the description of a new species of myzopoda (myzopodidae: chiroptera) from western madagascar
TRANSCRIPT

ARTICLE IN PRESS
www.elsevier.de/mambio
Please cite th
(Myzopodida
1616-5047/$ - see
doi:10.1016/j.mam
Original investigation
The description of a new species of Myzopoda(Myzopodidae: Chiroptera) from western Madagascar
By S.M. Goodman, F. Rakotondraparany and A. Kofoky
Field Museum of Natural History, Chicago, USA and WWF, Antananarivo, Departement de Biologie Animale,Universite d’Antananarivo, Antananarivo, Madagasikara Voakajy, Antananarivo, Madagascar
Receipt of Ms. 6.2.2006Acceptance of Ms. 2.8.2006
Abstract
A new species of Myzopoda (Myzopodidae), an endemic family to Madagascar that waspreviously considered to be monospecific, is described. This new species, M. schliemanni,occurs in the dry western forests of the island and is notably different in pelage coloration,external measurements and cranial characters from M. aurita, the previously described species,from the humid eastern forests. Aspects of the biogeography of Myzopoda and its apparentclose association with the plant Ravenala madagascariensis (Family Strelitziaceae) arediscussed in light of possible speciation mechanisms that gave rise to eastern and westernspecies.r 2006 Deutsche Gesellschaft fur Saugetierkunde. Published by Elsevier GmbH. All rights reserved.
Key words: Myzopoda, Madagascar, new species, biogeography
Introduction
Recent research on the mammal fauna ofMadagascar has and continues to revealremarkable discoveries. A considerable num-ber of new small mammal and primatespecies have been described in recent years(Goodman et al. 2003), and numerousother mammals, known to taxonomists,await formal description. Much of thisrecent information is based on renewedinterest in biological inventories ofthe remaining forested zones of the islandand the associated systematic studies ofspecimens obtained during these surveys. Insome cases these animals can be recognizedas new to science when first handled in thefield by specialists, while in cases of cryptic
is article as: S.M. Goodman et al.,
e: Chiroptera) from western..., Mamm
front matter r 2006 Deutsche Gesellschaft fur S
bio.2006.08.001
speciation molecular studies have been veryinformative to resolve questions of specieslimits (e.g., Olson et al. 2004; Yoder et al.2005). The bat fauna of the island is noexception – until a decade ago these animalsremained largely under studied and ongoingsurveys and taxonomic work have revealedthat they may be something on the order of30% richer in species than noted in the mostrecent treatment of this group (Peterson et al.1995). As with other mammals, in certaincases new species of bats are immediatelyrecognizable (e.g., Goodman and Cardiff2004) and in other cases molecular tools helpin defining species limits (Goodman et al.2006).
The description of a new species of Myzopoda
. biol. (2006), doi:10.1016/j.mambio.2006.08.001
augetierkunde. Published by Elsevier GmbH. All rights reserved.
Mamm. biol. ] (]]]]) ] � ]]]–]]]

ARTICLE IN PRESS
2 S.M. Goodman et al.
The chiropteran family Myzopodidae isendemic to Madagascar and as currentlyunderstood is monospecific, represented byMyzopoda aurita (Milne-Edwards andGrandidier, 1878). After the creation ofthe Myzopodidae for this species (Thomas1904), it was proposed, based on morpholo-gical grounds, to be part of the Vespertilio-noidea, which included the familiesNatalidae, Furipteridae, Thyropteridae,Vespertilionidae, Mystacinidae and Molossi-dae (Koopman 1994). More recently,using molecular studies, the Myzopodidaehas been placed in the Noctilionoidea (Teel-ing et al. 2005), which also comprises theFuripteridae, Thyropteridae, Mystacinidae,Noctilionidae, Mormoopidae and Phyllosto-midae. Only two families of bats in the worldare currently considered monospecific (Sim-mons 2005): the Craseonycteridae and theMyzopodidae.With regard to Myzopoda, Thomas (1904)referred to it as ‘‘exceedingly rare andremarkable’’, the latter point certainly asso-ciated with the large flat adhesive organs, orsuckers, attached to the thumb and hind foot;hence its vernacular name sucker-footed bat.These structures have been the subject ofdetailed morphological studies (Schliemann1970, 1971).Until about 15 years ago few observations orspecimens were available of Myzopoda. How-ever, on the basis of recent field surveys it hasbeen recorded at a variety of sites in theeastern humid forests of Madagascar (Schlie-mann and Goodman 2003; Fig. 1). There wasone previously collected specimen from Ma-hajanga, on the dry western side of the island(Schliemann and Maas 1978), whichindicated that the ecological breadth of thisgenus was greater than previously recog-nized. Over the past few years we havecaptured individuals of Myzopoda at threedifferent localities in the west. These animalsare notably different in pelage colorationthan those of the east, which lead us toinvestigate their taxonomic status. Usingdata on external, cranial and dental mor-phology we are able to document that thewestern population of Myzopoda is differentthan that of the east, and it is describedherein as a new species.
Please cite this article as: S.M. Goodman et al.,
(Myzopodidae: Chiroptera) from western..., Mamm
Material and methods
In order to examine patterns of morphologicalvariation in Myzopoda we have consulted specimenshoused in several natural history museums. Theacronyms of these institutions are: AMNH –American Museum of Natural History, New York;BMNH – The Natural History Museum, London(formerly The British Museum [Natural History]);FMNH – Field Museum of Natural History,Chicago; MNHN – Museum national d’Histoirenaturelle, Paris; PBZT – Parc Botanique et Zoolo-gique de Tsimbazaza, Antananarivo; RMNH –Rijksmuseum van Natuurlijke Histoire, Leiden;UADBA – Universite d’Antananarivo, Departe-ment de Biologie Animale, Antananarivo; UADP –Universite d’Antananarivo, Departement de Paleon-tologie, Antananarivo; USNM – National Museumof Natural History (formerly United States NationalMuseum of Natural History), Washington, DC,ZMB – Museum fur Naturkunde, HumboldtUniversitat zu Berlin [formerly Zoologisches Mu-seum], Berlin; and ZMH – Zoologisches MuseumHamburg, Hamburg. A list of Myzopoda specimensused in this study is presented as Table 1.We recorded five external measurements in milli-meters using a ruler from collected specimens beforepreparation. These included: total length, tail length,hind foot length (not including claw), ear length andforearm length. Further, we measured body mass ingrams using a spring balance. For certain specimensthese data were obtained directly from museumlabels or field catalogues, and in other cases theywere measured from liquid preserved specimens.SMG also took 10 wing and two foot measurementsusing a dial callipers from liquid preserved speci-mens: 3rd digit – metacarpal, 3rd digit – 1st phalanx,3rd digit – 2nd phalanx, 3rd digit – 3rd phalanx, 4thdigit – metacarpal, 4th digit – 1st phalanx, 4th digit –2nd phalanx, 5th digit – metacarpal, 5th digit – 1stphalanx, 5th digit – 2nd phalanx, tibia and calcar.Seven cranial or mandibular and four dentalmeasurements were made by SMG using digitalcalipers, accurate to the nearest 0.1mm: occipito-nasal length, from occipital condyles to anterior-most point of nasal bone; greatest zygomaticbreadth, width taken across zygomatic arches atthe widest point; interorbital breadth, dorsal widthat most constricted part of skull; mastoid width,greatest width across skull at mastoid processes;greatest braincase width, breadth at widest portionof braincase; lacrimal width, greatest width acrossrostrum at lacrimal projections; anterior palatalwidth (C1–C1), taken across the outer alveolarborders of the upper canines; maxillary toothrow(C1–M3), length from anterior alveolar border ofupper canine to posterior alveolar border of the
The description of a new species of Myzopoda
. biol. (2006), doi:10.1016/j.mambio.2006.08.001
ARTICLE IN PRESS
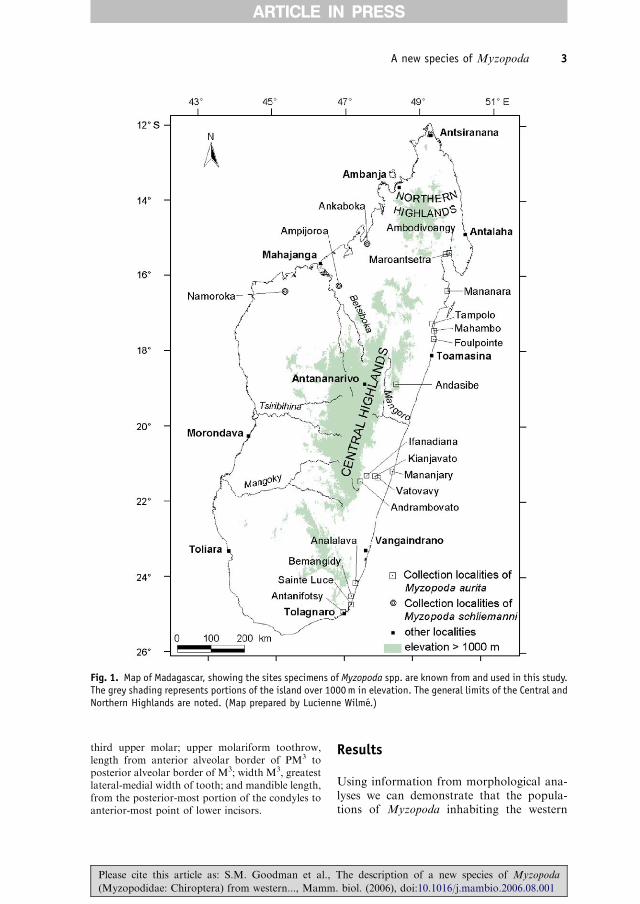
Fig. 1. Map of Madagascar, showing the sites specimens of Myzopoda spp. are known from and used in this study.The grey shading represents portions of the island over 1000 m in elevation. The general limits of the Central andNorthern Highlands are noted. (Map prepared by Lucienne Wilme.)
