the potential of chitosan-based gels containing intervertebral disc cells for nucleus pulposus...
TRANSCRIPT

ARTICLE IN PRESS
0142-9612/$ - se
doi:10.1016/j.bi
�Correspondfax: +1514 842
E-mail addr
Biomaterials 27 (2006) 388–396
www.elsevier.com/locate/biomaterials
The potential of chitosan-based gels containing intervertebraldisc cells for nucleus pulposus supplementation
Peter Roughleya,�, Caroline Hoemannb, Eric DesRosiersc,Fackson Mwaled, John Antonioud, Mauro Alinie
aGenetics Unit, Shriners Hospital for Children, and Department of Surgery, McGill University, Montreal, CanadabBiomedical and Chemical Engineering, Ecole Polytechnique, Montreal, Canada
cBioSyntech Canada Inc., Laval, CanadadLady David Institute, Jewish General Hospital, and Department of Surgery, McGill University, Montreal, Canada
eOrthopaedic Research Laboratory, McGill University, Montreal, Canada and AO Research Institute, Davos, Switzerland
Received 27 June 2005; accepted 30 June 2005
Available online 24 August 2005
Abstract
The suitability of chitosan-based hydrogels as scaffolds for the encapsulation of intervertebral disc (IVD) cells and the
accumulation of a functional extracellular matrix mimicking that of the nucleus pulposus (NP) was investigated. The specific
hypothesis under study was that the cationic chitosan would form an ideal environment in which large quantities of newly
synthesized anionic proteoglycan could be entrapped. Indeed, all the formulations of cell-seeded chitosan hydrogels, studied under
in vitro culture conditions, showed that the majority of proteoglycan produced by encapsulated NP cells was retained within the gel
rather than released into the culture medium. This was not always the case when annulus fibrosus cells were encapsulated, as unlike
the nucleus cells the annulus cells often did not survive when cultured in chitosan. The results support the concept that chitosan may
be a suitable scaffold for cell-based supplementation to help restore the function of the NP during the early stages of IVD
degeneration.
r 2005 Elsevier Ltd. All rights reserved.
Keywords: Chitin/chitosan; Cell encapsulation; Hydrogel; Intervertebral disc; Nucleus pulposus; Proteoglycan
1. Introduction
The intervertebral discs (IVDs) link adjacent verteb-rae within the cervical, thoracic and lumbar spine. Theyare fibrocartilaginous in nature [1], and provide thespine with its flexibility. Their unique properties alsoallow the mobile spine to resist excessive compression,bending and twisting [2]. In the human, the IVDs areseparated from the vertebral bone by the cartilageendplates, which in the juvenile are responsible forlongitudinal vertebral growth. In the adult, the IVDs are
e front matter r 2005 Elsevier Ltd. All rights reserved.
omaterials.2005.06.037
ing author. Tel.: +1514 282 7156;
5581.
ess: [email protected] (P. Roughley).
essentially avascular structures, which receive most oftheir nutrients from the vasculature of the vertebralbone by diffusion through the endplates. The size of theIVDs increases on progressing down the spine, with thelargest lumbar discs being approximately 1 cm thick and4 cm in diameter. The supply of nutrients to the cells atthe centre of these large discs is readily compromised,and is one factor leading to IVD degeneration [3–5].The IVDs are composed of two major structural
regions—the peripheral annulus fibrosus (AF) and thecentral nucleus pulposus (NP) [5]. The AF is a fibrosustissue consisting of concentric lamellae rich in collagenfibrils. The fibrils of each lamella are oriented parallel toone another and at 601 to the axis of the spine. Inadjacent lamellae, the direction of the collagen fibrils

ARTICLE IN PRESSP. Roughley et al. / Biomaterials 27 (2006) 388–396 389
alternates between left and right. This unique organiza-tion allows the AF to function as an intervertebralligament resisting vertebral motion. The NP has amore amorphous consistency, with randomly orientedcollagen fibrils and a large content of proteoglycan.This composition gives the NP a gelatinous appearancein the juvenile and healthy young adult, and providesit with the ability to resist compressive loads. Thelatter function is intimately related to the proteo-glycan (aggrecan) content of the NP [6]. Aggrecanis a large proteoglycan with numerous glycosamino-glycan (GAG) chains attached to its core protein [7].The high content of GAG in the NP provides theosmotic properties needed for the tissue to swell byimbibition of water and to thereby counter the effects ofcompression.Any event which decreases GAG content in the NP
will impair its functional ability. Such a decrease occursrelatively early during adult human life and may be aprelude to tissue degeneration [8]. During IVD degen-eration the NP becomes more fibrous in nature and thedemarcation between it and the inner AF becomesindistinct [9]. Subsequently, damage to the collagenfibrils results in cleft formation and tissue loss in the NP,which ultimately extends into the AF. This deteriorationof IVD structure and function in the lumbar spine iscommonly associated with low back pain [10,11]. Whilepoor IVD nutrition may be a major contributor to discdegeneration, biomechanical [12–15], biochemical[16–21] and genetic [22–24] influences may also play arole in some individuals.Current treatment of IVD degeneration ultimately
involves surgical excision of the damaged tissue,insertion of a cage or prosthesis to restore theintervertebral space, and vertebral bone fusion [25].While this may provide symptomatic relief, it also altersspine biomechanics and can exacerbate the degenerativeprocess in adjacent spinal segments. Ideally, biologicalrepair of the degenerating IVD would be preferable tosurgical excision, though at present there is noestablished procedure to achieve this result. Certainlythe type of biological repair needed would vary with theextent of IVD degeneration [26,27]. While late stagedegeneration might require complete replacement of thedisc with a tissue engineered construct, repair of theexisting tissue may be possible during early degene-ration. At this time it may be possible to supplementthe NP with additional extracellular matrix compo-nents, particularly aggrecan, to restore its diminishingfunction.One approach to such nuclear supplementation is to
inject cells seeded in a supporting biomolecular scaffoldinto the NP where they can be induced to produce anextracellular matrix rich in aggrecan. One featurerequired by such a scaffold is the ability to retain theaggrecan produced by the cells. Scaffolds that might
offer this potential include those formed from collagen[28,29], agarose [30], alginate [31,32], polyglycolic acid[33] and chitosan. Chitosan is a polymer of glucosamineand N-acetyl glucosamine derived by the partialdepolymerization and deacetylation of chitin extractedfrom the shells of crustaceans [34–36]. It can be inducedto form hydrogels, either by self-association or bycovalent cross-linking [37,38], and has been shown to becompatible with the entrapment of viable chondrocytes[39,40]. The cationic nature of the chitosan gel makes itan attractive scaffold to facilitate the entrapment of thehighly anionic aggrecan produced by chondrocytes orcells of the IVD.In the present work, cells from the bovine IVD were
encapsulated in chitosan-based gels and maintained intissue culture, in order to establish whether chitosan iscapable of retaining the aggrecan produced by the cellsand whether AF cells can be induced to behave in asimilar manner to NP cells.
