the subcellular distribution of rat liver serine-pyruvate aminotransferase
TRANSCRIPT

Biochem. J. (1982) 202,483-490 483Printed in Great Britain
The subcellular distribution of rat liver serine-pyruvate aminotransferase
Kathleen V. ROWSELL, Lamia M. R. AL-NAAMA* and Peter BENETTDepartment ofMedical Biochemistry, The Medical School, Manchester University, Manchester M13 9PT,
U.K.
(Received 5 August 1981/Accepted 9 November 1981)
1. The subcellular distribution of L-serine-pyruvate aminotransferase activity in rat liverwas investigated. About 80% was recovered from cell-free homogenates in a'total-particles' fraction and the remainder in the cytosol. 2. Subfractionation of theparticles by differential sedimentation and on sucrose density gradients showed adistribution for serine-pyruvate aminotransferase activity closely matching thatobserved for mitochondrial marker enzymes. 3. A study of the solubilization of enzymesfrom combined subcellular particles by digitonin at various concentrations alsoindicated a common subcellular location for serine-pyruvate aminotransferase andestablished mitochondrial enzymes. 4. The increase in liver serine-pyruvate amino-transferase activity induced by glucagonmitochondrial activity.
Accounts of the subcellular distribution of ratliver L-serine-pyruvate aminotransferase (EC2.6.1.51) vary. Without presenting supporting dataRowsell et al. (1972a) report the enzyme as whollyparticulate. Snell (1975, 1976) assigned 90% of theactivity to homogenate particles with 10% cytosolicand presented evidence that, in suckling rats at least,the particle association is with mitochondria.Noguchi et al. (1976, 1978) report recoveries of16% and 80% for the cytosol and combinedparticles respectively, 50-55% of the particle-boundenzyme being in the peroxisomes with the remainderin the mitochondria. Hoshino et al. (1977) claimedthat about 75% of the activity is in the cytosol andabout 25% in the mitochondria, though the latterwas a crude mitochondrial fraction (as described inthe present paper) containing presumably alsolysosomes and peroxisomes.
For a proper consideration of the role of ser-ine-pyruvate aminotransferase in liver metabolism,reliable knowledge as to its subcellular location isrequired. For example, it has been noted thatenzymes catalysing the reactions for the sequences[glyoxylate -.glycine -serine] and [hydroxypyru-vate -_D-glycerate --2-phosphoglycerateI are largelylocated in rat liver mitochondria (Rowsell et al.,1972a). [Kitagawa & Sugimoto (1979) reported acytosolic location for D-glycerate dehydrogenase,the enzyme responsible for the reduction of hydroxy-pyruvate, but deficiencies in the assay methodemployed make this a dubious claim]. If the enzyme
* Present address: University of Basrah, Basrah, Iraq.
injection was accounted for as an increased
for the linking reaction serine -.hydroxypyruvatewere also principally sited in the mitochondria, itwould be possible and reasonable to visualize asequence of reactions leading towards glucoseformation from glyoxylate, glycine or serine, theoverall operation being facilitated by the proximityof all the enzymes concerned. The subcellular distri-bution of rat liver serine-pyruvate aminotransferasewas re-investigated.
MethodsPreparation ofhomogenates
Male Sprague-Dawley rats (200-250g) starvedfor 24 h were stunned and bled to death from theblood vessels of the neck. To avoid contamination ofthe liver homogenates with blood cells, immediatelythe liver was perfused in situ with Ca2+-free Locke'ssolution for 3-4 min. Perfusion with Ca2+-free salinealso softens the intercellular cement and facilitatesthe disruption of the tissue (Anderson, 1953). Theliver was then quickly removed into a tared beaker inice, weighed and finely chopped with scissors. Thepreparation of homogenates (10% w/v) in un-buffered 0.4M-sucrose was as described by Rowsellet al. (1972a). Under these conditions, in ourexperience, mitochondria at least, judging by theirretention of glutamate dehydrogenase and appear-ance under the electron microscope, are morereliably maintained intact than when homogenatesare prepared in 0.25 M-sucrose, with or withoutadditives.
0306-3283/82/020483-08$01.50/1 ©) 1982 The Biochemical SocietyVol. 202

K. V. Rowsell, L. M. R. Al-Naama and P. Benett
SubcellularfractionationAll centrifugation procedures were performed at
2-40C and all fractions were stored on ice untilrequired for enzyme activity assays. Differentialsedimentation was carried out in the 8 x 25 ml anglerotor of the MSE65 centrifuge or, for the isolation ofnuclei, in the 8 x 50ml swing-out rotor of the Mistral4L centrifuge. For density-gradient fractionation the3 x 23 ml and 3 x 70ml swing-out rotors of theMSE65 were employed, or the B- 14 zonal rotor.
