thyroid disruptor 1,1,1-trichloro-2,2-bis(p-chlorophenyl)ethane (ddt) prevents internalization of...
TRANSCRIPT

REGULAR ARTICLE
Thyroid disruptor 1,1,1-trichloro-2,2-bis(p-chlorophenyl)ethane (DDT) prevents internalization of TSH receptor
S. Picchietti & M. Belardinelli & A. R. Taddei &A. M. Fausto & M. Pellegrino & R. Maggio & M. Rossi &F. Giorgi
Received: 5 September 2008 /Accepted: 19 December 2008 /Published online: 13 February 2009# Springer-Verlag 2009
Abstract The thyroid-stimulating hormone (TSH) receptor(TSHr) was made specifically fluorescent by insertion of atetracysteine motif (TSHr-FlAsH) into the C-terminal endand transiently transfected into COS-7 and HeLa cells. Theobservation that TSH administration caused the intracellularlevel of cAMP to increase in both TSHr-FlAsH-transfectedcell types indicated that the FlAsH binding motif did notalter normal TSHr functioning. When transfected into HeLacells and stimulated with TSH, the TSHr-FlAsH receptorexhibited a pronounced perinuclear labelling pattern,whereas labelling remained on the cell surface followingpre-incubation with 1,1,1-trichloro-2,2-bis(p-chlorophenyl)ethane (DDT). Chinese hamster ovary (CHO)-TSHr cellsprobed with anti-TSHr antibodies were fluorescent mainlyin the proximity of the plasma membrane, with fluores-cence being primarily restricted to a juxta-nuclear positionwhen exposed to 10 mU/ml TSH for 1 or 5 min. However,in the presence of DDT, the anti-TSHr fluorescencemaintained a peripheral location along the cell plasmamembrane, even if CHO-TSHr cells were stimulated withTSH for 1 and 5 min. To verify that DDT acted specifically
on the TSHr, CHO cells transfected with the A2a receptorwere used as controls. Following a 1-min stimulation with5’-(N-ethyl-carboxamido)-adenosine, A2a receptors weregradually internalized regardless of the presence of DDTin the culture medium. Finally, immunoelectron microscopyof CHO-TSHr cells showed that a 1-min exposure to TSHsufficed to displace anti-TSHr antibodies tagged with 10-nm gold particles into coated pits and vesicles but that theirsuperficial location was retained along the plasma mem-brane in the presence of DDT.
Keywords Thyroid-stimulating hormone receptor .
Endocytosis . 1,1,1-Trichloro-2,2-bis(p-chlorophenyl)ethane . A2a receptor . Chinese hamster ovary cells
Introduction
Much of what is presently known about the mechanismscontrolling cell proliferation and differentiation stems fromthe knowledge that cells can express a number of surfacereceptors capable of interacting with the external milieu.Depending on the nature of the ligand-receptor interaction,cells may respond specifically to a variety of signals carriedby many of the information-coding molecules present in theextracellular environment (Maggio et al. 2007). About 80%of the hormones, neurotransmitters and growth factorspresent in the bloodstream of eukaryotic organisms controlsa plethora of metabolic responses by interacting specificallywith G-protein-coupled receptors (GPCRs) (Bockaert andPin 1999). Within the large family of the GPCRs, theglycoprotein hormone receptors specific for luteinizinghormone, follicle-stimulating hormone and thyroid-stimu-lating hormone (TSH) share the common characteristics ofhaving a long ectodomain comprised of numerous leucine
Cell Tissue Res (2009) 336:31–40DOI 10.1007/s00441-008-0749-7
S. Picchietti :M. Belardinelli :A. R. Taddei :A. M. FaustoDepartment of Environmental Sciences, Tuscia University,Viterbo, Italy
M. PellegrinoDepartment of Human Physiology, University of Pisa,Pisa, Italy
R. MaggioDepartment of Experimental Medicine, University of L’Aquila,L’Aquila, Italy
M. Rossi : F. Giorgi (*)Department of Neuroscience, University of Pisa,56120 Pisa, Italye-mail: [email protected]

repeats and a seven-loop transmembrane domain (Hsu et al.2000). The study of their crystal structures has alsorevealed the existence of common mechanisms throughwhich glycoprotein hormone receptors might change con-formationally during activation and transmembrane signaltransduction (Fan and Hendrickson 2005; Sanders et al.2007). Following interaction with their specific hormones,these receptors uncouple from their associated G protein,which, in turn, activates a cascade of enzymes or ionchannel effectors (Lefkowits 2004; Vassart et al. 2004).
The TSH receptor (TSHr) plays a key role in controllingthe metabolic activity of the thyroid gland (Mirashi andMilgrom 1997). Following TSH stimulation of cellsexpressing TSHr (either in vivo thyroid cells or in vitrotransfected cells), the cAMP intracellular concentration ispromptly augmented (Davies et al. 2002) and a number ofcell functions, ranging from thyroglobulin exocytosis(Bjorkman et al. 1978), ion transport (Ericson and Nilsson2000) and release of thyroxine (Mori et al. 1988; Shimon etal. 2002), is consequently activated.
During the screening of several environmental factorsaltering thyroid homeostasis, Santini et al. (2003) foundthat a number of substances, including the insecticide 1,1,1-trichloro-2,2-bis(p-chlorophenyl)ethane (DDT), acted asthyroid disruptors. More recently, we have analysed indetail whether this effect on thyroid homeostasis occurs viainteraction with the TSHr. We have thus established thatDDT inhibits both the basal and the TSH-stimulatedaccumulation of cAMP in Chinese hamster ovary CHO-K1 cells stably transfected with TSHr (Rossi et al. 2007).Furthermore, DDT can still inhibit the basal TSHr activity,even when deprived of the extracellular domain, suggestingthat DDT interacts directly with the transmembrane domainof the TSHr. As controls, cells transfected with dopamineD1 and adenosine type A2a receptors have been shown torespond specifically to their respective agonists, even whenstimulated in the presence of DDT. These observations havebeen interpreted as indicating that DDT acts specifically asan inverse agonist of the TSHr itself (Rossi et al. 2007).
