the roles of geological history and colonization abilities in genetic differentiation between...
TRANSCRIPT

ORIGINALARTICLE
The roles of geological history andcolonization abilities in geneticdifferentiation between mammalianpopulations in the Philippine archipelago
Lawrence R. Heaney1*, Joseph S. Walsh, Jr1,2 and A. Townsend Peterson1
1Field Museum of Natural History, 1400 South
Lake Shore Drive, Chicago, USA, and2Undergraduate Program in Biological
Sciences, Northwestern University, Evanston,
IL, USA
*Correspondence: Lawrence R. Heaney, Field
Museum of Natural History, 1400 S. Lake Shore
Drive, Chicago, IL 60605, USA.
E-mail: [email protected]
Present address: A. Townsend Peterson, Natural
History Museum, The University of Kansas,
Lawrence, KS, USA.
ABSTRACT
Aim To test hypotheses that: (1) late Pleistocene low sea-level shorelines (rather
than current shorelines) define patterns of genetic variation among mammals on
oceanic Philippine islands; (2) species-specific ecological attributes, especially
forest fidelity and vagility, determine the extent to which common genetic
patterns are exhibited among a set of species; (3) populations show reduced
within-population variation on small, isolated oceanic islands; (4) populations
tend to be most highly differentiated on small, isolated islands; and (5) to assess
the extent to which patterns of genetic differentiation among multiple species are
determined by interactions of ecological traits and geological/geographic
conditions.
Location The Philippine Islands, a large group of oceanic islands in Southeast
(SE) Asia with unusually high levels of endemism among mammals.
Methods Starch-gel electrophoresis of protein allozymes of six species of small
fruit bats (Chiroptera, Pteropodidae) and one rodent (Rodentia, Muridae).
Results Genetic distances between populations within all species are not
correlated with distances between present-day shorelines, but are positively
correlated with distances between shorelines during the last Pleistocene period of
low sea level; relatively little intraspecific variation was found within these
‘Pleistocene islands’. Island area and isolation of oceanic populations have only
slight effects on standing genetic variation within populations, but populations
on some isolated islands have heightened levels of genetic differentiation, and
reduced levels of gene flow, relative to other islands. Species associated with
disturbed habitat (all of which fly readily across open habitats) show more genetic
variation within populations than species associated with primary rain forest (all
of which avoid flying out from beneath forest canopy). Species associated with
disturbed habitats, which tend to be widely distributed in SE Asia, also show
higher rates of gene flow and less differentiation between populations than species
associated with rain forest, which tend to be Philippine endemic species. One rain
forest bat has levels of gene flow and heterozygosity similar to the forest-living
rodent in our study.
Main conclusions The maximum limits of Philippine islands that were reached
during Pleistocene periods of low sea level define areas of relative genetic
homogeneity, whereas even narrow sea channels between adjacent but
permanently isolated oceanic islands are associated with most genetic variation
within the species. Moreover, the distance between ‘Pleistocene islands’ is
correlated with the extent of genetic distances within species. The structure of
genetic variation is strongly influenced by the ecology of the species,
predominantly as a result of their varying levels of vagility and ability to
tolerate open (non-forested) habitat. Readily available information on ecology
Journal of Biogeography (J. Biogeogr.) (2005) 32, 229–247
ª 2005 Blackwell Publishing Ltd www.blackwellpublishing.com/jbi 229

INTRODUCTION
The search for an understanding of the causes of differenti-
ation and diversification among island populations has been
an intellectual crucible in evolutionary biology. From the
original ruminations of Darwin and Wallace on the geographic
circumstances of speciation to observations of natural selection
in action, islands have provided a wealth of insight for
biologists (e.g. Grant, 1998; Hall & Holloway, 1998; Whittaker,
1998; Avise, 2000; Schluter, 2000). In particular, the presence
of high levels of endemism, and the processes and circum-
stances that produce those endemic species, has attracted
much attention. Clearly, part of the great appeal of island
systems is their relative simplicity: on islands, terrestrial
populations are discretely bounded, gene flow is likely to be
limited between them, island areas and between-island
distances are easily measured, and island communities tend
to have fewer species than mainland communities. Perhaps this
apparent simplicity has tended to wed biologists to simple
explanations for island phenomena. Biogeographers, in par-
ticular, have often divided into camps preferring either
historical (e.g. Rosen, 1975) or ecological (e.g. MacArthur &
Wilson, 1967) explanations of island phenomena. The
extensive bodies of work by both groups convinces us that
both ecological and historical factors must be important in
generating intraspecific and interspecific diversity in island
settings and that integration of the two perspectives is essential.
We agree with recent authors that historical and ecological
factors should be treated as complementary variables, rather
than competing hypotheses, in explaining patterns of geo-
graphic variation within species (e.g. Bermingham & Moritz,
1998; Whittaker, 1998; Heaney, 2000; Lomolino, 2000; Zink
et al., 2000; Riddle & Hafner, in press).
Recent developments, such as the use of DNA-based
population phylogenies, have been useful in resolving ques-
tions regarding the importance of historical and ecological
factors in influencing patterns of geographic variation in single
species (e.g. Avise, 2000). However, the particular history of
any single taxon will necessarily represent only a portion of
general patterns and causes of differentiation within any large,
historically complex region; this means that general patterns
often cannot be perceived, and general hypotheses often
cannot be tested, based on single species (Powers et al., 1991;
Riddle & Hafner, in press). A multi-species comparative
approach should be most useful for detecting the role of
ecological and historical factors in generating patterns of
variation and differentiation, helping to distinguish their
relative importance, and determining the manner in which
they interact to produce phylogenies and patterns of biological
diversity (e.g. Zink et al., 2000; Hewitt, 2001; Ricklefs &
Bermingham, 2001; Arbogast & Kenagy, 2001; Riddle &
Hafner, in press).
The Philippine archipelago is an exceptional theatre in
which to investigate the roles of past history and current
ecology in structuring geographic variation. The 7000 islands
originated as a set of de novo oceanic islands [with the
exception of one group that was united with mainland
Southeast (SE) Asia] of varying ages and geological histories,
as summarized below. It is an area of high biotic diversity and
exceptional endemism that is in critical need of conservation
(Myers, 1988; Wildlife Conservation Society of the Philippines,
1997; Heaney & Regalado, 1998; Mittermeier et al., 1999;
Holloway, 2003; Mey, 2003). While it is noteworthy that at
least 111 of the 170 native species of terrestrial mammals
(64%) are endemic (Heaney et al., 1998), it is still more
striking that 24 of 84 genera (29%) are endemic, implying
much in situ diversification, and phylogenetic studies suggest
that several large endemic clades are present among fruit bats
and murid rodents (Heaney & Rickart, 1990; Heaney, 2000;
Steppan et al., 2003). Each oceanic island that has remained
continuously isolated from its neighbouring islands is a unique
centre of mammalian endemism, with 25–80% of the non-
volant mammals endemic, even on islands of only a few
hundred square kilometres. Similar patterns are evident among
butterflies (Holloway, 2003) and trichopteran insects (Mey,
2003).
The manner in which this diversification has arisen among
Philippine mammals over evolutionary time, and the ecolog-
ical means by which it has been maintained, have been the
subject of diverse studies of biogeography, diversity gradients,
systematics, and population biology (e.g. Heaney, 1986, 1991a,
2000, 2001; Heideman & Heaney, 1989; Heaney & Rickart,
(habitat association and vagility) and geological circumstances (presence or
absence of Pleistocene land-bridges between islands, and distance between
oceanic islands during periods of low sea level) are combined to produce a simple
predictive model of likely patterns of genetic differentiation (and hence
speciation) among these mammals, and probably among other organisms, in
oceanic archipelagos.
Keywords
Biogeography, Chiroptera, differentiation, diversification, ecological traits, gene
flow, genetic variation, geology, Philippines, Rodentia.
L. R. Heaney et al.
230 Journal of Biogeography 32, 229–247, ª 2005 Blackwell Publishing Ltd

1990; Rickart et al., 1991, 1993; Heaney et al., 1998, 1999;
Steppan et al., 2003). Among these was an initial examination
of patterns of genetic differentiation in two species of fruit bats
(Peterson & Heaney, 1993); this analysis showed that Cyno-
pterus brachyotis, a species widespread in South East (SE) Asia
that occupies disturbed anthropogenic habitats, had high levels
of heterozygosity, high levels of gene flow, and low levels of
genetic differentiation. In contrast, Haplonycteris fischeri, a
Philippine endemic that occurs in primary rain forest, had low
levels of heterozygosity, low levels of gene flow, and high levels
of genetic differentiation. Further, we found that the two
species showed significantly similar geographic patterns of
genetic differentiation between populations, and that those
patterns were strongly influenced by the extent of island
shorelines during Pleistocene periods of low sea, but not by
current shorelines.
In this study, we extend those observations by increasing the
number of species (from 2 to 7) and the number of islands
(from 6 to 11). We include six fruit bat species because they are
speciose, abundant, and generally easily captured (Heideman
& Heaney, 1989). Several of these species are endemic to (but
widespread within) the archipelago, maximizing the likelihood
that general patterns could be detected. The 11 islands (Fig. 1)
represent many (although not all) of the distinct areas of
endemism in the Philippines, and a range of areas and degrees
of isolation; the number was limited by the availability of
suitably fresh frozen tissues. It should be noted that not all
species occur on all 11 of the islands, and in a few cases we
Figure 1 Philippine archipelago. Extent of
late Pleistocene landmasses (areas delimited
by present 120 m bathymetric contour) sha-
ded (redrawn from Heaney, 1986). Black dots
indicate the origin of samples used in this
study.
Geology, ecology, and genetic differentiation in Philippine mammals
Journal of Biogeography 32, 229–247, ª 2005 Blackwell Publishing Ltd 231