2. Methods
2.1. Source of intervertebral disc
Coccygeal IVDs were obtained from the tails of adult
bovine steers (2–4 years old). They were used for cell isolation
immediately after transportation to the laboratory from the
abattoir.
2.2. Isolation of disc cells
IVDs were dissected from the adjacent vertebral bodies, and
placed in Dulbecco’s modified Eagle medium (DMEM)
containing 44mM NaHCO3, 20mM HEPES buffer, 50 units/
ml penicillin G, 50 mg/ml streptomycin sulfate (all from Gibco
BRL), 150 mg/ml gentamycin sulfate (Sigma), and 2.5 mg/mlamphotericin B (fungizone, Gibco BRL). IVDs were separated
into AF and NP regions, which were divided into approxi-
mately 2mm pieces, and cells were then enzymatically isolated
from the tissue pieces. Tissue was incubated at 37 1C for
90min in serum-free DMEM, containing 0.2% pronase (from
Streptomyces griseus, Calbiochem-Novabiochem) and 0.004%
deoxyribonuclease I (DNase type II from bovine pancreas;
Sigma) at 2.5ml/g tissue. Tissue was then digested at 5ml/g
in DMEM/10% fetal calf serum (FCS; Fetal Clone III
from Hyclone Laboratories), containing 0.04% collagenase
II (from Clostridium histolyticum), 0.01% hyaluronidase (type
V from sheep testes) and 0.004% DNase (all from Sigma) for
AF, or 0.01% collagenase IA (from C. histolyticum), 0.01%
hyaluronidase and 0.004% DNase for NP. Digestion was
carried out with stirring at 37 1C for 9 h, after which digests
were filtered through a 70mm nylon sieve and cells were
recovered by centrifugation at 400g for 6min. Yields were
approximately 3� 106 cells/g tissue for both AF and NP. Cell
viability varied with each preparation, but was always in the
range 85–95% for both cell types, as measured by trypan blue
exclusion.

ARTICLE IN PRESSP. Roughley et al. / Biomaterials 27 (2006) 388–396390
2.3. Source of chitosan
Two sources of chitosan were used in this work. UltrasanTM,
ultrapure chitosan in free base form (MW 1125kDa, 79% or
95% deacetylated) was supplied by BioSyntech (Laval, Canada).
Protasans UP G213, a glutamate salt of chitosan (MW
300kDa, 85% deacetylated) was obtained from FMC Biopoly-
mer A.S. (Oslo, Norway). Different formulations of chitosan
gels were used in the present work to ensure that results were
independent of chitosan source and mode of cross-linking.
2.4. Encapsulation of cells in chitosan/glycerophosphate
(chitosan-GP) gels
A thermogelling chitosan solution was prepared by mixing
solutions of disodium b-glycerophosphate (GP) and Ultrasan
(95% deacetylated) solubilized in HCl. Ultrasan was dissolved
overnight in 0.1M hydrochloric acid, and the GP was prepared
in deionized water. Sterilization of the chitosan solution was
performed by autoclave, while the aqueous solution of GP was
filtered through a 0.22mm membrane. After sterilization, the
two solutions were cooled on ice, and mixed by adding the GP
drop-wise into the stirring chitosan solution. The resulting
mixture contained 2% w/w chitosan and 8% w/w GP. Such a
chitosan solution has been shown to remain liquid at
physiological pH at room temperature, but gels upon heating
to 37 1C [41]. Cell encapsulation was performed by gently
mixing this thermogelling solution at room temperature with
an equivalent volume of cell suspension in DMEM plus 10%
FCS until homogeneous (final cell density, 6� 106 cells/ml).
The thermogelling cell suspension was then poured into 48-
well culture plates (150ml/well, about 1� 106 cells in 1%
chitosan) and allowed to gel at 37 1C. After gelation, each well
was rinsed twice with fresh DMEM plus 10% FCS. Gels
containing 1.5% chitosan and 2� 106 cells/well were also
prepared.
2.5. Encapsulation of cells in chitosan/glycerophosphate/
hydroxyethyl cellulose (chitosan-GP/HEC) gels
A sterile chitosan-GP solution of 1.5% chitosan-135mM
disodium b-GP was prepared by dissolving 150mg Ultrasan
(79% deacetylated) in 9ml filter-sterilized 70mM HCl. This
solution was subjected to a 20min autoclave sterilization,
cooled to 4 1C, and 1ml filter-sterile 1.35M disodium b-GP(tissue culture grade, Sigma) was then added. Separately,
hydroxyethyl cellulose (HEC, Fluka Product Number 54290,
Sigma) was dissolved at 25mg/ml in DMEM and filter-
sterilized. A cell pellet containing NP or AF cells was
homogenously resuspended in 0.8ml HEC, then mixed with
4.8ml chitosan-GP at 4 1C (final cell density, 6� 106 cells/ml)
prior to pouring into a 60mm tissue culture Petri dish [42]. The
gel was allowed to solidify for 20min in a humidified, 5% CO2
cell culture incubator at 37 1C, and rinsed 5 times with
DMEM.
2.6. Encapsulation of cells in chitosan/genipin gels
A 2.5% Protasan solution (w/v) was prepared by adding the
Protasan powder progressively with stirring to 2ml serum-free
DMEM supplemented with 100mg/ml streptomycin, 100U/mlpenicillin and 50 ng/ml gentamicin, pH 7.4, at room tempera-
ture. Mixing was continued for 1 h until the Protasan was
completely dissolved. Subsequently, genipin powder (5% w/w
Protasan; Challenge Bioproducts Co., Taiwan) was added to
the chitosan solution and allowed to dissolve. Disc cells
suspended in 100ml serum-free DMEM were then added to the
Protasan/genipin solution (final cell density, 5� 106 cells/ml).
