
2plusmn
32
(23)
114
(1)
075
NS
Inte
r-th
erm
alai
rspe
ed12
8plusmn
37
(47)
134
plusmn3
0(2
1)12
0plusmn
30
(21)
127
plusmn3
6(2
3)17
0(1
)0
57N
S
Hor
izon
tal
appi
ng-g
lidi
ngi
ght
Gro
unds
peed
138
plusmn2
5(1
6)10
9plusmn
26
(9)
116
plusmn1
9(6
1)12
2plusmn
36
(68)
146
plusmn1
9(1
0)2
47
Air
spee
d11
1plusmn
23
(16)
103
plusmn2
3(9
)11
2plusmn
18
(58)
113
plusmn2
4(6
8)13
2plusmn
30
(10)
387
Mea
nplusmn
SD
(N)
Dif
fere
nces
wer
ete
sted
byA
NO
VA
S
igni
ca
nce
leve
l
plt
005
plt
001
H
oriz
onta
l
appi
ng-g
lidi
ng
ight
was
incl
uded
only
ifve
rtic
alsp
eeds
did
note
xcee
d0
plusmn0
5m
san
dif
noth
erm
alci
rcli
ngw
asob
serv
ed
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 391
rates did not decrease signi cantly (Spearman rank correlation N = 55r = shy 015 NS) Climbing rates in thermal circling below 1 ms occurredduring the whole day They did not differ between species groups (Table 2ANOVA F = 116 df = 3 N = 115 NS) Flight altitudes increased withincreasing climbing rates in birds lsquo ying along the coastrsquo (r = 049 N = 64p lt 00001) Average circling duration in completed soaring phases was106 plusmn 67 s (N = 80) and it was not related to climbing rate (r = 002N = 80 NS)
Mean airspeed and groundspeed in inter-thermal gliding did not differbetween the species groups (Table 2 ANOVA F = 057 and F = 075respectively df = 3 N = 112 NS) and means per species ranged between120-134 ms and 108-121 ms respectively Raptors increased airspeedsin inter-thermal gliding with increasing climbing rates (r = 052 N =113 p lt 00001) indicating that they maximised cross-country speeds insoaring-gliding ight (Pennycuick 1989 Spaar amp Bruderer 1997a) Meancross-country speeds in soaring-gliding did not differ between species groups(ANOVA cross-country airspeed F = 079 df = 3 N = 54 NS range 93-113 ms cross-country groundspeed F = 027 df = 3 N = 54 NS range76-93 ms)
Flight characteristics in horizontal apping-gliding
This ight style was considered if vertical speeds did not exceed 0 plusmn 05 msand no thermal circling was observed within a track Air- and groundspeedsdiffered between species groups (ANOVA airspeed F = 247 df = 4N = 161 p lt 005 groundspeed F = 387 df = 4 N = 164p lt 0005 Table 2) Ospreys had higher groundspeeds than harriers (GT2-test p lt 005) and higher ground- and airspeeds than black kites (p lt 005)Mean airspeeds per species ranged between 103-132 ms correspondinggroundspeeds between 109-146 ms
Flapping-gliding migrants reached signi cantly higher cross-country air-speeds (113 plusmn 22 ms N = 161) and groundspeeds (122 plusmn 31 ms N =164) than birds in soaring-gliding ight (airspeed 100 plusmn 36 ms N = 55Welch-test t = 249 df = 68 p lt 001 groundspeed 87 plusmn 25 msN = 55 t = 846 df = 114 p lt 00005)
In horizontal apping-gliding ight reactions to different wind con-ditions depended on migratory route raptors ying along the coast de-
392 MEYER SPAAR amp BRUDERER
Fig 8 Airspeed in horizontal apping-gliding ight versus tailwind- and sidewind-components for different migratory routes Linear regression (A) excluding one value inbrackets y = shy 0256x + 1128 R 2 = 0215 N = 56 p lt 0001 (B) r = shy 010 N = 57NS (C) r = shy 007 N = 85 NS (D) excluding two values in brackets y = 0191x+ 1072
R2 = 005 N = 84 p lt 005
creased airspeed with increasing tailwind-component (Fig 8A) but air-speed was not related to sidewind-component (Fig 8B) In contrast in rap-tors crossing the sea no relation was found between airspeed and tailwind-component (Fig 8C) but airspeed increased with increasing sidewind-component (Fig 8D)
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 393
Discussion
There are only few studies on the ight behaviour of migrating raptors cross-ing the sea mainly carried out at concentration points such as narrow wa-ter straits or peninsulas (Evans amp Lathbury 1972 Bernis 1975 Kerlinger1984) and none by tracking radar As these studies were limited to visualobservation mainly high altitudinal migration was often not detected (Ker-linger et al 1985) Our approach is different because (i) we observed raptormigration at an average coast site (ii) we used a tracking radar and (iii) wemeasured wind conditions at all ight levels
Migratory routes and in uence of environmental conditions
Although black kites and in larger numbers honey buzzards undertakelong sea-crossings in the Central Mediterranean (Beaman amp Galea 1974Agostini et al 1994) in our study no black kites and few honey buzzardswere observed attempting the sea crossing The majority of harriers falconsand ospreys crossed the sea Sea crossing honey buzzards occurred towardsthe end of the migratory season and were probably juveniles as in honeybuzzards adults migrate two weeks earlier than juveniles (Kjelleacuten 1992Agostini et al 1997) In contrast to the adults which often migrate in large ocks juveniles mainly migrate singly and have no experience about themigratory ight route This may lead to a higher proportion of juvenilescrossing the sea
Wind and thermal conditions affected migratory directions many harri-ers falcons and ospreys crossed the sea in favourable following and sidewinds but no sea crossings were observed in strong opposing winds This isreasonable because energy and time consumption for the sea crossing andtherefore the risk of exhaustion drastically increase in opposing winds How-ever tailwind-support did not fully explain the decisions on migratory routesbecause regularly birds followed the coast with a lower tailwind-supportcompared to crossing the sea These birds following the coast reached highclimbing rates in thermal circling and this might be compensating for thelower tailwind-support Under good thermal conditions birds were able tomigrate in soaring-gliding ight which consumes much less energy com-pared to apping-gliding ight when crossing the sea (see below Energyand time consumption) Anyway birds in bad shape for example those with
394 MEYER SPAAR amp BRUDERER
low fat reserves should follow the coast in order to minimise the risk ofstarvation to feed when ever it is possible and to y in soaring-gliding ight
At the same observation site migratory directions of nocturnal migrants ying below 2000 m above ground shift from 190deg to 250deg within the nightthe majority of the birds cross the sea when approaching the coast in the rsthalf of the night whereas they hesitate later on (Bruderer amp Liechti 1998)Thus the motivation to cross large barriers seems to decrease after somehours of migratory ight
Energy and time consumption
Energy consumption in apping ight disproportionately increases withbody mass while in soaring-gliding ight it is 2-3 times the basal metabolicrate (BMR) only (for herring gull Larus argentatus Baudinette amp Schmidt-Nielson 1974 Lustick et al 1978) In honey buzzards the average observedairspeed in apping-gliding ight was 111 ms and fat consumption perdistance (gkm) for an average honey buzzard (body mass in Table 3) atthis speed is about 6 times higher than in soaring-gliding ight (calculatedafter Pennycuick (1989 version 11) energy consumption in soaring-gliding ight = 2 times BMR body drag coef cient = 04) If the body dragcoef cient is set to 01 as recently proposed by Pennycuick (1997) energyconsumption in apping ight per km is reduced and energetic differencebetween soaring-gliding and apping ight become smaller but calculatedmaximum range speeds in apping ight are very high compared to ourobservations
We calculated energy and time consumption of different migratory routesbased on observed ight speeds for an average honey buzzard (Table 3) Notethat we did not take into account the time needed for refuelling consumedenergy (ie burned fat) and that we assumed zero wind Crossing the sea in apping-gliding ight (150 km distance) took on average about 4 hours andit was the time-minimising strategy for a honey buzzard (Table 3) Flyingalong the coast (250 km) required about 7 hours in soaring-gliding ightand 6 hours in apping-gliding ight Flying along the coast by soaring-gliding is the energy-minimising strategy for a honey buzzard and consumesabout 4 g fat (Table 3) Flapping-gliding ight along the coast is energeticallythe most expensive but never the fastest strategy Thus black kites andhoney buzzards migrated predominantly according to the energy minimising
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 395
TABLE 3 Observed airspeeds and corresponding estimated ight durationand fat consumption for an average honey buzzard when ying along the
coast or across the sea in different ight styles
Flight style Soaring-gliding Horizontal Horizontal apping-gliding apping-gliding
Flight route Flying along the coast Flying along the coast Crossing the seaDistance 250 km 250 km 150 km
Airspeed ms 101 (81-123) 108 (88-133) 108 (88-133)Flight duration hmin 652 (538-834) 626 (513-753) 351 (308-444)Fat consumption g 41 (34-55) 266 (230-298) 158 (138-179)for ight
Average honey buzzard = body mass 08 kg wing span 13 m wing area 025 m2 Medianand 25-75 range are given in brackets Calculations on fat consumption are based onPennycuickrsquos (1989) program of bird ight performance version 11 and birds are assumedto maximize cross-country speed in soaring-gliding ight (Spaar amp Bruderer 1997a)
strategy because a large proportion of the birds followed the coast in soaring-gliding ight
For species such as falcons or small harriers the difference in energyconsumption between apping and soaring-gliding ight is smaller Asthese species regularly use apping-gliding ight also over land or combinesoaring with apping-gliding ight differences in energy consumptionbetween land and sea route become even smaller and only short detours areenergetically favourable By combining soaring and apping-gliding ightsmaller raptors such as falcons or harriers are able to y under varyingenvironmental conditions to extend their migratory activity to times whenthermal updrafts are absent and to migrate even at night (Stark amp Liechti1993 Spaar amp Bruderer 1997b Spaar et al 1998)
Flight altitudes
Flight altitudes differed between the species groups partly due to differencesin migratory routes Birds crossing the sea ew considerably higher thanthose following the coast Kerlinger (1984) made similar observations atthe Delaware Bay birds crossing the bay ew at higher altitudes thanthose keeping on land He explained the higher ight altitudes by the bettervisibility of the other shore However the African coast is not visible fromour study site Some falcons even started crossing the sea after sunset
396 MEYER SPAAR amp BRUDERER
indicating that they continued migration into the night these birds had to y at least three hours until they reached the African coast The higher ightaltitudes of birds crossing the sea at our study site was mainly caused bytopographical features birds ying with southerly directions were forced toclimb higher up when crossing the mountains chains (1500 m height) about10-15 km northward of the coast and they afterwards maintained these ightlevels when approaching the sea
Flight altitudes of birds migrating along the coast were similar to altitudesobserved in North America and southern Israel with most raptors yingbelow 1000 m above ground and lt 5 above 1500 m (Kerlinger 19841989 Bruderer et al 1994 Spaar 1996)
Flight characteristics in soaring-gliding
Flight characteristics in soaring-gliding ight did not differ between thespecies groups (Table 2) As shown for raptor migration in Israel climb-ing rates do not depend primarily on morphological features but on environ-mental conditions (Spaar 1997) In general mean climbing rates in southernSpain were lower than in the semi-arid desert in Israel (Spaar 1997) Meanduration of the completed circling phases was shorter in Israel (88 plusmn 52 sSpaar 1995 1999) than in Spain In contrast to Israel (Spaar 1995 1997)circling time did not increase with increasing climbing rate Compared tothe semi-desert the development of thermal convection in southern Spainwas probably less stable and birds circled as long as possible in updraftsbecause they could not rely to encounter a next strong thermal within glid-ing distance This might also explain why inter-thermal gliding was oftensupported by apping it reduces sinking rate and prolongs gliding distance
In raptors migrating along the coast ight altitude and gliding airspeedswere related to thermal strength raptors circling in strong thermals hadhigher ight altitudes and higher inter-thermal gliding airspeeds than thosein weak thermals This is consistent with optimal ight theory for birds max-imising cross-country speed in soaring-gliding ight (Pennycuick 1989) asdemonstrated for steppe buzzards Buteo buteo vulpinus (Spaar amp Bruderer1997a)
Flight characteristics in horizontal apping-gliding
According to optimal ight theory maximum range speed decreases withfollowing winds and increases with head- and sidewinds (Pennycuick 1975
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 397
Liechti et al 1994) In raptors ying along the coast we observed decreas-ing airspeeds with increasing tailwind-support whereas birds crossing thesea did not reduce airspeeds Why did the birds react differently Sea cross-ings imply higher risks there is no possibility to land and weather (wind)conditions may change rapidly Higher airspeeds in following winds lead toshorter crossing times and lower the risk of unpredictable weather changesFurthermore drift by sidewinds may considerably extent crossing distancesand in order to reduce drift raptors increased airspeeds with increasingsindwind-components In contrast to sharp-shinned hawks Accipiter stria-tus avoiding crossings in strong lateral winds (Kerlinger 1985) we observedmany raptors to cross the sea in strong north-westerly (ie lateral) winds ifthey provided at least partial tailwind-support
Conclusions
Our observations at an average site at the southern coast of Spain showthat differences in water crossing behaviour are mainly species-speci cpresumably due to the costs for different ight strategies additionallyindividuals reacted to environmental factors such as wind and thermalconditions In medium sized raptors such as honey buzzard soaring-gliding ight along the coast is energy-minimising whereas crossing the sea istime-minimising Sea crossings occurred preferably with a good tailwind-support but also under strong sidewinds Raptors crossing the sea did notreduce airspeeds in following winds but increased airspeeds with increasingsidewinds They seemed to minimise the water crossing duration and theeffect of drift by sidewinds Under good thermal conditions birds regularlyfollowed the coast despite a lower tailwind-support compared to crossing thesea
References
Agostini N amp Logozzo D (1995) Autumn migration of honey buzzards in Southern Italymdash J Raptor Res 29 p 275-277
mdash mdash Logozzo D amp Colero C (1997) Migration of ocks of honey buzzards in SouthernItaly and Malta mdash J Raptor Res 31 p 84-86
mdash mdash Malara G Neri F Mollicone D amp Melotto S (1994) Flight strategies of honeybuzzards during spring migration across the Central Mediterranean mdash Avocetta 18p 73-76
398 MEYER SPAAR amp BRUDERER
Bannermann D-A amp Bannermann WM (1971) Handbook of the birds of Cyprus andmigrants of the Middle East mdash Oliver amp Boyd Edinburgh
Batschelet E (1981) Circular statistics in biology mdash Academic Press LondonBaudinette RV amp Schmidt-Nielsen K (1974) Energy costs of gliding ight in herring
gulls mdash Nature 248 p 83-84Beaman M amp Galea C (1974) The visible migration of raptors over the Maltese Islands
mdash Ibis 116 p 419-431Bernis F (1975) Migracion de Falconiformes y Ciconea spp por Gibraltar mdash IV Avance
sobre recuentos en el verano-otontildeo 1974 mdash Ardeola 21 p 581-592Bijlsma RG (1987) Bottleneck areas for migratory birds in the Mediterranean region mdash
ICBP Study report No 18 CambridgeBruderer B amp Liechti F (1998) Flight behaviour of nocturnally migrating birds in coastal
areas mdash crossing or coasting mdash J Avian Biol 29 p 499-510mdash mdash Blitzblau S amp Peter D (1994) Migration and ight behaviour of honey buzzards
Pernis apivorus in southern Israel observed by radar mdash Ardea 82 p 111-122mdash mdash Steuri T amp Baumgartner M (1995) Short-range high-precision surveillance of
nocturnal migration and tracking of single targets mdash Israel J Zool 41 p 207-220Evans PR amp Lathbury GW (1972) Raptor migration across the Strait of Gibraltar mdash Ibis
115 p 572-585Galea C amp Massa B (1985) Notes on the raptor migration across the central Mediter-
ranean mdash In Conservation studies on raptors (I Newton amp RD Chancellor eds)ICBP Technical Publication No 5 p 257-261
Gensboslashl B (1997) Greifvoumlgel mdash BLV VerlagsgesellschaftMuumlnchenGustin M amp Pizzari T (1998) Migratory pattern in the genus Circus sex and age
differential migration in Italy mdash Ornis Svecica 8 p 23-26Kerlinger P (1984) Flight behaviour of sharp-shinned hawks during migration 2 Over
water mdash Anim Beh 32 p 1029-1034mdash mdash (1985) Water-crossing behaviour of raptors during migration mdash Wilson Bull 97
p 109-113mdash mdash (1989) Flight strategies of migrating hawks mdash University of Chicago Press Chicagomdash mdash Bingman VP amp Able KP (1985) Comparative ight behaviour of migrating hawks
studied with tracking radar during autumn in central New York mdash Can J Zool 63p 755-761
Kjelleacuten N (1992) Differential timing of autumn migration between sex and age groups inraptors at Falsterbo Sweden mdash Orn Scan 23 p 420-434
Liechti F (1993) Naumlchtlicher Vogelzug im Herbst uumlber Suumlddeutschland Winddrift undKompensation mdash J Ornithol 134 p 373-404
mdash mdash Hedenstroumlm A amp Alerstam T (1994) Effects of sidewinds on optimal ight speedsof birds mdash J theor Biol 170 p 219-225
Lustick S Batterby B amp Kelty M (1978) Behavioural thermoregulation orientationtoward the sun in herring gulls mdash Sience 200 p 81-83
Pennycuick C (1975) Mechanics of ight mdash In Avian biology Vol 5 (DS Farner ampRJ King eds) Academic Press New York p 1-75
mdash mdash (1989) Bird ight performance a practical calculation manual mdash Oxford UniversityPress Oxford
mdash mdash (1997) Actual and lsquooptimumrsquo ight speeds eld data reassessedmdash J Exp Biol 200p 2355-2361
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 399
Richardson WJ (1978) Timing and amount of bird migration in relation to weather areview mdash Oikos 30 p 224-272
Spaar R (1995) Flight behaviour of steppe buzzard (Buteo buteo vulpinus) during springmigration in southern Israel a tracking radar study mdash Israel J Zool 41 p 489-500
mdash mdash (1996) Flight behaviour of migrating raptors in southern Israel mdash PhD thesisUniversity of Basel Verlag Schweizerische Vogelwarte Sempach
mdash mdash (1997) Flight strategies of migrating raptors a comparative study of interspecicvariation in ight characteristicsmdash Ibis 139 p 523-535
mdash mdash (1999) Flight behaviour of migrating raptors under varying environmental conditionsmdash In Proc 22 Int Ornithol Congr (N Adams amp R Slotow eds) University of NatalDurban p 1844-1862
mdash mdash amp Bruderer B (1997a) Optimal ight behaviour of soaring migrants A case studyof migrating steppe buzzards Buteo buteo vulpinus mdash Behav Ecol 8 p 288-297)
mdash mdash amp Bruderer B (1997b) Migration by apping or soaring ight strategies of marshMontagursquos and pallid harriers in southern Israel mdash Condor 99 p 458-469
mdash mdash Stark H amp Liechti F (1998) Migratory ight strategies of Levant sparrowhawkstime or energy minimization mdash Anim Behav 56 p 1185-1197
Stark H amp Liechti F (1993) Do Levant sparrowhawks Accipiter brevipes also migrate atnight mdash Ibis 135 p 233-236
Thiollay J-M (1977) Importance des populations de rapaces migrateurs en mediterraneeoccidentale mdash Alauda 45 p 115-121

388 MEYER SPAAR amp BRUDERER
Fig 5 Flight altitudes above ground (mean per track) in the course of the day (0 =sunrise 100 = sunset) Median 636 m for black kite 862 m for honey buzzard 910 m
for harriers 949 m for falcons and 951 m for osprey
Flight altitudes of honey buzzards black kites and harriers increased fromsunrise towards noon and were more or less stable thereafter (Fig 5) Falconsachieved high ight altitudes already soon after sunrise by apping-gliding ight In falcons ight altitudes differed between migratory routes birdscrossing the sea had higher ight altitudes than birds ying across the bayand along the coast (Fig 6 Kruskal-Wallis ANOVA H = 181 df =2 p = 00001 GT2-test crossing the sea versus ying across the bay(p lt 001) and versus along the coast (p lt 005)) In honey buzzards andharriers differences in ight altitudes between migratory routes showed thesame tendency but did not differ statistically (Fig 6)
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 389
Fig 6 Flight altitudes above ground (mean per track) for different migratory routes inhoney buzzard harriers and falcons Boxes are medians with 25-75 quartiles whiskersshow 10-90 range dots are outliers (ie outside the 15 box length from the upper and
lower value) GT2-test p lt 005 p lt 001
Fig 7 Mean climbing rates in thermal circling in the course of the day (0 = sunrise100 = sunset) Climbing rate did not differ between species groups (ANOVA F = 116
df = 4 N = 116 NS)
Flight characteristics in soaring-gliding
Thermal circling phases were recorded in 116 of 386 tracks and averageclimbing rate was 146 plusmn 084 ms (N = 116) Climbing rates increasedduring the morning hours (Fig 7 time of day lt 50 Spearman rankcorrelation N = 61 r = 034 p lt 001) Strong thermal convection aroundnoon allowed climbing rates of up to 45 ms From noon to sunset climbing
390 MEYER SPAAR amp BRUDERER
TA
BL
E2
Flig
htch
arac
teri
stic
sof
diffe
rent
spec
ies
grou
ps(m
s)
hone
ybu
zzar
dbl
ack
kite
harr
iers
falc
ons
ospr
eyF
Soar
ing
glid
ing
Cli
mbi
ngra
te1
35plusmn
077
(48)
171
plusmn0
97(2
2)1
54plusmn
087
(21)
134
plusmn0
84(2
4)1
86(1
)1
16N
SC
ross
-cou
ntry
grou
ndsp
eed
86
plusmn0
4(2
8)7
6plusmn
07
(7)
93
plusmn0
9(9
)9
3plusmn
10
(6)
89
(1)
079
NS
Cro
ss-c
ount
ryai
rspe
ed10
4plusmn
42
(28)
99
plusmn3
6(7
)9
3plusmn
25
(9)
95
plusmn3
1(1
0)11
3(1
)0
27N
SIn
ter-
ther
mal
grou
ndsp
eed
116
plusmn3
2(4
7)10
8plusmn
25
(21)
121
plusmn2
5(2
1)11
2plusmn
32
(23)
114
(1)
075
NS
Inte
r-th
erm
alai
rspe
ed12
8plusmn
37
(47)
134
plusmn3
0(2
1)12
0plusmn
30
(21)
127
plusmn3
6(2
3)17
0(1
)0
57N
S
Hor
izon
tal
appi
ng-g
lidi
ngi
ght
Gro
unds
peed
138
plusmn2
5(1
6)10
9plusmn
26
(9)
116
plusmn1
9(6
1)12
2plusmn
36
(68)
146
plusmn1
9(1
0)2
47
Air
spee
d11
1plusmn
23
(16)
103
plusmn2
3(9
)11
2plusmn
18
(58)
113
plusmn2
4(6
8)13
2plusmn
30
(10)
387
Mea
nplusmn
SD
(N)
Dif
fere
nces
wer
ete
sted
byA
NO
VA
S
igni
ca
nce
leve
l
plt
005
plt
001
H
oriz
onta
l
appi
ng-g
lidi
ng
ight
was
incl
uded
only
ifve
rtic
alsp
eeds
did
note
xcee
d0
plusmn0
5m
san
dif
noth
erm
alci
rcli
ngw
asob
serv
ed
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 391