A new species of Myzopoda 3
third upper molar; upper molariform toothrow,length from anterior alveolar border of PM3 toposterior alveolar border of M3; width M3, greatestlateral-medial width of tooth; and mandible length,from the posterior-most portion of the condyles toanterior-most point of lower incisors.
Please cite this article as: S.M. Goodman et al.,
(Myzopodidae: Chiroptera) from western..., Mamm
Results
Using information from morphological ana-lyses we can demonstrate that the popula-tions of Myzopoda inhabiting the western
The description of a new species of Myzopoda
. biol. (2006), doi:10.1016/j.mambio.2006.08.001

ARTICLE IN PRESS
Table 1. List of Myzopoda specimens examined (see Material and methods for a listing of museum acronyms).
Specimens of Myzopoda auritaProvince de Fianarantsoa: Andrambovato (USNM 449282); Ifanadiana (USNM 449315); Kianjavato (AMNH
257130; FMNH 185227; UADP 226; USNM 448886, 448929, 448930, 448931, 448932, 449283, 449285);Mananjary (MNHN 1907.618)
Province de Toamasina: Ambodivoangy (MNHN 1985.434), Andasibe (FMNH 184490); Foulpointe (FMNH183990); Mahambo (RMNH 26117); Mananara (ZMB 5866); Maroantsetra (MNHN 1985.654); Tampolo (FMNH165454, 179203, 179204, 179273), ‘‘Tamatave’’ ½¼ Toamasina� (BMNH 99.11.3.5; ZMH T1488, T1489, T1840,ZMB 23584)
Province de Toliara: Analalava (USNM 577065); Bemangidy (FMNH 85237, 92832); Sainte Luce (UADBA RBJ-200); USNM 578740, 578742, 578743, 578856, 578857, 578858)
No precise locality: ‘‘Madagascar’’ (MNHN 1997.770, holotype; ZMB 22843; ZMH T1487).
Specimens of Myzopoda schliemanniProvince de Mahajanga: SF d’Ampijoroa (FMNH 177327, holotype); Ankaboka (FMNH 185790, 185791,
185793, 185794); Parc National de Namoroka (UADBA RBJ 203, RBJ 204); ‘‘Majunga ½¼Mahajanga�, sur lacote ouest’’ (MNHN 1947.292).
4 S.M. Goodman et al.
and eastern portions of Madagascar aredifferent from one another. Even thoughthe type specimen of M. aurita (MNHN1997.770) bears the vague designation ofbeing from ‘‘Madagascar’’, with no furtherdetails in the type description (Milne-Ed-wards and Grandidier 1878), this specimen,based on color and external measurements(see Comparisons section), is from the east.Hence, the western individuals represent anew species that is described here.
Fig. 2. Different views of holotype of Myzopodaschliemanni (FMNH 177327). Upper left – dorsal viewof cranium, upper right – ventral view of cranium andlower – lateral view of cranium and mandible.(Photograph taken by John Weinstein, Field Museumimage number Z94433-05d.)
Myzopoda schliemanni new species
Holotype: FMNH 177327, male specimenprepared as skin, skull and partial post-cranial skeleton. Field number SMG 13624.Collected by S. M. Goodman on 18 April2003 at Province de Mahajanga [Parc Na-tional d’Ankarafantsika], SF [Station Forest-iere] d’Ampijoroa, Jardin Botanique A,16119.40S, 46148.40E, 160m (Fig. 1). The skinspecimen is in good condition; although as ithad considerable subcutaneous fat, there wassome seepage of grease onto the dorsal andlower ventral pelage. The basisphenoid-ba-sioccipital suture is fully ossified and it has afull permanent dentition. The left portion ofthe rostrum is partially broken at the level ofthe lacrimal canal and the winged-portions ofthe pterygoids are broken (Fig. 2).Paratypes: Three males and four females andall are adult, except as noted. MNHN 1947.