Differential sedimentationfractionationIn separate experiments the following fractiona-
tions were made. (1) The homogenate was centri-fuged at 150000g (raV. 6.9cm) for 100min to give aclear supernatant cytosol fraction and a sedimentedtotal-particles fraction. (2) A nuclear fraction wassedimented at 550g (raV 7.7 cm) for 2 min leaving asupernatant containing other subcellular particulatecomponents with the cytosol. The nuclear fractionwas washed three times by resuspension in 0.4M-sucrose containing 0.2 mM-CaCl2 and recentrifuga-tion as described above before a final resuspension inthe same medium. (3) Nuclei were sedimented asdescribed in (2) and discarded. The supernatant wascentrifuged at 30000g (raV 6.9cm) for 60min toobtain a clear cytosol fraction and sedimentedparticles. The latter were resuspended in 0.4 M-sucrose and centrifuged at 15400g (rav 6.9cm) for10min to sediment a crude mitochondrial fractionleaving a portion of the microsomes in the super-natant. (4) After removing the nuclei the remainingsupernatant was centrifuged at 61600g (raV 6.9 cm)for 15min to obtain a cytosol fraction containinglight microsomes and a sedimented particles frac-tion. The latter was resuspended in 0.4M-sucroseand centrifuged at 11 500g (raV 6.9 cm) for 0min tosediment a crude mitochondrial fraction leavingmicrosomes in the supernatant.A Teflon homogenizer operated by hand was used
to resuspend the sedimented fractions. The originalhomogenizing medium was used except with thesedimented nuclei.
Density-gradientfractionationSucrose gradients were used in attempts to
separate mitochondria, lysosomes and peroxisomes.In some experiments, to facilitate the separation oflysosomes, rats were pre-injected with Triton WR-1339 (supplied by Hilton Davis Chemicals Ltd.,Newcastle-upon-Tyne, U.K.) as described byLeighton et al. (1968). A 10% (w/v) homogenatewas prepared and centrifuged at 500g for 3min toremove nuclei. The supernatant remaining wasrecentrifuged at 26 700g (r,V 9.4 cm) for 10min tosediment a particulate fraction (described as 'crudemitochondria') for application to a sucrose gradient.
Continuous sucrose gradients were constructed
with an LKB Ultragrad 11300 gradient-makingdevice and discontinuous gradients by pipettingdiscrete layers of sucrose solution. Details are givenin the Results section. After centrifugation, sucrosesolution, at the highest concentration for the gradientused, was introduced through the bottom of thetubes at 1.5 ml/min and fractions were collectedfrom the top by using an ISCO density gradientfractionator model 185 (Instrumental SpecialtiesCo., Lincoln, NE, U.S.A.). Alternatively fractionswere isolated with a modified Beckman tube slicer asdescribed by Turner (1964). The sucrose con-centration (% w/w) of each fraction recovered wasmeasured at 200C on an Abbe refractometer,designed to read directly both refractive index andthe associated sucrose concentration, then by referr-ing to appropriate calibration curves the sucroseconcentration (% w/v) and density were derived.
Zonal centrifugationFor each experiment a crude mitochondrial
fraction was prepared from three rats, not pre-treated with Triton WR-1339, and resuspended toobtain the equivalent of 0.5 g of original tissue/ml in55% (w/v) sucrose. A portion (30ml) was appliedover a sucrose gradient [57-70% (w/v), 520mllcushioned by 80ml of 70% (w/v) sucrose, in thezonal rotor. An overlay of 50ml of 50% (w/v)sucrose completed the loading procedure. Aftercentrifugation at 58 000g (rav 4.5 cm) for 3 h, 70%(w/v) sucrose was pumped into the periphery inletsat 20 ml/min and 20 ml fractions were collected fromthe centre.
Digitonin treatment ofparticlesA 10% (w/v) digitonin solution was prepared by
heating digitonin in 0.4M-sucrose. This was mixedwith 0.4M-sucrose in centrifuge tubes to obtain aseries of 2.0ml portions containing 0-50mg ofdigitonin/ml which were then cooled to 0°C. Toeach tube 8.0ml of a rat liver crude mitochondrialpreparation was added, and, after mixing by gentleswirling, tubes were left at 00C for 15 min. The tubeswere then centrifuged at 30000g (rav 9.4cm) for30min. The supernatant and the resuspended pelletswere then taken for enzyme assays.
Enzyme assaysTo measure the recovery of different subcellular
components the following marker enzymes wereassayed: lactate dehydrogenase (EC 1.1.1.27;cytosol), glucose 6-phosphatase (EC 3.1.3.9;microsomal), acid phosphatase (EC 3.1.3.2; lyso-somal), glutamate dehydrogenase (EC 1.4.1.2)and cytochrome oxidase (EC 1.9.3.1) (mitochon-drial), urate oxidase (EC 1.7.3.3), D-amino acidoxidase (EC 1.4.3.3) and catalase (EC 1.11.1.6)(peroxisomal); DNA was used as a marker for
1982
484

Subcellular distribution of serine-pyruvate aminotransferase
nuclei. Except as detailed below, assay procedureswere as described by Rowsell et al. (1972a).
