thyroid hormone regulates tag-1 expression in the developing rat brain
TRANSCRIPT

Thyroid hormone regulates TAG-1 expression in thedeveloping rat brain
Manuel Alvarez-Dolado,1 Ange lica Figueroa,1 Serguei Kozlov,2,* Peter Sonderegger,2 Andrew J. Furley3 andAlberto MunÄoz1
1Instituto de Investigaciones BiomeÂdicas `Alberto Sols', Consejo Superior de Investigaciones Cientõ®cas (CSIC) ± Universidad
AutoÂnoma de Madrid (UAM), Arturo Duperier 4, 28029 Madrid, Spain2Department of Biochemistry, University of Zurich, CH-8057 Zurich, Switzerland3Developmental Genetics Programme, University of Shef®eld, Shef®eld S10 2TN, UK
Keywords: adhesion, axon, gene expression, hypothyroidism, neural differentiation
Abstract
TAG-1 is a member of the immunoglobulin superfamily of cell adhesion molecules thought to play important roles in neuronal
differentiation and the establishment of connectivity during brain development. Because these are processes also affected byhypothyroidism, we studied the effects of thyroid hormone deprivation and administration on TAG-1 expression in the developing
rat brain. By in situ hybridization, immunohistochemistry and Western blotting we found that TAG-1 RNA and protein levels are
upregulated in the hypothyroid brain. From embryonic day 20 to postnatal day (P) 15, elevated TAG-1 RNA was found in severalareas including the cerebral cortex, hippocampus and olfactory bulb. In agreement with this, TAG-1 protein was overexpressed in
the major ®bre tracts arising from these structures, including the corpus callosum, anterior and hippocampal commissures and
lateral olfactory tract. A similar overexpression of TAG-1 by hypothyroidism was detected in the cerebellum, but starting only atP15. In all cases, elevation of TAG-1 RNA and protein expression could be reversed by thyroid hormone treatment. These results
show that the deregulation of TAG-1 might contribute to the alterations caused by the lack of thyroid hormone during brain
development.
Introduction
The neural cell adhesion molecule (CAM) TAG-1, a member of the
immunoglobulin (Ig) superfamily (axonin-1 in chick; TAX-1/CNTN2
in human; Furley et al., 1990; Zuellig et al., 1992; Hasler et al., 1993),
is thought to play an important role in axonal growth and guidance,
neurite outgrowth and the formation of highly ordered neural
connections (Ruegg et al., 1989; Furley et al., 1990; Stoeckli et al.,
1991; Sonderegger & Rathjen, 1992; Stoeckli, 1997; Fitzli et al.,
2000). It is present as a surface component of the axonal "pathway
sensor" apparatus (Vogt et al., 1996), and has been implicated in the
migration and local differentiation of cerebellar granule cells (Fishell
& Hatten, 1991; Wolfer et al., 1994; Bailly et al., 1996; Buttiglione
et al., 1998). Although capable of homophilic binding, it is the
heterophilic interactions of TAG-1, particularly with L1/Ng-CAM and
NrCAM, that appear to be important in these processes (Kuhn et al.,
1991; Felsenfeld et al., 1994; Stoeckli et al., 1997).
TAG-1 is expressed on many classes of neurons from the earliest
stages of axonogenesis, when it can ®rst be found on cell bodies as
axons begin to form and later on growing axons and their growth
cones (Yamamoto et al., 1986; Dodd et al., 1988; Wolfer et al., 1994;
Yoshihara et al., 1995; Karagogeos et al., 1997). On several classes
of axon its expression is transient, notably on spinal commissural and
motor axons, where disappearance of TAG-1 coincides with axons
reaching intermediate or ®nal targets (Dodd et al., 1988).
Nonetheless, high levels of TAG-1 expression continue through to
the third postnatal week, most notably in the cerebellum and
neocortex where TAG-1 protein is found on differentiating granule
cells in the inner external granule cell layer (iEGL) and on
corticothalamic projections (Wolfer et al., 1994; Yoshihara et al.,
1995). However, even in these regions TAG-1 expression is precisely
controlled with the protein disappearing at speci®c stages of
axonogenesis.
Very little is known about how the expression of TAG-1 is
controlled, although in vitro promoter analysis studies have identi®ed
regions upstream of the ®rst exon that will direct expression in
transfected cultured lines (Giger et al., 1995; Kozlov et al., 1995).
Thyroid hormone (triiodothyronine, T3) and the prohormone
thyroxine (T4) plays a crucial role in CNS development
(Oppenheimer & Schwartz, 1997; Bernal & GuadanÄo, 1998), and is
known to regulate the expression of a number of proteins involved in
axonogenesis, including the IgCAMs MAG, NCAM and L1
(RodrõÂguez-PenÄa et al., 1993; Iglesias et al., 1996; Alvarez-Dolado
et al., 2000). Severe thyroid hormone de®ciency in humans leads to
cretinism, a syndrome associated with mental retardation and
neurological de®cits (DeLong, 1990). Hypothyroidism in rodents
causes a complex array of alterations which include abnormal
arborization and distribution of dendritic spines, de®cient neurite
outgrowth, aberrant patterns of connections, myelination defects,
abnormal cortical lamination, and a delay in the migration of
Correspondence: Dr A. MunÄoz, as above.Email: [email protected]
*Present address: Cancer and Developmental Biology Laboratory, ABL-BasicResearch Program, NCI-FCRDC, Bldg. 539, Room 121 A, PO Box B,Frederick, MD 21702±1201, USA
Received 30 March 2001, revised 14 July 2001, accepted 20 August 2001
European Journal of Neuroscience, Vol. 14, pp. 1209±1218, 2001 ã Federation of European Neuroscience Societies

cerebellar granule cells (Legrand, 1984; Dussault & Ruel, 1987). The
involvement of TAG-1 in many of the processes altered by
hypothyroidism, the fact that other IgCAMs are regulated by T3,
and the coincidence of the peaks of TAG-1 expression with T3 effects
in the CNS, led us to investigate the effects of thyroid status on
TAG-1. To our knowledge, this is the ®rst time that an epigenetic
factor, T3, has been shown to regulate TAG-1 expression.
Materials and methods
Preparation of hypothyroid animals
Wistar rats raised in our animal facilities were used. The maintenance
and handling of the animals were as recommended by the European
Communities Council Directive of November 24th, 1986 (86/609/
EEC) following the guidelines and approval of the Committee for
Animal Research of our Institute. All efforts were made to reduce the
number of animals used, and to use alternatives to in vivo techniques.
To induce embryonal and neonatal hypothyroidism, 2-mercapto-1-
methylimidazole (0.02% MMI, Sigma, St. Louis, MO, USA) and
potassium perchlorate (1% KClO4, Merck, Germany) were adminis-
tered in the drinking water of the dams from the ninth day after
conception until the animals were killed, always under adequate
anaesthesia (1 mL/100 mg body weight of a solution containing
2 mg/mL diazepam, 25 mg/mL ketamine and 0.1 mg/mL atropine).