Please cite this article as: S.M. Goodman et al.,
(Myzopodidae: Chiroptera) from western..., Mamm
292, female, ‘‘Majunga, sur la cote ouest’’, 12June 1940; UADBA RBJ 203 and 204, femaleand male (respectively), Province de Maha-janga, Parc National de Namoroka, An-driabe, about 5 km south of Namoroka(village), 16124030.600 and 45118039.500, 110m,
The description of a new species of Myzopoda
. biol. (2006), doi:10.1016/j.mambio.2006.08.001

ARTICLE IN PRESS
A new species of Myzopoda 5
13 October 2004; FMNH 185790, 185791,male and sub-adult female (respectively),Province de Mahajanga, Ankaboka,15102.240S, 47147.130E, 200m, 4 June 2000;FMNH 185793, 185794, male and female(respectively), Province de Mahajanga,Ankaboka, 15102.240S, 47147.130E, 200m,5 June 2000.Etymology: The species name has beenchosen to honor Professor Dr. HaraldSchliemann, University of Hamburg, inrecognition of his long interest in sucker-footed bats, including several importantanatomical studies (e.g., Schliemann 1970,1971). Professor Schliemann has recentlyretired from his university post after a longand fruitful career, including being theDirector of the Zoologisches Institut undZoologisches Museum, Hamburg. We sug-gest Schliemann’s sucker-footed bat as thevernacular name of this species and easternsucker-footed bat for M. aurita.Distribution: Myzopoda schliemanni isknown from three precise localities in thedry deciduous forest zone of western Mada-gascar from Ankaboka at the northern limitof its distribution to Namoroka at thesouthern limit (Fig. 1). These localities areapproximately 300 km apart and in lowlandhabitat. The holotype was obtained atAmpijoroa, which is also a lowland site andabout 80 km inland direct distance from thecoastal town of Mahajanga. The fourthcollection locality, of the Paris specimen(MNHN 1947. 292), is vague. On the basisof extrapolation of its preferred habitat type(see below), it is likely that this species has abroader distribution in the dry deciduouszones of the west, particularly further northto perhaps at least the Sambirano region inthe vicinity of Ambanja.Measurements of holotype: Measurementsare in mm and the body mass in g. Totallength, 107; tail length, 44; hind foot length,6; ear length, 30; forearm, 47; weight, 9.9;occipitonasal length, 14.9; greatest zygomaticbreadth, 11.7; interorbital breadth, 4.1; mas-toid width, 9.8; greatest braincase width, 8.8;lacrimal width, 5.9; anterior palatal width(C1–C1), 4.0; maxillary toothrow (C1–M3),5.9; upper molariform toothrow, 4.3; widthM3, 2.0; mandible length, 11.0; 3rd digit –
Please cite this article as: S.M. Goodman et al.,
(Myzopodidae: Chiroptera) from western..., Mamm
metacarpal length, 37.2; 3rd digit – 1stphalanx length, 16.5; 3rd digit – 2nd phalanxlength, 16.6; 3rd digit – 3rd phalanx length,11.7; 4th digit – metacarpal length, 36.0; 4thdigit – 1st phalanx length, 13.6; 4th digit –2nd phalanx length, 14.0; 5th digit – meta-carpal length, 37.2; 5th digit – 1st phalanxlength, 11.5; 5th digit – 2nd phalanx length,8.5; tibia length, 15.0; calcar length, 10.9.Description: Myzopoda schliemanni is arelatively small microchiropteran bat with aforearm length of 45–49mm. Like the othermember of this genus it has very prominentfunnel-shaped ears, which are not joined attheir base, and the external ear conchattaches close to the base of the mouth.There is an odd mushroom-shaped process atthe base of each ear that is probably thetragus (Thomas 1904; cf. Koopman 1994).Distinct large flat adhesive organs, or suck-ers, are attached to the thumb and hind foot.Second digit of wing reduced to the meta-carpal and a small phalanx. Tail elongatedand reaches terminus of extensive uropata-gium.The hairs of the dorsum have buff-browndistal tips merging to moderate mouse-graybases. The venter is a uniform light mouse-gray. The posterior portion of the braincasetends to be less bulbous than in M. aurita, thepalatine foramen is constricted and with anarrow ‘‘v-shape’’, the nasal sulcus is shallowand relatively broad, the nasals are narrowand rectangular in shape and the frontals arenot notably inflated. Dental formula in adultsis 2/3–1/1–3/3–3/3. It is notably smallerin several external measurements thanM. aurita.Variation: t-Tests were conducted of theexternal, cranial and dental measurementsof adult M. aurita (maximum n ¼ 36) andM. schliemanni (maximum n ¼ 7), andneither species was found to be sexuallydimorphic. In all subsequent comparisonsthe sexes are combined for each species.Considerable variation, as reflected by stan-dard deviation, was found in certain externalmeasurements for M. aurita (Tab. 2). Whenthe measurements are restricted to a singlecollector there is a notable reduction in thestandard deviation of total length and hindfoot length, while other measurements, such
The description of a new species of Myzopoda
. biol. (2006), doi:10.1016/j.mambio.2006.08.001

ARTICLE IN PRESS
Please cite this article as: S.M. Goodman et al., The description of a new species of Myzopoda
(Myzopodidae: Chiroptera) from western..., Mamm. biol. (2006), doi:10.1016/j.mambio.2006.08.001
Table
2.
Exte
rnal
mea
sure
men
ts(i
nm
m)
and
body
mas
s(i
ng)
ofM
yzop
oda
spp.
Sam
ples
are
ofad
ult
san
ddiv
ided
bet
wee
nea
ster
nan
dw
este
rnlo
caliti
es.
Des
crip
tive
stat
isti
cspre
sente
das
mea
n7
stan
dard
devi
atio
n,
min
imum
–m
axim
um
,n.
Sam
ples
mea
sure
dby
asi
ngle
fiel
dco
llec
tor
(SM
G)
are
init
alic
s.t-
Test
com
par
isons
are
pre
sente
dbet
wee
nth
eco
mple
tew
este
rnan
dea
ster
nsa
mple
s.M
easu
rem
ents
ofM
.au
rita
holo
type
wer
em
easu
red
direc
tly
from
liqu
idpre
serv
edsp
ecim
en.
Tota
lle
ngth
Tail
length
Hin
dfo
ot
length
Ear
length
Fore
arm
length
Body
mas
s
Myz
opod
asc
hliem
anni
Holo
type
FMNH
17732
7107
44
630
47
9.9
Com
bined
102.37
8.9
645.67
1.3
65.77
0.5
830.97
0.7
547.47
1.3
09.37
1.3
4sa
mple
s92–10
7,
n¼
344–47,
n¼
75–6,
n¼
630–32
,n¼
745–49,
n¼
77.8
–10
.3,
n¼
3
Myz
opod
aau
rita
Holo
type
MNH
N1997.
770
—51
635
48
—
Com
bined
110.37
4.4
746.97
3.0
37.47
1.7
033.97
1.4
548.87
1.3
78.67
0.8
5sa
mple
s101–1
18,
n¼
26
40–53,
n¼
36
5–101
,n¼
30
31–39
,n¼
27
46–52,
n¼
31
7.2
–10
.5,
n¼
26
112.77
1.5
347.37
3.0
65.37
0.5
832.77
1.1
547.77
1.5
29.37
0.2
5111–1
14,
n¼
344–50,
n¼
35–6,
n¼
332–34
,n¼
346–49,
n¼
39.0
–9.
5,
n¼
3
t¼
2:05,df¼
27
n.s
.t¼
2:07,df¼
32
t¼
2:03,df¼
36
n.s
.P¼
0:01
P¼
5:7�6
P¼
0:007
3rd
dig
it–
met
acar
pal
3rd
dig
it–
1st
phal
anx
3rd
dig
it–
2nd
phal
anx
3rd
dig
it–
3rd
phal
anx
4th
dig
it–
met
acar
pal
4th
dig
it–
1st
phal
anx
Myz
opod
asc
hliem
anni
Holo
type
FMNH
17732
737.2
16.5
16.6
11.7
36.0
13.6
Com
bined
36.07
1.0
816.87
0.5
718.27
0.7
411.87
1.9
135.67
1.4
414.17
0.3
9sa
mple
s34.2
–37.2
,n¼
816.1
–17.8
,n¼
816.6
–18.7
,n¼
810.6
–13.8
,n¼
733.5
–37.7
,n¼
813.6
–14.7
,n¼
8
Myz
opod
aau
rita
Holo
type
MNH
N1997.
770
38.2
18.0
20.5
12.3
38.7
14.7
6 S.M. Goodman et al.

ARTICLE IN PRESS
Please cite this article as: S.M. Goodman et al., The description of a new species of Myzopoda
(Myzopodidae: Chiroptera) from western..., Mamm. biol. (2006), doi:10.1016/j.mambio.2006.08.001
Com
bin
ed37.57
1.0
117.37
0.5
918.67
0.9
212.47
0.8
136.97
1.1
114.57
0.3
9sa
mple
s35.6
–39.0
,n¼
17
16.3
–18.6
,n¼
19
17.0
–20.5
,n¼
19
11.0
–13.7
,n¼
18
34.97
38.7
,n¼
17
13.5
–16.5
,n¼
19
t¼
2:07,df¼
23
n.s
.n.s
.n.s
.t¼
2:07,df¼
23
t¼
2:06,df¼
25
P¼
0:005
P¼
0:02
P¼
0:04
4th
dig
it–
2nd
phal
anx
5th
dig
it–
met
acar
pal
5th
dig
it–
1st
phal
anx
5th
dig
it–
2nd
phal
anx
Tibia
Calc
ar
Myz
opod
asc
hliem
anni
Holo
type
FMNH
17732
714.0
37.2
11.5
8.5
15.0
10.9
Com
bin
ed15.57
0.7
736.77
1.5
611.67
0.4
810.37
0.9
916.27
1.2
113.27
1.7
7sa
mple
s14.0
–16.6
,n¼
834.7
–38.9
,n¼
810.8
–12.4
,n¼
88.5
–11
.6,
n¼
814.8
–18.4
,n¼
810.9
–16.1
,n¼
8
Myz
opod
aau
rita
Holo
type
MNH
N1997.7
70
16.3
38.4
12.7
10.8
16.4
14.0
Com
bin
ed15.07
0.8
738.17
1.1
311.67
0.5
010.37
0.6
217.27
0.6
114.07
1.0
8sa
mple
s13.5
–16.5
,n¼
19
36.0
–40.0
,n¼
17
10.7
–12.7
,n¼
19
8.9
–11
.7,
n¼
19
16.0
–17.9
,n¼
15
12.6
–16.1
,n¼
14
n.s
.t¼
2:07,df¼
23
n.s
.n.s
.t¼
2:08,df¼
21
n.s
P¼
0:02
P¼
0:01
1Onthebasisofthis
rangeandassociatedvariance
itcanbeassumed
thatthesemeasurements
are
amixture
ofthose
withandwithoutthehindclaw.