lacked tissues for study from reference islands where the
species is known to occur. We also include Rattus everetti, a
Philippine endemic murid rodent that is the only non-volant
small mammal that occurs widely within the archipelago;
fortunately, it is sufficiently abundant that we were able to
obtain several adequate samples. We used protein electro-
phoresis in this study, an analytical tool of great power that is
technically difficult and not often used currently, but highly
appropriate for investigation of the issues of concern to us
here. Because protein electrophoresis requires relatively large
amounts of fresh or freshly-frozen tissue, our sample sizes for
the fruit bats are sometimes small. However, we have
endeavoured to use statistical analysis cautiously to avoid
falsely detecting patterns, especially in a multi-species com-
parison, with the result that those patterns that we describe
here are likely to be robust. We believe that such information
on population genetics is crucial to developing an under-
standing of the role of geography in influencing the process of
diversification in this highly biodiverse oceanic archipelago.
As part of this study, we test two hypotheses that we proposed
in the earlier paper. The first is that late Pleistocene shorelines of
oceanic islands (i.e. the maximum extent of dry-land islands)
are as important in structuring patterns of intraspecific
variation in the Philippines as they are in delimiting regions
of interspecific diversity. Previous investigation of patterns of
species distribution, diversity, and endemism in Philippine
mammals have shown that shorelines of islands that existed
during Pleistocene periods of low sea level form the primary
boundaries between the highly distinct faunal regions in the
archipelago (25–80% endemism among non-volant mammals
and 7–22% among fruit bats; Heaney, 1986, 1991a, 1993, in
press; Heaney et al., 1998). We predict that these geographic
barriers will also manifest themselves as significant partitions of
genetic variation between island populations common to all
seven species in this study. The second hypothesis is that
ecological attributes of species determine the extent to which
common patterns are exhibited. We expect that basic know-
ledge regarding distribution and habitat preferences of these
species derived from extensive field work in the Philippines
(e.g. Heideman & Heaney, 1989; Heaney et al., 1998, 1999;
Rickart et al., 1991, 1993) will provide indications of levels of
gene flow and consequent degree of differentiation between
populations. We also address three questions unresolved in the
previous study: (1) whether small, isolated island populations
tend to show reduced within-population variability, (2) whe-
ther small, isolated island populations tend to be more
genetically differentiated, and (3) whether the tendency to
develop genetically distinct populations, which we consider to
be an intrinsic component of the process of speciation, is
correlated with, and can be predicted from, readily measured
ecological and geographic/geological parameters.
Study species and site
This study examines seven species of Philippine mammals that
fall into three general ecological and geographic patterns: (1)
three species widespread in SE Asia, all of which are primarily
associated with disturbed habitat; (2) three species that are
endemic to the Philippines but widespread within the oceanic
archipelago and are primarily associated with forest (although
with variation, as noted below); and (3) one species that is a
Philippine endemic associated with forest, but restricted to one
Pleistocene island (Heaney et al., 1989, 1999; Heideman &
Heaney, 1989; Heaney, 1991a; Rickart et al., 1991, 1993).
The three fruit bats to which we refer as ‘widespread
species’ are found throughout SE Asia and are common in
disturbed anthropogenic habitats in the Philippines. They are
C. brachyotis, a small (30–35 g) frugivore; Macroglossus
minimus, a small (15–20 g) nectarivore; and Rousettus am-
plexicaudatus, a medium-sized (70–100 g) frugivore. In the
Philippines, these three bats forage in orchards, other agricul-
tural areas, and disturbed secondary forest. They are most
common at lower elevations and are usually absent in montane
rain forest. Rousettus amplexicaudatus is most common in
clearings and orchards, and are known to regularly fly long
distances (> 20 km night)1) to forage, often across open water
(e.g. Rickart et al., 1993). Macroglossus minimus is usually
found in association with wild or domestic abaca or banana
(Musa spp.) in open secondary forest or agricultural areas, but
also feed on mangroves that grow in patches in estuaries.
Cynopterus brachyotis prefers agricultural areas or secondary
forest, and is rare in primary rain forest except on one small
island (Maripipi) which lacks Philippine endemic fruit bats
(Rickart et al., 1993).
In contrast, the second cluster of three species are endemic
to the Philippines, but occur nearly throughout the archipel-
ago. They are members of endemic genera, generally are
common in relatively undisturbed rain forest, and are variable
in their presence in heavily disturbed areas lacking good
canopy cover. Haplonycteris fischeri, a small (15–20 g) frugiv-
orous bat, is the most habitat-restricted mammal in this
category; it is common beneath the canopy in primary rain
forest, scarce in secondary forest, and absent in open
agricultural areas. It is often the most abundant fruit bat in
mature forest at middle elevations. Ptenochirus jagori, a
medium-sized (70–90 g) frugivorous bat, is known to move
farther than H. fischeri (Heideman & Heaney, 1989) and
prefers primary rain forest but can maintain populations even
in degraded secondary rain forest. Rattus everetti, a large
(230–420 g) endemic Philippine rodent and the only non-
volant mammal in this study, prefers disturbed forest and
tolerates primary forest, but is absent away from forest
(Rickart et al., 1993; Heaney et al., 1999).
The third group is represented by one species of small
(35–40 g) frugivorous bat, P. minor. It is restricted to a single
Pleistocene island (Greater Mindanao; see below), is common
in primary or good secondary lowland rain forest, tolerates
second-growth, and is scarce outside of forest.
The Philippine archipelago is an especially interesting arena
for investigating biological diversification (Heaney, 1986,
1991a,b; Mitchell et al., 1986; Packham, 1996; Hall, 1998,
2002; Steppan et al., 2003). In brief, the first of the extant
L. R. Heaney et al.
232 Journal of Biogeography 32, 229–247, ª 2005 Blackwell Publishing Ltd

islands appeared as dry land during the early Oligocene, but
most of the extant islands have originated since the late
Miocene, and many during the Pliocene. One portion of the
archipelago, Palawan and associated small islands, probably
was joined to the Asian mainland during the middle or early
Pleistocene, and is part of the Sunda Shelf biogeographic
region, rather than the Philippine region (Esselstyn et al., in
press). The rest of the archipelago has never had a dry-land
connection to SE Asia, having arisen as a set of de novo islands
from the ocean floor, most often far to the SE of its current
location, as a result of tectonic and volcanic activity, gradual
coalescence, and variable but progressive uplift. Most current
topography is probably close to its maximum level. Since the
archipelago sits on an uplifted platform bracketed by deep
trenches, the depths between some islands are relatively
shallow. Thus, as Pleistocene sea level rose and fell in concert
with the development of continental ice sheets, certain groups
of islands have experienced repeated cycles of coalescence and
fragmentation. During the most recent period of low sea level
in the Pleistocene, c. 18,000 years ago (Fairbanks, 1989; Siddall
et al., 2003), many current islands merged into four large
islands: Greater Luzon, Greater Mindanao, Greater Negros-
Panay, and Greater Palawan (Fig. 1). The present configur-
ation of islands represents a phase of fragmentation due to
high sea level. Each of the ‘Pleistocene islands’ shown in Fig. 1
originated as a de novo oceanic island between c. 25 and
0.5 Ma, and (with the exception of Palawan) has had no dry-
land connection to other islands or continents. We call them
‘Pleistocene islands’ because they reached their largest size
during the late Pleistocene periods of low sea level, not because
they originated during the Pleistocene.
Eleven present-day islands from seven Pleistocene islands
are represented in this study (Fig. 1). During the late
Pleistocene period of low sea level, Luzon, Catanduanes, and
Polillo coalesced into Greater Luzon; Leyte and Biliran
coalesced with modern Mindanao into Greater Mindanao;
and Fuga and Barit also coalesced into a single landmass.
Negros was part of Greater Negros-Panay, while Mindoro,
Sibuyan, and Dalupiri each stood alone and have remained
unconnected to other islands. Each major Pleistocene island
has been documented as a centre of endemism. Greater Luzon
and Greater Mindanao, for example, have 70–80% endemism,
and Mindoro, Greater Negros-Panay, and Sibuyan each have
40–50% endemism, among native non-volant mammal species
(Heaney, 1993, in press).
MATERIALS AND METHODS
Sampling was conducted in and near large tracts of forest
(relative to the size of the islands); on all islands, most
deforestation near our sites dates from the last 10–30 years.
Thus, our estimates of genetic variation should not reflect the
effects of habitat destruction. Bats were collected in mist nets
and euthanized with lethal doses of sodium pentobarbital. Rats
were collected in Victor snap traps. Tissues were harvested
immediately and frozen in liquid nitrogen, and later stored at
the Field Museum in an ultracold freezer at )80 �C. Voucherspecimens were prepared and deposited at the Field Museum,
the Philippine National Museum, and the United States
National Museum of Natural History.
Protein electrophoresis protocols for C. brachyotis and
H. fischeri are described in Peterson & Heaney (1993) and all
data included here for these species are from that study. For
the remaining species, equal portions of heart, liver, and
skeletal muscle tissue were homogenized in a 1 mM disodium
EDTA/100 mM Trizma base/0.2 mM NAD, NADP, and ATP
buffer, centrifuged for 45 min at 12,000 rpm, and supernatants
drawn into capillary tubes for storage. Samples were electro-
phoresed for 4–6 h on 12% starch gels, depending on the
specific analysis desired. Gels were sliced horizontally, and each
slice stained using specific protein assays from Shaw & Prasad
(1970) and Harris & Hopkinson (1978). Each sample was
scored at 32 presumptive genetic loci (enzyme commission
numbers from International Union of Biochemistry and
Molecular Biology, 1992): AAT (2.6.1.1, 2 loci), ACN
(4.2.1.3, 2 loci), ACP (3.1.3.2), ADH (1.1.1.1, 3 loci), AK
(2.7.4.3, 2 loci), ATA (2.6.1.2), CK (2.7.3.2, 2 loci), EST
(3.1.1.1, 2 loci), G3PDH (1.1.1.8), G6PDH (1.1.1.49), GDA
(3.5.4.3), GPI (5.3.1.9), ICD (1.1.1.42, 2 loci), LDH (1.1.1.27, 2
loci), MDH (1.1.1.37, 2 loci), MPI (5.3.1.8), NP (2.4.2.1), PEP
(3.4.11; 5 loci, corresponding to PEP-A, -B, -C, -D, -S), PGD
(1.1.1.44), PGM (5.4.2.2), PK (2.7.1.40), and SOD (1.15.1.1).
For each species, all individuals were analysed on the same gel.
To assure correct assignment of homologies, reference indi-
viduals were included at multiple points on each gel.
Analyses were performed in BIOSYS-1 (Swofford &
Selander, 1981). Allele frequencies and three measures of
within-population variation – mean observed heterozygosity
(Hobs), number of alleles per locus (NALL), and percentage of
loci polymorphic (POLY; 5% criterion) – were calculated.
Departure from Hardy–Weinberg equilibrium was tested by
3 methods (the chi-square goodness-of-fit test, and an exact
probability test (Haldane, 1954); because our samples were
sometimes small, we also used a chi-square test with Levene
(1949) correction for small sample sizes) (Table 1; Appendix
S1 in Supplementary Material). Tests of association between
measures of within-population variation (Hobs, NALL, POLY)
with island area and isolation were performed with linear
regression and non-parametric Spearman rank correlation.
Tests for differences between species in levels of within-
population variation were performed with analysis of variance
and non-parametric Kruskal–Wallis tests.
Fixation indices (F-statistics; Wright, 1951, 1965) were used
to summarize the distribution of genetic variation within and
between populations. Confidence limits on estimates of FSTwere established by jack-knifing over loci as recommended by
Weir & Cockerham (1984); we employed a relatively conser-
vative experimentwise error rate (a ¼ 0.001). Hierarchical
fixation indices (Wright, 1978) and variance components
(Cockerham, 1969, 1973; Weir, 1990) were calculated based on
the Pleistocene connections among islands as follows: (Luzon,
Catanduanes, Polillo) (Leyte, Biliran) (Negros) (Mindoro)
Geology, ecology, and genetic differentiation in Philippine mammals
Journal of Biogeography 32, 229–247, ª 2005 Blackwell Publishing Ltd 233