Postnatal day (P) 0 animals were killed 8±12 h after birth. T4
treatment (Sigma) consisted of daily single intraperitoneal injections
of 1.8 mg/100 g body weight starting 4 days before death. Rats were
killed 24 h after the last T4 injection. For treatment of embryos
(5 mL/h, 3 mg T4/100 g body weight per day) an osmotic pump
(2ML2 model; Alzet, CA, USA) was subcutaneously implanted into
the dams (using the anaesthetic solution above). T3 treatments
(Sigma) consisted of a single intraperitoneal injection (1 mg/g body
weight) 8 h before the animals were killed. The results obtained in
the animal studies referred to consistent ®ndings based on the
analysis of sections from at least three animals per experimental
group. Hypothyroid animals showed an arrest of body weight
increase (24% on P15, 55% on P25) and lower levels of both T4
(from 1.46 to 0.022 ng/g, 66.3-fold) and T3 (from 4.4 to 0.2 ng/g; 22-
fold) in brain tissue as measured by a speci®c radioimmunoassay
similar to those described previously Morreale de Escobar et al.,
1985; GarcõÂa-FernaÂndez et al., 1997; Alvarez-Dolado et al., 2000).
Samples for in situ hybridization and immunohistochemistry
Under profound pentobarbitone anaesthesia (4 mg/100 g, Abbott,
Chicago, IL, USA), animals were perfused transcardially with saline
and then with cold 4% p-formaldehyde in 0.1 M sodium phosphate
(pH 7.4). The brains were removed quickly and cryoprotected in 4%
p-formaldehyde +30% sucrose (w/v) in phosphate-buffered saline
(PBS) at 4 °C. Subsequently, parallel serial 40 mm [for embryonal
day (E)20] or 25 mm (P0±P30) thick coronal sections were cut using a
cryostat (Leica Instruments, Germany).
Preparation of riboprobes
To obtain antisense or sense TAG-1 riboprobes a 1.6 kb fragment of
the rat TAG-1 cDNA subcloned into the Bluescript vector was
transcribed in vitro using T7 or T3 RNA polymerases, respectively, in
the presence of 50 mCi of 35S-UTP. Non-radioactive riboprobes were
prepared using a digoxigenin RNA labelling kit (Boehringer
Mannheim, Germany). Riboprobes were cleaned by centrifugation
through Chroma Spin-100 DEPC-H2O columns (Clontech, Palo Alto,
CA, USA).
In situ hybridization
In situ hybridization on ¯oating sections followed the procedure of
Gall & Isackson (1989). Sections were thawed, washed with PBS for
5 min and treated for an additional 10 min at room temperature under
free-¯oating conditions with 0.1% Triton X-100, 0.2 M HCl, 0.25%
acetic anhydride in 0.1 M triethanolamine, and post®xed with 4%
p-formaldehyde. The sections were then preincubated in hybridiz-
ation solution (0.6 M NaCl, 20 mM PIPES-Na, 10 mM EDTA, 50%
formamide, 0.2% SDS, 5 3 Denhardt's solution, 10% dextran
sulphate, 50 mM DTT, 250 mg/mL of sheared salmon sperm DNA
and 250 mg/mL of yeast tRNA) for 3±5 h at 55 °C, and later
incubated overnight at 55 °C in the same solution containing 250 ng/
mL of the digoxigenin-labelled riboprobe or the 35S-UTP-labelled
riboprobe (15 3 106 c.p.m./mL). Sections were consecutively
washed once in 2 3 standard saline citrate (1 3 SSC: 0.15 M
NaCl, 0.015 M sodium citrate) + 10 mM b-mercaptoethanol at room
temperature for 30 min, once in 5 3 TEN (50 mM Tris pH 7.5, 5 mM
EDTA, 0.5 M NaCl) supplemented with 4 mg/mL mRNAse at 37 °C
for 1 h, twice in 0.5 3 SSC + 50% formamide + 10 mM b-mer-
captoethanol at 55 °C for 1 h, once in 0.1 3 SSC + 10 mM b-
mercaptoethanol at 68 °C for 1 h, and ®nally once in PBS at room
temperature for 5 min. To develop the digoxigenin staining we used
the nucleic acid detection kit from Boehringer Mannheim (Germany).
For radioactive in situ hybridization, sections were mounted onto
slides, dehydrated by ethanol series (containing 0.3 M NH4Ac) and
exposed for 5 days to Hyper®lm b-MAX ®lms (Amersham-
Pharmacia Biotech, UK), developed with Kodak D19, and ®xed.
For anatomical abbreviations we followed those in Paxinos & Watson
(1998).
Immunohistochemistry
Tissue sections were incubated overnight with a 1 : 10 000±5000
dilution of a rabbit polyclonal anti-TAG-1 antibody (Dodd et al.,
1988; a gift from Dr T. Jessell) or with the 1C12 monoclonal anti-rat
TAG-1 antibody (Dodd et al., 1988; a gift from Dr D. Karagogeos) at
4 °C under free-¯oating conditions. Sections were incubated with a
goat biotinylated anti-rabbit or anti-mouse Ig (Vector Laboratories,
UK) at a dilution of 1 : 200 in 0.1 M PBS + 0.1% Triton X-100 + 3%
goat serum. Sections were then washed three times in 0.1 M
PBS + 0.1 M Triton X-100 and incubated for a further 30 min with
avidin-biotinylated horseradish peroxidase macromolecular complex
(ABC, Vector Laboratories, UK) at a dilution of 1 : 100 in 0.1 M
PBS. The IgG fraction obtained from nonimmunized rabbits was used
for control staining. The peroxidase activity was visualized using a
solution containing 0.5 mg/mL diaminobenzidine (Sigma) and 0.3%
H202 in 0.1 M PBS. Sections were washed with 0.1 M PBS, and
mounted onto slides. After air-drying, they were dehydrated in graded
alcohol solutions, cleared in xylene and ®nally coverslipped with
DePeX mounting medium (Sigma).
Western blotting
Membrane-enriched protein extracts for immunoblot analysis of L1
expression were obtained as described (Rettig et al., 1992). Brie¯y,
brain tissue (cerebral cortex or cerebellum) was homogenized in 7
volumes of chilled, high pH buffer (30 mM diethylamine, 1 mM
EDTA, pH 11.5), cleared by centrifugation at 900 3 g for 15 min at
4 °C, and ®nally centrifuged at 100 000 3 g for 30 min at the same
temperature. Aliquots of the pellets containing 25±50 mg protein
were electrophoresed in 6% polyacrylamide gels and transferred to
nylon PVDF membranes overnight using a vertical transfer apparatus
(Bio-Rad Laboratories, CA, USA) at 1 mA/cm2. Care was taken to
1210 M. Alvarez-Dolado et al.
ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 14, 1209±1218

ensure equal loading of individual extracts by measuring the protein
concentration. The ®lters were washed, blocked with Blotto (5%
skimmed milk in PBS), and incubated with the TG3 rabbit polyclonal
anti-TAG-1 antibody (generously donated by Dr T. Jessel) at a
1 : 10 000 dilution. The blots were developed by a peroxidase
reaction using the ECL detection system (Amersham-Pharmacia
Biotech, UK).
Results
In situ hybridization and immunohistochemical studies
To determine whether TAG-1 expression is regulated by thyroid
hormone in the developing rat brain, we performed in situ
hybridization and immunohistochemical studies of embryonal and
postnatal control and hypothyroid rats.