Statisticaltestsnotconducted.
A new species of Myzopoda 7

ARTICLE IN PRESS
8 S.M. Goodman et al.
as tail length, continue to show the same levelof variation (Tab. 2). Within M. schliemannithere is little variation in the dorsal buff-brown and ventral mouse-gray pelage colora-tion or in cranial and dental measurementsamongst the specimens obtained at severaldifferent localities. The pelage coloration ofM. aurita is a consistent dark brown.Comparisons: In the holotype skin specimenof M. schliemanni (FMNH 177327) fromAmpijoroa, the dorsum fur, which is longerand shaggier than in M. aurita, has buff-brown distal tips merging to moderatemouse-gray bases. The ventrum is a uniformlight mouse-gray (Fig. 3). In contrast, thepelage of M. aurita is characterized bybeing a uniform dark brown, bordering onblackish, on both the ventrum and dorsum(Fig. 3); although a few individuals have adark golden-brown tinge to the pelage. In thespecimens of M. schliemanni from Ankabokaand Namoroka, all of which were preservedin formaldehyde and subsequently trans-ferred to ethanol, the pelage coloration isconsistent with the holotype of M. schlie-manni and distinctly lighter than in indivi-duals of eastern M. aurita.Even though the holotype of M. aurita hasbeen stored in alcohol for over 130 years, thepelage is not completely foxed and the colorremains notably darker than individualsof this genus from western Madagascar.Further, the specimen’s external measure-ments fall within the range of the easternspecies (Tab. 2). Hence, we conclude that theholotype represents M. aurita as definedherein.The external measurements of M. schliemanniare distinctly smaller than M. aurita (Tab. 2).For total length, hind foot length, ear lengthand forearm length these differences arestatistically significant. Perhaps most notablein this regard is ear length, which shows lessoverlap between the two species than theother external measurements. Several of thewing bone measurements show statisticallysignificant differences between the two spe-cies, and in each case it is M. schliemanni thatis smaller than M. aurita (Tab. 2).The two species show broad overlap incranial and dental measurements (Tab. 3)and none of these comparisons are statisti-
Please cite this article as: S.M. Goodman et al.,
(Myzopodidae: Chiroptera) from western..., Mamm
cally significant. The tooth formula is iden-tical in both species. Further, no notabledifference was found in tooth or cusp shape.In M. schliemanni the cranium tends tobe slightly less bulbous in appearance thanin M. aurita. The palatine foramen inM. schliemanni is notably more constrictedand a narrow ‘‘v-shape’’ as compared to themore open and broad ‘‘U-shape’’ in M. aurita(Fig. 4). Further, there are distinct differencesin aspects of the rostral morphology in thetwo species (Fig. 5). In M. aurita the nasalsulcus is distinctly narrower and deeper thanin M. schliemanni. Further, in M. schliemannithe nasals form a largely parallel structure,while in M. aurita they are expanded poster-iorly and then constricted. The frontals arenotably more inflated in M. aurita than inM. schliemanni. The holotypes of M. schlie-manni and M. aurita were directly comparedand these differences are consistent betweenall adult specimens of the two taxa.Principal component analyses were con-ducted on external, wing bone, cranial, anddental measurements of the specimens ofMyzopoda spp. Of these four comparisons itwas only the external measurements thatshowed a clear separation of M. schliemanniand M. aurita (Fig. 6), in which the first twofactors explained 69.7% of the variance. Earlength and forearm length had the heaviestloadings (Tab. 4). These results are concor-dant with the univariate analyses.Natural history: It has been noted in theliterature that Myzopoda aurita is a denizenof moist tropical forests and is often foundassociated with the large broad-leaf plants,most notably Ravenala madagascariensis ofthe Family Strelitziaceae (Schliemann andMaas 1978; Koopman 1994; Gopfert andWasserthal 1995). This tree with banana-likeleaves, reaching 10–15m tall, often growsnaturally in areas with moist ground. Rave-nala is also one of the more importantpioneering plants in heavily degraded habi-tats, particularly the eastern biome, afterforest clearing and the successive passage offire (Blanc et al. 2003). It is presumed that thesucker-like structures on this bat’s wrists andankles are used to adhere to slick flat leaf-surfaces, aiding the bat in climbing up theleaves or adhering to a roosting position. In
The description of a new species of Myzopoda
. biol. (2006), doi:10.1016/j.mambio.2006.08.001

ARTICLE IN PRESS
Please cite this article as: S.M. Goodman et al., The description of a new species of Myzopoda
(Myzopodidae: Chiroptera) from western..., Mamm. biol. (2006), doi:10.1016/j.mambio.2006.08.001
Fig. 3. Image above, photograph ventral view of holotype of Myzopoda schliemanni (FMNH 177327) obtained on18 April 2003 in the Station Forestiere d’Ampijoroa, Parc National d’Ankarafantsika. (Photograph taken by StevenM. Goodman.) Image below, photograph ventral view of typical eastern M. aurita captured at Tampolo.(Photograph taken by Achille P. Raselimanana.)
A new species of Myzopoda 9
ARTICLE IN PRESS
Please cite this article as: S.M. Goodman et al., The description of a new species of Myzopoda
(Myzopodidae: Chiroptera) from western..., Mamm. biol. (2006), doi:10.1016/j.mambio.2006.08.001
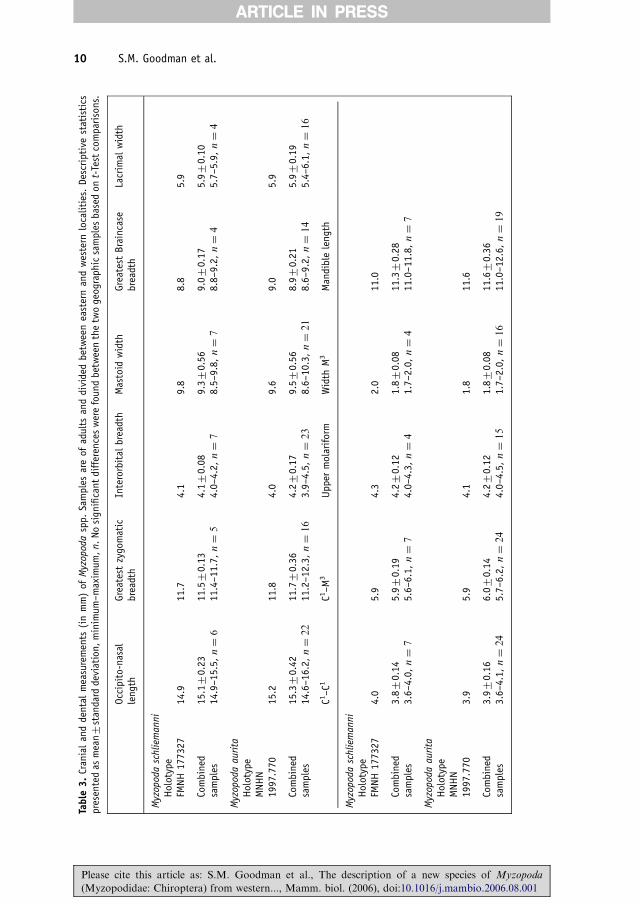
Table
3.
Cran
ial
and
denta
lm
easu
rem
ents
(in
mm
)of
Myz
opod
asp
p.