This solution was gently mixed to get a uniform suspension,
then poured into a 35mm plastic Petri dish and allowed to gel
at 37 1C.
2.7. Culture of chitosan/cell constructs
After gelling, cell-seeded scaffolds gelled in 48 well plates
were used for culture with 0.5ml culture medium. Cell-seeded
scaffolds gelled in Petri dishes were used to isolate 6mm
diameter and 2mm thick discs that were removed with a Keyes
Dermal Biopsy Punch (World Source, USA). The discs were
cultured in 24 well plates with 1.5ml medium. All cultures were
performed in DMEM containing vitamin C (50 mg/ml) at 37 1Cin a humidified CO2 incubator, with medium changes every 2
days. The medium was supplemented with either 10% FCS or
FCS plus 10 ng/ml recombinant human transforming growth
factor b1 (TGFb1; Biosource, Camarillo, CA). All cultureconditions were usually analyzed in duplicate with cells from
the same preparation, and all trends were confirmed using cells
from additional preparations. The results from one represen-
tative experiment are presented in the tables.
2.8. Analysis of GAG
GAG content was analysed using the dimethyl methylene
blue (DMMB) assay [43] after proteinase K digestion of the
construct or directly on the medium.
2.9. Analysis of cell viability
The viability of cells before and after entrapment in the
scaffold was evaluated using a fluorescence assay that
simultaneously determines live and dead cell numbers using
calcein AM (green) and ethidium homodimer (red), respec-
tively (LIVE/DEADs Viability/Cytotoxicity Assay; Molecu-
lar Probes, USA). The cells or cell-seeded scaffolds were
exposed to the probes for 30min at 37 1C, and cells were then
visualized using a Zeiss fluorescence microscope equipped with
a digital camera, following compression of the construct on a
microscope slide. For selected images, automatic cell counting
was performed using ImageJ (Java freeware).
2.10. Histology
Constructs were fixed in 2.5% v/v glutaraldehyde for 24 h at
4 1C, and then dehydrated with a 50–100%v/v ethanol series
with a final change in xylene, before embedding in paraffin.
Five micrometers sections were cut and mounted onto
positively charged slides (Fisher Scientific), which were heated
at 55 1C to ensure adherence of the sections. For staining,
sections were dewaxed in xylene, then rehydrated in a
100–50% ethanol series, and quickly rinsed in distilled water.

ARTICLE IN PRESSP. Roughley et al. / Biomaterials 27 (2006) 388–396 391
The chitosan scaffold was stained with Fast green and GAG/
proteoglycan was subsequently visualized by staining with
Safranin O.
3. Results
In initial experiments 1% or 1.5% chitosan hydrogelswere formed in the presence of b-GP in the bottom of48-well culture plates (chitosan-GP gels) with cellsseeded at 6� 106 or 12� 106 cells/ml. The initialhydrogels weighed about 150mg and contained either1� 106 or 2� 106 cells encapsulated in either 1% or1.5% chitosan. The results revealed that with bothchitosan concentrations about 60% of the proteoglycan(assessed as GAG) produced by NP cells was retainedby the constructs and not released into the culturemedium (Table 1). Doubling the cell density in thescaffold did not result in twice the amount ofaccumulated proteoglycan, although the GAG contentdid increase. With respect to a healthy bovine IVD, theproteoglycan concentration of the constructs after 20days represents about 8–12% of that occurring in thetissue [29]. When the cells from the AF were cultured ina similar manner, the results were strikingly different.The AP cells did not survive the culture period and weremostly dead by the end of 20 days. By this time,dissolution of the scaffold had occurred resulting inconsiderable weight loss and little proteoglycan waspresent. Over 90% of the proteoglycan that wasproduced earlier during the culture period was presentin the culture medium (Table 1).The chitosan/GP hydrogels did not possess a parti-
