the microbe-mineral environment and gypsum neogenesis in a weathered polar evaporite
TRANSCRIPT

The microbe–mineral environment and gypsum neogenesis ina weathered polar evaporiteC. S. COCKELL,1 G. R. OSINSKI,2 N. R. BANERJEE,3 K. T. HOWARD,4 I . GILMOUR5 AND
J. S. WATSON5
1Geomicrobiology Research Group, Centre for Earth, Planetary, Space and Astronomical Research (CEPSAR), Open University,
Milton Keynes, UK2Department of Earth Sciences ⁄ Physics and Astronomy, University of Western Ontario, London, ON, Canada3Department of Earth Sciences, University of Western Ontario, London, ON, Canada4Impacts and Astromaterials Research Centre, Department of Mineralogy, Natural History Museum, London, UK5Centre for Earth, Planetary, Space and Astronomical Research (CEPSAR), Open University, Milton Keynes, UK
ABSTRACT
Evaporitic deposits are a globally widespread habitat for micro-organisms. The microbe–mineral environment in
weathered and remobilized gypsum from exposed mid-Ordovician marine evaporite beds in the polar desert of
Devon Island, Nunavut, Canadian High Arctic was examined. The gypsum is characterized by internal green
zones of cyanobacterial colonization (dominated by Gloeocapsa ⁄ Aphanothece and Chroococcidiopsis spp. mor-
photypes) and abundant black zones, visible from the surface, that contain pigmented cyanobacteria and fungi.
Bioessential elements in the gypsum are primarily provided by allochthonous material from the present-day polar
desert. The disruption, uplift and rotation of the evaporite beds by the Haughton meteorite impact 39 Ma have
facilitated gypsum weathering and its accessibility as a habitat. No cultured cyanobacteria, bacteria and fungi
were halophilic consistent with the expectation that halophily is not required to persist in gypsum habitats.
Heterotrophic bacteria from the evaporite were slightly or moderately halotolerant, as were heterotrophs
isolated from soil near the gypsum outcrop showing that halotolerance is common in arctic bacteria in this
location. Psychrotolerant Arthrobacter species were isolated. No psychrophilic organisms were isolated. Two
Arthrobacter isolates from the evaporite were used to mediate gypsum neogenesis in the laboratory,
demonstrating a potential role for microbial biomineralization processes in polar environments.
Received 6 October 2009; accepted 23 March 2010
Corresponding author: Charles S. Cockell. Tel.: +44 1908 652588; fax: +44 1908 848022; e-mail: c.s.cockell@
open.ac.uk
INTRODUCTION
Microbial communities that inhabit evaporitic deposits have
been described from a diversity of global locations. The best
studied communities associated with evaporitic deposits are
those of the marine intertidal zone and salterns (e.g. Cau-
mette, 1993; Caumette et al., 1994; Rothschild et al., 1994),
where organisms inhabit the salt crusts produced from the
evaporation of seawater. Within some evaporitic deposits that
are sufficiently stable, a well-defined zonation of the interior
community occurs. Oren et al. (1995) described the zonation
of communities inhabiting gypsum deposits in a saltern pond
in Eilat, Israel, with distinctive differentiation occurring
between layers of oxygenic phototrophs, dominated by cyano-
bacteria such as Synechococcus and Phormidium spp., and
purple sulphur bacteria beneath them. This distinct zonation
was also demonstrated by fatty acid (Ionescu et al., 2007) and
molecular biological analysis (Sørensen et al., 2005). Synecho-
coccus are reported to be a dominant phototroph taxa in
the evaporites in Guerrero Negro, Baja, California, which is
composed of 22% gypsum (Rothschild et al., 1994).
Some organisms within evaporitic deposits or the salty
waters associated with them are not merely passive inhabit-
ants, but play an active role in biogeochemical processes, for
example fixing nitrogen (Boison et al., 2004) or precipitating
minerals. Douglas and Yang report the biogenic formation of
rosickyite, a thermodynamically unstable form of sulphur in
gypsum crusts in Death Valley, USA (Douglas & Yang, 2002;
� 2010 Blackwell Publishing Ltd 293
Geobiology (2010), 8, 293–308 DOI: 10.1111/j.1472-4669.2010.00240.x

Douglas, 2004). The extent to which the organisms within
salt deposits take part in processes such as microbial mineral
neogenesis is still poorly understood. As these processes could
potentially leave mineral biosignatures or preserve organic
remains, evaporites have been considered promising targets
for investigating life on the early Earth or possibly elsewhere
(Mancinelli et al., 2004; Edwards et al., 2005; Stivaletta
et al., 2005), particularly on Mars where large quantities of
sulphate deposits, including gypsum, are known to occur
(Clark et al., 1982; Rieder et al., 1997; Gendrin et al., 2005;
Langevin et al., 2005; Squyres et al., 2006).
Communities in evaporitic deposits have been described in
extreme environments, where the habitat can provide protec-
tion from the environment, in a similar way to which endoliths
are protected from extreme conditions by living in the interior
of rocks. For example, cyanobacteria, principally Chroococcidi-
opsis morphotypes, inhabit evaporitic deposits in the Atacama
Desert (Chile) (Wierzchos et al., 2006). The organisms inha-
bit halite mounds within the hyperarid core of the desert,
achieving metabolic activity during brief periods of moisture
availability, potentially assisted by halite deliquescence (Davil-
la et al., 2008). Chroococcidiopsis phylotypes have been
described from gypsum soil crusts in the Atacama Desert,
Mojave Desert (USA), Al-Jafr Basin (Jordan) (Dong et al.,
2007) and a gypsum outcrop in the Harz Mountains
(Germany) (Boison et al., 2004).
Polar environments also host organisms capable of inhabit-
ing salt crusts. Hughes & Lawley (2003) described the coloni-
zation of thin gypsum crusts on sandstone boulders on
Alexander Island, Antarctica by endolithic organisms, includ-
ing cyanobacteria, bacteria and fungi. Although primary pro-
ductivity was low, they showed that the gypsum provides
protection from UV radiation and desiccation. Parnell et al.
(2004) described the cyanobacterial colonization of selenite
(calcium sulphate) crystals in the Haughton impact structure,
Devon Island, Canadian High Arctic. These crystals were
formed by post-impact hydrothermal activity at the margins
of the crater and the organisms inhabit the space between the
selenite sheets.
Organisms that inhabit gypsum deposits would not neces-
sarily be expected to be halophilic as the solubility of gypsum
is low in water [2.4 g L)1 (0.014 M) at 25 �C], compared to
halite [360 g L)1 (6.16 M) at 25 �C]. However, Garcia-Pichel
et al. (2001) studied soil crusts of the Colorado Plateau, USA
and found that gypsum crusts were the most phylogenetically
divergent of the crusts studied, which they suggested might
be caused by the requirement for halotolerance. Thus, the
physiological requirements for living within gypsum require
further study.
In this work, we investigated the microbe–mineral environ-
ment in weathered and remobilized gypsum associated with
Ordovician evaporitic outcrops in a polar desert on Devon
Island in the Canadian High Arctic. We tested hypotheses
concerning the ability of organisms to take part in mineral
neogenesis and their physiological requirements.
MATERIALS AND METHODS
Study sites
Gypsum outcrops of the mid-Ordovician Bay Fiord Forma-
tion occur throughout the 39 Ma, 23-km diameter Haugh-
ton impact structure (Bischoff & Oskierski, 1988; Osinski
et al., 2005a). Samples of evaporites from a gypsum outcrop
at 75�23.85¢ N, 89�32.37¢ W (Fig. 1) were collected for
geomicrobiological analysis. The outcrop comprised a block
Fig. 1 Location of gypsum outcrop studied in this
article in relation to major lithological units in the
Haughton impact structure, Nunavut, Canadian
High Arctic (map after Osinski et al., 2005b).
294 C. S. COCKELL et al.
� 2010 Blackwell Publishing Ltd

of rotated ⁄ disrupted gypsum constituting part of the cen-
tral uplift draped by impact melt breccia in the east of the
Haughton impact structure in the Haughton River valley
(Fig. 2A,B). Samples were collected into aseptic bags and
maintained at ambient temperatures until return to the
laboratory.
Light microscopy
Light microscopy was used to examine the zonation of organ-
isms within the gypsum crust. Material from several represen-
tative parts of colonized gypsum was examined by crushing
the material gently into a fine powder. Some of this powder
was distributed onto a microscope slide and doubly distilled
H2O was added to suspend the material. The material was
examined using bright field microscopy on a Leica DMRP
fluorescence microscope (Leica Microsystems, Wetzlar,
Germany). Autofluorescence of cyanobacteria was examined
using a Leica N2.1 cube, which uses an excitation filter with a
bandpass of 515–560 nm and a long-band emission filter
(>590 nm).
To visualize prokaryotes, gypsum samples were stained by
soaking them in SYBR Green I DNA binding dye according
to the manufacturer’s instructions (Invitrogen, Paisley, UK).
Stained cells were observed fluorescing green using an excita-
tion waveband of 450–490 nm (Leica filter cube I3) and a
long-band emission cutoff filter of >515 nm. Images of
stained cells were superposed on bright field micrographs of
the same field of view to show relationships between stained
prokaryotes and cyanobacteria. Control tests on autoclaved
material showed that the stain did not non-specifically bind to
gypsum or other minerals present in the habitat.
Scanning electron microscopy and energy dispersive
spectroscopy
The location and characteristics of micro-organisms within
the gypsum and their mineral environment were examined by
scanning electron microscopy (SEM). Selected blocks (�0.3–
1 cm3) of gypsum encrustations from green colonized zones
were mounted on aluminium stubs with two-sided carbon
tape. Samples were carbon coated (15–20 nm thickness) and
were examined in secondary electron mode at a 20 kV acceler-
ating voltage and 7–15 mm working distance using a Quanta
3D dual beam FIBSEM (FEI, Hillsboro, OR, USA). Energy
dispersive spectroscopy (EDS), with a count time of 50–100 s
and data analysis with the software, Inca (Oxford Scientific
Instruments, Oxford, UK) were used to study qualitatively
the elemental composition of the samples.
To obtain high resolution images of crystals formed in gyp-
sum neogenesis experiments, a field emission gun (FEG)
SEM (ZEISS Supra 55-VP; Zeiss Microimaging, Gottingen,
Germany) was used. Samples were carbon coated (15–20 nm
thickness) and were examined in secondary electron mode at a
2–15 kV accelerating voltage and 7–10 mm working distance.
Analysis of the crystal composition was accomplished by EDS
as described above.
Transmission electron microscopy of gypsum sections
Transmission electron microscopy (TEM) was used to exam-
ine the relationship of organisms to the surface of the gypsum
and the presence of extracellular polysaccharide (EPS) around
organisms. Small fragments of gypsum from the green colo-
nized zone were dropped into 2.5% glutaraldehyde in 0.1 M
A B
C D
Fig. 2 Characteristics of the gypsum habitat. (A)
Gypsum outcrop at 75�2385¢ N, 89�3237¢ W emb-
edded within the melt-rocks of the Haughton
impact structure. Note rotation and disruption of
bedding planes (white dashed lines) caused by
impact (scale bar 1 m). (B) Close-up of black square
in (A) showing detail of bedding planes (scale bar
1 m). (C) Surface of outcrop showing precipitation
of gypsum along the vertical bedding planes and
fractures in which organisms preferentially grow
(scale bar 1 cm). Note visible black colouration,
which is biological. (D) Green zone of phototrophs
within reprecipitated gypsum (arrow) (scale bar
1 cm).
Microbe–mineral environment and gypsum neogenesis 295
� 2010 Blackwell Publishing Ltd