rates did not decrease signi cantly (Spearman rank correlation N = 55r = shy 015 NS) Climbing rates in thermal circling below 1 ms occurredduring the whole day They did not differ between species groups (Table 2ANOVA F = 116 df = 3 N = 115 NS) Flight altitudes increased withincreasing climbing rates in birds lsquo ying along the coastrsquo (r = 049 N = 64p lt 00001) Average circling duration in completed soaring phases was106 plusmn 67 s (N = 80) and it was not related to climbing rate (r = 002N = 80 NS)
Mean airspeed and groundspeed in inter-thermal gliding did not differbetween the species groups (Table 2 ANOVA F = 057 and F = 075respectively df = 3 N = 112 NS) and means per species ranged between120-134 ms and 108-121 ms respectively Raptors increased airspeedsin inter-thermal gliding with increasing climbing rates (r = 052 N =113 p lt 00001) indicating that they maximised cross-country speeds insoaring-gliding ight (Pennycuick 1989 Spaar amp Bruderer 1997a) Meancross-country speeds in soaring-gliding did not differ between species groups(ANOVA cross-country airspeed F = 079 df = 3 N = 54 NS range 93-113 ms cross-country groundspeed F = 027 df = 3 N = 54 NS range76-93 ms)
Flight characteristics in horizontal apping-gliding
This ight style was considered if vertical speeds did not exceed 0 plusmn 05 msand no thermal circling was observed within a track Air- and groundspeedsdiffered between species groups (ANOVA airspeed F = 247 df = 4N = 161 p lt 005 groundspeed F = 387 df = 4 N = 164p lt 0005 Table 2) Ospreys had higher groundspeeds than harriers (GT2-test p lt 005) and higher ground- and airspeeds than black kites (p lt 005)Mean airspeeds per species ranged between 103-132 ms correspondinggroundspeeds between 109-146 ms
Flapping-gliding migrants reached signi cantly higher cross-country air-speeds (113 plusmn 22 ms N = 161) and groundspeeds (122 plusmn 31 ms N =164) than birds in soaring-gliding ight (airspeed 100 plusmn 36 ms N = 55Welch-test t = 249 df = 68 p lt 001 groundspeed 87 plusmn 25 msN = 55 t = 846 df = 114 p lt 00005)
In horizontal apping-gliding ight reactions to different wind con-ditions depended on migratory route raptors ying along the coast de-
392 MEYER SPAAR amp BRUDERER
Fig 8 Airspeed in horizontal apping-gliding ight versus tailwind- and sidewind-components for different migratory routes Linear regression (A) excluding one value inbrackets y = shy 0256x + 1128 R 2 = 0215 N = 56 p lt 0001 (B) r = shy 010 N = 57NS (C) r = shy 007 N = 85 NS (D) excluding two values in brackets y = 0191x+ 1072
R2 = 005 N = 84 p lt 005
creased airspeed with increasing tailwind-component (Fig 8A) but air-speed was not related to sidewind-component (Fig 8B) In contrast in rap-tors crossing the sea no relation was found between airspeed and tailwind-component (Fig 8C) but airspeed increased with increasing sidewind-component (Fig 8D)
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 393
Discussion
There are only few studies on the ight behaviour of migrating raptors cross-ing the sea mainly carried out at concentration points such as narrow wa-ter straits or peninsulas (Evans amp Lathbury 1972 Bernis 1975 Kerlinger1984) and none by tracking radar As these studies were limited to visualobservation mainly high altitudinal migration was often not detected (Ker-linger et al 1985) Our approach is different because (i) we observed raptormigration at an average coast site (ii) we used a tracking radar and (iii) wemeasured wind conditions at all ight levels
Migratory routes and in uence of environmental conditions
Although black kites and in larger numbers honey buzzards undertakelong sea-crossings in the Central Mediterranean (Beaman amp Galea 1974Agostini et al 1994) in our study no black kites and few honey buzzardswere observed attempting the sea crossing The majority of harriers falconsand ospreys crossed the sea Sea crossing honey buzzards occurred towardsthe end of the migratory season and were probably juveniles as in honeybuzzards adults migrate two weeks earlier than juveniles (Kjelleacuten 1992Agostini et al 1997) In contrast to the adults which often migrate in large ocks juveniles mainly migrate singly and have no experience about themigratory ight route This may lead to a higher proportion of juvenilescrossing the sea
Wind and thermal conditions affected migratory directions many harri-ers falcons and ospreys crossed the sea in favourable following and sidewinds but no sea crossings were observed in strong opposing winds This isreasonable because energy and time consumption for the sea crossing andtherefore the risk of exhaustion drastically increase in opposing winds How-ever tailwind-support did not fully explain the decisions on migratory routesbecause regularly birds followed the coast with a lower tailwind-supportcompared to crossing the sea These birds following the coast reached highclimbing rates in thermal circling and this might be compensating for thelower tailwind-support Under good thermal conditions birds were able tomigrate in soaring-gliding ight which consumes much less energy com-pared to apping-gliding ight when crossing the sea (see below Energyand time consumption) Anyway birds in bad shape for example those with
394 MEYER SPAAR amp BRUDERER
low fat reserves should follow the coast in order to minimise the risk ofstarvation to feed when ever it is possible and to y in soaring-gliding ight
At the same observation site migratory directions of nocturnal migrants ying below 2000 m above ground shift from 190deg to 250deg within the nightthe majority of the birds cross the sea when approaching the coast in the rsthalf of the night whereas they hesitate later on (Bruderer amp Liechti 1998)Thus the motivation to cross large barriers seems to decrease after somehours of migratory ight
Energy and time consumption
Energy consumption in apping ight disproportionately increases withbody mass while in soaring-gliding ight it is 2-3 times the basal metabolicrate (BMR) only (for herring gull Larus argentatus Baudinette amp Schmidt-Nielson 1974 Lustick et al 1978) In honey buzzards the average observedairspeed in apping-gliding ight was 111 ms and fat consumption perdistance (gkm) for an average honey buzzard (body mass in Table 3) atthis speed is about 6 times higher than in soaring-gliding ight (calculatedafter Pennycuick (1989 version 11) energy consumption in soaring-gliding ight = 2 times BMR body drag coef cient = 04) If the body dragcoef cient is set to 01 as recently proposed by Pennycuick (1997) energyconsumption in apping ight per km is reduced and energetic differencebetween soaring-gliding and apping ight become smaller but calculatedmaximum range speeds in apping ight are very high compared to ourobservations
We calculated energy and time consumption of different migratory routesbased on observed ight speeds for an average honey buzzard (Table 3) Notethat we did not take into account the time needed for refuelling consumedenergy (ie burned fat) and that we assumed zero wind Crossing the sea in apping-gliding ight (150 km distance) took on average about 4 hours andit was the time-minimising strategy for a honey buzzard (Table 3) Flyingalong the coast (250 km) required about 7 hours in soaring-gliding ightand 6 hours in apping-gliding ight Flying along the coast by soaring-gliding is the energy-minimising strategy for a honey buzzard and consumesabout 4 g fat (Table 3) Flapping-gliding ight along the coast is energeticallythe most expensive but never the fastest strategy Thus black kites andhoney buzzards migrated predominantly according to the energy minimising
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 395
TABLE 3 Observed airspeeds and corresponding estimated ight durationand fat consumption for an average honey buzzard when ying along the
coast or across the sea in different ight styles
Flight style Soaring-gliding Horizontal Horizontal apping-gliding apping-gliding
Flight route Flying along the coast Flying along the coast Crossing the seaDistance 250 km 250 km 150 km
Airspeed ms 101 (81-123) 108 (88-133) 108 (88-133)Flight duration hmin 652 (538-834) 626 (513-753) 351 (308-444)Fat consumption g 41 (34-55) 266 (230-298) 158 (138-179)for ight
Average honey buzzard = body mass 08 kg wing span 13 m wing area 025 m2 Medianand 25-75 range are given in brackets Calculations on fat consumption are based onPennycuickrsquos (1989) program of bird ight performance version 11 and birds are assumedto maximize cross-country speed in soaring-gliding ight (Spaar amp Bruderer 1997a)
strategy because a large proportion of the birds followed the coast in soaring-gliding ight
For species such as falcons or small harriers the difference in energyconsumption between apping and soaring-gliding ight is smaller Asthese species regularly use apping-gliding ight also over land or combinesoaring with apping-gliding ight differences in energy consumptionbetween land and sea route become even smaller and only short detours areenergetically favourable By combining soaring and apping-gliding ightsmaller raptors such as falcons or harriers are able to y under varyingenvironmental conditions to extend their migratory activity to times whenthermal updrafts are absent and to migrate even at night (Stark amp Liechti1993 Spaar amp Bruderer 1997b Spaar et al 1998)
Flight altitudes
Flight altitudes differed between the species groups partly due to differencesin migratory routes Birds crossing the sea ew considerably higher thanthose following the coast Kerlinger (1984) made similar observations atthe Delaware Bay birds crossing the bay ew at higher altitudes thanthose keeping on land He explained the higher ight altitudes by the bettervisibility of the other shore However the African coast is not visible fromour study site Some falcons even started crossing the sea after sunset
396 MEYER SPAAR amp BRUDERER
indicating that they continued migration into the night these birds had to y at least three hours until they reached the African coast The higher ightaltitudes of birds crossing the sea at our study site was mainly caused bytopographical features birds ying with southerly directions were forced toclimb higher up when crossing the mountains chains (1500 m height) about10-15 km northward of the coast and they afterwards maintained these ightlevels when approaching the sea
Flight altitudes of birds migrating along the coast were similar to altitudesobserved in North America and southern Israel with most raptors yingbelow 1000 m above ground and lt 5 above 1500 m (Kerlinger 19841989 Bruderer et al 1994 Spaar 1996)
Flight characteristics in soaring-gliding
Flight characteristics in soaring-gliding ight did not differ between thespecies groups (Table 2) As shown for raptor migration in Israel climb-ing rates do not depend primarily on morphological features but on environ-mental conditions (Spaar 1997) In general mean climbing rates in southernSpain were lower than in the semi-arid desert in Israel (Spaar 1997) Meanduration of the completed circling phases was shorter in Israel (88 plusmn 52 sSpaar 1995 1999) than in Spain In contrast to Israel (Spaar 1995 1997)circling time did not increase with increasing climbing rate Compared tothe semi-desert the development of thermal convection in southern Spainwas probably less stable and birds circled as long as possible in updraftsbecause they could not rely to encounter a next strong thermal within glid-ing distance This might also explain why inter-thermal gliding was oftensupported by apping it reduces sinking rate and prolongs gliding distance
In raptors migrating along the coast ight altitude and gliding airspeedswere related to thermal strength raptors circling in strong thermals hadhigher ight altitudes and higher inter-thermal gliding airspeeds than thosein weak thermals This is consistent with optimal ight theory for birds max-imising cross-country speed in soaring-gliding ight (Pennycuick 1989) asdemonstrated for steppe buzzards Buteo buteo vulpinus (Spaar amp Bruderer1997a)
Flight characteristics in horizontal apping-gliding
According to optimal ight theory maximum range speed decreases withfollowing winds and increases with head- and sidewinds (Pennycuick 1975
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 397
Liechti et al 1994) In raptors ying along the coast we observed decreas-ing airspeeds with increasing tailwind-support whereas birds crossing thesea did not reduce airspeeds Why did the birds react differently Sea cross-ings imply higher risks there is no possibility to land and weather (wind)conditions may change rapidly Higher airspeeds in following winds lead toshorter crossing times and lower the risk of unpredictable weather changesFurthermore drift by sidewinds may considerably extent crossing distancesand in order to reduce drift raptors increased airspeeds with increasingsindwind-components In contrast to sharp-shinned hawks Accipiter stria-tus avoiding crossings in strong lateral winds (Kerlinger 1985) we observedmany raptors to cross the sea in strong north-westerly (ie lateral) winds ifthey provided at least partial tailwind-support
Conclusions
Our observations at an average site at the southern coast of Spain showthat differences in water crossing behaviour are mainly species-speci cpresumably due to the costs for different ight strategies additionallyindividuals reacted to environmental factors such as wind and thermalconditions In medium sized raptors such as honey buzzard soaring-gliding ight along the coast is energy-minimising whereas crossing the sea istime-minimising Sea crossings occurred preferably with a good tailwind-support but also under strong sidewinds Raptors crossing the sea did notreduce airspeeds in following winds but increased airspeeds with increasingsidewinds They seemed to minimise the water crossing duration and theeffect of drift by sidewinds Under good thermal conditions birds regularlyfollowed the coast despite a lower tailwind-support compared to crossing thesea
References
Agostini N amp Logozzo D (1995) Autumn migration of honey buzzards in Southern Italymdash J Raptor Res 29 p 275-277
mdash mdash Logozzo D amp Colero C (1997) Migration of ocks of honey buzzards in SouthernItaly and Malta mdash J Raptor Res 31 p 84-86
mdash mdash Malara G Neri F Mollicone D amp Melotto S (1994) Flight strategies of honeybuzzards during spring migration across the Central Mediterranean mdash Avocetta 18p 73-76
398 MEYER SPAAR amp BRUDERER
Bannermann D-A amp Bannermann WM (1971) Handbook of the birds of Cyprus andmigrants of the Middle East mdash Oliver amp Boyd Edinburgh
Batschelet E (1981) Circular statistics in biology mdash Academic Press LondonBaudinette RV amp Schmidt-Nielsen K (1974) Energy costs of gliding ight in herring
gulls mdash Nature 248 p 83-84Beaman M amp Galea C (1974) The visible migration of raptors over the Maltese Islands
mdash Ibis 116 p 419-431Bernis F (1975) Migracion de Falconiformes y Ciconea spp por Gibraltar mdash IV Avance
sobre recuentos en el verano-otontildeo 1974 mdash Ardeola 21 p 581-592Bijlsma RG (1987) Bottleneck areas for migratory birds in the Mediterranean region mdash
ICBP Study report No 18 CambridgeBruderer B amp Liechti F (1998) Flight behaviour of nocturnally migrating birds in coastal
areas mdash crossing or coasting mdash J Avian Biol 29 p 499-510mdash mdash Blitzblau S amp Peter D (1994) Migration and ight behaviour of honey buzzards
Pernis apivorus in southern Israel observed by radar mdash Ardea 82 p 111-122mdash mdash Steuri T amp Baumgartner M (1995) Short-range high-precision surveillance of
nocturnal migration and tracking of single targets mdash Israel J Zool 41 p 207-220Evans PR amp Lathbury GW (1972) Raptor migration across the Strait of Gibraltar mdash Ibis
115 p 572-585Galea C amp Massa B (1985) Notes on the raptor migration across the central Mediter-
ranean mdash In Conservation studies on raptors (I Newton amp RD Chancellor eds)ICBP Technical Publication No 5 p 257-261
Gensboslashl B (1997) Greifvoumlgel mdash BLV VerlagsgesellschaftMuumlnchenGustin M amp Pizzari T (1998) Migratory pattern in the genus Circus sex and age
differential migration in Italy mdash Ornis Svecica 8 p 23-26Kerlinger P (1984) Flight behaviour of sharp-shinned hawks during migration 2 Over
water mdash Anim Beh 32 p 1029-1034mdash mdash (1985) Water-crossing behaviour of raptors during migration mdash Wilson Bull 97
p 109-113mdash mdash (1989) Flight strategies of migrating hawks mdash University of Chicago Press Chicagomdash mdash Bingman VP amp Able KP (1985) Comparative ight behaviour of migrating hawks
studied with tracking radar during autumn in central New York mdash Can J Zool 63p 755-761
Kjelleacuten N (1992) Differential timing of autumn migration between sex and age groups inraptors at Falsterbo Sweden mdash Orn Scan 23 p 420-434
Liechti F (1993) Naumlchtlicher Vogelzug im Herbst uumlber Suumlddeutschland Winddrift undKompensation mdash J Ornithol 134 p 373-404
mdash mdash Hedenstroumlm A amp Alerstam T (1994) Effects of sidewinds on optimal ight speedsof birds mdash J theor Biol 170 p 219-225
Lustick S Batterby B amp Kelty M (1978) Behavioural thermoregulation orientationtoward the sun in herring gulls mdash Sience 200 p 81-83
Pennycuick C (1975) Mechanics of ight mdash In Avian biology Vol 5 (DS Farner ampRJ King eds) Academic Press New York p 1-75
mdash mdash (1989) Bird ight performance a practical calculation manual mdash Oxford UniversityPress Oxford
mdash mdash (1997) Actual and lsquooptimumrsquo ight speeds eld data reassessedmdash J Exp Biol 200p 2355-2361
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 399
Richardson WJ (1978) Timing and amount of bird migration in relation to weather areview mdash Oikos 30 p 224-272
Spaar R (1995) Flight behaviour of steppe buzzard (Buteo buteo vulpinus) during springmigration in southern Israel a tracking radar study mdash Israel J Zool 41 p 489-500
mdash mdash (1996) Flight behaviour of migrating raptors in southern Israel mdash PhD thesisUniversity of Basel Verlag Schweizerische Vogelwarte Sempach
mdash mdash (1997) Flight strategies of migrating raptors a comparative study of interspecicvariation in ight characteristicsmdash Ibis 139 p 523-535
mdash mdash (1999) Flight behaviour of migrating raptors under varying environmental conditionsmdash In Proc 22 Int Ornithol Congr (N Adams amp R Slotow eds) University of NatalDurban p 1844-1862
mdash mdash amp Bruderer B (1997a) Optimal ight behaviour of soaring migrants A case studyof migrating steppe buzzards Buteo buteo vulpinus mdash Behav Ecol 8 p 288-297)
mdash mdash amp Bruderer B (1997b) Migration by apping or soaring ight strategies of marshMontagursquos and pallid harriers in southern Israel mdash Condor 99 p 458-469
mdash mdash Stark H amp Liechti F (1998) Migratory ight strategies of Levant sparrowhawkstime or energy minimization mdash Anim Behav 56 p 1185-1197
Stark H amp Liechti F (1993) Do Levant sparrowhawks Accipiter brevipes also migrate atnight mdash Ibis 135 p 233-236
Thiollay J-M (1977) Importance des populations de rapaces migrateurs en mediterraneeoccidentale mdash Alauda 45 p 115-121

MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 389
Fig 6 Flight altitudes above ground (mean per track) for different migratory routes inhoney buzzard harriers and falcons Boxes are medians with 25-75 quartiles whiskersshow 10-90 range dots are outliers (ie outside the 15 box length from the upper and
lower value) GT2-test p lt 005 p lt 001
Fig 7 Mean climbing rates in thermal circling in the course of the day (0 = sunrise100 = sunset) Climbing rate did not differ between species groups (ANOVA F = 116
df = 4 N = 116 NS)
Flight characteristics in soaring-gliding
Thermal circling phases were recorded in 116 of 386 tracks and averageclimbing rate was 146 plusmn 084 ms (N = 116) Climbing rates increasedduring the morning hours (Fig 7 time of day lt 50 Spearman rankcorrelation N = 61 r = 034 p lt 001) Strong thermal convection aroundnoon allowed climbing rates of up to 45 ms From noon to sunset climbing
390 MEYER SPAAR amp BRUDERER
TA
BL
E2
Flig
htch
arac
teri
stic
sof
diffe
rent
spec
ies
grou
ps(m
s)
hone
ybu
zzar
dbl
ack
kite
harr
iers
falc
ons
ospr
eyF
Soar
ing
glid
ing
Cli
mbi
ngra
te1
35plusmn
077
(48)
171
plusmn0
97(2
2)1
54plusmn
087
(21)
134
plusmn0
84(2
4)1
86(1
)1
16N
SC
ross
-cou
ntry
grou
ndsp
eed
86
plusmn0
4(2
8)7
6plusmn
07
(7)
93
plusmn0
9(9
)9
3plusmn
10
(6)
89
(1)
079
NS
Cro
ss-c
ount
ryai
rspe
ed10
4plusmn
42
(28)
99
plusmn3
6(7
)9
3plusmn
25
(9)
95
plusmn3
1(1
0)11
3(1
)0
27N
SIn
ter-
ther
mal
grou
ndsp
eed
116
plusmn3
2(4
7)10
8plusmn
25
(21)
121
plusmn2
5(2
1)11
2plusmn
32
(23)
114
(1)
075
NS
Inte
r-th
erm
alai
rspe
ed12
8plusmn
37
(47)
134
plusmn3
0(2
1)12
0plusmn
30
(21)
127
plusmn3
6(2
3)17
0(1
)0
57N
S
Hor
izon
tal
appi
ng-g
lidi
ngi
ght
Gro
unds
peed
138
plusmn2
5(1
6)10
9plusmn
26
(9)
116
plusmn1
9(6
1)12
2plusmn
36
(68)
146
plusmn1
9(1
0)2
47
Air
spee
d11
1plusmn
23
(16)
103
plusmn2
3(9
)11
2plusmn
18
(58)
113
plusmn2
4(6
8)13
2plusmn
30
(10)
387
Mea
nplusmn
SD
(N)
Dif
fere
nces
wer
ete
sted
byA
NO
VA
S
igni
ca
nce
leve
l
plt
005
plt
001
H
oriz
onta
l
appi
ng-g
lidi
ng
ight
was
incl
uded
only
ifve
rtic
alsp
eeds
did
note
xcee
d0
plusmn0
5m
san
dif
noth
erm
alci
rcli
ngw
asob
serv
ed
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 391
rates did not decrease signi cantly (Spearman rank correlation N = 55r = shy 015 NS) Climbing rates in thermal circling below 1 ms occurredduring the whole day They did not differ between species groups (Table 2ANOVA F = 116 df = 3 N = 115 NS) Flight altitudes increased withincreasing climbing rates in birds lsquo ying along the coastrsquo (r = 049 N = 64p lt 00001) Average circling duration in completed soaring phases was106 plusmn 67 s (N = 80) and it was not related to climbing rate (r = 002N = 80 NS)
Mean airspeed and groundspeed in inter-thermal gliding did not differbetween the species groups (Table 2 ANOVA F = 057 and F = 075respectively df = 3 N = 112 NS) and means per species ranged between120-134 ms and 108-121 ms respectively Raptors increased airspeedsin inter-thermal gliding with increasing climbing rates (r = 052 N =113 p lt 00001) indicating that they maximised cross-country speeds insoaring-gliding ight (Pennycuick 1989 Spaar amp Bruderer 1997a) Meancross-country speeds in soaring-gliding did not differ between species groups(ANOVA cross-country airspeed F = 079 df = 3 N = 54 NS range 93-113 ms cross-country groundspeed F = 027 df = 3 N = 54 NS range76-93 ms)
Flight characteristics in horizontal apping-gliding
This ight style was considered if vertical speeds did not exceed 0 plusmn 05 msand no thermal circling was observed within a track Air- and groundspeedsdiffered between species groups (ANOVA airspeed F = 247 df = 4N = 161 p lt 005 groundspeed F = 387 df = 4 N = 164p lt 0005 Table 2) Ospreys had higher groundspeeds than harriers (GT2-test p lt 005) and higher ground- and airspeeds than black kites (p lt 005)Mean airspeeds per species ranged between 103-132 ms correspondinggroundspeeds between 109-146 ms