(Sibuyan) (Dalupiri) (Fuga, Barit). Estimates of overall gene
flow between populations (Nm) were derived from the
approximation FST ¼ 1/(1 + 4Nm) as recommended by Slat-
kin & Barton (1989), and also computed by the private alleles
method of Slatkin (1985) using the correction for sample size
of Slatkin (1985) as clarified in Slatkin & Barton (1989). Both
methods provide reasonable estimates of Nm; Slatkin & Barton
(1989) found that these two methods of estimating Nm
bracketed the true values of Nm in some simulations. Gene
flow among pairs of populations (M; Slatkin, 1993) was
calculated by the GST (Nei, 1973) and theta (1; Weir &
Cockerham, 1984) methods, using a program by Slatkin
(1993). We present the results for M as calculated by the GST
method (Nei, 1973, 1977) because estimates of gene flow most
commonly reported in the literature are calculated by this
method (e.g. it is the method used by BIOSYS-1; Swofford &
Selander, 1981). M between Pleistocene islands is simply the
mean of all estimates along that track.
Cavalli-Sforza & Edwards’ (1967) arc genetic distance (as
preferred by Wright, 1978) and Nei’s (1978) unbiased genetic
distance were calculated (Appendix S2). Geographic distances
were measured as nearest shore-to-shore distances from
bathymetric charts (Department of Defense, Defense Mapping
Agency charts, Part 2 – Hydrographic Products, Region 9 –
East Asia) at the smallest practical scales; distances between
Pleistocene shorelines were estimated using the 120 m bath-
ymetric contour from the same charts (Appendix S3). To test
the correspondence of genetic and geographic distances, we
used permutation-based matrix correlation tests (Mantel,
1967; Dietz, 1983). These tested the proportionality of
Cavalli-Sforza and Edwards’ arc genetic distance matrices
and geographic distance matrices (between both present-day
and inferred Pleistocene shorelines), using a FORTRAN
program (MATCORR.EXE) by Dietz (1983). We present the
results of the Spearman rank correlations for matrices rather
than the more commonly used Mantel test (which uses
Pearson product-moment correlation; Mantel, 1967) because
the Spearman test is less sensitive to the actual distance
measure used (Dietz, 1983) and it is more appropriate when
there is less certainty about the reliability of close ranks (Sokal
& Rohlf, 1981).
The ‘small oceanic islands’ in this study (Sibuyan, Barit,
Dalupiri, and Fuga) did not coalesce with large Pleistocene
islands at the last glacial maximum. Each originated as a de
novo oceanic island and has remained continuously isolated, as
described above. Peterson & Heaney (1993) found that
populations of Cynopterus and Haplonycteris on these small,
isolated oceanic islands showed weakly reduced within-
population variability, and predicted that reduced variability
would be found in other species. To test this hypothesis, we
compared Hobs, NALL, and POLY for each species between
large Pleistocene island populations and small oceanic island
populations using analysis of variance and the non-parametric
Kruskal–Wallis test. Peterson & Heaney (1993) also found that
populations on the small, isolated oceanic islands were
consistently distinct from other islands in the Philippines,
and predicted that this pattern would be evident in other
species. To test this hypothesis, we examined genetic distance
and M. All tracks leading to small oceanic islands were
distinguished from all tracks that did not include an oceanic
island (i.e. tracks within and between large Pleistocene islands).
For each species, small oceanic island tracks were compared
with large Pleistocene island tracks by the Kruskal–Wallis test,
which utilizes a conservative number of degrees of freedom.
Throughout our analyses, there are instances in which a
given hypothesis is tested across several species. In such cases,
Table 1 Genetic variation within populations (Hobs, observed heterozygosity) and island areas
Island Area (km2)
Widespread Endemic Narrow endemic
Rousettus
amplexicaudatus
Macroglossus
minimus
Cynopterus
brachyotis
Ptenochirus
jagori
Haplonycteris
fischeri
Rattus
everetti
Ptenochirus
minor
Luzon 104,688 0.145 0.058 0.065 0.022 0.054 0.025 –
Negros 12,704 0.111 0.051 0.083 – 0.056 – –
Mindoro 9736 – 0.050 – – – – –
Leyte 7213 0.115 0.042 0.060 0.032 0.000 0.048 0.027
Catanduanes 1430 0.118 0.025 0.071 0.024 0.036 0.017 –
Polillo 606 0.091 – – – – – –
Biliran 497 0.090 0.033 0.097 0.022 0.036 0.025 0.028
Sibuyan 448 0.103 0.063 0.040 0.034 0.022 – –
Fuga 93 0.090 – – – – – –
Dalupiri 62 0.075 – – – – – –
Barit 5 0.103 – – – – – –
Mean Hobs 0.104 0.046 0.069 0.027 0.034 0.029 0.028
Mean NALL 1.36 1.20 1.45 1.20 1.20 1.20 1.15
Mean POLY 25.5 14.3 26.2 10.3 19.0 11.5 12.0
Summary lines include mean Hobs, mean NALL (number of alleles per population), and mean POLY (proportion of loci polymorphic per popu-
lation).
L. R. Heaney et al.
234 Journal of Biogeography 32, 229–247, ª 2005 Blackwell Publishing Ltd

it is possible that no individual test of the hypothesis will prove
statistically significant although a significant overall effect
exists. The growing field of meta-analysis (e.g. Gurevitch et al.,
1992) provides many methods for combining results from
different studies. We restrict ourselves to Fisher’s relatively
simple method of combined probabilities (Fisher, 1954; Sokal
& Rohlf, 1981) for testing for overall significance. In our
application of the method, each species is treated as an
independent test of the hypothesis under consideration, e.g.
that Hobs is related to log(area). The sum across species of the
natural logarithms of the P-values for each such regression or
rank correlation is multiplied by )2. This value is distributed
as a chi-square with 2 k degrees of freedom, where k is the
number of separate tests and probabilities (in this case, the
number of species).
Relationships between populations were explored through
phenetic and phylogenetic methods using the genetic distance
measures noted above. UPGMA dendrograms (Sneath & Sokal,
1973) and Distance Wagner trees (Farris, 1972) were generated
in BIOSYS-1 (Swofford & Selander, 1981). Fitch (Fitch &
Margoliash, 1967) trees were generated in PHYLIP (Felsen-
stein, 1989). A maximum parsimony analysis of allele
frequencies was conducted using the FREQPARS program of
Swofford & Berlocher (1987). All of these methods yield
similar topological results, and so the UPGMA dendrograms
using Cavalli-Sforza and Edwards’ arc genetic distance are
presented by convention (Fig. 3).
RESULTS
Departures from Hardy–Weinberg equilibrium
The largest number of departures from Hardy–Weinberg
equilibrium was detected by the Levene (1949) correction for
small sample sizes to the chi-square test. This method detected
17 departures from Hardy–Weinberg equilibrium, of 225
possible polymorphic loci-populations. Many fewer were
detected by the chi-square test for goodness-of-fit and the
exact probability test. All departures were heterozygote defi-
ciencies of small degree and were concentrated in a few loci
[e.g. G6PDH (5), NP (3), and ICD (2)]. Removal of these loci
does not qualitatively affect any of the conclusions presented
here.
Population dendrograms
Two consistent trends are apparent in the UPGMA population
dendrograms (Fig. 3). First, the small oceanic island popula-
tions (Sibuyan, Dalupiri, Fuga, and Barit) are often the most
strongly differentiated from other populations. Sibuyan, in
particular, appears most genetically distinct in three of the five
taxa that are found on that island. The other trend that
emerges is that Biliran and Leyte, the pair of islands with the
shallowest ocean depth between them (< 10 m), are the least
differentiated in two of six taxa and are not well differentiated
in a third. The presumed Pleistocene hierarchy of island
relationships, however, emerges clearly only in the case of R.
everetti, the only non-volant mammal in this study. It should
be noted that cluster analysis of island populations based on
genetic distances constitutes an exploratory technique and is
not appropriate for testing the hypothesis that Pleistocene
shorelines form significant partitions of genetic variation
(Sneath & Sokal, 1973).
Variation within populations: geological correlates
Genetic variability within populations is theoretically related to
effective population size (Wright, 1931). Smaller populations
are expected to lose genetic variation via genetic drift more
rapidly than larger populations, thus achieving a lower
standing level of variation due to mutation-drift balance. This
expectation has received much attention in the conservation
biology literature (e.g. Soule 1976, 1987; Frankham, 1995,
1996; Lande, 1995; Berry, 1998), yet how often this phenom-
enon is important in wild populations is not clear. To test
whether this theoretical relationship is present in these
Philippine mammals, we used island area, which varies over
six orders of magnitude in this study, and the logarithm of
island area, as reasonable proxies for effective population size
(P. minor, represented by only two populations, was not
considered in this analysis). In only one of the six species was a
significant regression of within-population variation and
island area, detected (similar results were obtained for all
three measures of within-population variation; only the results
for Hobs are reported). Rousettus amplexicaudatus, a wide-
spread SE Asian fruit bat, displayed significant regressions of
Hobs vs. island area and the logarithm of island area
{Hobs ¼ 0.098 + 0.000(area), r ¼ 0.775, P ¼ 0.008;
Hobs ¼ 0.071 + 0.005[log (area)], r ¼ 0.735, P ¼ 0.015}.
Ptenochirus jagori displayed a significant regression of the
number of alleles on the logarithm of island area
{NALL ¼ 0.772 + 0.138[log(area)], r ¼ 0.963, P ¼ 0.008},
but NALL is a highly sample-size-dependent measure and
there was also a significant regression of sample size (N) on
island area for this species {N ¼ )14.883 + 7.964[log(area)],
r ¼ 0.938, P ¼ 0.019}, so we exercise caution in interpreting
this result. Fisher’s method of combined probabilities for all
species combined across measures did not yield any significant
overall results for relationships of Hobs, NALL, and POLY with
area or log(area). Non-parametric methods, including Spear-
man rank correlation (Sokal & Rohlf, 1981), did not show
more significant associations of within-population variability
measures and area than would be expected by chance for all
taxa other than R. amplexicaudatus, nor were any significant
overall trends detected using combined probabilities from
non-parametric tests.
We also considered the possibility that isolation of island
populations may have an effect on within-population varia-
bility through lowered frequency of migration. Schmitt et al.
(1995) showed that heterozygosity within island populations of
the fruit bat C. nusatenggara in Indonesia was correlated with
distance to the nearest large source population, and Hisheh
Geology, ecology, and genetic differentiation in Philippine mammals
Journal of Biogeography 32, 229–247, ª 2005 Blackwell Publishing Ltd 235