In this study, we have aimed at verifying whether DDTexerts any inhibitory effect on the intracellular trafficking ofthe TSHr in TSHr-transfected CHO and HeLa cells. Wehave investigated this hypothesis by modifying TSHr bythe insertion of a short tetracysteine sequence into its C-terminal domain. By this procedure, the receptor isrendered specifically fluorescent by interaction with themembrane-permeant fluorescein derivative, fluorescein ar-senical hairpin (FlAsH; Hoffman et al. 2005). Oncepermanently transfected into CHO cells or transientlyexpressed in HeLa cells, this modified TSHr has beenlocalized in vivo by dynamic confocal microscopy or bylight fluorescent microscopy following aldehyde fixation.The TSHr has also been localized immunocytochemically
with specific anti-TSHr antibodies in COS-7 and CHOcells. In each case, TSHr localization has been shown tovary in relation to DDT and/or TSH exposures. Our long-term aim is to use DDT as a lead compound to search forTSHr inhibitors potentially capable of acting as pharma-ceutical drugs counteracting hyperthyroidism.
Materials and methods
Materials
Tissue culture media and sera were from Sigma-Aldrich (St.Louis, Mo., USA) and Celbio (Milan, Italy). [3H]Adeninewas from PerkinElmer Life and Analytical Sciences(Boston, Mass., USA). Forskolin, Ro 20-1724, 3-isobutyl-1-methylxanthine (IBMX), 5’-(N-ethyl-carboxamido)-adeno-sine (NECA), 1,1,1-trichloro-2,2-bis(p-chlorophenyl)ethane(DDT) and 1,2-ethanedithiol (EDT) were from Sigma-Aldrich. DDTwas dissolved in a 99.5% dimethylsulphoxide(DMSO) and further diluted in Dulbecco’s modified Eagle’smedium (DMEM) to yield a final concentration of 100 μM.Control samples were treated with equivalent amounts ofDMSO at concentrations not higher than 12.73 mM. PlasmidDNA transfections were carried out by using either theDEAE-dextran (Yang and Yang 1997) or the lypofectamine(Sigma-Aldrich) procedures (Ourlin et al. 1997). Primaryantibodies for the localization of TSHr (both monoclonaland polyclonal) and the polyclonal anti-adenosin A2areceptor were purchased from Santa Cruz Biotechnology(Calif., USA). Bovine TSH (bTSH) was purchased fromSigma-Aldrich.
Eukaryotic expression vectors
Wild-type human TSHr cDNA inserted into the pcDNA3.1-Zeo(-) vector was a kind gift from Dr. M. Szkudlinski(Trophogen, Rockville, Md., USA). The construction ofwild-type TSHr was as described by Zhang et al. (2000). Thehuman adenosine A2a receptor cloned in a pcDNA3.1(-)vector was purchased from the Unité Mixte de RecherchecDNA Resource Centre. The cDNA for type V adenylcyclase, subcloned into a pXMD1 vector, was kindlyprovided by Dr. Z. Vogel (Weizman Institute of Sciences,Rehovot, Israel).
Cell culture
HeLa cells transiently transfected with the TSHr or CHO-K1 cells stably transfected with the TSHr (CHO-TSHr)were routinely employed (Perret et al. 1990). Control cellswere either CHO-K1 cells or CHO-K1 stably transfectedwith the adenosine A2a (CHO-A2a) receptor (Olah and
32 Cell Tissue Res (2009) 336:31–40

Stiles 1995). CHO cells were incubated in 24-well trays andmaintained at 37°C in a humidified atmosphere (containing5% CO2) in Dulbecco’s modified Eagle’s medium(DMEM), supplemented with 10% fetal bovine serum,100 U/ml penicillin, 100 μg/ml streptomycin, 2% L-glutamine, and 1% non-essential amino acids and growndirectly in 24-well plates (1.0×105 cells in each well).HeLa cells were incubated in these trays with DMEMmixed with F12 HAM supplemented with glutamine. CHO-TSHr and CHO-A2a cells were maintained in the constantpresence of the selecting agent G-418 (Sigma-Aldrich) at aconcentration of 0.6 mg per millilitre of culture medium.HeLa-TSHr or CHO-TSHr cells were seeded on a roundglass coverslip placed at the bottom of each well. By theend of each culture period, glass coverslips were mountedon glass slides by using the Anti-fade kit (Invitrogen,Carlsbad, Calif., USA).
Construction of TSHr-FlAsh pcDNA 3.1 plasmid
The TSHr-FlAsH was generated by inserting an exapeptidecontaining a tetracysteine motif (CCPGCC) into the C-terminal of the TSHr downstream of the leucine 764. Theresulting TSHr-FlAsH cDNA was subcloned in thepcDNA3.1Zeo(-) vector between the EcoRI and XhoI sites.
Transient transfection of COS-7 cell by the DEAE-Dextranmethod
Exponentially growing COS-7 cells were trypsinized,seeded at 5×105 cells in 10-cm Petri dishes and incubatedovernight in 10 ml growth medium. Then, 2–3 μg plasmidDNA was mixed with a 0.05% DEAE-Dextran (Sigma-Aldrich) solution in phosphate-buffered saline (PBS),swirled gently and added dropwise to the culture mediumfor 30 min at 37°C under 5% CO2. At the end of thisperiod, the medium was removed and the cells were rinsedtwice with fresh medium and incubated again for 2 h in thepresence of 80 μM chloroquine (Sigma-Aldrich). Cellswere eventually re-fed by incubation in a new medium for24 h at 37°C under 5% CO2. The following day, cells wereseeded at the appropriate ratio (70×103:100×103) in 24-well Petri dishes and incubated for an additional 24 h priorto being tested for adenylyl cyclase.
