the cog complex, rab6 and copi define a novel golgi retrograde trafficking pathway that is exploited...
TRANSCRIPT

The COG complex, Rab6 and COPI define a novel Golgiretrograde trafficking pathway that is exploited by SubAB toxin
Richard D. Smith1, Rose Willett1, Tetyana Kudlyk1, Irina Pokrovskaya1, Adrienne W.Paton2, James C. Paton2, and Vladimir V. Lupashin1
1Department of Physiology and Biophysics, UAMS, Little Rock, AR2School of Molecular and Biomedical Science, University of Adelaide, Australia
AbstractToxin trafficking studies provide valuable information about endogenous pathways of intracellulartransport. Subtilase cytotoxin (SubAB) is transported in a retrograde fashion through theendosome to the Golgi and then to the ER, where it specifically cleaves the ER chaperone BiP/GRP78. To identify the SubAB Golgi trafficking route, we have used siRNA-mediated silencingand immunofluorescence microscopy in HeLa and Vero cells. Knockdown of subunits of theConserved Oligomeric Golgi (COG) complex significantly delays SubAB cytotoxicity and blocksSubAB trafficking to the cis-Golgi. Depletion of Rab6 and β-COP proteins causes similar delay inSubAB-mediated GRP78 cleavage and did not augment the trafficking block observed in COGKD cells, indicating that all three Golgi factors operate on the same “fast” retrograde traffickingpathway. SubAB trafficking is completely blocked in cells deficient in the Golgi SNARESyntaxin5 and does not require the activity of endosomal sorting nexins SNX1 and SNX2.Surprisingly, depletion of Golgi tethers p115 and golgin-84 which regulates two previouslydescribed COPI vesicle-mediated pathways did not interfere with SubAB trafficking, indicatingthat SubAB is exploiting a novel COG/Rab6/COPI-dependent retrograde trafficking pathway.
KeywordsGolgi; SubAB toxin; retrograde traffic; Conserved Oligomeric Golgi complex; coiled-coil tether
INTRODUCTIONMany bacterial and plant toxins often exhibit a structural organization consisting of twofragments, called A and B (1). B subunits of these toxins recognize and bind to specificreceptors on the surface of mammalian cells and direct the internalization and trafficking ofan enzymatic A subunit. A medically important subset of bacterial toxins, called the AB5toxins, consists of hexameric assemblies comprising a single catalytically active A-subunitand a pentamer of B-subunits. Subtilase cytotoxin (SubAB) belongs to a new AB5 toxinfamily that is produced by a strain of Shiga toxigenic Escherichia coli (2). The Catalytic Asubunit of SubAB, which shares sequence homology to a subtilase–like serine protease ofBacillus anthracis, specifically cleaves and inactivates the ER lumen-localized chaperoneBiP/GRP78 (3). The SubAB-mediated GRP78 cleavage leads to a transient inhibition ofprotein synthesis, cell cycle arrest and 2 days later to cell death (4). SubAB is the only toxinknown to target a lumenal ER protein. The B subunits of SubAB bind to the surface of target
Corresponding author: Dr. Vladimir Lupashin, Department of Physiology and Biophysics, University of Arkansas for MedicalSciences, Biomed 261-2, Slot 505, 200 South Cedar St, Little Rock, AR, 72205, tel 501-603-1170, fax 501-686-8167,[email protected].
NIH Public AccessAuthor ManuscriptTraffic. Author manuscript; available in PMC 2010 October 1.
Published in final edited form as:Traffic. 2009 October ; 10(10): 1502–1517. doi:10.1111/j.1600-0854.2009.00965.x.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

cells via glycans displayed on glycoproteins, including beta1 integrin (5). Surprisingly,SubAB has a high degree of binding specificity for glycans terminating with α2,3-linkedresidues of the non-human sialic acid N-glycolylneuraminic acid (6), and therefore inhumans a SubAB receptor is generated by metabolic incorporation of an exogenous factorderived from food. Internalization of SubAB is exclusively clathrin-dependent (7). The othertwo well-studied AB5 toxins, Shiga toxin and Cholera toxin, are internalized in both aclathrin- and a cholesterol-dependent manner. On the way from the cell surface to the ER allthree toxins are passing through the Golgi apparatus, although the exact intra-Golgiretrograde trafficking pathways used by various toxins are not well understood.