et al. (1998) found similar results for Eonycteris spelaea in the
same area. The analogy with patterns of interspecific diversity
is clear: more isolated islands are predicted to receive fewer
colonists and tend to have fewer species than islands closer to
source pools of colonists (MacArthur & Wilson, 1967).
Similarly, one might expect that more isolated islands would
receive fewer migrants to augment their genetic diversity.
Various indices of isolation (distance to the nearest island,
distance to the nearest large island, distance to the nearest
Pleistocene island, and distance to the nearest large Pleistocene
island) were used in our study. No more significant results
than expected by chance were obtained for regressions of
measures of within-population variability on isolation indices,
for partial regressions of within-population variability on area
[and log(area)] plus isolation indices, nor for overall trends
across species from combined probabilities. Hence, our data
indicate that neither island area nor isolation of island
populations from sources of migrants have a substantial
overall effect on standing genetic variation within populations
in this system, in spite of variation in island area over six
orders of magnitude.
Variation within populations: ecological correlates
We have documented that the widespread SE Asian species
that occur in the Philippines usually occupy disturbed habitats,
while the Philippine endemic species prefer primary rain forest
(although some can maintain populations in disturbed forest;
e.g. Heideman & Heaney, 1989; Heaney et al., 1998, 1999;
Rickart et al., 1991, 1993). Our previous study of C. brachyotis
and H. fischeri found that the widespread C. brachyotis, which
prefers disturbed, open habitat, showed high levels of variation
within populations, and the endemic species (H. fischeri),
which preferred closed-canopy primary forest, showed low
levels of variation within populations (Peterson & Heaney,
1993). We postulated that this trend constituted a general
pattern. To test this proposition, we performed nested analyses
of variance on Hobs, NALL, and POLY in our seven species
(Table 1), where the levels are ecological category (widespread/
habitat tolerant species vs. endemic/forest-associated species
plus single-Pleistocene-island endemics), species within eco-
logical category, and island populations as the replicates within
species. For all three measures, both levels were significant at
P < 0.01. This result indicates that, while significant hetero-
geneity exists within ecological categories, the widespread SE
Asian species, which prefer disturbed habitat, display sig-
nificantly higher levels of within-population variability than
species endemic to the Philippines, which prefer primary
forest, as predicted.
Since heterozygosity and other measures of within-popula-
tion variation usually do not meet the assumptions of
parametric statistics (they are skewed positively in this case),
we also performed non-parametric statistical tests. Because no
appropriate non-parametric analogue for nested anova is
available, we took two approaches. First, we performed a
Kruskal–Wallis test using each island population of each
species as an independent observation and tested whether or
not widespread SE Asian species had greater within-population
variability than endemic species and the single-island Pleisto-
cene endemic. For all three measures, all tests are significant
(P < 0.01; Table 2). Second, a more conservative approach was
applied, treating each species as an independent observation.
Species means of the three measures of genetic variation were
used to test the hypothesis that widespread species (which
prefer disturbed habitat) have higher levels of within-popula-
tion genetic variation than endemic species (which prefer
primary forest; Table 2, with mean values in Table 1). Hobs was
significantly higher in widespread species (P < 0.05), and
NALL and POLY were nearly so (0.10 < P < 0.05), again
supporting the prediction.
Variation between populations: geological correlates
Wright (1943) provided the theoretical underpinnings for a
simple notion, that populations closer to one another should
be more similar to one another than populations farther apart
due to the homogenizing effects of gene flow. The quantitative
theory of ‘isolation by distance’ is mathematically complex and
can be difficult to test in its particulars, but numerous authors,
notably R. Sokal and colleagues (Sokal & Wartenberg, 1983;
Sokal, 1988; Livshits et al., 1991), have relied on non-
parametric, permutation-based, matrix correspondence tech-
niques to test for the presence of an isolation by distance
pattern. We used these methods to test statistically the
proportionality of matrices of genetic distances and geographic
distances between populations. We compared matrices of
genetic distance (results for Cavalli-Sforza and Edwards’ arc
genetic distance are presented in Table 3; other genetic
distance measures yielded similar results) with matrices of
geographic distance between present-day islands. The tests for
all species yield a similar and rather surprising result: matrices
of genetic distances between island populations are not
correlated with the matrix of geographic distances between
present-day islands in any of the six species, nor is an overall
Table 2 Comparisons of genetic variation within populations
between widespread SE Asian species and Philippine endemics
Hobs NALL POLY n
(a)
Widespread SE Asian 641 573 570 23
Philippine endemic 179 247 250 17
P-value 0.001 0.007 0.004
(b)
Widespread SE Asian 18 16.5 17 3
Philippine endemic 10 11.5 11 4
P-value 0.034 0.079 0.077
Sum of ranks and significance values from Kruskal–Wallis test on
observed heterozygosity (Hobs), number of alleles per locus (NALL),
and proportion of loci polymorphic (POLY), using (a) islands as
observations, and (b) species as observations.
L. R. Heaney et al.
236 Journal of Biogeography 32, 229–247, ª 2005 Blackwell Publishing Ltd
significant correlation detected using combined probabilities
(Table 3).
However, another relevant pattern requires testing. Patterns
of mammalian interspecific diversity in the Philippines are
strongly influenced by the boundaries of islands that existed
during maximal late Pleistocene sea-level lowering, which
represent maximum coalescence during the history of the
archipelago (Heaney, 1986, 1991a, 1993, in press; Steppan
et al., 2003). This pattern of interspecific diversity suggested
another test: we calculated matrix correspondence between
genetic distance matrices and the matrix of geographic
distances between Pleistocene shorelines, inferred from the
120 m bathymetric contour. Spearman rank correlation
yielded significant results for two of six taxa (M. minimus
and P. jagori) and nearly significant results (0.10 > P > 0.05)
for three additional taxa (R. amplexicaudatus, C. brachyotis,
and H. fischeri) for these Pleistocene distances (Table 3). Using
combined probabilities across species, we conclude that genetic
distances are not correlated with distances between present-day
shorelines, but are correlated with distances between late
Pleistocene shorelines. It appears, therefore, that the geological
history (including both long-term isolation between some
islands and Pleistocene coalescence among others) has created
a common pattern of geographic variation between popula-
tions across the six species, partitioned by late Pleistocene
shorelines. Specifically, the geographic distance between per-
manently isolated geo-historical units is positively correlated
with increasing genetic distance.
Variation between populations: ecological correlates
A final prediction about the extent of the common pattern of
differentiation between populations will be addressed here. We
believe that the presence of the widespread species on small,
isolated islands where no endemic species are found (e.g. Barit,
Dalupiri, and Fuga) and the observation that widespread
species will readily cross clearings but endemic species are
rarely found far from primary rain forest or out from beneath
good canopy cover (except P. jagori, as noted below), should
result in lower levels of gene flow between populations of the
endemic species on different islands. If this is correct,
population genetic theory predicts that endemic species will
display higher levels of between-population differentiation
than widespread species. To test this prediction, we examined
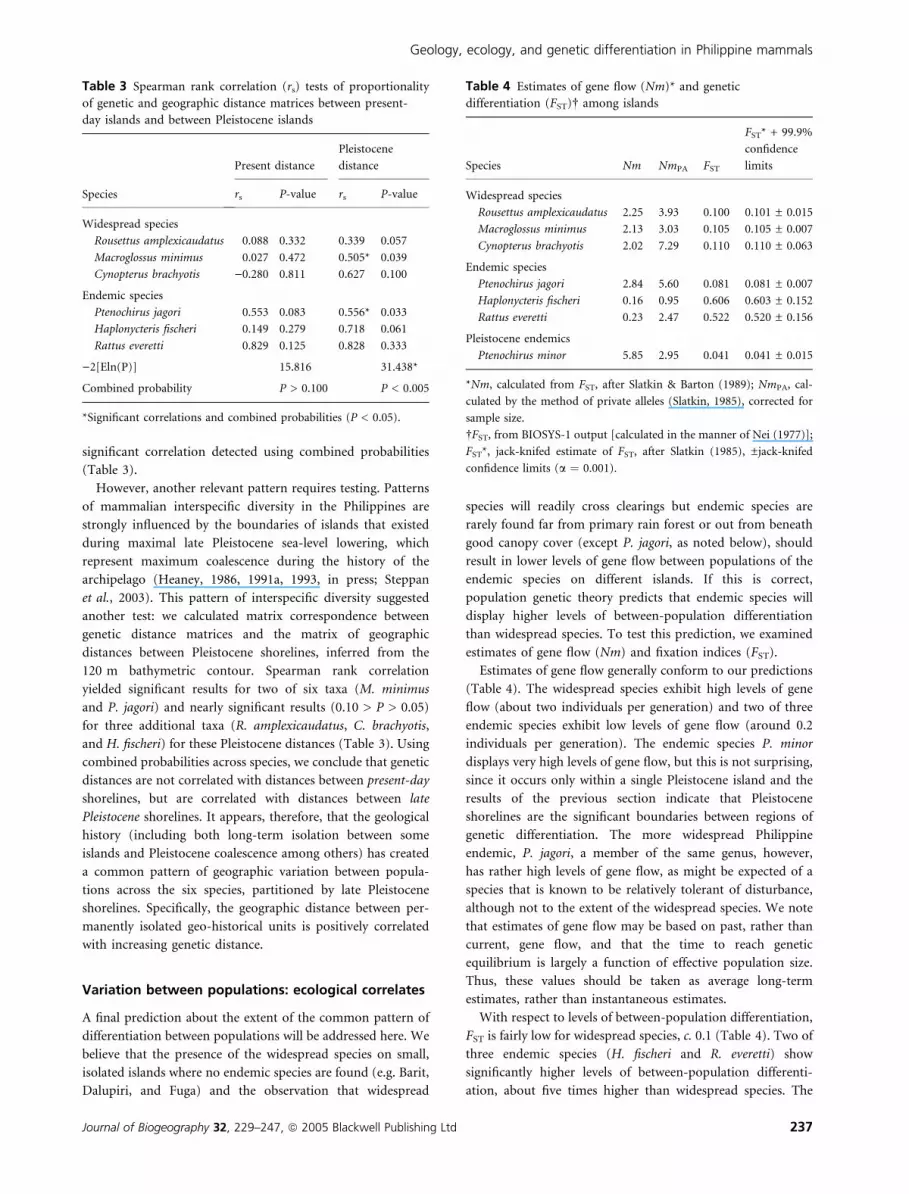
estimates of gene flow (Nm) and fixation indices (FST).
Estimates of gene flow generally conform to our predictions
(Table 4). The widespread species exhibit high levels of gene
flow (about two individuals per generation) and two of three
endemic species exhibit low levels of gene flow (around 0.2
individuals per generation). The endemic species P. minor
displays very high levels of gene flow, but this is not surprising,
since it occurs only within a single Pleistocene island and the
results of the previous section indicate that Pleistocene
shorelines are the significant boundaries between regions of
genetic differentiation. The more widespread Philippine
endemic, P. jagori, a member of the same genus, however,
has rather high levels of gene flow, as might be expected of a
species that is known to be relatively tolerant of disturbance,
although not to the extent of the widespread species. We note
that estimates of gene flow may be based on past, rather than
current, gene flow, and that the time to reach genetic
equilibrium is largely a function of effective population size.
Thus, these values should be taken as average long-term
estimates, rather than instantaneous estimates.
With respect to levels of between-population differentiation,
FST is fairly low for widespread species, c. 0.1 (Table 4). Two of
three endemic species (H. fischeri and R. everetti) show
significantly higher levels of between-population differenti-
ation, about five times higher than widespread species. The
Table 3 Spearman rank correlation (rs) tests of proportionality
of genetic and geographic distance matrices between present-
day islands and between Pleistocene islands
Species
Present distance
Pleistocene
distance
rs P-value rs P-value
Widespread species
Rousettus amplexicaudatus 0.088 0.332 0.339 0.057
Macroglossus minimus 0.027 0.472 0.505* 0.039
Cynopterus brachyotis )0.280 0.811 0.627 0.100
Endemic species
Ptenochirus jagori 0.553 0.083 0.556* 0.033
Haplonycteris fischeri 0.149 0.279 0.718 0.061
Rattus everetti 0.829 0.125 0.828 0.333
)2[Eln(P)] 15.816 31.438*
Combined probability P > 0.100 P < 0.005
*Significant correlations and combined probabilities (P < 0.05).
Table 4 Estimates of gene flow (Nm)* and genetic
differentiation (FST)� among islands
Species Nm NmPA FST
FST* + 99.9%
confidence
limits
Widespread species
Rousettus amplexicaudatus 2.25 3.93 0.100 0.101 ± 0.015
Macroglossus minimus 2.13 3.03 0.105 0.105 ± 0.007
Cynopterus brachyotis 2.02 7.29 0.110 0.110 ± 0.063
Endemic species
Ptenochirus jagori 2.84 5.60 0.081 0.081 ± 0.007
Haplonycteris fischeri 0.16 0.95 0.606 0.603 ± 0.152
Rattus everetti 0.23 2.47 0.522 0.520 ± 0.156
Pleistocene endemics
Ptenochirus minor 5.85 2.95 0.041 0.041 ± 0.015
*Nm, calculated from FST, after Slatkin & Barton (1989); NmPA, cal-
culated by the method of private alleles (Slatkin, 1985), corrected for
sample size.
�FST, from BIOSYS-1 output [calculated in the manner of Nei (1977)];
FST*, jack-knifed estimate of FST, after Slatkin (1985), ±jack-knifed
confidence limits (a ¼ 0.001).
Geology, ecology, and genetic differentiation in Philippine mammals
Journal of Biogeography 32, 229–247, ª 2005 Blackwell Publishing Ltd 237