cularly firm texture and did not lend themselves tomanipulation in culture. A different formulation wastherefore desirable, from which a firm texture gel results.Cross-linking of the chitosan provides one means ofincreasing gel rigidity and this was achieved by allowingpolymerization to proceed in the presence of hydro-xyethyl cellulose, to produce cell-seeded chitosan-GP/
Table 1
GAG retention in chitosan-GP hydrogel
Cell typea Chitosan (%) Cell no. x 106 Wet wt. (mg)
NP 1 1 13273
NP 1 2 14379
NP 1.5 1 13574
NP 1.5 2 12977
AF 1 1 43713
AF 1 2 4479
AF 1.5 1 69711
AF 1.5 2 73716
aCells were isolated from the nucleus pulposus (NP) or the annulus fibrosbGAG was analyzed by the DMMB assay in the chitosan construct (C) acThe percentage GAG retained in the construct relative to total synthesis
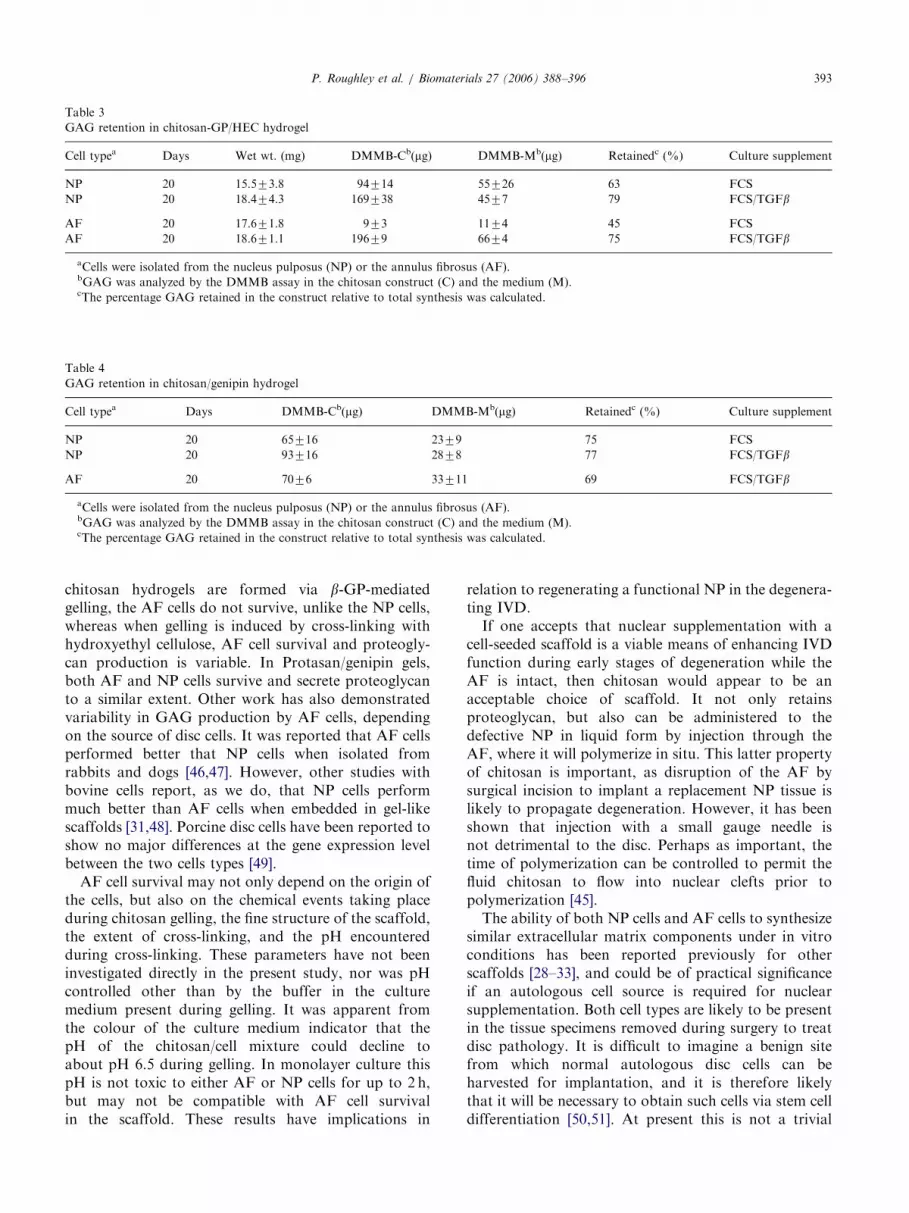
HEC gels [40,42]. This enabled uniform discs to bepunched from 2mm-thick sheets of the hydrogelprepared in a Petri dish. 1.5% chitosan-GP/HEC gelscontaining 6� 106 cells/ml were routinely used, and atypical 6mm disc weighed about 50mg upon initialisolation, and contained about 3� 105 cells. WhenNP cells were encapsulated, proteoglycan was foundto accumulate in the scaffold in the presence of FCS(Table 2). By day 20 about 70% of the synthesizedproteoglycan was retained by the scaffold. While theproteoglycan content of the scaffold was increased byday 40, it had not doubled, and the proportion ofretained material had declined, indicating that theability of the scaffold to retain the newly producedextracellular matrix does decline with time. During thistime period, cell number doubled up to day 20 and thenremained relatively constant. When cultures containedTGFb in addition to FCS to provide increased stimula-tion of proteoglycan synthesis, the degree of GAGretention by day 20 of culture could be increased to over80%. In all cases, the size of the NP cell-seededconstructs decreased during culture, particularly duringthe first 20 days, and the constructs became opaque andfirm (Fig. 1). By the end of culture the constructspossessed up to 15% of the GAG content of normaldisc. AF cells did not survive the culture conditions andby day 20 the constructs were once again small andtransparent (Fig. 1), with only minimal GAG beingpresent (Table 2). The presence of TGFb in addition toFCS did not improve AF cell survival or matrixproduction.However, this distinction between NP and AF cells
was not always apparent in the chitosan-GP/HECsystem. During a subsequent encapsulation in chito-san-GP/HEC, using a different batch of HEC, NP cellsbehaved in a similar manner to that described above,illustrating the reproducible behaviour of these cells inchitosan. In contrast, the AF cells survived theencapsulation and culture conditions in this case. Whencultured in the presence of FCS alone, the AF cells did
DMMB-Cb (mg) DMMB-Mb (mg) Retainedc (%)
54272 373768 59
884715 577724 61
572754 402732 59
6647100 446716 60
1878 167735 10
29712 28575 9
1874 203762 8
2771 302724 8
us (AF).
nd the medium (M).
was calculated.

ARTICLE IN PRESS
Table 2
GAG retention in chitosan-GP/HEC hydrogel
Cell typea Days Wet wt. (mg) DMMB-Cb(mg) DMMB-Mb(mg) Retainedc (%) Culture supplement
NP 20 28.0 106 46 70 FCS
NP 40 23.6 143 108 57 FCS
NP 20 17.3 135 23 85 FCS/TGFbNP 40 17.6 127 63 67 FCS/TGFb
AF 20 21.0 o1 37 — FCS
AF 40 17.2 o1 46 — FCS
AF 20 20.9 o1 37 — FCS/TGFbAF 40 22.3 3 49 6 FCS/TGFb
aCells were isolated from the nucleus pulposus (NP) or the annulus fibrosus (AF).bGAG was analyzed by the DMMB assay in the chitosan construct (C) and the medium (M).cThe percentage GAG retained in the construct relative to total synthesis was calculated.
Fig. 1. Appearance of cell-seeded chitosan-GP/HEC scaffolds. Scaf-
folds were seeded with either nucleus pulposus (NP) or annulus
fibrosus (AF) cells, then cultured for 20 days in the presence of FCS or
FCS plus TGFb.