Studies of bacterial toxins can provide valuable information about pathways of intracellulartransport. Retrograde transport of both Shiga and Cholera toxins to the Golgi and the ERseems to be dependent on different Rab and ARF GTPases, SNAREs and vesicular tetheringproteins (8–15). Comparison of different toxins reveals differences indicating the presenceof more than one pathway between early endosomes and the ER through the Golgi apparatus(16). Specific SubAB-mediated GRP78 cleavage provides an extremely sensitive andquantitative assay (7) to measure subtilase cytotoxin arrival at the ER in both control cellsand in cells deficient in putative components of the Golgi retrograde trafficking machinery.This should allow identification of SubAB intra-Golgi trafficking pathway(s). We haverecently found that co-depletion of Rab6 and Conserved Oligomeric Golgi (COG) vesiculartethering factor suppresses the Golgi fragmentation phenotype that is associated with theCOG knockdown (KD) (17). This epistatic assay provides the evidence that two proteins actin the same pathway.
In this work we have used both single and double KD strategies to uncover a novel Rab6,COG and COPI-dependent intra-Golgi retrograde trafficking pathway that is exploited bySubAB toxin for fast delivery to the ER.
RESULTSIn both HeLa and Vero cells, Brefeldin A treatment completely blocks SubAB-mediatedcleavage of GRP78 (7), indicating that the functional Golgi apparatus is essential for SubABtrafficking from plasma membrane to endoplasmic reticulum (ER). We have previouslyidentified the COG protein complex as an important vesicle tethering component of theintra-Golgi retrograde trafficking machinery that is responsible for the localization of Golgiglycosyltransferases and for retrograde delivery of Shiga toxin B subunit (15,18). To test thehypothesis that the COG complex is involved in the retrograde trafficking of SubAB toxin,we have utilized a shRNA (short hairpin RNA) strategy to create a stable Cog4p knock-down (Cog4 sKD) HeLa GalNAc T2-GFP cell line. The level of Cog4 protein in Cog4 sKDcells was reduced by more than 85%, as compared to the actin loading control (Figure 1A).Golgi membranes in Cog4 sKD cells appeared to be partially fragmented (Figure 1B) that isconsistent with the phenotype previously described for Cog3 siRNA KD cells (15). Bothcontrol and Cog4 sKD cells were incubated with purified SubAB toxin (3 µg/ml) for threehours. Kinetics of GRP78 cleavage was used to estimate SubAB trafficking efficiency. Incontrol HeLa cells GRP78 cleavage started at the 60 min time point, with more than 80% ofGRP78 cleaved after 120 min of incubation (Figure 1C–D, Control). In sharp contrast,GRP78 cleavage in Cog4 sKD cells was significantly delayed and started 120 min aftertoxin application (Figure 1C–D, Cog4 KD). Three hours after the start of the assay less than60% of the GRP78 was cleaved in Cog4 sKD cells indicating that Cog4p is required for the“fast” delivery of the SubAB to the ER. The block in SubAB trafficking in Cog4 sKD cellswas not complete and GRP78 was completely cleaved 5 hours after toxin application (datanot shown), indicating that in HeLa cells SubAB is also using a Cog4p-independent “slow”delivery pathway. Importantly, the delay in GRP78 cleavage observed in Cog4 sKD cells
Smith et al. Page 2
Traffic. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

was not due to the Golgi fragmentation, since a treatment of HeLa cells with Nocodazole, adrug that affects microtubule function and causes severe Golgi fragmentation, demonstratedthe SubAB-mediated GRP78 cleavage kinetics were similar to that of the untreated cells(Figure 1D, Nocodazole). Incubation of HeLa cells with labelled cytotoxin at 4°C revealedthat the same amount of SubAB binds to mock-treated and Cog4 KD cells (Figure 1E),indicating that COG complex knock-down did not compromise the plasma membrane-localized SubAB receptor(s).
