the role of flt3 in haematopoietic malignancies
TRANSCRIPT

Haematopoiesis involves the complex regulation of pro-liferation and differentiation of many cell types — fromthe parent stem cell to the functional components of thebone marrow and peripheral blood, including the ery-throid, myeloid, lymphoid and platelet lineages (FIG. 1).Despite the many genetic checks and balances that are inplace to regulate haematopoesis, mutations of crucialregulatory genes can occur that disrupt normalhaematopoiesis and promote leukemogenesis.
The FMS-like tyrosine kinase 3 (FLT3) gene encodesa membrane-bound receptor tyrosine kinase (RTK)that has a crucial role in normal haematopoiesis.Recently, FLT3 mutations have been found in patientswith acute lymphoblastic leukaemia (ALL; 1–3% ofpatients), myelodysplasia (5–10%) and acute myeloidleukaemia (AML; 15–35%), making FLT3 one of themost frequently mutated genes in haematologicalmalignancies. A flurry of research has increased ourunderstanding of the biology of these FLT3 mutationsand how they affect clinical outcome, but many ques-tions concerning the aetiology of the mutations andhow they cause leukaemia remain unanswered. Whatregulatory pathways does mutated FLT3 disrupt andhow does the activation of FLT3 mutants differ fromthat of wild-type FLT3? What is the prognostic strengthof mutations in FLT3 and should we use them to guidetreatment decisions? Is FLT3 a reasonable target forsmall-molecule inhibitors? Exploring these issues could
lead to a better understanding of leukemogenesis, more-specific prognostic classifications for haematologicalmalignancies and, most importantly, new treatmentsthat target the pathways involved.
Structure and expression of FLT3The Flt3 (also known as fetal liver kinase 2, Flk2) gene wascloned independently by two groups using enrichedmouse fetal liver cells1 and placenta cells2. The cloning ofthe human FLT3 gene soon followed3,4. The human FLT3gene is located on chromosome 13q12, and has 85%amino-acid sequence homology with mouse Flt3 (REF. 5).FLT3 encodes an RTK of 993 amino acids in length thatbelongs to the RTK subclass III family, which comprisesmembrane-bound receptors with five immunoglobulin-like extracellular domains, a transmembrane domain, ajuxtamembrane domain and two intracellular tyrosine-kinase domains (TKDs) linked by a kinase-insertdomain6 (FIG. 2). All members of the RTK subclass IIIfamily have important roles in the proliferation, differen-tiation and survival of normal haematopoietic cells7.Other members of the RTK subclass III family includethe macrophage colony-stimulating factor (M-CSF)receptor (FMS), Steel factor receptor (KIT) and the recep-tors for platelet-derived growth factors A and B(PDGFRA and PDGFRB)6.
Originally, the FLT3 gene was thought to contain 21exons, mimicking the intron–exon boundaries of KIT.
THE ROLE OF FLT3 INHAEMATOPOIETIC MALIGNANCIESDerek L. Stirewalt and Jerald P. Radich
Normal haematopoietic cells use complex systems to control proliferation, differentiation and celldeath. The control of proliferation is, in part, accomplished through the ligand-induced stimulationof receptor tyrosine kinases, which signal to downstream effectors through the RAS pathway.Recently, mutations in the FMS-like tyrosine kinase 3 (FLT3) gene, which encodes a receptortyrosine kinase, have been found to be the most common genetic lesion in acute myeloidleukaemia (AML), occurring in ~25% of cases. Exploring the mechanism by which these FLT3mutations cause uncontrolled proliferation might lead to a better understanding of how cellsbecome cancerous and provide insights for the development of new drugs.
Clinical Research Division,Fred Hutchinson CancerResearch Center and theDivision of Oncology,University of Washington,Seattle, WA 98109, USA.Correspondence to D.L.S.e-mail: [email protected]:10.1038/nrc1169
650 | SEPTEMBER 2003 | VOLUME 3 www.nature.com/reviews/cancer
R E V I E W S

NATURE REVIEWS | CANCER VOLUME 3 | SEPTEMBER 2003 | 651
R E V I E W S
normally express FLT3 (REF. 20), but FLT3 expression isuncommon in this type of leukaemia. Recently,Armstrong et al.21 found that patients with ALL withtranslocations involving the mixed-lineage leukaemia(MLL) gene had a unique gene-expression profile of adistinct subtype of acute leukaemia. Overexpression ofthe wild-type FLT3 gene distinguished this subtypewith MLL translocations from other types of ALL andAML21. These findings raise the possibility that specificgenetic disturbances, such as MLL translocations,might influence the expression of FLT3.
Structure and expression of FLT3 ligandAfter cloning the Flt3 gene, two groups independentlyused soluble mouse Flt3 to clone the gene encodingmouse Flt3 ligand (Flt3l)22,23. The mouse Flt3l cDNAwas then used to clone the human FLT3L gene23,24.Mouse and human FLT3L encode proteins of 231 and235 amino acids, respectively15. There is 72% identity atthe amino-acid level between mouse and humanFLT3L. The identity seems to be greatest (73%) for theextracellular region of the protein and least (52%) forthe cytoplasmic domain15.
The FLT3L gene encodes a type 1 transmembraneprotein, which contains an amino-terminal signallingpeptide, four extracellular helical domains, spacer andtether regions, a transmembrane domain and a smallcytoplasmic domain15,25. Other type 1 transmembraneproteins include KIT ligand (Steel factor or stem-cellfactor) and M-CSF (CSF1) — all three proteins havesimilar intron–exon boundaries, sizes and structures,which indicates that they are ancestrally related15.Ligand activity resides mainly in the extracellulardomains, with the spacer and tether regions linking theactive extracellular component to the transmembranedomain and, thereby, anchoring the ligand to the cell.The function of the cytoplasmic domain remainsunknown, but it does not seem to have a crucial role instimulation of FLT3 (REF. 15).
There are several different isoforms of both mouseand human FLT3L22–24,26. Human FLT3L occurs mainlyas the full-length transmembrane isoform, which can beproteolytically cleaved in the spacer and tether region togenerate a second soluble isoform23,24. This second isoform does not contain the carboxyl end of the trans-membrane isoform and consists mainly of the extracel-lular domains of the protein. A third, rare isoform ofFLT3L has an alternatively spliced exon 6 that creates apremature stop codon, resulting in a slightly differentsoluble form of the protein15.
Unlike human FLT3L, the most abundant isoformof mouse Flt3l is a 220-amino-acid, membrane-asso-ciated protein. This membrane-associated isoformresults from the failure to splice out an intron, creat-ing a reading-frame shift that encodes a stretch ofhydrophobic amino acids15. As a result, the mem-brane-associated isoform does not include the spacer,tether, transmembrane or cytoplasmic domains thatnormally anchor the ligand26. Instead, the stretch ofhydrophobic amino acids anchors the ligand to thecell surface. The absence of the spacer and tether
However, recent studies indicate that the intron–exonboundaries of the first three extracellular immunoglob-ulin domains differ between FLT3 and KIT, with FLT3encoding eight exons in this region compared with fiveexons for KIT 8. Two forms of human FLT3 have beendescribed — a 158–160-kDa membrane-bound proteinthat is glycosylated at N-linked glycosylation sites in theextracellular domain and an unglycosylated 130–143-kDa protein that is not membrane bound9,10 (FIG. 2).
In normal human cells, expression of FLT3 occursmainly in early myeloid and lymphoid progenitors11,and not in erythroid cells12, megakaryocytes13 or mastcells14 (FIG. 1). FLT3 messenger RNA has also been foundin other lympho-haematopoietic organs, such as theliver, spleen, thymus and placenta. However, expressionin these organs might be due to contamination fromhaematopoietic cells rather than actual expression in theorgan tissue3,15.
Many types of leukaemia express FLT3. In mosttypes, FLT3 expression by the leukaemic cell mirrorsthat by the normal cell counterpart (FIG. 1). So, FLT3 isexpressed by normal myeloid and lymphoid progeni-tors, and by the leukaemic cells of 70–90% of patientswith AML10,11 (which originates from primitive, multi-potent colony-forming cells or myeloid-committedcolony-forming unit cells16) and ALL (the common B-cell form of ALL derives from immature, progenitor Bcells17). B-cell chronic lymphocytic leukaemia (CLL)probably arises from a more mature lymphocyte popu-lation18, and CLL cells do not express FLT3. Similarly,although chronic myeloid leukaemia (CML) arisesfrom an early progenitor stem cell19, a large percentageof CML cells retain differentiation capacity, and expres-sion of FLT3 is lost during differentiation. However,FLT3 expression does not always correlate with the pre-sumed cell of origin of the leukaemia. For example,most T-cell ALLs evolve from early thymocytes, which
Summary
• FMS-like tyrosine kinase 3 (FLT3) is a receptor tyrosine kinase (RTK) involved in theproliferation, differentiation and apoptosis of haematopoietic cells. It is mainlyexpressed by early myeloid and lymphoid progenitor cells.
• Many cells of the haematopoietic system produce FLT3 ligand (FLT3L), whichpromotes dimerization and activation of FLT3. The activated receptor then activatesthe phosphatidylinositol 3-kinase (PI3K) and RAS signal-transduction cascades.
• The FLT3 internal tandem duplication (ITD) results from a head-to-tail duplication of3–400 base pairs in exons 14 or 15, which encode the juxtamembrane domain of FLT3.
• Point mutations in FLT3 occur in heavily conserved areas of the intracellular tyrosine-kinase domain (TKD), homologous to point mutations that are seen in other RTKssuch as KIT and FMS.
• FLT3 mutations are the most frequent genetic lesion seen in acute myeloid leukaemia(AML). The prevalence of FLT3 ITDs is 15–35%, with an additional 5–10% of patientshaving FLT3 TKD mutations.
• Both types of FLT3 mutation cause ligand-independent activation of the receptor andactivation of downstream signalling pathways.
• The presence of a FLT3 ITD is associated with poor clinical outcome in both paediatricand adult patients with AML.
• Several drugs that target FLT3 are in early clinical trials.

652 | SEPTEMBER 2003 | VOLUME 3 www.nature.com/reviews/cancer
R E V I E W S
FLT3L is expressed by most tissues, includinghaematopoietic organs (spleen, thymus, peripheral bloodand bone marrow), prostate, ovary, kidney, lung, colon,small intestine, testis, heart and placenta, with the highestlevel of expression by peripheral-blood mononuclearcells. The brain is one of the few tissues without demon-strable expression of FLT3L. Most immortalizedhaematopoietic cell lines express FLT3L28,29, and expres-sion has even been detected in immortalized colorectalcancer cell lines30. There are no large studies of expressionof FLT3L in primary tumour samples.
The expression of FLT3L by a wide variety of tis-sues is in contrast to the limited expression pattern ofFLT3, which is found mainly in early haematopoieticprogenitor cells. This indicates that the expression of
regions also prohibits the normal proteolytic cleavagethat occurs in the human full-length transmembraneisoform25,26. A soluble variant of mouse Flt3l, similarto the soluble variant of human FLT3L with an alter-natively spliced exon 6, has also been identified. Thebiological relevance of these various isoforms isunknown, and studies have not looked in any detailat whether specific isoforms are preferentiallyexpressed by subtypes of tissue. All human andmouse isoforms of FLT3L are biologically active,although the relative activity of the different isoformsremains to be clarified15. Unlike other RTK ligands(for example, Steel factor), FLT3L does not havespecies specificity27; soluble human FLT3L can activatemouse Flt3, and vice-versa.
T cell
Self-renewingpluripotent stem cell
Committed stem cell
Early lymphoid progenitor
Early myeloid progenitor
NK cell
EarlyNK cell
B cell
PreB
ProB
PreT
ProT
Platelets
Monocyte
CFU-G CFU-M
CFU-E
CFU-MK
CFU-GM
CFU-B
CFU-D
MK
Granulocyte Dendritic cell Mast cell/basophil
–
–
– +/–
? –
+/– +/– +
+/–
–
+/–
+/– +
+ +
– – +/–
– – ?
+
–RBC
Figure 1 | Expression of FLT3 in normal haematopoiesis. The maturation and differentiation of cells during normalhaematopoiesis is shown, indicating how expression of FMS-like tyrosine kinase 3 (FLT3) is linked to this process (shown as +, –,+/– or ? (unknown)). FLT3 is mainly expressed by early myeloid and lymphoid progenitor cells, with some expression by the moremature monocytic lineage cells. Colony-forming units for the erythroid (CFU-E), megakaryocytic (CFU-MK), granulocytic–monocytic(CFU-GM), basophilic (CFU-B), granulocytic (CFU-G), monocytic (CFU-M), and dendritic (CFU-D) lineages are shown. NK cell,natural killer cell; RBC, red blood cell.