sodium cacodylate pH 7.4. The gypsum fragments were
post-fixed in 2% osmium tetroxide in sodium cacodylate,
dehydrated through an ascending series of acetone concentra-
tions, infiltrated in Epon (Hexion, Columbus, OH, USA)
minus accelerator at 60 �C with six changes. The samples were
finally embedded in complete Epon and polymerized at 60 �Cfor 48 h.
Ultrathin sections of �70 nm were cut using a microtome
diamond knife and collected on copper slot grids with a
formvar film. The sections were counter-stained with uranyl
acetate and Reynolds lead citrate before examination in a JEM
1400 transmission electron microscope (Jeol, Tokyo, Japan)
operating at 80 kV. Digital images were acquired using an
AMT XR60 digital camera (AMT Imaging, Danvers, MA,
USA).
X-ray diffraction
X-ray diffraction was used to study the parent material in the
outcrop and the reprecipitated gypsum. An INEL X-ray
diffractometer (XRD) (Inel, Strasbourg, France) with a
curved position sensitive detector was used for XRD analyses
at the Natural History Museum, London. A Ge monochro-
mator allowed selection of only CuKa radiation. Samples were
ground to a 35 lm powder and mounted on single-crystal
quartz substrates that were rotated continuously during
analysis. Post-monochromator slits were used to restrict the
beam to 0.24 · 5.00 mm and patterns were collected with
the beam at an incident angle of 3.6� to the flat-topped
sample. Analyses were conducted over 30 min. Silicon and
silver behenate were used as external standards in this
study. Calibration and data collection were performed using
Diffgrab�. WinXPow� was used for phase identification.
X-ray fluorescence
X-ray fluorescence (XRF) was used to examine the elemental
composition of the parent and reprecipitated gypsum and thus
the potential supply of nutrients to the enclosed biota. Sam-
ples of source evaporite taken from beneath the colonized
zones and reprecipitated gypsum deposits from green colo-
nized zones were sampled for XRF analysis. The samples were
crushed into an homogenous powder in a mortar and pestle
and pressed into discs using a Methyl methacrylate binder.
Analysis was carried out using an XRF Philips PW1480 Spec-
trometer (PANalytical, Almelo, The Netherlands) with semi-
quantitative software.
Raman spectroscopy
Raman spectroscopy was used to investigate the mineralogy of
the parent material and reprecipitated gypsum, to examine
whether the parent material contained anhydrite and to cross-
correlate spectra to XRD and XRF data. Raman was carried
out using a Horiba Jobin Yvon LabRAM (Horiba Jobin Yvon,
Stanmore, UK) and samples were excited using a 514.5-nm
(green) laser at 0.7 mW. For each material, three separate
samples of �10 g were powdered in a mortar and pestle and
pooled for analysis to obtain spectra representative of the
mean composition of the material. The footprint area of analy-
sis was a 2-lm-diameter spot. Spectra were the mean of five
separate spectra, each acquired over 20 s. Data were gathered
in the program LABSPEC (Horiba Jobin Yvon).
Penetration of light into the samples
The penetration of light into the reprecipitated gypsum was
examined to determine the light environment for photo-
trophs and the protection that they receive from ultraviolet
(UV) radiation. Light transmission spectra (250–750 nm)
were measured with an optical microspectrophotometer
(Craic Technologies, San Dimas, CA, USA), linked to a Leica
DMR microscope. The light source was a 75-W xenon-arc
lamp and the light was focussed onto an area of 10 · 10 lm.
Two representative samples of thin consolidated gypsum crust
were examined of 0.9-mm thickness each. Spectra at 10 differ-
ent spots were acquired and a mean spectrum was calculated.
The penetration of light at 300 nm (midway through the
UVB radiation region), 360 nm (midway through the UVA
radiation region) and at 680 nm (taken as the chlorophyll a
absorption maximum) was calculated.
Sulphur isotopic analysis
Sulphur isotopic analysis was carried out to determine if
gypsum reprecipitation was microbially mediated. Samples for
isotopic analysis were prepared by collecting �2 g from four
independent gypsum samples and crushing them into an
homogenous powder using a mortar and pestle. Two sets of
samples were prepared for analysis in this way. Samples of unal-
tered source evaporite, reprecipitated gypsum near the colo-
nized zones (uncolonized by phototrophs) and heavily
colonized (from the green colonized zones) gypsum were
prepared. Sulphur isotope analysis was undertaken by
EA-IRMS using a Europa Scientific 20-20 isotope ratio mass
spectrometer linked to a Sercon elemental analyser (Sercon,
Crewe, UK). Tin capsules containing reference or sample
material plus vanadium pentoxide catalyst were loaded into an
automatic sampler and dropped into a furnace held at
1080 �C and combusted in the presence of oxygen. The
combusted gases were treated in a helium stream over com-
bustion catalysts (tungstic oxide ⁄ zirconium oxide). Water was
removed using a Nafion� membrane (DuPont, Willmington,
DE, USA). The reference material used for 34S isotope analysis
was IA-R036 (barium sulphate, d34SV-CDT = +20.74&).
Samples of IA-R036, IA-R025 (barium sulphate, d34SV-CDT =
+8.53&) and IA-R026 (silver sulphide, d34SV-CDT =
+3.96&) were used for calibration and correction of the
296 C. S. COCKELL et al.
� 2010 Blackwell Publishing Ltd