The COG complex is composed of eight subunits that are grouped into two sub-complexes:Lobe A (Cog1–4) and Lobe B (Cog5–8) (19–21).To check whether other subunits of theCOG complex are necessary for the fast intracellular trafficking of SubAB toxin, HeLa cellswere transfected with siRNAs that specifically down-regulated biosynthesis of Cog2p,Cog3p, Cog4p, Cog6p and Cog8p. In all cases a significant (96% for COG3 KD, 92% forCOG4 KD, 91% for COG6 KD, 88% for COG8 KD) reduction in the protein level oftargeted proteins was achieved for all COG subunits as estimated by immunoblot analysisusing Odyssey LiCOR System (Figure 2A). Due to a lack of anti-Cog2p antibodies, theefficiency of COG2 knock-down was estimated by RT-PCR (Figure 2B). In all COG KDcells the rate of SubAB-mediated GRP78 cleavage was significantly decreased (Figure 2C),indicating that the entire COG complex is important for a fast SubAB intracellular delivery.Expression of siRNA-resistant mouse Cog3p (15) partially restored a SubAB-mediatedGRP78 cleavage in COG3 KD HeLa cells (Figure 2D–E); similar rescue of SubABtrafficking was observed in Cog4KD cells transfected with Cog4-myc siRNA-resistantplasmid and in COG8KD cells transfected with Cog8-myc siRNA-resistant plasmid(Supplementary figure 1). To test for possible synergy between Lobe A and Lobe B subunitsin SubAB delivery, GRP78 cleavage in Cog3/Cog6 and Cog3/Cog4 double knock-downcells was analyzed (Figure 2F). The delay in GRP78 cleavage was found to be moresignificant in double COG subunit KD HeLa cells as compared to a single COG subunit KDcells. It was more pronounced in Cog3/Cog4 KD cells lacking two subunits of the samelobe, indicating that the increased delay in SubAB trafficking was due to more efficientsilencing achieved in a double KD procedure, rather than to the synergy between COGcomplex sub-complexes. Very limited (less than 20%) GRP78 cleavage was observed inCOG double KD cells two hours after incubation with toxin, signifying the essential role ofthe COG complex in a fast PM-Golgi-ER delivery of SubAB toxin. Importantly, we foundthat at the 180 min incubation time point, up to 20% of GRP78 was cleaved even in the mostsevere Cog3/Cog4 double KD cells (Figure 2F). SubAB-mediated GRP78 cleavagecontinued slowly, reaching up to 80% after six hours of incubation (data not shown). Thisslowly sustained GRP78 cleavage is likely to indicate existence of a secondary “slow” COGcomplex-independent SubAB delivery pathway in HeLa cells.
COG complex deficiency could affect SubAB cytotoxicity either directly, via regulation ofretrograde membrane carriers that carry SubAB to ER, or indirectly, by affectinglocalization and/or expression of SubAB receptors on the cell surface. We have shownpreviously that the depletion of Cog3p did not block the delivery of both plasma membraneand lysosomal proteins (15,18). In agreement to this result, the depletion of other COGsubunits did not significantly effect the anterograde protein trafficking in HeLA cells(Supplementary figure 2) To visualize SubAB trafficking we have labelled SubAB withHiLyte Fluor 555 (HF555) dye and with Alexa Fluor 647(AF647) and analyzed SubAB-HF555 and SubAB-AF647 intracellular trafficking by the immunofluorescence (IF)approach. Labelling of SubAB with either HiLyte Fluor 555 or Alexa Fluor 647 dyes did notchange the kinetics of SubAB-mediated GRP78 cleavage (data not shown); hence SubAB-HF555/AF647 trafficking is likely to be identical to the trafficking of the unlabelledsubtilase cytotoxin.
Smith et al. Page 3
Traffic. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

We have initially tried to use SubAB-HF555 in HeLa cells and found that after 60 min and120 min of incubation, the majority of SubAB-HF555 did not localize with GalNAc-T2-GFP-positive Golgi structures, indicating that in HeLa cells SubAB is being distributedthroughout a variety of endocytic compartments (data not shown). As a result, the pattern ofSubAB intracellular trafficking in HeLa cells was not interpretable with a straightforward IFapproach. This is in agreement with an earlier observation that that in several cell types,including HeLa cells, not more than 5–10% of endocytosed toxin is transported in thedirection of the Golgi apparatus (22).