E20
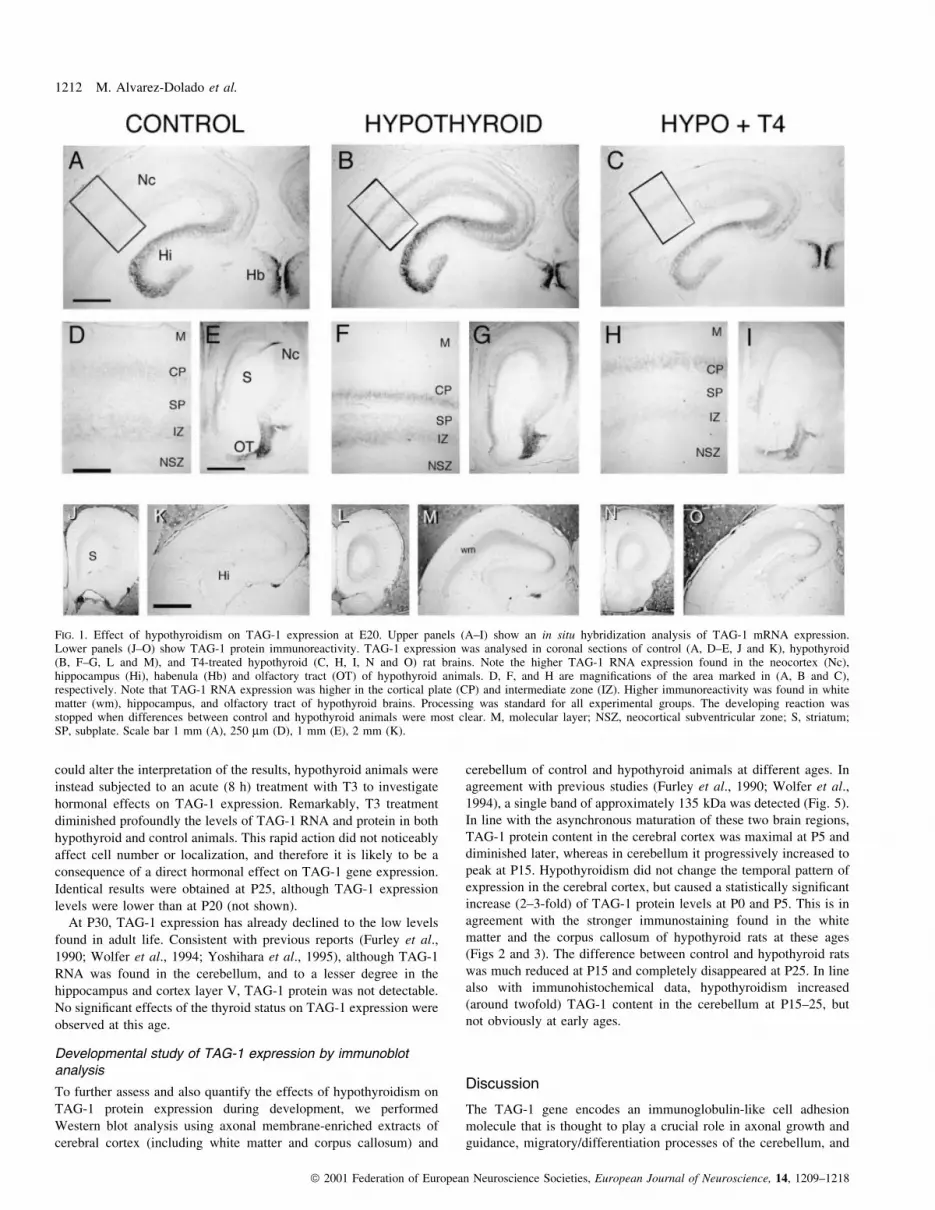
In agreement with previous reports (Yoshihara et al., 1995), in situ
hybridization analysis showed that in control animals TAG-1
transcripts were prominent in the cortical plate (CP), intermediate
zone (IZ) of the neocortex, hippocampus, olfactory bulb, and
habenula (Fig. 1A, D and E). Lower levels of TAG-1 expression
were detected in the hypothalamic nuclei and the prospective EGL of
the cerebellum. Hypothyroidism did not change the regional pattern
of expression, but caused an increase in TAG-1 RNA expression in
the neocortex, hippocampus, olfactory bulb, and habenula (compare
Fig. 1B, F and G with Fig. A, D and E). In contrast, no differences
were observed in other brain regions, particularly the hypothalamus
and prospective cerebellum (not shown). The effect of in vivo
hormonal treatment was also investigated. As shown in Fig. 1C, H
and I, T4 administration to hypothyroid dams diminished TAG-1
RNA expression almost to normal levels in affected regions.
TAG-1 immunoreactivity was detected almost exclusively in
axonal compartments (Fig. 1J and K). As described previously
(Wolfer et al., 1994), at this age TAG-1 protein was found in the
white matter of the neocortex, internal capsule, corpus callosum,
anterior commissure, hippocampus, ®mbria, and lateral olfactory tract
of control rats. Cell bodies in the cortical plate were TAG-1-negative,
but stained axons entering this zone from the subplate were found. In
hypothyroid animals, TAG-1 immunostaining was increased substan-
tially in the corpus callosum, white mater of the neocortex, internal
capsule, hippocampus, and olfactory tract (compare Fig. 1L and M
with Fig. J and K). No signi®cant differences were observed in other
brain regions. In concordance with RNA data, T4 treatment of
hypothyroid dams led to a partial normalization of TAG-1 protein
expression (Fig. 1N and O). These results indicated that alterations in
the thyroid status modulate TAG-1 RNA and protein expression at
this embryonal age.
P0
At P0, TAG-1 RNA expression in control rats followed the same
pattern as at E20 in all brain regions (Fig. 2A and D). In hypothyroid
rats a clear upregulation of TAG-1 transcripts was apparent in the
prospective layer V of all areas of the neocortex, dentate gyrus, CA2±
3 of the hippocampus, and anterior olfactory bulb (Fig. 2B and F). In
contrast, no obvious differences were observed in other regions such
as the habenula (not shown) or the external granular layer (EGL) of
the cerebellum (Fig. 2C and E).
Both the pattern and intensity of TAG-1 immunoreactivity were
similar to those found at E20 (Fig. 2G, H and L), with the addition
that it was also detected strongly in the inner rim of the EGL of the
cerebellum (Fig. 2K). In hypothyroid rats, in agreement with RNA
results, there was an signi®cant increase in TAG-1 protein levels in
the corpus callosum, anterior and hippocampal commissures, ®mbria,
lateral olfactory tract, and, to a lesser extent, in the forceps major and
white matter of the cerebral cortex of hypothyroid rats (compare
Fig. 2G, H and L with Fig. I, J and N). However, once again, no
obvious differences were observed in the cerebellum (Fig. 2K and
M), although by this time, as noted above, TAG-1 expression levels
are very high in the EGL anyway.
P5
At this age, TAG-1 RNA expression was restricted to layer V of the
cerebral cortex, layers II±III of the retrosplenial cortex, hippocampus,
and the EGL of the cerebellum of euthyroid animals (Fig. 3A, D, G
and H). Hypothyroidism induced an abnormal high level of TAG-1
transcripts in the retrosplenial cortex and, also but to a lesser degree,
in layer V of the cerebral cortex and hippocampus (compare Fig. 3B,
E and I with Fig. A, D and G). Daily treatment of hypothyroid
animals with T4 from P1 to P5 led to a normalization of TAG-1 RNA
levels in these regions (Fig. 3C, F and K). At this age, the normal
high level of expression of TAG-1 in the EGL of the cerebellum
remained apparently unresponsive to changes in thyroid status
(Fig. 3H, J and L), a result also found at P10 (not shown).