Flapping-gliding migrants reached signi cantly higher cross-country air-speeds (113 plusmn 22 ms N = 161) and groundspeeds (122 plusmn 31 ms N =164) than birds in soaring-gliding ight (airspeed 100 plusmn 36 ms N = 55Welch-test t = 249 df = 68 p lt 001 groundspeed 87 plusmn 25 msN = 55 t = 846 df = 114 p lt 00005)
In horizontal apping-gliding ight reactions to different wind con-ditions depended on migratory route raptors ying along the coast de-
392 MEYER SPAAR amp BRUDERER
Fig 8 Airspeed in horizontal apping-gliding ight versus tailwind- and sidewind-components for different migratory routes Linear regression (A) excluding one value inbrackets y = shy 0256x + 1128 R 2 = 0215 N = 56 p lt 0001 (B) r = shy 010 N = 57NS (C) r = shy 007 N = 85 NS (D) excluding two values in brackets y = 0191x+ 1072
R2 = 005 N = 84 p lt 005
creased airspeed with increasing tailwind-component (Fig 8A) but air-speed was not related to sidewind-component (Fig 8B) In contrast in rap-tors crossing the sea no relation was found between airspeed and tailwind-component (Fig 8C) but airspeed increased with increasing sidewind-component (Fig 8D)
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 393
Discussion
There are only few studies on the ight behaviour of migrating raptors cross-ing the sea mainly carried out at concentration points such as narrow wa-ter straits or peninsulas (Evans amp Lathbury 1972 Bernis 1975 Kerlinger1984) and none by tracking radar As these studies were limited to visualobservation mainly high altitudinal migration was often not detected (Ker-linger et al 1985) Our approach is different because (i) we observed raptormigration at an average coast site (ii) we used a tracking radar and (iii) wemeasured wind conditions at all ight levels
Migratory routes and in uence of environmental conditions
Although black kites and in larger numbers honey buzzards undertakelong sea-crossings in the Central Mediterranean (Beaman amp Galea 1974Agostini et al 1994) in our study no black kites and few honey buzzardswere observed attempting the sea crossing The majority of harriers falconsand ospreys crossed the sea Sea crossing honey buzzards occurred towardsthe end of the migratory season and were probably juveniles as in honeybuzzards adults migrate two weeks earlier than juveniles (Kjelleacuten 1992Agostini et al 1997) In contrast to the adults which often migrate in large ocks juveniles mainly migrate singly and have no experience about themigratory ight route This may lead to a higher proportion of juvenilescrossing the sea
Wind and thermal conditions affected migratory directions many harri-ers falcons and ospreys crossed the sea in favourable following and sidewinds but no sea crossings were observed in strong opposing winds This isreasonable because energy and time consumption for the sea crossing andtherefore the risk of exhaustion drastically increase in opposing winds How-ever tailwind-support did not fully explain the decisions on migratory routesbecause regularly birds followed the coast with a lower tailwind-supportcompared to crossing the sea These birds following the coast reached highclimbing rates in thermal circling and this might be compensating for thelower tailwind-support Under good thermal conditions birds were able tomigrate in soaring-gliding ight which consumes much less energy com-pared to apping-gliding ight when crossing the sea (see below Energyand time consumption) Anyway birds in bad shape for example those with
394 MEYER SPAAR amp BRUDERER
low fat reserves should follow the coast in order to minimise the risk ofstarvation to feed when ever it is possible and to y in soaring-gliding ight
At the same observation site migratory directions of nocturnal migrants ying below 2000 m above ground shift from 190deg to 250deg within the nightthe majority of the birds cross the sea when approaching the coast in the rsthalf of the night whereas they hesitate later on (Bruderer amp Liechti 1998)Thus the motivation to cross large barriers seems to decrease after somehours of migratory ight
Energy and time consumption
Energy consumption in apping ight disproportionately increases withbody mass while in soaring-gliding ight it is 2-3 times the basal metabolicrate (BMR) only (for herring gull Larus argentatus Baudinette amp Schmidt-Nielson 1974 Lustick et al 1978) In honey buzzards the average observedairspeed in apping-gliding ight was 111 ms and fat consumption perdistance (gkm) for an average honey buzzard (body mass in Table 3) atthis speed is about 6 times higher than in soaring-gliding ight (calculatedafter Pennycuick (1989 version 11) energy consumption in soaring-gliding ight = 2 times BMR body drag coef cient = 04) If the body dragcoef cient is set to 01 as recently proposed by Pennycuick (1997) energyconsumption in apping ight per km is reduced and energetic differencebetween soaring-gliding and apping ight become smaller but calculatedmaximum range speeds in apping ight are very high compared to ourobservations
We calculated energy and time consumption of different migratory routesbased on observed ight speeds for an average honey buzzard (Table 3) Notethat we did not take into account the time needed for refuelling consumedenergy (ie burned fat) and that we assumed zero wind Crossing the sea in apping-gliding ight (150 km distance) took on average about 4 hours andit was the time-minimising strategy for a honey buzzard (Table 3) Flyingalong the coast (250 km) required about 7 hours in soaring-gliding ightand 6 hours in apping-gliding ight Flying along the coast by soaring-gliding is the energy-minimising strategy for a honey buzzard and consumesabout 4 g fat (Table 3) Flapping-gliding ight along the coast is energeticallythe most expensive but never the fastest strategy Thus black kites andhoney buzzards migrated predominantly according to the energy minimising
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 395
TABLE 3 Observed airspeeds and corresponding estimated ight durationand fat consumption for an average honey buzzard when ying along the
coast or across the sea in different ight styles
Flight style Soaring-gliding Horizontal Horizontal apping-gliding apping-gliding
Flight route Flying along the coast Flying along the coast Crossing the seaDistance 250 km 250 km 150 km
Airspeed ms 101 (81-123) 108 (88-133) 108 (88-133)Flight duration hmin 652 (538-834) 626 (513-753) 351 (308-444)Fat consumption g 41 (34-55) 266 (230-298) 158 (138-179)for ight
Average honey buzzard = body mass 08 kg wing span 13 m wing area 025 m2 Medianand 25-75 range are given in brackets Calculations on fat consumption are based onPennycuickrsquos (1989) program of bird ight performance version 11 and birds are assumedto maximize cross-country speed in soaring-gliding ight (Spaar amp Bruderer 1997a)
strategy because a large proportion of the birds followed the coast in soaring-gliding ight
For species such as falcons or small harriers the difference in energyconsumption between apping and soaring-gliding ight is smaller Asthese species regularly use apping-gliding ight also over land or combinesoaring with apping-gliding ight differences in energy consumptionbetween land and sea route become even smaller and only short detours areenergetically favourable By combining soaring and apping-gliding ightsmaller raptors such as falcons or harriers are able to y under varyingenvironmental conditions to extend their migratory activity to times whenthermal updrafts are absent and to migrate even at night (Stark amp Liechti1993 Spaar amp Bruderer 1997b Spaar et al 1998)
Flight altitudes
Flight altitudes differed between the species groups partly due to differencesin migratory routes Birds crossing the sea ew considerably higher thanthose following the coast Kerlinger (1984) made similar observations atthe Delaware Bay birds crossing the bay ew at higher altitudes thanthose keeping on land He explained the higher ight altitudes by the bettervisibility of the other shore However the African coast is not visible fromour study site Some falcons even started crossing the sea after sunset
396 MEYER SPAAR amp BRUDERER
indicating that they continued migration into the night these birds had to y at least three hours until they reached the African coast The higher ightaltitudes of birds crossing the sea at our study site was mainly caused bytopographical features birds ying with southerly directions were forced toclimb higher up when crossing the mountains chains (1500 m height) about10-15 km northward of the coast and they afterwards maintained these ightlevels when approaching the sea
Flight altitudes of birds migrating along the coast were similar to altitudesobserved in North America and southern Israel with most raptors yingbelow 1000 m above ground and lt 5 above 1500 m (Kerlinger 19841989 Bruderer et al 1994 Spaar 1996)
Flight characteristics in soaring-gliding
Flight characteristics in soaring-gliding ight did not differ between thespecies groups (Table 2) As shown for raptor migration in Israel climb-ing rates do not depend primarily on morphological features but on environ-mental conditions (Spaar 1997) In general mean climbing rates in southernSpain were lower than in the semi-arid desert in Israel (Spaar 1997) Meanduration of the completed circling phases was shorter in Israel (88 plusmn 52 sSpaar 1995 1999) than in Spain In contrast to Israel (Spaar 1995 1997)circling time did not increase with increasing climbing rate Compared tothe semi-desert the development of thermal convection in southern Spainwas probably less stable and birds circled as long as possible in updraftsbecause they could not rely to encounter a next strong thermal within glid-ing distance This might also explain why inter-thermal gliding was oftensupported by apping it reduces sinking rate and prolongs gliding distance
In raptors migrating along the coast ight altitude and gliding airspeedswere related to thermal strength raptors circling in strong thermals hadhigher ight altitudes and higher inter-thermal gliding airspeeds than thosein weak thermals This is consistent with optimal ight theory for birds max-imising cross-country speed in soaring-gliding ight (Pennycuick 1989) asdemonstrated for steppe buzzards Buteo buteo vulpinus (Spaar amp Bruderer1997a)
Flight characteristics in horizontal apping-gliding
According to optimal ight theory maximum range speed decreases withfollowing winds and increases with head- and sidewinds (Pennycuick 1975
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 397
Liechti et al 1994) In raptors ying along the coast we observed decreas-ing airspeeds with increasing tailwind-support whereas birds crossing thesea did not reduce airspeeds Why did the birds react differently Sea cross-ings imply higher risks there is no possibility to land and weather (wind)conditions may change rapidly Higher airspeeds in following winds lead toshorter crossing times and lower the risk of unpredictable weather changesFurthermore drift by sidewinds may considerably extent crossing distancesand in order to reduce drift raptors increased airspeeds with increasingsindwind-components In contrast to sharp-shinned hawks Accipiter stria-tus avoiding crossings in strong lateral winds (Kerlinger 1985) we observedmany raptors to cross the sea in strong north-westerly (ie lateral) winds ifthey provided at least partial tailwind-support
Conclusions
Our observations at an average site at the southern coast of Spain showthat differences in water crossing behaviour are mainly species-speci cpresumably due to the costs for different ight strategies additionallyindividuals reacted to environmental factors such as wind and thermalconditions In medium sized raptors such as honey buzzard soaring-gliding ight along the coast is energy-minimising whereas crossing the sea istime-minimising Sea crossings occurred preferably with a good tailwind-support but also under strong sidewinds Raptors crossing the sea did notreduce airspeeds in following winds but increased airspeeds with increasingsidewinds They seemed to minimise the water crossing duration and theeffect of drift by sidewinds Under good thermal conditions birds regularlyfollowed the coast despite a lower tailwind-support compared to crossing thesea
References
Agostini N amp Logozzo D (1995) Autumn migration of honey buzzards in Southern Italymdash J Raptor Res 29 p 275-277
mdash mdash Logozzo D amp Colero C (1997) Migration of ocks of honey buzzards in SouthernItaly and Malta mdash J Raptor Res 31 p 84-86
mdash mdash Malara G Neri F Mollicone D amp Melotto S (1994) Flight strategies of honeybuzzards during spring migration across the Central Mediterranean mdash Avocetta 18p 73-76
398 MEYER SPAAR amp BRUDERER
Bannermann D-A amp Bannermann WM (1971) Handbook of the birds of Cyprus andmigrants of the Middle East mdash Oliver amp Boyd Edinburgh
Batschelet E (1981) Circular statistics in biology mdash Academic Press LondonBaudinette RV amp Schmidt-Nielsen K (1974) Energy costs of gliding ight in herring
gulls mdash Nature 248 p 83-84Beaman M amp Galea C (1974) The visible migration of raptors over the Maltese Islands
mdash Ibis 116 p 419-431Bernis F (1975) Migracion de Falconiformes y Ciconea spp por Gibraltar mdash IV Avance
sobre recuentos en el verano-otontildeo 1974 mdash Ardeola 21 p 581-592Bijlsma RG (1987) Bottleneck areas for migratory birds in the Mediterranean region mdash
ICBP Study report No 18 CambridgeBruderer B amp Liechti F (1998) Flight behaviour of nocturnally migrating birds in coastal
areas mdash crossing or coasting mdash J Avian Biol 29 p 499-510mdash mdash Blitzblau S amp Peter D (1994) Migration and ight behaviour of honey buzzards
Pernis apivorus in southern Israel observed by radar mdash Ardea 82 p 111-122mdash mdash Steuri T amp Baumgartner M (1995) Short-range high-precision surveillance of
nocturnal migration and tracking of single targets mdash Israel J Zool 41 p 207-220Evans PR amp Lathbury GW (1972) Raptor migration across the Strait of Gibraltar mdash Ibis
115 p 572-585Galea C amp Massa B (1985) Notes on the raptor migration across the central Mediter-
ranean mdash In Conservation studies on raptors (I Newton amp RD Chancellor eds)ICBP Technical Publication No 5 p 257-261
Gensboslashl B (1997) Greifvoumlgel mdash BLV VerlagsgesellschaftMuumlnchenGustin M amp Pizzari T (1998) Migratory pattern in the genus Circus sex and age
differential migration in Italy mdash Ornis Svecica 8 p 23-26Kerlinger P (1984) Flight behaviour of sharp-shinned hawks during migration 2 Over
water mdash Anim Beh 32 p 1029-1034mdash mdash (1985) Water-crossing behaviour of raptors during migration mdash Wilson Bull 97
p 109-113mdash mdash (1989) Flight strategies of migrating hawks mdash University of Chicago Press Chicagomdash mdash Bingman VP amp Able KP (1985) Comparative ight behaviour of migrating hawks
studied with tracking radar during autumn in central New York mdash Can J Zool 63p 755-761
Kjelleacuten N (1992) Differential timing of autumn migration between sex and age groups inraptors at Falsterbo Sweden mdash Orn Scan 23 p 420-434
Liechti F (1993) Naumlchtlicher Vogelzug im Herbst uumlber Suumlddeutschland Winddrift undKompensation mdash J Ornithol 134 p 373-404
mdash mdash Hedenstroumlm A amp Alerstam T (1994) Effects of sidewinds on optimal ight speedsof birds mdash J theor Biol 170 p 219-225
Lustick S Batterby B amp Kelty M (1978) Behavioural thermoregulation orientationtoward the sun in herring gulls mdash Sience 200 p 81-83
Pennycuick C (1975) Mechanics of ight mdash In Avian biology Vol 5 (DS Farner ampRJ King eds) Academic Press New York p 1-75
mdash mdash (1989) Bird ight performance a practical calculation manual mdash Oxford UniversityPress Oxford
mdash mdash (1997) Actual and lsquooptimumrsquo ight speeds eld data reassessedmdash J Exp Biol 200p 2355-2361
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 399
Richardson WJ (1978) Timing and amount of bird migration in relation to weather areview mdash Oikos 30 p 224-272
Spaar R (1995) Flight behaviour of steppe buzzard (Buteo buteo vulpinus) during springmigration in southern Israel a tracking radar study mdash Israel J Zool 41 p 489-500
mdash mdash (1996) Flight behaviour of migrating raptors in southern Israel mdash PhD thesisUniversity of Basel Verlag Schweizerische Vogelwarte Sempach
mdash mdash (1997) Flight strategies of migrating raptors a comparative study of interspecicvariation in ight characteristicsmdash Ibis 139 p 523-535
mdash mdash (1999) Flight behaviour of migrating raptors under varying environmental conditionsmdash In Proc 22 Int Ornithol Congr (N Adams amp R Slotow eds) University of NatalDurban p 1844-1862
mdash mdash amp Bruderer B (1997a) Optimal ight behaviour of soaring migrants A case studyof migrating steppe buzzards Buteo buteo vulpinus mdash Behav Ecol 8 p 288-297)
mdash mdash amp Bruderer B (1997b) Migration by apping or soaring ight strategies of marshMontagursquos and pallid harriers in southern Israel mdash Condor 99 p 458-469
mdash mdash Stark H amp Liechti F (1998) Migratory ight strategies of Levant sparrowhawkstime or energy minimization mdash Anim Behav 56 p 1185-1197
Stark H amp Liechti F (1993) Do Levant sparrowhawks Accipiter brevipes also migrate atnight mdash Ibis 135 p 233-236
Thiollay J-M (1977) Importance des populations de rapaces migrateurs en mediterraneeoccidentale mdash Alauda 45 p 115-121

390 MEYER SPAAR amp BRUDERER
TA
BL
E2
Flig
htch
arac
teri
stic
sof
diffe
rent
spec
ies
grou
ps(m
s)
hone
ybu
zzar
dbl
ack
kite
harr
iers
falc
ons
ospr
eyF
Soar
ing
glid
ing
Cli
mbi
ngra
te1
35plusmn
077
(48)
171
plusmn0
97(2
2)1
54plusmn
087
(21)
134
plusmn0
84(2
4)1
86(1
)1
16N
SC
ross
-cou
ntry
grou
ndsp
eed
86
plusmn0
4(2
8)7
6plusmn
07
(7)
93
plusmn0
9(9
)9
3plusmn
10
(6)
89
(1)
079
NS
Cro
ss-c
ount
ryai
rspe
ed10
4plusmn
42
(28)
99
plusmn3
6(7
)9
3plusmn
25
(9)
95
plusmn3
1(1
0)11
3(1
)0
27N
SIn
ter-
ther
mal
grou
ndsp
eed
116
plusmn3
2(4
7)10
8plusmn
25
(21)
121
plusmn2
5(2
1)11
2plusmn
32
(23)
114
(1)
075
NS
Inte
r-th
erm
alai
rspe
ed12
8plusmn
37
(47)
134
plusmn3
0(2
1)12
0plusmn
30
(21)
127
plusmn3
6(2
3)17
0(1
)0
57N
S
Hor
izon
tal
appi
ng-g
lidi
ngi
ght
Gro
unds
peed
138
plusmn2
5(1
6)10
9plusmn
26
(9)
116
plusmn1
9(6
1)12
2plusmn
36
(68)
146
plusmn1
9(1
0)2
47
Air
spee
d11
1plusmn
23
(16)
103
plusmn2
3(9
)11
2plusmn
18
(58)
113
plusmn2
4(6
8)13
2plusmn
30
(10)
387
Mea
nplusmn
SD
(N)
Dif
fere
nces
wer
ete
sted
byA
NO
VA
S
igni
ca
nce
leve
l
plt
005
plt
001
H
oriz
onta
l
appi
ng-g
lidi
ng
ight
was
incl
uded
only
ifve
rtic
alsp
eeds
did
note
xcee
d0
plusmn0
5m
san
dif
noth
erm
alci
rcli
ngw
asob
serv
ed
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 391
rates did not decrease signi cantly (Spearman rank correlation N = 55r = shy 015 NS) Climbing rates in thermal circling below 1 ms occurredduring the whole day They did not differ between species groups (Table 2ANOVA F = 116 df = 3 N = 115 NS) Flight altitudes increased withincreasing climbing rates in birds lsquo ying along the coastrsquo (r = 049 N = 64p lt 00001) Average circling duration in completed soaring phases was106 plusmn 67 s (N = 80) and it was not related to climbing rate (r = 002N = 80 NS)
Mean airspeed and groundspeed in inter-thermal gliding did not differbetween the species groups (Table 2 ANOVA F = 057 and F = 075respectively df = 3 N = 112 NS) and means per species ranged between120-134 ms and 108-121 ms respectively Raptors increased airspeedsin inter-thermal gliding with increasing climbing rates (r = 052 N =113 p lt 00001) indicating that they maximised cross-country speeds insoaring-gliding ight (Pennycuick 1989 Spaar amp Bruderer 1997a) Meancross-country speeds in soaring-gliding did not differ between species groups(ANOVA cross-country airspeed F = 079 df = 3 N = 54 NS range 93-113 ms cross-country groundspeed F = 027 df = 3 N = 54 NS range76-93 ms)
Flight characteristics in horizontal apping-gliding
This ight style was considered if vertical speeds did not exceed 0 plusmn 05 msand no thermal circling was observed within a track Air- and groundspeedsdiffered between species groups (ANOVA airspeed F = 247 df = 4N = 161 p lt 005 groundspeed F = 387 df = 4 N = 164p lt 0005 Table 2) Ospreys had higher groundspeeds than harriers (GT2-test p lt 005) and higher ground- and airspeeds than black kites (p lt 005)Mean airspeeds per species ranged between 103-132 ms correspondinggroundspeeds between 109-146 ms
Flapping-gliding migrants reached signi cantly higher cross-country air-speeds (113 plusmn 22 ms N = 161) and groundspeeds (122 plusmn 31 ms N =164) than birds in soaring-gliding ight (airspeed 100 plusmn 36 ms N = 55Welch-test t = 249 df = 68 p lt 001 groundspeed 87 plusmn 25 msN = 55 t = 846 df = 114 p lt 00005)
In horizontal apping-gliding ight reactions to different wind con-ditions depended on migratory route raptors ying along the coast de-
392 MEYER SPAAR amp BRUDERER
Fig 8 Airspeed in horizontal apping-gliding ight versus tailwind- and sidewind-components for different migratory routes Linear regression (A) excluding one value inbrackets y = shy 0256x + 1128 R 2 = 0215 N = 56 p lt 0001 (B) r = shy 010 N = 57NS (C) r = shy 007 N = 85 NS (D) excluding two values in brackets y = 0191x+ 1072
R2 = 005 N = 84 p lt 005
creased airspeed with increasing tailwind-component (Fig 8A) but air-speed was not related to sidewind-component (Fig 8B) In contrast in rap-tors crossing the sea no relation was found between airspeed and tailwind-component (Fig 8C) but airspeed increased with increasing sidewind-component (Fig 8D)
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 393
Discussion
There are only few studies on the ight behaviour of migrating raptors cross-ing the sea mainly carried out at concentration points such as narrow wa-ter straits or peninsulas (Evans amp Lathbury 1972 Bernis 1975 Kerlinger1984) and none by tracking radar As these studies were limited to visualobservation mainly high altitudinal migration was often not detected (Ker-linger et al 1985) Our approach is different because (i) we observed raptormigration at an average coast site (ii) we used a tracking radar and (iii) wemeasured wind conditions at all ight levels
Migratory routes and in uence of environmental conditions
Although black kites and in larger numbers honey buzzards undertakelong sea-crossings in the Central Mediterranean (Beaman amp Galea 1974Agostini et al 1994) in our study no black kites and few honey buzzardswere observed attempting the sea crossing The majority of harriers falconsand ospreys crossed the sea Sea crossing honey buzzards occurred towardsthe end of the migratory season and were probably juveniles as in honeybuzzards adults migrate two weeks earlier than juveniles (Kjelleacuten 1992Agostini et al 1997) In contrast to the adults which often migrate in large ocks juveniles mainly migrate singly and have no experience about themigratory ight route This may lead to a higher proportion of juvenilescrossing the sea