Transient trasfection of HeLa cell by the lypofectaminemethod
HeLa cells were cultured at a density of 7×104 cells/wellon 14-mm diameter round coverslips (sterilized and de-greased with 80% ethanol and absolute ethanol). Thefollowing day, cells were transfected with the lipofectaminesolution and incubated for an additional 24 h under 5%
CO2 at 37°C. The lipofectamine solution was prepared bymixing 0.5 μl lipofectamine and 0.4 μg TSH-FlAsHplasmid in 250 μl DMEM mixed with F12 HAMsupplemented with glutamine and non-essential aminoacids. The next day, the lipofectamine solution wasreplaced with fresh medium containing fetal bovine serumand antibiotics and maintained until the adenylyl cyclaseassay was performed.
FlAsH assay
HeLa cells were assayed for FlAsH following exposure to1 μM FlAsH, 12.5 μM EDT and 0.05% DMSO in 1 g/l glucose in PBS. Initially, 667 μM FlAsH, 8.3 mM EDTand 33.3% DMSO were allowed to equilibrate for 15 min atroom temperature in the darkness. Subsequently, thissolution was diluted with 1 g/l glucose/PBS to yield a finalconcentration of 1 μM FlAsH, 12.5 μM EDT and 0.05%DMSO. Following an additional 10-min equilibration atroom temperature, 300 μl of this solution was added to theHeLa cells. They were further incubated for an additional1 h at 37°C under 5% CO2 in the presence either of DMSOalone as control cells or of DMSO containing 100 μM DDTas experimental cells. Cells were then washed twice with300 μM EDT, 25 μM BAL wash buffer containing either100 μM DDT or 1% DMSO. The washing solution waseventually replaced with glucose/PBS and 10 mU/ml TSHand maintained in culture medium for 10 min at roomtemperature. The reaction was blocked by fixing the HeLacells for 1 h with 1% glutaraldehyde, 2% paraformaldehyde(Electron Microscopy Sciences, Hatfield, Pa., USA) in cold(4°C) phosphate buffer at pH 7.2. Following fixation,coverslips were transferred to cold phosphate buffer andeventually mounted on glass slides with 20 μl Anti-fadesolution. Mounted glass slides were sealed with nail polish,stored overnight at −20°C and then analysed by scanningconfocal microscopy. To detect the tetracysteine-boundFlAsH, fixed HeLa cells were excited at 488 nm with anArgon laser and the resulting signal that was filtered byscanning confocal microscopy at 515–530 nm emission wasevaluated with an image analyser program.
cAMP accumulation
cAMP accumulation in intact cells was measured inquadruplicate experiments by a two-step column separationprocedure (Johnson and Salomon 1991). The experimentalprotocol for this functional assay was as follows: CHO-K1,CHO-TSHr or CHO-A2a cells were preincubated for 2 hwith 250 μl medium supplemented with 1.25 μCi [3H]adenine at 37°C in a humidified atmosphere. At the end ofthis preincubation period, cells were stimulated with theirrespective agonist for an additional 10 min at the indicated
Cell Tissue Res (2009) 336:31–40 33

concentrations in the presence of the phosphodiesteraseinhibitors Ro 20-1724 (0.5 mM) and IBMX (1 mM). Thereaction was then blocked with perchloric acid containing0.1 mM unlabelled cAMP, followed by neutralization withKOH. The amount of [3H]cAMP formed was determined asdescribed by Avidor-Reiss et al. (1995). Decays per minute(dpm) values of [3H]cAMP recovered from the columnswere normalized to their respective protein contents, whichon average were 50.3±3.4 μg.
Fluorescence immunocytochemistry
CHO-K1, CHO-TSHr and CHO-A2a cells were cultured onglass coverslips at 37°C in a humidified atmosphere(containing 5% CO2) in Dulbecco’s modified Eagle’smedium supplemented with 5% fetal bovine serum,100 U/ml penicillin, 100 μg/ml streptomycin, 2% L-glutamine and 1% non-essential amino acids. On the thirdday of culture, the entire medium was substituted with freshmedium containing no serum in order to enhance receptorexpression. Following an additional day, cells were exposedfor 100 min to DDT (Sigma-Aldrich) at a final concentra-tion of 100 μM. Controls were run with CHO-K1 cellsexposed to the same DDT concentration and with CHO-TSHr or CHO-A2a cells not exposed to DDT. In someexperiments, both experimental and control cells wereexposed for 30 min to the fluorescent probe Lysotracker(InVitrogen) at a concentration of 50 nM to label acidifiedlysosomal or late endosomal vesicles. TSH was eventuallyadded at a concentration of 1 mU/ml and allowed to act onexperimental and control cells for either 1 or 5 min. Theagonist NECA was used under the same experimentalconditions at a final concentration of 1 μM. By the end ofeach incubation period, cells were washed with PBS andfixed with 4% paraformaldehyde in 0.05 M phosphatebuffer at pH 7.2. Cells were permeabilized with 0.05%Tween-20 and 0.5% bovine serum albumin (BSA) in PBSand then incubated for 90 min at room temperature in aPBS solution containing either mouse anti-TSHr (125 μg/ml) diluted 1:20 or rabbit anti-A2a receptor (200 μg/ml)diluted 1:150. Following incubation with primary anti-bodies, cells were washed three times with PBS containing0.1% Tween-20, plus an additional wash with PBS alone,and incubated for 1 h at room temperature with a secondaryantibody (fluorescein-isothiocyanate-labelled goat anti-mouse IgG or anti-rabbit IgG; InVitrogen). Cells incubatedwith secondary antibodies alone were used as negativecontrols. After the completion of all incubation steps, cellswere extensively washed with H2O and finally mounted onglass slides with anti-fading agents. Staining was assessedwith a Zeiss fluorescence microscope equipped with acolour video camera (Axio Cam MRC, Milano Italy)controlled by AxioVision software.