A less complicated intracellular trafficking pattern of SubAB was observed previously inVero cells (3). In Vero cells significantly lower SubAB initial concentration (0.05 µg/ml)was sufficient to completely cleave GRP78 in 60 min (Figure 3B, control). Vero cellstransfected with COG3 siRNA demonstrated substantial down-regulation of Cog3p (Figure3A). Again, as in the HeLa case, depletion of Cog3p in Vero cells significantly delayedSubAB-mediated GRP78 cleavage (Figure 3B). Incubation of both control and Cog3 KDVero cells with SubAB-HF555 at 4°C resulted in a comparable binding of labelled toxin tothe plasma membrane (Figure 3C–D, 0 min row). Upon incubation at 37°C the SubAB-HF555 was rapidly internalized and in more than 80% of control cells the toxin was mostlylocated in a perinuclear region where it co-localized with cis-Golgi marker giantin (Figure3C, 30 min, arrows). This major co-localization of internalized SubAB-HF555 with cis-Golgi was persistent in control Vero cells 60 min after toxin trafficking started (Figure 4C,60 min). Labelled SubAB toxin was rapidly endocytosed in Cog3 KD Vero cells, however,in a striking contrast with control cells, the majority of internalized cytotoxin was found atthe cell periphery at both 30 and 60 min time points (Figure 3D). Less than 15% of Cog3KD cells demonstrated perinuclear localization of SubAB-HF555. However, even in thesecells the perinuclear toxin was not co-localized with the cis-Golgi marker giantin (Figure3D, 60 min, arrowhead), indicating that COG complex malfunction causes accumulation ofinternalized cytotoxin in pre-cis-Golgi trafficking intermediates.
To gain more understanding about SubAB-positive pre-cis-Golgi compartments bothCOG3KD and mock-treated Vero cells were incubated with fluorescently labelled SubAB(SubAB-HF555 and SubAB-AF647) and stained for the markers for both endocytic andsecretory pathways (Figure 4). In control cells SubAB upon incubation at for 60 min at 37°Cwas partially co-localized with trans-Golgi marker TGN46 and did not show any significantco-localization with endosomal-lysosomal compartments. In COG3 KD Vero cells some(~15–20%) of internalized SubAB was co-localized with TGN46 (Figure 4, TGN46 row,arrow), indicating that COG depletion did not block SubAB delivery to TGN/trans-Golgi.No significant co-localization of SubAB was observed with early endosomal protein EEA1(Figure 4, EEA1 row), late endosomal protein Rab7 (Figure 4, rab7 row) and lysosomalmarker Lamp2 (Figure 4, Lamp2 row). We also were unable to find a significant SubAB co-localization with Snx2, Vps26,Rab11 and PDI (Supplementary figure 3 and data not shown).Similar results were obtained for COG8 KD cells (data not shown).
To identify other Golgi proteins that may regulate intracellular trafficking of SubAB, wehave analyzed known COG complex-interacting trafficking factors.
We have demonstrated previously that the COG complex is acting downstream of the trans-Golgi-localized Rab6 protein and that the epistatic depletion of Rab6 inhibited the Golgi-disruptive effect of COG inactivation by siRNA or antibodies (17). Here we found that thedepletion of Rab6 in HeLa cells affected the kinetics of SubAB-mediated GRP78 cleavagein a manner analogous to COG depletion (Figure 5). Efficient double Rab6/Cog3 KD(Figure 5A) resulted in delay in SubAB delivery similar to that observed in the single Rab6KD cells (Figure 5B) indicating that both Rab6 and the COG complex operate on the same
Smith et al. Page 4
Traffic. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

retrograde pathway of SubAB delivery. Rab6 siRNA was not efficient in Vero cells (datanot shown), therefore we tested if the overexpression of GDP-restricted Rab6 mutant wouldinterfere with SubAB trafficking. Vero cells were co-transfected with pCFP and pRab6S21Nplasmids and treated with fluorescently-labelled SubAB. We found that in CFP-positivecells internalized SubAB was preferentially co-localized with EEA1 and not with giantin(Figure 5C), indicating that Rab6 malfunction was interfered with the endosome-to-Golgileg of SubAB retrograde trafficking itinerary.