Wind and thermal conditions affected migratory directions many harri-ers falcons and ospreys crossed the sea in favourable following and sidewinds but no sea crossings were observed in strong opposing winds This isreasonable because energy and time consumption for the sea crossing andtherefore the risk of exhaustion drastically increase in opposing winds How-ever tailwind-support did not fully explain the decisions on migratory routesbecause regularly birds followed the coast with a lower tailwind-supportcompared to crossing the sea These birds following the coast reached highclimbing rates in thermal circling and this might be compensating for thelower tailwind-support Under good thermal conditions birds were able tomigrate in soaring-gliding ight which consumes much less energy com-pared to apping-gliding ight when crossing the sea (see below Energyand time consumption) Anyway birds in bad shape for example those with
394 MEYER SPAAR amp BRUDERER
low fat reserves should follow the coast in order to minimise the risk ofstarvation to feed when ever it is possible and to y in soaring-gliding ight
At the same observation site migratory directions of nocturnal migrants ying below 2000 m above ground shift from 190deg to 250deg within the nightthe majority of the birds cross the sea when approaching the coast in the rsthalf of the night whereas they hesitate later on (Bruderer amp Liechti 1998)Thus the motivation to cross large barriers seems to decrease after somehours of migratory ight
Energy and time consumption
Energy consumption in apping ight disproportionately increases withbody mass while in soaring-gliding ight it is 2-3 times the basal metabolicrate (BMR) only (for herring gull Larus argentatus Baudinette amp Schmidt-Nielson 1974 Lustick et al 1978) In honey buzzards the average observedairspeed in apping-gliding ight was 111 ms and fat consumption perdistance (gkm) for an average honey buzzard (body mass in Table 3) atthis speed is about 6 times higher than in soaring-gliding ight (calculatedafter Pennycuick (1989 version 11) energy consumption in soaring-gliding ight = 2 times BMR body drag coef cient = 04) If the body dragcoef cient is set to 01 as recently proposed by Pennycuick (1997) energyconsumption in apping ight per km is reduced and energetic differencebetween soaring-gliding and apping ight become smaller but calculatedmaximum range speeds in apping ight are very high compared to ourobservations
We calculated energy and time consumption of different migratory routesbased on observed ight speeds for an average honey buzzard (Table 3) Notethat we did not take into account the time needed for refuelling consumedenergy (ie burned fat) and that we assumed zero wind Crossing the sea in apping-gliding ight (150 km distance) took on average about 4 hours andit was the time-minimising strategy for a honey buzzard (Table 3) Flyingalong the coast (250 km) required about 7 hours in soaring-gliding ightand 6 hours in apping-gliding ight Flying along the coast by soaring-gliding is the energy-minimising strategy for a honey buzzard and consumesabout 4 g fat (Table 3) Flapping-gliding ight along the coast is energeticallythe most expensive but never the fastest strategy Thus black kites andhoney buzzards migrated predominantly according to the energy minimising
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 395
TABLE 3 Observed airspeeds and corresponding estimated ight durationand fat consumption for an average honey buzzard when ying along the
coast or across the sea in different ight styles
Flight style Soaring-gliding Horizontal Horizontal apping-gliding apping-gliding
Flight route Flying along the coast Flying along the coast Crossing the seaDistance 250 km 250 km 150 km
Airspeed ms 101 (81-123) 108 (88-133) 108 (88-133)Flight duration hmin 652 (538-834) 626 (513-753) 351 (308-444)Fat consumption g 41 (34-55) 266 (230-298) 158 (138-179)for ight
Average honey buzzard = body mass 08 kg wing span 13 m wing area 025 m2 Medianand 25-75 range are given in brackets Calculations on fat consumption are based onPennycuickrsquos (1989) program of bird ight performance version 11 and birds are assumedto maximize cross-country speed in soaring-gliding ight (Spaar amp Bruderer 1997a)
strategy because a large proportion of the birds followed the coast in soaring-gliding ight
For species such as falcons or small harriers the difference in energyconsumption between apping and soaring-gliding ight is smaller Asthese species regularly use apping-gliding ight also over land or combinesoaring with apping-gliding ight differences in energy consumptionbetween land and sea route become even smaller and only short detours areenergetically favourable By combining soaring and apping-gliding ightsmaller raptors such as falcons or harriers are able to y under varyingenvironmental conditions to extend their migratory activity to times whenthermal updrafts are absent and to migrate even at night (Stark amp Liechti1993 Spaar amp Bruderer 1997b Spaar et al 1998)
Flight altitudes
Flight altitudes differed between the species groups partly due to differencesin migratory routes Birds crossing the sea ew considerably higher thanthose following the coast Kerlinger (1984) made similar observations atthe Delaware Bay birds crossing the bay ew at higher altitudes thanthose keeping on land He explained the higher ight altitudes by the bettervisibility of the other shore However the African coast is not visible fromour study site Some falcons even started crossing the sea after sunset
396 MEYER SPAAR amp BRUDERER
indicating that they continued migration into the night these birds had to y at least three hours until they reached the African coast The higher ightaltitudes of birds crossing the sea at our study site was mainly caused bytopographical features birds ying with southerly directions were forced toclimb higher up when crossing the mountains chains (1500 m height) about10-15 km northward of the coast and they afterwards maintained these ightlevels when approaching the sea
Flight altitudes of birds migrating along the coast were similar to altitudesobserved in North America and southern Israel with most raptors yingbelow 1000 m above ground and lt 5 above 1500 m (Kerlinger 19841989 Bruderer et al 1994 Spaar 1996)
Flight characteristics in soaring-gliding
Flight characteristics in soaring-gliding ight did not differ between thespecies groups (Table 2) As shown for raptor migration in Israel climb-ing rates do not depend primarily on morphological features but on environ-mental conditions (Spaar 1997) In general mean climbing rates in southernSpain were lower than in the semi-arid desert in Israel (Spaar 1997) Meanduration of the completed circling phases was shorter in Israel (88 plusmn 52 sSpaar 1995 1999) than in Spain In contrast to Israel (Spaar 1995 1997)circling time did not increase with increasing climbing rate Compared tothe semi-desert the development of thermal convection in southern Spainwas probably less stable and birds circled as long as possible in updraftsbecause they could not rely to encounter a next strong thermal within glid-ing distance This might also explain why inter-thermal gliding was oftensupported by apping it reduces sinking rate and prolongs gliding distance
In raptors migrating along the coast ight altitude and gliding airspeedswere related to thermal strength raptors circling in strong thermals hadhigher ight altitudes and higher inter-thermal gliding airspeeds than thosein weak thermals This is consistent with optimal ight theory for birds max-imising cross-country speed in soaring-gliding ight (Pennycuick 1989) asdemonstrated for steppe buzzards Buteo buteo vulpinus (Spaar amp Bruderer1997a)
Flight characteristics in horizontal apping-gliding
According to optimal ight theory maximum range speed decreases withfollowing winds and increases with head- and sidewinds (Pennycuick 1975
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 397
Liechti et al 1994) In raptors ying along the coast we observed decreas-ing airspeeds with increasing tailwind-support whereas birds crossing thesea did not reduce airspeeds Why did the birds react differently Sea cross-ings imply higher risks there is no possibility to land and weather (wind)conditions may change rapidly Higher airspeeds in following winds lead toshorter crossing times and lower the risk of unpredictable weather changesFurthermore drift by sidewinds may considerably extent crossing distancesand in order to reduce drift raptors increased airspeeds with increasingsindwind-components In contrast to sharp-shinned hawks Accipiter stria-tus avoiding crossings in strong lateral winds (Kerlinger 1985) we observedmany raptors to cross the sea in strong north-westerly (ie lateral) winds ifthey provided at least partial tailwind-support
Conclusions
Our observations at an average site at the southern coast of Spain showthat differences in water crossing behaviour are mainly species-speci cpresumably due to the costs for different ight strategies additionallyindividuals reacted to environmental factors such as wind and thermalconditions In medium sized raptors such as honey buzzard soaring-gliding ight along the coast is energy-minimising whereas crossing the sea istime-minimising Sea crossings occurred preferably with a good tailwind-support but also under strong sidewinds Raptors crossing the sea did notreduce airspeeds in following winds but increased airspeeds with increasingsidewinds They seemed to minimise the water crossing duration and theeffect of drift by sidewinds Under good thermal conditions birds regularlyfollowed the coast despite a lower tailwind-support compared to crossing thesea
References
Agostini N amp Logozzo D (1995) Autumn migration of honey buzzards in Southern Italymdash J Raptor Res 29 p 275-277
mdash mdash Logozzo D amp Colero C (1997) Migration of ocks of honey buzzards in SouthernItaly and Malta mdash J Raptor Res 31 p 84-86
mdash mdash Malara G Neri F Mollicone D amp Melotto S (1994) Flight strategies of honeybuzzards during spring migration across the Central Mediterranean mdash Avocetta 18p 73-76
398 MEYER SPAAR amp BRUDERER
Bannermann D-A amp Bannermann WM (1971) Handbook of the birds of Cyprus andmigrants of the Middle East mdash Oliver amp Boyd Edinburgh
Batschelet E (1981) Circular statistics in biology mdash Academic Press LondonBaudinette RV amp Schmidt-Nielsen K (1974) Energy costs of gliding ight in herring
gulls mdash Nature 248 p 83-84Beaman M amp Galea C (1974) The visible migration of raptors over the Maltese Islands
mdash Ibis 116 p 419-431Bernis F (1975) Migracion de Falconiformes y Ciconea spp por Gibraltar mdash IV Avance
sobre recuentos en el verano-otontildeo 1974 mdash Ardeola 21 p 581-592Bijlsma RG (1987) Bottleneck areas for migratory birds in the Mediterranean region mdash
ICBP Study report No 18 CambridgeBruderer B amp Liechti F (1998) Flight behaviour of nocturnally migrating birds in coastal
areas mdash crossing or coasting mdash J Avian Biol 29 p 499-510mdash mdash Blitzblau S amp Peter D (1994) Migration and ight behaviour of honey buzzards
Pernis apivorus in southern Israel observed by radar mdash Ardea 82 p 111-122mdash mdash Steuri T amp Baumgartner M (1995) Short-range high-precision surveillance of
nocturnal migration and tracking of single targets mdash Israel J Zool 41 p 207-220Evans PR amp Lathbury GW (1972) Raptor migration across the Strait of Gibraltar mdash Ibis
115 p 572-585Galea C amp Massa B (1985) Notes on the raptor migration across the central Mediter-
ranean mdash In Conservation studies on raptors (I Newton amp RD Chancellor eds)ICBP Technical Publication No 5 p 257-261
Gensboslashl B (1997) Greifvoumlgel mdash BLV VerlagsgesellschaftMuumlnchenGustin M amp Pizzari T (1998) Migratory pattern in the genus Circus sex and age
differential migration in Italy mdash Ornis Svecica 8 p 23-26Kerlinger P (1984) Flight behaviour of sharp-shinned hawks during migration 2 Over
water mdash Anim Beh 32 p 1029-1034mdash mdash (1985) Water-crossing behaviour of raptors during migration mdash Wilson Bull 97
p 109-113mdash mdash (1989) Flight strategies of migrating hawks mdash University of Chicago Press Chicagomdash mdash Bingman VP amp Able KP (1985) Comparative ight behaviour of migrating hawks
studied with tracking radar during autumn in central New York mdash Can J Zool 63p 755-761
Kjelleacuten N (1992) Differential timing of autumn migration between sex and age groups inraptors at Falsterbo Sweden mdash Orn Scan 23 p 420-434
Liechti F (1993) Naumlchtlicher Vogelzug im Herbst uumlber Suumlddeutschland Winddrift undKompensation mdash J Ornithol 134 p 373-404
mdash mdash Hedenstroumlm A amp Alerstam T (1994) Effects of sidewinds on optimal ight speedsof birds mdash J theor Biol 170 p 219-225
Lustick S Batterby B amp Kelty M (1978) Behavioural thermoregulation orientationtoward the sun in herring gulls mdash Sience 200 p 81-83
Pennycuick C (1975) Mechanics of ight mdash In Avian biology Vol 5 (DS Farner ampRJ King eds) Academic Press New York p 1-75
mdash mdash (1989) Bird ight performance a practical calculation manual mdash Oxford UniversityPress Oxford
mdash mdash (1997) Actual and lsquooptimumrsquo ight speeds eld data reassessedmdash J Exp Biol 200p 2355-2361
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 399
Richardson WJ (1978) Timing and amount of bird migration in relation to weather areview mdash Oikos 30 p 224-272
Spaar R (1995) Flight behaviour of steppe buzzard (Buteo buteo vulpinus) during springmigration in southern Israel a tracking radar study mdash Israel J Zool 41 p 489-500
mdash mdash (1996) Flight behaviour of migrating raptors in southern Israel mdash PhD thesisUniversity of Basel Verlag Schweizerische Vogelwarte Sempach
mdash mdash (1997) Flight strategies of migrating raptors a comparative study of interspecicvariation in ight characteristicsmdash Ibis 139 p 523-535
mdash mdash (1999) Flight behaviour of migrating raptors under varying environmental conditionsmdash In Proc 22 Int Ornithol Congr (N Adams amp R Slotow eds) University of NatalDurban p 1844-1862
mdash mdash amp Bruderer B (1997a) Optimal ight behaviour of soaring migrants A case studyof migrating steppe buzzards Buteo buteo vulpinus mdash Behav Ecol 8 p 288-297)
mdash mdash amp Bruderer B (1997b) Migration by apping or soaring ight strategies of marshMontagursquos and pallid harriers in southern Israel mdash Condor 99 p 458-469
mdash mdash Stark H amp Liechti F (1998) Migratory ight strategies of Levant sparrowhawkstime or energy minimization mdash Anim Behav 56 p 1185-1197
Stark H amp Liechti F (1993) Do Levant sparrowhawks Accipiter brevipes also migrate atnight mdash Ibis 135 p 233-236
Thiollay J-M (1977) Importance des populations de rapaces migrateurs en mediterraneeoccidentale mdash Alauda 45 p 115-121

MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 391
rates did not decrease signi cantly (Spearman rank correlation N = 55r = shy 015 NS) Climbing rates in thermal circling below 1 ms occurredduring the whole day They did not differ between species groups (Table 2ANOVA F = 116 df = 3 N = 115 NS) Flight altitudes increased withincreasing climbing rates in birds lsquo ying along the coastrsquo (r = 049 N = 64p lt 00001) Average circling duration in completed soaring phases was106 plusmn 67 s (N = 80) and it was not related to climbing rate (r = 002N = 80 NS)
Mean airspeed and groundspeed in inter-thermal gliding did not differbetween the species groups (Table 2 ANOVA F = 057 and F = 075respectively df = 3 N = 112 NS) and means per species ranged between120-134 ms and 108-121 ms respectively Raptors increased airspeedsin inter-thermal gliding with increasing climbing rates (r = 052 N =113 p lt 00001) indicating that they maximised cross-country speeds insoaring-gliding ight (Pennycuick 1989 Spaar amp Bruderer 1997a) Meancross-country speeds in soaring-gliding did not differ between species groups(ANOVA cross-country airspeed F = 079 df = 3 N = 54 NS range 93-113 ms cross-country groundspeed F = 027 df = 3 N = 54 NS range76-93 ms)
Flight characteristics in horizontal apping-gliding
This ight style was considered if vertical speeds did not exceed 0 plusmn 05 msand no thermal circling was observed within a track Air- and groundspeedsdiffered between species groups (ANOVA airspeed F = 247 df = 4N = 161 p lt 005 groundspeed F = 387 df = 4 N = 164p lt 0005 Table 2) Ospreys had higher groundspeeds than harriers (GT2-test p lt 005) and higher ground- and airspeeds than black kites (p lt 005)Mean airspeeds per species ranged between 103-132 ms correspondinggroundspeeds between 109-146 ms
Flapping-gliding migrants reached signi cantly higher cross-country air-speeds (113 plusmn 22 ms N = 161) and groundspeeds (122 plusmn 31 ms N =164) than birds in soaring-gliding ight (airspeed 100 plusmn 36 ms N = 55Welch-test t = 249 df = 68 p lt 001 groundspeed 87 plusmn 25 msN = 55 t = 846 df = 114 p lt 00005)
In horizontal apping-gliding ight reactions to different wind con-ditions depended on migratory route raptors ying along the coast de-
392 MEYER SPAAR amp BRUDERER
Fig 8 Airspeed in horizontal apping-gliding ight versus tailwind- and sidewind-components for different migratory routes Linear regression (A) excluding one value inbrackets y = shy 0256x + 1128 R 2 = 0215 N = 56 p lt 0001 (B) r = shy 010 N = 57NS (C) r = shy 007 N = 85 NS (D) excluding two values in brackets y = 0191x+ 1072
R2 = 005 N = 84 p lt 005
creased airspeed with increasing tailwind-component (Fig 8A) but air-speed was not related to sidewind-component (Fig 8B) In contrast in rap-tors crossing the sea no relation was found between airspeed and tailwind-component (Fig 8C) but airspeed increased with increasing sidewind-component (Fig 8D)
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 393
Discussion
There are only few studies on the ight behaviour of migrating raptors cross-ing the sea mainly carried out at concentration points such as narrow wa-ter straits or peninsulas (Evans amp Lathbury 1972 Bernis 1975 Kerlinger1984) and none by tracking radar As these studies were limited to visualobservation mainly high altitudinal migration was often not detected (Ker-linger et al 1985) Our approach is different because (i) we observed raptormigration at an average coast site (ii) we used a tracking radar and (iii) wemeasured wind conditions at all ight levels
Migratory routes and in uence of environmental conditions
Although black kites and in larger numbers honey buzzards undertakelong sea-crossings in the Central Mediterranean (Beaman amp Galea 1974Agostini et al 1994) in our study no black kites and few honey buzzardswere observed attempting the sea crossing The majority of harriers falconsand ospreys crossed the sea Sea crossing honey buzzards occurred towardsthe end of the migratory season and were probably juveniles as in honeybuzzards adults migrate two weeks earlier than juveniles (Kjelleacuten 1992Agostini et al 1997) In contrast to the adults which often migrate in large ocks juveniles mainly migrate singly and have no experience about themigratory ight route This may lead to a higher proportion of juvenilescrossing the sea
Wind and thermal conditions affected migratory directions many harri-ers falcons and ospreys crossed the sea in favourable following and sidewinds but no sea crossings were observed in strong opposing winds This isreasonable because energy and time consumption for the sea crossing andtherefore the risk of exhaustion drastically increase in opposing winds How-ever tailwind-support did not fully explain the decisions on migratory routesbecause regularly birds followed the coast with a lower tailwind-supportcompared to crossing the sea These birds following the coast reached highclimbing rates in thermal circling and this might be compensating for thelower tailwind-support Under good thermal conditions birds were able tomigrate in soaring-gliding ight which consumes much less energy com-pared to apping-gliding ight when crossing the sea (see below Energyand time consumption) Anyway birds in bad shape for example those with
394 MEYER SPAAR amp BRUDERER
low fat reserves should follow the coast in order to minimise the risk ofstarvation to feed when ever it is possible and to y in soaring-gliding ight
At the same observation site migratory directions of nocturnal migrants ying below 2000 m above ground shift from 190deg to 250deg within the nightthe majority of the birds cross the sea when approaching the coast in the rsthalf of the night whereas they hesitate later on (Bruderer amp Liechti 1998)Thus the motivation to cross large barriers seems to decrease after somehours of migratory ight
Energy and time consumption
Energy consumption in apping ight disproportionately increases withbody mass while in soaring-gliding ight it is 2-3 times the basal metabolicrate (BMR) only (for herring gull Larus argentatus Baudinette amp Schmidt-Nielson 1974 Lustick et al 1978) In honey buzzards the average observedairspeed in apping-gliding ight was 111 ms and fat consumption perdistance (gkm) for an average honey buzzard (body mass in Table 3) atthis speed is about 6 times higher than in soaring-gliding ight (calculatedafter Pennycuick (1989 version 11) energy consumption in soaring-gliding ight = 2 times BMR body drag coef cient = 04) If the body dragcoef cient is set to 01 as recently proposed by Pennycuick (1997) energyconsumption in apping ight per km is reduced and energetic differencebetween soaring-gliding and apping ight become smaller but calculatedmaximum range speeds in apping ight are very high compared to ourobservations
We calculated energy and time consumption of different migratory routesbased on observed ight speeds for an average honey buzzard (Table 3) Notethat we did not take into account the time needed for refuelling consumedenergy (ie burned fat) and that we assumed zero wind Crossing the sea in apping-gliding ight (150 km distance) took on average about 4 hours andit was the time-minimising strategy for a honey buzzard (Table 3) Flyingalong the coast (250 km) required about 7 hours in soaring-gliding ightand 6 hours in apping-gliding ight Flying along the coast by soaring-gliding is the energy-minimising strategy for a honey buzzard and consumesabout 4 g fat (Table 3) Flapping-gliding ight along the coast is energeticallythe most expensive but never the fastest strategy Thus black kites andhoney buzzards migrated predominantly according to the energy minimising
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 395
TABLE 3 Observed airspeeds and corresponding estimated ight durationand fat consumption for an average honey buzzard when ying along the
coast or across the sea in different ight styles
Flight style Soaring-gliding Horizontal Horizontal apping-gliding apping-gliding
Flight route Flying along the coast Flying along the coast Crossing the seaDistance 250 km 250 km 150 km
Airspeed ms 101 (81-123) 108 (88-133) 108 (88-133)Flight duration hmin 652 (538-834) 626 (513-753) 351 (308-444)Fat consumption g 41 (34-55) 266 (230-298) 158 (138-179)for ight