Immunoelectron microscopy
CHO-TSHr cells were cultured until confluence on high-density large-pore (3 μm) filters in cell-culture inserts(Becton Dickinson, Mountain View, Calif., USA) andtreated with DDT and/or TSH as specified above. Cellswere then fixed with 4% paraformaldehyde, 0.5% glutaral-dehyde in 50 mM phosphate buffer pH 7.2, rinsed in thesame buffer, dehydrated in an ascending series of ethanols,embedded in LRWhite methacrylate resin, placed in gelatincapsules and allowed to polymerize at room temperature inthe presence of benzoyl peroxide. For immunogold stain-ing, ultrathin sections were obtained with an LKB ultrami-crotome (LKB, Uppsala, Sweden) and placed on uncoatednickel grids. Sections were first blocked with a solution of0.5% BSA in 0.05 M TRIS-HCl buffer at pH 7.6 and thenincubated with mouse anti-human TSHr (1:200). Gridswere washed in 0.05 M TRIS-HCl buffer at pH 7.6 andincubated with 20-nm gold-labelled goat anti-rabbit or anti-mouse IgG antibodies (British BioCell International, UK)and eventually counterstained with uranyl acetate. Sectionswere observed in a JEM EX II transmission electronmicroscope (Jeol Italia). Sections incubated with thesecondary antibody alone served as controls.
Results
Fluorescent detection of TSHr
To establish the possible displacement of TSHr in trans-fected cells after TSH stimulation and/or DDT inhibition,the receptor itself was made specifically fluorescent by theFlAsH procedure. Accordingly, a stretch of six amino acidscontaining a tetracysteine motif (TSHr-FlAsH) was intro-duced into the C-terminal end of the TSHr, as specified inthe Materials and methods section. Initially, the TSHr-FlAsH receptor was transiently transfected into COS-7 cellsand tested for its ability to increase the intracellular level ofcAMP. TSH administration caused the intracellular level ofcAMP to increase in TSHr-FlAsH-transfected COS-7 cellsin much the same way as in COS-7 cells transfected withthe wild-type TSHr (Fig. 1). We took this observation toindicate that insertion of the FlAsH binding motif did notalter normal TSHr functioning. Having established thisequivalence in COS7 cells, a stable CHO-TSHr-FlAsH cellline was created and a number of CHO-positive clones wereselected for their highest cAMP response to TSH stimula-tion. However, similar intracellular staining patterns werefound in CHO-TSHr-FlAsH and CHO-K1 cell types,indicating the existence of a fluorescent background. Toovercome this difficulty, the TSHr-FlAsH transfectionprocedure was optimized for HeLa cells as described by
34 Cell Tissue Res (2009) 336:31–40

Stroffekova et al. (2001). The HeLa-TSHr-FlAsH cellsresponded to TSH stimulation as efficiently as HeLa cellstransfected with the wild-type TSHr alone (Fig. 1). At thesame time, they appeared to exhibit fluorescent labellingpatterns that varied, both in intensity and spatial distribu-tion, depending on the culturing conditions adopted(Fig. 2). In the absence of any stimulation, HeLa cells hada diffuse background staining, whereas they exhibited apronounced perinuclear labelling pattern when stimulatedwith TSH (10 mU/ml), probably indicating the internaliza-tion of the TSHr-FlAsH complex. On the contrary, labellingremained associated with the cell surface in cases in whichTSH stimulation was preceded by exposure to 100 μMDDT, indicating that TSHr-FlAsH internalization could becompletely abolished under these latter conditions.
Immunocytochemical detection of TSHr trafficking
To provide additional evidence on TSHr trafficking uponexposure to TSH and/or DDT, CHO-TSHr cells were probed
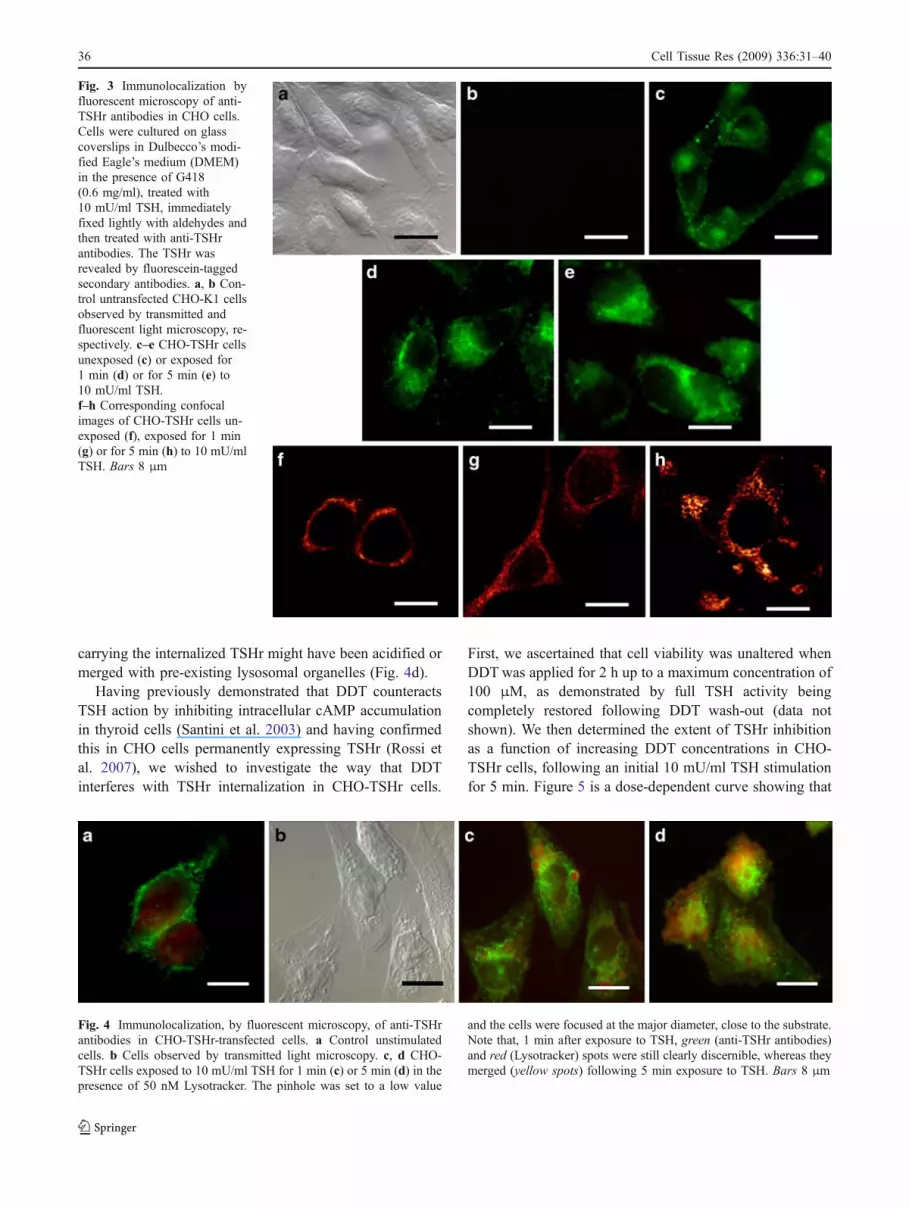
with anti-TSHr antibodies. Control CHO-k1 cells were voidof any fluorescent staining when exposed to these specificantibodies (see Fig. 3a, b). By contrast, CHO cells transfectedwith the TSHr (CHO-TSHr) were highly fluorescent alongthe plasma membrane when exposed to anti-TSHr antibodies(Fig. 3c), thus indicating the specificity of the immunelocalization of the TSHr. Essentially similar results wereobtained regardless of the monoclonal or polyclonal natureof the antibody employed for this immune localization.