The COG complex genetically and physically interacts with subunits of COPI vesicular coatthat operates in both intra-Golgi and Golgi to ER retrograde membrane trafficking steps(15,23,24), and COG and COPI may work in concert to ensure the proper retention andretrieval of a subset of proteins in the Golgi (25). To downregulate β-COP we used apreviously characterized siRNA in HeLa cells (26). Efficient (more than 90%) siRNA-mediated depletion of β-COP did not affect the intracellular level of Cog3p (Figure 6A), butcaused a significant delay in SubAB-mediated GRP78 cleavage (Figure 6B). Again, doubleβ-COP/Cog3 KD resulted in delay in SubAB delivery similar to that observed in the singleβ-COP KD cells (Figure 6B), demonstrating that both COPI and COG complexes control thesame retrograde SubAB delivery pathway.
Knock-down of β-COP was in some way detrimental for HeLa cells (only 50% of cells wererecovered after 72 of β-COP KD) and resulted in an extensive Golgi fragmentation (data notshown). Therefore we have tested if the less invasive COPI malfunction caused byoverexpressioin of GTP-locked mutant for on Arf1(27) would interfere with SubABtrafficking. Vero cells were transfected with pArf1Q71L-CFP plasmid and treated withfluorescently-labelled SubAB. We found that CFP-Arf1Q71L expression blockedfluorescently-labelled SubAB trafficking to Giantin-positive cis-Golgi membranes (Figure6C), confirming the essential role of COPI in retrograde trafficking of subtilase cytotoxin.As in the case of COG3 KD cells, in Arf1Q71L expressing Vero cells SubAB was trapped intrafficking intermediates that did not co-localize with any tested endocytic markers (data notshown).
The COG complex genetically and physically interacts with a subset of Golgi-operatedSNARE molecules (15,23,24). In mammalian cells, hCog4p and hCog6p interact with Qa-SNARE Syntaxin5a (28). We have used previously described siRNA (29) to knockdownboth long and short isoforms of Syntaxin5 (Figure 7A) and found that SubAB-mediatedGRP78 cleavage was almost completely blocked (Figure 7B). Less than 8% of GRP78 wascleaved even after 180 min of incubation of HeLa cells with SubAB toxin, indicating thatSyntaxin 5 is required for both fast (COG complex-dependent) and slow (COG complex-independent) intracellular delivery of subtilase cytotoxin. In Syntaxin5 KD Vero cellsfluorescently-labelled SubAB failed to co-localize with both cis (Giantin) and trans (P230)Golgi compartments (Figure 7C) and was accumulated in peripherally–located smallmembrane structures, indicating an early block in retrograde trafficking.