18O contribution to the SO ion beam. IA-R036, IA-R025 and
IA-R026 are in-house standards calibrated and traceable to
NBS-127 (barium sulphate, d34SCDT = +20.3&) and IAEA-
S-1 (silver sulphide, d34SV-CDT = )0.3&). For quality control
purposes, test samples of IA-R036 and IAEA-SO-5 (barium
sulphate, d34SV-CDT = +0.50&) were measured. The data are
reported in the usual delta notation relative to V-CDT. The
error was ±0.5 per mil.
Thermochemolysis–GC–MS analysis
Thermochemolysis–GC–MS analysis was used to study the
major lipids present in the gypsum. Approximately 350 mg of
crushed colonized gypsum was dissolved in deionized water
(8 mL), sonicated and centrifuged. This procedure was
repeated five times. The residue was then freeze-dried. To
analyse microbial lipids in the gypsum, an aliquot of the
freeze-dried residue was placed in a quartz pyrolysis tube
plugged at either end with quartz wool; 10 lL of 25% tetram-
ethylammonium hydroxide in methanol was added to the
sample and the methanol was allowed to evaporate for 12 h.
The sample was heated using a CDS 5000 Pyroprobe fitted
with a 1500 valve interface held at 250 �C (CDS Analytical,
Oxford, PA, USA) and coupled to a GC-MS. GC-MS analysis
was carried out using an Agilent Technologies 6890 gas
chromatograph coupled to a 5973 mass spectrometer. Separa-
tion was performed on a S.G.E. (UK) BPX-5 column (30 m
length, 0.25 mm internal diameter and 0.25 lm film
thickness). Helium at a column flow rate of 1.1 mL min)1
was used as the carrier gas. Injection was at a 5:1 split and the
injector temperature was 270 �C. The GC oven temperature
was held for 1 min at 50 �C and then programmed at
5 �C min)1 to 310 �C. The final temperature was held for
9 min.
Microbial physiology
Enrichments and isolations were carried out to investigate the
physiology of organisms within the material and to determine
their degree of halotolerance or halophily. Cyanobacteria were
cultured by scattering fragments of crushed gypsum (5 g in
total) from green and black colonized zones onto BG-11
(Rippka et al., 1979) agar plates (2% Bacteriological Agar No.
1; Oxoid, Basingstoke, UK) with 50 lg mL)1 cycloheximide
to prevent fungal growth. Selection for halotolerant ⁄ halo-
philic species was accomplished by scattering fragments of
material onto BG-11 plates with amended NaCl concentra-
tions of 0.05, 0.1, 0.5, 1, 2 and 4 M. Plates were incubated at
4 and 21 �C under natural day ⁄ night light cycles for 2 months
until colonies had formed.
BG-11 was found to provide a suitable medium for the
cultivation of many fungal species from the gypsum when
employed without cycloheximide. Halotolerance ⁄ halophily of
fungal species was tested in an identical manner to the cyano-
bacteria. Plates were incubated at 4 and 21 �C for 5 weeks.
Selected isolates of fungi were also grown on 2% Malt agar.
Heterotrophic bacteria were isolated by scattering frag-
ments of crushed gypsum (5 g in total) from colonized zones
onto 1 ⁄ 10 Tryptone Soya Agar (TSA) plates made with
0.06 g L)1 TSA broth (Oxoid), 0.2 g CaSO4.2H2O and 2%
Bacteriological Agar No. 1 (Oxoid). Plates were incubated at
4 and 21 �C for 5 weeks.
To test for halophily (the requirement for salt to allow
growth), any organisms that grew on the NaCl-amended
BG-11 and 1 ⁄ 10 TSA plates were subcultured onto NaCl-free
plates and colony formation was monitored after 3 weeks. To
determine the maximum halotolerance of heterotrophic bac-
teria, isolates obtained on the NaCl-amended plates were
grown on plates with 0.1, 0.5, 1, 2 and 4 M NaCl and colony
formation was monitored after 3 weeks.
To investigate for halotolerance of bacteria in the surround-
ing arctic environment, soil samples were collected �50 m
east of the outcrop. Three samples were obtained by collecting
a soil core of �5 cm depth and pooling the samples. The soil
was collected into aseptic bags and maintained at ambient
temperature. Upon return to the laboratory 2 g of this soil
was added to 100 mL of TSA broth and shaken vigorously for
�2 min. Half mL aliquots of this solution (10 and 100X dilu-
tions) were spread onto TSA plates. After 2 weeks, sixty-four
distinct colonies (selected based on varying colour or mor-
phology) were removed from the plates and spread onto TSA
plates without NaCl and plates amended with 0.1, 1, 2 and
4 M NaCl to test for halotolerance. Colony formation was
monitored after 3 weeks.
All media and other reagents were obtained from Sigma
(Sigma-Aldrich, Gillingham, UK).
Identification of isolates
Cyanobacteria were identified by morphological criteria using
bright field microscopy on a Leica DMRP fluorescence micro-
scope (Leica Microsystems) at the Open University (Milton
Keynes, UK). Fungi were identified based on morphological
criteria at CABI (Centre for Agricultural Bioscience Interna-
tional; Egham, UK).
Selected heterotrophic bacterial isolates were identified by
16S rRNA gene partial sequencing. Microbial isolates were
identified by direct PCR from colonies. DNA template was
prepared by dispersing a bacterial colony in 50 lL of 10 mM
TE buffer, pH 7.5, and freeze-thawing at )20 �C. PCRs
were performed with the primers pA (5¢-AGA GTT TGA
TCC TGG CTC AG-3¢) and pH (5¢-AAG GAG GTG ATC
CAG CCG CA-3¢). The reaction mixture contained: 5 lL
of template DNA; 200 lM dNTP; 1 lM primers; 1.25 U of
Taq DNA polymerase; 1X PCR Buffer [20 mM Tris–HCl
(pH 8.4), 50 mM KCl] and 2.5 mM MgCl (Invitrogen).
Amplification consisted of incubation at 94 �C for 15 min
(which was used to assist cell disruption). This was followed
Microbe–mineral environment and gypsum neogenesis 297
� 2010 Blackwell Publishing Ltd

by 35 cycles of 1 min at 94 �C, 1 min at 55 �C and a 1-min
extension at 72 �C, with a final extension of 5 min at
72 �C. The PCR products were run on a 0.8% (w ⁄ v) agar
gel and extracted using a GenElute Gel Extraction kit
(Sigma-Aldrich, Poole, UK) according to the manufacturer’s
instructions. DNA sequencing of the PCR products was car-
ried out directly by DNA Vision (Liege, Belgium).
Sequences (up to 750 bp) were phylogenetically classified
and their nearest 16S rRNA gene sequences identified in the
Ribosomal Database Project (RDP) using the Classifier and
Sequence Match tools.
A phylogenetic tree of Arthrobacter isolates was
constructed. A region of 507 nucleotides (Escherichia coli
numbering 210–716 covering 16S rRNA variable regions
V2–V4) was used for tree construction with putative close
phylogenetic relatives obtained in RDP. A tree was first con-
structed using the neighbour-joining method (Saitou & Nei,
1987) and Kimura 2-parameter for distance correction
(Kimura, 1980) using the PHYLIP v. 3.67 inference package
(DNADIST and NEIGHBOR). SEQBOOT (1000 bootstraps) and
CONSENSE were used to provide confidence estimates for the
tree topology for a consensus tree. This tree was used as an
input tree to obtain a maximum-likelihood tree with distance
estimates using DNAML. Trees were visualized with NJPLOT
and TREEVIEW.
Nucleotide sequence accession numbers
The partial 16S rRNA sequences of the bacterial isolates
(Table 2) were deposited in GenBank (accession numbers
FJ706080–FJ706092).
Bacterial gypsum neogenesis by Arthrobacter
To test the hypothesis that Arthrobacter species could take
part in gypsum neogenesis, Arthrobacter isolates were grown
on CaSO4.2H2O-saturated 1 ⁄ 10 TSA agar plates. Two iso-
lates that formed bright white colonies were selected on the
basis that white colonies had the greatest chance of signify-
ing gypsum precipitation. The isolates were grown on 1 ⁄ 10
TSA agar plates for 2 weeks with no CaSO4 supplement to
ensure that there was no residual gypsum. A CaSO4.2H2O
saturated 1 ⁄ 10 TSA broth was prepared by vigorous shaking
of 10 g of CaSO4.2H2O in 500 mL 1 ⁄ 10 TSA which had
been previously sterilized and cooled. The medium was left
to settle overnight at 21 �C and was then filtered through a
0.2-lm filter. The isolates were inoculated into 50 mL of
the medium in sterilized and dried glass bottles and the
bottles were incubated at 21 �C for 4 weeks. At the end of
the incubation period, 2 mL of medium was filtered onto
0.2-lm black nitrocellulose filters to dryness and the filter
was examined by FEG-SEM as described above. All experi-
ments were prepared in replicate. Replicate uninoculated
controls were prepared.
RESULTS
Characteristics of colonized gypsum
Gypsum outcrops are found widely distributed across the
Haughton impact structure (Osinski et al., 2005a). They are
often disrupted and rotated and have been in this state since
an asteroid or comet impact �39 Ma (Sherlock et al., 2005).
The outcrop studied here exhibited conspicuous impact-rota-
tion of the bedding planes (Fig. 2A,B); the bedding prior to
impact was subhorizontal with a 3–5� westerly dip. The repre-
cipitated colonized gypsum was localized along the edges of
the near-vertical bedding planes (Fig. 2C) through which
precipitation in the form of snow-melt or rain can percolate.
The surface of the outcrop exhibited other irregular gypsum
reprecipitation textures (Fig. 2D). With the unaided eye,
phototrophs displayed separation into three distinct zones.
Black regions on the surface and within the gypsum corre-
spond to highly pigmented organisms (Fig. 2C,D). Beneath
the surface at a depth of between�2 and 5 mm the organisms
formed green zones of growth, although the depth of the
zones was irregular depending on the crystallinity and density
of the gypsum precipitate (Fig. 2D). Regions of pink pigmen-
tation were much more rarely observed within the gypsum,
which were usually associated with green zones. Unlike previ-
ously studied gypsum evaporites (e.g., Oren et al., 1995),
these zones were not developed into a well-defined vertically
stratified community.
Light microscopy
Within the black-zoned regions of the gypsum were abundant
pigmented cyanobacteria of size �5–10 lm (Fig. 3D). The
cells sometimes formed small colonies, but were mainly iso-
lated coccoid forms. Within the black zones, black fungal con-
idia were observed associated with patches of clear fungal
filaments.
Within green regions of the evaporite (Fig. 2D), the
primary morphotype corresponded to Gloeocapsa ⁄ Aphanot-
hece spp. (Fig. 3A). The cells were small (�1–2 lm) and grew
in a colonial growth habit surrounded by a conspicuous polys-
accharide sheath. Very rarely cyanobacterial filaments were
also observed, which were often encrusted with presumptive
gypsum crystals (Fig. 3C). Also abundant were organisms dis-
playing Chroococcidiopsis-like morphotypes (Fig. 3B). The
cells had the appearance of single cells, but many of them were
heavily pigmented colonies. Some displayed tetrad growth
patterns. All of these morphotypes displayed autofluores-
cence.
SYBR Green I staining revealed the presence of abundant
prokaryotes (Fig. 3E,F). The organisms were often, but
not exclusively, associated with cyanobacteria and dead cell
matter. They displayed both coccoid and rod morpho-
logies.
298 C. S. COCKELL et al.
� 2010 Blackwell Publishing Ltd