Average honey buzzard = body mass 08 kg wing span 13 m wing area 025 m2 Medianand 25-75 range are given in brackets Calculations on fat consumption are based onPennycuickrsquos (1989) program of bird ight performance version 11 and birds are assumedto maximize cross-country speed in soaring-gliding ight (Spaar amp Bruderer 1997a)
strategy because a large proportion of the birds followed the coast in soaring-gliding ight
For species such as falcons or small harriers the difference in energyconsumption between apping and soaring-gliding ight is smaller Asthese species regularly use apping-gliding ight also over land or combinesoaring with apping-gliding ight differences in energy consumptionbetween land and sea route become even smaller and only short detours areenergetically favourable By combining soaring and apping-gliding ightsmaller raptors such as falcons or harriers are able to y under varyingenvironmental conditions to extend their migratory activity to times whenthermal updrafts are absent and to migrate even at night (Stark amp Liechti1993 Spaar amp Bruderer 1997b Spaar et al 1998)
Flight altitudes
Flight altitudes differed between the species groups partly due to differencesin migratory routes Birds crossing the sea ew considerably higher thanthose following the coast Kerlinger (1984) made similar observations atthe Delaware Bay birds crossing the bay ew at higher altitudes thanthose keeping on land He explained the higher ight altitudes by the bettervisibility of the other shore However the African coast is not visible fromour study site Some falcons even started crossing the sea after sunset
396 MEYER SPAAR amp BRUDERER
indicating that they continued migration into the night these birds had to y at least three hours until they reached the African coast The higher ightaltitudes of birds crossing the sea at our study site was mainly caused bytopographical features birds ying with southerly directions were forced toclimb higher up when crossing the mountains chains (1500 m height) about10-15 km northward of the coast and they afterwards maintained these ightlevels when approaching the sea
Flight altitudes of birds migrating along the coast were similar to altitudesobserved in North America and southern Israel with most raptors yingbelow 1000 m above ground and lt 5 above 1500 m (Kerlinger 19841989 Bruderer et al 1994 Spaar 1996)
Flight characteristics in soaring-gliding
Flight characteristics in soaring-gliding ight did not differ between thespecies groups (Table 2) As shown for raptor migration in Israel climb-ing rates do not depend primarily on morphological features but on environ-mental conditions (Spaar 1997) In general mean climbing rates in southernSpain were lower than in the semi-arid desert in Israel (Spaar 1997) Meanduration of the completed circling phases was shorter in Israel (88 plusmn 52 sSpaar 1995 1999) than in Spain In contrast to Israel (Spaar 1995 1997)circling time did not increase with increasing climbing rate Compared tothe semi-desert the development of thermal convection in southern Spainwas probably less stable and birds circled as long as possible in updraftsbecause they could not rely to encounter a next strong thermal within glid-ing distance This might also explain why inter-thermal gliding was oftensupported by apping it reduces sinking rate and prolongs gliding distance
In raptors migrating along the coast ight altitude and gliding airspeedswere related to thermal strength raptors circling in strong thermals hadhigher ight altitudes and higher inter-thermal gliding airspeeds than thosein weak thermals This is consistent with optimal ight theory for birds max-imising cross-country speed in soaring-gliding ight (Pennycuick 1989) asdemonstrated for steppe buzzards Buteo buteo vulpinus (Spaar amp Bruderer1997a)
Flight characteristics in horizontal apping-gliding
According to optimal ight theory maximum range speed decreases withfollowing winds and increases with head- and sidewinds (Pennycuick 1975
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 397
Liechti et al 1994) In raptors ying along the coast we observed decreas-ing airspeeds with increasing tailwind-support whereas birds crossing thesea did not reduce airspeeds Why did the birds react differently Sea cross-ings imply higher risks there is no possibility to land and weather (wind)conditions may change rapidly Higher airspeeds in following winds lead toshorter crossing times and lower the risk of unpredictable weather changesFurthermore drift by sidewinds may considerably extent crossing distancesand in order to reduce drift raptors increased airspeeds with increasingsindwind-components In contrast to sharp-shinned hawks Accipiter stria-tus avoiding crossings in strong lateral winds (Kerlinger 1985) we observedmany raptors to cross the sea in strong north-westerly (ie lateral) winds ifthey provided at least partial tailwind-support
Conclusions
Our observations at an average site at the southern coast of Spain showthat differences in water crossing behaviour are mainly species-speci cpresumably due to the costs for different ight strategies additionallyindividuals reacted to environmental factors such as wind and thermalconditions In medium sized raptors such as honey buzzard soaring-gliding ight along the coast is energy-minimising whereas crossing the sea istime-minimising Sea crossings occurred preferably with a good tailwind-support but also under strong sidewinds Raptors crossing the sea did notreduce airspeeds in following winds but increased airspeeds with increasingsidewinds They seemed to minimise the water crossing duration and theeffect of drift by sidewinds Under good thermal conditions birds regularlyfollowed the coast despite a lower tailwind-support compared to crossing thesea
References
Agostini N amp Logozzo D (1995) Autumn migration of honey buzzards in Southern Italymdash J Raptor Res 29 p 275-277
mdash mdash Logozzo D amp Colero C (1997) Migration of ocks of honey buzzards in SouthernItaly and Malta mdash J Raptor Res 31 p 84-86
mdash mdash Malara G Neri F Mollicone D amp Melotto S (1994) Flight strategies of honeybuzzards during spring migration across the Central Mediterranean mdash Avocetta 18p 73-76
398 MEYER SPAAR amp BRUDERER
Bannermann D-A amp Bannermann WM (1971) Handbook of the birds of Cyprus andmigrants of the Middle East mdash Oliver amp Boyd Edinburgh
Batschelet E (1981) Circular statistics in biology mdash Academic Press LondonBaudinette RV amp Schmidt-Nielsen K (1974) Energy costs of gliding ight in herring
gulls mdash Nature 248 p 83-84Beaman M amp Galea C (1974) The visible migration of raptors over the Maltese Islands
mdash Ibis 116 p 419-431Bernis F (1975) Migracion de Falconiformes y Ciconea spp por Gibraltar mdash IV Avance
sobre recuentos en el verano-otontildeo 1974 mdash Ardeola 21 p 581-592Bijlsma RG (1987) Bottleneck areas for migratory birds in the Mediterranean region mdash
ICBP Study report No 18 CambridgeBruderer B amp Liechti F (1998) Flight behaviour of nocturnally migrating birds in coastal
areas mdash crossing or coasting mdash J Avian Biol 29 p 499-510mdash mdash Blitzblau S amp Peter D (1994) Migration and ight behaviour of honey buzzards
Pernis apivorus in southern Israel observed by radar mdash Ardea 82 p 111-122mdash mdash Steuri T amp Baumgartner M (1995) Short-range high-precision surveillance of
nocturnal migration and tracking of single targets mdash Israel J Zool 41 p 207-220Evans PR amp Lathbury GW (1972) Raptor migration across the Strait of Gibraltar mdash Ibis
115 p 572-585Galea C amp Massa B (1985) Notes on the raptor migration across the central Mediter-
ranean mdash In Conservation studies on raptors (I Newton amp RD Chancellor eds)ICBP Technical Publication No 5 p 257-261
Gensboslashl B (1997) Greifvoumlgel mdash BLV VerlagsgesellschaftMuumlnchenGustin M amp Pizzari T (1998) Migratory pattern in the genus Circus sex and age
differential migration in Italy mdash Ornis Svecica 8 p 23-26Kerlinger P (1984) Flight behaviour of sharp-shinned hawks during migration 2 Over
water mdash Anim Beh 32 p 1029-1034mdash mdash (1985) Water-crossing behaviour of raptors during migration mdash Wilson Bull 97
p 109-113mdash mdash (1989) Flight strategies of migrating hawks mdash University of Chicago Press Chicagomdash mdash Bingman VP amp Able KP (1985) Comparative ight behaviour of migrating hawks
studied with tracking radar during autumn in central New York mdash Can J Zool 63p 755-761
Kjelleacuten N (1992) Differential timing of autumn migration between sex and age groups inraptors at Falsterbo Sweden mdash Orn Scan 23 p 420-434
Liechti F (1993) Naumlchtlicher Vogelzug im Herbst uumlber Suumlddeutschland Winddrift undKompensation mdash J Ornithol 134 p 373-404
mdash mdash Hedenstroumlm A amp Alerstam T (1994) Effects of sidewinds on optimal ight speedsof birds mdash J theor Biol 170 p 219-225
Lustick S Batterby B amp Kelty M (1978) Behavioural thermoregulation orientationtoward the sun in herring gulls mdash Sience 200 p 81-83
Pennycuick C (1975) Mechanics of ight mdash In Avian biology Vol 5 (DS Farner ampRJ King eds) Academic Press New York p 1-75
mdash mdash (1989) Bird ight performance a practical calculation manual mdash Oxford UniversityPress Oxford
mdash mdash (1997) Actual and lsquooptimumrsquo ight speeds eld data reassessedmdash J Exp Biol 200p 2355-2361
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 399
Richardson WJ (1978) Timing and amount of bird migration in relation to weather areview mdash Oikos 30 p 224-272
Spaar R (1995) Flight behaviour of steppe buzzard (Buteo buteo vulpinus) during springmigration in southern Israel a tracking radar study mdash Israel J Zool 41 p 489-500
mdash mdash (1996) Flight behaviour of migrating raptors in southern Israel mdash PhD thesisUniversity of Basel Verlag Schweizerische Vogelwarte Sempach
mdash mdash (1997) Flight strategies of migrating raptors a comparative study of interspecicvariation in ight characteristicsmdash Ibis 139 p 523-535
mdash mdash (1999) Flight behaviour of migrating raptors under varying environmental conditionsmdash In Proc 22 Int Ornithol Congr (N Adams amp R Slotow eds) University of NatalDurban p 1844-1862
mdash mdash amp Bruderer B (1997a) Optimal ight behaviour of soaring migrants A case studyof migrating steppe buzzards Buteo buteo vulpinus mdash Behav Ecol 8 p 288-297)
mdash mdash amp Bruderer B (1997b) Migration by apping or soaring ight strategies of marshMontagursquos and pallid harriers in southern Israel mdash Condor 99 p 458-469
mdash mdash Stark H amp Liechti F (1998) Migratory ight strategies of Levant sparrowhawkstime or energy minimization mdash Anim Behav 56 p 1185-1197
Stark H amp Liechti F (1993) Do Levant sparrowhawks Accipiter brevipes also migrate atnight mdash Ibis 135 p 233-236
Thiollay J-M (1977) Importance des populations de rapaces migrateurs en mediterraneeoccidentale mdash Alauda 45 p 115-121
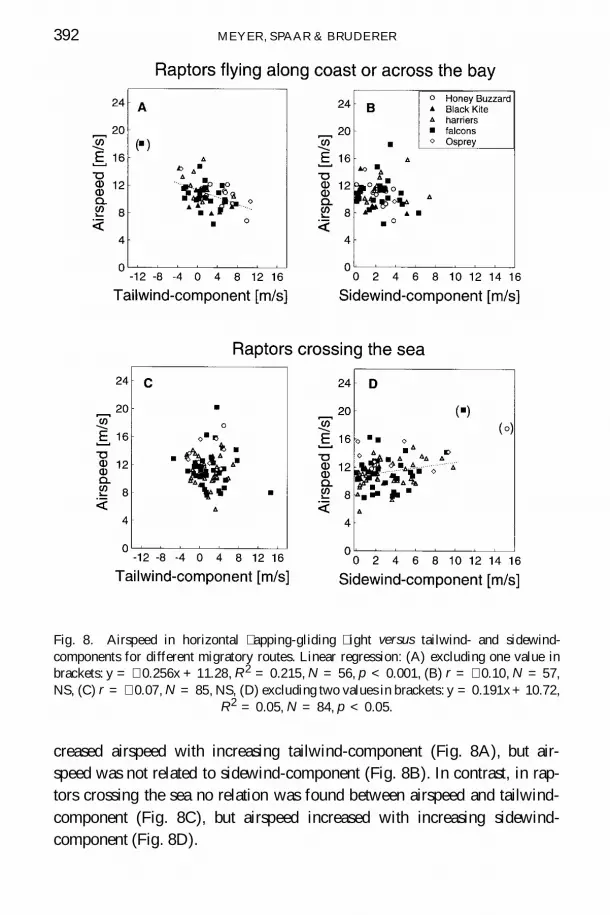
392 MEYER SPAAR amp BRUDERER
Fig 8 Airspeed in horizontal apping-gliding ight versus tailwind- and sidewind-components for different migratory routes Linear regression (A) excluding one value inbrackets y = shy 0256x + 1128 R 2 = 0215 N = 56 p lt 0001 (B) r = shy 010 N = 57NS (C) r = shy 007 N = 85 NS (D) excluding two values in brackets y = 0191x+ 1072
R2 = 005 N = 84 p lt 005
creased airspeed with increasing tailwind-component (Fig 8A) but air-speed was not related to sidewind-component (Fig 8B) In contrast in rap-tors crossing the sea no relation was found between airspeed and tailwind-component (Fig 8C) but airspeed increased with increasing sidewind-component (Fig 8D)
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 393
Discussion
There are only few studies on the ight behaviour of migrating raptors cross-ing the sea mainly carried out at concentration points such as narrow wa-ter straits or peninsulas (Evans amp Lathbury 1972 Bernis 1975 Kerlinger1984) and none by tracking radar As these studies were limited to visualobservation mainly high altitudinal migration was often not detected (Ker-linger et al 1985) Our approach is different because (i) we observed raptormigration at an average coast site (ii) we used a tracking radar and (iii) wemeasured wind conditions at all ight levels
Migratory routes and in uence of environmental conditions
Although black kites and in larger numbers honey buzzards undertakelong sea-crossings in the Central Mediterranean (Beaman amp Galea 1974Agostini et al 1994) in our study no black kites and few honey buzzardswere observed attempting the sea crossing The majority of harriers falconsand ospreys crossed the sea Sea crossing honey buzzards occurred towardsthe end of the migratory season and were probably juveniles as in honeybuzzards adults migrate two weeks earlier than juveniles (Kjelleacuten 1992Agostini et al 1997) In contrast to the adults which often migrate in large ocks juveniles mainly migrate singly and have no experience about themigratory ight route This may lead to a higher proportion of juvenilescrossing the sea
Wind and thermal conditions affected migratory directions many harri-ers falcons and ospreys crossed the sea in favourable following and sidewinds but no sea crossings were observed in strong opposing winds This isreasonable because energy and time consumption for the sea crossing andtherefore the risk of exhaustion drastically increase in opposing winds How-ever tailwind-support did not fully explain the decisions on migratory routesbecause regularly birds followed the coast with a lower tailwind-supportcompared to crossing the sea These birds following the coast reached highclimbing rates in thermal circling and this might be compensating for thelower tailwind-support Under good thermal conditions birds were able tomigrate in soaring-gliding ight which consumes much less energy com-pared to apping-gliding ight when crossing the sea (see below Energyand time consumption) Anyway birds in bad shape for example those with
394 MEYER SPAAR amp BRUDERER
low fat reserves should follow the coast in order to minimise the risk ofstarvation to feed when ever it is possible and to y in soaring-gliding ight
At the same observation site migratory directions of nocturnal migrants ying below 2000 m above ground shift from 190deg to 250deg within the nightthe majority of the birds cross the sea when approaching the coast in the rsthalf of the night whereas they hesitate later on (Bruderer amp Liechti 1998)Thus the motivation to cross large barriers seems to decrease after somehours of migratory ight
Energy and time consumption
Energy consumption in apping ight disproportionately increases withbody mass while in soaring-gliding ight it is 2-3 times the basal metabolicrate (BMR) only (for herring gull Larus argentatus Baudinette amp Schmidt-Nielson 1974 Lustick et al 1978) In honey buzzards the average observedairspeed in apping-gliding ight was 111 ms and fat consumption perdistance (gkm) for an average honey buzzard (body mass in Table 3) atthis speed is about 6 times higher than in soaring-gliding ight (calculatedafter Pennycuick (1989 version 11) energy consumption in soaring-gliding ight = 2 times BMR body drag coef cient = 04) If the body dragcoef cient is set to 01 as recently proposed by Pennycuick (1997) energyconsumption in apping ight per km is reduced and energetic differencebetween soaring-gliding and apping ight become smaller but calculatedmaximum range speeds in apping ight are very high compared to ourobservations
We calculated energy and time consumption of different migratory routesbased on observed ight speeds for an average honey buzzard (Table 3) Notethat we did not take into account the time needed for refuelling consumedenergy (ie burned fat) and that we assumed zero wind Crossing the sea in apping-gliding ight (150 km distance) took on average about 4 hours andit was the time-minimising strategy for a honey buzzard (Table 3) Flyingalong the coast (250 km) required about 7 hours in soaring-gliding ightand 6 hours in apping-gliding ight Flying along the coast by soaring-gliding is the energy-minimising strategy for a honey buzzard and consumesabout 4 g fat (Table 3) Flapping-gliding ight along the coast is energeticallythe most expensive but never the fastest strategy Thus black kites andhoney buzzards migrated predominantly according to the energy minimising
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 395
TABLE 3 Observed airspeeds and corresponding estimated ight durationand fat consumption for an average honey buzzard when ying along the
coast or across the sea in different ight styles
Flight style Soaring-gliding Horizontal Horizontal apping-gliding apping-gliding
Flight route Flying along the coast Flying along the coast Crossing the seaDistance 250 km 250 km 150 km
Airspeed ms 101 (81-123) 108 (88-133) 108 (88-133)Flight duration hmin 652 (538-834) 626 (513-753) 351 (308-444)Fat consumption g 41 (34-55) 266 (230-298) 158 (138-179)for ight
Average honey buzzard = body mass 08 kg wing span 13 m wing area 025 m2 Medianand 25-75 range are given in brackets Calculations on fat consumption are based onPennycuickrsquos (1989) program of bird ight performance version 11 and birds are assumedto maximize cross-country speed in soaring-gliding ight (Spaar amp Bruderer 1997a)
strategy because a large proportion of the birds followed the coast in soaring-gliding ight
For species such as falcons or small harriers the difference in energyconsumption between apping and soaring-gliding ight is smaller Asthese species regularly use apping-gliding ight also over land or combinesoaring with apping-gliding ight differences in energy consumptionbetween land and sea route become even smaller and only short detours areenergetically favourable By combining soaring and apping-gliding ightsmaller raptors such as falcons or harriers are able to y under varyingenvironmental conditions to extend their migratory activity to times whenthermal updrafts are absent and to migrate even at night (Stark amp Liechti1993 Spaar amp Bruderer 1997b Spaar et al 1998)
Flight altitudes
Flight altitudes differed between the species groups partly due to differencesin migratory routes Birds crossing the sea ew considerably higher thanthose following the coast Kerlinger (1984) made similar observations atthe Delaware Bay birds crossing the bay ew at higher altitudes thanthose keeping on land He explained the higher ight altitudes by the bettervisibility of the other shore However the African coast is not visible fromour study site Some falcons even started crossing the sea after sunset
396 MEYER SPAAR amp BRUDERER
indicating that they continued migration into the night these birds had to y at least three hours until they reached the African coast The higher ightaltitudes of birds crossing the sea at our study site was mainly caused bytopographical features birds ying with southerly directions were forced toclimb higher up when crossing the mountains chains (1500 m height) about10-15 km northward of the coast and they afterwards maintained these ightlevels when approaching the sea
Flight altitudes of birds migrating along the coast were similar to altitudesobserved in North America and southern Israel with most raptors yingbelow 1000 m above ground and lt 5 above 1500 m (Kerlinger 19841989 Bruderer et al 1994 Spaar 1996)
Flight characteristics in soaring-gliding
Flight characteristics in soaring-gliding ight did not differ between thespecies groups (Table 2) As shown for raptor migration in Israel climb-ing rates do not depend primarily on morphological features but on environ-mental conditions (Spaar 1997) In general mean climbing rates in southernSpain were lower than in the semi-arid desert in Israel (Spaar 1997) Meanduration of the completed circling phases was shorter in Israel (88 plusmn 52 sSpaar 1995 1999) than in Spain In contrast to Israel (Spaar 1995 1997)circling time did not increase with increasing climbing rate Compared tothe semi-desert the development of thermal convection in southern Spainwas probably less stable and birds circled as long as possible in updraftsbecause they could not rely to encounter a next strong thermal within glid-ing distance This might also explain why inter-thermal gliding was oftensupported by apping it reduces sinking rate and prolongs gliding distance
In raptors migrating along the coast ight altitude and gliding airspeedswere related to thermal strength raptors circling in strong thermals hadhigher ight altitudes and higher inter-thermal gliding airspeeds than thosein weak thermals This is consistent with optimal ight theory for birds max-imising cross-country speed in soaring-gliding ight (Pennycuick 1989) asdemonstrated for steppe buzzards Buteo buteo vulpinus (Spaar amp Bruderer1997a)
Flight characteristics in horizontal apping-gliding
According to optimal ight theory maximum range speed decreases withfollowing winds and increases with head- and sidewinds (Pennycuick 1975
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 397
Liechti et al 1994) In raptors ying along the coast we observed decreas-ing airspeeds with increasing tailwind-support whereas birds crossing thesea did not reduce airspeeds Why did the birds react differently Sea cross-ings imply higher risks there is no possibility to land and weather (wind)conditions may change rapidly Higher airspeeds in following winds lead toshorter crossing times and lower the risk of unpredictable weather changesFurthermore drift by sidewinds may considerably extent crossing distancesand in order to reduce drift raptors increased airspeeds with increasingsindwind-components In contrast to sharp-shinned hawks Accipiter stria-tus avoiding crossings in strong lateral winds (Kerlinger 1985) we observedmany raptors to cross the sea in strong north-westerly (ie lateral) winds ifthey provided at least partial tailwind-support
Conclusions
Our observations at an average site at the southern coast of Spain showthat differences in water crossing behaviour are mainly species-speci cpresumably due to the costs for different ight strategies additionallyindividuals reacted to environmental factors such as wind and thermalconditions In medium sized raptors such as honey buzzard soaring-gliding ight along the coast is energy-minimising whereas crossing the sea istime-minimising Sea crossings occurred preferably with a good tailwind-support but also under strong sidewinds Raptors crossing the sea did notreduce airspeeds in following winds but increased airspeeds with increasingsidewinds They seemed to minimise the water crossing duration and theeffect of drift by sidewinds Under good thermal conditions birds regularlyfollowed the coast despite a lower tailwind-support compared to crossing thesea
References
Agostini N amp Logozzo D (1995) Autumn migration of honey buzzards in Southern Italymdash J Raptor Res 29 p 275-277
mdash mdash Logozzo D amp Colero C (1997) Migration of ocks of honey buzzards in SouthernItaly and Malta mdash J Raptor Res 31 p 84-86