In control euthyroid rats TAG-1 immunoreactivity diminished
substantially with respect to earlier ages except in the cerebellum,
where by contrast, expression increased (as has been reported by
Wolfer et al., 1994). TAG-1 protein was detected in the corpus
callosum, anterior and dorsal hippocampal commissures, and the
iEGL of the cerebellum (Fig. 3M, P and Q). In accordance with RNA
results, TAG-1 levels in hypothyroid rats were higher than in controls
in the corpus callosum and the anterior and hippocampal commis-
sures (compare Fig. 3N and R with Fig. M and P) but remained
unchanged in the cerebellum (Fig. 3Q). Also in agreement with in
situ hybridization data, T4 treatment substantially reduced TAG-1
protein levels in responsive areas (Fig. 3O, T and U).
P15±30
By digoxigenin in situ hybridization, a high level of TAG-1
expression at P15 was found to be con®ned to the cerebellum as in
previous studies (Furley et al., 1990; Yoshihara et al., 1995; data not
shown). In comparison to earlier ages, TAG RNA levels were slightly
higher in both the iEGL and IGL. Unexpectedly, considering the
apparent lack of effect in these regions at earlier ages and in contrast
to results obtained at E20 and P0±P5, hypothyroidism caused an
increase in TAG-1 RNA expression. A relative increase in expression
of TAG-1 in the hypothyroid cerebellum was also found by
conventional radioactive in situ (not shown). Using this technique,
TAG-1 RNA was detected in other brain regions but, in contrast to
earlier ages, its expression levels in these regions were not obviously
affected by thyroid status (not shown).
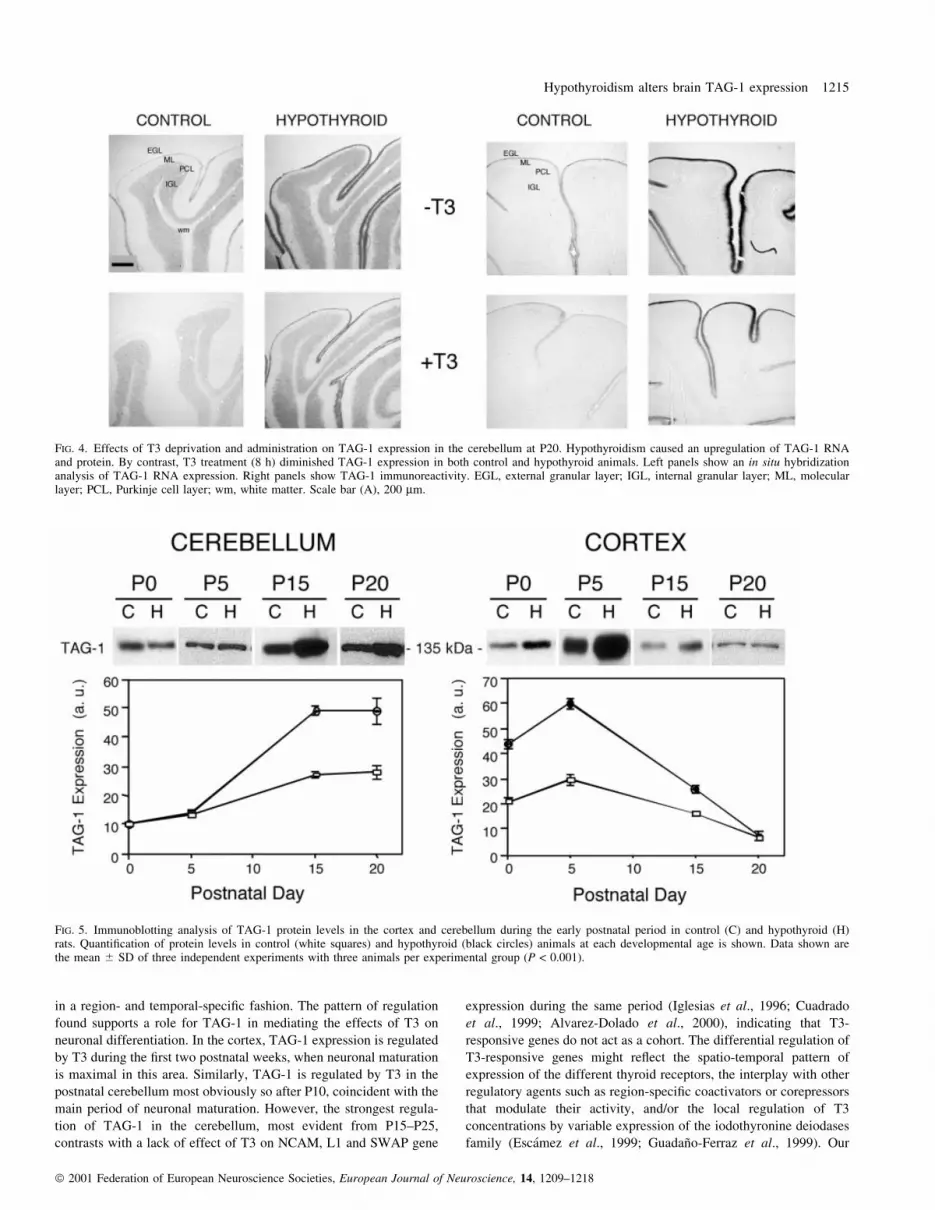
We extended our analysis of the cerebellum to P20 and P25. At
P20, TAG-1 RNA was detected in the EGL and IGL (Fig. 4, left),
whereas TAG-1 immunostaining was restricted to short processes
only in the EGL (Fig. 4, right). TAG-1 RNA and protein expression
were substantially higher in hypothyroid rats as compared to control.
At these ages it is well documented that the number of cells in the
EGL is higher in hypothyroid rats than in control due to a marked
delay in the migration of granular cells to the IGL (Clos et al., 1980;
Legrand, 1984; Dussault & Ruel, 1987). To avoid the complication
that this altered migration might be affected by prolonged T4
treatment (normally 4 days) such that changes in cell localization
Hypothyroidism alters brain TAG-1 expression 1211
ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 14, 1209±1218
could alter the interpretation of the results, hypothyroid animals were
instead subjected to an acute (8 h) treatment with T3 to investigate
hormonal effects on TAG-1 expression. Remarkably, T3 treatment
diminished profoundly the levels of TAG-1 RNA and protein in both
hypothyroid and control animals. This rapid action did not noticeably
affect cell number or localization, and therefore it is likely to be a
consequence of a direct hormonal effect on TAG-1 gene expression.
Identical results were obtained at P25, although TAG-1 expression
levels were lower than at P20 (not shown).
At P30, TAG-1 expression has already declined to the low levels
found in adult life. Consistent with previous reports (Furley et al.,
1990; Wolfer et al., 1994; Yoshihara et al., 1995), although TAG-1
RNA was found in the cerebellum, and to a lesser degree in the
hippocampus and cortex layer V, TAG-1 protein was not detectable.
No signi®cant effects of the thyroid status on TAG-1 expression were
observed at this age.
Developmental study of TAG-1 expression by immunoblotanalysis
To further assess and also quantify the effects of hypothyroidism on
TAG-1 protein expression during development, we performed
Western blot analysis using axonal membrane-enriched extracts of
cerebral cortex (including white matter and corpus callosum) and
cerebellum of control and hypothyroid animals at different ages. In
agreement with previous studies (Furley et al., 1990; Wolfer et al.,
1994), a single band of approximately 135 kDa was detected (Fig. 5).
In line with the asynchronous maturation of these two brain regions,
TAG-1 protein content in the cerebral cortex was maximal at P5 and
diminished later, whereas in cerebellum it progressively increased to
peak at P15. Hypothyroidism did not change the temporal pattern of
expression in the cerebral cortex, but caused a statistically signi®cant
increase (2±3-fold) of TAG-1 protein levels at P0 and P5. This is in
agreement with the stronger immunostaining found in the white
matter and the corpus callosum of hypothyroid rats at these ages
(Figs 2 and 3). The difference between control and hypothyroid rats
was much reduced at P15 and completely disappeared at P25. In line
also with immunohistochemical data, hypothyroidism increased
(around twofold) TAG-1 content in the cerebellum at P15±25, but
not obviously at early ages.