P. Roughley et al. / Biomaterials 27 (2006) 388–396392
not produce much proteoglycan, but when the culturemedium was also supplemented the TGFb, proteoglycanproduction and retention within the scaffold resembledthat observed with the NP cells (Table 3). Thus, AFcells, although temperamental in the chitosan/b-GPsystem, have the potential to behave as NP cells underappropriate conditions.To determine whether other chitosan preparations
and cross-linking agents could generate a suitablescaffold for proteoglycan retention with NP and AFcells, Protasan G213 was cross-linked with genipin [44].Preliminary experiments had revealed that 2.5% Prota-san solutions cross-linked with 5% genipin gave optimal
gelling characteristics and cell survival upon encapsula-tion [45]. This system was also compatible with theproduction of uniform cell-seeded discs for in vitroculture work. NP and AF cells were again encapsulatedand maintained in culture for 20 days. In contrast toprevious studies using b-GP-induced chitosan gelling,both AF cells and NP cells survived in the chitosan/genipin scaffolds, with greater than 90% viability byday 20. In this scaffold both cell types produced proteo-glycan that was retained. In the case of NP cells, about75% of the proteoglycan was retained within thescaffold, while with AF cells the retention was onlyslightly decreased (Table 4).The chitosan scaffolds were also examined by
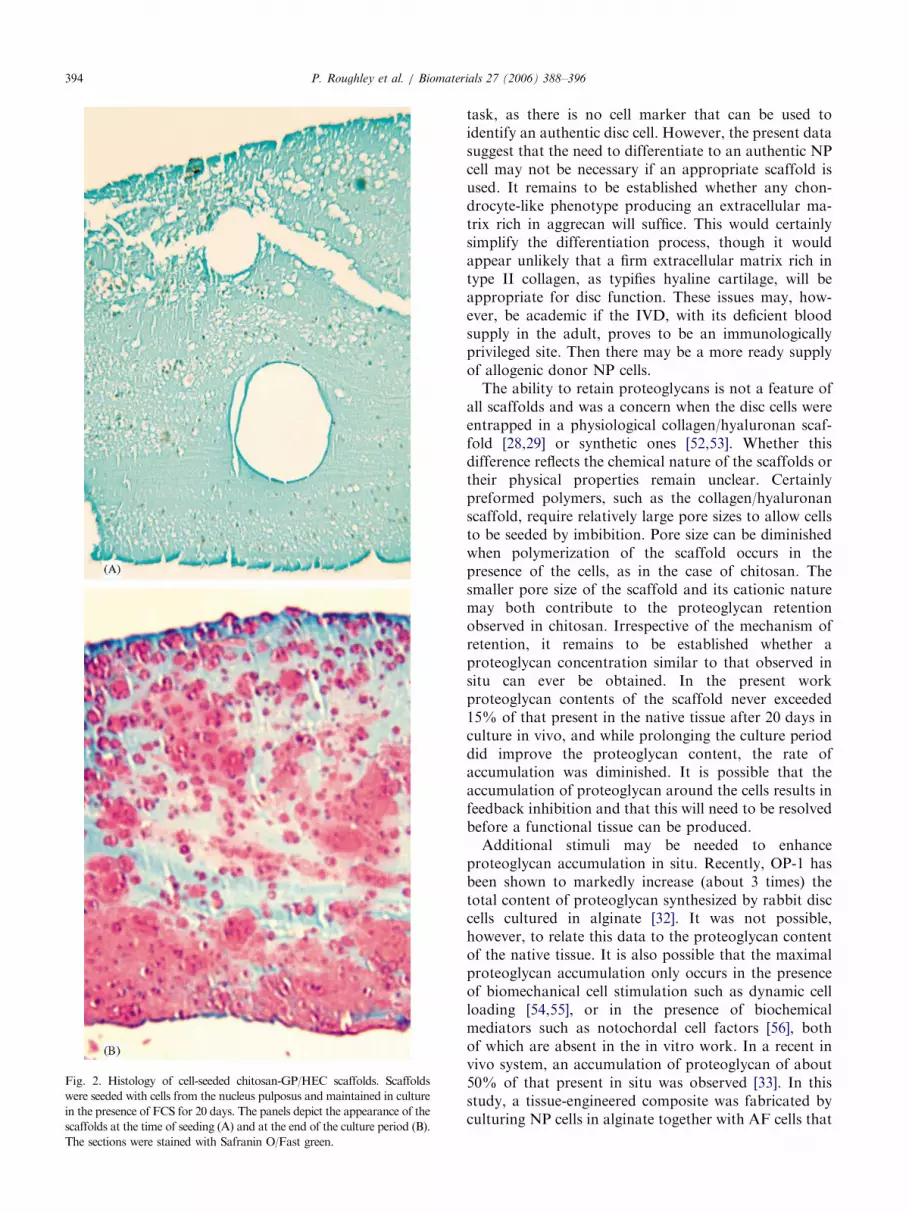
histology to identify the location of the entrapped cellsand the distribution of the retained proteoglycan.Safranin O staining was used to detect the anionicGAG chains of the proteoglycan. This stain does notbind to the cationic chitosan. Analysis of the cross-linked chitosan disc immediately after cell encapsulationrevealed a relatively uniform distribution of cellsthroughout the scaffold (Fig. 2). After 20 days ofculture in DMEM/FCS, Safranin O staining wasprominent around the NP cells, and in places mergedto form a continuous matrix. It is apparent that cells inboth the centre and periphery of the chitosan discs wereproducing proteoglycan. Similar findings were observedwith all three the chitosan formulations for NP cells.
4. Discussion
The present data indicate that chitosan hydrogels areable to retain the majority of the proteoglycan thatentrapped NP cells produce. Furthermore, NP cellsurvival and proteoglycan retention appears to berelatively independent of the method used to generatethe hydrogel. In contrast, AF cells do not behave in asimilar manner in all hydrogel formulations. When the
ARTICLE IN PRESS
Table 3
GAG retention in chitosan-GP/HEC hydrogel
Cell typea Days Wet wt. (mg) DMMB-Cb(mg) DMMB-Mb(mg) Retainedc (%) Culture supplement
NP 20 15.573.8 94714 55726 63 FCS
NP 20 18.474.3 169738 4577 79 FCS/TGFb
AF 20 17.671.8 973 1174 45 FCS
AF 20 18.671.1 19679 6674 75 FCS/TGFb
aCells were isolated from the nucleus pulposus (NP) or the annulus fibrosus (AF).bGAG was analyzed by the DMMB assay in the chitosan construct (C) and the medium (M).cThe percentage GAG retained in the construct relative to total synthesis was calculated.
Table 4
GAG retention in chitosan/genipin hydrogel
Cell typea Days DMMB-Cb(mg) DMMB-Mb(mg) Retainedc (%) Culture supplement
NP 20 65716 2379 75 FCS
NP 20 93716 2878 77 FCS/TGFb
AF 20 7076 33711 69 FCS/TGFb
aCells were isolated from the nucleus pulposus (NP) or the annulus fibrosus (AF).bGAG was analyzed by the DMMB assay in the chitosan construct (C) and the medium (M).cThe percentage GAG retained in the construct relative to total synthesis was calculated.