Mineralogy of the habitat
Scanning electron microscopy–EDS showed that the unal-
tered evaporite comprised a consolidated matrix of calcium
sulphate (Fig. 4A). The material contained other minor com-
ponents, which EDS showed to be comprised the major ele-
ments Mg, Si, Al, Fe and K (spectrum 1). The reprecipitated
gypsum (Fig. 4B) had a greater porosity than the parent mate-
rial, which provides space for microbial colonization. Under
SEM, organisms were observed as large amorphous masses,
consistent with the colonial growth forms (Fig. 4B) observed
by bright field microscopy (Fig. 3). The reprecipitated gyp-
sum contained grains of alumino-silicate materials containing
Mg, Si, Al, Fe and K (spectrum 2), some of which were on the
surface of the gypsum, suggesting an allochthonous source.
In some newly precipitated regions, the gypsum was pure
(spectrum 3) consistent with the white appearance of the gyp-
sum by visual inspection. The integrated spectrum shown
(spectrum 4) was representative of the reprecipitated gypsum.
By XRD, the original evaporite material and the newly
precipitated material on its surface exhibited peaks corre-
sponding to gypsum (CaSO4.2H2O). No anhydrite was
detected and no other major minerals were detected (data not
shown). Minor alumino-silicate phases are inferred on the
basis of the SEM–EDS spectra.
X-ray fluorescence data are shown in Table 1. Newly pre-
cipitated gypsum was depleted in non-gypsum elements,
except magnesium and barium, in which it was enriched.
Transmission electron microscopy of gypsum sections
Transmission electron microscopy revealed the presence of
numerous cell clusters associated with the gypsum surface.
Large gypsum crystals fell out of the sections, presumably
because of a weak connection with the resin, leaving cells pre-
viously on the gypsum surface associated with white empty
zones (Fig. 5). Most cells displayed abundant EPS production
(Fig. 5A). Many cells also had small gypsum crystals, which
did survive sectioning, associated with their surfaces (Fig. 5B).
Raman spectroscopy
The major peaks associated with the newly precipitated gyp-
sum and the underlying unweathered material corresponded
A B
C D
E F
Fig. 3 Micro-organisms within the gypsum. (A–C)
Micrographs of cyanobacterial morphotypes inhab-
iting the green zone in the gypsum (scale bar
10 lm), (D) micrograph of dominant morphotype
within the back zoned regions of the gypsum (scale
bar 20 lm), (E,F) SYBR Green I staining of prokary-
otes associated with gypsum crystals and cyanobac-
teria. Presumptive heterotrophs associated with
phototrophs are highlighted by an arrow. Photo-
trophs are highlighted with a dashed arrow. The
SYBR Green I image is superposed with the back-
ground of the bright field image to show location of
gypsum crystals and phototrophs. In image (E), the
cyanobacteria are the same morphotype as in (D).
The dark pigmentation prevents visualization by
SYBR Green I in contrast to (F) where the cyanobac-
terium [morphotype as in (B)] has stained [scale bar
in (E) and (F) is 10 lm].
Microbe–mineral environment and gypsum neogenesis 299
� 2010 Blackwell Publishing Ltd

to gypsum (identical with previously reported peaks at 416,
495, 621, 671, 1006, 1136 cm)1; Sharma et al., 2009)
(Fig. 6). The major peak at 1006 cm)1 is attributed to the
symmetric stretching vibrational mode of the sulphate ion.
No peaks corresponding to other minerals were observed.
Both the reprecipitated gypsum and the unweathered parent
material showed characteristic peaks attributed to O-H
stretching bands of water at 3406 and 3498 cm)1 showing
that the parent material is predominantly gypsum, not anhy-
drite. The parent material had a pronounced, but broad, peak
at 1600 cm)1 and a shoulder at �1375 cm)1. These features
most closely correspond to the organic G (aromatic carbons)
and D bands, respectively (Schopf et al., 2002; Jehlicka et al.,
2003; Guedes et al., 2005), showing the presence of organic
material in the Ordovician parent evaporite material.
Sulphur isotopic analysis
The mean d34S for the three samples was: source evaporite,
29.38&; reprecipitated uncolonized gypsum 29.91&; colo-
nized gypsum 30.25&.
Penetration of light into the gypsum
Light was effectively attenuated through gypsum crusts.
Through a 0.9-mm-thick gypsum crust, the mean penetration
through 10 different locations at 300 nm was 0.23% (±0.12),
at 360 nm it was 0.34% (±0.18) and at 680 nm it was 0.5%
(±0.24).
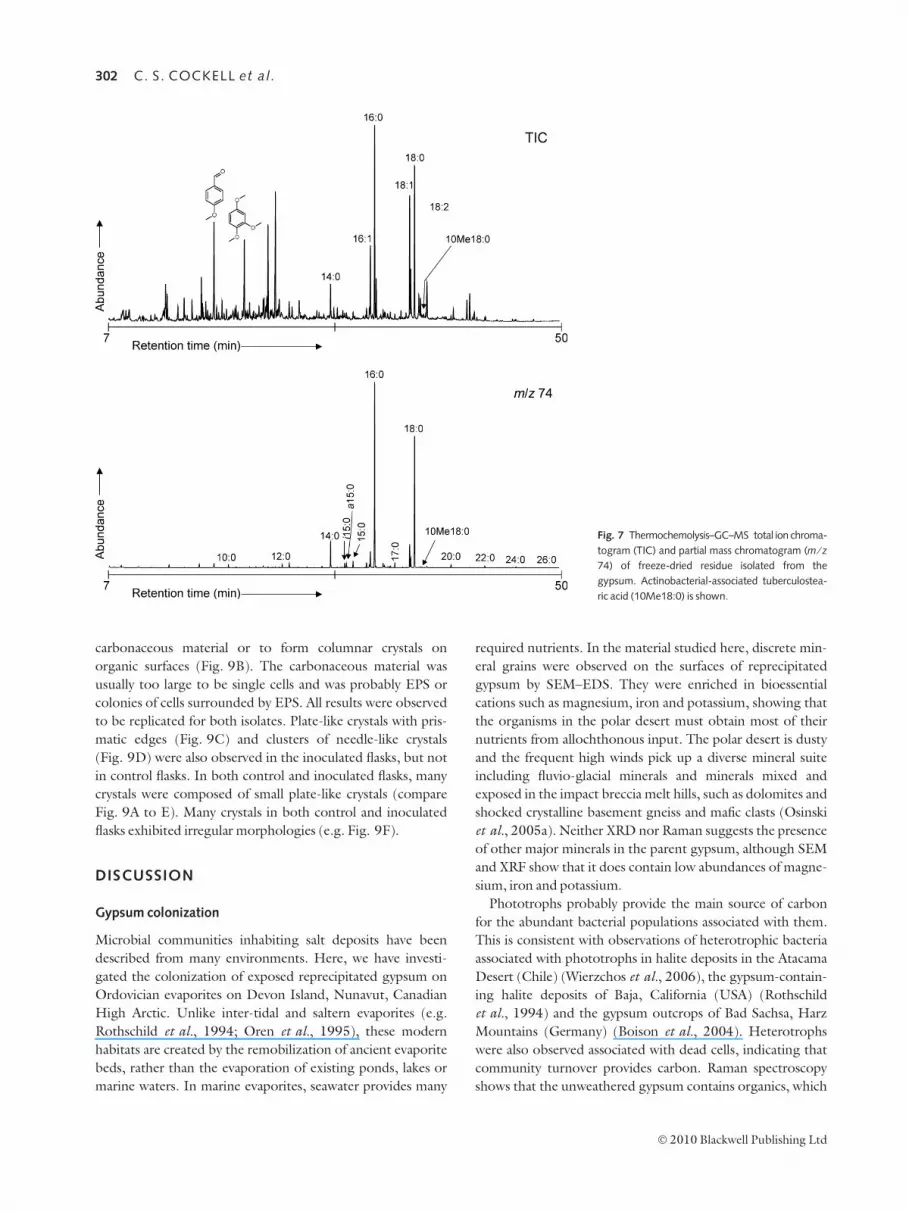
Thermochemolysis–GC–MS analysis of microbial
community
Thermochemolysis of organic material within the reprecipitat-
ed gypsum liberates a number of different compounds. The
most abundant compounds are fatty acids (as methyl esters)
which have an even predominance and range from C10 to C26.
10-methyl octanoic acid (also known as tuberculostearic acid)
is present (Fig. 7) which is a cell wall lipid produced by
Actinobacteria; the C18 homologue is also present
(O’Donnell et al., 1982; Brennan, 1988; Macnaughton &
O’Donnell, 1994). The presence of relatively high molecular
weight (>C18) fatty acids is also a common component of
actinobacterial cell walls.
Microbial isolations and physiology
On BG11 plates, cyanobacterial isolates were obtained on
plates amended with NaCl up to 0.1 M concentration, but not
on plates with 0.5 M NaCl concentration or higher. All organ-
isms obtained in NaCl-amended plates grew on unamended
plates showing that they were halotolerant, but not halophilic.
No cyanobacteria grew at 4 �C.
Diverse fungal species grew on BG-11 plates. One species
formed conspicuous and large quantities of black conidia,
which are morphologically similar to the black conidia
observed in field samples. Morphological criteria showed this
0Full scale 4416 cts
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
0Full scale 255 cts
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
0Full scale 2311 cts
1 2 3 4 5 6 7 8 0Full scale 4193 cts
1 2 3 4 5 6 7 8
1
A
2 B 3
4
Fig. 4 SEM–EDS analysis of minerals in the habitat surrounding a typical
cyanobacterial colonist. (A) A control area of unaltered Ordovician evaporitic
material under the habitat (scale bar 50 lm). EDS spectrum 1 corresponds to
unaltered material. Note presence of trace alumino-silicate material in evapo-
rite. (B) An example region of colonized gypsum (green zone in Fig. 2D; scale
bar 50 lm). Arrow is dried colonial form of cyanobacterium in rock. Spectrum 2
is of representative grains of presumptive allochthonous alumino-silicates, spec-
trum 3 is reprecipitated gypsum (note more pure form of gypsum than control
spectrum 1), spectrum 4 is the integrated spectrum of minerals around the
organisms.
Table 1 XRF data of reprecipitated gypsum from green colonized zones and
parent evaporite. Data are mean of three samples (SD < 5% for each analyte)
Analyte Reprecipitated gypsum (%) Parent evaporite (%)
Ca 25.23 26.52
SO4 68.74 70.54
BaO 3.9 –
SiO2 0.8 1.4
MgO 0.5 0.3
SrO 0.4 0.4
Al2O3 0.2 0.4
K2O 0.06 0.2
Fe2O3 0.07 0.2
Cl 0.02 0.03
300 C. S. COCKELL et al.
� 2010 Blackwell Publishing Ltd