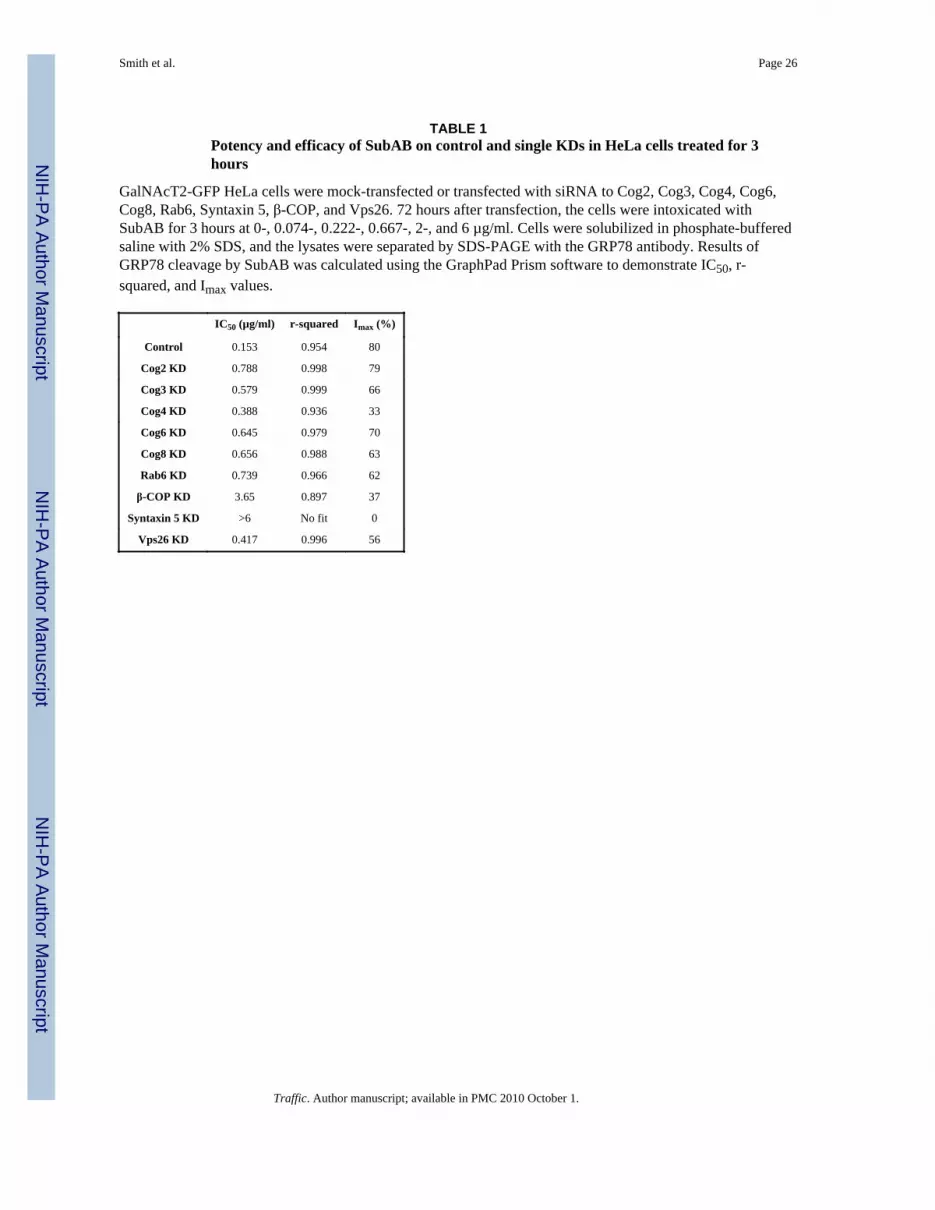
Knock-down of Cog2, Cog3, Cog6,Cog8 and Rab6 resulted in a four-fold increase ofpotency or Inhibitory Concentration of 50% (IC50) for SubAB in HeLa cells (Table 1 andSupplementary figure 4). More dramatic IC50 increase was detected in β-COP KD cells,while even a significant increase (up to 6 µg/ml) in SubAB concentration applied onSyntaxin5 KD cells has not resulted in any GRP78 cleavage.
It was reported recently that Cog2p physically interacts with another cis-Golgi vesicle-tethering factor p115 (30), and that p115 stimulates formation of Syntaxin 5-containingSNARE complex in vitro (31). In addition, p115/SNARE interaction was shown to beessential for Golgi biogenesis (32). On the other hand, we have shown previously that the
Smith et al. Page 5
Traffic. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

epistatic depletion of Rab6 did not inhibit the Golgi-disruptive effect of p115 inactivation bysiRNA (17), indicating that Rab6 and p115 operate on independent Golgi traffickingpathways. Here we used the same siRNA–mediated p115 silencing to test for a possiblep115 requirement in SubAB trafficking in HeLa cells. P115 KD cells lacked the signal forp115 protein and demonstrated the characteristic Golgi fragmentation phenotype (Figure8A,C), but rather surprisingly, did not show any inhibition or delay in SubAB-mediatedGRP78 cleavage (Figure 8B), indicating that SubAB is delivered to the ER via a p115-independent pathway.
The Warren lab recently proposed that p115-giantin and golgin-84-CASP Golgi tethersdefine two distinct sub-populations of COPI vesicles (33). Since p115 was dispensable forthe fast COPI-dependent delivery of SubAB through the Golgi stack, we tested golgin-84involvement in cytotoxin trafficking. Interestingly, golgin-84 KD cells lacked the signal forgolgin-84 protein and demonstrated the characteristic Golgi fragmentation phenotype inHeLa cells (Figure 8A, C), but again, as in the case of p115 KD cells, did not show anyinhibition or delay in SubAB-mediated GRP78 cleavage (Figure 8B), indicating that SubABis delivered to the ER via a golgin-84-independent pathway. This data indicate that SubABis exploiting a novel p115/golgin-84-independent intra-Golgi trafficking pathway. Anotherpossibility is that the small residual amounts of coiled-coil Golgi tethers in p115 KD andgolgin 84 KD cells were sufficient to fully support SubAB retrograde delivery.