Before making assays for glutamate dehydro-genase activity, all subcellular fractions were left at00C for 3h with 0.1% (v/v) Triton X-100. Theactivity thus measured was the same as that assayedafter sonication at 0-20C for 10min [with maxi-mum power on the MSE Ultrasonic (disintegrator)Model 150W], but activation by Triton was moreconvenient where several fractions required assay.Cytochrome oxidase was assayed by observing there-oxidation of reduced cytochrome c, as describedby Smith (1960), with fraction samples clarified bysonication or by treatment with Triton X-100 asdescribed above. D-Amino acid oxidase was assayedby measuring the pyruvate formed aerobically fromD-alanine. Assay mixtures (8.0 ml) contained 20mM-D-alanine, 50mM-potassium phosphate buffer,pH 7.4, and tissue preparation and were incubated at370C under oxygen. Samples (2.0 ml) were removedat 0, 15 and 30min into 2.Oml of 10% (w/v)trichloroacetic acid. After centrifugation supernat-ants were analysed for pyruvate with lactatedehydrogenase. Catalase was assayed by measuringthe decrease in H202 absorbance at 240nm, asdescribed by Luck (1963). The catalase was solu-bilized by pretreating fraction samples at 0°C with1% (v/v) Triton X-100 and in the presence of 0.33%bovine serum albumin to protect the enzyme frominactivation by the detergent (see Leighton et al.,1968). Samples were diluted to a suitable con-centration with 0.05M-potassium phosphate buffer,pH 7.0, immediately before assay.
L-Serine-pyruvate aminotransferase was assayedas described by Rowsell et al. (1979) and L-alanine-glyoxylate aminotransferase by measuringthe pyruvate formed with lactate dehydrogenase inthe presence of Tris at pH8.4, as described byRowsell et al. (1972b).
Checks were made on the reliability of the assayprocedures, for use in subcellular fractionationstudies, and some sucrose interference with theserine-pyruvate aminotransferase assay, (i.e. withthe determination of hydroxypyruvate), was found.When blank assays (no tissue extract) were con-ducted with samples of sucrose at high concen-tration (>60%), a slight re-oxidation of NADH wasobserved (a decrease in A340 of about 0.003) whenglycerate dehydrogenase was added to HCl04-treated, neutralized assay samples. This was attribut-able apparently to hydrolysis products of sucroseobtained on exposure to HCl04. The effect was notseen with sucrose made up and maintained atneutrality. To correct for this in subcellular-frac-tionation studies, a blank serine-pyruvate amino-transferase assay was made for each fractioninvestigated using the same volume and sucroseconcentration as in the test assay.
Vol. 202
Table 1. Whole homogenate enzyme activities in rat liverActivities are expressed as umol of substrate utilizedor product formed/min per g of liver + S.E.M. (no. ofobservations), except for catalase activity which isgiven as mmol/min per g. DNA concentrations were2220 + 200 (n = 3),ug/g of liver.
EnzymeLactate dehydrogenaseGlutamate dehydrogenaseCytochrome oxidaseGlucose 6-phosphataseAcid phosphataseUrate oxidaseD-Amino acid oxidaseCatalaseAlanine-glyoxylate aminotransferaseSerine-pyruvate aminotransferase
Activity302+14(10)33.5 + 2.2 (20)28.5 ± 2.2 (12)7.43 +0.7 (12)6.42 +0.3 (16)0.75 +0.08 (12)0.32 +0.10 (6)552 + 1.5 (4)3.76 +0.38 (12)
0.083 + 0.004 (20)
Except with cytochrome oxidase and catalase allenzyme activity values were based on linear pro-gress curves. Whole homogenate activities for theenzymes assayed in the present work are given inTable 1.
Results
There was no significant difference in serine-pyruvate aminotransferase activity when homogen-ates were made from the same rat liver in 0.4M-sucrose or in 0.1 M-potassium phosphate buffer,pH 7.4. When the activity was assayed on anhomogenate freshly prepared in 0.4M-sucrose andafter 3 h storage at 00C there was no significantdecrease (<2%). It appeared there would be littleloss of activity during fractionation.
Subcellular fractions obtained by differential sedi-mentation
Reproducibly somewhat more than 80% of theserine-pyruvate aminotransferase activity wasrecovered in a total-particles fraction and theremainder in the cytosol [Table 2, (1)]. Samples of atotal-particles preparation were sonicated at sub-maximal power for periods up to 15 min. Aftercentrifugation at 160OOOg for 100min the super-natant and the resuspended pellet were assayed forserine-pyruvate aminotransferase. A solubilizationof the enzyme was observed but at a somewhatslower rate than for glutamate dehydrogenase; yetthe recovery of serine-pyruvate aminotransferase inthe cytosol clearly exceeded that of glutamatedehydrogenase and of other particle marker en-zymes. We conclude that the serine-pyruvateaminotransferase activity found in the cytosol islocated there in the intact liver cell; it does notoriginate in the disruption of a subcellular organellein the course of homogenization or fractionation.