Sam
ple
sar
eof
adult
san
ddiv
ided
bet
wee
nea
ster
nan
dw
este
rnlo
caliti
es.
Des
crip
tive
stat
isti
cspre
sente
das
mea
n7
stan
dard
devi
atio
n,m
inim
um
–m
axim
um
,n.No
signifi
cant
dif
fere
nce
sw
ere
found
bet
wee
nth
etw
oge
ogr
aphic
sam
ple
sbas
edon
t-Te
stco
mpar
isons.
Occ
ipit
o-n
asal
length
Gre
ates
tzy
gom
atic
bre
adth
Inte
rorb
ital
bre
adth
Mas
toid
wid
thGre
ates
tBra
inca
sebre
adth
Lacr
imal
wid
th
Myz
opod
asc
hliem
anni
Holo
type
FMNH
17732
714.9
11.7
4.1
9.8
8.8
5.9
Com
bined
15.17
0.2
311.57
0.1
34.17
0.0
89.37
0.5
69.07
0.1
75.97
0.1
0sa
mple
s14.9
–15.5
,n¼
611.4
–11.7
,n¼
54.0
–4.
2,
n¼
78.5
–9.
8,
n¼
78.8
–9.2
,n¼
45.7
–5.
9,
n¼
4
Myz
opod
aau
rita
Holo
type
MNH
N1997.
770
15.2
11.8
4.0
9.6
9.0
5.9
Com
bined
15.37
0.4
211.77
0.3
64.27
0.1
79.57
0.5
68.97
0.2
15.97
0.1
9sa
mple
s14.6
–16.2
,n¼
22
11.2
–12.3
,n¼
16
3.9
–4.
5,
n¼
23
8.6
–10
.3,
n¼
21
8.6
–9.2
,n¼
14
5.4
–6.
1,
n¼
16
C1–C1
C1–M
3U
ppe
rm
ola
rifo
rmW
idth
M3
Man
dib
lele
ngth
Myz
opod
asc
hliem
anni
Holo
type
FMNH
17732
74.0
5.9
4.3
2.0
11.0
Com
bined
3.87
0.1
45.97
0.1
94.27
0.1
21.87
0.0
811.37
0.2
8sa
mple
s3.6
–4.
0,
n¼
75.6
–6.
1,
n¼
74.0
–4.
3,
n¼
41.7
–2.
0,
n¼
411.0
–11.8
,n¼
7
Myz
opod
aau
rita
Holo
type
MNH
N1997.
770
3.9
5.9
4.1
1.8
11.6
Com
bined
3.97
0.1
66.07
0.1
44.27
0.1
21.87
0.0
811.67
0.3
6sa
mple
s3.6
–4.
1,
n¼
24
5.7
–6.
2,
n¼
24
4.0
–4.
5,
n¼
15
1.7
–2.
0,
n¼
16
11.0
–12.6
,n¼
19
10 S.M. Goodman et al.

ARTICLE IN PRESS
Fig. 4. Views of the palatine foramen in (left) Myzopoda schliemanni (Holotype FMNH 177327) and (right)M. aurita (FMNH 179273). The difference in the foramen shape in these two specimens is consistent betweenthe two species. The left pterygoid (as viewed here) of M. schliemanni is broken and bent laterally and the rightone is completely missing. (Drawing by Rebecca Kramer.)
A new species of Myzopoda 11
1948, the late Harry Hoogstraal found thisspecies in a Ravenala leaf in southeasternMadagascar, in this case a young uncoiledone (Schliemann and Goodman 2003; fieldcatalogue in the FMNH), and althoughnumerous field researchers have subsequentlysearched the leaves of this plant for Myzopo-da, this is the only direct observation ofwhich we are aware to support this associa-tion. Captive individuals readily climb upRavenala leaves (Gopfert and Wasserthal1995).An interesting plant–bat relationship occursin the genus Thyroptera, a member of theNewWorld family Thyropteridae, that showsremarkable parallel development of suctiondiscs on the wrists and ankles as found inMyzopoda. Thyroptera, composed of threedistinct species, are known to use theirsuckers to grip the slick leaves of Heliconia(Family Musaceae) and Calathea (FamilyMarantaceae), which they use as day roostsin often heavily degraded forest (Findley andWilson 1974; Vonhof and Fenton 2004).Analyses of the functional morphology ofthese appendage structures and molecularstudies of the phylogenetic relationships ofThyroptera and Myzopoda (Schliemann 1970,1971; Teeling et al. 2005) reveal that theseremarkable adaptations are convergent be-tween the two genera. Even though there isonly one direct published observation of
Please cite this article as: S.M. Goodman et al.,
(Myzopodidae: Chiroptera) from western..., Mamm
Myzopoda roosting in Ravenala leaves, con-text and inference would imply that theaxiom about the association of this bat andplant is correct. Thus, we assume that thepreferred natural day roost sites of Myzopodaare plants with broad and smooth leaves,such as Ravenala or other plants such asPandanus (Family Pandanaceae), certainspecies of palms and ferns, introduced Musa(Family Musaceae), and Typhonodorum(Family Araceae).For the specimens of M. aurita obtained inthe eastern biome and for which we havedetails on the specific situation where theywhere captured, the majority were within orclose to stands of Ravenala and generally atelevations below 50m. In a vast portion ofthe east, from at least Vangaindrano toMaroantsetra, much of the former extensivelowland forests have been cleared and densestands of Ravenala are the dominant pioneer-ing species in the resulting secondary habitat.Myzopoda has been recorded in this zone atseveral different sites and based on extra-polation presumably has a broad distributionin heavily degraded lowland eastern areas,particularly those dominated by Ravenala.Our records of Myzopoda from westernMadagascar are associated with locally mesicconditions and for most cases in closeproximity to Ravenala or other plants withbroad and smooth leaves. The holotype of
The description of a new species of Myzopoda
. biol. (2006), doi:10.1016/j.mambio.2006.08.001

ARTICLE IN PRESS
Fig. 5. Views of the upper rostrum in (left) Myzopodaschliemanni (Holotype FMNH 177327) and (right)M. aurita (FMNH 179273). The differences in the nasalsulcus, nasal shape and frontals in these two speci-mens are consistent between the two species.(Drawing by Rebecca Kramer.)
12 S.M. Goodman et al.
M. schliemanni was captured in a mist netat the edge of a marsh, near the StationForestiere d’Ampijoroa, which included Ra-venala and Raphia palms, albeit the former
Please cite this article as: S.M. Goodman et al.,
(Myzopodidae: Chiroptera) from western..., Mamm
not a common member of the local plantcommunity. The Ankaboka sites where thisspecies was netted contain highly degradedlowland areas and the animals were capturedover streams in gallery forest. AlthoughRavenala was not specifically noted in thiszone, it can be found in the general vicinity.Finally, at Namoroka both specimens werecaptured by hand while roosting in a cave(Kofoky et al. 2006) and an additionalindividual was netted over a stream in amixed marsh woodland area. The habitatimmediately surrounding the cave site wasdry deciduous forest, and about 750–1000mdeeper in the forest was a permanent riversystem with Raphia palms and large strap-leaf Pandanus. A third individual captured atNamoroka and released was obtained inhabitat heavily modified by human activitiesand included an extensive marsh system withRaphia, Pandanus and a few examples ofRavenala, and mango trees on slightly higherground.The holotype of M. schliemanni was obtainedon 18 April and had notably large deposits ofyellow sub-cutaneous fat. Further, the testi-cles were scrotal, measured 5� 3mm, and theepididymides were convoluted. The scrotumcoloration was noted as being, ‘‘dull yellowwith dark spots’’. Amongst the four speci-mens obtained at Ankaboka in early Junethere was one sub-adult with an unossifiedskull and partial non-permanent dentition.One of the animals handled at Namoroka inmid-October was a female with developedmammae. Nothing definitive can be statedabout the reproductive season in this species,but it would appear that it is not confined toa narrow period of the year.Several male specimens of M. aurita werenoted as having scrotal testes, including oneindividual on 14 March in which theymeasured 4� 3mm and individuals on 14April and 24 May that measured 5� 4mmand with convoluted epididymides. Thus, itwould appear that reproductive males retaintheir testicles in a descended position for atleast 3 months per year.In two cases of male M. aurita with enlargedtestes it was recorded in the field cataloguethat the scrotum was ‘‘light beige in colora-tion with black spots around testicles’’. It has
The description of a new species of Myzopoda
. biol. (2006), doi:10.1016/j.mambio.2006.08.001

ARTICLE IN PRESS
-1.5-1.0-0.50.00.51.01.52.02.53.0
-2.5 -1.5 -0.5 0.5 1.5 2.5-2.5-2.0-1.5-1.0-0.50.00.51.01.52.02.5
-2.5
-1.5
-0.5
0.5
1.5
2.5
-2.5 -2.0 -1.5 -1.0 -0.5 0.0 0.5 1.0 1.5 2.0-2.5-2.0-1.5-1.0-0.50.00.51.01.52.0
-3.5 -2.5 -1.5 -0.5 0.5 1.5 2.5 3.5
-2.5 -1.5 -0.5 0.5 1.5 2.5
Fig. 6. Projection of factor 1 (x-axis) and factor 2 (y-axis) in principal component analysis of external (upperleft), wing bone (upper right), cranial (lower left) and dental (lower right) measurements of Myzopoda aurita(black squares) and M. schliemanni (light squares). Loading contributions of variables to each axis are shown inTable 4.