geographically restricted P. minor displays very low levels of
differentiation within its single Pleistocene island (although we
have few sampling sites), a trend that we discuss below. The
more widespread endemic species, P. jagori, presents an
illustrative exception to the other endemic species. It displays
low levels of within-population variation, but unlike the other
endemic species, its habitat tolerances are moderately broad
(i.e. it often forages in open habitats and readily maintains
populations in secondary forest), and it displays high levels of
overall gene flow. We interpret this result as evidence that it is
the not the categorization of a species as an endemic per se that
is usually associated with low gene flow, but rather with the
usual (but not ubiquitous) tendency of endemic species to
have low tolerance for disturbed, open habitat, since P. jagori
apparently has the broadest habitat tolerance of any endemic
Philippine fruit bat (although less than the widespread SE
Asian species). Morphological data (Walsh, 1998) show a
similar but less pronounced trend, in that levels of within-
population variation in P. jagori are similar to those of other
endemic species, while levels of between-population variation
resemble those of widespread species.
Wright’s (1978) hierarchical F-statistics (Table 5) show how
overall variation between populations may be apportioned into
variation between Pleistocene islands and variation between
present-day islands within Pleistocene islands. Two of three
Philippine endemics show the overwhelming proportion of
their between-population variation structured by the bound-
aries of Pleistocene islands (H. fischeri, 92%; R. everetti, 97%).
They exhibit almost no variation between present-day islands
within Pleistocene islands. This lack of differentiation between
populations within Pleistocene islands is consistent with the
observation of low FST in P. minor, which occurs only on a
single Pleistocene island. The third endemic species, P. jagori,
also displays over half of its variation between populations at
the between-Pleistocene-island level (59%), consistent with its
moderately broad habitat preferences. Two of three widespread
species also display substantial proportions of their between-
population genetic variation at the between-Pleistocene-island
level (C. brachyotis, 79%; M. minimus, 34%). In contrast,
R. amplexicaudatus shows relatively little of its between-
population genetic variation structured according to the
boundaries of Pleistocene islands (7%).
Overall patterns
Two less commonly employed modes of analysis will be
discussed here to help further illuminate the pattern and extent
of genetic differentiation in these Philippine mammals.
Cockerham’s (1973) extension of Wright’s method of variance
components is used to show the complete breakdown of total
genetic variation (Table 6), including variation within present
island populations which Wright’s hierarchical F-statistics do
not illustrate. Additionally, Slatkin’s (1993) M-statistics are
used to estimate gene flow between all pairs of islands (Fig. 2).
Some clear trends emerge. First, almost no differentiation is
found between present-day island populations within Pleisto-
cene islands for these species (Table 6). Second, there is an
overall pattern of genetic differentiation between Pleistocene
island groups. Two of three endemic species exhibit substantial
proportions of their total genetic variation at the between-
Pleistocene-island level (H. fischeri and R. everetti). Most of the
widespread species also show more variation between Pleisto-
cene islands than between present-day islands within Pleisto-
cene islands, as does the endemic species, P. jagori. Only
R. amplexicaudatus appears to display a pattern of variation in
which Pleistocene shorelines do not form the primary parti-
tions of genetic variation.
Several further generalizations about patterns of gene flow
between populations as inferred from allozyme data can be
discovered by estimating gene flow (Nm) between all pairs of
populations using M-statistics (Slatkin, 1993; Fig. 2). The first
general pattern that emerges is that levels of gene flow are
higher between Leyte and Biliran than between Luzon and
Catanduanes for most species, including all endemic species.
This pattern of genetic similarity is consistent with two factors:
(1) Leyte and Biliran are presently closer to one another than
are Luzon and Catanduanes and may experience higher rates of
present-day gene flow; and (2) Leyte and Biliran are separated
by a much shallower ocean channel than are Luzon and
Catanduanes (Heaney, 1986) and genetic similarity may reflect
the more recent separation of Leyte and Biliran. The second
general pattern that emerges is that gene flow to Sibuyan
appears almost uniformly attenuated (except in M. minimus).
Table 5 Wright’s (1978) hierarchical F-statistics illustrating
genetic differentiation among Pleistocene islands vs. present-day
island populations
Species Level FXY
Variance
component
Percentage
variance
Widespread species
Rousettus amplexicaudatus GPI 0.003 0.0113 7
ISL 0.045 0.1449 93
Total 0.048 0.1562 100
Macroglossus minimus GPI 0.021 0.0280 34
ISL 0.040 0.0536 66
Total 0.061 0.0816 100
Cynopterus brachyotis GPI 0.070 0.0855 79
ISL 0.021 0.0226 21
Total 0.088 0.1081 100
Endemic species
Ptenochirus jagori GPI 0.029 0.0223 59
ISL 0.020 0.0152 41
Total 0.048 0.0375 100
Haplonycteris fischeri GPI 0.537 0.7818 92
ISL 0.100 0.0675 8
Total 0.583 0.8493 100
Rattus everetti GPI 0.492 0.8279 97
ISL 0.030 0.0253 3
Total 0.507 0.8532 100
GPI, among Pleistocene islands; ISL, among present-day islands within
Pleistocene islands.
L. R. Heaney et al.
238 Journal of Biogeography 32, 229–247, ª 2005 Blackwell Publishing Ltd

This could be the result of smaller or more isolated islands
being more difficult targets for migrants to hit, although the
decrease in levels of migration does not appear to be sufficient
to influence levels of within-population variability, as noted
below. Finally, endemic species display lower levels of gene
flow between Pleistocene islands than within Pleistocene
Table 6 Cockerham’s (1973) method of complete variance partitioning. Each level is expressed as a percentage of the total variance
Widespread Endemic Narrow endemic
Rousettus
amplexi-caudatus (16/29)
Macroglossus
minimus (15/30)
Cynopterus
brachyotis (9/14)
Ptenochirus
jagori (15/29)
Haplonycteris
fischeri (10/14)
Rattus
everetti (10/26)
Ptenochirus
minor (7/29)
Variable loci only
GPI 2.4 3.6 6.7 3.8 30.7 23.9 –
ISL 4.8 3.4 2.6 1.4 0.8 1.3 3.5
W/I 92.8 93.0 90.7 94.8 68.5 74.8 96.5
Total 100.0 100.0 100.0 100.0 100.0 100.0 100.0
All loci (monomorphic loci calculated as 100% at lowest level)
GPI 1.4 1.8 4.3 1.9 21.9 9.2 –
ISL 2.6 1.7 1.7 0.7 0.6 0.5 0.8
W/I 96.0 96.5 94.0 97.4 77.5 90.3 99.2
Total 100.0 100.0 100.0 100.0 100.0 100.0 100.0
Results are tabled separately for all loci and for variable loci only. Numbers in parentheses indicate the number of variable loci over the total number
of loci for each taxon. Negative variance components are treated as equal to zero.
GPI, between Pleistocene islands; ISL, between present-day islands within Pleistocene islands; W/I, within present-day islands.
Figure 2 Estimates of gene flow between
selected populations [M after Slatkin (1993)].
Estimates of gene flow between Pleistocene
islands are averages of all estimates along that
track. Abbreviations are: BIL, Biliran; CAT,
Catanduanes; LEY, Leyte; LUZ, Luzon; NEG,
Negros; SIB, Sibuyan. Philippine endemic
species are on the right, non-endemics are on
the left.
Geology, ecology, and genetic differentiation in Philippine mammals
Journal of Biogeography 32, 229–247, ª 2005 Blackwell Publishing Ltd 239

islands, with one exception (H. fischeri: Greater Luzon/Greater
Negros-Panay). Widespread species show no clear differences
in estimates of gene flow within and between Pleistocene
shorelines. This result indicates that the permanent oceanic
barriers between Pleistocene islands are of greater conse-
quence, especially for endemic species, than recent seawater
barriers within Pleistocene islands which have been inundated
by sea-level rise during the past 18,000 years. This is true in
spite of the fact that some of the more recently formed water
gaps within Pleistocene islands are larger than distances
between Pleistocene islands (Appendix S3).
These overall patterns show the influences of both geological
and ecological processes: the Pleistocene history of the
Philippine archipelago has created a common pattern of
significant differentiation between, but not within, Pleistocene
islands. Further, widespread species associated with disturbed
habitats show less variation between and more variation within
populations than species endemic to the Philippines associated
with primary rain forest.
Differentiation of small oceanic island populations
In general, the populations on the small, isolated oceanic
islands in this study (Sibuyan, Barit, Dalupiri, and Fuga)
displayed only weakly reduced within-population variability as
might be expected due to their small size and isolation. Hobs,
NALL, and POLY were significantly lower in oceanic island
populations only for R. amplexicaudatus, and combining
probabilities across the five relevant species did not detect a
significant overall result. There was, however, a consistent
trend for small oceanic island populations to be more strongly
genetically differentiated than large Pleistocene island popula-
tions. Cavalli-Sforza and Edwards arc genetic distance and M
were significantly higher and lower, respectively (both
P < 0.01), along tracks leading to small oceanic islands than
along other island tracks in three of five species (C. brachyotis,
H. fischeri, and R. amplexicaudatus) and nearly so in one
species (P. jagori; both P � 0.10). Combining probabilities
yields a significant overall result for both measures (P < 0.001)
and we conclude that tracks to small, isolated oceanic islands
are characterized by higher genetic distances and lower
estimates of gene flow than tracks between large Pleistocene
islands.
DISCUSSION
Geological history vs. ecology?
This investigation of genetic variation in Philippine mammals
makes two general points clear. First, geological history is of
paramount importance in structuring patterns of variation
between populations of the seven species we studied. The
geological history of the Philippine archipelago has been
characterized by the long-term uplift and short-term (Pleisto-
cene) coalescence and fragmentation of groups of islands. The
maximal late Pleistocene shorelines define biogeographic units
with no history of dry-land connections to other islands or
continents. Populations of mammals on modern islands within
‘Pleistocene islands’ have been separated from one another for
c. 18,000 years; these show little or no genetic variation
between them. In contrast, populations on islands separated by
permanent barriers to dispersal have developed substantial
genetic differentiation. This pattern is concordant with
patterns of interspecific diversity in Philippine mammals; late
Pleistocene shorelines delimit faunal regions for mammals
with high levels of endemism among non-volant mammals (up
to 80%) and moderate levels of endemism among fruit bats
(up to 21%; Heaney, 1986, 1991a, 1993, in press).
The importance of Pleistocene history in structuring
genetic variation within mammalian species in the islands
of Wallacea has also been addressed by Schmitt et al. (1995),
Hisheh et al. (1998), and Maharadatunkamsi et al. (2003).
Their examination of differentiation in the fruit bats
C. nusatengarra and E. spelaea from the Lesser Sunda Islands
of Indonesia concluded that patterns of genetic distances
between island populations are associated with recent
colonization from west to east along the island arc (we note
they are also consistent with the isolation-by-distance effect),
and are more closely correlated with distances between
Pleistocene shorelines than with distances between present
shorelines, although the overall level of population subdivi-
sion that they recorded was markedly lower than in this
study. In contrast to Schmitt et al.’s (1995) and Hisheh
et al.’s (1998) results, we detected little evidence of a
relationship of island area (which is presumably related to
effective population size) or island isolation (which is
presumably related to levels of gene flow) to amounts of
variation within populations. Similarly, Juste et al. (2000)
found that, of several insular populations of an African fruit
bat, Eidolon helvum, the most isolated of these showed much
greater genetic differentiation than the others, in association
with reduced gene flow.
Second, the ecological attributes of individual species
influence the extent to which, and the manner in which,
common historical signals are expressed. Our knowledge of
natural history and distributions of these mammals success-
fully predicted whether or not they would demonstrate large or
small degrees of genetic differentiation between populations
(including, in most respects, the ecologically intermediate
P. jagori) and whether or not they would display relatively high
or low levels of variation within populations. This result has
important implications for understanding evolution – any
attempt to make general statements about rates of genetic
change between populations must take into account the
ecological attributes of the species in question. We also stress
that these insights into the common regional pattern of
variation and particularly into the differences in expression of
that pattern are unlikely to have been gained by the examina-
tion of a single species. Generalizations about the patterns of
geographic variation and their causes are best achieved
through comparative studies such as this one, and similar
studies (e.g. Caccone, 1985; Waples, 1987; Brumfield &
L. R. Heaney et al.
240 Journal of Biogeography 32, 229–247, ª 2005 Blackwell Publishing Ltd

Capparella, 1996; Zink et al., 2000; Riddle & Hafner, in press).
Thus, we conclude that geological history and ecological traits
produce interactive processes, and must be considered
simultaneously for accurate conclusions about general genetic
processes in nature to be reached.
Small islands + isolation = speciation?
One of the motivations for studying geographic variation is the
identification of especially distinctive populations. To the
degree that the origin of new species is an extension of
divergence between populations within species, identifying
divergent populations can provide insight into the processes of
speciation. In his classic volume, Mayr (1963) identified
‘geographical isolates’ as those populations most likely to
produce new species; discussing island populations in partic-
ular, he characterized geographical isolates as typically inhab-
iting islands of small area and differing environmental
conditions, having low population sizes, possessing distinctive
morphologies or behaviours, and being spatially or temporally
separated from other populations by barriers to gene flow. For
the Philippine mammals in this study, populations inhabiting
the small, isolated oceanic islands (Sibuyan, Barit, Dalupiri,
and Fuga) that were not connected as part of any large
Pleistocene island appear to qualify as geographical isolates.
These small oceanic islands are much smaller in area than the
large Pleistocene islands and have never had dry-land connec-
tions to them. All of them except Sibuyan also have depauperate
mammal faunas (Heaney et al., 1998, L.R. Heaney et al.,
unpubl. data). These island populations do not show evidence
of substantially reduced genetic variation within any given
species. We assume that effective population size is limited by
island area. While these small oceanic islands are indeed small
by most standards, they do not appear to be small enough to
erode within-population genetic variability as has been sugges-
ted for other small island populations of mammals (Berry, 1986,
1998). Using the population density estimates of Heideman &
Heaney (1989) for fruit bats on Negros Island as first
approximations, it seems likely that bat populations on even
the smallest of our islands number at least in the thousands – far
above the numbers typically believed to cause noticeable loss of
genetic variation (Lande & Barrowclough, 1987).
Although the populations on small, isolated islands do not
show reduced genetic within-island variation within a given
species, they are more genetically distinct. For each species, all
oceanic island populations possess unique alleles, and the
population of H. fischeri from Sibuyan is fixed for one unique
allele (Appendix S1). Further, the genetic distances between
populations evident in Fig. 3 are greater between Pleistocene
islands than within them, and are typically greatest to the small
oceanic islands. Whether this genetic distinctiveness is due to
selective or neutral factors is not evident from our data;
certainly, it is conceivable that the differing biotic and abiotic
environmental conditions on these small oceanic islands may
impose different selective pressures on these populations.
However, the rate of fixation of genes due to genetic drift is a
function of the inverse of effective population size (Kimura,
1983) and it may be that these small oceanic island populations
are sufficiently small to have accelerated rates of fixation of
new alleles relative to the large island populations.
It is unlikely that the genetic distinctiveness of small isolated
island populations is solely a reflection of the phylogenetic
history of these populations. Peterson & Heaney (1993)
discussed the importance of establishing population-level
phylogenies for understanding the biogeographic history of
these taxa in the Philippines. They objected to interpreting
UPGMA dendrograms, which are the results of cluster analyses
of distance data utilizing mid-point rooting, as representations
of phylogeny, preferring that estimates of phylogenetic
relationships be based on parsimony analyses of character
data using outgroup comparisons. Additional objections to the
use of allozymes in phylogenetic analyses are that allozyme
frequencies are temporally unstable (Gaines et al., 1978;
Crother, 1990) and that transitions between electromorphs
Figure 3 UPGMA dendrograms of island populations based on
genetic distance (Cavalli-Sforza & Edwards (1967) arc distance).
Abbreviations are: BAR, Barit; BIL, Biliran; CAT, Catanduanes;
DAL, Dalupiri; FUG, Fuga; LEY, Leyte; LUZ, Luzon; MRO,
Mindoro; NEG, Negros; POL, Polillo; SIB, Sibuyan. All
dendrograms drawn to same scale.
Geology, ecology, and genetic differentiation in Philippine mammals
Journal of Biogeography 32, 229–247, ª 2005 Blackwell Publishing Ltd 241