485

K. V. Rowsell, L. M. R. Al-Naama and P. Benett
Table 2. Enzyme activities ofsubcellularfractions isolated by differential sedimentationFractions were prepared from rat liver homogenates by differential sedimentation as described in the Methods section[procedures (1)-(4)]. Percentages of the original whole homogenate values are given.
Serine-pyruvateamino-
Experiment transferase(1)Total-particles
fractionCytosol fraction
Lactatedehydro-genase
Glutamatedehydro-genase
82 12 98 99
17 90 0.4 0
Cytochrome Glucose Acid Urateoxidase 6-phosphatase phosphatase oxidase DNA
99
2
93 92
6 0
Recovery(2)Nuclear fractionSupernatant fraction
Recovery(3)Crude mitochondrial
fractionMicrosomal fractionCytosol fraction
Recovery(4)Crude mitochondrial
fractionMicrosomal fractionCytosol + light
microsomal fraction
99 102 98.4
692
592
494
98 97 98 88
69 15 81
12 15 1225 78 6
106 108
69
99
7 84
10 1519 74
57
98 96 96
A small proportion of the serine-pyruvate amino-transferase activity was recovered in nuclear frac-tions [Table 2, (2)] and in microsomal fractions[Table 2, (3) and (4)] but contamination with markerenzymes for other subcellular components castsdoubt on a genuine location there. The results inTable 2 suggest a distribution for serine-pyruvateaminotransferase between the cytosol and particlesmainly other than nuclei or microsomes. A majorassociation with crude mitochondrial fractions was
regularly observed. These fractions contained a highproportion of the mitochondria and the lysosomesand would also be expected to include peroxisomes.
Subcellular fractions obtained by centrifugation on
density gradientsCrude liver mitochondrial fractions prepared from
rats pretreated with Triton WR-1339 were sub-jected to fractionation on a sucrose gradientdesigned to maximize the separation of lysosomesfrom mitochondria. The bulk of the acid phos-
phatase was then recoverable in the upper fractionsaccompanied by relatively little serine-pyruvateaminotransferase activity (Table 3). Other experi-ments were made with gradients designed to in-crease the separation of mitochondria from peroxi-somes (Table 4). In neither type of experiment was itpossible to obtain mitochondria uncontaminatedwith peroxisomes, but repeatedly it was observedthat the distribution of serine-pyruvate amino-transferase activity along the gradients fairly closelyparalleled those of the mitochondrial marker en-zymes and this was not observed with the peroxi-
somal or lysosomal markers.Zonal centrifugation experiments were made in
further attempts to improve the separation ofmitochondria and peroxisomes. As indicated by thedistribution of glutamate dehydrogenase and cyto-chrome oxidase, mitochondria were recovered in arelatively restricted band centred on a density of1.20 g/ml. However, the peroxisomal marker en-
zymes (catalase, D-amino acid oxidase and urate
1982
99
286
101
082
82
99 92
498
7625
102
21
101
659
95
Recovery
66
2019
105
70
2415
16
6828
112 109
486

Subcellular distribution of serine-pyruvate aminotransferase
Table 3. Enzyme activities of subcellular fractions isolated by centrifugation on a sucrose gradient designed to separatelysosomes and mitochondria
A crude mitochondrial fraction was prepared from a rat pre-injected with Triton WR-1339, as described in theMethods section, and resuspended to obtain the equivalent of 0.2 g of original tissue/ml in 0.4M-sucrose. Portions(10ml) were applied over continuous sucrose gradients [15.4-80% (w/v), 52ml] and after centrifugation at 9300Og(rav. 9.4 cm) for 90 min fractions were collected sequentially from the top of the gradient. Enzyme activities recoveredare presented as percentages of crude mitochondrial activities and as relative concentrations (R), i.e. % recovery/%gradient volume.
FractionGlutamate
dehydrogenase
(Yo) R0 01.6 0.061.0 0.05
93 3.2
Cytochrome Acidoxidase phosphatase
(%) R (%) R0 0 7.5 0.361 0.03 76 2.62 0.10 7.0 0.33
93 3.2 9.7 0.33
Urate Serine-pyruvateoxidase aminotransferase
(%) R (%) R00 0 00 0 4 0.140 0 12 0.5784 2.9 81 2.8
Table 4. Enzyme activities of subcellular fractions isolated by centrifugation on a sucrose gradient designed to separatemitochondria andperoxisomes
A crude mitochondrial fraction was prepared as described in Table 2. It was resuspended to obtain the equivalent of0.2 g of original tissue/ml in 50% (w/v) sucrose, density 1.19 g/ml. Portions (15 ml) were applied over continuoussucrose gradients [58-70% (w/v), 35ml] and then overlaid with 12ml of 0.4M-sucrose. After centrifugation at7800Og (ray. 11.3cm) for 180min, fractions were collected from the top of the gradient. Enzyme activities areexpressed as in Table 3.