A new species of Myzopoda 13
already been noted in the literature that maleMyzopodidae have pigmented portions oftheir external sexual organs (Kermott andTimm 1988), which in this case was scattereddark brown pigment to the tunica vaginalissurrounding the epididymis. The function ofthe colored testicular pouch is uncertain, butmay be associated with thermoregulation,visual communication, or a shield againstultraviolet radiation. Given that the pigmen-tation in Myzopoda forms an irregularpattern, rather than being equally distribu-tion across the scrotal region and seemsconfined to adult males in breeding condi-tion, this would indicate a probable socialcommunication function, rather than relatedto temperature regulation or a radiationshield.
Discussion
Given the putative association mentionedabove between Ravenala and Myzopoda, we
Please cite this article as: S.M. Goodman et al.,
(Myzopodidae: Chiroptera) from western..., Mamm
examine this relationship in finer detail tounderstand potential speciation mechanismsin Myzopoda. Blanc et al. (2003) indicatedthat the upper elevational limit of Ravenala issomewhere between 1000 and 1100m. Theresults of elevational transects of plantcommunities in the eastern humid forestsare concordant with this assessment. Rave-nala, as a member of non-disturbed forest,drops out between 800 and 1000m, and thispattern is relatively consistent across the 121latitudinal range of the eastern forests (Lewiset al. 1996; Messmer et al. 2000). However,Ravenala can be found in isolated patches,particularly in wet valley bottoms associatedwith degraded habitats (savoka), up to about1300m (Humbert 1965). This plant is com-mon along the complete length of the easternlowland area, with a distinctly more clumpeddistribution in drier zones at the northern endof the island near Antsiranana, where it canbe found in wet valley bottoms, and thenwest to the Sambirano region near Ambanja,and then becomes distinctly less common
The description of a new species of Myzopoda
. biol. (2006), doi:10.1016/j.mambio.2006.08.001
ARTICLE IN PRESS
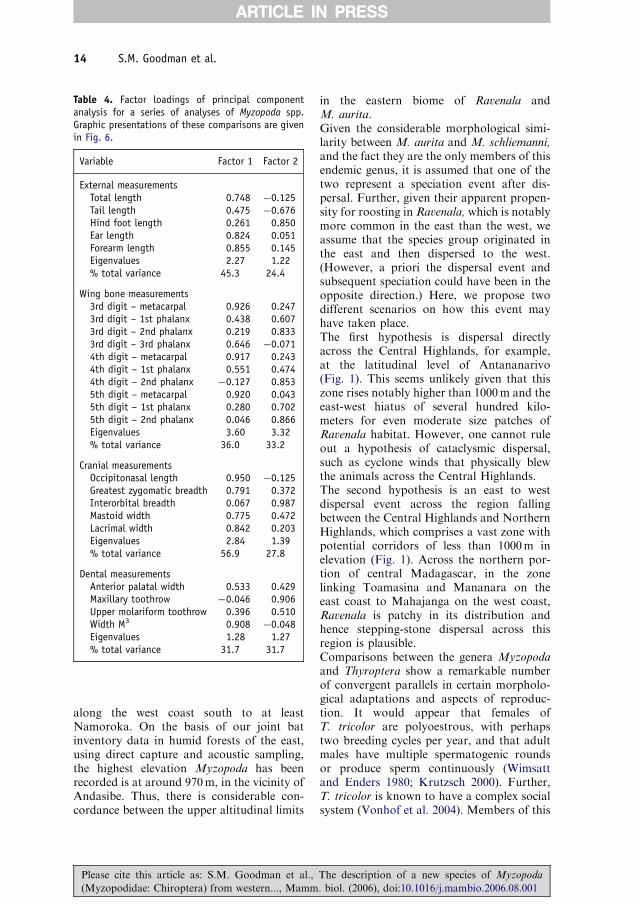
Table 4. Factor loadings of principal componentanalysis for a series of analyses of Myzopoda spp.Graphic presentations of these comparisons are givenin Fig. 6.
Variable Factor 1 Factor 2
External measurementsTotal length 0.748 �0.125Tail length 0.475 �0.676Hind foot length 0.261 0.850Ear length 0.824 0.051Forearm length 0.855 0.145Eigenvalues 2.27 1.22% total variance 45.3 24.4
Wing bone measurements3rd digit – metacarpal 0.926 0.2473rd digit – 1st phalanx 0.438 0.6073rd digit – 2nd phalanx 0.219 0.8333rd digit – 3rd phalanx 0.646 �0.0714th digit – metacarpal 0.917 0.2434th digit – 1st phalanx 0.551 0.4744th digit – 2nd phalanx �0.127 0.8535th digit – metacarpal 0.920 0.0435th digit – 1st phalanx 0.280 0.7025th digit – 2nd phalanx 0.046 0.866Eigenvalues 3.60 3.32% total variance 36.0 33.2
Cranial measurementsOccipitonasal length 0.950 �0.125Greatest zygomatic breadth 0.791 0.372Interorbital breadth 0.067 0.987Mastoid width 0.775 0.472Lacrimal width 0.842 0.203Eigenvalues 2.84 1.39% total variance 56.9 27.8
Dental measurementsAnterior palatal width 0.533 0.429Maxillary toothrow �0.046 0.906Upper molariform toothrow 0.396 0.510Width M3 0.908 �0.048Eigenvalues 1.28 1.27% total variance 31.7 31.7