mdash mdash Malara G Neri F Mollicone D amp Melotto S (1994) Flight strategies of honeybuzzards during spring migration across the Central Mediterranean mdash Avocetta 18p 73-76
398 MEYER SPAAR amp BRUDERER
Bannermann D-A amp Bannermann WM (1971) Handbook of the birds of Cyprus andmigrants of the Middle East mdash Oliver amp Boyd Edinburgh
Batschelet E (1981) Circular statistics in biology mdash Academic Press LondonBaudinette RV amp Schmidt-Nielsen K (1974) Energy costs of gliding ight in herring
gulls mdash Nature 248 p 83-84Beaman M amp Galea C (1974) The visible migration of raptors over the Maltese Islands
mdash Ibis 116 p 419-431Bernis F (1975) Migracion de Falconiformes y Ciconea spp por Gibraltar mdash IV Avance
sobre recuentos en el verano-otontildeo 1974 mdash Ardeola 21 p 581-592Bijlsma RG (1987) Bottleneck areas for migratory birds in the Mediterranean region mdash
ICBP Study report No 18 CambridgeBruderer B amp Liechti F (1998) Flight behaviour of nocturnally migrating birds in coastal
areas mdash crossing or coasting mdash J Avian Biol 29 p 499-510mdash mdash Blitzblau S amp Peter D (1994) Migration and ight behaviour of honey buzzards
Pernis apivorus in southern Israel observed by radar mdash Ardea 82 p 111-122mdash mdash Steuri T amp Baumgartner M (1995) Short-range high-precision surveillance of
nocturnal migration and tracking of single targets mdash Israel J Zool 41 p 207-220Evans PR amp Lathbury GW (1972) Raptor migration across the Strait of Gibraltar mdash Ibis
115 p 572-585Galea C amp Massa B (1985) Notes on the raptor migration across the central Mediter-
ranean mdash In Conservation studies on raptors (I Newton amp RD Chancellor eds)ICBP Technical Publication No 5 p 257-261
Gensboslashl B (1997) Greifvoumlgel mdash BLV VerlagsgesellschaftMuumlnchenGustin M amp Pizzari T (1998) Migratory pattern in the genus Circus sex and age
differential migration in Italy mdash Ornis Svecica 8 p 23-26Kerlinger P (1984) Flight behaviour of sharp-shinned hawks during migration 2 Over
water mdash Anim Beh 32 p 1029-1034mdash mdash (1985) Water-crossing behaviour of raptors during migration mdash Wilson Bull 97
p 109-113mdash mdash (1989) Flight strategies of migrating hawks mdash University of Chicago Press Chicagomdash mdash Bingman VP amp Able KP (1985) Comparative ight behaviour of migrating hawks
studied with tracking radar during autumn in central New York mdash Can J Zool 63p 755-761
Kjelleacuten N (1992) Differential timing of autumn migration between sex and age groups inraptors at Falsterbo Sweden mdash Orn Scan 23 p 420-434
Liechti F (1993) Naumlchtlicher Vogelzug im Herbst uumlber Suumlddeutschland Winddrift undKompensation mdash J Ornithol 134 p 373-404
mdash mdash Hedenstroumlm A amp Alerstam T (1994) Effects of sidewinds on optimal ight speedsof birds mdash J theor Biol 170 p 219-225
Lustick S Batterby B amp Kelty M (1978) Behavioural thermoregulation orientationtoward the sun in herring gulls mdash Sience 200 p 81-83
Pennycuick C (1975) Mechanics of ight mdash In Avian biology Vol 5 (DS Farner ampRJ King eds) Academic Press New York p 1-75
mdash mdash (1989) Bird ight performance a practical calculation manual mdash Oxford UniversityPress Oxford
mdash mdash (1997) Actual and lsquooptimumrsquo ight speeds eld data reassessedmdash J Exp Biol 200p 2355-2361
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 399
Richardson WJ (1978) Timing and amount of bird migration in relation to weather areview mdash Oikos 30 p 224-272
Spaar R (1995) Flight behaviour of steppe buzzard (Buteo buteo vulpinus) during springmigration in southern Israel a tracking radar study mdash Israel J Zool 41 p 489-500
mdash mdash (1996) Flight behaviour of migrating raptors in southern Israel mdash PhD thesisUniversity of Basel Verlag Schweizerische Vogelwarte Sempach
mdash mdash (1997) Flight strategies of migrating raptors a comparative study of interspecicvariation in ight characteristicsmdash Ibis 139 p 523-535
mdash mdash (1999) Flight behaviour of migrating raptors under varying environmental conditionsmdash In Proc 22 Int Ornithol Congr (N Adams amp R Slotow eds) University of NatalDurban p 1844-1862
mdash mdash amp Bruderer B (1997a) Optimal ight behaviour of soaring migrants A case studyof migrating steppe buzzards Buteo buteo vulpinus mdash Behav Ecol 8 p 288-297)
mdash mdash amp Bruderer B (1997b) Migration by apping or soaring ight strategies of marshMontagursquos and pallid harriers in southern Israel mdash Condor 99 p 458-469
mdash mdash Stark H amp Liechti F (1998) Migratory ight strategies of Levant sparrowhawkstime or energy minimization mdash Anim Behav 56 p 1185-1197
Stark H amp Liechti F (1993) Do Levant sparrowhawks Accipiter brevipes also migrate atnight mdash Ibis 135 p 233-236
Thiollay J-M (1977) Importance des populations de rapaces migrateurs en mediterraneeoccidentale mdash Alauda 45 p 115-121

MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 393
Discussion
There are only few studies on the ight behaviour of migrating raptors cross-ing the sea mainly carried out at concentration points such as narrow wa-ter straits or peninsulas (Evans amp Lathbury 1972 Bernis 1975 Kerlinger1984) and none by tracking radar As these studies were limited to visualobservation mainly high altitudinal migration was often not detected (Ker-linger et al 1985) Our approach is different because (i) we observed raptormigration at an average coast site (ii) we used a tracking radar and (iii) wemeasured wind conditions at all ight levels
Migratory routes and in uence of environmental conditions
Although black kites and in larger numbers honey buzzards undertakelong sea-crossings in the Central Mediterranean (Beaman amp Galea 1974Agostini et al 1994) in our study no black kites and few honey buzzardswere observed attempting the sea crossing The majority of harriers falconsand ospreys crossed the sea Sea crossing honey buzzards occurred towardsthe end of the migratory season and were probably juveniles as in honeybuzzards adults migrate two weeks earlier than juveniles (Kjelleacuten 1992Agostini et al 1997) In contrast to the adults which often migrate in large ocks juveniles mainly migrate singly and have no experience about themigratory ight route This may lead to a higher proportion of juvenilescrossing the sea
Wind and thermal conditions affected migratory directions many harri-ers falcons and ospreys crossed the sea in favourable following and sidewinds but no sea crossings were observed in strong opposing winds This isreasonable because energy and time consumption for the sea crossing andtherefore the risk of exhaustion drastically increase in opposing winds How-ever tailwind-support did not fully explain the decisions on migratory routesbecause regularly birds followed the coast with a lower tailwind-supportcompared to crossing the sea These birds following the coast reached highclimbing rates in thermal circling and this might be compensating for thelower tailwind-support Under good thermal conditions birds were able tomigrate in soaring-gliding ight which consumes much less energy com-pared to apping-gliding ight when crossing the sea (see below Energyand time consumption) Anyway birds in bad shape for example those with
394 MEYER SPAAR amp BRUDERER
low fat reserves should follow the coast in order to minimise the risk ofstarvation to feed when ever it is possible and to y in soaring-gliding ight
At the same observation site migratory directions of nocturnal migrants ying below 2000 m above ground shift from 190deg to 250deg within the nightthe majority of the birds cross the sea when approaching the coast in the rsthalf of the night whereas they hesitate later on (Bruderer amp Liechti 1998)Thus the motivation to cross large barriers seems to decrease after somehours of migratory ight
Energy and time consumption
Energy consumption in apping ight disproportionately increases withbody mass while in soaring-gliding ight it is 2-3 times the basal metabolicrate (BMR) only (for herring gull Larus argentatus Baudinette amp Schmidt-Nielson 1974 Lustick et al 1978) In honey buzzards the average observedairspeed in apping-gliding ight was 111 ms and fat consumption perdistance (gkm) for an average honey buzzard (body mass in Table 3) atthis speed is about 6 times higher than in soaring-gliding ight (calculatedafter Pennycuick (1989 version 11) energy consumption in soaring-gliding ight = 2 times BMR body drag coef cient = 04) If the body dragcoef cient is set to 01 as recently proposed by Pennycuick (1997) energyconsumption in apping ight per km is reduced and energetic differencebetween soaring-gliding and apping ight become smaller but calculatedmaximum range speeds in apping ight are very high compared to ourobservations
We calculated energy and time consumption of different migratory routesbased on observed ight speeds for an average honey buzzard (Table 3) Notethat we did not take into account the time needed for refuelling consumedenergy (ie burned fat) and that we assumed zero wind Crossing the sea in apping-gliding ight (150 km distance) took on average about 4 hours andit was the time-minimising strategy for a honey buzzard (Table 3) Flyingalong the coast (250 km) required about 7 hours in soaring-gliding ightand 6 hours in apping-gliding ight Flying along the coast by soaring-gliding is the energy-minimising strategy for a honey buzzard and consumesabout 4 g fat (Table 3) Flapping-gliding ight along the coast is energeticallythe most expensive but never the fastest strategy Thus black kites andhoney buzzards migrated predominantly according to the energy minimising
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 395
TABLE 3 Observed airspeeds and corresponding estimated ight durationand fat consumption for an average honey buzzard when ying along the
coast or across the sea in different ight styles
Flight style Soaring-gliding Horizontal Horizontal apping-gliding apping-gliding
Flight route Flying along the coast Flying along the coast Crossing the seaDistance 250 km 250 km 150 km
Airspeed ms 101 (81-123) 108 (88-133) 108 (88-133)Flight duration hmin 652 (538-834) 626 (513-753) 351 (308-444)Fat consumption g 41 (34-55) 266 (230-298) 158 (138-179)for ight
Average honey buzzard = body mass 08 kg wing span 13 m wing area 025 m2 Medianand 25-75 range are given in brackets Calculations on fat consumption are based onPennycuickrsquos (1989) program of bird ight performance version 11 and birds are assumedto maximize cross-country speed in soaring-gliding ight (Spaar amp Bruderer 1997a)
strategy because a large proportion of the birds followed the coast in soaring-gliding ight
For species such as falcons or small harriers the difference in energyconsumption between apping and soaring-gliding ight is smaller Asthese species regularly use apping-gliding ight also over land or combinesoaring with apping-gliding ight differences in energy consumptionbetween land and sea route become even smaller and only short detours areenergetically favourable By combining soaring and apping-gliding ightsmaller raptors such as falcons or harriers are able to y under varyingenvironmental conditions to extend their migratory activity to times whenthermal updrafts are absent and to migrate even at night (Stark amp Liechti1993 Spaar amp Bruderer 1997b Spaar et al 1998)
Flight altitudes
Flight altitudes differed between the species groups partly due to differencesin migratory routes Birds crossing the sea ew considerably higher thanthose following the coast Kerlinger (1984) made similar observations atthe Delaware Bay birds crossing the bay ew at higher altitudes thanthose keeping on land He explained the higher ight altitudes by the bettervisibility of the other shore However the African coast is not visible fromour study site Some falcons even started crossing the sea after sunset
396 MEYER SPAAR amp BRUDERER
indicating that they continued migration into the night these birds had to y at least three hours until they reached the African coast The higher ightaltitudes of birds crossing the sea at our study site was mainly caused bytopographical features birds ying with southerly directions were forced toclimb higher up when crossing the mountains chains (1500 m height) about10-15 km northward of the coast and they afterwards maintained these ightlevels when approaching the sea
Flight altitudes of birds migrating along the coast were similar to altitudesobserved in North America and southern Israel with most raptors yingbelow 1000 m above ground and lt 5 above 1500 m (Kerlinger 19841989 Bruderer et al 1994 Spaar 1996)
Flight characteristics in soaring-gliding
Flight characteristics in soaring-gliding ight did not differ between thespecies groups (Table 2) As shown for raptor migration in Israel climb-ing rates do not depend primarily on morphological features but on environ-mental conditions (Spaar 1997) In general mean climbing rates in southernSpain were lower than in the semi-arid desert in Israel (Spaar 1997) Meanduration of the completed circling phases was shorter in Israel (88 plusmn 52 sSpaar 1995 1999) than in Spain In contrast to Israel (Spaar 1995 1997)circling time did not increase with increasing climbing rate Compared tothe semi-desert the development of thermal convection in southern Spainwas probably less stable and birds circled as long as possible in updraftsbecause they could not rely to encounter a next strong thermal within glid-ing distance This might also explain why inter-thermal gliding was oftensupported by apping it reduces sinking rate and prolongs gliding distance
In raptors migrating along the coast ight altitude and gliding airspeedswere related to thermal strength raptors circling in strong thermals hadhigher ight altitudes and higher inter-thermal gliding airspeeds than thosein weak thermals This is consistent with optimal ight theory for birds max-imising cross-country speed in soaring-gliding ight (Pennycuick 1989) asdemonstrated for steppe buzzards Buteo buteo vulpinus (Spaar amp Bruderer1997a)
Flight characteristics in horizontal apping-gliding
According to optimal ight theory maximum range speed decreases withfollowing winds and increases with head- and sidewinds (Pennycuick 1975
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 397
Liechti et al 1994) In raptors ying along the coast we observed decreas-ing airspeeds with increasing tailwind-support whereas birds crossing thesea did not reduce airspeeds Why did the birds react differently Sea cross-ings imply higher risks there is no possibility to land and weather (wind)conditions may change rapidly Higher airspeeds in following winds lead toshorter crossing times and lower the risk of unpredictable weather changesFurthermore drift by sidewinds may considerably extent crossing distancesand in order to reduce drift raptors increased airspeeds with increasingsindwind-components In contrast to sharp-shinned hawks Accipiter stria-tus avoiding crossings in strong lateral winds (Kerlinger 1985) we observedmany raptors to cross the sea in strong north-westerly (ie lateral) winds ifthey provided at least partial tailwind-support
Conclusions
Our observations at an average site at the southern coast of Spain showthat differences in water crossing behaviour are mainly species-speci cpresumably due to the costs for different ight strategies additionallyindividuals reacted to environmental factors such as wind and thermalconditions In medium sized raptors such as honey buzzard soaring-gliding ight along the coast is energy-minimising whereas crossing the sea istime-minimising Sea crossings occurred preferably with a good tailwind-support but also under strong sidewinds Raptors crossing the sea did notreduce airspeeds in following winds but increased airspeeds with increasingsidewinds They seemed to minimise the water crossing duration and theeffect of drift by sidewinds Under good thermal conditions birds regularlyfollowed the coast despite a lower tailwind-support compared to crossing thesea
References
Agostini N amp Logozzo D (1995) Autumn migration of honey buzzards in Southern Italymdash J Raptor Res 29 p 275-277
mdash mdash Logozzo D amp Colero C (1997) Migration of ocks of honey buzzards in SouthernItaly and Malta mdash J Raptor Res 31 p 84-86
mdash mdash Malara G Neri F Mollicone D amp Melotto S (1994) Flight strategies of honeybuzzards during spring migration across the Central Mediterranean mdash Avocetta 18p 73-76
398 MEYER SPAAR amp BRUDERER
Bannermann D-A amp Bannermann WM (1971) Handbook of the birds of Cyprus andmigrants of the Middle East mdash Oliver amp Boyd Edinburgh
Batschelet E (1981) Circular statistics in biology mdash Academic Press LondonBaudinette RV amp Schmidt-Nielsen K (1974) Energy costs of gliding ight in herring
gulls mdash Nature 248 p 83-84Beaman M amp Galea C (1974) The visible migration of raptors over the Maltese Islands
mdash Ibis 116 p 419-431Bernis F (1975) Migracion de Falconiformes y Ciconea spp por Gibraltar mdash IV Avance
sobre recuentos en el verano-otontildeo 1974 mdash Ardeola 21 p 581-592Bijlsma RG (1987) Bottleneck areas for migratory birds in the Mediterranean region mdash
ICBP Study report No 18 CambridgeBruderer B amp Liechti F (1998) Flight behaviour of nocturnally migrating birds in coastal
areas mdash crossing or coasting mdash J Avian Biol 29 p 499-510mdash mdash Blitzblau S amp Peter D (1994) Migration and ight behaviour of honey buzzards
Pernis apivorus in southern Israel observed by radar mdash Ardea 82 p 111-122mdash mdash Steuri T amp Baumgartner M (1995) Short-range high-precision surveillance of
nocturnal migration and tracking of single targets mdash Israel J Zool 41 p 207-220Evans PR amp Lathbury GW (1972) Raptor migration across the Strait of Gibraltar mdash Ibis
115 p 572-585Galea C amp Massa B (1985) Notes on the raptor migration across the central Mediter-
ranean mdash In Conservation studies on raptors (I Newton amp RD Chancellor eds)ICBP Technical Publication No 5 p 257-261
Gensboslashl B (1997) Greifvoumlgel mdash BLV VerlagsgesellschaftMuumlnchenGustin M amp Pizzari T (1998) Migratory pattern in the genus Circus sex and age
differential migration in Italy mdash Ornis Svecica 8 p 23-26Kerlinger P (1984) Flight behaviour of sharp-shinned hawks during migration 2 Over
water mdash Anim Beh 32 p 1029-1034mdash mdash (1985) Water-crossing behaviour of raptors during migration mdash Wilson Bull 97
p 109-113mdash mdash (1989) Flight strategies of migrating hawks mdash University of Chicago Press Chicagomdash mdash Bingman VP amp Able KP (1985) Comparative ight behaviour of migrating hawks
studied with tracking radar during autumn in central New York mdash Can J Zool 63p 755-761
Kjelleacuten N (1992) Differential timing of autumn migration between sex and age groups inraptors at Falsterbo Sweden mdash Orn Scan 23 p 420-434
Liechti F (1993) Naumlchtlicher Vogelzug im Herbst uumlber Suumlddeutschland Winddrift undKompensation mdash J Ornithol 134 p 373-404
mdash mdash Hedenstroumlm A amp Alerstam T (1994) Effects of sidewinds on optimal ight speedsof birds mdash J theor Biol 170 p 219-225
Lustick S Batterby B amp Kelty M (1978) Behavioural thermoregulation orientationtoward the sun in herring gulls mdash Sience 200 p 81-83
Pennycuick C (1975) Mechanics of ight mdash In Avian biology Vol 5 (DS Farner ampRJ King eds) Academic Press New York p 1-75
mdash mdash (1989) Bird ight performance a practical calculation manual mdash Oxford UniversityPress Oxford
mdash mdash (1997) Actual and lsquooptimumrsquo ight speeds eld data reassessedmdash J Exp Biol 200p 2355-2361
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 399
Richardson WJ (1978) Timing and amount of bird migration in relation to weather areview mdash Oikos 30 p 224-272
Spaar R (1995) Flight behaviour of steppe buzzard (Buteo buteo vulpinus) during springmigration in southern Israel a tracking radar study mdash Israel J Zool 41 p 489-500
mdash mdash (1996) Flight behaviour of migrating raptors in southern Israel mdash PhD thesisUniversity of Basel Verlag Schweizerische Vogelwarte Sempach
mdash mdash (1997) Flight strategies of migrating raptors a comparative study of interspecicvariation in ight characteristicsmdash Ibis 139 p 523-535
mdash mdash (1999) Flight behaviour of migrating raptors under varying environmental conditionsmdash In Proc 22 Int Ornithol Congr (N Adams amp R Slotow eds) University of NatalDurban p 1844-1862
mdash mdash amp Bruderer B (1997a) Optimal ight behaviour of soaring migrants A case studyof migrating steppe buzzards Buteo buteo vulpinus mdash Behav Ecol 8 p 288-297)
mdash mdash amp Bruderer B (1997b) Migration by apping or soaring ight strategies of marshMontagursquos and pallid harriers in southern Israel mdash Condor 99 p 458-469
mdash mdash Stark H amp Liechti F (1998) Migratory ight strategies of Levant sparrowhawkstime or energy minimization mdash Anim Behav 56 p 1185-1197
Stark H amp Liechti F (1993) Do Levant sparrowhawks Accipiter brevipes also migrate atnight mdash Ibis 135 p 233-236
Thiollay J-M (1977) Importance des populations de rapaces migrateurs en mediterraneeoccidentale mdash Alauda 45 p 115-121

394 MEYER SPAAR amp BRUDERER
low fat reserves should follow the coast in order to minimise the risk ofstarvation to feed when ever it is possible and to y in soaring-gliding ight
At the same observation site migratory directions of nocturnal migrants ying below 2000 m above ground shift from 190deg to 250deg within the nightthe majority of the birds cross the sea when approaching the coast in the rsthalf of the night whereas they hesitate later on (Bruderer amp Liechti 1998)Thus the motivation to cross large barriers seems to decrease after somehours of migratory ight
Energy and time consumption
Energy consumption in apping ight disproportionately increases withbody mass while in soaring-gliding ight it is 2-3 times the basal metabolicrate (BMR) only (for herring gull Larus argentatus Baudinette amp Schmidt-Nielson 1974 Lustick et al 1978) In honey buzzards the average observedairspeed in apping-gliding ight was 111 ms and fat consumption perdistance (gkm) for an average honey buzzard (body mass in Table 3) atthis speed is about 6 times higher than in soaring-gliding ight (calculatedafter Pennycuick (1989 version 11) energy consumption in soaring-gliding ight = 2 times BMR body drag coef cient = 04) If the body dragcoef cient is set to 01 as recently proposed by Pennycuick (1997) energyconsumption in apping ight per km is reduced and energetic differencebetween soaring-gliding and apping ight become smaller but calculatedmaximum range speeds in apping ight are very high compared to ourobservations