NATURE REVIEWS | CANCER VOLUME 3 | SEPTEMBER 2003 | 653
R E V I E W S
5 minutes of stimulation and reaching a maximumafter 15 minutes32. Degraded by-products of the lig-and–receptor complex are seen as early as 20 minutesafter stimulation32. So, the entire process of activation,internalization and degradation of FLT3 occurs rapidlyin cells, similar to the rapid activation and turnover ofother RTK subclass III receptors such as KIT34. The rateof FLT3 production, the rapidity of degradation of theactivated FLT3L–FLT3 complex and the downstreameffects of activation probably participate in a complexfeedback loop that regulates normal receptor activity.
Exposure to FLT3L is a crucial component of theregulation of FLT3 activity. Although serum levels ofFLT3L are usually low in normal adults35, they areinducible. Patients who have low white-blood-cellcounts secondary to either haematopoietic disease (forexample, FANCONI ANAEMIA or APLASTIC ANAEMIA) orchemotherapy have markedly increased levels of FLT3Lin the serum, indicating that the systemic release of FLT3L might be an attempt to promotehaematopoiesis35. However, although systemic adminis-tration of FLT3L has been shown to mobilize primitivehaematopoietic stem cells in mouse models36, clinicaluse of FLT3L in humans has not been attempted.
FLT3 is a more rate-limiting step in determining thetissue specificity of FLT3 activation than is expressionof FLT3L. All expression data, however, must be inter-preted with caution, because most studies have exam-ined expression of FLT3L mRNA, which might notfully predict the associated protein levels.
Regulation of FLT3 activation by FLT3LMost unstimulated FLT3 receptors reside as monomersin the plasma membrane (FIG. 2). In this inactive state,the conformation of the receptor might result in stericinhibition of dimerization and exposure of phosphorylacceptor sites in the TKD by the juxtamembranedomain; a similar mechanism of inhibition betweenregulatory domains and catalytic domains has beenfound in other families of tyrosine kinases31. After stim-ulation with FLT3L, membrane-bound FLT3 quicklychanges conformation, forming a homodimer andexposing phosphoryl acceptor sites in the TKD32 (FIG. 2).Dimerization stabilizes this conformational change,which further enhances activation of the receptor33.Phosphorylation of FLT3 occurs within 5–15 minutesafter FLT3L binding. The FLT3L–FLT3-phosphatecomplex is then rapidly internalized, starting within
FANCONI ANAEMIA
An autosomal-dominantdisorder that is characterized byskeletal growth abnormalitiesand a high incidence ofmalignancies, especially of thebone marrow.
APLASTIC ANAEMIA
An acquired injury to the bonemarrow that causes a decrease innormal haematopoesis. Thisleads to potentially fatalcomplications from lownumbers of red and white bloodcells and platelets.
G G
G
GG
G G
G G
Transcription
TranslationGlycosylation
FLT3L binding
Phosphorylation
Internalizationand degradaton
60S
40S
5′
mRNA
Protein
KI3′
L
E EE
E E
TM
JM
K K
P P P
P
P
P P P P
Figure 2 | Structure and activation of wild-type FLT3. Transcription of the FMS-like tyrosine kinase 3 (FLT3) gene produces FLT3mRNA, which is translated to FLT3 protein. FLT3 contains five extracellular immunoglobulin-like domains (E), a transmembranedomain (TM), a juxtamembrane domain (JM) and two tyrosine-kinase domains (K) that are linked through the tyrosine-kinase insert(KI). Cytoplasmic FLT3 undergoes glycosylation (G), which promotes localization of the receptor to the membrane. Wild-type FLT3remains as a monomeric, inactivated protein on the cell surface until FLT3 ligand (L), probably in a dimeric form, binds the receptorand induces receptor dimerization. FLT3 dimerization promotes phosphorylation (P) of the tyrosine-kinase domains, therebyactivating the receptor and downstream effectors. The dimerized receptors are quickly internalized and degraded.

654 | SEPTEMBER 2003 | VOLUME 3 www.nature.com/reviews/cancer
R E V I E W S
interacting or forming complexes with numerous otherproteins including GRB2 (REF. 41), GRB2-associatedbinder 2 (GAB2)43 and SHIP 40. FLT3 and these adap-tor-protein complexes probably stimulate downstreameffectors in the RAS–RAF–MEK–ERK pathway41,44 andPI3K pathway 41,43,45 (FIG. 3). However, unlike theFMS–Flt3 chimeric receptor, wild-type human FLT3does not directly bind the p85 subunit of PI3K. Instead,it forms complex associations with many proteins, suchas GRB2, GAB2, SHIP, SHP2 (a protein tyrosine phos-phatase), CBL (a proto-oncogene) and CBLB (CBL-related protein), that ultimately act on p85 (REFS 41,43,45)
(FIG. 3). FLT3 activation also modulates crucial down-stream signal transducers and activators of transcrip-tion, such as STAT5A46, and prevents the upregulationof expression of genes that can induce apoptosis, suchas BCL2-associated X protein (BAX)47. However, all ofthe signalling-pathway data must be interpreted withcaution, because most of these experiments were car-ried out in either mouse cell models or human cell linesartificially induced by gene transduction. In addition,FLT3 pathways seem to be highly species and tissue spe-cific38,41, which indicates that it might be difficult togeneralize the above findings to normal humanhaematopoietic cells. Nevertheless, these studies showthat FLT3 has a crucial role in many of the regulatoryprocesses of haematopoietic cells, including phospho-lipid metabolism, transcription, proliferation andapoptosis, and they have established a possible linkbetween FLT3 and the RAS pathway — which isthought to be involved in leukemogenesis48–50.
FLT3 function in normal cellsActivation of FLT3 by FLT3L promotes the in vitrogrowth of early progenitor cells51–53. FLT3-mediatedresponses are highly dependent on the cell type andother growth factors that are acting on the cell.Gabbianelli et al.12 and others53 found that stimulationwith FLT3L without the addition of other growth fac-tors promoted the monocytic differentiation of earlyhaematopoietic progenitor cells, without a marked pro-liferative response. However, FLT3L stimulation in com-bination with other growth factors such as interleukin-3(IL-3), G-CSF, CSF1, GM-CSF, erythropoietin (EPO)and KIT ligand produced a more vigorous proliferativeresponse54 and greatly enhanced the development ofgranulocytic–monocytic colony-forming units12,53. Inexperiments comparing these various combinations ofgrowth factors, Rusten et al.53 found that the combina-tion of FLT3L, KIT ligand, and IL-3 produced the mostrobust proliferative effect, with the growth of primitiveand more committed myeloid progenitor cells.Stimulation with FLT3L has no significant effect on ery-thropoiesis or megakaryopoiesis 12, owing to the factthat FLT3 is not expressed by these cell lineages.
Stimulation of FLT3 is also important in lymphoctyedevelopment. FLT3L stimulation in combination with IL-7 and IL-11 promoted the long-term clonal expan-sion and differentiation of uncommitted mousehaematopoietic progenitor cells51. Similar experimentsusing human fetal bone marrow found that stimulation
The low concentrations of FLT3L in the serum of nor-mal adults and the proximity of cells that express FLT3Lat a high level to early progenitor haematopoietic cellsindicate that a PARACRINE feedback loop might controlFLT3 activation. Primitive haematopoietic cells would bestimulated by the local secretion of FLT3L or by directcontact with FLT3L on the cell surface of neighbouringmononuclear cells. In addition, the co-expression of bothFLT3L and FLT3 by the same cells indicates that anAUTOCRINE feedback loop might function in somehaematopoietic cells28, so that an individual cell can deter-mine its own level of FLT3 activation. Such autocrineactivation has been proposed for other growth-factorreceptors, such as granulocyte–macrophage colony-stimulating factor (GM-CSF) and its receptor37.
Many questions regarding the activation of FLT3by FLT3L and degradation of the FLT3L–FLT3 com-plex remain unanswered. Do cells require long-termexposure to FLT3L to induce the desired effects ofFLT3 activation? Do the kinetics of FLT3 activationand degradation differ between the various types ofnormal haematopoietic cell or between normal andmalignant haematopoietic cells?
The FLT3 signal-transduction pathwayBefore the discovery of FLT3L, the RTK–RAS signal-transduction pathway was studied using chimeric recep-tors containing the extracellular ligand-binding domainsof human FMS fused to the intracellular catalyticdomains of mouse Flt3 (REFS 38,39). Activation of thechimeric receptor with CSF1 transformed mouse celllines led to a full proliferative response in the absence ofother growth factors38,39. Co-precipitation assays foundthat the chimeric FMS–Flt3 protein directly associatedwith the Src-homology 2 (SH2) domain of the p85 sub-unit of phosphatidylinositol 3-kinase (PI3K; a regulatoryprotein of phospholipid metabolism) and with growthfactor receptor-bound protein 2 (Grb2; an adaptor pro-tein)38,39. This physical association also extended to othereffectors of phospholipid metabolism and proliferation,including phospholipase Cγ1 (Plcγ1; a regulatory proteinof phosphoinositol metabolism), SH2-containingsequence proteins (SHCs; coupling proteins that areinvolved in numerous phopholipid and proliferativepathways) and GTPase-activating protein (Gap; animportant regulatory protein of the proliferativeRas–Raf–Mek–Erk pathway)38,39. These interactions ledto phosphorylation of the associated proteins and activa-tion of downstream effectors such as Vav (which isinvolved in haematopoiesis)38,39. More recently, the stim-ulated FMS–Flt3 receptor was found to be associatedwith SH2-domain-containing inositol phosphatase(Ship)40. Although Ship is involved in phosopholipidmetabolism, it might also act as a negative regulator ofproliferation by competitively binding phosphorylatedSHC proteins that would otherwise activate theRas–Raf–Mek–Erk pathway41.
The cloning of FLT3L allowed more-direct study ofthe molecular pathways involved in FLT3activation22,24. Similar to findings with the chimericmodels, SHC proteins were rapidly phosphorylated42,
PARACRINE
An effect in a cell that is causedby stimulation by hormonessecreted from another cell.
AUTOCRINE
An effect in a cell that is causedby stimulation by hormonessecreted from the same cell.

NATURE REVIEWS | CANCER VOLUME 3 | SEPTEMBER 2003 | 655
R E V I E W S
with FLT3L in combination with IL-7 promoted stro-mal-cell-independent growth of pro-B cells and the differentiation of pro-B cells to pre-B cells55. FLT3L-mediated activation might also induce the proliferationand differentiation of early T cells23,56. A combination ofIL-3, IL-6, IL-7 and FLT3L generated a two–sixfoldincrease in the number of the most primitive thymicprogenitors compared with cells grown under the sameconditions without FLT3L56.
Given the widespread expression of FLT3 and thefunctional effects observed, it is surprising that Flt3-knockout mice had relatively normal haematopoiesis,without gross morphological changes in the bone mar-row57. The only apparent cellular abnormality in Flt3-knockout mice was a small reduction in the size of thepro-B-cell and pre-B-cell compartments. Transplantationof a mixture of early haematopoietic progenitor cellsexpressing either non-functioning Flt3 or wild-type Flt3led to reconstitution of the bone marrow with cells thatmostly expressed wild-type Flt3, which indicates thatthese cells had a significant growth advantage57. Thisadvantage occurred across all cell lineages, indicating thatFlt3 disruption caused a global disturbance in thehaematopoietic system57. In addition, mice with knock-outs of both Kit and Flt3 developed lethal haematopoieticdeficiencies. So, FLT3 has an important, but not absolute,role in normal haematopoiesis, working in conjunctionwith other growth factors to foster the proliferation anddifferentiation of myeloid and lymphoid cells. These find-ings indicate that selective FLT3 inhibitors might blockinappropriate FLT3 activation in leukaemia cells withouttoo severe haematopoietic side-effects.
FLT3 and the immune systemStimulation with FLT3L promotes dendritic-cell devel-opment58,59, leading to the clonal expansion of dendriticcells in the bone marrow, spleen, lymph nodes andperipheral blood60,61. Given the central role of dendriticcells as antigen-presenting cells in the immune system,these findings indicate that stimulation with FLT3Lmight enhance the immune response to malignancies. Itis of note that FLT3L-mediated clonal expansion of den-dritic cells promotes the regression of both solid andhaematopoietic tumours in mouse models62–66.
Studies in humans combining FLT3L stimulationwith immunization with cancer antigens have yieldedmixed results67,68. Fong et al.68 examined the ability ofFLT3L-mobilized dendritic cells to induce an immuneresponse against a carcinoembryonic antigen (CEA)self-peptide, which is often overexpressed in solid malig-nancies. Twelve patients with solid tumors (nine withcolon tumours, one with a tumour of the rectum andtwo with lung tumours) participated in the Phase Istudy. Stimulation with FLT3L expanded the dendritic-cell population by 20-fold in vivo and increased the pro-duction of cytotoxic CD8+ T cells specific for the CEApeptide. In addition, two of the 12 patients had com-plete resolution of their metastatic disease, and onepatient had a partial response. Clinical responses corre-lated with the clonal expansion of CD8+ T cells specificfor the CEA peptide.
Proteins
Transcription
Translation
60S
40S
5′
mRNA
3′
CBLPI3K
SHIP
4E-BP1
elF4E
SHP2 GAB2
mTOR
S6K
PDK1
AKT
GRB2
RAS
RAF
MEK
ERK
RSK
CREB
ELK
P38
STATBCL2 BAD
P P
SHC
P
P P
L
Figure 3 | The FLT3 signalling cascade. Although the FMS-like tyrosine kinase 3 (FLT3)signalling cascade has not been definitively characterized, this figure shows some of thecomplex associations and downstream effects that probably occur after activation of FLT3.Binding of FLT3 ligand (L) to FLT3 triggers the PI3K (phosphatidylinositol 3-kinase) and RASpathways, leading to increased cell proliferation and the inhibition of apoptosis. PI3K activityis probably regulated through various interactions between FLT3, SH2-containing sequenceproteins (SHCs) and one or more other proteins, such as SH2-domain-containing inositolphosphatase (SHIP), SH2-domain-containing protein tyrosine phosphatase 2 (SHP2), CBL(a proto-oncogene) and GRB2-binding protein (GAB2). Activated PI3K stimulatesdownstream proteins such as 3-phosphoinositide-dependent protein kinase 1 (PDK1),protein kinase B (PKB/AKT) and the mammalian target of rapamycin (mTOR), which initiatethe transcription and translation of crucial regulatory genes through the activation of p70 S6kinase (S6K) and the inhibition of eukaryotic initiation factor 4E-binding protein (4E-BP1). In addition, PI3K activation blocks apoptosis through phosphorylation of the pro-apoptoticBCL2-family protein BAD (BCL2 antagonist of cell death). Activated FLT3 also associateswith GRB2 through SHC, so activating RAS. RAS activation stimulates downstreameffectors such as RAF, MAPK/ERK kinases (MEKs), extracellular-signal-regulated kinase(ERK), and the 90-kDa ribosomal protein S6 kinase (RSK). These downstream effectorsactivate cyclic adenosine monophosphate-response element binding protein (CREB), ELKand signal transducer and activators of transcription (STATs), which lead to the transcriptionof genes involved in proliferation. Both pathways probably also interact with many other anti-apoptotic and cell-cycle proteins, such as WAF1, KIP1 and BRCA1. Solid arrows depictdirect associations between proteins, whereas dashed arrows depict associations that havebeen indicated indirectly.