species to most closely resemble Stachybotrys chartarum. A
common fungus observed on the plates formed septate dema-
tiaceous mycelia. It failed to sporulate and colonies reached
only 1.3 cm diameter after 50 days on 2% Malt agar plates. It
was morphologically similar to other rock-inhabiting fungi
belonging to the Herpotrichiellaceae. Fungi with black coni-
dia were obtained on plates amended with NaCl up to 1 M
concentration, but none were obtained at 2 or 4 M. No fungi
grew at 4 �C.
All heterotrophic bacteria isolated on plates saturated with
CaSO4 also grew on non-CaSO4-amended plates showing
that none required saturated CaSO4 concentrations to grow.
No heterotrophic bacteria were isolated on 4 M NaCl-
amended plates. One isolate (N5; Table 2) was isolated on
2 M NaCl-amended plates. Four isolates were obtained on
1 M NaCl-amended plates (N6–N9; Table 2). At 0.5 M NaCl,
similar morphology colonies were obtained as observed at 1 M
NaCl. At 0.1 M NaCl, plates were covered in many small red
colonies (N12; Table 2) in addition to those morphotypes
observed at higher concentrations. An actinobacterial mor-
photype was also obtained (N13; Table 2). At 0.05 M NaCl, a
variety of white and cream-coloured colonies grew in addition
to organisms observed at higher concentrations of NaCl.
None of 16 heterotrophic organisms selected from NaCl-
amended plates (N5–N9, N12, N13 and nine colonies from
the 0.05 M plates) were halophilic (all were able to grow on
unamended plates). Of the 16 heterotrophic bacteria obtained
on NaCl-amended plates and studied for their maximum salt
tolerance, all were able to grow at 0.1 M NaCl. Thirteen could
grow at 0.5 M NaCl (81%), 10 could grow at 1 M NaCl (62%),
one could grow at 2 M NaCl (N5; Table 2) and none grew at
4 M NaCl. The isolate that grew at 2 M NaCl could not grow
at 3.4 M NaCl, showing that it was moderately, but not extre-
mely, halotolerant. Four heterotrophic bacterial isolates were
obtained on plates incubated at 4 �C (N1–N4; Table 2).
These isolates were able to grow at 21 �C and grew faster than
at 4 �C, showing that they were psychrotolerant. No psychro-
philic bacteria were isolated.
Many of the isolates we obtained (Table 2) belonged to the
Arthrobacter. The partial 16S rRNA sequence phylogenetic
tree of the Arthrobacter isolates is shown in Fig. 8.
Of sixty-four heterotrophic isolates obtained from arctic
soil, all could grow at 0.1 M, 51 (81%) could grow at 0.5 M
NaCl, 29 (45%) could grow on 1 M NaCl and one isolate grew
on 2 M NaCl. None could grow at 4 M NaCl.
Bacterial gypsum neogenesis by Arthrobacter
Two Arthrobacter isolates that formed white colonies on
CaSO4-saturated TSA plates were examined for gypsum
crystal formation. In CaSO4-saturated 1 ⁄ 10 TSA solution,
gypsum crystals were observed in biological and control flasks.
However, in inoculated bottles (e.g. isolate N10; Fig. 9A,B),
gypsum crystals were observed to form along the periphery of
A B
Fig. 5 Transmission electron micrographs of
(unknown) prokaryotes associated with gypsum
surfaces. (A) Abundant extracellular polysaccharide
(EPS) (arrow) produced around two cells. The
empty area below corresponds to the location of
gypsum (scale bar 500 nm). (B) Cell showing gyp-
sum crystals (arrow) attached to the cell membrane
(cf. Fig. 3C) (scale bar 500 nm).
Fig. 6 Raman spectra of parent evaporite beds
(upper scan) and reprecipitated gypsum (lower
scan). Major peaks correspond to gypsum. Both
materials show characteristic O-H stretching bands
in water (shown to the right) at 3406 and
3498 cm)1. The parent material also shows a broad
peak at 1600 cm)1 (arrow).
Microbe–mineral environment and gypsum neogenesis 301
� 2010 Blackwell Publishing Ltd
carbonaceous material or to form columnar crystals on
organic surfaces (Fig. 9B). The carbonaceous material was
usually too large to be single cells and was probably EPS or
colonies of cells surrounded by EPS. All results were observed
to be replicated for both isolates. Plate-like crystals with pris-
matic edges (Fig. 9C) and clusters of needle-like crystals
(Fig. 9D) were also observed in the inoculated flasks, but not
in control flasks. In both control and inoculated flasks, many
crystals were composed of small plate-like crystals (compare
Fig. 9A to E). Many crystals in both control and inoculated
flasks exhibited irregular morphologies (e.g. Fig. 9F).
DISCUSSION
Gypsum colonization
Microbial communities inhabiting salt deposits have been
described from many environments. Here, we have investi-
gated the colonization of exposed reprecipitated gypsum on
Ordovician evaporites on Devon Island, Nunavut, Canadian
High Arctic. Unlike inter-tidal and saltern evaporites (e.g.
Rothschild et al., 1994; Oren et al., 1995), these modern
habitats are created by the remobilization of ancient evaporite
beds, rather than the evaporation of existing ponds, lakes or
marine waters. In marine evaporites, seawater provides many
required nutrients. In the material studied here, discrete min-
eral grains were observed on the surfaces of reprecipitated
gypsum by SEM–EDS. They were enriched in bioessential
cations such as magnesium, iron and potassium, showing that
the organisms in the polar desert must obtain most of their
nutrients from allochthonous input. The polar desert is dusty
and the frequent high winds pick up a diverse mineral suite
including fluvio-glacial minerals and minerals mixed and
exposed in the impact breccia melt hills, such as dolomites and
shocked crystalline basement gneiss and mafic clasts (Osinski
et al., 2005a). Neither XRD nor Raman suggests the presence
of other major minerals in the parent gypsum, although SEM
and XRF show that it does contain low abundances of magne-
sium, iron and potassium.
Phototrophs probably provide the main source of carbon
for the abundant bacterial populations associated with them.
This is consistent with observations of heterotrophic bacteria
associated with phototrophs in halite deposits in the Atacama
Desert (Chile) (Wierzchos et al., 2006), the gypsum-contain-
ing halite deposits of Baja, California (USA) (Rothschild
et al., 1994) and the gypsum outcrops of Bad Sachsa, Harz
Mountains (Germany) (Boison et al., 2004). Heterotrophs
were also observed associated with dead cells, indicating that
community turnover provides carbon. Raman spectroscopy
shows that the unweathered gypsum contains organics, which
Fig. 7 Thermochemolysis–GC–MS total ion chroma-
togram (TIC) and partial mass chromatogram (m ⁄ z74) of freeze-dried residue isolated from the
gypsum. Actinobacterial-associated tuberculostea-
ric acid (10Me18:0) is shown.
302 C. S. COCKELL et al.
� 2010 Blackwell Publishing Ltd

is lost in the reprecipitated gypsum. This material has either
leached away or it may also provide a source of organics for
heterotrophic micro-organisms. Another potential source of
organic material is the weathered products of dolomite (e.g.
Allen Bay Formation) in this region of Devon Island, which
contains bitumen and hydrocarbon fluid inclusions (Parnell
et al., 2007) and may be one component of the allochthonous
alumino-silicate material in the reprecipitated gypsum.
Micro-organisms do not play a major role in biogeochemi-
cal cycling of the gypsum itself, suggested by the largely
unchanged sulphur isotopic composition. This is consistent
with the aerobic environment in the evaporite which will
generally preclude sulphate reduction, although the small
enrichment in 34S might suggest localized anaerobic zones of
sulphate reduction or other mechanisms leading to loss of
light sulphur.
Physiological requirements
We were unable to isolate halophilic cyanobacteria, fungi or
heterotrophic bacteria in the gypsum. This is consistent with
theoretical expectations on the water activity (aw) in gypsum,
which provides a better measure of the biological availability
of water than solubility. Water potential is the effective water
content expressed as its mole fraction given by the van’t Hoff
equation:
w ¼ �m/CRT ; ð1Þ
where m is the number of particles in solution per mole-
cule (2 for gypsum), / is the activity coefficient (which is
assumed to be 1 for dilute solutions such as gypsum in
water), C is the concentration in mol kg)1, R is the
gas constant (8.31 J K)1 mol)1) and T is temperature in
Kelvin.
Water activity is computed from the inverse Kelvin equa-
tion:
aw ¼ expMwwRT
� �; ð2Þ
where Mw is the molecular mass of water (0.018 kg mol)1).
Substituting eqn (2) into eqn (1) gives
aw ¼ expð�MwmwCÞ: ð3Þ
The solubility of gypsum at 18 �C is 2.6 g L)1 and its molecu-
lar mass is 172.17 kg mol)1 giving a saturated solution a con-
centration of 0.015 molal (solubility drops to 2.23 g L)1 at
0 �C, which would be experienced in the Arctic, which
increases slightly the water activity) (Marion & Farren, 1997,
1999). Thus, the aw of a saturated gypsum solution at
18 �C = 0.9995.
Table 2 Identity of bacterial isolates obtained in this work
Isolate
number
Genbank
accession
number Description ⁄ Phylum Closest uncultured match (similarity) Closest cultured match (similarity)
N1 FJ706080 Psychrotolerant strain 1
Actinobacteria
DQ125753 Uncultured bacterium
clone AKAU3866 (95%)
EU029648 Arthrobacter sp. MY14015
from Minyong glacier (95%)
N2 FJ706081 Psychrotolerant strain 2
Actinobacteria
AM268326 Microbacterium
phyllosphaerae (98%)
EU468057 Uncultured bacterium clone
CE2_b03_2 (98%)
N3 FJ706082 Psychrotolerant strain 3
Actinobacteria
EF651060 Uncultured Arthrobacter
sp. clone AUVE_03D09 (91%)
EU157717 Arthrobacter sp. 31 (91%)
N4 FJ706083 Psychrotolerant strain 4
Actinobacteria
DQ125724 Uncultured bacterium
clone AKAU3822 (98%)
EF601820 Arthrobacter sp. WX4 (98%)
N5 FJ706084 Isolate from 2 M NaCl plate
Actinobacteria
FJ429561 Uncultured Streptomyces
sp. clone 2537 (100%)
FJ406045 Streptomyces albidoflavus
strain AS 4.1291 (100%)
N6 FJ706085 Isolate 1 from 1 M NaCl plate
Actinobacteria
EU536500 Uncultured bacterium
clone nbt214 h04 (99%)
AM990781 Kocuria sp. MOLA 5 (99%)
N7 FJ706086 Isolate from 1 M NaCl plate
Actinobacteria
AM167988 Uncultured Arthrobacter
sp.(98%)
AJ315069 Arthrobacter tumbae (99%)
N8 FJ706087 Isolate from 1 M NaCl plate
Actinobacteria
EF614070 Uncultured bacterium
clone NGD49 (96%)
EF471900 Arthrobacter sp. GW10-3
(96%)
N9 FJ706088 Isolate from 1 M NaCl plate
Actinobacteria
EU979035 Uncultured
actinobacterium clone g26 (92%)
DQ830985 Aestuariimicrobium
kwangyangensis strain R47 (93%)
N10 FJ706089 Gypsum neogenesis isolate 1
Actinobacteria
EF614054 Uncultured bacterium
clone NGD34 (99%)
AM933512 Arthrobacter sp. FB21
(99%)
N11 FJ706090 Gypsum neogenesis isolate 2
Actinobacteria
EU153020 Uncultured bacterium
clone G22-20 (99%)
AF479354 Glacial ice bacterium
G50-TB7 (Arthrobacter) (99%)
N12 FJ706091 Common red colony
a-proteobacteria
EF071489 Uncultured
Acetobacteraceae (98%)
AF170742 Paracraurococcus sp.
QSSC5-3 (Antarctic quartz rocks)
(97%)
N13 FJ706092 Common actinobacterium EU132912 Uncultured soil bacterium
(96%)
AY253863 Kribella flavida (97%)
Microbe–mineral environment and gypsum neogenesis 303
� 2010 Blackwell Publishing Ltd