14 S.M. Goodman et al.
along the west coast south to at leastNamoroka. On the basis of our joint batinventory data in humid forests of the east,using direct capture and acoustic sampling,the highest elevation Myzopoda has beenrecorded is at around 970m, in the vicinity ofAndasibe. Thus, there is considerable con-cordance between the upper altitudinal limits
Please cite this article as: S.M. Goodman et al.,
(Myzopodidae: Chiroptera) from western..., Mamm
in the eastern biome of Ravenala andM. aurita.Given the considerable morphological simi-larity between M. aurita and M. schliemanni,and the fact they are the only members of thisendemic genus, it is assumed that one of thetwo represent a speciation event after dis-persal. Further, given their apparent propen-sity for roosting in Ravenala, which is notablymore common in the east than the west, weassume that the species group originated inthe east and then dispersed to the west.(However, a priori the dispersal event andsubsequent speciation could have been in theopposite direction.) Here, we propose twodifferent scenarios on how this event mayhave taken place.The first hypothesis is dispersal directlyacross the Central Highlands, for example,at the latitudinal level of Antananarivo(Fig. 1). This seems unlikely given that thiszone rises notably higher than 1000m and theeast-west hiatus of several hundred kilo-meters for even moderate size patches ofRavenala habitat. However, one cannot ruleout a hypothesis of cataclysmic dispersal,such as cyclone winds that physically blewthe animals across the Central Highlands.The second hypothesis is an east to westdispersal event across the region fallingbetween the Central Highlands and NorthernHighlands, which comprises a vast zone withpotential corridors of less than 1000m inelevation (Fig. 1). Across the northern por-tion of central Madagascar, in the zonelinking Toamasina and Mananara on theeast coast to Mahajanga on the west coast,Ravenala is patchy in its distribution andhence stepping-stone dispersal across thisregion is plausible.Comparisons between the genera Myzopodaand Thyroptera show a remarkable numberof convergent parallels in certain morpholo-gical adaptations and aspects of reproduc-tion. It would appear that females ofT. tricolor are polyoestrous, with perhapstwo breeding cycles per year, and that adultmales have multiple spermatogenic roundsor produce sperm continuously (Wimsattand Enders 1980; Krutzsch 2000). Further,T. tricolor is known to have a complex socialsystem (Vonhof et al. 2004). Members of this
The description of a new species of Myzopoda
. biol. (2006), doi:10.1016/j.mambio.2006.08.001

ARTICLE IN PRESS
A new species of Myzopoda 15
genus also have relatively complex geogra-phical ranges (Gregorin et al. 2006). Given allof the interesting repeated convergent pat-terns between Myzopoda and Thyroptera,research on the natural history and repro-ductive system in the former genus is almostcertain to uncover some fascinating details.On the basis of limited information,M. schliemanni lives in portions of the centrallowland west where there are relatively mesicconditions, associated with marshes, rivers ortransitional humid-dry forests. Although twoof the three sites this species is knownfrom are protected areas (Parc Nationald’Ankarafantsika and Parc National deNamoroka), it appears, based on our inven-tory work, to be sparsely dispersed in thecentral western portions of the island (Good-man et al. 2005). The forest habitat at theAnkaboka sites, as well as one of the twosites at Namoroka, were notably disturbed byhuman activities and by extrapolation thisspecies appears to be resilient to certain levelsof anthropogenic perturbation. Across itsknown range, measuring about 300 km inlength, there is considerable non-continuoushabitat similar to sites in which it has beenfound, and we suspect that the threats to thisspecies’ long-term existence are not notablysevere.In their assessment of the conservation statusof the world’s Microchiroptera, Hutson et al.(2001) considered M. aurita to be a species ofspecial concern, largely associated with itbeing the sole member of an endemic familyand that little information was availableabout its distribution. Over the course ofthe past decades, largely as a result ofbiological inventories, we now know thatthis species is more broadly distributed thanpreviously realized and inhabits areas ofextensive secondary habitats of Ravenala.While it is true that M. aurita is currentlyonly known from in or around three pro-tected areas (Parc National de Marojejy, ParcNational de Masoala and Reserve Specialed’Andasibe; Pont and Armstrong 1990;Russ et al. 2001; Schliemann and Goodman2003; Tab. 1), inferred levels of threat maybe less severe then previously assumedgiven its ability to live in highly disturbedareas. Finally, based on the description of a
Please cite this article as: S.M. Goodman et al.,
(Myzopodidae: Chiroptera) from western..., Mamm
second species of Myzopoda herein, thefamily Myzopodidae is no longer monospe-cific and the sole family of bats withthis level of uniqueness in the world is theCraseonycteridae.The discovery of M. schliemanni and newinformation on the distribution and conser-vation status of M. aurita are based on recentsurvey work. In a few years considerableprogress has been made on numerous aspectsof the chiropteran fauna of the island. Thisunderlines the importance of basic scientificresearch for establishing the priorities forconservation programs and assessments ofpresumed rare and possibly endangeredanimals.
Acknowledgements
We are grateful to the Direction des Eaux etForets and Association Nationale pour laGestion des Aires protegees for issuingcollection and exportation permits. BP Con-servation Programme, Darwin Initiative(162/10/024), John D. and Catherine T.MacArthur Foundation, Rufford SmallGrants and Volkswagen Foundation pro-vided funding for portions of the fieldwork.For access to specimens under their care, weare indebted to Nancy Simmons (AMNH),Paulina Jenkins (BMNH), Jean-Marc Ponsand Ronan Kirsch (MNHN), Chris Smeenk(RMNH), Olga Ramilijaona and DanielRakotondravony (UADBA), Berthe Rako-tosamimanana (UADP), Michael Carleton(USNM), Robert Asher (ZMB) and HaraldSchliemann (ZMH). We are grateful to FanjaRatrimomanarivo and Angelos Tianarifidyfor providing some additional specimenmaterial for this study and Julie Razafima-nahaka, Daudet Andriafidison, and RoselineRampilamanana for aid with fieldwork inNamoroka. Several friends and colleaguesprepared or made available figures: RebeccaKramer (Figs. 4 and 5), Achille P. Raselima-nana (Fig. 3b), John Weinstein (Fig. 2), andLucienne Wilme (Fig. 1). Jorg Ganzhornkindly translated the summary into German.For comments on an earlier version of thisstudy we are grateful to Jon Russ andRichard Jenkins.
The description of a new species of Myzopoda
. biol. (2006), doi:10.1016/j.mambio.2006.08.001

ARTICLE IN PRESS
16 S.M. Goodman et al.
Zusammenfassung
Beschreibung einer neuen Art von Myzopoda (Myzopodidae: Chiroptera) von West-Madagaskar
Die Myzopodidae, eine endemische Familie von Fledermausen auf Madagaskar, galt bisher alsmonospezifisch. In Madagaskar wurde eine neue Art, M. schliemanni, gefunden. Diese Art aus denTrockenwaldern West-Madagaskars unterscheidet sich in der Fellfarbung, externen morphologischenMaXen und Schadelmerkmalen von M. aurita, der bisher einzig beschriebenen Art der Familie.Myzopoda aurita ist aus den immergrunen Regenwaldern des Ostens Madagaskars bekannt.Biogeographische Muster der Verteilung von Myzopoda und ihre enge Assoziation mit dem ‘‘Baumdes Reisenden (Ravenala madagascariensis; Strelitziaceae)’’ werden in Zusammenhang mitSpeziationsmechanismen diskutiert, die zur Entstehung der westlichen und ostlichen Artbeigetragen haben.r 2006 Deutsche Gesellschaft fur Saugetierkunde. Published by Elsevier GmbH. All rights reserved.
References
Blanc, P.; Hladik, A.; Rabenandrianina, N.;Robert, J.S.; Hladik, C.M. (2003): Strelitzia-
ceae: the variants of Ravenala in natural andanthropogenic habitats. In: The Natural His-tory of Madagascar. Ed. by S.M. Goodmanand J.P. Benstead. Chicago: The University ofChicago Press. Pp. 472–476.
Findley, J. S.; Wilson, D. E. (1974): Observationson the Neotropical disk-winged bat, Thyroptera
bicolor Spix. J. Mammalogy 55, 562–571.Goodman, S. M.; Cardiff, S. G. (2004): A new
species of Chaerephon (Molossidae) from Ma-
dagascar with notes on other members of thefamily. Acta Chiropterol. 6, 227–248.
Goodman, S.M.; Ganzhorn, J.U.; Rakotondrav-ony, D. (2003): Introduction to the mammals.
In: The Natural History of Madagascar. Ed. byS.M. Goodman and J.P. Benstead. Chicago:The University of Chicago Press. Pp.1159–1186.
Goodman, S.M.; Cardiff, S. G.; Ranivo, J.;
Russell, A. L.; Yoder, A. D. (2006): A newspecies of Emballonura (Chiroptera: Emballo-nuridae) from the dry regions of Madagascar.Amer. Mus. Novitates (in press).
Goodman, S. M.; Andriafidison, D.; Andrianai-voarivelo, R.; Cardiff, S. G.; Ifticene, E.;Jenkins, R. K. B.; Kofoky, A.; Mbohoahy, T.;Rakotondravony, D.; Ranivo, J.; Ratrimoma-narivo, F.; Razafimanahaka, J.; Racey, P. A.