P. Roughley et al. / Biomaterials 27 (2006) 388–396 393
chitosan hydrogels are formed via b-GP-mediatedgelling, the AF cells do not survive, unlike the NP cells,whereas when gelling is induced by cross-linking withhydroxyethyl cellulose, AF cell survival and proteogly-can production is variable. In Protasan/genipin gels,both AF and NP cells survive and secrete proteoglycanto a similar extent. Other work has also demonstratedvariability in GAG production by AF cells, dependingon the source of disc cells. It was reported that AF cellsperformed better that NP cells when isolated fromrabbits and dogs [46,47]. However, other studies withbovine cells report, as we do, that NP cells performmuch better than AF cells when embedded in gel-likescaffolds [31,48]. Porcine disc cells have been reported toshow no major differences at the gene expression levelbetween the two cells types [49].AF cell survival may not only depend on the origin of
the cells, but also on the chemical events taking placeduring chitosan gelling, the fine structure of the scaffold,the extent of cross-linking, and the pH encounteredduring cross-linking. These parameters have not beeninvestigated directly in the present study, nor was pHcontrolled other than by the buffer in the culturemedium present during gelling. It was apparent fromthe colour of the culture medium indicator that thepH of the chitosan/cell mixture could decline toabout pH 6.5 during gelling. In monolayer culture thispH is not toxic to either AF or NP cells for up to 2 h,but may not be compatible with AF cell survivalin the scaffold. These results have implications in
relation to regenerating a functional NP in the degenera-ting IVD.If one accepts that nuclear supplementation with a
cell-seeded scaffold is a viable means of enhancing IVDfunction during early stages of degeneration while theAF is intact, then chitosan would appear to be anacceptable choice of scaffold. It not only retainsproteoglycan, but also can be administered to thedefective NP in liquid form by injection through theAF, where it will polymerize in situ. This latter propertyof chitosan is important, as disruption of the AF bysurgical incision to implant a replacement NP tissue islikely to propagate degeneration. However, it has beenshown that injection with a small gauge needle isnot detrimental to the disc. Perhaps as important, thetime of polymerization can be controlled to permit thefluid chitosan to flow into nuclear clefts prior topolymerization [45].The ability of both NP cells and AF cells to synthesize
similar extracellular matrix components under in vitroconditions has been reported previously for otherscaffolds [28–33], and could be of practical significanceif an autologous cell source is required for nuclearsupplementation. Both cell types are likely to be presentin the tissue specimens removed during surgery to treatdisc pathology. It is difficult to imagine a benign sitefrom which normal autologous disc cells can beharvested for implantation, and it is therefore likelythat it will be necessary to obtain such cells via stem celldifferentiation [50,51]. At present this is not a trivial
ARTICLE IN PRESS
Fig. 2. Histology of cell-seeded chitosan-GP/HEC scaffolds. Scaffolds
were seeded with cells from the nucleus pulposus and maintained in culture
in the presence of FCS for 20 days. The panels depict the appearance of the
scaffolds at the time of seeding (A) and at the end of the culture period (B).
The sections were stained with Safranin O/Fast green.
P. Roughley et al. / Biomaterials 27 (2006) 388–396394
task, as there is no cell marker that can be used toidentify an authentic disc cell. However, the present datasuggest that the need to differentiate to an authentic NPcell may not be necessary if an appropriate scaffold isused. It remains to be established whether any chon-drocyte-like phenotype producing an extracellular ma-trix rich in aggrecan will suffice. This would certainlysimplify the differentiation process, though it wouldappear unlikely that a firm extracellular matrix rich intype II collagen, as typifies hyaline cartilage, will beappropriate for disc function. These issues may, how-ever, be academic if the IVD, with its deficient bloodsupply in the adult, proves to be an immunologicallyprivileged site. Then there may be a more ready supplyof allogenic donor NP cells.The ability to retain proteoglycans is not a feature of
all scaffolds and was a concern when the disc cells wereentrapped in a physiological collagen/hyaluronan scaf-fold [28,29] or synthetic ones [52,53]. Whether thisdifference reflects the chemical nature of the scaffolds ortheir physical properties remain unclear. Certainlypreformed polymers, such as the collagen/hyaluronanscaffold, require relatively large pore sizes to allow cellsto be seeded by imbibition. Pore size can be diminishedwhen polymerization of the scaffold occurs in thepresence of the cells, as in the case of chitosan. Thesmaller pore size of the scaffold and its cationic naturemay both contribute to the proteoglycan retentionobserved in chitosan. Irrespective of the mechanism ofretention, it remains to be established whether aproteoglycan concentration similar to that observed insitu can ever be obtained. In the present workproteoglycan contents of the scaffold never exceeded15% of that present in the native tissue after 20 days inculture in vivo, and while prolonging the culture perioddid improve the proteoglycan content, the rate ofaccumulation was diminished. It is possible that theaccumulation of proteoglycan around the cells results infeedback inhibition and that this will need to be resolvedbefore a functional tissue can be produced.Additional stimuli may be needed to enhance
proteoglycan accumulation in situ. Recently, OP-1 hasbeen shown to markedly increase (about 3 times) thetotal content of proteoglycan synthesized by rabbit disccells cultured in alginate [32]. It was not possible,however, to relate this data to the proteoglycan contentof the native tissue. It is also possible that the maximalproteoglycan accumulation only occurs in the presenceof biomechanical cell stimulation such as dynamic cellloading [54,55], or in the presence of biochemicalmediators such as notochordal cell factors [56], bothof which are absent in the in vitro work. In a recent invivo system, an accumulation of proteoglycan of about50% of that present in situ was observed [33]. In thisstudy, a tissue-engineered composite was fabricated byculturing NP cells in alginate together with AF cells that

ARTICLE IN PRESSP. Roughley et al. / Biomaterials 27 (2006) 388–396 395
were seeded into a polyglycolic acid/polylactic acidpolymer. Finally, the construct was implanted in thesubcutaneous space of athymic mice for 12 weeks. Inthis closed system, proteoglycan cannot freely diffuseout of the construct, as in most of the in vitro studiesdescribed until now, and therefore accumulation canoccur within the alginate gel. However, one shouldappreciate that, while the latter approach is aimed atreplacing the entire IVD (annulus and nucleus tissues atthe same time), our approach is intended to supplementor replace only the NP tissue.Finally one must accept that while nuclear supple-
mentation may be technically feasible, the environmentof the degenerate disc may be unsuitable for thegeneration of adequate repair. It is commonly believedthat inadequate disc nutrition in the adult predisposesthe tissue to degenerative changes [4,5]. If this is the casethen supplementation of the NP with additional cellsmay be of immediate benefit but ultimately destined forfailure unless a means can be found to improve discnutrition. Only time will tell if this proves to be the case,but in the meantime nuclear supplementation continuesto be a technique worthy of investigation, with chitosanas a potential scaffold.