This high water activity is equivalent to a NaCl concentra-
tion of 0.01 M and would not be expected to select for halo-
tolerance or halophily (Grant, 2004) and is not sufficiently
high to select even for slight halotolerance (0.2–0.85 M NaCl;
as defined by DasSarma & Arora, 2001).
Despite this observation, many of the heterotrophic bacte-
rial isolates were moderately halotolerant (tolerant of salt
concentration >0.85 M and <3.4 M), as were heterotrophs
from the surrounding soil, showing that halotolerance is a
widespread attribute in arctic bacteria. Statistical comparisons
between the soil and gypsum isolates are not reliable because
it is not possible to tell which isolates in the gypsum crust are
active and which ones are blown in from the surrounding
environment and are inactive; furthermore some soils in the
region contain gypsum (Bowden & Parnell, 2007). Studies
on halotolerance are generally confined to hypersaline envi-
ronments, so we do not know how widespread halotolerance
is in soil bacteria generally. In polar environments, a contribu-
tory selection pressure could plausibly be freeze-thaw cycles
that might form briny micro-environments within soils.
Although living in gypsum does not require halophily, the
habitat does select for specific organisms. The diversity of
Gloeocapsa morphotypes observed in black epilithic biofilms
on arctic rocks (Fig. 3D; Cockell et al., 2002) is not observed
in the gypsum at such high abundance and filamentous cyano-
bacteria are rare. Parnell et al. (2004) similarly observed a low
diversity of cyanobacteria belonging to Gloeocapsa in selenite
from the same region, although Nostoc sp. was commonly
observed in that material. Factors other than the requirement
for halophily are responsible for species selection. For exam-
ple, the gypsum has a lower concentration of bioessential
nutrients compared to rock surfaces; a different nutrient regi-
men might act as a selection pressure. The gypsum-encrusted
prokaryotes and cyanobacterial filaments observed by trans-
mission electron and bright field microscopy, respectively, and
the gypsum neogenesis shown in Arthrobacter isolates suggest
that growing in the presence of gypsum precipitation is a
challenge for organisms. Despite the high water activity in a
saturated gypsum solution, the material is highly porous.
These factors probably make the gypsum a desiccating envi-
ronment, which is consistent with the copious EPS produced
by many organisms observed by TEM. In principle, the anhy-
drite from which these gypsum deposits formed would also
make the habitat desiccating. Although trace quantities of this
mineral cannot be ruled out, the XRD and Raman analysis
show that it cannot be a significant contributor to desiccation
in the outcrop studied here.
No psychrophilic isolates were obtained. Tang et al. (1997)
showed that many arctic cyanobacteria had optimum meso-
philic growth temperatures, which suggests successful coloni-
zation through slow persistent growth rather than optimum
adaptation to low temperatures. Our data show that many
arctic bacteria and fungi also possess mesophilic growth
optima.
Within the gypsum, the organisms are protected from UV
radiation. In the High Arctic, the minimum zenith angle
experienced in July when the temperatures are highest and
rain water is available is 51�, so that the maximum instanta-
neous UV radiation flux is lower than in temperate or tropical
evaporitic habitats, but the 24-h photoperiod makes the
potential cumulative dose as high (Cockell et al., 2001).
Parnell et al. (2004) reported a high level of UV penetration
to organisms within selenite sheets with a reduction of only
25% through a 1-mm-thick sheet. This is caused by the poor
UV radiation absorption of calcium sulphate and the trans-
parent nature of selenite crystals. The organisms within the
habitat described here gain advantage from the precipitation
Fig. 8 Phylogenetic tree of gypsum Arthrobacter isolates and phylogenetic
relatives. A maximum-likelihood tree was generated using a neighbour-joining
tree (1000 bootstraps) as the input tree and using a mask of 507 nucleotide
positions (Escherichia coli numbering 210–716 covering V2–V4). Aquifex sp.
(accession number AB304892) was used as an outgroup. The scale bar
represents the number of changes per nucleotide position. The black arrows
correspond to Arthrobacter isolates obtained in this study. The two white-
headed arrows are the isolates used to demonstrate gypsum neogenesis.
304 C. S. COCKELL et al.
� 2010 Blackwell Publishing Ltd

of small gypsum crystals which can effectively scatter UV radi-
ation (Cockell & Raven, 2004). Our data are consistent with
Hughes & Lawley (2003) who observed the effective attenua-
tion of UV radiation in a 1.2-mm-thick Antarctic gypsum
crust (0.005% of the UVB and 0.05% of UVA). Concomi-
tantly, however, the organisms’ exposure to photosyntheti-
cally active radiation will be lower than it would be in selenite
(Parnell et al., 2004) as this region of the spectrum is also
scattered.
Gypsum neogenesis by Arthrobacter
Many of the isolates that were obtained belonged to the
Arthrobacter. Arthrobacter, Gram-positive, pleomorphic,
non-spore forming members of the Actinobacteria, are known
to be an extremophilic genus of bacteria adapted to survival in
dry, cold and oligotrophic environments (Cacciari & Lippi,
1986; Reddy et al., 2002). They have been isolated from the
surface of rocks (Gounot, 1967; Heyrman et al., 2005) and
from permafrost (Vishnivetskaya et al., 2006) and are known
to sequester metals from rocks (Brantley et al., 2001).
Arthrobacter have previously been isolated from meadow soils
on Devon Island (Nelson & Parkinson, 1978a,b). Our obser-
vations that the dominant cultured psychrotolerant and halo-
tolerant organisms in this study belonged to the Arthrobacter
are consistent with these reports. The identification of the
lipid, tuberculostearic acid, which is associated with the
Actinobacteria (Brennan, 1988; Macnaughton & O’Donnell,
1994), is consistent with the importance of Actinobacteria in
this habitat. This lipid is formed by Arthrobacter spp. (O’Don-
nell et al., 1982). The recent report that Gram-positive
Actinobacteria constitute the major component of bacterial
communities in carbonates in this region of the Arctic (Lacelle
et al., 2009) is consistent with these findings.
In view of these observations, we hypothesized that in the
polar environment Arthrobacter might be one group playing a
role in mediating mineralization of the gypsum habitat.
Microbially mediated mineralization processes are known to
occur in the arctic (Konhauser et al., 2005). The data
obtained here show that Arthrobacter species can enhance
gypsum neogenesis.
In the inoculated experiments, crystals were observed to
have formed on carbonaceous materials. They were composed
of small plate-like crystals consistent with gypsum crystals on
cell surfaces observed by TEM and bright field microscopy.
The morphology of the crystals was indistinguishable from
similar crystals observed in controls. Small crystals were
observed to have formed in both abiotic and biotic experi-
ments. Precipitation within the controls suggests either
nucleation of crystals around abiotic particles or growth of
crystals around gypsum particles <0.2 lm that passed through
the filter. This would have occurred in the inoculated bottles,
A B
C D
E F
Fig. 9 Evidence for gypsum neogenesis. (A) Forma-
tion of gypsum crystals on an organic template in
culture N10 (scale bar 10 lm). Arrows in close-up,
(B) (scale bar 4 lm), are gypsum (EDS analysis).
Smooth material underlying crystals is carbona-
ceous (EDS analysis showed only carbon and
oxygen as constituents), (C) (scale bar 10 lm)
and (D) (scale bar 50 lm) show plate-like and
needle-like crystals, respectively, which are com-
mon crystal morphologies in inoculated experi-
ments. (E) and (F) show platy and irregular crystal
morphologies, respectively, which are seen in both
inoculated and abiotic experiments (compare E to
A) (scale bar 10 lm).
Microbe–mineral environment and gypsum neogenesis 305
� 2010 Blackwell Publishing Ltd