FractionA
Density Volume of total(g/ml) (ml) volume
(a) 1.19 31.5 51(b) 1.23 4.7 8(c) 1.25 5.5 9
Glutamate Cytochrome Acid Urate D-Amino Serine-pyruvatedehydrogenase oxidase phosphatase oxidase acid oxidase aminotransferase
(%) R (%) R (%o) R (%) R (%o) R (%o) R60 1.18 55 1.08 88 1.73 36 0.71 53 1.04 55 1.0827 3.38 3 1 3.84 2 0.25 14.5 1.8 10 1.25 27 3.3812 1.30 16 1.78 2 0.22 34 3.73 34 3.78 15 1.67
oxidase) were recovered to some degree throughoutthe gradient. Total enzyme recoveries were >90%,and we can only conclude that with this technique, inour hands, the peroxisomes had suffered damage.Nevertheless the findings were again consistent witha mitochondrial location for serine-pyruvate amino-transferase. Its activity distribution, together withthat of alanine-glyoxylate aminotransferase, closelymatched those of the mitochondrial markerenzymes.
Differential extraction ofparticle-bound enzymes bydigitonin
Crude mitochondrial preparations were treatedwith different concentrations of digitonin in 0.4M-sucrose. After centrifugation, supernatants andresuspended pellets were assayed for serine-pyru-vate aminotransferase activity and for mitochon-drial and peroxisomal marker enzymes (Fig. 1).Assays for urate oxidase were not included since, as
Vol. 202
a peroxisomal 'core' enzyme (see de Duve &Baudhuin, 1966), it was expected it would not besolubilized by digitonin.
There was some release of catalase from theparticles even in the absence of digitonin. (Thissuggests that there will be a loss of this enzyme fromthe peroxisomes in the course of subcellular frac-tionation and that catalase is not therefore an idealperoxisomal marker.) But at the lowest digitoninconcentration investigated the release of catalasewas greatly increased and for 5mg of digitonin/mlthere was almost a total loss from the particles withan equivalent recovery in the supernatant. The lossof D-amino acid oxidase from the particles showedthe same dependence on digitonin but unaccount-ably no activity was measurable in the super-natants.
Higher concentrations of digitonin were needed tobring about the solubilization of glutamate de-hydrogenase, and for increasing digitonin concen-
Density(g/ml)
(a) 1.05(b) 1.10(c) 1.15(d) 1.23
Volume(ml)13181318
of totalvolume
21292129
487
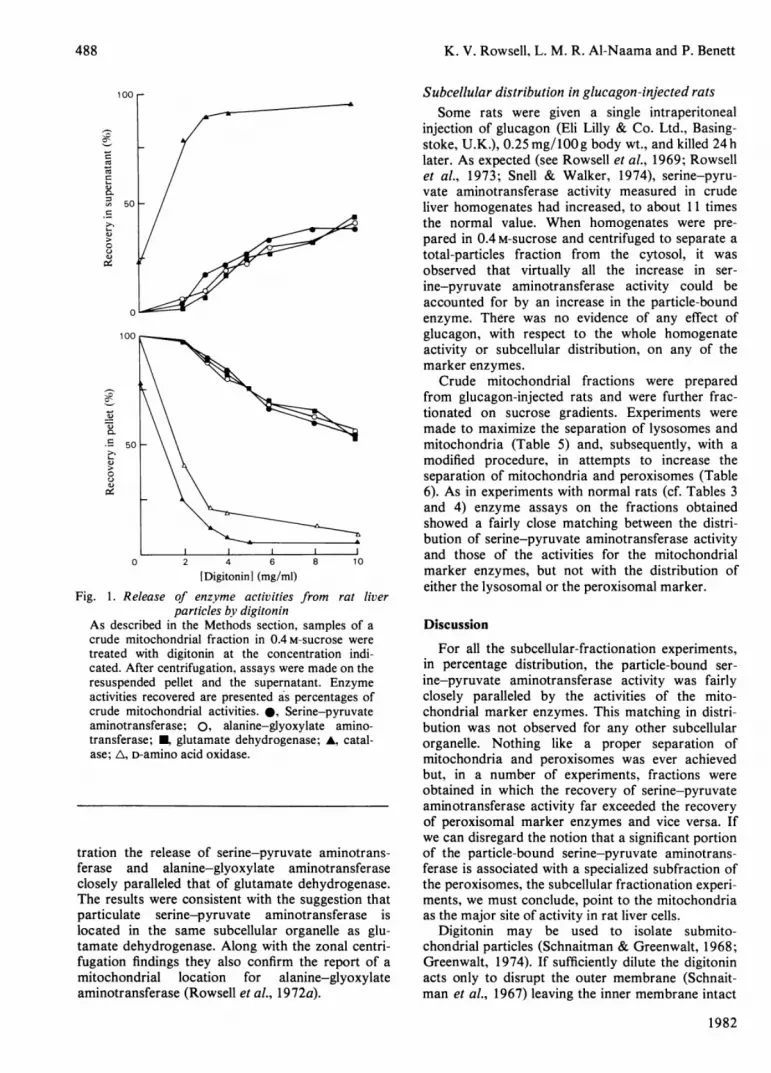
K. V. Rowsell, L. M. R. Al-Naama and P. Benett
u: 50 t\
50
0~
0
500
50
0 2 4 6
[Digitonin] (mg/mi)
Fig. 1. Release of enzyme activities fiparticles by digitonin
As described in the Methods section,
crude mitochondrial fraction in 0.4 m-s5treated with digitonin at the concent
cated. After centrifugation, assays were
resuspended pellet and the supernataiactivities recovered are presented a's pe'
crude mitochondrial activities. 0, Serir
aminotransferase; 0, alanine-glyoxyl
transferase; glutamate dehydrogenasase; A, D-amino acid oxidase.