Upon exposure to 10 mU/ml TSH for 1 min, CHO TSHrcells exhibited an entirely different labelling pattern, theTSHr being displaced from the plasma membrane to ajuxta-nuclear position (Fig. 3d). Prolonging the TSHstimulation for up to 5 minutes increased both the intensityof fluorescent staining (Fig. 3e) and the cytoplasmicspreading of the TSHr. When these observations wereextended to the confocal microscope, the TSHr was seen tobe mainly associated with the plasma membrane in theabsence of any TSH stimulation (Fig. 3f), was displacedinto part of the cytoplasm after 10 mU/ml TSH stimulationfor 1 min (Fig. 3g) and eventually gathered in spot-likecytoplasmic clusters after up to 5 min of TSH stimulation(Fig. 3h). Taken together, these observations indicated thatTSH exposure caused the TSHr to be progressivelywithdrawn from the plasma membrane and to be internal-ized by endocytosis. TSH effects on CHO-TSHr cells werefurther investigated following exposure to Lysotracker, afluorescent acidotropic probe known to label specificallylate endosome and lysosomes. As can clearly be seen inFig. 4a, unstimulated CHO-TSHr cells exhibited a predom-inantly cell-membrane-associated fluorescence, whereascells exposed for 1 min to TSH exhibited an intracellulargreen labelling pattern still clearly distinguishable from thered lysosomal staining (Fig. 4b, c). However, within 5 min,CHO-TSHr cells appeared to be enriched by a few yellowspots, indicating that some of the endosomal vesicles
Fig. 1 Effects of TSH stimula-tion on COS-7 and HeLa cellstransiently transfected with TSHreceptor (TSHr)-FlAsH (TSHr-FlAsH) or wild-type TSHr(TSHr-wt). Both cell types wereexposed to TSH for 10 min at a10 or 20 mU/ml concentration.As controls, untransfectedCOS-7 and HeLa cells wereexposed for the same lengths oftime to TSH. cAMP concentra-tion was determined andexpressed as decays per minute(dpm) per microgram protein(FlAsH fluorescein arsenicalhairpin)
Fig. 2 Fluorescent visualization, by confocal microscopy, of the TSHras modified by the insertion of a tetracysteine motif into the carboxy-terminal following exposure to FlAsH according to the procedure ofHoffmann et al. (2005). a Control for untransfected HeLa cells. bTSHr-FlAsH-transfected HeLa cells exposed for 10 min to 10 mU/mlTSH. c TSHr-FlAsH-transfected HeLa cells exposed for 10 min to10 mU/ml TSH, following DDT treatment for 40 min at aconcentration of 100 μM. Bars 20 μm
Cell Tissue Res (2009) 336:31–40 35
carrying the internalized TSHr might have been acidified ormerged with pre-existing lysosomal organelles (Fig. 4d).
Having previously demonstrated that DDT counteractsTSH action by inhibiting intracellular cAMP accumulationin thyroid cells (Santini et al. 2003) and having confirmedthis in CHO cells permanently expressing TSHr (Rossi etal. 2007), we wished to investigate the way that DDTinterferes with TSHr internalization in CHO-TSHr cells.
First, we ascertained that cell viability was unaltered whenDDTwas applied for 2 h up to a maximum concentration of100 μM, as demonstrated by full TSH activity beingcompletely restored following DDT wash-out (data notshown). We then determined the extent of TSHr inhibitionas a function of increasing DDT concentrations in CHO-TSHr cells, following an initial 10 mU/ml TSH stimulationfor 5 min. Figure 5 is a dose-dependent curve showing that
Fig. 4 Immunolocalization, by fluorescent microscopy, of anti-TSHrantibodies in CHO-TSHr-transfected cells. a Control unstimulatedcells. b Cells observed by transmitted light microscopy. c, d CHO-TSHr cells exposed to 10 mU/ml TSH for 1 min (c) or 5 min (d) in thepresence of 50 nM Lysotracker. The pinhole was set to a low value
and the cells were focused at the major diameter, close to the substrate.Note that, 1 min after exposure to TSH, green (anti-TSHr antibodies)and red (Lysotracker) spots were still clearly discernible, whereas theymerged (yellow spots) following 5 min exposure to TSH. Bars 8 μm
Fig. 3 Immunolocalization byfluorescent microscopy of anti-TSHr antibodies in CHO cells.Cells were cultured on glasscoverslips in Dulbecco’s modi-fied Eagle’s medium (DMEM)in the presence of G418(0.6 mg/ml), treated with10 mU/ml TSH, immediatelyfixed lightly with aldehydes andthen treated with anti-TSHrantibodies. The TSHr wasrevealed by fluorescein-taggedsecondary antibodies. a, b Con-trol untransfected CHO-K1 cellsobserved by transmitted andfluorescent light microscopy, re-spectively. c–e CHO-TSHr cellsunexposed (c) or exposed for1 min (d) or for 5 min (e) to10 mU/ml TSH.f–h Corresponding confocalimages of CHO-TSHr cells un-exposed (f), exposed for 1 min(g) or for 5 min (h) to 10 mU/mlTSH. Bars 8 μm
36 Cell Tissue Res (2009) 336:31–40

TSH activity, as measured by the intracellular accumulationof cAMP, progressively diminishes when CHO-TSHr cellsare exposed to increasing DDT concentrations. Figure 6 is aseries of four confocal images showing CHO-TSHr cellsexposed to increasing concentrations of DDT. Under these
culturing conditions, the intracellular labelling of the cellsis also progressively reduced as the DDT concentrationincreases. To achieve this level of analysis, CHO cells hadto be sectioned according to an optical plane enhancingtheir intracellular organelle content with respect to theirmembrane contours.