cannot be reliably ordered (Lewontin, 1991). As noted by
Peterson & Heaney (1993), the distinctive small oceanic island
populations are very unlikely to represent basal divisions vs.
other populations in a phylogenetic sense due to their
geographic setting (well away from likely avenues of coloniza-
tion) and to the fact that they are among the geologically most
recent of the Philippine islands (Steppan et al., 2003). This
implies, for example, that the long branches associated with
Sibuyan Island (Fig. 3) are due to rapid evolution on Sibuyan,
not to earlier isolation than other populations. Our interpret-
ation could be tested by use of intraspecific phylogenies
generated from DNA sequences, as currently being developed
by T. Roberts (pers. comm.).
Gene flow: endemism or habitat requirements?
This report yields two additional insights particularly relevant
to our understanding of mammals and the Philippine biota.
First, it seems an intuitively obvious expectation that a non-
volant mammal should have rates of dispersal across water
barriers orders of magnitude lower than those of a volant
mammal. It is therefore remarkable that the endemic rodent
R. everetti shows levels of gene flow between Pleistocene islands
comparable to that of the endemic fruit bat H. fischeri. One
likely explanation for this unanticipated result is that the
tolerance for disturbed, open habitat by H. fischeri is far less
than that of R. everetti (Rickart et al., 1993; Heaney et al., 1998,
1999): although it is far more effective for small mammals to
fly over water barriers than to swim across them, some bat
species may cross-water gaps less often than some rodents.
This inference suggests the hypothesis that habitat affinity may
be as important as mode of dispersal in accounting for
variation in levels of gene flow across different taxa.
Second, the pattern of variation within and between
populations exhibited by P. jagori highlights distinctive aspects
of its historical and ecological traits. It is an endemic species
that is common in good quality rain forest habitat, and it
displays low levels of within-population variation. However, it
also persists well in degraded forest and often flies in cleared
areas, and it displays high gene flow and little variation
between populations. This combination of low within- and
between-population variability is often considered to be the
earmark of a recent colonizer. It is possible that P. jagori was
formerly more restricted in distribution, perhaps on a single
Pleistocene island like its sister-taxon, P. minor, and has only
recently spread throughout the Philippine archipelago. The
data presented here cannot resolve this question, and even if
the colonization of the whole of the Philippine archipelago by
P. jagori has been a relatively recent event, some differentiation
between populations has occurred. The Sibuyan population
appears relatively distinct based on the allozyme data presented
here (Fig. 2), and morphological data (Walsh, 1998) indicate
that on the islands of Greater Mindanao, where P. jagori is
sympatric with the smaller P. minor, P. jagori has evolved
much larger body size than is found in its other island
populations. Investigations currently underway on the
phylogenetic relationships of populations of Ptenochirus and
related cynopterine fruit bats by T. Roberts using DNA
sequencing (pers. comm.) may provide additional insight into
the historical association of genetic variation and geographic
distribution in these taxa.
A simple geographical/ecological model of genetic
differentiation in oceanic archipelagos
The previous analyses suggest that patterns of genetic variation
in these Philippine mammals are strongly influenced by two
ecological variables, namely their vagility (especially the ability
to fly) and the ability to tolerate open habitats (which in the
Philippines are synonymous with heavily disturbed habitats).
These variables interact simultaneously in any given species to
largely determine its colonizing ability. These two variables, in
turn, are associated with varying levels of genetic differenti-
ation, as discussed further below.
Among our study species, C. brachyotis and M. minimus are
moderately strong fliers, and R. amplexicaudatus is a very
strong flier; all three prefer open habitat, and we characterize
them as having high colonizing ability. One of the endemic
bats (P. jagori) prefers closed-canopy, primary forest, but is a
strong flier and maintains populations in open, disturbed
forest well; we characterize it overall as having moderate
colonizing ability. Two species, H. fischeri and R. everetti, have
low colonizing ability, but for different reasons: H. fischeri can
fly, but is a relatively weak flier and rarely will fly out from
under good canopy cover, while R. everetti does well in
disturbed forest (although not in intensive agricultural areas)
but cannot fly. Ptenochirus minor prefers good canopy cover
but tolerates second growth, and so may have better colon-
izing ability overall than the prior two species, but its
restriction to a single Pleistocene island (Greater Mindanao)
implies limited abilities. We further note that C. brachyotis,
M. minimus, and R. amplexicaudatus occur on even the most
isolated of Philippine islands, whereas P. jagori and R. everetti
are absent from some of the most isolated islands (including
Barit, Batan, Dalupiri, and Fuga) and H. fischeri is absent from
those islands and also from less isolated Siquijor and
Camiguin (north of Mindanao; Lepiten 1997; Heaney et al.,
1998), providing direct empirical evidence of their limited
dispersal abilities.
We have found three categories of Philippine islands with
respect to geological history: (1) the current islands, many of
which aggregated during Pleistocene periods of low sea level
(as shown in Fig. 1); within such aggregates, these share very
similar faunas; (2) large Pleistocene islands that are surroun-
ded by deep water, which have been isolated continuously
throughout their existence but are not very distant from one
another; each has a very distinctive fauna, with 40–80%
endemism (Heaney, 1998, 2000); and (3) Pleistocene islands
that are especially small and isolated by great distance, some of
which are depauperate. The last two categories differ primarily
in degree, but in the Philippines, they are fairly discrete, as can
be seen in Fig. 1.
L. R. Heaney et al.
242 Journal of Biogeography 32, 229–247, ª 2005 Blackwell Publishing Ltd

We summarize the two variables of colonizing ability and
geographic isolation for our seven study species along two axes
in Fig. 4. We present each as a categorical variable rather than
a continuous variable because, while conceptually each is
continuous, our data demonstrate that they appear to behave
in the Philippines in a discontinuous fashion because of
historical circumstances; in general, the world is often neither
homogeneous nor continuous with respect to these variables.
Our data (Table 2, Fig. 2) show C. brachyotis, M. minimus,
and R. amplexicaudatus to have consistently high gene flow both
within and between Pleistocene islands (Nm of 4–14), although
C. brachyotis has Nm as low as 1.2–1.7 on some isolated islands
(such as Sibuyan). P. jagori, as expected, hasNm at intermediate
levels (3–5.5), and H. fischeri and R. everetti have Nm values
always far less than 1.0 between Pleistocene islands.
The converse of gene flow, genetic differentiation, is
calculated from the same data set as gene flow, and is
effectively its inverse, so that species with high gene flow have
low differentiation (FST values), and the converse (Tables 2
and 4). In other words, in the absence of high levels of gene
flow (defined as Nm less than 1.0 by Wright, 1931), substantial
genetic differentiation takes place.
Our data demonstrate that these two variables have
combined to have a strong association with rates of genetic
differentiation, as we show in the third axis in Fig. 4. Within a
Pleistocene island (Fig. 4, bottom row), none of our study
species showed genetic differentiation. On separate but adja-
cent Pleistocene islands (Fig. 5, middle row), H. fischeri and
R. everetti show moderately high genetic differentiation, but
the other species very low differentiation. On distant oceanic
islands (Fig. 5, top row), H. fischeri and R. everetti again show
heightened differentiation due to low colonizing ability.
Somewhat to our surprise, C. brachyotis shows moderate
differentiation on isolated islands, perhaps indicating that our
estimates of colonizing ability are too high. We found the
species with the highest colonizing ability (R. amplexicaudatus)
to show virtually no genetic differentiation on distant Pleis-
tocene islands, one that is smaller and a somewhat weaker flier
(M. minimus) to show slightly more, and a species with
medium colonizing ability (P. jagori) to show somewhat
greater genetic differentiation on the same islands.
Most crucially for the study of diversification, a comparison
of species on a given set of isolated oceanic islands shows
that species with high colonizing ability (R. amplexicaudatus
and M. minimus) exhibit little genetic differentiation, species
with moderate colonizing ability (P. jagori and apparently
C. brachyotis) exhibit a moderate amount of genetic
differentiation, and species with little colonizing ability
(H. fischeri, R. everetti) on the same islands exhibit high
genetic differentiation. In other words, the likelihood of any
given species developing substantial genetic differentiation
increases as it reaches progressively more isolated islands, but a
species with ecological traits that reduce its colonizing ability
(e.g. an aversion to flying out from beneath the canopy or an
inability to fly) will differentiate on less isolated islands than a
species with ecological traits that increase the rate of gene flow.
This simple description of patterns of interaction between
colonization ability and isolation, and the degree of genetic
differentiation that is associated, may be used as a model to
make predictions for other Philippine mammals, for other
Philippine organisms, and for organisms in other oceanic
archipelagos. For example, we predict that insectivorous bats in
the Philippines will show similar patterns: widespread species
that do well in disturbed habitats, such as those in the genera
Taphozous, Miniopterus, Myotis, Pipistrellus, and Scotophilus
should be very similar to C. brachyotis, M. minimus, and
R. amplexicaudatus, but Philippine endemic species in the
genera Rhinolophus and some Hipposideros that live in primary
forest should be similar to Haplonycteris and Ptenochirus. If
there are animals with still less dispersal ability than H. fischeri
and R. everetti, perhaps such as the many native shrews and
mice, or frogs with very low dispersal, we predict that they will
show even lower levels of colonizing ability and higher rates of
genetic differentiation; this prediction is at least generally
supported by the findings of Brown & Guttman (2002) and
Steppan et al. (2003). We predict that Philippine birds will
show very similar patterns to these bats overall, with widespread
species that favour disturbed habitats having genetic patterns
that resemble those of C. brachyotis, M. minimus, and
R. amplexicaudatus, whereas those that require primary forest
and/or that have limited flight will have genetic patterns that
resemble those of H. fischeri. In this study, we found all species
to have high colonizing ability and low differentiation within
Pleistocene islands, but we note that some other taxa may have
Figure 4 Graphical model of the interaction of colonizing ability
and degree of isolation on the extent of genetic differentiation by
six species of fruit bats and one murid rodent in the oceanic
Philippines. Isolation is low (within islands that merged during
Pleistocene periods of low sea level), medium (between islands
separated continuously by deep water, but by channels that are
narrow), or high (between islands separated continuously by deep
water and broad channels). Cb, Cynopterus brachyotis, Hf, Hapl-
onycteris fischeri, Mm, Macroglossus minimus, Pj, Ptenochirus
jagori, Pm, Ptenochirus minor, Ra, Rousettus amplexicaudatus,
Re, Rattus everetti.
Geology, ecology, and genetic differentiation in Philippine mammals
Journal of Biogeography 32, 229–247, ª 2005 Blackwell Publishing Ltd 243