tration the release of serine-pyruvatcferase and alanine-glyoxylate amiclosely paralleled that of glutamate deThe results were consistent with the siparticulate serine-pyruvate aminotilocated in the same subcellular orgEtamate dehydrogenase. Along with thefugation findings they also confirm ttmitochondrial location for alaniaminotransferase (Rowsell et al., 1972w
Subcellular distribution in glucagon-injected ratsSome rats were given a single intraperitoneal
injection of glucagon (Eli Lilly & Co. Ltd., Basing-stoke, U.K.), 0.25 mg/lOOg body wt., and killed 24hlater. As expected (see Rowsell et al., 1969; Rowsellet al., 1973; Snell & Walker, 1974), serine-pyru-vate aminotransferase activity measured in crudeliver homogenates had increased, to about 11 times
Zt_ - the normal value. When homogenates were pre-pared in 0.4 M-sucrose and centrifuged to separate atotal-particles fraction from the cytosol, it wasobserved that virtually all the increase in ser-ine-pyruvate aminotransferase activity could beaccounted for by an increase in the particle-boundenzyme. There was no evidence of any effect ofglucagon, with respect to the whole homogenateactivity or subcellular distribution, on any of themarker enzymes.
Crude mitochondrial fractions were preparedfrom glucagon-injected rats and were further frac-tionated on sucrose gradients. Experiments weremade to maximize the separation of lysosomes andmitochondria (Table 5) and, subsequently, with amodified procedure, in attempts to increase theseparation of mitochondria and peroxisomes (Table6). As in experiments with normal rats (cf. Tables 3and 4) enzyme assays on the fractions obtainedshowed a fairly close matching between the distri-
A bution of serine-pyruvate aminotransferase activityand those of the activities for the mitochondrialmarker enzymes, but not with the distribution ofeither the lysosomal or the peroxisomal marker.
rom rat liver
-amples of a Discussionucrose were
For all the subcellular-fractionation experiments,made on the in percentage distribution, the particle-bound ser-nt. Enzyme ine-pyruvate aminotransferase activity was fairlyrcentages of closely paralleled by the activities of the mito-ne-pyruvate chondrial marker enzymes. This matching in distri-late amino- bution was not observed for any other subcellularse; A, catal- organelle. Nothing like a proper separation of
mitochondria and peroxisomes was ever achievedbut, in a number of experiments, fractions wereobtained in which the recovery of serine-pyruvateaminotransferase activity far exceeded the recoveryof peroxisomal marker enzymes and vice versa. Ifwe can disregard the notion that a significant portion
e aminotrans- of the particle-bound serine-pyruvate aminotrans-inotransferase ferase is associated with a specialized subfraction ofehydrogenase. the peroxisomes, the subcellular fractionation experi-uggestion that ments, we must conclude, point to the mitochondriaransferase is as the major site of activity in rat liver cells.anelle as glu- Digitonin may be used to isolate submito-zonal centri- chondrial particles (Schnaitman & Greenwalt, 1968;
ie report of a Greenwalt, 1974). If sufficiently dilute the digitoninne-glyoxylate acts only to disrupt the outer membrane (Schnait-a). man et al., 1967) leaving the inner membrane intact
1982
488
Subcellular distribution of serine-pyruvate aminotransferase
Table 5. Enzyme activities of subcellularfractions isolatedfrom glucagon-injected rats on a sucrose gradient designed toseparate lysosomes and mitochondria
A crude mitochondrial fraction was prepared from two rats pre-injected with Triton WR-1339 (see the Methodssection) and 24h after glucagon injection (see the Results section). It was resuspended to obtain the equivalent of0.2g of original tissue/ml in 0.4M-sucrose. Portions (5ml) were applied over discontinuous sucrose gradientsr40-80% (w/v), 15 ml] and, after centrifugation at 93 000g (ray. 9.4 cm) for 180min, fractions were isolated with theuse of a tube slicer. Enzyme activities are as expressed in Table 3.