Discussion
The TAG-1 gene encodes an immunoglobulin-like cell adhesion
molecule that is thought to play a crucial role in axonal growth and
guidance, migratory/differentiation processes of the cerebellum, and
FIG. 1. Effect of hypothyroidism on TAG-1 expression at E20. Upper panels (A±I) show an in situ hybridization analysis of TAG-1 mRNA expression.Lower panels (J±O) show TAG-1 protein immunoreactivity. TAG-1 expression was analysed in coronal sections of control (A, D±E, J and K), hypothyroid(B, F±G, L and M), and T4-treated hypothyroid (C, H, I, N and O) rat brains. Note the higher TAG-1 RNA expression found in the neocortex (Nc),hippocampus (Hi), habenula (Hb) and olfactory tract (OT) of hypothyroid animals. D, F, and H are magni®cations of the area marked in (A, B and C),respectively. Note that TAG-1 RNA expression was higher in the cortical plate (CP) and intermediate zone (IZ). Higher immunoreactivity was found in whitematter (wm), hippocampus, and olfactory tract of hypothyroid brains. Processing was standard for all experimental groups. The developing reaction wasstopped when differences between control and hypothyroid animals were most clear. M, molecular layer; NSZ, neocortical subventricular zone; S, striatum;SP, subplate. Scale bar 1 mm (A), 250 mm (D), 1 mm (E), 2 mm (K).
1212 M. Alvarez-Dolado et al.
ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 14, 1209±1218

the formation of highly ordered neural connections (Ruegg et al.,
1989; Furley et al., 1990; Stoeckli, 1997; Sonderegger, 1998). Here
we show that thyroid hormone, a major epigenetic factor during
development, regulates TAG-1 expression during brain maturation in
the rat. To our knowledge, this is the ®rst identi®cation of a systemic
factor that can regulate TAG-1.
As in the case of other T3 target genes (GarcõÂa-FernaÂndez et al.,
1997; Alvarez-Dolado et al., 1998, 1999, 2000), TAG-1 is regulated
FIG. 2. Upregulation of TAG-1 expression by hypothyroidism in newborn rats. Upper panels (A±F) show an in situ hybridization analysis of TAG-1 RNAexpression. Lower panels (G±N) show TAG-1 protein immunoreactivity. Note the higher TAG-1 RNA expression in layer V of the neocortex (arrow in B),dentate gyrus (DG) and CA regions of the hippocampus, and anterior olfactory nuclei (AON) of hypothyroid rats. Immunoreactivity was found elevated incorpus callosum (cc), forceps major (fmj), ®mbria (®), lateral olfactory tract (lo), and anterior commissure (aca) of hypothyroid rats. EGL, external granularlayer; RC, retrosplenial cortex. Scale bar 1 mm (A and C), 1 mm (C±F), 2 mm (G); 1 mm (H), 600 mm (K±N).
Hypothyroidism alters brain TAG-1 expression 1213
ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 14, 1209±1218

FIG. 3. Upregulation of TAG-1 expression by hypothyroidism at P5. Upper panels (A±L) show an in situ hybridization analysis of TAG-1 RNA expression.Lower panels (M±U) show TAG-1 protein immunoreactivity. Note the elevated TAG-1 RNA expression in layers II±III of the retrosplenial cortex (RC), andlayer V (arrows in G, Y and K) of the cortex in hypothyroid rats. Immunoreactivity was found elevated in the dorsal hippocampal commissure (dhc), corpuscallosum, and anterior commissure (ac). T4 treatment of hypothyroid animals was as described in the methods section and normalized the TAG-1 expressionlevels. Scale bar 1 mm (A), 250 mm (D), 1 mm (G), 500 mm (H), 1 mm (P).
1214 M. Alvarez-Dolado et al.
ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 14, 1209±1218
in a region- and temporal-speci®c fashion. The pattern of regulation
found supports a role for TAG-1 in mediating the effects of T3 on
neuronal differentiation. In the cortex, TAG-1 expression is regulated
by T3 during the ®rst two postnatal weeks, when neuronal maturation
is maximal in this area. Similarly, TAG-1 is regulated by T3 in the
postnatal cerebellum most obviously so after P10, coincident with the
main period of neuronal maturation. However, the strongest regula-
tion of TAG-1 in the cerebellum, most evident from P15±P25,
contrasts with a lack of effect of T3 on NCAM, L1 and SWAP gene
expression during the same period (Iglesias et al., 1996; Cuadrado
et al., 1999; Alvarez-Dolado et al., 2000), indicating that T3-
responsive genes do not act as a cohort. The differential regulation of
T3-responsive genes might re¯ect the spatio-temporal pattern of
expression of the different thyroid receptors, the interplay with other
regulatory agents such as region-speci®c coactivators or corepressors
that modulate their activity, and/or the local regulation of T3
concentrations by variable expression of the iodothyronine deiodases
family (EscaÂmez et al., 1999; GuadanÄo-Ferraz et al., 1999). Our
FIG. 5. Immunoblotting analysis of TAG-1 protein levels in the cortex and cerebellum during the early postnatal period in control (C) and hypothyroid (H)rats. Quanti®cation of protein levels in control (white squares) and hypothyroid (black circles) animals at each developmental age is shown. Data shown arethe mean 6 SD of three independent experiments with three animals per experimental group (P < 0.001).
FIG. 4. Effects of T3 deprivation and administration on TAG-1 expression in the cerebellum at P20. Hypothyroidism caused an upregulation of TAG-1 RNAand protein. By contrast, T3 treatment (8 h) diminished TAG-1 expression in both control and hypothyroid animals. Left panels show an in situ hybridizationanalysis of TAG-1 RNA expression. Right panels show TAG-1 immunoreactivity. EGL, external granular layer; IGL, internal granular layer; ML, molecularlayer; PCL, Purkinje cell layer; wm, white matter. Scale bar (A), 200 mm.
Hypothyroidism alters brain TAG-1 expression 1215
ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 14, 1209±1218

results show an abnormally high expression of TAG-1 induced by
hypothyroidism in several brain regions. Whether the regulation of
TAG-1 by T3 is direct or indirect remains unclear, although the tight
correlation of RNA and protein levels in response to thyroid status,
and the rapid response to acute T3 treatment in the cerebellum
suggests a simple regulatory mechanism.
It is known that TAG-1 is expressed during the formation of callosal
and commissural projections, playing an important role in axon
guidance and fasciculation (Dodd et al., 1988; Wolfer et al., 1994;
Stoeckli, 1997; Sonderegger, 1998). The injection of soluble TAG-1 or
anti-TAG-1 antibodies into the developing spinal cord has dramatic
effects on spinal commissural axon fasciculation and path®nding, most
likely due to alterations in interactions with the IgCAMs L1/NgCAM
and Nr-CAM (Stoeckli & Landmesser, 1995), to which TAG-1 is
known to bind heterophilically (Kuhn et al., 1991; Felsenfeld et al.,
1994; BruÈmmendorf & Rathjen, 1998; Volkmer et al., 1998).