656 | SEPTEMBER 2003 | VOLUME 3 www.nature.com/reviews/cancer
R E V I E W S
However, Disis et al.67 were unable to show a signifi-cant beneficial immune response as a result of FLT3Lstimulation in their Phase I study. Ten patients (ninewith breast cancer and one with ovarian cancer, all over-expressing HER2/NEU) were vaccinated withHER2/NEU peptides while receiving either FLT3L, orFLT3L and GM-CSF. FLT3L alone was ineffective at pro-moting cytotoxic T-cell responses specific forHER2/NEU, and it led to the development of autoim-mune disease, perhaps through the clonal expansion ofinterferon-γ-secreting T cells67. However, unlike thestudy by Fong et al.68, FLT3L-mobilized dendritic cellswere not infused before vaccination in this study, whichmight have compromised immune responses.
Although pre-clinical and early clinical studies indi-cate an immunological benefit from FLT3L-mediatedpriming of dendritic cells under some circumstances,any therapeutic use of FLT3L must be carefully weighedagainst the potential side effect of autoimmune disease.In addition, there is a possibility of stimulating malig-nant cell growth through activation of the RAS–mito-gen-activated protein kinase pathway, especially inhaematological malignancies. After all, FLT3L stimulatesthe proliferation of immortalized AML cell lines andprimary AML samples47,69, and FLT3L stimulation ofmalignant haematopoietic cells could potentially pro-vide them with a growth advantage over normalhaematopoietic cells.
FLT3 mutations in haematopoietic malignanciesNakao et al.70 first described FLT3 mutations inhaematopoietic malignancies in 1996, which stimu-lated a large amount of research characterizing thefrequency and relevance of FLT3 mutations70–78. Themost common form of FLT3 mutation is an internaltandem duplication (ITD) in exons 14 and 15 (previ-ously known as exons 11 and 12), which occurs in15–35% of patients with AML70–77,79–81 and 5–10% ofpatients with myelodysplasia (MDS)78,82. FLT3 ITDsare formed when a fragment of the juxtamembrane-domain coding sequence is duplicated and inserted ina direct head-to-tail orientation (FIG. 4; BOX.1). Thelength of the ITD varies from 3 to ≥400 base pairs,and the reading frame of the transcript is always pre-served, either by a faithful in-frame duplication or bythe insertion of nucleotides at the ITD junction tomaintain the original reading frame75.
The second most common type of FLT3 mutation isa missense point mutation in exon 20 (previouslyknown as exon 17) of the TKD. TKD mutations occurin patients with AML (5–10%), MDS (2–5%) and ALL(1–3%)76,83,84, and the most common nucleotide substi-tution (GAT → TAT) changes an aspartic acid to tyro-sine (D835Y). Other mutations in codon 835, such asdeletions, have also been described, but these are far lesscommon. In addition, point mutations, deletions andinsertions in the codons surrounding codon 835 havealso been found76,83,85. Similar to the ITDs, all TKDmutations maintain the same open reading frame. Asingle patient will occasionally have both an ITD andTKD point mutation of FLT3, but most patients have
PM
Internalduplications Point mutations
InsertionsDeletions
ITD
A
Ba b
c d
Inactive wild-type monomers
Adaptor protein
Unique adaptorprotein
Active ITD and wild-type dimer Active monomer with point mutation
Active wild-type dimer
ITD
?
?
P
P
P
P
PP
P P P
P
P P
P
P
TM
JM
K1 K2
L
Figure 4 | Structure and location of FLT3 mutations. A | Internal tandem duplications(ITDs) occur in exons 14 or 15 of the juxtamembrane domain (JM), which lies directly betweenthe transmembrane domain (TM) and the first tyrosine-kinase domain (K1). Point mutations,insertions and deletions are found in exon 20 of K2. B | A comparison of the possible modesof activation of wild-type FLT3 and mutant FLT3. a | Without ligand (L), receptors remainmostly in the inactivated monomeric form. The conformation of the monomeric receptorprobably involves repulsive forces that prevent dimerization and possibly block adaptorproteins (green) from docking. b | After ligand binding, conformational changes in FLT3promote dimerization, phosphorylation (P) and adaptor-protein docking, transmitting signalsto downstream effectors (green arrow). c | ITDs probably promote ligand-independentdimerization and activation of FLT3 through changing the conformation of the expressedreceptor. Experiments indicate that ITD-containing FLT3 might activate both similar (blue) andunique (red) pathways compared with wild-type FLT3. The conformational changesassociated with ITDs might change the structure of the receptor such that unique adaptorproteins can now dock. d | Missense mutations in TKD2 might also promote ligand-independent activation. It is unknown if dimerization is required or if the downstream effectorsare similar to those activated through the wild-type cascade.

NATURE REVIEWS | CANCER VOLUME 3 | SEPTEMBER 2003 | 657
R E V I E W S
It has been hypothesized that the development ofAML requires at least two classes of genetic abnormalitythat independently disrupt at least two regulatoryprocesses in haematopoietic cells. For example, one pri-mary genetic event such as the PML–RARA transloca-tion might impair differentiation, whereas another ‘hit’such as an FLT3 ITD promotes proliferation andsurvival95. This theory derives from Knudson’s ‘two hithypothesis’, which proposes that most cancers require atleast two mutations, with the first being either germinalor somatic, whereas the second is always somatic96.However, although this concept is intellectually satisfy-ing, it is unclear how FLT3 mutations actually cooperateto cause cancer. For example, FLT3 mutations disruptdifferentiation, proliferation and apoptosis71,87,89,91, andthe components of these regulatory pathways have atremendous amount of both redundancy and ‘cross-talk’between pathways (FIG. 3). Therefore, a single mutationmight not completely disrupt an entire pathway or itsregulatory function, or might not simply affect a singlefunctional pathway. In some cases, there might be a‘pathway dosage’ effect, whereby different mutationsmust act on the same regulatory pathway to promote amore complete disruption of its normal activity. Forexample, both FLT3 ITDs and t(15;17) disrupt differenti-ation91,95, but these mutations are often found together inpatients with AML. The implications of these findingsfor treatment are important. If multiple genetic abnor-malities are required to induce AML, then the removal ofone of these abnormalities by novel therapeutic agentsmight be sufficient to partially reverse the survivaladvantage of the malignant clone.
Simple models of FLT3 ITDs indicate inappropriatesignal activation of its usual transduction pathway.However, it might be that FLT3 ITDs recruit new path-ways as part of their pathogenic properties. Mutant FLT3activates many downstream effectors, including RAS,ERK, protein kinase B (PKB/AKT) and STAT5 (REFS
71,87,89). In mouse transduction models, wild-type andmutant Flt3 affect these downstream regulatory proteinsin a different manner89. For example, Flt3l stimulation ofmouse haematopoietic cells transduced with wild-typeFlt3 results in only weak phosphorylation of Stat5, with-out any evidence of Stat5 binding to DNA, whereashaematopoietic cells transduced with Flt3 ITD have high-level Stat5 phosphorylation and Stat5 DNA binding 89.Recently, Mizuki et al.97 used DNA microarrays to com-pare the RNA-expression profiles of mouse haematopoi-etic cell lines transduced with either wild-type Flt3 or Flt3ITD. Marked differences in gene expression were foundbetween the wild-type and ITD Flt3-transduced cells.Pu.1 and C/EBPα — two proteins that are involved in theregulation of haematopoietic-cell differentiation and pro-liferation — were significantly repressed in Flt3 ITD-transduced cells but were expressed in the wild-type Flt3-transduced cells97. Together, these data indicate thatactivation of FLT3 ITD and wild-type receptors mightaffect downstream effectors in unique ways. If so, thesedifferences, however subtle, might be used to develop tar-geted treatments for patients with AML with FLT3 ITDsand for patients with other haematopoietic malignancies.
only one type of FLT3 mutation76,86. In total, ~25–45%of patients with AML will have some form of mutationin FLT3, making disturbances in this gene the mostcommon form of genetic abnormality in AML.
Biology of FLT3 ITDs. In vitro transduction studies haveshown that FLT3 ITDs promote ligand-independentdimerization, auto-phosphorylation and constitutiveactivation of the receptor71,87–89. How FLT3 ITDs pro-mote constitutive activation is unknown. Experimentsindicate that mutations in the juxtamembrane domainprobably eliminate the naturally occurring repressiveregions of FLT3, which normally prevent dimerizationwithout ligand stimulation (FIG. 4B). Various mutations(insertions, deletions or duplications) in several regionsof the juxtamembrane domain cause constitutive acti-vation of the receptor. In addition, FLT3 engineeredwith an ITD, but without a TKD region, dimerizes withwild-type receptors without the addition of ligand andcan activate wild-type FLT388. A similar repressivesequence has been identified in the juxtamembranedomain of vascular endothelial growth factor receptor 1(VEGFR1), and disruption of this repressive sequencepromotes activation of a VEGFR1–VEGFR2 chimericprotein90. So, the loss of repressor function with subse-quent constitutive receptor activation might be a rela-tively common event. Another possible explanation isthat FLT3 ITDs disrupt the intrinsic negative regulatoryeffects of the juxtamembrane domain on its own TKDs,whereby FLT3 ITDs change the conformation of themonomeric receptor, exposing the phosphoryl acceptorsites in the TKDs. Once these sites are exposed, phos-phoryl donors dock and activate the receptor. In thismodel, receptors with FLT3 ITDs would not requiredimerization for activation. It should be emphasizedthat these models of mutant FLT3 activation have notbeen completely proven.
The constitutive activation induced by FLT3 ITDspromotes ligand-independent proliferation and blocksmyeloid differentiation of early haematopoietic cells inmouse model systems71,87,88,91. Recently, Kelly et al.92
found that FLT3 ITDs induce a myeloproliferative-likedisease in mouse bone-marrow transplant models.These experiments indicate that FLT3 ITDs are suffi-cient to promote a phenotypical change associated withenhanced proliferation, but they are not sufficient toinduce overt acute leukaemia. Other genetic lesions thatare common in acute leukaemia have been found toinduce a similar myeloproliferative (but not leukaemic)phenotype in mouse models. Of particular interest ist(15;17) — a balanced, reciprocal chromosome translo-cation of the promyelocytic leukaemia (PML) gene at15q22 and the retinoic acid receptor-α (RARA) gene at17q12 — which causes a myeloproliferative-like syn-drome in mouse models, with ~15–20% of the miceprogressing to a leukaemia-like disease93,94. In humanAML, FLT3 ITDs are often found in patients witht(15;17). It is of note that mice transplanted withhaematopoietic cells with both t(15;17) and a FLT3 ITDuniversally develop a leukaemia-like syndrome, with amarked reduction in the latency time of disease onset95.