Once the overall effects of the DDT concentrations wereascertained, we attempted to determine, by fluorescent lightmicroscopy, the way that DDT exposure affects thelocalization of the TSHr in CHO cells. Control CHO-TSHrcells were characterized by a prevailing intracellularfluorescent labelling following a 5-min exposure to TSH(Fig. 7a, d). By contrast, exposure to TSH for 1 or 5 min inthe presence of DDT resulted in a fluorescence mainlyalong the cell contour, thus indicating that the TSHr had notbeen internalized and persisted on the plasma membrane(Fig. 7b, c, e, f). When CHO-TSHr cells were exposed toDDT following pre-labelling with Lysotracker, none of theanti-TSHr green fluorescent spots merged with any of thered labelled organelles, indicating that endocytic internali-zation of the TSHr had been inhibited (data not shown).
To verify that the DDT effects on TSHr internalization intransfected CHO cells were specifically exerted on thereceptor itself and did not simply interfere with generalfunctions compromising cell vitality, the above experimentswere repeated with CHO cells transfected with theadenosine A2a receptor, a glycoprotein receptor belongingto the same GPCRs family as the TSHr and likewiseendowed with a seven-pass transmembrane domain. Theanti-A2a antibodies identified, as seen by immunofluore-sence microscopy, a number of spots along the plasmamembrane of CHO-A2a transfected cells (Fig. 7g). Follow-ing stimulation for 1 or 5 min with the specific agonist
Fig. 6 Immunolocalization by confocal microscopy of anti-TSHrantibodies in CHO-TSHr-transfected cells exposed for 5 min to10 mU/ml TSH following a period of 100 min in the presence ofincreasing concentrations of DDT from 0 to 100 μM. Cells werecultured in DMEM in the presence of DDT and TSH, mildly fixed in
aldehydes and observed at an optical plane focused on the main part ofthe cell body to demonstrate primarily the endosomal compartment.They were eventually treated with anti-TSHr antibodies, the receptorbeing revealed by fluorescein-tagged secondary antibodies. Bars20 μm
Fig. 5 Dose-response curve expressing the effects of DDT exposureon CHO-TSHr cells stimulated with 10 mU/ml TSH. CHO-TSHr cellswere treated as specified and exposed for the same lengths of time todoses of DDT ranging from 0 to 100 μM. Data on cAMPconcentrations are from three independent experiments and expressedas dpm per μg of proteins
Cell Tissue Res (2009) 336:31–40 37

NECA, A2a receptors were seen to be displaced from theplasma membrane and were gradually internalized(Fig. 7h). However, unlike TSHrs that were retained alongthe cell surface upon exposure to DDT, the adenosine A2areceptors were still internalized, even if stimulated withNECA in the presence of DDT (Fig. 7i). We consider thatthese observations confirm our previous results (Rossi et al.2007) showing that DDT specifically inhibits the internal-ization of the TSHr by interacting directly with it or withthe phospholipid rafts that anchor its transmembranedomains to the plasma membrane.
To verify that TSHr internalization did indeed occur viareceptor-mediated endocytosis, TSHrs were localized byimmunoelectron microscopy in CHO-TSHr cells treatedwith TSH alone or following DDT exposure. As shown inFig. 8a, a 1-min exposure to TSH sufficed to displace anti-TSHr antibodies tagged with 10-nm gold particles fromtheir superficial location to coated pits and vesicles. Bycontrast, when CHO-TSHr cells were treated for the same
length of time with TSH in the presence of DDT, anti-TSHrantibodies labelled with gold particles retained theirsuperficial location along the plasma membrane (Fig. 8b).As expected, the number of endocytic vesicles that weregold-labelled after a 5-min exposure to TSH increased inCHO-TSHr cells, whereas in the presence of DDT,essentially no change was observed (data not shown).
Discussion
The observations described in this work can be summarizedas follows. CHO cells transfected with the TSHr respond toTSH stimulation by internalizing the receptor itself. Within1 min of TSH exposure, the receptor has gained a juxta-nuclear location in CHO-TSHr cells, whereas at least 5 minare required for the receptor to appear in late endosomes ormerge with part of the lysosomal compartment. Observa-tions by confocal and electron microscopy have confirmed
Fig. 7 Immunolocalization, by fluorescent microscopy, of anti-TSHrantibodies in CHO-TSHr-transfected cells exposed for 5 min to10 mU/ml TSH following a period of 100 min in the presence of100 μM DDT. a, d Control CHO-TSHr cells stimulated with 10 mU/ml TSH for 5 min with no DDT added. b–f CHO-TSHr cellsstimulated with 10 mU/ml TSH for 1 min (b, e) and 5 min (c, f) in thepresence of 100 μM DDT (b, c transmitted light, e, f correspondingfluorescent images). g–i CHO cells transfected with the A2a receptor.g Unstimulated control CHO-A2a cells. h CHO-A2a cells exposed for
1 min to 1 μM NECA. i CHO-A2a cells exposed for 1 min to 1 μMNECA in the presence of 100 μM DDT. Cells were cultured inDMEM in the presence of DDT and 10 mU/ml TSH, mildly fixed inaldehydes and treated with antibodies. Their respective receptors wererevealed by fluorescently tagged secondary antibodies. Under theseculturing conditions, green fluorescent anti-A2a spots were internal-ized following NECA stimulation, regardless of the presence of DDTin the culture medium. Bars 8 μm
38 Cell Tissue Res (2009) 336:31–40

that receptor internalization in CHO-TSHr cells occurs byendocytosis, primarily through the formation of coated pitsand vesicles along the plasma membrane. However, since anumber of fluorescent spots can still be detected inunstimulated CHO cells, endocytosis probably occurs evenin the absence of any TSH stimulation, possibly as a resultof TSHr constitutive activity (Parnot et al. 2002). Internal-ization may thus be seen as one of the functional correlatesthat follow receptor activation caused by ligand binding andintracellular elevation of the cAMP level (Rossi et al. 2007).