Interestingly, hypothyroidism impairs the maturation of axons in
both the corpus callosum and the anterior commissure (Gravel et al.,
1990; Berbel et al., 1994; GuadanÄo-Ferraz et al., 1994). Hypothyroid
rats display inappropriate callosal connections from cortical layers II±
III, with a widespread tangential distribution of callosally projecting
neurons in the visual, parietal and auditory areas (Gravel & Hawkes,
1990; Berbel et al., 1993; Lucio et al., 1997). Although the major
elevation in TAG-1 mRNA expression that we see in the cortex at E20
is in layer V and not in II/III, and the majority of axons in the corpus
callosum are thought to derive from layer II/III, both our protein data,
and that of others (Wolfer et al., 1994), clearly demonstrate signi®cant
levels of TAG-1 protein on ®bres in the corpus callosum (and anterior
commissure) from as early as E16 and remaining until postnatal stages
(Figs 2 and 3). Whether this protein is derived from layer II/III neurons
in the retrosplenial cortex (where TAG-1 mRNA is detected), from the
subset of layer V neurons known to project in the corpus callosum
(Kasper et al., 1994a, 1994b), or perhaps from a transient population of
callosal axons that also might contribute to the path®nding of layer II/
III axons (Ding & Elberger, 2001) is unclear. However, our
immunohistochemistry clearly demonstrates that levels of TAG-1 on
axons in the corpus callosum and anterior commissure increase in
hypothyroid animals (Figs 2 and 3). This increase might directly affect
the path®nding of the overexpressing axons, be they layer II/III axons
or an another axon subset on which layer II/III are dependent.
Additionally, because TAG-1 is naturally produced in two isoforms,
membrane-bound and secreted (Ruegg et al., 1989; Furley et al.,
1990), it is conceivable that the TAG-1 overexpression during
hypothyroidism causes an abnormal accumulation of the soluble
TAG-1 isoform which might alter the interactions with other CAMs
necessary for the formation of this major ®bre tract. We have also
found that L1/Ng-CAM expression is abnormally high in the main
commissural processes of hypothyroid rats (Alvarez-Dolado et al.,
2000). Together, the alteration of TAG-1 and L1/Ng-CAM levels in
these ®bre tracts might contribute to the anomalies found in the
development of commissural and callosal projections during hypo-
thyroidism.
In the hypothyroid brain, many neurons are inappropriately located
for their birthdate causing an abnormal laminar distribution (Lucio
et al., 1997 and references therein). The extracellular environment of
the intermediate zone and subplate during cortical development is
critical for adequate neuronal migration and guidance of thalamic
axons (Rauch, 1997). The expression of the extracellular matrix
proteins Reelin and Tenascin-C have been reported to be altered by
hypothyroidism during cortical development (Alvarez-Dolado et al.,
1998, 1999). The expression of TAG-1 is coincident with that of
Reelin and Tenascin-C, which in turn are expressed coincident with
the maturation of thalamocortical axon pathways and cortical plate
neurons (D'Arcangelo et al., 1995; Fukuda et al., 1997; GoÈtz et al.,
1997). Because TAG-1 is secreted from neurons and is a possible
target for integrin binding (Ruegg et al., 1989; Furley et al., 1990;
Felsenfeld et al., 1994), the overexpression of these molecules in
hypothyroid animals might alter the balance of neuronal interactions
with the extracellular matrix, leading to altered migration and
guidance.
Both T3 and TAG-1 have also been implicated in neuronal
migration and neurite outgrowth of cerebellar granule cells in rodents
(Legrand, 1984; DeLong, 1990; Bailly et al., 1996; Buttiglione et al.,
1998). Expression of TAG-1 begins at the earliest stages of granule
cell differentiation, as these cells become postmitotic in the iEGL, but
seems to disappear shortly after the cells begin to migrate inward
(Furley et al., 1990; Kuhar et al., 1993; Bailly et al., 1996), at which
time F3/contactin becomes expressed (AJF and G. Gennarini,
unpublished observations). A functional interaction between TAG-1
and F3 has been proposed to be necessary for these differentiation
steps (Buttiglione et al., 1998), although in vitro experiments suggest
that TAG-1 is not necessary for the migration process itself (Fishell &
Hatten, 1991). Indeed, one possibility is that downregulation of TAG-
1 might be necessary to allow full inward migration to the IGL to
occur. In this respect it is interesting to note that F3 expression, which
seems to mark the next stage in differentiation, is not affected by
thyroid status (C. Navarro-Yubero, MA-D, G. Gennarini and AM,
unpublished data), supporting the idea that T3 speci®cally affects
granule cells at the stage in differentiation in which they normally
express TAG-1. Thus the delay in granule cell migration seen in
hypothyroid animals (Clos et al., 1980; Legrand, 1984; Dussault &
Ruel, 1987) could be due, in part, to a failure of granule cells to turn
off TAG-1 at an appropriate point in differentiation.
Finally, it has long been suggested that T3 exerts its effects on
brain development during the postnatal period (Schwartz et al.,
1997). However, our data show that TAG-1 expression is also
affected by hypothyroidism during late embryonic stages. Similar
results were recently obtained for reelin, a gene which is down-
regulated by thyroid hormone deprivation before birth (Alvarez-
Dolado et al., 1999) and NSP and Oct-1 genes (Dowling et al., 2000).
Together, these results demonstrate that in contrast to what was
previously thought, the rat brain is responsive to thyroid hormone
during the late gestational period.
In summary, our results show that thyroid hormone de®ciency
causes an asynchronous abnormal upregulation of TAG-1 gene
expression in several areas of the developing rat brain. Changes in
TAG-1 protein levels may contribute to explain at least partially the
alterations in neuronal maturation and connectivity and the delayed
cerebellar cell migration found in the hypothyroid brain.
Acknowledgements
We are grateful to Fernando NuÂnÄez and Pablo SenÄor for the care of animalsand to Margarita GonzaÂlez and Teresa MartõÂnez for their expert technical help.This work was supported by grants from European Union (Biomed-2, BMH4-CT97-2653) to A.M., P.S., and A.J.F., and Plan Nacional de Salud, Ministeriode Ciencia y TecnologõÂa of Spain (SAF98-0060) to A.M. M.A.-D. wassupported by the abovementioned EU project, and A.F. by a fellowship fromPlan Nacional.
Abbreviations
CAM, cell adhesion molecule; CNS, central nervous system; E, embryonicday; EGL, external granule layer; iEGL, inner external granule layer; GPI,glycosyl-phosphatidylinositol; IGL, internal granular layer; IgCAM, immuno-globulin cell adhesion molecules; P, postnatal day; T3, triiodothyronine; T4,thyroxine.
1216 M. Alvarez-Dolado et al.
ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 14, 1209±1218

References
Alvarez-Dolado, M., Cuadrado, A., Navarro-Yubero, C., Sonderegger, P.,Furley, A.J., Bernal, J. & MunÄoz, A. (2000) Regulation of the L1 celladhesion molecule by thyroid hormone in the developing brain. Mol. Cell.Neurosci., 16, 499±514.
Alvarez-Dolado, M., GonzaÂlez-Sancho, J., Bernal, J. & MunÄoz, A. (1998)Developmental expression of tenascin-C is altered by hypothyroidism in therat brain. Neuroscience, 84, 309±322.
Alvarez-Dolado, M., Ruiz, M., del RõÂo, J.A., AlcaÂntara, S., Burgaya, F.,Sheldon, M., Nakajima, K., Bernal, J., Howell, B., Curran, T., Soriano, E. &MunÄoz, A. (1999) Thyroid hormone regulates reelin and dab1 expressionduring brain development. J. Neurosci., 19, 6979±6993.