658 | SEPTEMBER 2003 | VOLUME 3 www.nature.com/reviews/cancer
R E V I E W S
studies have involved sample populations thatreceived various treatments, including transplanta-tion in some cases. In addition, the methods used todetect FLT3 ITDs varied. Most studies used PCRamplification of exons 14 and 15 followed byagarose-gel electrophoresis, although other tech-niques such as single-strand conformational analysisor gene scan (BOX 1) might be more sensitive73,106.Prospective studies in uniformly treated patientcohorts will hopefully clarify the clinical relevance ofFLT3 ITDs in adults.
Despite attempts to sort patients with AMLaccording to risk (‘risk stratification’), on the basis oftheir presenting clinical and laboratory characteris-tics, outcomes after chemotherapy for similarpatients are highly variable. Why are patients withAML with FLT3 ITDs sometimes cured with standardchemotherapy, but most often are not? Perhaps notall FLT3 ITDs are equal, although there are no data tosupport this hypothesis. Other possibilities are thatFLT3 ITDs are innocent by-standers (which isunlikely given the experimental data cited above) orthat other genetic abnormalities associated with FLT3ITDs confound the analyses, by making the outcomeeither worse or better. FLT3 ITDs have been associ-ated with other known prognostic abnormalitiessuch as t(15;17), t(6;9) and MLL mutations76,105.Some of these genetic abnormalities, such as t(15;17),are associated with a good prognosis, whereas others,such as t(6;9), are associated with poor clinical out-come107. Knowledge of how FLT3 ITDs interact withother cytogenetic abnormalities might provide a bio-logical insight into leukemogenesis and clarify theuncertainties of risk stratification.
Two studies of patients with AML with t(15;17)found no significant impact of FLT3 ITDs on clinicaloutcome, but there was a trend (not statistically signifi-cant) towards shorter disease-free survival and higherrelapse rates in patients with an FLT3 mutation72,108. Inaddition, both studies found a significant associationbetween FLT3 ITDs and leukocytosis, which indicatesthat the FLT3 ITD has a proliferative biological effect
Biology of FLT3 TKD point mutations. Mutations inthe TKD region of FLT3 also promote constitutivephosphorylation of the receptor and ligand-indepen-dent cell growth83–85. It is not known how FLT3 TKDmutations activate the receptor or whether the activa-tion of TKD-mutant FLT3 more closely resemblesthat of wild-type FLT3 or of FLT3 ITD. There is ~80%identity between the TKDs of FMS, KIT and FLT3(REF. 6), and TKD mutations have also been found inboth FMS and KIT98–100. In TKD-mutated FMS andKIT, the tyrosine-kinase activity is highly dependenton the type of amino-acid substitution in the TKD,such that some point mutations cause a markedincrease in activity, whereas others inhibitactivity101,102. These experiments are in sharp contrastto the mutational experiments in the juxtamembranedomain of FLT3, which show that any mutation in thejuxtamembrane region causes an increase inactivity88. On the basis of experiments with other RTKsubclass III receptors, FLT3 TKD mutations probablyincrease the level of intrinsic tyrosine-kinase activity,rather than interfering with repressive domains andpromoting dimerization.
Clinical relevance of FLT3 ITDs. FLT3 mutations are ofclinical importance70–77,79–82,103,104 (TABLE 1). FLT3 ITDshave been strongly associated with LEUKOCYTOSIS, high blastcounts, normal cytogenetics, t(15;17) and t(6;9). Recently,one study indicated that FLT3 ITDs might also be associ-ated with duplications and/or double-strand DNA breaksin the breakpoint cluster region of the MLL gene105.
The prevalence of FLT3 ITDs in patients withAML increases with age, ranging from 5–15% in pae-diatric patients to 25–35% in adults. Most studies inpaediatric patients with AML have found that FLT3ITDs are strong, independent predictors of poor clin-ical outcome74,103,104 (TABLE 1). Studies in adult patientswith AML are not as conclusive but still show anoverall trend for poor clinical outcome in patientswith FLT3 ITDs75–77,80 (TABLE 1). However, all of thesestudies potentially suffer from the inherent biases ofretrospective analyses. For example, most of these
LEUKOCYTOSIS
An abnormal increase in thenumber of white blood cells.
Table 1 | Clinical relevance of FLT3 ITDs in patients with AML*
Number of Frequency of Mean age CR DFS OS 5-year DFS 5-year OS Referencepatients in FLT3 ITDs (%) of patients wild-type:ITD wild-type:ITD wild-type:ITD wild-type:ITD wild-type:ITDstudy (years) (%) (months) (months) (%) (%)
64 11 6 93:57 N.D. N.D. 69:14 N.D. 104
94 5 6.8 N.D. N.D. N.D. 60:20 N.D. 103
91 17 10.5 74:40 N.D. N.D. N.D. N.D. 74
201 23 N.D. 75:71 (N.S.) N.D. N.D. 50:18 45:14 80
979 20 N.D. 67:71 (N.S.) 18:13 14:10 (N.S.) N.D. N.D. 76
106 13 41 N.D. N.D. 29:13 N.D. N.D. 77
854 27 41 84:78 N.D. N.D. 46:30 44:32 81
1003 23 57 70:70 (N.S.) 13:7 15:12 (N.S.) N.D. N.D. 75
140 27 67 40:55 (N.S.) 8:8 (N.S.) 7:8 (N.S.) N.D. N.D. 73
*All results are statistically significant unless labelled as (N.S.). ML, acute myeloid leukaemia; CR, complete response; DFS, disease-free survival; FLT3, FMS-liketyrosine kinase 3; ITD, internal tandem duplication; N.D., not determined; N.S., not statistically significant (P > 0.05); OS, overall survival.

NATURE REVIEWS | CANCER VOLUME 3 | SEPTEMBER 2003 | 659
R E V I E W S
LOSS OF HETEROZYGOSITY
(LOH). A loss of one of thealleles at a given locus as a resultof a genomic change, such asmitotic deletion, geneconversion or chromosomemissegregration.
HOMOLOGOUS
RECOMBINATION
The process by which segmentsof DNA are exchanged betweentwo DNA duplexes that sharehigh sequence similarity.
strong over-representation of the FLT3 ITD, indicatingthat LOH might not account for most of the differ-ences in ITD:wild-type ratios. It has been proposedthat an increased ITD:wild-type ratio might be due toHOMOLOGOUS RECOMBINATION of the FLT3 ITD allele,thereby creating more copies of the FLT3 ITD allelethan of the wild-type allele76. Gene amplification of thewild-type FLT3 gene has been shown in one leukaemiacell line110, but this has not been examined in actualpatient samples. The most plausible explanation for thevariation of allelic ratios between leukaemia cases liesin the observation that, in many cases, leukaemia mightbe a polyclonal process. Some sub-populations of AMLcells might have an FLT3 ITD, whereas others containonly wild-type FLT3. For example, FLT3 mutational
in cases of AML with t(15;17). FLT3 ITDs are mainlyfound in the heterozygous state, but they are some-times found in a single-copy, homozygous state afterloss of the normal FLT3 allele. The FLT3 ‘allelic ratio’ ofmutated to wild-type FLT3 might explain some of theprognostic variability of FLT3 ITDs. Several studieshave indicated that patients with AML having highITD:wild-type FLT3 ratios have the worst clinicalprognosis76,109. Whitman et al.109 found that threepatients with an extremely high ITD:wild-type ratiohad LOSS OF HETEROZYGOSITY (LOH) of the wild-type FLT3allele, which indicates that LOH might explain some ofthe differences in the ITD:wild-type ratio betweenpatients. However, Thiede et al.76 were unable to detectLOH of the wild-type allele in nine patients with
Box 1 | The detection of FLT3 mutations
Fortunately, the detection of mutations in the FMS-like tyrosine kinase 3 (FLT3) gene is relatively straightforward, fastand cheap. Internal tandem duplications (ITDs) of FLT3 can usually be detected by gel electrophoresis of thepolymerase-chain-reaction (PCR) products generated from amplifying exons 14 and 15 of the FLT3 gene, in which mostITDs occur (see figure, part a). The PCR primers will generate a wild-type band and a larger FLT3 ITD band, which canbe separated by gel electrophoresis.The detection of FLT3 point mutations is more complicated, but can also be achieved by PCR. The double-strand PCRproduct is denatured, then run on a single-strand conformational polymorphism (SSCP) gel, which allows the singlestrands to re-anneal, forming a unique secondary structure. A base-pair mutation results in a different secondarystructure, which migrates differently on the gel than the structure of the wild-type sequence (see figure, part b). Theabnormal bands can then be excised from the gel and sequenced for confirmation.
There is some evidence that the ratio of mutant to wild-type alleles might determine the pathogenesis of FLT3mutations. This can be assessed by fluorescent-labelled PCR, whereby the labelled PCR products are separatedaccording to size by migration through a capillary system, with the intensity of the products (relating to relativeabundance) determined by laser excitation. In part c of the figure, lane A shows a PCR of a wild-type FLT3 allele; thecorresponding fluorescent signal is shown below. Lane B and panel B show a PCR of a leukaemia sample with a lowITD:wild-type FLT3 ratio (0.9). Lane C and panel C show a PCR of a leukaemia sample with mainly mutated FLT3(giving a high ITD:wild-type ratio of 2.4) and lane D and panel D show a sample with equal amounts of the wild-typeand mutated FLT3 allele (images courtesy of S. Meschinchi).
a Duplication of exon 14 in FLT3 ITD c ITD: wild-type FLT3 allelic ratio
b FLT3 point mutations in exon 20
Normal:
Normal
Exon 14 Intron Exon 15
Patient: Exon 14
ITD
Intron Exon 15
Patient samples
FLT3 ITDWT
Normal Patient samples
240 280
0.0A
0.9B
2.4C
0.5D
320 360 400
A B C D
Allelic ratio

660 | SEPTEMBER 2003 | VOLUME 3 www.nature.com/reviews/cancer
R E V I E W S
FLT3 inhibitorsAfter the success of imatinib (Glivec; Novartis) — asmall-molecule tyrosine-kinase inhibitor (TKI) used totreat CML111 — there has been a tremendous effort todiscover other small-molecule inhibitors that could beefficacious for other malignancies. Given the high fre-quency of activating FLT3 mutations in patients withAML, FLT3 and its downstream pathway are attractivetargets for directed inhibition. Recent experiments indi-cate that MLL rearrangements might exert some of theirneoplastic effects through upregulation of expression ofwild-type FLT3 (REF. 21). Therefore, TKIs specific forFLT3 might have an even broader use in AML and/orother haematopoietic malignancies than first predictedon the basis of FLT3 mutation analyses. Non-specificTKIs (such as herbimycin A, AG1295, and AG1296)block the constitutive activation of FLT3 ITDs, as well asother proteins such as heat shock protein 90 (HSP90),thereby inhibiting the growth of cells that express FLT3ITDs112–115. However, many of these non-specific com-pounds are highly toxic in humans, prompting searchesfor more selective compounds for clinical use.
Several promising TKIs have been identified recentlythat might change the treatment of haematopoieticmalignancies (TABLE 2). CEP-701 and CEP-5214(Cephalon) are two orally bioavailable TKIs derivedfrom indolocarbazole116,117. Both compounds preferen-tially inhibit autophosphorylation of wild-type andmutant FLT3, and show limited inhibition of KIT, FMSand PDGFR. Levis et al.116 found that CEP-701 is cyto-toxic to leukaemia blasts from patients with AML in vitro and in vivo, and prolongs the survival of miceinjected with BaF3/ITD cells. A Phase I/II trial is nowunderway in patients with refractory and relapsed AMLwith FLT3 mutations, and preliminary results have beenpresented in abstract form at the American Society ofHematology118. In eight patients, CEP-701 seemed to berelatively well tolerated, with nausea, fatigue and neu-tropaenia being the most commonly described side-effects. CEP-701 dosage has now been titrated to 80 mgtwice a day. At this early stage, it is difficult to discern thepotential efficacy of CEP-701, but one patient did nor-malize his peripheral-blood counts, with <5% blasts inhis bone marrow, which indicates that this agent mightbe beneficial in some patients with AML.
Sugen has also identified several compounds(SU5416, SU5614 and SU11248) that block FLT3 activa-tion119–121. Unlike the compounds developed byCephalon, these agents also inhibit other RTKs such asKIT, PDGFR and VEGFR119–121. After initial Phase I tri-als, Giles et al.122 recently examined SU5416 at a dose of145 mg m–2 in 55 patients with refractory or relapsedAML regardless of FLT3 status. The main toxicitiesincluded headaches (14% of patients), dyspnoea (14%),infusion-related reactions (11%) and thromboticepisodes (7%). Three patients (5%) had a partialresponse. An early Phase I trial examined SU11248 in 32patients with advanced AML. Again, FLT3 mutationswere not necessary for inclusion in the trial. SU11248was relatively well tolerated, with fatigue being the maindose-limiting toxicity121,123. Approximately half of the
status can change between diagnosis and relapse, withsome patients developing a new or different FLT3mutation on relapse, whereas other patients lose theirFLT3 mutation altogether106,108. If a leukaemia con-tained two clones with different FLT3 mutation status(one with cells containing an FLT3 ITD, and one withwild-type FLT3), then patients having a higher percent-age of clones with FLT3 ITDs would have higherITD:wild-type ratios. If the cells with the FLT3 ITD aremore resistant to therapy, then patients with higherITD:wild-type ratios would be more likely to have apoor clinical outcome than patients with a lower ratio.
It is not known how a high FLT3 ITD:wild-typeallelic ratio affects mRNA and protein expression ofthe mutant and wild-type alleles. It would be expectedthat those samples with a high FLT3 ITD:wild-typeallelic ratio would also have a higher level of FLT3 ITDmRNA and of FLT3 ITD protein expression. Recently,Libura et al.105 examined the relative mRNA expres-sion of 31 patients with AML with FLT3 ITDs com-pared with 100 patients with AML with wild-typeFLT3. Overall, the mean FLT3 mRNA expression ofthe ITD group was not significantly different fromthat of the wild-type group, but there was a widerange of FLT3 expression within the patients withFLT3 ITD and wild-type FLT3. As noted previously,wild-type FLT3 expression is increased in high-riskpatients with MLL translocations21. Taken together,these studies indicate that overexpression of FLT3, beit wild-type or mutated, might be a common threadin high-risk acute leukaemia.
Clinical relevance of FLT3 TKD mutations. Althoughboth FLT3 ITDs and FLT3 TKD mutations cause lig-and-independent activation in vitro, the clinical rele-vance of these mutations might be different. Onestudy of 201 patients with AML found no significantassociation between FLT3 TKD mutations and age,white-blood-cell counts, leukaemic-blast counts orcytogenetics; however, there was a trend for shorterdisease-free survival for patients with FLT3 TKDmutations83. In a larger study of 979 patients, sub-group analyses in patients of less than 60 years of agefound that FLT3 TKD mutations were associated witha higher white-blood-cell count, higher blast percent-age and a relative reduction in overall survival76, butno significant differences in disease-free or overallsurvival could be established for the entire AML pop-ulation. Libura et al.105 have found that, similar toFLT3 ITDs, FLT3 TKD mutations occur more com-monly in AML samples with MLL duplications or adouble-strand break in the BCR of the MLL gene. Inaddition, samples with FLT3 TKD mutations tend tohave higher levels of FLT3 expression than sampleswith either FLT3 ITDs or wild-type FLT3 (REF. 105). It isnot known how these latest findings factor into anyprognostic relevance of FLT3 TKD mutations, but asFLT3 TKD mutations are less frequent than FLT3ITDs, extremely large studies will be required to deter-mine the impact, if any, of FLT3 TKD mutations onclinical outcome.