We calculated energy and time consumption of different migratory routesbased on observed ight speeds for an average honey buzzard (Table 3) Notethat we did not take into account the time needed for refuelling consumedenergy (ie burned fat) and that we assumed zero wind Crossing the sea in apping-gliding ight (150 km distance) took on average about 4 hours andit was the time-minimising strategy for a honey buzzard (Table 3) Flyingalong the coast (250 km) required about 7 hours in soaring-gliding ightand 6 hours in apping-gliding ight Flying along the coast by soaring-gliding is the energy-minimising strategy for a honey buzzard and consumesabout 4 g fat (Table 3) Flapping-gliding ight along the coast is energeticallythe most expensive but never the fastest strategy Thus black kites andhoney buzzards migrated predominantly according to the energy minimising
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 395
TABLE 3 Observed airspeeds and corresponding estimated ight durationand fat consumption for an average honey buzzard when ying along the
coast or across the sea in different ight styles
Flight style Soaring-gliding Horizontal Horizontal apping-gliding apping-gliding
Flight route Flying along the coast Flying along the coast Crossing the seaDistance 250 km 250 km 150 km
Airspeed ms 101 (81-123) 108 (88-133) 108 (88-133)Flight duration hmin 652 (538-834) 626 (513-753) 351 (308-444)Fat consumption g 41 (34-55) 266 (230-298) 158 (138-179)for ight
Average honey buzzard = body mass 08 kg wing span 13 m wing area 025 m2 Medianand 25-75 range are given in brackets Calculations on fat consumption are based onPennycuickrsquos (1989) program of bird ight performance version 11 and birds are assumedto maximize cross-country speed in soaring-gliding ight (Spaar amp Bruderer 1997a)
strategy because a large proportion of the birds followed the coast in soaring-gliding ight
For species such as falcons or small harriers the difference in energyconsumption between apping and soaring-gliding ight is smaller Asthese species regularly use apping-gliding ight also over land or combinesoaring with apping-gliding ight differences in energy consumptionbetween land and sea route become even smaller and only short detours areenergetically favourable By combining soaring and apping-gliding ightsmaller raptors such as falcons or harriers are able to y under varyingenvironmental conditions to extend their migratory activity to times whenthermal updrafts are absent and to migrate even at night (Stark amp Liechti1993 Spaar amp Bruderer 1997b Spaar et al 1998)
Flight altitudes
Flight altitudes differed between the species groups partly due to differencesin migratory routes Birds crossing the sea ew considerably higher thanthose following the coast Kerlinger (1984) made similar observations atthe Delaware Bay birds crossing the bay ew at higher altitudes thanthose keeping on land He explained the higher ight altitudes by the bettervisibility of the other shore However the African coast is not visible fromour study site Some falcons even started crossing the sea after sunset
396 MEYER SPAAR amp BRUDERER
indicating that they continued migration into the night these birds had to y at least three hours until they reached the African coast The higher ightaltitudes of birds crossing the sea at our study site was mainly caused bytopographical features birds ying with southerly directions were forced toclimb higher up when crossing the mountains chains (1500 m height) about10-15 km northward of the coast and they afterwards maintained these ightlevels when approaching the sea
Flight altitudes of birds migrating along the coast were similar to altitudesobserved in North America and southern Israel with most raptors yingbelow 1000 m above ground and lt 5 above 1500 m (Kerlinger 19841989 Bruderer et al 1994 Spaar 1996)
Flight characteristics in soaring-gliding
Flight characteristics in soaring-gliding ight did not differ between thespecies groups (Table 2) As shown for raptor migration in Israel climb-ing rates do not depend primarily on morphological features but on environ-mental conditions (Spaar 1997) In general mean climbing rates in southernSpain were lower than in the semi-arid desert in Israel (Spaar 1997) Meanduration of the completed circling phases was shorter in Israel (88 plusmn 52 sSpaar 1995 1999) than in Spain In contrast to Israel (Spaar 1995 1997)circling time did not increase with increasing climbing rate Compared tothe semi-desert the development of thermal convection in southern Spainwas probably less stable and birds circled as long as possible in updraftsbecause they could not rely to encounter a next strong thermal within glid-ing distance This might also explain why inter-thermal gliding was oftensupported by apping it reduces sinking rate and prolongs gliding distance
In raptors migrating along the coast ight altitude and gliding airspeedswere related to thermal strength raptors circling in strong thermals hadhigher ight altitudes and higher inter-thermal gliding airspeeds than thosein weak thermals This is consistent with optimal ight theory for birds max-imising cross-country speed in soaring-gliding ight (Pennycuick 1989) asdemonstrated for steppe buzzards Buteo buteo vulpinus (Spaar amp Bruderer1997a)
Flight characteristics in horizontal apping-gliding
According to optimal ight theory maximum range speed decreases withfollowing winds and increases with head- and sidewinds (Pennycuick 1975
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 397
Liechti et al 1994) In raptors ying along the coast we observed decreas-ing airspeeds with increasing tailwind-support whereas birds crossing thesea did not reduce airspeeds Why did the birds react differently Sea cross-ings imply higher risks there is no possibility to land and weather (wind)conditions may change rapidly Higher airspeeds in following winds lead toshorter crossing times and lower the risk of unpredictable weather changesFurthermore drift by sidewinds may considerably extent crossing distancesand in order to reduce drift raptors increased airspeeds with increasingsindwind-components In contrast to sharp-shinned hawks Accipiter stria-tus avoiding crossings in strong lateral winds (Kerlinger 1985) we observedmany raptors to cross the sea in strong north-westerly (ie lateral) winds ifthey provided at least partial tailwind-support
Conclusions
Our observations at an average site at the southern coast of Spain showthat differences in water crossing behaviour are mainly species-speci cpresumably due to the costs for different ight strategies additionallyindividuals reacted to environmental factors such as wind and thermalconditions In medium sized raptors such as honey buzzard soaring-gliding ight along the coast is energy-minimising whereas crossing the sea istime-minimising Sea crossings occurred preferably with a good tailwind-support but also under strong sidewinds Raptors crossing the sea did notreduce airspeeds in following winds but increased airspeeds with increasingsidewinds They seemed to minimise the water crossing duration and theeffect of drift by sidewinds Under good thermal conditions birds regularlyfollowed the coast despite a lower tailwind-support compared to crossing thesea
References
Agostini N amp Logozzo D (1995) Autumn migration of honey buzzards in Southern Italymdash J Raptor Res 29 p 275-277
mdash mdash Logozzo D amp Colero C (1997) Migration of ocks of honey buzzards in SouthernItaly and Malta mdash J Raptor Res 31 p 84-86
mdash mdash Malara G Neri F Mollicone D amp Melotto S (1994) Flight strategies of honeybuzzards during spring migration across the Central Mediterranean mdash Avocetta 18p 73-76
398 MEYER SPAAR amp BRUDERER
Bannermann D-A amp Bannermann WM (1971) Handbook of the birds of Cyprus andmigrants of the Middle East mdash Oliver amp Boyd Edinburgh
Batschelet E (1981) Circular statistics in biology mdash Academic Press LondonBaudinette RV amp Schmidt-Nielsen K (1974) Energy costs of gliding ight in herring
gulls mdash Nature 248 p 83-84Beaman M amp Galea C (1974) The visible migration of raptors over the Maltese Islands
mdash Ibis 116 p 419-431Bernis F (1975) Migracion de Falconiformes y Ciconea spp por Gibraltar mdash IV Avance
sobre recuentos en el verano-otontildeo 1974 mdash Ardeola 21 p 581-592Bijlsma RG (1987) Bottleneck areas for migratory birds in the Mediterranean region mdash
ICBP Study report No 18 CambridgeBruderer B amp Liechti F (1998) Flight behaviour of nocturnally migrating birds in coastal
areas mdash crossing or coasting mdash J Avian Biol 29 p 499-510mdash mdash Blitzblau S amp Peter D (1994) Migration and ight behaviour of honey buzzards
Pernis apivorus in southern Israel observed by radar mdash Ardea 82 p 111-122mdash mdash Steuri T amp Baumgartner M (1995) Short-range high-precision surveillance of
nocturnal migration and tracking of single targets mdash Israel J Zool 41 p 207-220Evans PR amp Lathbury GW (1972) Raptor migration across the Strait of Gibraltar mdash Ibis
115 p 572-585Galea C amp Massa B (1985) Notes on the raptor migration across the central Mediter-
ranean mdash In Conservation studies on raptors (I Newton amp RD Chancellor eds)ICBP Technical Publication No 5 p 257-261
Gensboslashl B (1997) Greifvoumlgel mdash BLV VerlagsgesellschaftMuumlnchenGustin M amp Pizzari T (1998) Migratory pattern in the genus Circus sex and age
differential migration in Italy mdash Ornis Svecica 8 p 23-26Kerlinger P (1984) Flight behaviour of sharp-shinned hawks during migration 2 Over
water mdash Anim Beh 32 p 1029-1034mdash mdash (1985) Water-crossing behaviour of raptors during migration mdash Wilson Bull 97
p 109-113mdash mdash (1989) Flight strategies of migrating hawks mdash University of Chicago Press Chicagomdash mdash Bingman VP amp Able KP (1985) Comparative ight behaviour of migrating hawks
studied with tracking radar during autumn in central New York mdash Can J Zool 63p 755-761
Kjelleacuten N (1992) Differential timing of autumn migration between sex and age groups inraptors at Falsterbo Sweden mdash Orn Scan 23 p 420-434
Liechti F (1993) Naumlchtlicher Vogelzug im Herbst uumlber Suumlddeutschland Winddrift undKompensation mdash J Ornithol 134 p 373-404
mdash mdash Hedenstroumlm A amp Alerstam T (1994) Effects of sidewinds on optimal ight speedsof birds mdash J theor Biol 170 p 219-225
Lustick S Batterby B amp Kelty M (1978) Behavioural thermoregulation orientationtoward the sun in herring gulls mdash Sience 200 p 81-83
Pennycuick C (1975) Mechanics of ight mdash In Avian biology Vol 5 (DS Farner ampRJ King eds) Academic Press New York p 1-75
mdash mdash (1989) Bird ight performance a practical calculation manual mdash Oxford UniversityPress Oxford
mdash mdash (1997) Actual and lsquooptimumrsquo ight speeds eld data reassessedmdash J Exp Biol 200p 2355-2361
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 399
Richardson WJ (1978) Timing and amount of bird migration in relation to weather areview mdash Oikos 30 p 224-272
Spaar R (1995) Flight behaviour of steppe buzzard (Buteo buteo vulpinus) during springmigration in southern Israel a tracking radar study mdash Israel J Zool 41 p 489-500
mdash mdash (1996) Flight behaviour of migrating raptors in southern Israel mdash PhD thesisUniversity of Basel Verlag Schweizerische Vogelwarte Sempach
mdash mdash (1997) Flight strategies of migrating raptors a comparative study of interspecicvariation in ight characteristicsmdash Ibis 139 p 523-535
mdash mdash (1999) Flight behaviour of migrating raptors under varying environmental conditionsmdash In Proc 22 Int Ornithol Congr (N Adams amp R Slotow eds) University of NatalDurban p 1844-1862
mdash mdash amp Bruderer B (1997a) Optimal ight behaviour of soaring migrants A case studyof migrating steppe buzzards Buteo buteo vulpinus mdash Behav Ecol 8 p 288-297)
mdash mdash amp Bruderer B (1997b) Migration by apping or soaring ight strategies of marshMontagursquos and pallid harriers in southern Israel mdash Condor 99 p 458-469
mdash mdash Stark H amp Liechti F (1998) Migratory ight strategies of Levant sparrowhawkstime or energy minimization mdash Anim Behav 56 p 1185-1197
Stark H amp Liechti F (1993) Do Levant sparrowhawks Accipiter brevipes also migrate atnight mdash Ibis 135 p 233-236
Thiollay J-M (1977) Importance des populations de rapaces migrateurs en mediterraneeoccidentale mdash Alauda 45 p 115-121

MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 395
TABLE 3 Observed airspeeds and corresponding estimated ight durationand fat consumption for an average honey buzzard when ying along the
coast or across the sea in different ight styles
Flight style Soaring-gliding Horizontal Horizontal apping-gliding apping-gliding
Flight route Flying along the coast Flying along the coast Crossing the seaDistance 250 km 250 km 150 km
Airspeed ms 101 (81-123) 108 (88-133) 108 (88-133)Flight duration hmin 652 (538-834) 626 (513-753) 351 (308-444)Fat consumption g 41 (34-55) 266 (230-298) 158 (138-179)for ight
Average honey buzzard = body mass 08 kg wing span 13 m wing area 025 m2 Medianand 25-75 range are given in brackets Calculations on fat consumption are based onPennycuickrsquos (1989) program of bird ight performance version 11 and birds are assumedto maximize cross-country speed in soaring-gliding ight (Spaar amp Bruderer 1997a)
strategy because a large proportion of the birds followed the coast in soaring-gliding ight
For species such as falcons or small harriers the difference in energyconsumption between apping and soaring-gliding ight is smaller Asthese species regularly use apping-gliding ight also over land or combinesoaring with apping-gliding ight differences in energy consumptionbetween land and sea route become even smaller and only short detours areenergetically favourable By combining soaring and apping-gliding ightsmaller raptors such as falcons or harriers are able to y under varyingenvironmental conditions to extend their migratory activity to times whenthermal updrafts are absent and to migrate even at night (Stark amp Liechti1993 Spaar amp Bruderer 1997b Spaar et al 1998)
Flight altitudes
Flight altitudes differed between the species groups partly due to differencesin migratory routes Birds crossing the sea ew considerably higher thanthose following the coast Kerlinger (1984) made similar observations atthe Delaware Bay birds crossing the bay ew at higher altitudes thanthose keeping on land He explained the higher ight altitudes by the bettervisibility of the other shore However the African coast is not visible fromour study site Some falcons even started crossing the sea after sunset
396 MEYER SPAAR amp BRUDERER
indicating that they continued migration into the night these birds had to y at least three hours until they reached the African coast The higher ightaltitudes of birds crossing the sea at our study site was mainly caused bytopographical features birds ying with southerly directions were forced toclimb higher up when crossing the mountains chains (1500 m height) about10-15 km northward of the coast and they afterwards maintained these ightlevels when approaching the sea
Flight altitudes of birds migrating along the coast were similar to altitudesobserved in North America and southern Israel with most raptors yingbelow 1000 m above ground and lt 5 above 1500 m (Kerlinger 19841989 Bruderer et al 1994 Spaar 1996)
Flight characteristics in soaring-gliding
Flight characteristics in soaring-gliding ight did not differ between thespecies groups (Table 2) As shown for raptor migration in Israel climb-ing rates do not depend primarily on morphological features but on environ-mental conditions (Spaar 1997) In general mean climbing rates in southernSpain were lower than in the semi-arid desert in Israel (Spaar 1997) Meanduration of the completed circling phases was shorter in Israel (88 plusmn 52 sSpaar 1995 1999) than in Spain In contrast to Israel (Spaar 1995 1997)circling time did not increase with increasing climbing rate Compared tothe semi-desert the development of thermal convection in southern Spainwas probably less stable and birds circled as long as possible in updraftsbecause they could not rely to encounter a next strong thermal within glid-ing distance This might also explain why inter-thermal gliding was oftensupported by apping it reduces sinking rate and prolongs gliding distance
In raptors migrating along the coast ight altitude and gliding airspeedswere related to thermal strength raptors circling in strong thermals hadhigher ight altitudes and higher inter-thermal gliding airspeeds than thosein weak thermals This is consistent with optimal ight theory for birds max-imising cross-country speed in soaring-gliding ight (Pennycuick 1989) asdemonstrated for steppe buzzards Buteo buteo vulpinus (Spaar amp Bruderer1997a)
Flight characteristics in horizontal apping-gliding
According to optimal ight theory maximum range speed decreases withfollowing winds and increases with head- and sidewinds (Pennycuick 1975
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 397
Liechti et al 1994) In raptors ying along the coast we observed decreas-ing airspeeds with increasing tailwind-support whereas birds crossing thesea did not reduce airspeeds Why did the birds react differently Sea cross-ings imply higher risks there is no possibility to land and weather (wind)conditions may change rapidly Higher airspeeds in following winds lead toshorter crossing times and lower the risk of unpredictable weather changesFurthermore drift by sidewinds may considerably extent crossing distancesand in order to reduce drift raptors increased airspeeds with increasingsindwind-components In contrast to sharp-shinned hawks Accipiter stria-tus avoiding crossings in strong lateral winds (Kerlinger 1985) we observedmany raptors to cross the sea in strong north-westerly (ie lateral) winds ifthey provided at least partial tailwind-support
Conclusions
Our observations at an average site at the southern coast of Spain showthat differences in water crossing behaviour are mainly species-speci cpresumably due to the costs for different ight strategies additionallyindividuals reacted to environmental factors such as wind and thermalconditions In medium sized raptors such as honey buzzard soaring-gliding ight along the coast is energy-minimising whereas crossing the sea istime-minimising Sea crossings occurred preferably with a good tailwind-support but also under strong sidewinds Raptors crossing the sea did notreduce airspeeds in following winds but increased airspeeds with increasingsidewinds They seemed to minimise the water crossing duration and theeffect of drift by sidewinds Under good thermal conditions birds regularlyfollowed the coast despite a lower tailwind-support compared to crossing thesea
References
Agostini N amp Logozzo D (1995) Autumn migration of honey buzzards in Southern Italymdash J Raptor Res 29 p 275-277
mdash mdash Logozzo D amp Colero C (1997) Migration of ocks of honey buzzards in SouthernItaly and Malta mdash J Raptor Res 31 p 84-86
mdash mdash Malara G Neri F Mollicone D amp Melotto S (1994) Flight strategies of honeybuzzards during spring migration across the Central Mediterranean mdash Avocetta 18p 73-76
398 MEYER SPAAR amp BRUDERER
Bannermann D-A amp Bannermann WM (1971) Handbook of the birds of Cyprus andmigrants of the Middle East mdash Oliver amp Boyd Edinburgh
Batschelet E (1981) Circular statistics in biology mdash Academic Press LondonBaudinette RV amp Schmidt-Nielsen K (1974) Energy costs of gliding ight in herring
gulls mdash Nature 248 p 83-84Beaman M amp Galea C (1974) The visible migration of raptors over the Maltese Islands
mdash Ibis 116 p 419-431Bernis F (1975) Migracion de Falconiformes y Ciconea spp por Gibraltar mdash IV Avance
sobre recuentos en el verano-otontildeo 1974 mdash Ardeola 21 p 581-592Bijlsma RG (1987) Bottleneck areas for migratory birds in the Mediterranean region mdash
ICBP Study report No 18 CambridgeBruderer B amp Liechti F (1998) Flight behaviour of nocturnally migrating birds in coastal
areas mdash crossing or coasting mdash J Avian Biol 29 p 499-510mdash mdash Blitzblau S amp Peter D (1994) Migration and ight behaviour of honey buzzards
Pernis apivorus in southern Israel observed by radar mdash Ardea 82 p 111-122mdash mdash Steuri T amp Baumgartner M (1995) Short-range high-precision surveillance of
nocturnal migration and tracking of single targets mdash Israel J Zool 41 p 207-220Evans PR amp Lathbury GW (1972) Raptor migration across the Strait of Gibraltar mdash Ibis
115 p 572-585Galea C amp Massa B (1985) Notes on the raptor migration across the central Mediter-
ranean mdash In Conservation studies on raptors (I Newton amp RD Chancellor eds)ICBP Technical Publication No 5 p 257-261
Gensboslashl B (1997) Greifvoumlgel mdash BLV VerlagsgesellschaftMuumlnchenGustin M amp Pizzari T (1998) Migratory pattern in the genus Circus sex and age
differential migration in Italy mdash Ornis Svecica 8 p 23-26Kerlinger P (1984) Flight behaviour of sharp-shinned hawks during migration 2 Over
water mdash Anim Beh 32 p 1029-1034mdash mdash (1985) Water-crossing behaviour of raptors during migration mdash Wilson Bull 97
p 109-113mdash mdash (1989) Flight strategies of migrating hawks mdash University of Chicago Press Chicagomdash mdash Bingman VP amp Able KP (1985) Comparative ight behaviour of migrating hawks
studied with tracking radar during autumn in central New York mdash Can J Zool 63p 755-761
Kjelleacuten N (1992) Differential timing of autumn migration between sex and age groups inraptors at Falsterbo Sweden mdash Orn Scan 23 p 420-434
Liechti F (1993) Naumlchtlicher Vogelzug im Herbst uumlber Suumlddeutschland Winddrift undKompensation mdash J Ornithol 134 p 373-404
mdash mdash Hedenstroumlm A amp Alerstam T (1994) Effects of sidewinds on optimal ight speedsof birds mdash J theor Biol 170 p 219-225
Lustick S Batterby B amp Kelty M (1978) Behavioural thermoregulation orientationtoward the sun in herring gulls mdash Sience 200 p 81-83
Pennycuick C (1975) Mechanics of ight mdash In Avian biology Vol 5 (DS Farner ampRJ King eds) Academic Press New York p 1-75
mdash mdash (1989) Bird ight performance a practical calculation manual mdash Oxford UniversityPress Oxford
mdash mdash (1997) Actual and lsquooptimumrsquo ight speeds eld data reassessedmdash J Exp Biol 200p 2355-2361
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 399
Richardson WJ (1978) Timing and amount of bird migration in relation to weather areview mdash Oikos 30 p 224-272
Spaar R (1995) Flight behaviour of steppe buzzard (Buteo buteo vulpinus) during springmigration in southern Israel a tracking radar study mdash Israel J Zool 41 p 489-500
mdash mdash (1996) Flight behaviour of migrating raptors in southern Israel mdash PhD thesisUniversity of Basel Verlag Schweizerische Vogelwarte Sempach
mdash mdash (1997) Flight strategies of migrating raptors a comparative study of interspecicvariation in ight characteristicsmdash Ibis 139 p 523-535
mdash mdash (1999) Flight behaviour of migrating raptors under varying environmental conditionsmdash In Proc 22 Int Ornithol Congr (N Adams amp R Slotow eds) University of NatalDurban p 1844-1862
mdash mdash amp Bruderer B (1997a) Optimal ight behaviour of soaring migrants A case studyof migrating steppe buzzards Buteo buteo vulpinus mdash Behav Ecol 8 p 288-297)