Bailly, Y., Kyriakopoulou, K., Delhaye-Bouchaud, N., Mariani, J. &Karagogeos, D. (1996) Cerebellar granule cell differentiation in mutantand X-irradiated rodents revealed by the neural adhesion molecule TAG-1.J. Comp. Neurol., 369, 150±161.
Berbel, P., GuadanÄo-Ferraz, A., Angulo, A. & Cerezo, J. (1994) Role ofthyroid hormones in the maturation of interhemisferic connections in rats.Behav. Brain Res., 64, 9±14.
Berbel, P., GuadanÄo-Ferraz, A., MartõÂnez, M., Quiles, J., Balboa, R. &Innocenti, G. (1993) Organization of auditory callosal connections inhypothyroid rats. Eur. J. Neurosci., 5, 1465±1478.
Bernal, J. & GuadanÄo-Ferraz, A. (1998) Thyroid hormone and thedevelopment of the brain. Curr. Opin. Endocrinol. Diab., 5, 296±302.
BruÈmmendorf, T. & Rathjen, F. (1998) Molecular interactions involvingimmunoglobulin superfamily adhesion proteins. In Sonderegger, P. (ed.), IgSuperfamily Molecules in the Nervous System. Harwood AcademicPublishers, Amsterdam, pp. 23±56.
Buttiglione, M., Revest, J.-M., Pavlou, O., Karagogeos, D., Furley, A.J.,Rougon, G. & Faivre-Sarrailh, C. (1998) A functional interaction betweenthe neuronal adhesion molecules TAG-1 and F3 modulates neuriteoutgrowth and fasciculation of cerebellar granule cells. J. Neurosci., 18,6853±6870.
Clos, J., Legrand, C. & Legrand, J. (1980) Effects of thyroid state on theformation and early morphological development of Bergmann glia in thedeveloping rat cerebellum. Dev. Neurosci., 3, 199±208.
Cuadrado, A., Bernal, J. & MunÄoz, A. (1999) Identi®cation of the mammalianhomolog of the splicing regulator Suppressor-of-white-apricot as a thyroidhormone regulated gene. Mol. Brain Res., 71, 332±340.
D'Arcangelo, G., Miao, G., Chen, S.-C., Soares, H., Morgan, J. & Curran, T.(1995) A protein related to extracellular matrix proteins deleted in themouse mutant Reeler. Nature, 374, 719±723.
DeLong, G. (1990) The effect of iodine de®ciency on neuromusculardevelopment. IDD Newsletter, 6, 1±9.
Ding, S.L. & Elberger, A.J. (2001) Postnatal development of biotinylateddextran amine-labeled corpus callosum axons projecting from the visual andauditory cortices to the visual cortex of the rat. Exp. Brain Res., 136, 179±193.
Dodd, J., Morton, S.B., Karagogeos, D., Yamamoto, M. & Jessell, T.M. (1988)Spatial regulation of axonal glycoprotein expression on subsets ofembryonic spinal neurons. Neuron, 1, 105±116.
Dowling, A.L.S., Martz, G.U., Leonard, J.L. & Zoeller, R.T. (2000) Acutechanges in maternal thyroid hormone induce rapid and transient changes ingene expression in fetal rat brain. J. Neurosci., 20, 2255±2265.
Dussault, J. & Ruel, J. (1987) Thyroid hormones and brain development.Annu. Rev. Physiol., 49, 321±334.
EscaÂmez, M.J., GuadanÄo-Ferraz, A., Cuadrado, A. & Bernal, J. (1999) Type 3iodothyronine deiodinase is selectively expressed in areas related to sexualdifferentiation in the newborn rat brain. Endocrinology, 140, 5443±5446.
Felsenfeld, D., Hynes, M., Skoler, K., Furley, A.J. & Jessell, T. (1994) TAG-1can mediate homophilic binding, but neurite outgrowth on TAG-1 requieresan L1-like molecule and beta 1 integrins. Neuron, 12, 675±690.
Fishell, G. & Hatten, M.E. (1991) Astrotactin provides a receptor system forCNS neuronal migration. Development, 113, 755±765.
Fitzli, D., Stoeckli, E.T., Kunz, S., Siribour, K., Rader, C., Kunz, B., Kozlov,S.V., Buchstaller, A., Lane, R.P., Suter, D.M., Dreyer, W.J. & Sonderegger,P. (2000) A direct interaction of axonin-1 with NgCAM-related celladhesion molecule (NrCAM) results in guidance, but not growth ofcommissural axons. J. Cell Biol., 149, 951±968.
Fukuda, T., Kawano, H., Ohyama, K., Li, H.-P., Takeda, Y., Oohira, A. &Kawamura, K. (1997) Immunohistochemical localization of neurocan andL1 in the formation of thalamocortical pathway of developing rats. J. Comp.Neurol., 382, 141±152.
Furley, A.J., Morton, S., Manalo, D., Karagogeos, D., Dodd, J. & Jessel, T.
(1990) The axonal glycoprotein TAG-1 is an immunoglobulin superfamilymember with neurite outgrowth-promoting activity. Cell, 61, 157±170.
Gall, C. & Isackson, P. (1989) Limbic seizures increase neuronal production ofmessenger RNA for nerve growth factor. Science, 245, 758±761.
GarcõÂa-FernaÂndez, L.F., Rausell, E., Urade, Y., Hayaishi, O., Bernal, J. &MunÄoz, A. (1997) Hypothyroidism alters the expression of prostaglandin D2synthase/b-trace in speci®c areas of the developing rat brain. Eur. J.Neurosci., 9, 1566±1573.
Giger, R.J., Vogt, L., Zuellig, R.A., Rader, C., Henehan-Beatty, A., Wolfer,D.P. & Sonderegger, P. (1995) The gene of chicken axonin-1. Completestructure and analysis of the promoter. Eur. J. Biochem., 227, 617±628.
GoÈtz, M., Bolz, J., JoÈster, A. & Faissner, A. (1997) Tenascin-C synthesis andin¯uence on axonal growth during rat development. Eur. J. Neurosci., 9,496±506.
Gravel, C. & Hawkes, R. (1990) Maturation of the corpus callosum of the rat:I. In¯uence of thyroid hormones on the topography of callosal projections.J. Comp. Neurol., 291, 128±146.
Gravel, C., Sasseville, R. & Hawkes, R. (1990) Maturation of the corpuscallosum of the rat II: In¯uence of the thyroid hormones on the number andmaturation of axons. J. Comp. Neurol., 291, 147±161.
GuadanÄo-Ferraz, A., EscaÂmez, M.-J., Rausell, E. & Bernal, J. (1999)Expression of type 2 iodothyronine deiodinase in hypothyroid rat brainindicates an important role of thyroid hormone in the development ofspeci®c primary sensory systems. J. Neurosci., 19, 3430±3439.
GuadanÄo-Ferraz, A., Escobar del Rey, F., Morreale de Escobar, G., Innocenti,G. & Berbel, P. (1994) The development of the anterior commissure innormal and hypothyroid rats. Dev. Brain Res., 81, 293±308.
Hasler, T., Rader, C., Stoeckli, E.T., Zuellig, R. & Sonderegger, P. (1993)cDNA cloning, structural features, and eucaryotic expression of humanTAG-1/axonin-1. Eur. J. Biochem., 211, 329±339.
Iglesias, T., CaubõÂn, J., Stunnenberg, H., Zaballos, A., Bernal, J. & MunÄoz, A.(1996) Thyroid hormone-dependent transcriptional repression of neural celladhesion molecule during brain maturation. EMBO J., 15, 4307±4316.