NATURE REVIEWS | CANCER VOLUME 3 | SEPTEMBER 2003 | 661
R E V I E W S
translocations that overexpress wild-type FLT3 (REFS
21,110). These findings indicate that PKC412 (and pos-sibly other TKIs) might be effective for the treatmentof a wide range of haematopoietic malignancies thatoverexpress FLT3, even if they do not have activatingFLT3 mutations. If similar experiments can validatethese findings in other types of cell line and primaryAML samples, then the therapeutic use of TKIs mightbroaden to include other malignancies, such as AMLand B-cell ALL, or any haematopoietic disease thatoverexpresses FLT3.
Compared with the results of initial clinical trials ofimatinib for the treatment of CML, early reports ofFLT3 TKIs for the treatment of AML might seemunimpressive. This should not be surprising. AML is anaggressive, heterogeneous disease, with various cytoge-netic abnormalities and mutations. CML, especially inthe chronic phase, is far less aggressive and morehomogeneous, uniformly characterized by a singlegenetic abnormality, the Philadelphia chromosome,which is a reciprocal translocation of the BCR and ABLgenes (t(9;22)). In animal models, the BCR–ABLtranslocation is necessary and sufficient to promoteleukemogenesis; moreover, there is evidence thatBCR–ABL alone both drives differentiation and blocksapoptosis, so that disruption of the activity of theBCR–ABL fusion protein could undermine theleukaemic process. By contrast, FLT3 mutations aloneare not sufficient to promote leukemogenesis in mousemodels95. Finally, all studies of FLT3 inhibitors haveinvolved heavily pre-treated patients who have failedchemotherapy — a therapy-resistant and severely illgroup of patients. So, in this patient cohort, a reductionin blast counts can be considered a success, and this is
patients had some form of haematological response asshown by a >50% reduction in their percentage ofleukaemia blasts.
MLN518 (otherwise known as CT53518 fromMillenium) is a novel piperarazinyl quinazoline thatinhibits the growth of FLT3 ITD-transformed cells in vitro and in vivo124. Similar to the Sugen compounds,MLN518 also inhibits wild-type FLT3, PDGFRB andKIT. In a Phase I study of patients with relapsed orrefractory AML, preliminary findings showed that twoof six patients had >50% reduction in the number ofbone blasts. This study is currently in progress withadditional patients125.
Lastly, PKC412 (Novartis) is a benzoylstauro-sporine — initially developed as a VEGFR inhibitor —that blocks the activity of wild-type and mutant FLT3(REFS 21,126). On the basis of Phase I dosing studies insolid tumours, a Phase II trial has begun studyingPKC412 at a dose of 75 mg three times a day inpatients with relapsed or refractory AML with FLT3mutations127. All patients were poor candidates formyelosuppressive chemotherapy and had a poor per-formance status of less than 3. Mild nausea was themost common side effect in the first eight patients, butthree of the eight patients discontinued use of the drugfor reasons other than progression of disease (liver tox-icity, fatal pulmonary toxicity and severe lethargy). It isunclear if these complications were related to drugadministration. No partial or complete responses wereobtained in this heavily pre-treated and debilitatedpatient population. Analysis of results is continuing,and additional data should be available in the nearfuture. Of additional interest is the fact that PKC412has been found to kill leukaemia cells with MLL
Table 2 | Tyrosine-kinase inhibitors in early clinical trials
Compound Company Route Receptor Side effects Responses Clinical developmentinhibition
CEP-701 Cephalon Oral FLT3 (wild-type Nausea, fatigue 1 of 8 patients: Phase II trial in adult and mutant) and neutropaenia <5% blasts in AML*; accrual in progress
bone marrow
SU5416 Sugen IV FLT3 (wild-type Headaches, dyspnoea, 3 of 55 patients: Phase II trial in adult AMLand mutant), infusion reaction and partial remission and high-grade MDSKIT, FMS and thrombosis with 6–25% bone-PDGFR marrow blasts
SU11248 Sugen Oral FLT3 (wild-type Fatigue 32 patients, 13 of Phase I trial in adult AML;and mutant), 16 evaluable patients: accrual in progressKIT, FMS and >50% reduction inPDGFR peripheral blasts
MLN518 Millenium Oral FLT3 (wild-type Not reported yet 8 patients, 2 of 3 Phase I trial in adult AMLand mutant), evaluable patients: and high-grade MDS;KIT and PDGFR 50% reduction in accrual in progress
marrow blasts
PKC412 Novartis Oral FLT3 (wild-type Nausea; two deaths of 1 of 8 patients: Phase II trial in adult AMLand mutant), uncertain aetiology; >50% reduction inPKC, VEGFR and pulmonary toxicity; marrow blastsPDGFR lethargy
L-000021649 Merck Oral FLT3 (wild-type Not reported yet Not reported yet Phase I trial in paediatricand mutant) AML and ALL: startingand VEGFR accrual
* Patients with AML had to have either an FLT3 internal tandem duplication or a mutation in the tyrosine-kinase domain. ALL, acute lymphocytic luekaemia; AML, acutemyeloid leukaemia; FLT3, FMS-like tyrosine kinase 3; IV, intravenous; MDS, myelodysplasia; PDGFR, platelet-derived growth factor receptor; PKC, protein kinase C; VEGFR,vascular endothelial growth factor receptor.
662 | SEPTEMBER 2003 | VOLUME 3 www.nature.com/reviews/cancer
R E V I E W S
Future directionsMany questions remain to be answered with regardsto the biology of FLT3 and its role in leukemogenesis.There is the fundamental question of how FLT3mutations actually promote leukemogenesis, andwhat other molecular pathways are recruited or com-promised for this process to occur. As discussed, pre-liminary data indicate that FLT3 mutations are notsufficient for leukemogenesis but might cooperatewith other genetic disturbances to facilitate its devel-opment. First, do subtle differences between the path-ways that are activated by wild-type FLT3 and mutantFLT3 account for the leukemogenic potential of FLT3mutations? Initial evidence indicates that there mightbe biological differences between activated wild-typeand mutant FLT3. If identified, these differencescould be exploited in future treatments. Second, whyare FLT3 mutations common in AML but not B-cellALL (given that normal B cells express FLT3). Doesthis reflect the baseline mitotic or apoptotic activityof myeloid versus lymphoid progenitors, or does itreflect fundamental molecular differences in thepathway that give myeloid cells, but not lymphoidcells, with FLT3 mutations a proliferative advantage?Exploring these curious phenomena might providean insight into the aetiology of FLT3 mutations, aswell as the general problem of cell-type-specificmutations. In addition, a more fundamental under-standing of how various mutations influence inter-related pathways should allow drug targeting at crucial biological ‘checkpoints’. Microarray analyses,which allow the examination of expression of thou-sands of genes simultaneously, will have a crucial rolein showing the similarities and differences in geneexpression due to different mutations. However,many of the complex interactions in the FLT3 path-way are probably not regulated at the level of eithermRNA or protein expression, rather by changes inthe phosphorylation of proteins or complexes ofproteins. A detailed interrogation of such a complexsystem will require methods that examine both geneexpression and protein activation.
The clinical relevance of FLT3 mutations also needsto be explored in more detail in trials with patientsreceiving various treatments. Larger prospective trialswill better clarify the effects of FLT3 mutations inpatients with AML treated with standard chemother-apy or dose-intensification chemotherapy. These stud-ies also need to be expanded to include other treatmentschemes, such as autologous and allogeneic transplan-tation. It will be crucial to couple the FLT3 mutationanalyses to FLT3 expression results and mutationalanalyses of other genes, such as KIT, FMS and RAS, thatact on similar pathways (BOX 2). These studies mightprovide an insight into the relationships between vari-ous mutated genes and begin to genetically characterizeleukaemia, identifying subgroups of patients that canbe prognostically classified on the basis of mutationstatus. As most FLT3 mutations occur in patients withnormal cytogenetics, any prognostic relevance of thesemutations could help dictate future treatment plans.
supported by accompanying molecular studies show-ing that these inhibitors do indeed block activation ofthe mutant receptor. Therefore, there is guarded opti-mism that FLT3 inhibitors might become a successfultherapeutic tool for patients with leukaemia, particu-larly AML. Moreover, the recent discovery that theRTK–RAS signalling pathway is frequently mutated inAML (BOX 2) has promoted the strategy of findinginhibitors for targets further downstream of FLT3. Suchinhibitors might equally block inappropriate activationby either FLT3 or RAS mutations.
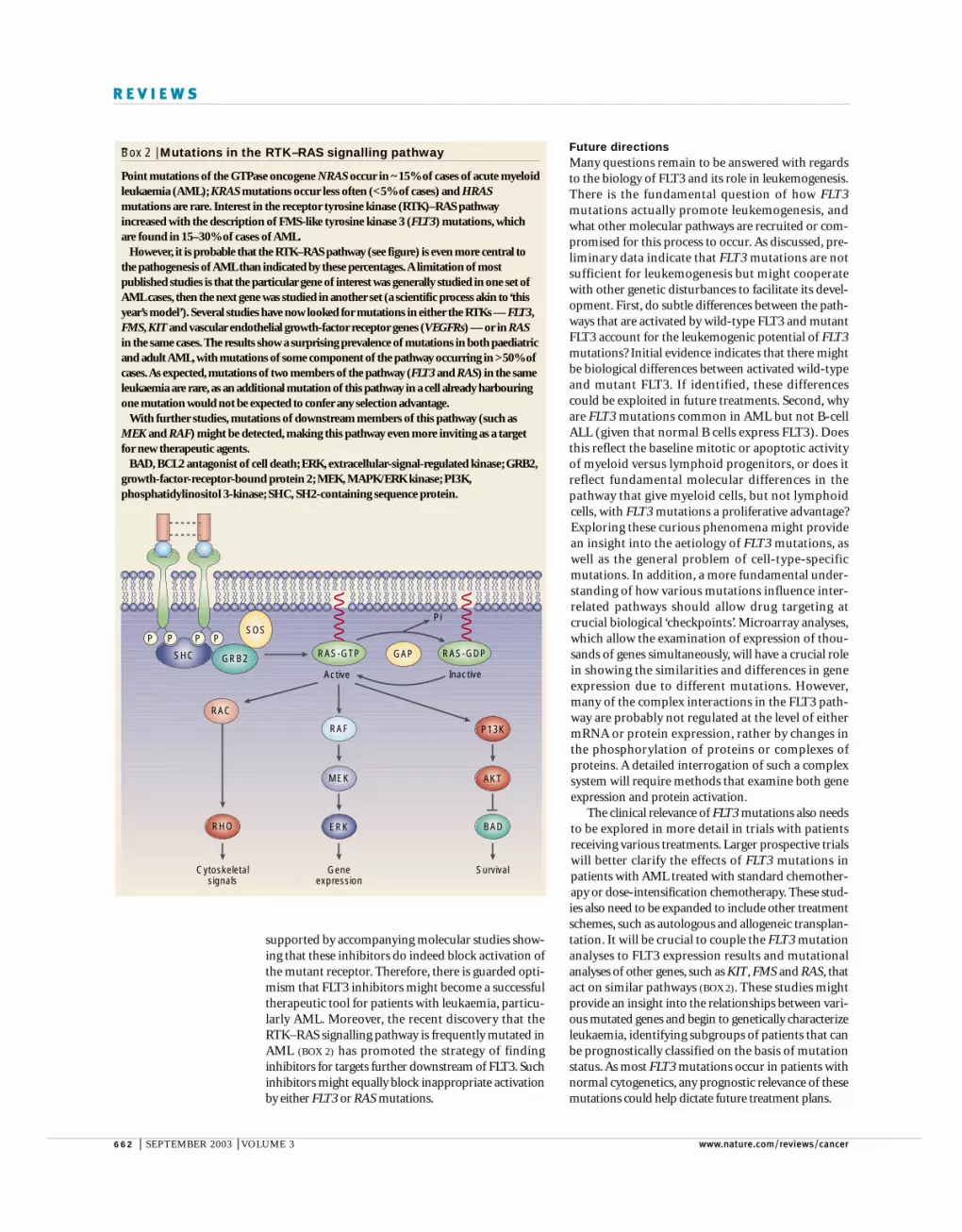
–Box 2 | Mutations in the RTK–RAS signalling pathway
Point mutations of the GTPase oncogene NRAS occur in ~15% of cases of acute myeloidleukaemia (AML); KRAS mutations occur less often (<5% of cases) and HRASmutations are rare. Interest in the receptor tyrosine kinase (RTK)–RAS pathwayincreased with the description of FMS-like tyrosine kinase 3 (FLT3) mutations, whichare found in 15–30% of cases of AML.
However, it is probable that the RTK–RAS pathway (see figure) is even more central tothe pathogenesis of AML than indicated by these percentages.A limitation of mostpublished studies is that the particular gene of interest was generally studied in one set ofAML cases, then the next gene was studied in another set (a scientific process akin to ‘thisyear’s model’). Several studies have now looked for mutations in either the RTKs — FLT3,FMS, KIT and vascular endothelial growth-factor receptor genes (VEGFRs) — or in RASin the same cases. The results show a surprising prevalence of mutations in both paediatricand adult AML, with mutations of some component of the pathway occurring in >50% ofcases.As expected, mutations of two members of the pathway (FLT3 and RAS) in the sameleukaemia are rare, as an additional mutation of this pathway in a cell already harbouringone mutation would not be expected to confer any selection advantage.
With further studies, mutations of downstream members of this pathway (such asMEK and RAF) might be detected, making this pathway even more inviting as a targetfor new therapeutic agents.
BAD, BCL2 antagonist of cell death; ERK, extracellular-signal-regulated kinase; GRB2,growth-factor-receptor-bound protein 2; MEK, MAPK/ERK kinase; PI3K,phosphatidylinositol 3-kinase; SHC, SH2-containing sequence protein.
P P P P
GRB2
Active Inactive
Pi
RAF
MEK
ERK
GAP
RAC
RHO BAD
SOS
Geneexpression
Cytoskeletalsignals
Survival
RAS-GDPRAS-GTPSHC
P13K
AKT