Fraction
Density Volume of total(g/ml) (ml) volume
(a) 1.05 5.3 26.5(b) 1.10 1.5 7.5(c) 1.19 2.3 11.5(d) 1.20 5.5 27.5(e) 1.23 2.4 12.0(f) 1.27 3.0 15.0
Glutamatedehyrogenase
(%) R0 01.1 0.15
29 2.5264 2.332.3 0.190 0
Cytochromeoxidase
(%) R0 02.8 0.37
41 3.5752 1.892.7 0.220 0
Acidphosphatase
(%) R38 1.434 4.513 1.138 0.290 00 0
Urate Serine-pyruvateoxidase aminotransferase
(%) R (%) R0 0 0 00 0 1.5 0.208.2 0.71 32 2.78
33 1.20 63 2.2928 2.33 3.2 0.2710 0.67 0 0
Table 6. Enzyme activities of subcellular fractions isolated from glucagon-injected rats on a sucrose gradient designedto separate mitochondria andperoxisomes
A crude mitochondrial fraction was prepared from two rats 24h after glucagon injection (see the Results section)but without Triton pre-treatment. It was resuspended to obtain the equivalent of 0.2g of original tissue/ml in 50%(w/v) sucrose. The fractionation procedure was as described in Table 5 except that the discontinuous sucrosegradient used was 60-80% (w/v). Enzyme activities are expressed as in Table 3.
Fraction__A 5 Glutamate% dehydrogenase
Density Volume of total ,
(g/ml) (ml) volume (%) R(a) 1.19 7 35 91 2.60(b) 1.23 4 20 4.5 0.23(c) 1.24 3 15 1.3 0.09(d) 1.25 1.7 8.5 0 0(e) 1.27 4.3 21.5 0 0
Cytochrome Acidoxidase phosphatase
(%) R (%) R92 2.63 81 2.311.5 0.08 12 0.600.9 0.06 5 0.330 0 0 00 0 0 0
Urateoxidase
(%) R32 0.909.1 0.4619 1.2712 1.4110 0.47
Serine-pyruvateaminotransferase
A
(%) R86 2.468.7 0.441.5 0.100 00 0
and enclosing the matrix enzymes. As the con-centration of digitonin is increased the inner mem-brane too becomes permeable or is disrupted and thematrix contents leak out (Schnaitman et al., 1967;Cassidy et al., 1972). Digitonin is known to act alsoon the peroxisomal membrane to release solubleenzymes such as D-amino acid oxidase and catal-ase, though not urate oxidase, which is associatedwith the insoluble core of the peroxisomes (de Duve& Baudhuin, 1966). It appears from the presentexperiments that the critical concentration of digi-tonin required to solubilize the mitochondrial matrixenzymes (e.g. glutamate dehydrogenase) is higherthan that needed for the liberation of the solubleperoxisomal enzymes. It is also clear that the releaseof serine-pyruvate aminotransferase, with theapplication of increasing concentrations of digi-
tonin, much more closely parallels the release of themitochondrial matrix marker than it does that of theperoxisomal enzymes. The results of the digitoninexperiments agree with the present findings onseparated subcellular fractions in showing mito-chondria as the likely site of the particle-boundserine-pyruvate aminotransferase in rat liver.
The present finding that the bulk (>80%) of ratliver serine-pyruvate aminotransferase activity isparticle-bound accords with the observations ofSnell (1975, 1976) and Noguchi et al. (1976). Incontrast Hoshino et al. (1977) found the largerproportion (about 75%) to be cytosolic. Thisdisagreement may be attributable to the differencesin the homogenization medium employed. Hoshinoet al. (1977) used O.1M-sucrose containing 0.1M-potassium phosphate buffer, pH 7.6, and 1 mM-
Vol. 202
489

490 K. V. Rowsell, L. M. R. Al-Naama and P. Benett
EDTA. We have used 0.4 M-sucrose with noaddition, basing this procedure on earlier findingsthat salt additions to the sucrose are conducive tothe leakage of mitochondrial enzymes (Turner,1964).We have no satisfactory explanation for the
discrepancy between present findings and those ofNoguchi et al. (1978) who report a somewhat higherrecovery of serine-pyruvate aminotransferase inperoxisomes than in mitochondria after zonalcentrifugation on a sucrose gradient.Our results, indicating a distribution of liver
serine-pyruvate aminotransferase activity betweenmitochondria (about 80%) and cytosol (about 20%),are essentially in agreement with Snell's (1975,1976) findings for suckling rats. It appears that thisdistribution is not a special feature of the neonatalstage of development. And the distribution, we find,is still between mitochondria and cytosol when liverserine-pyruvate aminotransferase activity is greatlyincreased in rats by glucagon injection; the addi-tional activity is apparently in the mitochondria.Fukushima et al. (1978) also conclude that thisactivity is largely located in mitochondria from bothnormal and glucagon-treated rats, but insufficientdetail is given on the distribution of serine-pyruvateaminotransferase and marker enzymes to allow areally critical assessment of their findings.