far lower dispersal abilities and be influenced by more subtle
isolating barriers (such as different types of forest), and
therefore show higher rates of genetic differentiation within a
given modern island (perhaps including frogs and some
montane non-volant small mammals, flightless montane inver-
tebrates, etc. that live in montane and mossy forest; Heaney &
Rickart, 1990; Heaney, 2001). We predict their relative levels of
gene flow and differentiation will be structured similarly to the
bats but on a smaller geographic scale, with equivalent
ecological correlates regarding habitat fidelity and vagility.
Finally, we predict that species on the oceanic islands of
Wallacea as a whole, and in other oceanic archipelagos, will
show similar patterns of association between colonizing ability,
isolation, and differentiation. Ideal test cases would include the
birds (Lack, 1976; Ricklefs & Bermingham, 2001) and lizards
(Losos & Schluter, 2000; Harmon et al., 2003) of the Caribbean,
and the fauna of the Sea of Cortez (Case et al., 2002).
CONCLUSION
For much of the last three decades, most studies of biological
diversity in island ecosystems have approached the topic from
either an ecological or an historical/geological perspective,
typified by the ecologically-oriented equilibrium model of
MacArthur & Wilson (1967) and by the historically-oriented
vicariance model (e.g. Rosen, 1975). Moreover, there is
generally a tendency for biological research to be deliberately
conducted in very simple systems so that the impact of specific
processes may be finely parsed. This study has been useful in
demonstrating that the question of ‘geological history or
ecology’ is based on a false premise that only one or the other
is a significant factor; in this case, both are highly important, and
we predict that both will be found to be equally important in all
other oceanic archipelagos. In this case, current genetic patterns
are profoundly influenced by historical (geological) factors, but
ecological features of the various species, such as habitat
association and vagility, strongly affect rates of gene flow and,
therefore, degree of divergence among populations. Indeed, we
found that habitat association may in some cases be even more
important in determining patterns of genetic variation within
species than whether a species is volant or non-volant.
Few of our findings could have been derived from studying
any single one of the seven species, and few could have been
derived had we worked in a simpler set of islands. While very
specific and narrowly-oriented studies of simple island systems
or single species may be useful and often necessary in adding
clarity, studies that emphasize broad contexts, multiple species,
and complex regions are likely to be essential for discovering
broad patterns and processes. Indeed, this study would have
been strengthened by including even more species and islands.
The complexity of global biological diversity will best be
understood by combining historical and ecological questions
and perspectives, and considering them for many species
simultaneously; the challenge is not to determine which single
model or process predominates, but rather to determine how
the processes interact under varying circumstances, and how
narrow/focused models can be integrated into comprehensive
models that accurately portray the very real complexity of
living systems (Case & Cody, 1982; Heaney, 1986, 2000;
Whittaker, 1998; Zink et al., 2000; Hewitt, 2001; Arbogast &
Kenagy, 2002; Riddle & Hafner, in press).
ACKNOWLEDGEMENTS
We gratefully acknowledge the assistance of many colleagues
and institutions with the field portion of this study, especially
A. C. Alcala, D. S. Balete, C. Catibog-Sinha, R. I. Crombie,
C. Custodio, R. Fernandez, P. C. Gonzales, P. D. Heideman,
J. S. H. Klompen, M. Laranjo, M. V. Lepiten-Tabao,W. Pollisco,
E. A. Rickart, C. A. Ross, B. R. Tabaranza, Jr, R. C. B. Utzurrum,
the Protected Areas and Wildlife Bureau of the Philippines, the
Department of Environment and Natural Resources of the
Philippines, Silliman University, the Haribon Foundation, and
the Philippine National Museum. Permits were provided by the
Philippine Department of Natural Resources. We thank E. A.
Rickart, P. D. Heideman, T. J. McIntyre, R. S. Thorington, Jr,
R. S. Hoffmann, M. J. Carleton, J. H. Brown, G. G. Musser,
J. M. Bates, and S. J. Hackett, who have all played important
roles in the development of the ideas presented here. We thank
John Bates, Shannon Hackett, Trina Roberts, and two anony-
mous reviewers for their helpful comments on earlier drafts of
the manuscript. These studies were supported in part by the US
National Science Foundation (BSR-8514223), the Smithsonian
Institution Office of Fellowships and Grants, the Ellen Thorne
Smith and Barbara Brown Funds of the Field Museum, and the
John D. and Catherine T. MacArthur Foundation’s World
Environment and Resources Program (90-9272A).
SUPPLEMENTARY MATERIAL
The following material is available from http://www.blackwell
publishing.com/products/journals/suppmat/JBI/JBI1120/
JBI1120sm.htm
Appendix S1 Allele frequencies for Philippine mammals.
Appendix S2 Genetic distance matrices.
Appendix S3 Matrix of geographic distances between islands.
REFERENCES
Arbogast, B.S. & Kenagy, G.J. (2001) Comparative phylogeo-
graphy as an integrative approach to historical biogeo-
graphy. Journal of Biogeography, 28, 819–825.
Avise, J.C. (2000) Phylogeography, the history and formation of
species. Harvard University Press, Cambridge.
Bermingham, E. & Moritz, C. (1998) Comparative phylogeo-
graphy: concepts and applications. Molecular Ecology, 7,
367–369.
Berry, R.J. (1986) Genetics of insular populations of mammals,
with particular reference to differentiation and founder
effects in British small mammals. Biological Journal of the
Linnean Society, 28, 205–230.
L. R. Heaney et al.
244 Journal of Biogeography 32, 229–247, ª 2005 Blackwell Publishing Ltd

Berry, R.J. (1998) Evolution of small mammals. Evolution on
islands (ed. by P.R. Grant), pp. 35–50. Oxford University
Press, Oxford.
Brown, R.M. & Guttman, S.I. (2002) Phylogenetic systematics
of the Rana signata complex of Philippine and Bornean
stream frogs: reconsideration of Huxley’s modification of
Wallace’s Line at the Oriental-Australian faunal zone inter-
face. Biological Journal of the Linnean Society, 76, 393–461.
Brumfield, R.T. & Capparella, A.P. (1996) Historical diversi-
fication of birds in northwestern South America: a mole-
cular perspective on the role of vicariant events. Evolution,
50, 1607–1624.
Caccone, A. (1985) Gene flow in cave arthropods: a qualitative
and quantitative approach. Evolution, 39, 1223–1235.
Case, T.J. & Cody, M.L. (1982) Synthesis: pattern and process
in island biogeography. Island biogeography in the Sea of
Cortez (ed. by T.J. Case and M.L. Cody), pp. 307–341.
University of California Press, Berkeley.
Case, T.J., Cody, M.L. & Ezcurra, E. (eds) (2002) A new island
biogeography of the Sea of Cortez. Oxford University Press,
Oxford.
Cavalli-Sforza, L.L. & Edwards, A.W.F. (1967) Phylogenetic
analysis: models and estimation procedures. Evolution, 21,
550–570.
Cockerham, C.C. (1969) Variance of gene frequencies. Evolu-
tion, 23, 72–84.
Cockerham, C.C. (1973) Analysis of gene frequencies. Genetics,
74, 679–700.
Crother, B.I. (1990) Is ‘some better than none’ or do allele
frequencies contain phylogenetically useful information?
Cladistics, 6, 277–281.
Dietz, E.J. (1983) Permutation tests for association between
distance matrices. Systematic Zoology, 32, 21–26.
Esselstyn, J.A., Widmann, P. & Heaney, L.R. (in press) The
mammals of Palawan Island, Philippines. Proceedings of the
Biological Society of Washington, in press.
Fairbanks, R.G. (1989) A 17,000-year glacio-eustatic sea level
record: influence of glacial melting rates in the Younger Dryas
event and deep-ocean circulation. Science, 342, 637–642.
Farris, J.S. (1972) Estimating phylogenetic trees from distance
data. American Naturalist, 106, 645–668.
Felsenstein, J. (1989) PHYLIP (phylogeny inference package),
Version 3.2 manual. University of Washington, Seattle, WA.
Fisher, R.A. (1954) Statistical methods for research workers, 12th
edn. Oliver and Boyd, Edinburgh.
Fitch, W.M. & Margoliash, E. (1967) Construction of phylo-
genetic trees. Science, 155, 279–284.
Frankham, R. (1995) Inbreeding and extinction: a threshold
effect. Conservation Biology, 9, 792–799.
Frankham, R. (1996) Relationship of genetic variation to
population size in wildlife. Conservation Biology, 10, 1500–
1508.
Gaines, M.S., McClenaghan, L.R., Jr & Rose, R.K. (1978)
Temporal patterns of allozymic variation in fluctuating
populations of Microtus ochrogaster. Evolution, 32, 723–739.
Grant, P.R. (1998) Evolution on islands. Oxford University
Press, Oxford.
Gurevitch, J., Morrow, L.L., Wallace, A. & Walsh, J.S. 1992. A
meta-analysis of competition in field experiments. American
Naturalist, 140, 539–572.
Haldane, J.B.S. (1954) A test for randomness of mating.
Journal of Genetics, 52, 631–635.
Hall, R. (1998) The plate tectonics of Cenozoic SE Asia and the
distribution of land and sea. Biogeography and geological
evolution of SE Asia (ed. by R. Hall and J.D. Holloway).
Backhuys Publishers, Leiden.
Hall, R. (2002) Cenozoic geological and plate tectonic evolu-
tion of SE Asia and the SW Pacific: computer-based
reconstructions, model and animations. Journal of Asian
Earth Sciences, 20, 353–431.
Hall, R. & Holloway, J.D. (eds) (1998) Biogeography and geo-
logical evolution of SE Asia. Backhuys Publishers, Leiden.
Harmon, L.J., Schulte, J.A., II, Larson, A. & Losos, J.B. 2003.
Tempo and mode of evolutionary radiation in Iguanian
lizards. Science, 301, 961–964.
Harris, H. & Hopkinson, D.A. (1978) Handbook of enzyme
electrophoresis in human genetics. North Holland Publishing
Co., Amsterdam.
Heaney, L.R. (1986) Biogeography of mammals in SE Asia:
estimates of rates of colonization, extinction, and speciation.
Biological Journal of the Linnean Society, 28, 127–165.
Heaney, L.R. (1991a) An analysis of patterns of distribution
and species richness among Philippine fruit bats (Pteropo-
didae). Bulletin of the American Museum of Natural History,
206, 145–167.
Heaney, L.R. (1991b) A synopsis of climatic and vegetational
change in Southeast Asia. Climatic Change, 19, 53–61.
Heaney, L.R. (1993) Biodiversity patterns and conservation of
mammals in the Philippines. Asia Life Sciences, 2, 261–274.
Heaney, L.R. (2000) Dynamic disequilibrium: a long-term,
large-scale perspective on the equilibrium model of island
biogeography. Global Ecology and Biogeography, 9, 59–74.
Heaney, L.R. (2001) Small mammal diversity along elevational
gradients in the Philippines: an assessment of patterns and
hypotheses. Global Ecology and Biogeography, 10, 15–39.
Heaney, L.R. (in press) Conservation in oceanic archipelagos.
Frontiers of biogeography, recent advances in the geography of
nature (ed. by M.V. Lomolino and L.R. Heaney). Sinauer
Associates, Sunderland.
Heaney, L.R., Heideman, P.D., Rickart, E.A., Utzurrum, R.B. &
Klompen, J.S.H. (1989) Elevational zonation of mammals in
the central Philippines. Journal of Tropical Ecology, 5, 259–
280.
Heaney, L.R. & Regalado, J.C. (1998) Vanishing treasures of the
Philippine rain forest. The Field Museum, Chicago, IL.
Heaney, L.R. & Rickart, E.A. (1990) Correlations of clades and
clines: geographic, elevational, and phylogenetic distribution
patterns among Philippine mammals. Vertebrates in the
tropics (ed. by G. Peters and R. Hutterer), pp. 321–332.
Museum Alexander Koenig, Bonn.
Geology, ecology, and genetic differentiation in Philippine mammals
Journal of Biogeography 32, 229–247, ª 2005 Blackwell Publishing Ltd 245