so that some of the crystals observed in the biological experi-
ments are likely to be abiotic.
The biologically mediated gypsum neogenesis that was
observed could plausibly be triggered by two processes: (i) the
nucleation of gypsum around bacteria whereby the bacteria
merely act as particles for gypsum growth as has been observed
with cyanobacteria (Braithwaite & Whitton, 1987), or (ii) the
more specific formation of gypsum crystals on cell membranes
or EPS. A plausible mechanism might be the binding of Ca2+
to negatively charged bacterial cell surfaces (Beveridge & Fyfe,
1985; Ferris, 1990; Mitchell & Ferris, 2006) or negatively
charged moieties on EPS (Tourney & Ngwenya, 2008), caus-
ing localized supersaturation and precipitation of gypsum. A
similar mechanism was proposed for the precipitation of gyp-
sum on the cell wall of the cyanobacterium Synechococcus sp. in
Green Lake, New York (Thompson & Ferris, 1990). This
process may also cause the permanent entrapment of micro-
organisms within the reprecipitated gypsum. The fact that we
observed gypsum crystal formation on organic matter in the
biotic experiment suggests that metabolic activity is not
required to cause gypsum neogenesis.
Implications for impact crater geobiology
Finally, we note the association of the gypsum with an impact
crater. Impact events are known to have an important influ-
ence on the geomicrobiological environment. They can
increase the abundance of lithic habitats for micro-organisms
by fracturing rocks and in some instances increasing their
translucence for phototrophs (Cockell et al., 2002, 2005).
The communities observed in the gypsum deposits are associ-
ated with evaporite beds disrupted by an asteroid or comet
impact 39 Ma and now emplaced in impact breccia deposits.
The impact-induced rotation and disruption of the beds, such
as in the unit examined here, have weakened the bedding
planes, thus improving water penetration, facilitating gypsum
mobilization and subsequent colonization.
Impact hydrothermal systems, such as the one established
by the Haughton impact event (Osinski et al., 2001), remobi-
lize and precipitate salts. At Haughton, a habitat was created
in the recrystallized hydrothermally precipitated selenite (Par-
nell et al., 2004), which is colonized by cyanobaacteria, analo-
gously to the habitat described in this study.
The crater cavity created by impact events can lead to the
formation of salty, evaporitic habitats if the crater cavity ponds
water. For example, a hypersaline lake exists within the bowl
of the 50 000-year-old 1-km diameter Tswaing impact crater
in South Africa (Schoeman & Ashton, 1982; Ashton & Scho-
eman, 1983). In addition to surface habitats, the confinement
of seawater from the shallow marine Chesapeake Bay impact
�35 Ma has trapped briny habitats in the deep subsurface
(Sanford, 2005; Cockell et al., 2009; Sanford et al., 2009).
Thus, the data presented here are another instance of the
diverse ways in which impact events can influence the distribu-
tion and accessibility of salt deposits as habitats for a micro-
biota.
ACKNOWLEDGMENTS
We would like to thank Pascal Lee and the Haughton-Mars
Project for work in the field site and the Canadian Space
Agency (Canadian Analogue Research Network) for funding.
We thank Brady Carter of Decagon Devices, Pullman, WA for
discussions on water activity and Gordon Imlach, Open
University, for assistance with the FEG-SEM. Preparation of
the TEM samples was carried out by Heather Davies at the
Open University. We thank two anonymous referees who
provided helpful review comments.
REFERENCES
Ashton PJ, Schoeman FR (1983) Limnological studies on the Pretoria
Salt Pan, a hypersaline maar lake. 1. Morphometric, physical and
chemical features. Hydrobiologia 99, 61–73.
Beveridge TJ, Fyfe WS (1985) Metal fixation by bacterial cell walls.Canadian Journal of Earth Sciences 22, 1893–1898.
Bischoff L, Oskierski W (1988) The surface structure of the
Haughton Impact Crater, Devon Island, Canada. Meteoritics23, 209–220.
Boison G, Mergel A, Jolker H, Bothe H (2004) Bacterial life and
dinitrogen fixation at a gypsum rock. Applied and EnvironmentalMicrobiology 70, 7070–7077.
Bowden SA, Parnell J (2007) Intracrystalline lipids within sulfates
from the Haughton impact structure – implications for survival of
lipids on Mars. Icarus 187, 422–429.
Braithwaite CJR, Whitton BA (1987) Gypsum and halite associatedwith the cyanobacterium Entophysalis. Geomicrobiology Journal 5,
53–55.
Brantley SL, Liermann L, Bau M, Wu S (2001) Uptake of trace metalsand rare earth elements from hornblende by a soil bacterium.
Geomicrobiology Journal 18, 37–61.
Brennan PJ (1988) Mycobacterium and other actinomycetes. In
Microbial Lipids, Vol. 1 (eds Ratledge C, Wilkinson SG). AcademicPress, London, pp. 203–298.
Cacciari I, Lippi D (1986) Arthrobacters, successful arid soil bacteria.
A review. Arid Soil Research and Rehabilitation 1, 1–30.
Caumette P (1993) Ecology and physiology of phototrophic bacteriaand sulfate-reducing bacteria in marine salterns. Experientia 49,
473–481.
Caumette P, Matheron R, Raymond N, Relexans JC (1994) Microbial
mats in the hypersaline ponds of Mediterranean salterns (Salin-de-Giraud, France). FEMS (Federation of European MicrobiologicalSocieties) Microbiology – Ecology 13, 273–286.
Clark B, Baird A, Weldon R, Tsusaki D, Schnabel L, Candelaria M(1982) Chemical composition of Martian fines. Journal of Geophysi-cal Research 87, 10,059–10,068.
Cockell CS, Raven JA (2004) Zones of photosynthetic potential on
Mars and the early Earth. Icarus 169, 300–310.Cockell CS, Scherer K, Horneck G, Rettberg P, Facius R, Gugg-
Helminger A, Lee P (2001) Exposure of arctic field scientists to
ultraviolet radiation assessed using personal dosimeters. Photochem-istry and Photobiology 74, 570–578.
Cockell CS, Lee P, Osinski G, Horneck G, Broady P (2002) Impact-
induced microbial endolithic habitats. Meteoritics & PlanetaryScience 37, 1287–1298.
306 C. S. COCKELL et al.
� 2010 Blackwell Publishing Ltd