Acknowledgements
This work was supported by the Shriners of NorthAmerica, the Canadian Arthritis Network, the Cana-dian Institutes for Health Research and the AOFoundation.
References
[1] Benjamin M, Evans EJ. Fibrocartilage. J Anat 1990;171:1–15.
[2] Hukins DWL, Meakin JR. Relationship between structure and
mechanical function of the tissues of the intervertebral joint. Am
Zool 2000;40:42–52.
[3] Buckwalter JA. Aging and degeneration of the human inter-
vertebral disc. Spine 1995;20:1307–14.
[4] Horner HA, Urban JPG. 2001 Volvo Award winner in basic
science studies: effect of nutrient supply on the viability of cells
from the nucleus pulposus of the intervertebral disc. Spine
2001;26:2543–9.
[5] Roberts S, Urban JPG. Degeneration of intervertebral disc.
Arthritis Res Ther 2003;5:120–30.
[6] Oegema TR. Biochemistry of the intervertebral disc. Clin Sports
Med 1993;12:419–39.
[7] Watanabe H, Yamada Y, Kimata K. Roles of aggrecan, a large
chondroitin sulfate proteoglycan, in cartilage structure and
function. J Biochem (Tokyo) 1998;124:687–93.
[8] Pearce RH, Grimmer BJ. Target tissue models: the proteoglycans
and degeneration of the human intervertebral disc. J Rheumatol
1983;10(Suppl. 11):108–10.
[9] Thompson JP, Pearce RH, Schechter MT, Adams ME, Tsang
IKY, Bishop PB. Preliminary evaluation of a scheme for grading
the gross morphology of the human intervertebral disc. Spine
1990;15:411–5.
[10] Vanharanta H. The intervertebral disc: a biologically active tissue
challenging therapy. Ann Med 1994;26:395–9.
[11] Luoma K, Riihimaki H, Luukkonen R, Raininko R, Viikari-
Juntura E, Lamminen A. Low back pain in relation to lumbar
disc degeneration. Spine 2000;25:487–92.
[12] Ishihara H, McNally DS, Urban JPG, Hall AC. Effects of
hydrostatic pressure on matrix synthesis in different regions of the
intervertebral disk. J Appl Physiol 1996;80:839–46.
[13] Handa T, Ishihara H, Ohshima H, Osada R, Tsuji H, Obata K.
Effects of hydrostatic pressure on matrix synthesis and matrix
metalloproteinase production in the human lumbar intervertebral
disc. Spine 1997;22:1085–91.
[14] Hutton WC, Elmer WA, Boden SD, Hyon S, Toribatake Y,
Tomita K, et al. The effect of hydrostatic pressure on
intervertebral disc metabolism. Spine 1999;24:1507–15.
[15] Hsieh AH, Lotz JC. Prolonged spinal loading induces matrix
metalloproteinase-2 activation in intervertebral discs. Spine
2003;28:1781–8.
[16] Kang JD, Stefanovic-Racic M, McIntyre LA, Georgescu HI,
Evans CH. Toward a biochemical understanding of human
intervertebral disc degeneration and herniation—contributions of
nitric oxide, interleukins, prostaglandin E2, and matrix metallo-
proteinases. Spine 1997;22:1065–73.
[17] Goupille P, Jayson MIV, Valat JP, Freemont AJ. Matrix
metalloproteinases: The clue to intervertebral disc degeneration?
Spine 1998;23:1612–26.
[18] Oegema TR, Johnson SL, Aguiar DJ, Ogilvie JW. Fibronectin
and its fragments increase with degeneration in the human
intervertebral disc. Spine 2000;25:2742–7.
[19] Liu GZ, Ishihara H, Osada R, Kimura T, Tsuji H. Nitric oxide
mediates the change of proteoglycan synthesis in the human
lumbar intervertebral disc in response to hydrostatic pressure.
Spine 2001;26:134–41.
[20] Ahn SH, Cho YW, Ahn MW, Jang SH, Sohn YK, Kim HS.
mRNA expression of cytokines and chemokines in herniated
lumbar intervertebral discs. Spine 2002;27:911–7.
[21] Roberts S, Caterson B, Menage J, Evans EH, Jaffray DC,
Eisenstein SM. Matrix metalloproteinases and aggrecanase—their
role in disorders of the human intervertebral disc. Spine
2000;25:3005–13.
[22] Annunen S, Paassilta P, Lohiniva J, Perala M, Pihlajamaa T,
Karppinen J, et al. An allele of COL9A2 associated with
intervertebral disc disease. Science 1999;285:409–12.
[23] Kawaguchi Y, Osada R, Kanamori M, Ishihara H, Ohmori K,
Matsui H, et al. Association between an aggrecan gene
polymorphism and lumbar disc degeneration. Spine 1999;24:
2456–60.
[24] Ala-Kokko L. Genetic risk factors for lumbar disc disease. Ann
Med 2002;34:42–7.
[25] Guyer RD, Ohnmeiss DD. Intervertebral disc prostheses. Spine
2003;28:S15–23.
[26] An HS, Thonar EJMA, Masuda K. Biological repair of
intervertebral disc. Spine 2003;28:S86–92.
[27] Cassinelli EH, Hall RA, Kang JD. Biochemistry of intervertebral
disc degeneration and the potential for gene therapy applications.
Spine J 2001;1:205–14.
[28] Rong Y, Sugumarn G, Silbert JE, Spector M. Proteoglycans
synthesized by canine intervertebral disc cells grown in a type I
collagen-glycosaminoglycan matrix. Tissue Eng 2002;8:1037–47.
[29] Alini M, Li W, Markovic P, Aebi M, Spiro RC, Roughley PJ. The
potential and limitations of a cell-seeded collagen/hyaluronan
scaffold to engineer an intervertebral disc-like matrix. Spine
2003;28:446–53.