Heaney, L.R., Balete, D.S., Dolar, L., Alcala, A.C., Dans, A.,
Gonzales, P.C., Ingle, N., Lepiten, M.V., Oliver, W., Rickart,
E.A., Tabaranza, B.R., Jr & Utzurrum, R.C.B. (1998) A
synopsis of the mammalian fauna of the Philippine Islands.
Fieldiana Zoology New Series, 88, 1–61.
Heaney, L.R., Balete, D.S., Rickart, E.A., Utzurrum, R.C.B. &
Gonzales, P.C. (1999) Mammalian diversity on Mt. Isarog, a
threatened center of endemism on southern Luzon Island,
Philippines. Fieldiana New Series, 95, 1–62.
Heideman, P.D. & Heaney, L.R. (1989) Population biology and
estimates of abundance of fruit bats (Pteropodidae) in
Philippine submontane rainforest. Journal of Zoology (Lon-
don), 218, 565–586.
Hewitt, G.M. (2001) Speciation, hybrid zones and phyloge-
ography – or seeing genes in space and time. Molecular
Ecology, 10, 907–913.
Hisheh, S., Westerman, M. & Schmitt, L.H. (1998) Biogeo-
graphy of the Indonesian archipelago: mitochondrial DNA
variation in the fruit bat, Eonycteris spelaea. Biological
Journal of the Linnean Society, 65, 329–345.
Holloway, J.D. (2003) Biological images of geological history:
through a glass darkly or brightly face to face? Journal of
Biogeography, 30, 165–180.
International Union of Biochemistry and Molecular Biology
(1992) Enzyme nomenclature. Academic Press, SanDiego, CA.
Juste, J., Ibanez, C. & Machordom, A. 2000. Morphological
and allozyme variation of Eidolon helvum (Mammalia:
Megachiroptera) in the islands of the Gulf of Ghana. Bio-
logical Journal of the Linnean Society, 71, 359–378.
Kimura, M. (1983) The neutral theory of molecular evolution.
Cambridge University Press, Cambridge.
Lack, D. (1976) Island biology illustrated by the land birds of
Jamaica. University of California Press, Berkeley.
Lande, R. (1995) Mutation and conservation. Conservation
Biology, 9, 782–791.
Lande, R. & Barrowclough, G.F. (1987) Effective population
size, genetic variation, and their use in population man-
agement. Viable populations for conservation (ed. by Soule,
M.), pp. 87–123. Cambridge University Press, Cambridge.
Lepiten, M.V. (1997) The mammals of Siquijor Island, central
Philippines. Sylvatrop, 5, 1–17.
Levene, H. (1949) On a matching problem arising in genetics.
Annals of Mathematics and Statistics, 20, 91–94.
Lewontin, R. (1991) Twenty-five years ago in genetics: elec-
trophoresis in the development of evolutionary genetics:
milestone or millstone? Genetics, 128, 657–662.
Livshits, G., Sokal, R.R. & Kobyliansky, E. (1991) Genetic
affinities of Jewish populations. American Journal of Human
Genetics, 49, 131–146.
Lomolino, M.V. (2000) A call for a new paradigm of island
biogeography. Global Ecology and Biogeography, 9, 1–6.
Losos, J.B. & Schluter, D. (2000) Analysis of an evolutionary
species-area relationship. Nature, 408, 847–850.
MacArthur, R.H. & Wilson, E.O. (1967) The theory of island
biogeography. Princeton University Press, Princeton.
Maharadatunkamsi, Hisheh, S., Kitchener, D.J. & Schmitt,
L.H. (2003) Relationships between morphology, genetics
and geography in the cave fruit bat Eonycteris spelaea
(Dobson, 1871) from Indonesia. Biological Journal of the
Linnean Society, 79, 511–522.
Mantel, N. (1967) The detection of disease clustering and a
generalized regression approach. Cancer Research, 27, 209–
220.
Mayr, E. (1963) Animal species and evolution. Belknap Press,
Cambridge, MA.
Mey, W. (2003) Insular radiation of the genus Hydropsyche
(Insecta, Trichoptera: Hydropsychidae) Pictet, 1834 in the
Philippines and its implications for the biogeography of
Southeast Asia. Journal of Biogeography, 30, 227–236.
Mitchell, A.H.G., Hernandez, F. & De La Cruz, A.P. (1986)
Cenozoic evolution of the Philippine archipelago. Journal of
Southeast Asian Earth Sciences, 1, 3–22.
Mittermeier, R.A., Myers, N. & Mittermeier, C.G. (1999)
Hotspots, earth’s biologically richest and most endangered
terrestrial ecoregions. CEMEX, Mexico City.
Myers, N. (1988) Environmental degradation and some eco-
nomic consequences in the Philippines. Environmental
Conservation, 15, 205–213.
Nei, M. (1973) Analysis of gene diversity in subdivided pop-
ulations. Proceedings of the National Academy of Science
USA, 70, 3321–3323.
Nei, M. (1977) F-statistics and analysis of gene diversity in
subdivided populations. Annals of Human Genetics, 41, 225–
233.
Nei, M. (1978) Estimation of average heterozygosity and genetic
distance from a small number of individuals. Genetics, 89,
583–590.
Packham, G. (1996) Cenozoic SE Asia: reconstructing its
aggregation and reorganization. Tectonic evolution of
Southeast Asia, Vol. 106 (ed. by R. Hall and D. Blundell),
123–152. Geological Society Special Publication, The Geo-
logical Society, London.
Peterson, A.T. & Heaney, L.R. (1993) Genetic differentiation
in Philippine bats of the genera Cynopterus and Haplo-
nycteris. Biological Journal of the Linnean Society, 49, 203–
218.
Powers, D.A., Lauerman, T., Crawford, D. & DiMichele, L.
(1991) Genetic mechanisms for adapting to a changing
environment. Annual Review of Genetics, 25, 629–659.
Rickart, E.A., Heaney, L.R. & Utzurrum, R.C.B. (1991) Dis-
tribution and ecology of small mammals along an elevational
transect in southeastern Luzon. Journal of Mammalogy, 72,
458–469.
Rickart, E.A., Heaney, L.R., Heideman, P.D. & Utzurrum,
R.C.B. (1993) The distribution and ecology of mammals on
Leyte, Biliran, and Maripipi Islands, Philippines. Fieldiana:
Zoology, New Series, 72, 1–62.
Ricklefs, R.E. & Bermingham, E. (2001) Nonequilibrium
diversity dynamics of the Lesser Antillean avifauna. Science,
294, 1522–1524.
L. R. Heaney et al.
246 Journal of Biogeography 32, 229–247, ª 2005 Blackwell Publishing Ltd

Riddle, B.R. & Hafner, D.J. (in press) The past and future roles
of phylogeography in historical biogeography. Frontiers of
biogeography (ed. by M.V. Lomolino and L.R. Heaney).
Sinauer Associates, Sunderland.
Rosen, D.E. (1975) A vicariance model of Caribbean bioge-
ography. Systematic Zoology, 24, 431–464.
Schluter, D. (2000) The ecology of adaptive radiation. Oxford
University Press, Oxford.
Schmitt, L.H., Kitchener, D.J. & How, R.A. (1995) A genetic
perspective of mammalian variation and evolution in the
Indonesian archipelago: biogeographic correlates in the fruit
bat Cynopterus. Evolution, 49, 399–412.
Shaw, C.R. & Prasad, R. (1970) Starch-gel electrophoresis of
enzymes: a compilation of recipes. Biochemical Genetics, 4,
297–320.
Siddall, M., Rohling, E.J., Almogi-Labin, A., Hemleben, C.,
Meischner, D., Schmelzer, I. & Smeed, D.A. (2003) Sea-level
fluctuations during the last glacial cycle. Nature, 423, 853–
858.
Slatkin, M. (1985) Rare alleles as indicators of gene flow.
Evolution, 39, 53–65.
Slatkin, M. (1993) Isolation by distance in equilibrium and
nonequilibrium populations. Evolution, 47, 264–279.
Slatkin, M. & Barton, N. (1989) A comparison of three indirect
methods for estimating average level of gene flow. Evolution,
43, 1349–1368.
Sneath, P.H.A. & Sokal, R.R. (1973) Numerical taxonomy.
W. H. Freeman, San Francisco.
Sokal, R.R. (1988) Genetic, geographic, and linguistic distances
in Europe. Proceedings of the National Academy of Science
USA, 85, 1722–1726.
Sokal, R.R. & Rohlf, F.J. (1981) Biometry, 2nd edn. W. H.
Freeman and Co., New York.
Sokal, R.R. & Wartenberg, D.E. (1983) A test of spatial auto-
correlation analysis using an isolation-by-distance model.
Genetics, 105, 219–237.
Soule, M.E. (1976) Allozyme variation, its determinants in
space and time. Molecular evolution (ed. by F.J. Ayala), pp.
60–77. Sinauer Associates, Sunderland.
Soule, M.E. (1987) Viable populations for conservation. Cam-
bridge University Press, Cambridge.
Steppan, S.J., Zawadski, C. & Heaney, L.R. (2003) Molecular
phylogenyof the endemicPhilippine rodentApomys (Muridae)
and the dynamics of diversification in an oceanic archipelago.
Biological Journal of the Linnean Society, 80, 699–715.
Swofford, D.L. & Berlocher, S.H. (1987) Inferring evolutionary
trees from gene frequency data under the principle of max-
imum parsimony. Systematic Zoology, 36, 293–325.
Swofford, D.L. & Selander, R.B. (1981) BIOSYS-1: A FOR-
TRAN program for the comprehensive analysis of electro-
phoretic data in population genetics and systematics. Journal
of Heredity, 72, 281–283.
Walsh, J.S., Jr (1998) Geographic variation in Philippine fruit
bats (Mammalia: Pteropodidae) and systematics of the
cynopterine section. Unpublished PhD Dissertation,
University of Chicago, Chicago, IL, 257pp.
Waples, R.S. (1987) A multispecies approach to the analysis of
gene flow in marine shore fishes. Evolution, 41, 385–400.
Weir, B.S. (1990) Genetic data analysis. Sinauer Associates, Inc.
Publishers, Sunderland.
Weir, B.S. & Cockerham, C.C. (1984) Estimating F-statistics for
the analysis of population structure.Evolution, 38, 1358–1370.
Whittaker, R.J. (1998) Island biogeography: ecology, evolution
and conservation. Oxford University Press, Oxford, 285pp.
Wildlife Conservation Society of the Philippines (1997)
Philippine red data book. Bookmark, Manila, Philippines,
240pp.
Wright, S. (1931) Evolution in Mendelian populations. Gen-
etics, 16, 97–159.
Wright, S. (1943) Isolation by distance. Genetics, 28, 114–138.
Wright, S. (1951) The genetical structure of populations. An-
nals of Eugenics, 15, 323–354.
Wright, S. (1965) The interpretation of population structure
by F-statistics with special regard to systems of mating.
Evolution, 19, 395–420.
Wright, S. (1978) Evolution and genetics of populations, Vol. 4.
Variability within and among natural populations. University
of Chicago Press, Chicago, IL.
Zink, R.M., Blackwell-Rago, R.C. & Ronquist, F. (2000) The
shifting roles of dispersal and vicariance in biogeography.
Proceedings of the Royal Society of London Series B – Biolo-
gical Sciences, 267, 497–503.
BIOSKETCHES
Lawrence Heaney is Curator and Head of the Division of Mammals at the Field Museum. His research is focused on the evolution,
ecology and conservation of mammal diversity in island and island-like ecosystems, especially in Southeast Asia and the western USA.
Joseph Walsh is a Lecturer in the Undergraduate Program in Biological Sciences at Northwestern University and a Research
Associate in Zoology at the Field Museum. He has worked on the systematics and biogeography of Southeast Asian fruit bats and is
currently involved in restoration of tallgrass prairie and oak savannas in northern Illinois.
Townsend Peterson is Associate Professor in Ecology & Evolutionary Biology, and Curator of Ornithology in the Natural History
Museum, University of Kansas. His research focuses on ecological and historical factors shaping species’ geographic distributions.
Editor: Philip Stott
Geology, ecology, and genetic differentiation in Philippine mammals
Journal of Biogeography 32, 229–247, ª 2005 Blackwell Publishing Ltd 247