mdash mdash amp Bruderer B (1997b) Migration by apping or soaring ight strategies of marshMontagursquos and pallid harriers in southern Israel mdash Condor 99 p 458-469
mdash mdash Stark H amp Liechti F (1998) Migratory ight strategies of Levant sparrowhawkstime or energy minimization mdash Anim Behav 56 p 1185-1197
Stark H amp Liechti F (1993) Do Levant sparrowhawks Accipiter brevipes also migrate atnight mdash Ibis 135 p 233-236
Thiollay J-M (1977) Importance des populations de rapaces migrateurs en mediterraneeoccidentale mdash Alauda 45 p 115-121

396 MEYER SPAAR amp BRUDERER
indicating that they continued migration into the night these birds had to y at least three hours until they reached the African coast The higher ightaltitudes of birds crossing the sea at our study site was mainly caused bytopographical features birds ying with southerly directions were forced toclimb higher up when crossing the mountains chains (1500 m height) about10-15 km northward of the coast and they afterwards maintained these ightlevels when approaching the sea
Flight altitudes of birds migrating along the coast were similar to altitudesobserved in North America and southern Israel with most raptors yingbelow 1000 m above ground and lt 5 above 1500 m (Kerlinger 19841989 Bruderer et al 1994 Spaar 1996)
Flight characteristics in soaring-gliding
Flight characteristics in soaring-gliding ight did not differ between thespecies groups (Table 2) As shown for raptor migration in Israel climb-ing rates do not depend primarily on morphological features but on environ-mental conditions (Spaar 1997) In general mean climbing rates in southernSpain were lower than in the semi-arid desert in Israel (Spaar 1997) Meanduration of the completed circling phases was shorter in Israel (88 plusmn 52 sSpaar 1995 1999) than in Spain In contrast to Israel (Spaar 1995 1997)circling time did not increase with increasing climbing rate Compared tothe semi-desert the development of thermal convection in southern Spainwas probably less stable and birds circled as long as possible in updraftsbecause they could not rely to encounter a next strong thermal within glid-ing distance This might also explain why inter-thermal gliding was oftensupported by apping it reduces sinking rate and prolongs gliding distance
In raptors migrating along the coast ight altitude and gliding airspeedswere related to thermal strength raptors circling in strong thermals hadhigher ight altitudes and higher inter-thermal gliding airspeeds than thosein weak thermals This is consistent with optimal ight theory for birds max-imising cross-country speed in soaring-gliding ight (Pennycuick 1989) asdemonstrated for steppe buzzards Buteo buteo vulpinus (Spaar amp Bruderer1997a)
Flight characteristics in horizontal apping-gliding
According to optimal ight theory maximum range speed decreases withfollowing winds and increases with head- and sidewinds (Pennycuick 1975
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 397
Liechti et al 1994) In raptors ying along the coast we observed decreas-ing airspeeds with increasing tailwind-support whereas birds crossing thesea did not reduce airspeeds Why did the birds react differently Sea cross-ings imply higher risks there is no possibility to land and weather (wind)conditions may change rapidly Higher airspeeds in following winds lead toshorter crossing times and lower the risk of unpredictable weather changesFurthermore drift by sidewinds may considerably extent crossing distancesand in order to reduce drift raptors increased airspeeds with increasingsindwind-components In contrast to sharp-shinned hawks Accipiter stria-tus avoiding crossings in strong lateral winds (Kerlinger 1985) we observedmany raptors to cross the sea in strong north-westerly (ie lateral) winds ifthey provided at least partial tailwind-support
Conclusions
Our observations at an average site at the southern coast of Spain showthat differences in water crossing behaviour are mainly species-speci cpresumably due to the costs for different ight strategies additionallyindividuals reacted to environmental factors such as wind and thermalconditions In medium sized raptors such as honey buzzard soaring-gliding ight along the coast is energy-minimising whereas crossing the sea istime-minimising Sea crossings occurred preferably with a good tailwind-support but also under strong sidewinds Raptors crossing the sea did notreduce airspeeds in following winds but increased airspeeds with increasingsidewinds They seemed to minimise the water crossing duration and theeffect of drift by sidewinds Under good thermal conditions birds regularlyfollowed the coast despite a lower tailwind-support compared to crossing thesea
References
Agostini N amp Logozzo D (1995) Autumn migration of honey buzzards in Southern Italymdash J Raptor Res 29 p 275-277
mdash mdash Logozzo D amp Colero C (1997) Migration of ocks of honey buzzards in SouthernItaly and Malta mdash J Raptor Res 31 p 84-86
mdash mdash Malara G Neri F Mollicone D amp Melotto S (1994) Flight strategies of honeybuzzards during spring migration across the Central Mediterranean mdash Avocetta 18p 73-76
398 MEYER SPAAR amp BRUDERER
Bannermann D-A amp Bannermann WM (1971) Handbook of the birds of Cyprus andmigrants of the Middle East mdash Oliver amp Boyd Edinburgh
Batschelet E (1981) Circular statistics in biology mdash Academic Press LondonBaudinette RV amp Schmidt-Nielsen K (1974) Energy costs of gliding ight in herring
gulls mdash Nature 248 p 83-84Beaman M amp Galea C (1974) The visible migration of raptors over the Maltese Islands
mdash Ibis 116 p 419-431Bernis F (1975) Migracion de Falconiformes y Ciconea spp por Gibraltar mdash IV Avance
sobre recuentos en el verano-otontildeo 1974 mdash Ardeola 21 p 581-592Bijlsma RG (1987) Bottleneck areas for migratory birds in the Mediterranean region mdash
ICBP Study report No 18 CambridgeBruderer B amp Liechti F (1998) Flight behaviour of nocturnally migrating birds in coastal
areas mdash crossing or coasting mdash J Avian Biol 29 p 499-510mdash mdash Blitzblau S amp Peter D (1994) Migration and ight behaviour of honey buzzards
Pernis apivorus in southern Israel observed by radar mdash Ardea 82 p 111-122mdash mdash Steuri T amp Baumgartner M (1995) Short-range high-precision surveillance of
nocturnal migration and tracking of single targets mdash Israel J Zool 41 p 207-220Evans PR amp Lathbury GW (1972) Raptor migration across the Strait of Gibraltar mdash Ibis
115 p 572-585Galea C amp Massa B (1985) Notes on the raptor migration across the central Mediter-
ranean mdash In Conservation studies on raptors (I Newton amp RD Chancellor eds)ICBP Technical Publication No 5 p 257-261
Gensboslashl B (1997) Greifvoumlgel mdash BLV VerlagsgesellschaftMuumlnchenGustin M amp Pizzari T (1998) Migratory pattern in the genus Circus sex and age
differential migration in Italy mdash Ornis Svecica 8 p 23-26Kerlinger P (1984) Flight behaviour of sharp-shinned hawks during migration 2 Over
water mdash Anim Beh 32 p 1029-1034mdash mdash (1985) Water-crossing behaviour of raptors during migration mdash Wilson Bull 97
p 109-113mdash mdash (1989) Flight strategies of migrating hawks mdash University of Chicago Press Chicagomdash mdash Bingman VP amp Able KP (1985) Comparative ight behaviour of migrating hawks
studied with tracking radar during autumn in central New York mdash Can J Zool 63p 755-761
Kjelleacuten N (1992) Differential timing of autumn migration between sex and age groups inraptors at Falsterbo Sweden mdash Orn Scan 23 p 420-434
Liechti F (1993) Naumlchtlicher Vogelzug im Herbst uumlber Suumlddeutschland Winddrift undKompensation mdash J Ornithol 134 p 373-404
mdash mdash Hedenstroumlm A amp Alerstam T (1994) Effects of sidewinds on optimal ight speedsof birds mdash J theor Biol 170 p 219-225
Lustick S Batterby B amp Kelty M (1978) Behavioural thermoregulation orientationtoward the sun in herring gulls mdash Sience 200 p 81-83
Pennycuick C (1975) Mechanics of ight mdash In Avian biology Vol 5 (DS Farner ampRJ King eds) Academic Press New York p 1-75
mdash mdash (1989) Bird ight performance a practical calculation manual mdash Oxford UniversityPress Oxford
mdash mdash (1997) Actual and lsquooptimumrsquo ight speeds eld data reassessedmdash J Exp Biol 200p 2355-2361
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 399
Richardson WJ (1978) Timing and amount of bird migration in relation to weather areview mdash Oikos 30 p 224-272
Spaar R (1995) Flight behaviour of steppe buzzard (Buteo buteo vulpinus) during springmigration in southern Israel a tracking radar study mdash Israel J Zool 41 p 489-500
mdash mdash (1996) Flight behaviour of migrating raptors in southern Israel mdash PhD thesisUniversity of Basel Verlag Schweizerische Vogelwarte Sempach
mdash mdash (1997) Flight strategies of migrating raptors a comparative study of interspecicvariation in ight characteristicsmdash Ibis 139 p 523-535
mdash mdash (1999) Flight behaviour of migrating raptors under varying environmental conditionsmdash In Proc 22 Int Ornithol Congr (N Adams amp R Slotow eds) University of NatalDurban p 1844-1862
mdash mdash amp Bruderer B (1997a) Optimal ight behaviour of soaring migrants A case studyof migrating steppe buzzards Buteo buteo vulpinus mdash Behav Ecol 8 p 288-297)
mdash mdash amp Bruderer B (1997b) Migration by apping or soaring ight strategies of marshMontagursquos and pallid harriers in southern Israel mdash Condor 99 p 458-469
mdash mdash Stark H amp Liechti F (1998) Migratory ight strategies of Levant sparrowhawkstime or energy minimization mdash Anim Behav 56 p 1185-1197
Stark H amp Liechti F (1993) Do Levant sparrowhawks Accipiter brevipes also migrate atnight mdash Ibis 135 p 233-236
Thiollay J-M (1977) Importance des populations de rapaces migrateurs en mediterraneeoccidentale mdash Alauda 45 p 115-121

MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 397
Liechti et al 1994) In raptors ying along the coast we observed decreas-ing airspeeds with increasing tailwind-support whereas birds crossing thesea did not reduce airspeeds Why did the birds react differently Sea cross-ings imply higher risks there is no possibility to land and weather (wind)conditions may change rapidly Higher airspeeds in following winds lead toshorter crossing times and lower the risk of unpredictable weather changesFurthermore drift by sidewinds may considerably extent crossing distancesand in order to reduce drift raptors increased airspeeds with increasingsindwind-components In contrast to sharp-shinned hawks Accipiter stria-tus avoiding crossings in strong lateral winds (Kerlinger 1985) we observedmany raptors to cross the sea in strong north-westerly (ie lateral) winds ifthey provided at least partial tailwind-support
Conclusions
Our observations at an average site at the southern coast of Spain showthat differences in water crossing behaviour are mainly species-speci cpresumably due to the costs for different ight strategies additionallyindividuals reacted to environmental factors such as wind and thermalconditions In medium sized raptors such as honey buzzard soaring-gliding ight along the coast is energy-minimising whereas crossing the sea istime-minimising Sea crossings occurred preferably with a good tailwind-support but also under strong sidewinds Raptors crossing the sea did notreduce airspeeds in following winds but increased airspeeds with increasingsidewinds They seemed to minimise the water crossing duration and theeffect of drift by sidewinds Under good thermal conditions birds regularlyfollowed the coast despite a lower tailwind-support compared to crossing thesea
References
Agostini N amp Logozzo D (1995) Autumn migration of honey buzzards in Southern Italymdash J Raptor Res 29 p 275-277
mdash mdash Logozzo D amp Colero C (1997) Migration of ocks of honey buzzards in SouthernItaly and Malta mdash J Raptor Res 31 p 84-86
mdash mdash Malara G Neri F Mollicone D amp Melotto S (1994) Flight strategies of honeybuzzards during spring migration across the Central Mediterranean mdash Avocetta 18p 73-76
398 MEYER SPAAR amp BRUDERER
Bannermann D-A amp Bannermann WM (1971) Handbook of the birds of Cyprus andmigrants of the Middle East mdash Oliver amp Boyd Edinburgh
Batschelet E (1981) Circular statistics in biology mdash Academic Press LondonBaudinette RV amp Schmidt-Nielsen K (1974) Energy costs of gliding ight in herring
gulls mdash Nature 248 p 83-84Beaman M amp Galea C (1974) The visible migration of raptors over the Maltese Islands
mdash Ibis 116 p 419-431Bernis F (1975) Migracion de Falconiformes y Ciconea spp por Gibraltar mdash IV Avance
sobre recuentos en el verano-otontildeo 1974 mdash Ardeola 21 p 581-592Bijlsma RG (1987) Bottleneck areas for migratory birds in the Mediterranean region mdash
ICBP Study report No 18 CambridgeBruderer B amp Liechti F (1998) Flight behaviour of nocturnally migrating birds in coastal
areas mdash crossing or coasting mdash J Avian Biol 29 p 499-510mdash mdash Blitzblau S amp Peter D (1994) Migration and ight behaviour of honey buzzards
Pernis apivorus in southern Israel observed by radar mdash Ardea 82 p 111-122mdash mdash Steuri T amp Baumgartner M (1995) Short-range high-precision surveillance of
nocturnal migration and tracking of single targets mdash Israel J Zool 41 p 207-220Evans PR amp Lathbury GW (1972) Raptor migration across the Strait of Gibraltar mdash Ibis
115 p 572-585Galea C amp Massa B (1985) Notes on the raptor migration across the central Mediter-
ranean mdash In Conservation studies on raptors (I Newton amp RD Chancellor eds)ICBP Technical Publication No 5 p 257-261
Gensboslashl B (1997) Greifvoumlgel mdash BLV VerlagsgesellschaftMuumlnchenGustin M amp Pizzari T (1998) Migratory pattern in the genus Circus sex and age
differential migration in Italy mdash Ornis Svecica 8 p 23-26Kerlinger P (1984) Flight behaviour of sharp-shinned hawks during migration 2 Over
water mdash Anim Beh 32 p 1029-1034mdash mdash (1985) Water-crossing behaviour of raptors during migration mdash Wilson Bull 97
p 109-113mdash mdash (1989) Flight strategies of migrating hawks mdash University of Chicago Press Chicagomdash mdash Bingman VP amp Able KP (1985) Comparative ight behaviour of migrating hawks
studied with tracking radar during autumn in central New York mdash Can J Zool 63p 755-761
Kjelleacuten N (1992) Differential timing of autumn migration between sex and age groups inraptors at Falsterbo Sweden mdash Orn Scan 23 p 420-434
Liechti F (1993) Naumlchtlicher Vogelzug im Herbst uumlber Suumlddeutschland Winddrift undKompensation mdash J Ornithol 134 p 373-404
mdash mdash Hedenstroumlm A amp Alerstam T (1994) Effects of sidewinds on optimal ight speedsof birds mdash J theor Biol 170 p 219-225
Lustick S Batterby B amp Kelty M (1978) Behavioural thermoregulation orientationtoward the sun in herring gulls mdash Sience 200 p 81-83
Pennycuick C (1975) Mechanics of ight mdash In Avian biology Vol 5 (DS Farner ampRJ King eds) Academic Press New York p 1-75
mdash mdash (1989) Bird ight performance a practical calculation manual mdash Oxford UniversityPress Oxford
mdash mdash (1997) Actual and lsquooptimumrsquo ight speeds eld data reassessedmdash J Exp Biol 200p 2355-2361
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 399
Richardson WJ (1978) Timing and amount of bird migration in relation to weather areview mdash Oikos 30 p 224-272
Spaar R (1995) Flight behaviour of steppe buzzard (Buteo buteo vulpinus) during springmigration in southern Israel a tracking radar study mdash Israel J Zool 41 p 489-500
mdash mdash (1996) Flight behaviour of migrating raptors in southern Israel mdash PhD thesisUniversity of Basel Verlag Schweizerische Vogelwarte Sempach
mdash mdash (1997) Flight strategies of migrating raptors a comparative study of interspecicvariation in ight characteristicsmdash Ibis 139 p 523-535
mdash mdash (1999) Flight behaviour of migrating raptors under varying environmental conditionsmdash In Proc 22 Int Ornithol Congr (N Adams amp R Slotow eds) University of NatalDurban p 1844-1862
mdash mdash amp Bruderer B (1997a) Optimal ight behaviour of soaring migrants A case studyof migrating steppe buzzards Buteo buteo vulpinus mdash Behav Ecol 8 p 288-297)
mdash mdash amp Bruderer B (1997b) Migration by apping or soaring ight strategies of marshMontagursquos and pallid harriers in southern Israel mdash Condor 99 p 458-469
mdash mdash Stark H amp Liechti F (1998) Migratory ight strategies of Levant sparrowhawkstime or energy minimization mdash Anim Behav 56 p 1185-1197
Stark H amp Liechti F (1993) Do Levant sparrowhawks Accipiter brevipes also migrate atnight mdash Ibis 135 p 233-236
Thiollay J-M (1977) Importance des populations de rapaces migrateurs en mediterraneeoccidentale mdash Alauda 45 p 115-121

398 MEYER SPAAR amp BRUDERER
Bannermann D-A amp Bannermann WM (1971) Handbook of the birds of Cyprus andmigrants of the Middle East mdash Oliver amp Boyd Edinburgh
Batschelet E (1981) Circular statistics in biology mdash Academic Press LondonBaudinette RV amp Schmidt-Nielsen K (1974) Energy costs of gliding ight in herring
gulls mdash Nature 248 p 83-84Beaman M amp Galea C (1974) The visible migration of raptors over the Maltese Islands
mdash Ibis 116 p 419-431Bernis F (1975) Migracion de Falconiformes y Ciconea spp por Gibraltar mdash IV Avance
sobre recuentos en el verano-otontildeo 1974 mdash Ardeola 21 p 581-592Bijlsma RG (1987) Bottleneck areas for migratory birds in the Mediterranean region mdash
ICBP Study report No 18 CambridgeBruderer B amp Liechti F (1998) Flight behaviour of nocturnally migrating birds in coastal
areas mdash crossing or coasting mdash J Avian Biol 29 p 499-510mdash mdash Blitzblau S amp Peter D (1994) Migration and ight behaviour of honey buzzards
Pernis apivorus in southern Israel observed by radar mdash Ardea 82 p 111-122mdash mdash Steuri T amp Baumgartner M (1995) Short-range high-precision surveillance of
nocturnal migration and tracking of single targets mdash Israel J Zool 41 p 207-220Evans PR amp Lathbury GW (1972) Raptor migration across the Strait of Gibraltar mdash Ibis
115 p 572-585Galea C amp Massa B (1985) Notes on the raptor migration across the central Mediter-
ranean mdash In Conservation studies on raptors (I Newton amp RD Chancellor eds)ICBP Technical Publication No 5 p 257-261
Gensboslashl B (1997) Greifvoumlgel mdash BLV VerlagsgesellschaftMuumlnchenGustin M amp Pizzari T (1998) Migratory pattern in the genus Circus sex and age
differential migration in Italy mdash Ornis Svecica 8 p 23-26Kerlinger P (1984) Flight behaviour of sharp-shinned hawks during migration 2 Over
water mdash Anim Beh 32 p 1029-1034mdash mdash (1985) Water-crossing behaviour of raptors during migration mdash Wilson Bull 97
p 109-113mdash mdash (1989) Flight strategies of migrating hawks mdash University of Chicago Press Chicagomdash mdash Bingman VP amp Able KP (1985) Comparative ight behaviour of migrating hawks
studied with tracking radar during autumn in central New York mdash Can J Zool 63p 755-761
Kjelleacuten N (1992) Differential timing of autumn migration between sex and age groups inraptors at Falsterbo Sweden mdash Orn Scan 23 p 420-434
Liechti F (1993) Naumlchtlicher Vogelzug im Herbst uumlber Suumlddeutschland Winddrift undKompensation mdash J Ornithol 134 p 373-404
mdash mdash Hedenstroumlm A amp Alerstam T (1994) Effects of sidewinds on optimal ight speedsof birds mdash J theor Biol 170 p 219-225
Lustick S Batterby B amp Kelty M (1978) Behavioural thermoregulation orientationtoward the sun in herring gulls mdash Sience 200 p 81-83
Pennycuick C (1975) Mechanics of ight mdash In Avian biology Vol 5 (DS Farner ampRJ King eds) Academic Press New York p 1-75
mdash mdash (1989) Bird ight performance a practical calculation manual mdash Oxford UniversityPress Oxford
mdash mdash (1997) Actual and lsquooptimumrsquo ight speeds eld data reassessedmdash J Exp Biol 200p 2355-2361
MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 399
Richardson WJ (1978) Timing and amount of bird migration in relation to weather areview mdash Oikos 30 p 224-272
Spaar R (1995) Flight behaviour of steppe buzzard (Buteo buteo vulpinus) during springmigration in southern Israel a tracking radar study mdash Israel J Zool 41 p 489-500
mdash mdash (1996) Flight behaviour of migrating raptors in southern Israel mdash PhD thesisUniversity of Basel Verlag Schweizerische Vogelwarte Sempach
mdash mdash (1997) Flight strategies of migrating raptors a comparative study of interspecicvariation in ight characteristicsmdash Ibis 139 p 523-535
mdash mdash (1999) Flight behaviour of migrating raptors under varying environmental conditionsmdash In Proc 22 Int Ornithol Congr (N Adams amp R Slotow eds) University of NatalDurban p 1844-1862
mdash mdash amp Bruderer B (1997a) Optimal ight behaviour of soaring migrants A case studyof migrating steppe buzzards Buteo buteo vulpinus mdash Behav Ecol 8 p 288-297)
mdash mdash amp Bruderer B (1997b) Migration by apping or soaring ight strategies of marshMontagursquos and pallid harriers in southern Israel mdash Condor 99 p 458-469
mdash mdash Stark H amp Liechti F (1998) Migratory ight strategies of Levant sparrowhawkstime or energy minimization mdash Anim Behav 56 p 1185-1197
Stark H amp Liechti F (1993) Do Levant sparrowhawks Accipiter brevipes also migrate atnight mdash Ibis 135 p 233-236
Thiollay J-M (1977) Importance des populations de rapaces migrateurs en mediterraneeoccidentale mdash Alauda 45 p 115-121

MIGRATING RAPTORS FLIGHT DIRECTION AND BEHAVIOUR 399
Richardson WJ (1978) Timing and amount of bird migration in relation to weather areview mdash Oikos 30 p 224-272
Spaar R (1995) Flight behaviour of steppe buzzard (Buteo buteo vulpinus) during springmigration in southern Israel a tracking radar study mdash Israel J Zool 41 p 489-500
mdash mdash (1996) Flight behaviour of migrating raptors in southern Israel mdash PhD thesisUniversity of Basel Verlag Schweizerische Vogelwarte Sempach
mdash mdash (1997) Flight strategies of migrating raptors a comparative study of interspecicvariation in ight characteristicsmdash Ibis 139 p 523-535
mdash mdash (1999) Flight behaviour of migrating raptors under varying environmental conditionsmdash In Proc 22 Int Ornithol Congr (N Adams amp R Slotow eds) University of NatalDurban p 1844-1862
mdash mdash amp Bruderer B (1997a) Optimal ight behaviour of soaring migrants A case studyof migrating steppe buzzards Buteo buteo vulpinus mdash Behav Ecol 8 p 288-297)
mdash mdash amp Bruderer B (1997b) Migration by apping or soaring ight strategies of marshMontagursquos and pallid harriers in southern Israel mdash Condor 99 p 458-469
mdash mdash Stark H amp Liechti F (1998) Migratory ight strategies of Levant sparrowhawkstime or energy minimization mdash Anim Behav 56 p 1185-1197
Stark H amp Liechti F (1993) Do Levant sparrowhawks Accipiter brevipes also migrate atnight mdash Ibis 135 p 233-236
Thiollay J-M (1977) Importance des populations de rapaces migrateurs en mediterraneeoccidentale mdash Alauda 45 p 115-121