NATURE REVIEWS | CANCER VOLUME 3 | SEPTEMBER 2003 | 663
R E V I E W S
Cancer is a genetic disease. It is unlikely, however,that every defined mutation will have a unique prognos-tic impact, cause a singular aberration in a cellular path-way or require a specific targeted therapy. The carefulstudy of specific mutations such as in FLT3, and howthese mutations are different from or similar to otherclasses of mutation, will probably lead to new schemesof cancer classification and new therapies directed atspecific subsets of molecular lesion.
Finally, new TKIs are being developed rapidly.These need to be quickly screened in pre-clinical tri-als, choosing those that have the greatest potential forsuccess, and then moved rapidly to Phase I trials.Ultimately, the pre-clinical biological data availableso far indicate that TKIs alone might not be sufficientfor the treatment of leukaemia; therefore, the use ofcombinations of TKIs with other agents will probablyneed to be pursued.
1. Matthews, W., Jordan, C. T., Wiegand, G. W., Pardoll, D. &Lemischka, I. R. A receptor tyrosine kinase specific tohematopoietic stem and progenitor cell-enrichedpopulations. Cell 65, 1143–1152 (1991).
2. Rosnet, O., Marchetto, S., deLapeyriere, O. & Birnbaum, D.Murine Flt3, a gene encoding a novel tyrosine kinasereceptor of the PDGFR/CSF1R family. Oncogene 6,1641–1650 (1991).References 1 and 2 describe the initial cloning andexpression of the mouse Fms-like tyrosine kinase 3(Flt3) gene.
3. Rosnet, O. et al. Human FLT3/FLK2 gene: cDNA cloningand expression in hematopoietic cells. Blood 82,1110–1119 (1993).
4. Small, D. et al. STK-1, the human homolog of Flk-2/Flt-3, isselectively expressed in CD34+ human bone marrow cellsand is involved in the proliferation of early progenitor/stemcells. Proc. Natl Acad. Sci. USA 91, 459–463 (1994).References 3 and 4 describe the initial cloning andexpression of the human FLT3 gene.
5. Rosnet, O. et al. Close physical linkage of the FLT1 andFLT3 genes on chromosome 13 in man and chromosome 5in mouse. Oncogene 8, 173–179 (1993).
6. Agnes, F. et al. Genomic structure of the downstream part ofthe human FLT3 gene: exon/intron structure conservationamong genes encoding receptor tyrosine kinases (RTK) ofsubclass III. Gene 145, 283–288 (1994).
7. Rosnet, O. & Birnbaum, D. Hematopoietic receptors ofclass III receptor-type tyrosine kinases. Crit. Rev. Oncog. 4,595–613 (1993).
8. Abu-Duhier, F. M. et al. Genomic structure of human FLT3:implications for mutational analysis. Br. J. Haematol. 113,1076–1077 (2001).References 6 and 8 discuss the exon and intronstructure of FLT3, comparing FLT3 with othermembers of the receptor tyrosine kinase subclass IIIfamily.
9. Lyman, S. D. et al. Characterization of the protein encodedby the flt3 (flk2) receptor-like tyrosine kinase gene.Oncogene 8, 815–822 (1993).
10. Carow, C. E. et al. Expression of the hematopoietic growthfactor receptor FLT3 (STK-1/Flk2) in human leukemias.Blood 87, 1089–1096 (1996).
11. Rosnet, O. et al. Human FLT3/FLK2 receptor tyrosine kinaseis expressed at the surface of normal and malignanthematopoietic cells. Leukemia 10, 238–248 (1996).
12. Gabbianelli, M. et al. Multi-level effects of flt3 ligand onhuman hematopoiesis: expansion of putative stem cells andproliferation of granulomonocytic progenitors/monocyticprecursors. Blood 86, 1661–1670 (1995).
13. Ratajczak, M. Z. et al. FLT3/FLK-2 (STK-1) ligand does notstimulate human megakaryopoiesis in vitro. Stem Cells 14,146–150 (1996).
14. Hjertson, M., Sundstrom, C., Lyman, S. D., Nilsson, K. &Nilsson, G. Stem cell factor, but not flt3 ligand, inducesdifferentiation and activation of human mast cells. Exp.Hematol. 24, 748–754 (1996).
15. Lyman, S. D. & Jacobsen, S. E. c-kit ligand and Flt3 ligand:stem/progenitor cell factors with overlapping yet distinctactivities. Blood 91, 1101–1134 (1998).An excellent review that outlines the cloning,structure, biology and possible clinical implications ofFLT3 ligand (FLT3L). Comparisons between FLT3L andKIT ligand are made, which help to furthercharacterize the importance of FLT3L.
16. Sabbath, K. D., Ball, E. D., Larcom, P., Davis, R. B. & Griffin,J. D. Heterogeneity of clonogenic cells in acute myeloblasticleukemia. J. Clin. Invest. 75, 746–753 (1985).
17. Nadler, L. M. et al. B cell origin of non-T cell acutelymphoblastic leukemia. A model for discrete stages ofneoplastic and normal pre-B cell differentiation. J. Clin.Invest. 74, 332–340 (1984).
18. Rubartelli, A., Sitia, R., Zicca, A., Grossi, C. E. & Ferrarini, M.Differentiation of chronic lymphocytic leukemia cells:
correlation between the synthesis and secretion ofimmunoglobulins and the ultrastructure of the malignantcells. Blood 62, 495–504 (1983).
19. Martin, P. J. et al. Involvement of the B-lymphoid system inchronic myelogenous leukaemia. Nature 287, 49–50 (1980).
20. Uckun, F. M. et al. Clinical features and treatment outcomeof childhood T-lineage acute lymphoblastic leukemiaaccording to the apparent maturational stage of T-lineageleukemic blasts: a Children’s Cancer Group study. J. Clin.Oncol. 15, 2214–2221 (1997).
21. Armstrong, S. A. et al. MLL translocations specify a distinctgene expression profile that distinguishes a unique leukemia.Nature Genet. 30, 41–47 (2002).
22. Lyman, S. D. et al. Molecular cloning of a ligand for theflt3/flk-2 tyrosine kinase receptor: a proliferative factor forprimitive hematopoietic cells. Cell 75, 1157–1167 (1993).
23. Hannum, C. et al. Ligand for FLT3/FLK2 receptor tyrosinekinase regulates growth of haematopoietic stem cells and isencoded by variant RNAs. Nature 368, 643–648 (1994).
24. Lyman, S. D. et al. Cloning of the human homologue of themurine flt3 ligand: a growth factor for early hematopoieticprogenitor cells. Blood 83, 2795–2801 (1994).
25. Lyman, S. D. et al. Structural analysis of human and murineflt3 ligand genomic loci. Oncogene 11, 1165–1172 (1995).
26. Lyman, S. D. et al. Identification of soluble and membrane-bound isoforms of the murine flt3 ligand generated byalternative splicing of mRNAs. Oncogene 10, 149–157(1995).
27. Lyman, S. D., Brasel, K., Rousseau, A. M. & Williams, D. E.The flt3 ligand: a hematopoietic stem cell factor whoseactivities are distinct from steel factor. Stem Cells 12 (Suppl. 1),99–107; 108–110 (1994).
28. Brasel, K. et al. Expression of the flt3 receptor and its ligandon hematopoietic cells. Leukemia 9, 1212–1218 (1995).
29. Meierhoff, G. et al. Expression of FLT3 receptor and FLT3-ligand in human leukemia–lymphoma cell lines. Leukemia 9,1368–1372 (1995).
30. Spagnoli, G. C. et al. FLT3 ligand gene expression andprotein production in human colorectal cancer cell lines andclinical tumor specimens. Int. J. Cancer 86, 238–243(2000).
31. Gonfloni, S., Weijland, A., Kretzschmar, J. & Superti-Furga, G.Crosstalk between the catalytic and regulatory domainsallows bidirectional regulation of Src. Nature Struct. Biol. 7,281–286 (2000).
32. Turner, A. M., Lin, N. L., Issarachai, S., Lyman, S. D. &Broudy, V. C. FLT3 receptor expression on the surface ofnormal and malignant human hematopoietic cells. Blood 88,3383–3390 (1996).
33. Weiss, A. & Schlessinger, J. Switching signals on or off byreceptor dimerization. Cell 94, 277–280 (1998).
34. Yee, N. S., Langen, H. & Besmer, P. Mechanism of kit ligand,phorbol ester, and calcium-induced down-regulation of c-kitreceptors in mast cells. J. Biol. Chem. 268, 14189–14201(1993).
35. Lyman, S. D. et al. Plasma/serum levels of flt3 ligand are lowin normal individuals and highly elevated in patients withFanconi anemia and acquired aplastic anemia. Blood 86,4091–4096 (1995).
36. Ashihara, E. et al. FLT-3 ligand mobilizes hematopoieticprimitive and committed progenitor cells into blood in mice.Eur. J. Haematol. 60, 86–92 (1998).
37. Rogers, S. Y., Bradbury, D., Kozlowski, R. & Russell, N. H.Evidence for internal autocrine regulation of growth in acutemyeloblastic leukemia cells. Exp. Hematol. 22, 593–598(1994).
38. Dosil, M., Wang, S. & Lemischka, I. R. Mitogenic signallingand substrate specificity of the Flk2/Flt3 receptor tyrosinekinase in fibroblasts and interleukin-3-dependenthematopoietic cells. Mol. Cell. Biol. 13, 6572–6585 (1993).
39. Rottapel, R. et al. Substrate specificities and identification ofa putative binding site for PI3K in the carboxy tail of themurine Flt3 receptor tyrosine kinase. Oncogene 9,1755–1765 (1994).
References 38 and 39 were two of the first papers todescribe the downstream effectors of wild-type FLT3,using a chimeric receptor.
40. Marchetto, S. et al. SHC and SHIP phosphorylation andinteraction in response to activation of the FLT3 receptor.Leukemia 13, 1374–1382 (1999).
41. Zhang, S., Mantel, C. & Broxmeyer, H. E. Flt3 signalinginvolves tyrosyl-phosphorylation of SHP-2 and SHIP andtheir association with Grb2 and Shc in Baf3/Flt3 cells. J. Leukocyte Biol. 65, 372–380 (1999).
42. Lavagna-Sevenier, C., Marchetto, S., Birnbaum, D. &Rosnet, O. FLT3 signaling in hematopoietic cells involvesCBL, SHC and an unknown P115 as prominent tyrosine-phosphorylated substrates. Leukemia 12, 301–310 (1998).
43. Zhang, S. & Broxmeyer, H. E. Flt3 ligand induces tyrosinephosphorylation of gab1 and gab2 and their associationwith shp-2, grb2, and PI3 kinase. Biochem. Biophys. Res.Commun. 277, 195–199 (2000).
44. Srinivasa, S. P. & Doshi, P. D. Extracellular signal-regulatedkinase and p38 mitogen-activated protein kinase pathwayscooperate in mediating cytokine-induced proliferation of aleukemic cell line. Leukemia 16, 244–253 (2002).
45. Zhang, S. & Broxmeyer, H. E. p85 subunit of PI3 kinasedoes not bind to human Flt3 receptor, but associates withSHP2, SHIP, and a tyrosine-phosphorylated 100-kDaprotein in Flt3 ligand-stimulated hematopoietic cells.Biochem. Biophys. Res. Commun. 254, 440–445 (1999).
46. Zhang, S. et al. Essential role of signal transducer andactivator of transcription (Stat)5a but not Stat5b for Flt3-dependent signaling. J. Exp. Med. 192, 719–728 (2000).
47. Lisovsky, M. et al. Flt3 ligand stimulates proliferation andinhibits apoptosis of acute myeloid leukemia cells: regulationof Bcl-2 and Bax. Blood 88, 3987–3997 (1996).
48. Radich, J. P. et al. N-ras mutations in adult de novo acutemyelogenous leukemia: prevalence and clinical significance.Blood 76, 801–807 (1990).
49. Nakagawa, T. et al. Multiple point mutation of N-ras and K-ras oncogenes in myelodysplastic syndrome and acutemyelogenous leukemia. Oncology 49, 114–122 (1992).
50. Bustelo, X. R., Ledbetter, J. A. & Barbacid, M. Product ofvav proto-oncogene defines a new class of tyrosine proteinkinase substrates. Nature 356, 68–71 (1992).
51. Ray, R. J., Paige, C. J., Furlonger, C., Lyman, S. D. &Rottapel, R. Flt3 ligand supports the differentiation of early B cell progenitors in the presence of interleukin-11 andinterleukin-7. Eur. J. Immunol. 26, 1504–1510 (1996).
52. Veiby, O. P., Lyman, S. D. & Jacobsen, S. E. Combinedsignaling through interleukin-7 receptors and flt3 but not c-kit potently and selectively promotes B-cell commitmentand differentiation from uncommitted murine bone marrowprogenitor cells. Blood 88, 1256–1265 (1996).
53. Rusten, L. S., Lyman, S. D., Veiby, O. P. & Jacobsen, S. E.The FLT3 ligand is a direct and potent stimulator of the growthof primitive and committed human CD34+ bone marrowprogenitor cells in vitro. Blood 87, 1317–1325 (1996).
54. Shah, A. J., Smogorzewska, E. M., Hannum, C. & Crooks, G. M.Flt3 ligand induces proliferation of quiescent human bonemarrow CD34+CD38– cells and maintains progenitor cells invitro. Blood 87, 3563–3570 (1996).
55. Namikawa, R., Muench, M. O., de Vries, J. E. & Roncarolo,M. G. The FLK2/FLT3 ligand synergizes with interleukin-7 inpromoting stromal-cell-independent expansion anddifferentiation of human fetal pro-B cells in vitro. Blood 87,1881–1890 (1996).
56. Moore, T. A. & Zlotnik, A. Differential effects of Flk-2/Flt-3ligand and stem cell factor on murine thymic progenitorcells. J. Immunol. 158, 4187–4192 (1997).
57. Mackarehtschian, K. et al. Targeted disruption of the flk2/flt3gene leads to deficiencies in primitive hematopoieticprogenitors. Immunity 3, 147–161 (1995).
58. Saunders, D. et al. Dendritic cell development in culture fromthymic precursor cells in the absence ofgranulocyte/macrophage colony-stimulating factor. J. Exp.Med. 184, 2185–2196 (1996).