(2005): The distribution and conservation ofbats in the dry regions of Madagascar. Anim.Conserv. 8, 153–165.
Gopfert, M. C.; Wasserthal, L. T. (1995): Notes on
echolocation calls, food and roosting behaviourof the Old World sucker-footed bat Myzopoda
aurita (Chiroptera, Myzopodidae). Z. Sauge-tierkunde 60, 1–8.
Please cite this article as: S.M. Goodman et al.,
(Myzopodidae: Chiroptera) from western..., Mamm
Gregorin, R.; Gonc-alves, E.; Lim, B. K.; En-gstrom, M. D. (2006): New species of disk-
winged bat Thyroptera and range extension forT. discifera. J. Mammalogy 87, 238–246.
Hutson, A. M.; Mickleburgh, S. P.; Racey, P.(2001): Microchiropteran Bats – Global StatusSurvey and Conservation Action Plan. Gland:
IUCN/SSC Chiroptera Specialist Group.Humbert, H. (1965): Description des types de
vegetation. In: Notice de la carte de Madagas-car. Ed. by H. Humbert and G. Cours-Darne.Travaux de la Section Scientifique et Technique
de l’Institut Franc-ais de Pondichery, hors serie6, 46–78.
Kermott, L. H.; Timm, R. M. (1988): Scrotalmelanins in bats (Chiroptera): description,
distribution and function. J. Zool. (London)214, 519–532.
Kofoky, A. F.; Andriafidison, D.; Razafimanaha-ka, H. J.; Rampilamanana, R. L.; Jenkins, R.K. B. (2006): The first observation of a
Myzopoda sp. (Myzopodidae) roosting in wes-tern Madagascar. African Bat Conserv. News-lett. 9, 5–6.
Koopman, K.F. (1994): Chiroptera: Systematics.
In: Handbook of Zoology. Vol. 8, Ed. by J.Niethammer, H. Schliemann and D. Starck.Walter de Gruyter: Berlin. Pp. 1–217.
Krutzsch, P.H. (2000): Anatomy’ physiology andcyclicity of the male reproductive tract. In:
Reproductive Biology of Bats. Ed. by E.G.Crichton and P.H. Krutzsch. San Diego:Academic Press. Pp. 91–155.
Lewis, B. A.; Phillipson, P. B.; Andrianarisata, M.;
Rahajasoa, G.; Rakotomalaza, P. J.; Randriam-
bololona, M.; McDonagh, J. F. (1996): A studyof the botanical structure, composition, anddiversity of the eastern slopes of the Reserve
The description of a new species of Myzopoda
. biol. (2006), doi:10.1016/j.mambio.2006.08.001

ARTICLE IN PRESS
A new species of Myzopoda 17
Naturelle Integrale d’Andringitra, Madagascar.In: A Floral and Faunal Inventory of the EasternSlopes of the Reserve Naturelle Integraled’Andringitra, Madagascar: With Reference toElevational Variation. Ed. by S. M. Goodman.Fieldiana: Zoology, new series 85, 24–75.
Messmer, N.; Rakotomalaza, P. J.; Gautier, L.(2000): Structure and floristic composition ofthe begetation of the Parc National de Mar-ojejy, Madagascar. In: A Floral and FaunalInventory of the Parc National de Marojejy,Madagascar: With Reference to ElevationalVariation. Ed. by S. M. Goodman. Fieldiana:Zoology, new series 97, 41–104.
Milne-Edwards, A.; Grandidier, A. (1878): Notesur un nouveau genre de Chiroptere. Bull. Soc.Philomath. Paris 2, 220–221.
Olson, L. E.; Goodman, S. M.; Yoder, A. D.(2004): Illumination of cryptic species bound-aries in long-tailed shrew tenrecs (Mammalia:Tenrecidae; Microgale), with new insights intogeographic variation and distributional con-straints. Biol. J. Linn. Soc. 83, 1–22.
Peterson, R. L.; Eger, J. L.; Mitchell, L. (1995):Chiropteres. Vol. 84. Faune de Madagascar.Paris: Museum National d’Histoire Naturelle.Pp. 1–204.
Pont, S. M.; Armstrong, J. D. (1990): A study ofthe bat fauna of the Reserve Naturelle Integral[sic] de Marojejy in north-east Madagascar.Report of the Aberdeen University expeditionto Madagascar 1989. Aberdeen: Department ofZoology, University of Aberdeen.
Russ, J.; Bennett, D.; Ross, K.; Kofoky, A. (2001):The Bats of Madagascar: A Field Guide withDescriptions of Echolocation Calls. Glossop,UK: Viper Press.
Schliemann, H. (1970): Bau und Funktion derHaftorgane von Thyroptera und Myzopoda
(Vespertilionoidea, Microchiroptera, Mamma-lia). Z. Wiss. Zool. 181, 353–400.
Schliemann, H. (1971): Die Haftorgane vonThyroptera und Myzopoda (MicrochiropteraMammalia) – Gedanken zu ihr Entstehung alsParallelbildungen. Z. Zool. Syst. Evol.-Forsch.9, 61–80.
Schliemann, H.; Goodman, S.M. (2003): Myzopo-
da aurita, Old World sucker-footed bat. In: TheNatural History of Madagascar. Ed. by S.M.Goodman and J.P. Benstead. Chicago: TheUniversity of Chicago Press. Pp. 1303–1306.
Please cite this article as: S.M. Goodman et al.,
(Myzopodidae: Chiroptera) from western..., Mamm
Schliemann, H.; Maas, B. (1978): Myzopoda aurita.Mammalian Species 116, 1–2.
Simmons, N. B. (2005): Order Chiroptera. In:Mammal Species of the World: A Taxonomicand Geographic Reference. 3rd ed. Ed. by D. E.Wilson and D. M. Reeder. Baltimore: JohnsHopkins University Press. Pp. 312–529.
Teeling, E. C.; Springer, M. S.; Madsen, O.; Bates,P.; O’Brien, S. J.; Murphy, W. J. (2005): Amolecular phylogeny for bats illuminates bio-geography and the fossil record. Science 307,
580–584.Thomas, O. (1904): On the osteology and systema-
tic position of the rare Malagasy bat Myzopoda
aurita. Proc. Zool. Soc. London 1904, 2–6.Vonhof, M. J.; Fenton, M. B. (2004): Roost
availability and population size of Thyroptera
tricolor, a leaf-roosting bat, in north-easternCosta Rica. J. Trop. Ecol. 20, 291–305.
Vonhof, M. J.; Whitehead, H.; Fenton, M. B.(2004): Analysis of Spix’s disc-winged batassociation patterns and roosting home rangesreveal a novel social structure among bats.Anim. Behav. 68, 507–521.
Wimsatt, W. A.; Enders, A. C. (1980): Structureand morphogenesis of the uterus, placenta, andparaplacental organs of the Neotropical disc-winged bat Thyroptera tricolor Spix (Micro-chiroptera: Thyropteridae). Am. J. Anat. 159,209–243.
Yoder, A. D.; Hanley, C.; Heckman, K.; Raso-loarison, R.; Russell, A.; Ranivo, J.; Olson,L. E.; Soarimalala, V.; Karanth, K. P.; Rase-limanana, A. P.; Goodman, S. M. (2005): Amultidimensional approach for detecting spe-cies patterns in Malagasy vertebrates. Proc.Natl. Acad. Sci. USA 102, 6587–6594.
Authors’ addresses:
Steven M. Goodman, Field Museum of NaturalHistory, 1400 South Lake Shore Drive, Chicago, IL60605, USA and WWF, BP 738, Antananarivo (101),Madagascar Felix Rakotondraparany, Departementde Biologie Animale, Faculte des Sciences, Universited’Antananarivo, B.P. 906, Antananarivo (101), Ma-dagascar Amyot Kofoky, Madagasikara Voakajy,B.P. 5181, Antananarivo (101), Madagascar(e-mail: [email protected] (F. Rakoton-draparany), [email protected] (A. Kofoky).
The description of a new species of Myzopoda
. biol. (2006), doi:10.1016/j.mambio.2006.08.001