Karagogeos, D., Pourquie, C., Kyriakopoulou, K., Tavian, M., Stallcup, W.,Peault, B. & Pourquie, O. (1997) Expression of the cell adhesion proteinsBEN/SC1/DM-GRASP and TAG-1 de®nes early steps of axogenesis in thehuman spinal cord. J. Comp. Neurol., 397, 415±427.
Kasper, E.M., Larkman, A.U., Lubke, J. & Blakemore, C. (1994a) Pyramidalneurons in layer 5 of the rat visual cortex. I. Correlation among cellmorphology, intrinsic electrophysiological properties, and axon targets. J.Comp. Neurol., 339, 459±474.
Kasper, E.M., Lubke, J., Larkman, A.U. & Blakemore, C. (1994b) Pyramidalneurons in layer 5 of the rat visual cortex. III. Differential maturation ofaxon targeting, dendritic morphology, and electrophysiological properties.J. Comp. Neurol., 339, 495±518.
Kozlov, S.V., Giger, R.J., Hasler, T., Korvatska, E., Schorderet, D.F. &Sonderegger, P. (1995) The human TAX1 gene encoding the axon-associated cell adhesion molecule TAG-1/axonin: genomic structure andbasic promoter. Genomics, 30, 141±148.
Kuhar, S.G., Feng, L., Vidan, S., Ross, M.E., Hatten, M.E. & Heintz, N.(1993) Changing patterns of gene expression de®ne four stages of cerebellargranule neuron differentiation. Development, 117, 97±104.
Kuhn, R., Stoeckli, E.T., Condrau, M., Rathjen, F. & Sonderegger, P. (1991)Neurite outgrowth on inmobilized axonin-1 is mediated by a heterophilicinteraction with L1. J. Cell Biol., 115, 1113±1126.
Legrand, J. (1984) Effects of thyroid hormones on central nervous system. InYanai, J. (ed.), Neurobehavioural Teratology. Elsevier Science Publishers,Amsterdam, pp. 331±363.
Lucio, R., GarcõÂa, J., Cerezo, J., Pacheco, P., Innocenti, G. & Berbel, P. (1997)The development of auditory callosal connections in normal andhypothyroid rats. Cereb. Cortex, 7, 303±316.
Morreale de Escobar, G., Pastor, R., ObregoÂn, M. & Escobar del Rey, F.(1985) Effects of maternal hypothyroidism on the weight and thyroidhormone content of rat embryonic tissues, before and after onset of fetalthyroid function. Endocrinology, 117, 1890±1896.
Oppenheimer, J. & Schwartz, H. (1997) Molecular basis of thyroid hormone-dependent brain development. Endocr. Rev., 18, 462±475.
Paxinos, G. & Watson, C. (1998) The Rat Brain in Stereotaxic Coordinates,4th edn. Academic Press, San Diego.
Rauch, U. (1997) Modeling an extracellular environment for axonalpath®nding and fasciculation in the central nervous system. Cell TissueRes., 290, 349±356.
Rettig, W., Hoffman, S., Su, S. & Garin-Chesa, P. (1992) Species diversity ofneuronectin and cytotactin expression patterns in the vertebrate centralnervous system. Brain Res., 590, 219±228.
Hypothyroidism alters brain TAG-1 expression 1217
ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 14, 1209±1218

RodrõÂguez-PenÄa, A., Ibarrola, N., InÄiguez, M., MunÄoz, A. & Bernal, J. (1993)Neonatal hypothyroidism affects the timely expression of myelin-associatedglycoprotein in the rat. J. Clin. Invest., 91, 812±817.
Ruegg, M., Stoeckli, E.T., Kuhn, T., Heller, M., Zuellig, R. & Sonderegger, P.(1989) Puri®cation of axonin-1, a protein that is secreted from axons duringneurogenesis. EMBO J., 8, 55±63.
Schwartz, H., Ross, E. & Oppenheimer, J. (1997) Lack of effect of thyroidhormone on late fetal rat brain development. Endocrinology, 138, 3119±3124.
Sonderegger, P. (1998). Ig Superfamily Molecules in the Nervous System.Harwood Academic Publishers, Amsterdam.
Sonderegger, P. & Rathjen, F. (1992) Regulation of axonal growth in thevertebrate nervous system by interactions between glycoproteins belongingto two subgroups of the immunoglobulin duperfamily. J. Cell Biol., 119,1387±1394.
Stoeckli, E.T. (1997) Molecular mechanisms of growth cone guidance: stopand go? Cell Tissue Res., 290, 441±449.
Stoeckli, E.T., Kuhn, T., Duc, C., Ruegg, M. & Sonderegger, P. (1991) Theaxonally secreted protein axonin-1 is a potent substratum for neurite growth.J. Cell Biol., 112, 449±455.
Stoeckli, E.T. & Landmesser, L. (1995) Axonin-1, Nr-CAM, and Ng-CAMplay different roles in the in vivo guidance of chick commissural neurons.Neuron, 14, 1165±1179.
Stoeckli, E.T., Sonderegger, P., Pollerberg, G.E. & Landmesser, L.T. (1997)Interference with axonin-1 and NrCAM interactions unmasks a ¯oor-plateactivity inhibitory for commissural axons. Neuron, 18, 209±221.
Vogt, L., Giger, R., Ziegler, U., Kunz, B., Buchstaller, A., Hermens, W.,Kaplitt, M., Rosenfeld, M., Pfaff, D., Verhaagen, J. & Sonderegger, P.(1996) Continuous renewal of the axonal pathway sensor apparatus byinsertion of new sensor molecules into the growth cone membrane. Curr.Biol., 6, 1153±1158.
Volkmer, H., Zacharias, U., NoÈrenberg, U. & Rathjen, F. (1998) Dissection ofcomplex molecular interactions of neurofascin with Axonin-1, F11, andTenascin-R, which promote attachment and neurite formation of tectal cells.J. Cell Biol., 142, 1083±1093.
Wolfer, D., Henehan-Beatty, A., Stoeckli, E.T., Sonderegger, P. & Lipp, H.-P.(1994) Distribution of TAG-1/axonin-1 in ®bre tracts and migratory streamsof the developing mouse nervous system. J. Comp. Neurobiol., 345, 1±32.
Yamamoto, M., Boyer, A.M., Crandall, J.E., Edwards, M. & Tanaka, H.(1986) Distribution of stage speci®c neurite-associated proteins in thedeveloping murine nervous system recognized by a monoclonal antibody. J.Neurosci., 6, 3576±3594.
Yoshihara, Y., Kawasaki, M., Tamada, A., Nagata, S., Kagamiyama, H. &Mori, K. (1995) Overlapping and differential expression of BIG-2, BIG-1,TAG-1, and F3: four members of an axon-associated cell adhesion moleculesubgroup of the immunoglobulin superfamily. J. Neurobiol., 28, 51±69.
Zuellig, R.A., Rader, C., Schroeder, A., Kalousek, M.B., Vonbohlen, F.,Osterwalder, T., Inan, C., Stoeckli, E.T., Halbach, F., Affolter, H.U., Fritz,A., Hafen, E. & Sonderegger, P. (1992) The axonally secreted cell-adhesionmolecule, axonin-1± primary structure, immunoglobulin-like and®bronectin-type-iii-like domains and glycosyl-phosphatidylinositolanchorage. Eur. J. Biochem., 204, 453±463.
1218 M. Alvarez-Dolado et al.
ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 14, 1209±1218