664 | SEPTEMBER 2003 | VOLUME 3 www.nature.com/reviews/cancer
R E V I E W S
59. Maraskovsky, E. et al. Dramatic increase in the numbers offunctionally mature dendritic cells in Flt3 ligand-treated mice:multiple dendritic cell subpopulations identified. J. Exp. Med.184, 1953–1962 (1996).
60. Morse, M. A. et al. Preoperative mobilization of circulatingdendritic cells by Flt3 ligand administration to patients withmetastatic colon cancer. J. Clin. Oncol. 18, 3883–3893(2000).
61. Manfra, D. J. et al. Conditional expression of murine flt3ligand leads to expansion of multiple dendritic cell subsets inperipheral blood and tissues of transgenic mice. J. Immunol.170, 2843–2852 (2003).
62. Lynch, D. H. et al. Flt3 ligand induces tumor regression andantitumor immune responses in vivo. Nature Med. 3,625–631 (1997).
63. Chen, K. et al. Antitumor activity and immunotherapeuticproperties of Flt3-ligand in a murine breast cancer model.Cancer Res. 57, 3511–3516 (1997).
64. Chakravarty, P. K. et al. Flt3-ligand administration afterradiation therapy prolongs survival in a murine model ofmetastatic lung cancer. Cancer Res. 59, 6028–6032(1999).
65. Pawlowska, A. B. et al. In vitro tumor-pulsed or in vivo Flt3ligand-generated dendritic cells provide protection againstacute myelogenous leukemia in nontransplanted orsyngeneic bone marrow-transplanted mice. Blood 97,1474–1482 (2001).
66. Parajuli, P. et al. Immunization with wild-type p53 genesequences coadministered with Flt3 ligand induces anantigen-specific type 1 T-cell response. Cancer Res. 61,8227–8234 (2001).
67. Disis, M. L. et al. Flt3 ligand as a vaccine adjuvant inassociation with HER-2/neu peptide-based vaccines inpatients with HER-2/neu-overexpressing cancers. Blood 99,2845–2850 (2002).
68. Fong, L. et al. Altered peptide ligand vaccination with Flt3ligand expanded dendritic cells for tumor immunotherapy.Proc. Natl Acad. Sci. USA 98, 8809–8814 (2001).
69. Dehmel, U. et al. Effects of FLT3 ligand on human leukemiacells. I. Proliferative response of myeloid leukemia cells.Leukemia 10, 261–270 (1996).
70. Nakao, M. et al. Internal tandem duplication of the flt3 genefound in acute myeloid leukemia. Leukemia 10, 1911–1918(1996).The first article to describe ITDs of FLT3 in leukaemia,leading the way for future articles that examined theprognostic impact of FLT3 mutations.
71. Kiyoi, H. et al. Internal tandem duplication of the FLT3 geneis a novel modality of elongation mutation which causesconstitutive activation of the product. Leukemia 12,1333–1337 (1998).One of the first papers to show that FLT3 ITDs causeconstitutive activation and ligand-independent growthin mouse transduction models. It led to the use ofmouse transduction models for molecular-biologyinvestigations of the mutated receptor and potentialFLT3 inhibitors.
72. Kiyoi, H. et al. Internal tandem duplication of FLT3associated with leukocytosis in acute promyelocyticleukemia. Leukemia Study Group of the Ministry of Healthand Welfare (Kohseisho). Leukemia 11, 1447–1452(1997).
73. Stirewalt, D. L. et al. FLT3, RAS and TP53 mutations inelderly patients with acute myeloid leukemia. Blood 97,3589–3595 (2001).The only study to examine specifically the prognosticimpact of FLT3 ITDs in older adults with AML, whichshowed that single-strand conformational analyseswere more sensitive than previously published meansfor detecting FLT3 ITDs.
74. Meshinchi, S. et al. Prevalence and prognostic significanceof FLT3 internal tandem duplication in pediatric acutemyeloid leukemia. Blood 97, 89–94 (2001).
75. Schnittger, S. et al. Analysis of FLT3 length mutations in1003 patients with acute myeloid leukemia: correlationto cytogenetics, FAB subtype, and prognosis in theAMLCG study and usefulness as a marker for thedetection of minimal residual disease. Blood 100, 59–66(2002).
76. Thiede, C. et al. Analysis of FLT3-activating mutations in 979patients with acute myelogenous leukemia: association withFAB subtypes and identification of subgroups with poorprognosis. Blood 99, 4326–4335 (2002).References 74–76 are three of the largest studies toexamine FLT3 ITDs in paediatric and adult AML.
77. Abu-Duhier, F. M. et al. FLT3 internal tandem duplicationmutations in adult acute myeloid leukaemia define a high-risk group. Br. J. Haematol. 111, 190–195 (2000).
78. Horiike, S. et al. Tandem duplications of the FLT3 receptorgene are associated with leukemic transformation ofmyelodysplasia. Leukemia 11, 1442–1446 (1997).
79. Xu, F. et al. Tandem duplication of the FLT3 gene is found inacute lymphoblastic leukaemia as well as acute myeloidleukaemia but not in myelodysplastic syndrome or juvenilechronic myelogenous leukaemia in children. Br. J. Haematol.105, 155–162 (1999).
80. Kiyoi, H. et al. Prognostic implication of FLT3 and N-RASgene mutations in acute myeloid leukemia. Blood 93,3074–3080 (1999).
81. Kottaridis, P. D. et al. The presence of a FLT3 internaltandem duplication in patients with acute myeloid leukemia(AML) adds important prognostic information to cytogeneticrisk group and response to the first cycle of chemotherapy:analysis of 854 patients from the United Kingdom MedicalResearch Council AML 10 and 12 trials. Blood 98,1752–1759 (2001).
82. Yokota, S. et al. Internal tandem duplication of the FLT3gene is preferentially seen in acute myeloid leukemia andmyelodysplastic syndrome among various hematologicalmalignancies. A study on a large series of patients and celllines. Leukemia 11, 1605–1609 (1997).
83. Yamamoto, Y. et al. Activating mutation of D835 within theactivation loop of FLT3 in human hematologic malignancies.Blood 97, 2434–2439 (2001).
84. Abu-Duhier, F. M. et al. Identification of novel FLT-3 Asp835mutations in adult acute myeloid leukaemia. Br. J. Haematol.113, 983–988 (2001).References 83 and 84 describe new missensemutations in exon 17 (now referred to as exon 20) ofFLT3 in patients with leukaemia.
85. Spiekermann, K. et al. A new and recurrent activating lengthmutation in exon 20 of the FLT3 gene in acute myeloidleukemia. Blood 100, 3423–3425 (2002).
86. Meshinchi, S. et al. Activating mutations of RTK/ras signaltransduction pathway in pediatric acute myeloid leukemia.Blood 102, 1474–1479 (2003).
87. Hayakawa, F. et al. Tandem-duplicated Flt3 constitutivelyactivates STAT5 and MAP kinase and introducesautonomous cell growth in IL-3-dependent cell lines.Oncogene 19, 624–631 (2000).One of the first studies to describe the downstreameffects of constitutively activated FLT3 ITDs.
88. Kiyoi, H., Ohno, R., Ueda, R., Saito, H. & Naoe, T.Mechanism of constitutive activation of FLT3 with internaltandem duplication in the juxtamembrane domain.Oncogene 21, 2555–2563 (2002).This paper showed that FLT3 ITDs could constitutivelyactivate wild-type FLT3, providing an insight into themechanism of action of the mutant receptor.
89. Mizuki, M. et al. Flt3 mutations from patients with acutemyeloid leukemia induce transformation of 32D cellsmediated by the Ras and STAT5 pathways. Blood 96,3907–3914 (2000).
90. Gille, H. et al. A repressor sequence in the juxtamembranedomain of Flt-1 (VEGFR-1) constitutively inhibits vascularendothelial growth factor-dependent phosphatidylinositol 3′-kinase activation and endothelial cell migration. EMBO J.19, 4064–4073 (2000).
91. Zheng, R., Friedman, A. D. & Small, D. Targeted inhibition ofFLT3 overcomes the block to myeloid differentiation in32Dcl3 cells caused by expression of FLT3/ITD mutations.Blood 100, 4154–4161 (2002).
92. Kelly, L. M. et al. FLT3 internal tandem duplicationmutations associated with human acute myeloidleukemias induce myeloproliferative disease in a murinebone marrow transplant model. Blood 99, 310–318(2002).
93. Brown, D. et al. A PMLRARα transgene initiates murineacute promyelocytic leukemia. Proc. Natl Acad. Sci. USA94, 2551–2556 (1997).This study eloquently examines the molecular biologyand function of FLT3 ITDs, through blocking theireffects in transduction models. The results show thatFLT3 ITDs probably affect many different cellularprocesses, including differentiation, which had notbeen investigated extensively previously.
94. Grisolano, J. L., Wesselschmidt, R. L., Pelicci, P. G. & Ley, T. J.Altered myeloid development and acute leukemia in transgenicmice expressing PML-RARα under control of cathepsin Gregulatory sequences. Blood 89, 376–387 (1997).
95. Kelly, L. M. et al. PML/RARα and FLT3-ITD induce an APL-like disease in a mouse model. Proc. Natl Acad. Sci. USA99, 8283–8288 (2002).This paper shows that the combination of an FLT3 ITDand t(15;17) causes a leukaemia-like phenotype inmice, which supports the theory that multiplegenomic abnormalities might be necessary in mostleukaemia cells.
96. Knudson, A. G. Jr, Hethcote, H. W. & Brown, B. W. Mutationand childhood cancer: a probabilistic model for theincidence of retinoblastoma. Proc. Natl Acad. Sci. USA 72,5116–5120 (1975).
97. Mizuki, M. et al. Suppression of myeloid transcriptionfactors and induction of STAT response genes by AML-specific Flt3 mutations. Blood 101, 3164–3173(2002).
98. Beghini, A. et al. C-kit mutations in core binding factorleukemias. Blood 95, 726–727 (2000).
99. Sperr, W. R. et al. Systemic mastocytosis associated withacute myeloid leukaemia: report of two cases and detectionof the c-kit mutation Asp-816 to Val. Br. J. Haematol. 103,740–749 (1998).
100. Tsujimura, T. et al. Ligand-independent activation of c-kitreceptor tyrosine kinase in a murine mastocytoma cell lineP-815 generated by a point mutation. Blood 83, 2619–2626(1994).
101. Morley, G. M., Uden, M., Gullick, W. J. & Dibb, N. J. Cellspecific transformation by c-fms activating loop mutations isattributable to constitutive receptor degradation. Oncogene18, 3076–3084 (1999).
102. Moriyama, Y. et al. Role of aspartic acid 814 in the functionand expression of c-kit receptor tyrosine kinase. J. Biol.Chem. 271, 3347–3350 (1996).
103. Iwai, T. et al. Internal tandem duplication of the FLT3 geneand clinical evaluation in childhood acute myeloid leukemia.The Children’s Cancer and Leukemia Study Group, Japan.Leukemia 13, 38–43 (1999).
104. Kondo, M. et al. Prognostic value of internal tandemduplication of the FLT3 gene in childhood acutemyelogenous leukemia. Med. Pediatr. Oncol. 33, 525–529(1999).
105. Libura, M. et al. FLT3 and MLL intragenic abnormalities inAML reflect a common category of genotoxic stress. BloodJun 5 2003 (doi: 10.1182/blood-2003-01-0162).This paper was the first to examine the possiblecorrelations between MLL abnormalities andexpression of FLT3 in patient samples.
106. Shih, L. Y. et al. Internal tandem duplication of FLT3 inrelapsed acute myeloid leukemia: a comparativeanalysis of bone marrow samples from 108 adultpatients at diagnosis and relapse. Blood 100,2387–2392 (2002).
107. Alsabeh, R., Brynes, R. K., Slovak, M. L. & Arber, D. A.Acute myeloid leukemia with t(6;9) (p23;q34): associationwith myelodysplasia, basophilia, and initial CD34 negativeimmunophenotype. Am. J. Clin. Pathol. 107, 430–437(1997).
108. Noguera, N. I. et al. Alterations of the FLT3 gene in acutepromyelocytic leukemia: association with diagnosticcharacteristics and analysis of clinical outcome in patientstreated with the Italian AIDA protocol. Leukemia 16,2185–2189 (2002).
109. Whitman, S. P. et al. Absence of the wild-type allele predictspoor prognosis in adult de novo acute myeloid leukemiawith normal cytogenetics and the internal tandemduplication of FLT3: a cancer and leukemia group B study.Cancer Res. 61, 7233–7239 (2001).One of the first studies to indicate that the ITD:wild-type ratio of FLT3 might have prognostic impact inpatients with AML.
110. Armstrong, S. A. et al. Inhibition of FLT3 in MLL.Validation of a therapeutic target identified by geneexpression based classification. Cancer Cell. 3, 173–183(2003).This study was the first to show that inhibition ofFLT3 over-expression in a cell with MLLabnormalities could block the growth of thesecells, and it provided the rationale for examiningFLT3 inhibitors in other leukaemia cells that over-express FLT3.
111. Druker, B. J. & Lydon, N. B. Lessons learned from thedevelopment of an abl tyrosine kinase inhibitor forchronic myelogenous leukemia. J. Clin. Invest. 105, 3–7(2000).
112. Zhao, M. et al. In vivo treatment of mutant FLT3-transformedmurine leukemia with a tyrosine kinase inhibitor. Leukemia14, 374–378 (2000).
113. Levis, M., Tse, K. F., Smith, B. D., Garrett, E. & Small, D.A FLT3 tyrosine kinase inhibitor is selectively cytotoxic toacute myeloid leukemia blasts harboring FLT3 internaltandem duplication mutations. Blood 98, 885–887(2001).References 112 and 113 were the first to show thatactivation-mutant FLT3 could be blocked withtyrosine-kinase inhibitors, and that this inhibitioncaused apoptosis.
114. Tse, K. F. et al. Inhibition of the transforming activity of FLT3internal tandem duplication mutants from AML patients bya tyrosine kinase inhibitor. Leukemia 16, 2027–2036(2002).
115. Minami, Y. et al. Selective apoptosis of tandemly duplicatedFLT3-transformed leukemia cells by Hsp90 inhibitors.Leukemia 16, 1535–1540 (2002).