A number of earlier reports (Heldin et al. 1994) havedemonstrated that TSH stimulation of transfected CHOcells leads to receptor internalization. However, thisevidence has been obtained by exposing the TSHr directlyto the TSH conjugated with gold particles (Baratti-Elbaz etal. 1999) or by modifying the receptor itself through theinsertion of the yellow fluorescent protein (Singh et al.2004). In each case, the structural complexity conferred onthe TSHr by these chemical modifications may hinder ordelay its mobility during endocytic internalization. In thepresent work, the localization of the TSHr has beenroutinely ascertained by adopting two different procedures:by immunolocalization with primary mono- or polyclonalantibodies that bind to secondary antibodies conjugated togold particles in transfected CHO cells and by modifyingthe TSHr through insertion of a short tetra-cysteinesequence into its C-terminal domain in transfected HeLacells (Hoffman et al. 2005). Under both conditions, TSHr
can be visualized along the cell surface or is internalized,depending on the experimental treatment. This demon-strates that the modified TSHr is as efficient as the wild-type receptor in causing cAMP elevation and endocyticinternalization whenever exposed to TSH. As we havepreviously established that DDT inhibition of transfectedCHO cells occurs via direct interaction with the TSHr(Rossi et al. 2007), the present study has aimed at verifyingwhether it could also interfere with the process of receptorinternalization. Our data, based on immune and fluorescentreceptor localization, clearly show that TSHr internalizationis greatly inhibited in the presence of DDT, as demonstratedby its persistence along the cell membrane of CHO-TSHrcells. However, unlike TSHr, the internalization of the A2areceptor is not affected by DDT exposure under similarculturing conditions. We interpret these observations asindicating that DDT acts directly on the TSHr and that thestructural differences between the TSHr and adenosine A2areceptors suffice to explain their differential response toDDT exposure. Earlier observations from our laboratoryhave clearly demonstrated that the TSHr activity can still beinhibited by DDT, even if the TSHr is deprived of theextracellular domain, thus indicating that the insecticide, inall likelihood, acts as an inverse agonist directly on theheptahelical domain of the receptor. Indeed, neitherremoval of the TSHr ectodomain by trypsin treatment nortransfection with a trunkated TSHr void of the ectodomainsuffice to reduce the inhibitory effect exerted by DDT on
Fig. 8 Electron immunolocalization of anti-TSHr antibodies in CHO-TSHr-transfected cells following a 1-min exposure to 10 mU/ml TSH(a) or following a 5-min TSH exposure in the presence of 100 μMDDT (b). Cells were cultured in DMEM in the presence of TSH and/or DDT, mildly fixed in aldehydes and embedded in methacrylate.Thin sections from these cells were treated with anti-TSHr antibodies
and the receptor was revealed by 10-nm gold-tagged secondaryantibodies (N nucleus). Note that gold particles were located in coatedand endosomic vesicles in the absence of DDT (arrows in a), whereasthey remained restricted to the cell plasma membrane in the presenceof 100 μM DDT (arrows in b). Bars 300 μm (a), 450 μm (b)
Cell Tissue Res (2009) 336:31–40 39

CHO-TSHr-transfected cells (Rossi et al. 2007). Interest-ingly, a similar inverse agonist action has been shown to beexerted by the positive allosteric modulator 3,3-difluoro-benzaldazine on the transmembrane domain of the metab-otropic glutamate receptor 5, even in the absence of theextracellular domain (Goudet et al. 2004).
In conclusion, our observations are consonant with amodel whereby DDT acts directly and reversibly ontransfected Cos-7, HeLa and CHO cells to inhibit theinternalization of TSHr. At the morphological level, thisinhibitory effect is clearly evidenced by the receptorpersistence along the cell plasma membrane, thus indicatingthat a blocked TSHr is actually made incapable ofinteracting with the molecular apparatus sustaining endo-cytic internalization. A number of molecules structurallysimilar to DDT and capable of inhibiting the activity of theTSHr more efficiently are currently being tested in ourlaboratories (preliminary observations). We hope that thisstrategy may eventually lead to the adoption of suitablepharmaceutical drugs to counteract hyperthyroidism.