[30] Gruber HE, Leslie K, Ingram J, Norton HJ, Hanley EN. Cell-
based tissue engineering for the intervertebral disc: in vitro studies
of human disc cell gene expression and matrix production within
selected cell carriers. Spine J 2004;4:44–55.

ARTICLE IN PRESSP. Roughley et al. / Biomaterials 27 (2006) 388–396396
[31] Horner HA, Roberts S, Bielby RC, Menage J, Evans H, Urban
JPG. Cells from different regions of the intervertebral disc. Spine
2002;27:1018–28.
[32] Masuda K, takegami K, An H, Kumano F, Chiba K, Andersson
GB, Schmid T, Thonar E. Recombinant osteogenic protein-1
upregulates extracellular matrix metabolism by rabbit annulus
fibrosus and nucleus pulposus cells cultured in alginate beads.
J Orthop Res 2003;21:922–30.
[33] Mizuno H, Roy AK, Vacanti CA, Kojima K, Ueda M, Bonassar
LJ. Tissue-engineered composites of annulus fibrosus and nucleus
pulposus for intervertebral disc replacment. Spine 2004;29:
1290–8.
[34] Suh JKF, Matthew HWT. Application of chitosan-based poly-
saccharide biomaterials in cartilage tissue engineering: a review.
Biomaterials 2000;21:2589–98.
[35] Khor E, Lim LY. Implantable applications of chitin and chitosan.
Biomaterials 2003;24:2339–49.
[36] Singla AK, ChawlaM. Chitosan: some pharmaceutical and biological
properties—an update. J Pharm Pharmacol 2001;53:1047–67.
[37] Berger J, Reist M, Mayer JM, Felt O, Peppas NA, Gurny R.
Structure and interactions in covalently and ionically crosslinked
chitosan hydrogels for biomedical applications. Eur J Pharm
Biopharm 2004;57:19–34.
[38] Berger J, Reist M, Mayer JM, Felt O, Gurny R. Structure and
interactions in chitosan hydrogels formed by complexation or
aggregation for biomedical applications. Eur J Pharm Biopharm
2004;57:35–52.
[39] Chenite A, Chaput C, Wang D, Combes C, Buschmann MD,
Hoemann CD, et al. Novel injectable neutral solutions of chitosan
form biodegradable gels in situ. Biomaterials 2000;21:2155–61.
[40] Hoemann CD, Sun J, Chrzanowski V, Buschmann MD. A
multivalent assay to detect glycosaminoglycan, protein, collagen,
RNA, and DNA content in milligram samples of cartilage or
hydrogel-based repair cartilage. Anal Biochem 2002;300:1–10.
[41] Chenite A, Buschmann MD, Wang D, Chaput C, Kandani N.
Rheological characterization of thermogelling chitosan/glycerol
phosphate solutions. Carbohydr Polym 2001;46:39–47.
[42] Hoemann CD, Sun J, Legare A, McKee MD, Buschmann MD.
Tissue engineering of cartilage using an injectable and adhesive
chitosan-based cell delivery vehicle. Osteoarthritis Cart 2005;
13:318–29.
[43] Farndale RW, Buttle DJ, Barrett AJ. Improved quantitation and
discrimination of sulphated glycosaminoglycans by use of
dimethylmethylene blue. Biochim Biophys Acta 1986;883:173–7.
[44] Mi FL, Tan YC, Liang HF, Sung HW. In vivo biocompatibility
and degradability of a novel injectable chitosan-based implant.
Biomaterials 2002;23:181–91.
[45] Mwale F, Iordanova M, Demers CN, Steffen T, Roughley P,
Antoniou J. Biological evaluation of chitosan salts cross-linked to
genipin as a scaffold for disc tissue engineering. Tissue Eng
2005;11:130–40.
[46] Maldonado BA, Oegema TR. Initial characterization of the
metabolism of intervertebral disc cells encapsulated in micro-
spheres. J Orthop Res 1992;10:677–90.
[47] Chiba K, Andersson GB, Masuda K, Thonar EJ. Metabolism of
the extracellular matrix formed by intervertebral disc cells
cultured in alginate. Spine 1997;22:2885–93.
[48] Heathfield TF, Marchesi F, Aebi M, Alini M. Metabolic studies
of bovine intervertebral disc cells cultured in alginate beads:
variation with age and location. Trans Orthop Res Soc 1996;
21:678.
[49] Boyd LM, Chen J, Kraus VB, Setton LA. Conditioned medium
differentially regulates matrix protein gene expression in cells of
the intervertebral disc. Spine 2004;29:2217–22.
[50] Sakai D, Mochida J, Yamamoto Y, Nomura T, Okuma M,
Nishimura K, Nakai T, Ando K, Hotta T. Transplantation of
mesenchymal stem cells embedded in atelocollagen gel to the
intervertebral disc: a potential therapeutic model for disc
degeneration. Biomaterials 2003;24:3531–41.
[51] Risbud MV, Albert TJ, Guttapalli A, Vresilovic EJ, Hillibrand
AS, Vaccaro AR, Shapiro IM. Differentiation of mesenchymal
stem cells towards a nucleus pulposus-like phenotype in vitro:
implications for cell-based transplantation therapy. Spine 2004;
29:2627–32.
[52] Grad S, Zhou L, Gogolewski S, Alini M. Chondrocytes seeded
onto poly-L/DL-lactide (80%/20%) porous scaffolds: a biochem-
ical evaluation. J Biomed Mater Res 2003;66:571–9.
[53] Grad S, Kupcsik L, Gorna K, Gogolewski S, Alini M. The use of
biodegradable polyurethane scaffolds for cartilage tissue engi-
neering: potential and limitations. Biomaterials 2003;24:5163–71.
[54] MacLean JJ, Lee CR, Grad S, Ito K, Alini M, Iatridis JC. Effects
of immobilization and dynamic compression on intervertebral
disc cell gene expression in vivo. Spine 2003;28:973–81.
[55] Walsh AJL, Lotz JC. Biological response of the intervertebral disc
to dynamic loading. J Biomech 2004;37:329–37.
[56] Hunter CJ, Matyas JR, Duncan NA. The notochordal cell in the
nucleus pulposus: a review in the context of tissue engineering.
Tissue Eng 2003;9:667–77.