NATURE REVIEWS | CANCER VOLUME 3 | SEPTEMBER 2003 | 665
R E V I E W S
116. Levis, M. et al. A FLT3-targeted tyrosine kinase inhibitor iscytotoxic to leukemia cells in vitro and in vivo. Blood 99,3885–3891 (2002).
117. Levis, M. et al. FLT3-targeted inhibitors kill FLT3-dependent modeled cells, leukemia-derived cell lines, andprimary AML blasts in vitro and in vivo. Blood 89, 721a(2001).
118. Smith, B. D. et al. Single agent CEP-701, a novel FLT-3inhibitor, shows initial response in patients withrefactory acute myeloid leukemia. Blood 100, 314a(2002).
119. O’Farrell, A. M. et al. SU11248 is a novel FLT3 tyrosinekinase inhibitor with potent activity in vitro and in vivo. Blood101, 3597–3605 (2003).
120. Yee, K. W. et al. SU5416 and SU5614 inhibit kinase activityof wild-type and mutant FLT3 receptor tyrosine kinase.Blood 100, 2941–2949 (2002).
121. Foran, J. et al. An innovative single dose clinical studyshows potent inhibition of FLT3 phosphorylation bySU11248 in vivo: a clinical and pharmacodynamic studyin AML patients. Blood 100, 2196a (2002).
122. Giles, F. J. et al. SU5416, a small molecule tyrosinekinase receptor inhibitor, has biologic activity in patientswith refractory acute myeloid leukemia ormyelodysplastic syndromes. Blood 102, 795–801(2003).One of the first articles to show inhibition of growth and induction of apoptosis in FLT3 ITD-transducedcells using more-selective tyrosine-kinase inhibitors.
123. Foran, J. et al. A phase I study of repeated oral dosingwith SU11248 for the treatment of patients with acutemyeloid leukemia who have failed, or are not eligible forconvential chemotherapy. Blood 100, 2195a (2002).
124. Kelly, L. M. et al. CT53518, a novel selective FLT3 antagonistfor the treatment of acute myelogenous leukemia (AML).Cancer Cell 1, 421–432 (2002).
125. Heinrich, M. C. et al. A “first in man” study of the safety andPK/PD of an oral FLT3 inhibitor (MLN518) in patients withAML or high risk myelodysplasia. Blood 100, 1305a(2002).
126. Weisberg, E. et al. Inhibition of mutant FLT3 receptors inleukemia cells by the small molecule tyrosine kinase inhibitorPKC412. Cancer Cell 1, 433–443 (2002).
127. Stone, R. M. et al. PKC412, an oral FLT3 inhibitor, hasactivity in mutant FLT3 acute myeloid leukemia (AML): aphase II clinical trial. Blood 100, 316a (2002).
AcknowledgementsThis work was supported by grants from the National Institutesof Health.
Online links
DATABASESThe following terms in this article are linked online to:Cancer.gov: http://www.cancer.gov/search/leukaemiaLocusLink: http://www.ncbi.nlm.nih.gov/LocusLink/ABL | BAX | BCR | CBL | CBLB | CSF1 | EPO | FLT3 | Flt3 | FLT3L |Flt3l | GAB2 | G-CSF | GM-CSF | Grb2 | HER2/NEU | HSP90 | IL-3| IL-6 | IL-7 | IL-11 | FMS | KIT | KIT ligand | MLL | p85 | PDGFRA |PDGFRB | Plcγ1 | PML | RARA | Ship | SHP2 | STAT5A | Vav |VEGFR1 | VEGFR2Access to this interactive links box is free online.

O N L I N E
BiographyDerek L. Stirewalt and Jerald P. Radich are members of the ClinicalResearch Division at the Fred Hutchinson Cancer Research Center. TheRadich laboratory is interested in the genetics of leukaemia, with partic-ular interest in the interface of genetics and patient care. Much of theirwork has been on the detection of minimal residual disease as a predic-tor of relapse in patients with acute and chronic leukaemia. Morerecently, their work has focused on the gene-expression profiles that areassociated with response and progression in leukaemia. In addition, thelab is active in the area of signal-transduction changes in acuteleukaemia, especially stemming from mutations in the FLT3 gene.
Linksleukaemiahttp://www.cancer.gov/cancerinfo/types/leukemia
ABLhttp://www.ncbi.nlm.nih.gov/LocusLink/LocRpt.cgi?l=25BAXhttp://www.ncbi.nlm.nih.gov/LocusLink/LocRpt.cgi?l=581BCRhttp://www.ncbi.nlm.nih.gov/LocusLink/LocRpt.cgi?l=613CBLhttp://www.ncbi.nlm.nih.gov/LocusLink/LocRpt.cgi?l=867CBLBhttp://www.ncbi.nlm.nih.gov/LocusLink/LocRpt.cgi?l=868CSF1http://www.ncbi.nlm.nih.gov/LocusLink/LocRpt.cgi?l=1435EPOhttp://www.ncbi.nlm.nih.gov/LocusLink/LocRpt.cgi?l=2056FLT3http://www.ncbi.nlm.nih.gov/LocusLink/LocRpt.cgi?l=2322Flt3http://www.ncbi.nlm.nih.gov/LocusLink/LocRpt.cgi?l=14255FLT3Lhttp://www.ncbi.nlm.nih.gov/LocusLink/LocRpt.cgi?l=2323Flt3lhttp://www.ncbi.nlm.nih.gov/LocusLink/LocRpt.cgi?l=14256GAB2http://www.ncbi.nlm.nih.gov/LocusLink/LocRpt.cgi?l=9846G-CSFhttp://www.ncbi.nlm.nih.gov/LocusLink/LocRpt.cgi?l=1440GM-CSFhttp://www.ncbi.nlm.nih.gov/LocusLink/LocRpt.cgi?l=1437Grb2http://www.ncbi.nlm.nih.gov/LocusLink/LocRpt.cgi?l=14784HER2/NEUhttp://www.ncbi.nlm.nih.gov/LocusLink/LocRpt.cgi?l=2064HSP90http://www.ncbi.nlm.nih.gov/LocusLink/list.cgi?/&Q=hspca[sym]%20or%20hspcb[sym]&ORG=HsIL-3http://www.ncbi.nlm.nih.gov/LocusLink/LocRpt.cgi?l=3562IL-6http://www.ncbi.nlm.nih.gov/LocusLink/LocRpt.cgi?l=3569IL-7http://www.ncbi.nlm.nih.gov/LocusLink/LocRpt.cgi?l=3574IL-11http://www.ncbi.nlm.nih.gov/LocusLink/LocRpt.cgi?l=3589FMS
http://www.ncbi.nlm.nih.gov/LocusLink/LocRpt.cgi?l=1436KIThttp://www.ncbi.nlm.nih.gov/LocusLink/LocRpt.cgi?l=3815KIT ligandhttp://www.ncbi.nlm.nih.gov/LocusLink/LocRpt.cgi?l=4254MLLhttp://www.ncbi.nlm.nih.gov/LocusLink/LocRpt.cgi?l=4297p85http://www.ncbi.nlm.nih.gov/LocusLink/list.cgi?/&Q=pik3r1%20or%20pik3r2&ORG=MmPDGFRAhttp://www.ncbi.nlm.nih.gov/LocusLink/LocRpt.cgi?l=5156PDGFRBhttp://www.ncbi.nlm.nih.gov/LocusLink/LocRpt.cgi?l=5159Plcγ1http://www.ncbi.nlm.nih.gov/LocusLink/LocRpt.cgi?l=18803PMLhttp://www.ncbi.nlm.nih.gov/LocusLink/LocRpt.cgi?l=5371RARAhttp://www.ncbi.nlm.nih.gov/LocusLink/LocRpt.cgi?l=5914Shiphttp://www.ncbi.nlm.nih.gov/LocusLink/LocRpt.cgi?l=16331SHP2http://www.ncbi.nlm.nih.gov/LocusLink/LocRpt.cgi?l=5781STAT5Ahttp://www.ncbi.nlm.nih.gov/LocusLink/LocRpt.cgi?l=6776Vavhttp://www.ncbi.nlm.nih.gov/LocusLink/LocRpt.cgi?l=22324VEGFR1http://www.ncbi.nlm.nih.gov/LocusLink/LocRpt.cgi?l=2321VEGFR2http://www.ncbi.nlm.nih.gov/LocusLink/LocRpt.cgi?l=3791