References
Avidor-Reiss T, Bayewitch M, Levy R, Matus-Leibovitch N, Nevo I,Vogel Z (1995) Adenylyl cyclase supersensitization in mu-opioidreceptor-transfected Chinese hamster ovary cells followingchronic opioid treatment. J Biol Chem 270:29732–29738
Baratti-Elbaz C, Ghinea N, Lahuna O, Loosfelt H, Pichon C, MilgromE (1999) Internalization and recycling pathways of the thyrotro-pin receptor. Mol Endocrinol 13:1751–1765
Bjorkman U, Ekholm R, Ericson LE (1978) Effects of thyrotropin onthyroglobulin exocytosis and iodination in the rat thyroid gland.Endocrinology 102:460–470
Bockaert J, Pin J (1999) Molecular tinkering of G protein coupledreceptors: an evolutionary success. EMBO J 18:1723–1729
Davies T, Marians R, Latif R (2002) The TSH receptor reveals itself. JClin Invest 110:161–164
Ericson LE, Nilsson M (2000) Deactivation of TSH receptor signalingin filter-cultured pig thyroid epithelial cells. Am J PhysiolEndocrinol Metab 278:E611–E619
Fan QR, Hendrickson WA (2005) Structure of human follicle-stimulating hormone in complex with its receptor. Nature433:269–277
Goudet C, Gaven F, Kniazeff J, Vol C, Liu J, Cohen-Gonsaud M,Acher F, Prézeau L, Pin JP (2004) Heptahelical domain ofmetabotropic glutamate receptor 5 behaves like rhodopsin-likereceptors. Proc Natl Acad Sci USA 101:378–383
Heldin N-E, Gustavsson B, Hermansson A, Westermark B (1994)Thyrotropin (TSH)-induced receptor internalization in nonthyroidal cells transfected with a human TSH-receptor comple-mentary deoxyribonucleic acid. Endocrinology 134:2032–2036
Hoffmann C, Gaietta G, Bünemann M, Adams SR, Oberdorff-MaassS, Behr B, Vilardaga J-P, Tsien RY, Ellisman MH, Lohse MJ(2005) A FlAsH-based FRET approach to determine G protein−coupled receptor activation in living cells. Nat Methods 2:171–176
Hsu SY, Kudo M, Chen T, Nakabaiashy K, Bhalla A, Van der SpekPJ, Van Dujin M, Hsueh AJ (2000) The three subfamilies ofleucine rich repeat containing G protein-coupled receptors
(LGR): identification of LGR6 and LGR7 and the signallingmechanism for LGR7. Mol Endocrinol 14:1257–1271
Johnson RA, Salomon Y (1991) Assay of adenylyl cyclase catalyticactivity. Methods Enzymol 195:3–21
Lefkowits RJ (2004) Historical review: a brief history and personalretrospective of seven-transmembrane receptors. Trends Pharma-col 25:413–422
Maggio R, Innamorati G, Parenti M (2007) G-protein-coupledreceptor oligomerization provides the framework for signaldiscrimination. J Neurochem 103:1741–1752
Mirashi M, Milgrom E (1997) The TSH receptor. In: Weetman AP,Grossman A (eds) Pharmacotherapeutics of the thyroid gland.Springer, Berlin Heidelberg New York, pp 33–73
Mori M, Tajima K, Miyagawa J, Shimizu M, Mashita K, Sugihara S,Hamaoka T, Fujiwara H, Tarui S (1988) Release of 3,5,3′-triiodothyronine, thyroxine and thyroglobulin from TSH-stimu-lated mouse thyroids in the perfusion system. Cell Mol Life Sci44:766–768
Olah ME, Stiles GL (1995) Adenosine receptor subtypes: character-ization and therapeutic regulation. Annu Rev Pharmacol Toxicol35:581–606
Ourlin JC, Vilarem MJ, Daujat M, Harricane MC, Domergue J,Joyeux H, Baulieux J, Maurel P (1997) Lipid-mediated transfec-tion of normal adult human hepatocytes in primary culture. AnalBiochem 247:34–44
Parnot C, Miserey-Lenkei S, Bardin S, Corvol P, Clauser E (2002)Lessons from constitutively active mutants of G protein-coupledreceptors. Trends Endocrinol Metab 13:336–343
Perret J, Ludgate M, Libert F (1990) Stable expression of the humanTSH receptor in CHO cells and characterization of differentiallyexpressing clones. Biochem Biophys Res Comm 171:1044–1050
Rossi M, Dimida A, Dell’anno MT, Trincavelli ML, Agretti P, GiorgiF, Corsini GU, Pinchera A, Vitti P, Tonacchera M, Maggio R(2007) The thyroid disruptor 1,1,1-trichloro-2,2-bis(p-chloro-phenyl)-ethane appears to be an uncompetitive inverse agonistfor the thyrotropin receptor. J Pharmacol Exp Ther 320:465–474
Sanders J, Chirgadze DY, Sanders P, Baker S, Sullivan A, BhardwajaA, Bolton J, Reeve M, Nakatake N, Evans M, Richards T, PowellM, Miguel RN, Blundell TL, Furmaniak J, Smith BR (2007)Crystal structure of the TSH receptor in complex with a thyroid-stimulating autoantibody. Thyroid 17:395–410
Santini F, Vitti P, Ceccarini G, Mammoli C, Rosellini V, Pelosini C,Marsili A, Tonacchera M, Agretti P, Santoni T (2003) In vitroassay of thyroid disruptors affecting TSH-stimulated adenylatecyclase activity. J Endocrinol Invest 26:950–955
Shimon I, Cohen O, Lubetsky A, Olchovsky D (2002) Thyrotropinsuppression by thyroid hormone replacement is correlated withthyroxine level normalization in central hypothyroidism. Thyroid12:823–827
Singh SP, McDonald D, Hope TJ, Prabhakar BS (2004) Uponthyrotropin binding the thyrotropin receptor is internalized andlocalized to endosome. Endocrinology 145:1003–1010
Stroffekova K, Proenza C, Beam KG (2001) The protein-labelingreagent FLASH-EDT2 binds not only to CCXXCC motifs butalso non-specifically to endogenous cysteine-rich proteins.Pflugers Arch 442:859–866
Vassart G, Pardo L, Costagliola S (2004) A molecular dissection of theglycoprotein hormone receptors. Trends Biochem Sci 29:119–126
Yang YW, Yang JC (1997) Studies of DEAE-dextran-mediated genetransfer. Biotechnol Appl Biochem 25:47–51
Zhang M, Tong KP, Fremont V, Chen J, Narayan P, Puett D,Weintraub BD, Szkudlinski MW (2000) The extracellular domainsuppresses constitutive activity of the transmembrane domain ofthe human TSH receptor: implications for hormone-receptorinteraction and antagonist design. Endocrinology 141:3514–3517
40 Cell Tissue Res (2009) 336:31–40