Cockell CS, Lee P, Broady P, Lim DSS, Osinski GR, Parnell J, Koeberl
C, Pesonen L, Salminen J (2005) Effects of asteroid and comet
impacts on habitats for lithophytic organisms – a synthesis. Meteor-itics & Planetary Science 40, 1901–1914.
Cockell CS, Gronstal AL, Voytek MA, Kirshtein JD, Finster K,
Sanford WE, Glamoclija M, Gohn GS, Powars DS, Horton JW(2009) Microbial abundance in the deep subsurface of the
Chesapeake Bay impact crater: relationship to lithology and impact
processes. In The ICDP-USGS Deep Drilling Project in the Chesa-peake Bay Impact Structure: Results from the Eyreville Core Holes(eds Gohn GS, Koeberl C, Miller KG, Reimold WU). GeologicalSociety of America, Special Paper 458, 941–950.
DasSarma S, Arora P (2001) Halophiles. Encyclopedia of Life Sciences.John Wiley & Sons, Malden, MA.
Davilla AF, Gomez-Silva B, de los Rios A, Ascaso C, Olivares H,
McKay CP, Wierzchos J (2008) Facilitation of endolithic micro-
bial survival in the hyperarid core of the Atacama Desert by
mineral deliquescence. Journal of Geophysical Research 113,G01028.
Dong H, Rech JA, Jiang H, Sun H, Buck BJ (2007) Endolithic cyano-
bacteria in soil gypsum, occurrences in Atacama (Chile), Mojave(United States), and Al-Jafr Basin (Jordan) deserts. Journal ofGeophysical Research 112, G02030.
Douglas S (2004) Microbial biosignatures in evaporite deposits:
evidence from Death Valley, California. Planetary and Space Science52, 223–227.
Douglas S, Yang HX (2002) Mineral biosignatures in evaporites: pres-
ence of rosickyite in an endoevaporitic microbial community from
Death Valley, California. Geology 30, 1075–1078.Edwards HGM, Jorge Villar SE, Parnell J, Cockell CS, Lee P (2005)
Raman spectroscopic analysis of cyanobacterial gypsum halotrophs
and relevance for sulfate deposits on Mars. Analyst 130, 917–923.
Ferris FG (1990) Immobilization and mineralization of metallic ions
by bacteria. Energy Sources 12, 371–375.
Garcia-Pichel F, Lopez-Cortes A, Nubel U (2001) Phylogenetic andmorphological diversity of cyanobacteria in soil desert crusts from
the Colorado Plateau. Applied and Environmental Microbiology67, 1902–1910.
Gendrin A, Mangold N, Bibring JP, Langevin Y, Gondet B, Poulet F,Bonello G, Quantin C, Mustard J, Arvidson R, LeMouelic S (2005)
Sulfates in Martian layered terrains, The OMEGA ⁄ Mars Express
view. Science 307, 1587–1591.Gounot AM (1967) Role biologique des Arthrobacter dans les limons
souterrains. Annales de l’Institut Pasteur 113, 923–945.
Grant WD (2004) Life at low water activities. Philosophical Transac-tions of the Royal Society of London 359, 1249–1267.
Guedes A, Noronha F, Prieto C (2005) Characterisation of dispersed
organic matter from lower Palaeozoic metasedimentary rocks by
organic petrography, X-ray diffraction and micro-Raman
spectroscopy analyses. International Journal of Coal Geology 62,237–249.
Heyrman J, Verbeeeren J, Schumann P, Swings J, De Vos P (2005)
Six novel Arthrobacter species isolated from deteriorated mural
paintings. International Journal of Systematic and EvolutionaryMicrobiology 55, 1457–1464.
Hughes KA, Lawley B (2003) A novel Antarctic microbial endolithic
community within gypsum crusts. Environmental Microbiology 5,555–565.
Ionescu D, Lipski A, Altendorf K, Oren A (2007) Characterization of
the endoevaporitic microbial communities in a hypersaline gypsum
crust by fatty acid analysis. Hydrobiologia 576, 15–26.Jehlicka J, Urban O, Pokorny J (2003) Raman spectroscopy of carbon
and solid bitumens in sedimentary and metamorphic rocks. Spectro-
chimica Acta Part A: Molecular and Biomolecular Spectroscopy 59,
2341–2352.
Kimura M (1980) A simple method for estimating evolutionary ratesof base substitutions through comparative studies of nucleotide
sequences. Journal of Molecular Evolution 16, 111–120.
Konhauser KO, Fyfe WS, Schultze-Lam S, Ferris FG, Beveridge TJ(2005) Iron phosphate precipitation by epilithic microbial biofilms
in Arctic Canada. Canadian Journal of Earth Sciences 31, 1320–
1324.
Lacelle D, Pellerin A, Clark ID, Lauriol B, Fortin D (2009)(Micro)morphological, inorganic-organic isotope geochemistry
and microbial populations in endostromatolites (cf. fissure
calcretes), Haughton impact structure, Devon Island, Canada:
the influence of geochemical pathways on the preservation ofisotope biomarkers. Earth and Planetary Science Letters 281,
202–214.
Langevin Y, Poulet F, Bibring JP, Gondet B (2005) Sulfates in the
north polar region of Mars detected by OMEGA ⁄ Mars Express.Science 307, 1584–1586.
Macnaughton SJ, O’Donnell AG (1994) Tuberculostearic acid as a
means of estimating the recovery (using dispersion and differentialcentrifugation) of actinomycetes from soil. Journal of Microbiologi-cal Methods 20, 69–77.
Mancinelli RL, Fahlen TF, Landheim R, Klovstad MR (2004) Brines
and evaporites, analogs for Martian life. Advances in Space Research33, 1244–1246.
Marion GM, Farren RE (1997) Gypsum solubility at subzero temper-
atures. American Journal of Soil Science Society 61, 1666–1671.
Marion GM, Farren RE (1999) Mineral solubilities in the Na-K-Mg-Ca-Cl-SO4-H2O system, a re-evaluation of the sulfate chemistry in
the Spencer-Møller-Weare model. Geochimica et CosmochimicaActa 63, 1305–1318.
Mitchell AC, Ferris FG (2006) The influence of Bacillus pasteurii on
the nucleation and growth of calcium carbonate. GeomicrobiologyJournal 23, 213–226.
Nelson LM, Parkinson D (1978a) Growth characteristics of three bac-terial isolates from an arctic soil. Canadian Journal of Microbiology24, 909–914.
Nelson LM, Parkinson D (1978b) Effect of starvation on survival of
three bacterial isolates from an arctic soil. Canadian Journal ofMicrobiology 24, 1460–1467.
O’Donnell AG, Goodfellow M, Minnikin DE (1982) Lipids in the
classification of Nocardioides, reclassification of Arthrobacter simplex(Jensen) lochhead in the genus Nocardioides (Prauser) emend.
O’Donnell et al. as Nocardioides simplex comb. nov. Archives ofMicrobiology 133, 323–329.
Oren A, Kuhl M, Karsten U (1995) An endoevaporitic microbial matwithin a gypsum crust, zonation of phototrophs, photopigments
and light penetration. Marine Ecology Progress Series 128, 151–
159.
Osinski GR, Spray JG, Lee P (2001) Impact-induced hydrothermalactivity within the Haughton impact structure: generation of a tran-
sient, warm, wet oasis. Meteoritics & Planetary Science 36, 731–
745.
Osinski GR, Lee P, Spray JG, Parnell J, Lim DSS, Bunch TE, CockellCS, Glass B (2005a) Geological overview and cratering model for
the Haughton impact structure, Devon Island, Canadian High Arc-
tic. Meteoritics & Planetary Science 40, 1759–1776.Osinski GR, Lee P, Parnell J, Spray JG, Baron MT (2005b) A case
study of impact-induced hydrothermal activity: the Haughton
impact structure, Devon Island, Canadian High Arctic. Meteoritics& Planetary Science 40, 1859–1877.
Parnell J, Lee P, Cockell CS, Osinski GR (2004) Microbial coloniza-
tion in impact-generated hydrothermal sulphate deposits, Haugh-
Microbe–mineral environment and gypsum neogenesis 307
� 2010 Blackwell Publishing Ltd

ton impact structure, and implications for sulphates on Mars. Inter-national Journal of Astrobiology 3, 247–256.
Parnell J, Bowden SA, Osinski GR, Lee P, Green P, Taylor C, BaronM (2007) Organic geochemistry of impactites from the Haughton
impact structure, Devon island, Nunavut, Canada. Geochimica etCosmochimica Acta 71, 1800–1819.
Reddy GSN, Prakash JSS, Matsumoto GI, Stackebrandt E, Shivaji S
(2002) Arthrobacter roseus sp. nov., a psychrophilic bacterium iso-
lated from an Antarctic cyanobacterial mat sample. InternationalJournal of Systematic and Evolutionary Microbiology 52, 1017–1021.
Rieder R, Economou T, Wanke H, Turkevich A, Crisp J, Bruckner J,
Dreibus G, McSween HY Jr (1997) The chemical composition of
martian soil and rocks returned by the mobile Alpha Proton X-raySpectrometer: preliminary results from the X-ray mode. Science278, 1771–1778.
Rippka R, Deruelles J, Waterbury JB, Herdman M, Stanier RY
(1979) Generic assignments, strain histories and properties ofpure cultures of cyanobacteria. Journal of General Microbiology111, 1–61.
Rothschild LJ, Giver LJ, White MR, Mancinelli RL (1994) Metabolicactivity of microorganisms in evaporites. Journal of Phycology 30,
431–438.
Saitou N, Nei M (1987) The neighbor-joining method, a new
method for reconstructing phylogenetic trees. Molecular Biologyand Evolution 4, 406–425.
Sanford WE (2005) A simulation of the hydrothermal response to the
Chesapeake Bay bolide impact. Geofluids 5, 185–201.
Sanford WE, Voytek MA, Powars DS, Jones BF, Cozzarelli IM, Cock-ell CS, Eganhouse RP (2009) Pore-water chemistry from the
ICGP-USGS corehole in the Chesapeake Bay impact structure –
implications for paleohydrology, microbial habitat, and waterresources. In The ICDP-USGS Deep Drilling Project in the Chesa-peake Bay Impact Structure: Results from the Eyreville Core Holes(eds Gohn GS, Koeberl C, Miller KG, Reimold WU). GeologicalSociety of America, Special Paper 458, 867–890.
Schoeman FR, Ashton PJ (1982) The diatom flora of the Preto-
ria Salt Pan, Transvaal, Republic of South Africa. Bacillaria 5,
63–99.
Schopf JW, Kudryavtsev AB, Agresti DG, Wdowiak TJ, Czaja AD
(2002) Laser-Raman imagery of Earth’s earliest fossils. Nature416, 73–76.
Sharma SK, Misra AK, Lucey PG, Lentz RCF (2009) A combined
Raman and LIBS instrument for characterising minerals with
532 nm laser excitation. Spectrochimica Acta Part A: Molecularand Biomolecular Spectroscopy 73, 468–476.
Sherlock SC, Kelley SP, Parnell J, Green P, Lee P, Osinsk GR, Cockell
CS (2005) Re-evaluating the age of the Haughton impact event.
Meteoritics & Planetary Science 40, 1777–1787.Sørensen KB, Canfield DE, Teske AP, Oren A (2005) Community
composition of a hypersaline endoevaporitic microbial mat. Appliedand Environmental Microbiology 71, 7352–7365.
Squyres SW, Knoll AH, Arvidson RE, Clark BC, Grotzinger JP, JolliffBL, McLennan SM, Tosca N, Bell JF III, Calvin WM, Farrand WH,
Glotch TD, Golombek MP, Herkenhoff KE, Johnson JR, Kling-
elhofer G, McSween HY, Yen AS (2006) Two years at Meridiani
Planum: results from the Opportunity Rover. Science 313, 1403–1407.
Stivaletta N, Barbieri R, Marinangeli L, Ori GG, Bosco M (2005)
Fossil endolithic cyanobacteria in evaporites: implications forastrobiological exploration. LPSC Abstracts XXXVI, 1354.
Tang EPY, Tremblay R, Vincent WF (1997) Cyanobacterial
dominance of polar freshwater ecosystems. Are high-latitude
mat-formers adapted to low temperature? Journal of Phycology33, 171–181.
Thompson JB, Ferris FG (1990) Cyanobacterial precipitation of gyp-
sum, calcite, and magnesite from natural alkaline lake water. Geology18, 995–998.
Tourney J, Ngwenya BT (2008) Bacterial EPS as a mediator of cal-
cium carbonate morphology and polymorphism. MineralogicalMagazine 72, 291.
Vishnivetskaya TA, Petrova MA, Urbance J, Ponder M, Moyer CL,
Gilichinsky DA, Tiedje JM (2006) Bacterial community in ancient
Siberian permafrost as characterised by culture and culture-inde-
pendent methods. Astrobiology 6, 400–414.Wierzchos J, Ascaso C, McKay CP (2006) Endolithic Cyanobacteria
in Halite Rocks from the Hyperarid Core of the Atacama Desert
Astrobiology 6, 423–436.
308 C. S. COCKELL et al.
� 2010 Blackwell Publishing Ltd