With the rat some observations have suggestedserine-pyruvate aminotransferase as the liver en-zyme principally involved in initiating glucoseformation from serine, but other findings favourserine dehydratase (see e.g. Rowsell et al., 1973;Sandoval & Sols, 1974). This uncertainty appears tobe resolved by the results of experiments made onperfused rat livers and with the rat in vivo whichindicate that gluconeogenesis from serine is pre-dominantly via hydroxypyruvate in the normally-fedrat but that a much higher rate (approx. x 8) in 24 hstarved rats depends on almost equal contributionsvia pyruvate and hydroxypyruvate (Bhatia et al.,1975). Present work suggests that serine conversioninto hydroxypyruvate may occur largely in themitochondria, and the increase in mitochondrialserine-pyruvate aminotransferase activity in res-ponse to glucagon is consistent with a gluco-neogenic role for the enzyme sited there, though thisis not apparently the mechanism responsible for anenhanced flux via hydroxypyruvate in starved rats.The improved capacity for glucose formation fromserine after 24h starvation is not accompanied byany increase in liver homogenate serine-pyruvateaminotransferase activity. One explanation would bethat the portion of the extra gluconeogenesis
attributable to hydroxypyruvate formation was dueto activation of the aminotransferase and thispossibility deserves investigation with both themitochondrial and cytosolic enzymes.
References
Anderson, N. G. (1953) Science 117, 627-628Bhatia, S. C., Bhatia, S. & Rous, S. (1975) Life Sci. 17,
267-274Cassidy W. E., Leiter, E. H., Berquist, A. & Wagner, R.
P. (1972) J. Cell Biol. 53, 66-72de Duve, C. & Baudhuin, P. (1966) Physiol. Rev. 46,
323-357Fukushima, M., Aihara, Y. & Ichiyama, A. (1978) J.
Biol. Chem. 253, 1187-1194Greenwalt, J. W. (1974) Methods Enzymol. 31, 310-323Hoshino, J., Kiihne, U., Filjak, B. & Kr6ger, H. (1977) Z.
Naturforsch. 32c, 249-253Kitagawa, Y. & Sugimoto, E. (1979) Biochim. Biophys.
Acta 582, 276-282Leighton, F., Poole, B., Beaufay, H., Baudhuin, P.,
Coffey, J. W., Fowler, S. D. & de Duve, C. (1968) J.CellBiol. 37, 482-513
Luck, H. (1963) in Methods of Enzymatic Analysis(Bergmeyer, H.-U., ed.), pp. 885-894, Academic Press,New York and London
Noguchi, T., Okuno, E. & Kido, R. (1976) Biochem. J.159, 607-613
Noguchi, T., Minatogawa, Y., Takada, Y., Okuno, E. &Kido, R. (1978) Biochem. J. 170, 173-175
Rowsell, E. V, Snell, K., Carnie, J. A. & Al-Tai, A. H.(1969)Biochem.J. 115, 1071-1073
Rowsell, E. V., Snell, K., Carnie, J. A. & Rowsell, K. V.(1972a)Biochem.J. 127, 155-165
Rowsell, E. V., Carnie, J. A., Snell, K. & Taktak, B.(1972b) Int. J. Biochem. 3, 247-257
Rowsell, E. V., Al-Tai, A. H., Carnie, J. A. & Rowsell, K.V. (1973) Biochem. J. 134, 349-351
Rowsell, E. V., Carnie, J. A., Wahbi, S. D., Al-Tai, A. H.& Rowsell, K. V. (1979) Comp. Biochem. Physiol.63B, 543-555
Sandoval, I. V. & Sols, A. (1974) Eur. J. Biochem. 43,609-616
Schnaitman, C. & Greenwalt, J. W. (1968) J. Cell Biol.38, 158-175
Schnaitman, C., Erwin, V. G. & Greenwalt, J. W. (1967)J. CellBiol. 32, 719-735
Smith, L. (1960) Methods Biochem. Anal. 2,426-434Snell, K. (1975) FEBS Lett. 55, 202-205Snell, K. (1976) in The Use of Isolated Liver Cells and
Kidney Tubules in Metabolic Studies (Tager, J. M.,Soling, H. D. & Williamson, J. R., eds.), pp. 118-121,North-Holland, Amsterdam
Snell, K. & Walker, D. G. (1974) Biochem. J. 144,519-531
Turner, K. V. (1964) Ph.D. Thesis, University ofManchester
1982