We have reported previously that Cog3 KD HeLa cells are deficient in retrograde traffickingof Shiga-like toxin B (SLTB) subunit (15). Previous reports suggested that SubAB andSLTB retrograde trafficking pathways are similar, but not identical (7). Indeed we haveobserved that in Vero cells incubated with both SubAB-HF555 and SLTB-AF647, STBLaccumulation in perinuclear Golgi presided the arrival of SubAB to the same area (data notshown). We also found different sensitivity of toxins retrograde trafficking to the level ofCOG3 knock-down (Figure 9). At the intermediate level of COG3 KD fast delivery ofSubAB to Golgi was blocked, while STLB was delivered normally (Figure 9, partial COG3KD row). In a contrast, cells that maintain a very low level of Cog3p, both SubAB andSLTB were not delivered to the Golgi (Figure 9, COG3 KD).
Endosomal-to Golgi trafficking of Shiga toxin is also regulated by Rab6 (9,34) and bysorting nexins SNX1 and SNX2 (35,36). Combined depletion of SNX1 and SNX2 in Verocells gave a total inhibition of Shiga toxin transport to the trans-Golgi network by 80% (36).Having established the requirement for both COG complex and Rab6 in SubAB trafficking,we tested whether SNX1 and/or SNX2 are essential for this process in HeLa cells. siRNA-mediated inhibition was efficient in knocking down SNX1, SNX2 and both SNX1/SNX2(Figure 10A), but in all cases SubAB-mediated GRP78 cleavage was indistinguishable fromcontrol cells (Figure 10B), indicating that SubAB is delivered to the ER via the SNX1/SNX2-independent pathway. Interestingly, we found that the knock-down of anotherretromer component, Vps26 in HeLa cells (figure 10A) moderately inhibited SubAB-mediated GRP78 cleavage (Figure 10C), indicating that the retromer complex is notcompletely disposable for SubAB retrograde delivery.
DISCUSSIONAnalysis of SubAB trafficking in HeLa and Vero cells revealed several key Golgi residentproteins that are indispensable for the efficient delivery of the subtilase cytotoxin to the ER.The COG complex has been previously implicated in the retrograde trafficking of severalGolgi enzymes, SNAREs and Shiga toxin B subunit (15,18,28). Our experiments revealedthat down-regulation of any subunit of the COG complex significantly delayed SubABdelivery to the ER. The delay in SubAB delivery was very similar in cells deficient for either
Smith et al. Page 6
Traffic. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Lobe A (Cog 2, 3 and 4) or Lobe B (Cog6 and 8) of the COG complex, suggesting that theentire COG complex is required for the toxin delivery. The defect in SubAB trafficking wasmost apparent in cells deficient for two COG subunits, indicating that the residual COGcomplex activity that remained in cells after the knockdown of any single subunit was stillsufficient to support at least some SubAB trafficking. Binding to the cell surface and initialinternalization of labelled SubAB was normal in COG-deficient cells, whereas trafficking tocis-Golgi was dramatically inhibited, indicating that the COG complex is required for eitherintra-Golgi or endosome-to-Golgi trafficking steps. In COG3 KD Vero cells the internalizedSubAB did not show any significant co-localization with markers of endosomal/lysosomalpathway indicating a post-endosomal trafficking block. Some SubAB was able to arrive atthe perinuclear area and co-localize with TGN46 in Cog3 KD cells, but failed to co-localizewith the cis-Golgi marker giantin, indicating an intra-Golgi trafficking defect.
Importantly, COG deficiencies only notably delayed, but did not completely block theSubAB-mediated GRP78 cleavage, indicating the existence of multiple retrograde pathwaysexploited by SubAB toxin. The incomplete SubAB trafficking block in COG KD cells wasactually beneficial to this study, and allowed us to analyze other components of the intra-Golgi trafficking machinery that may work on the same or parallel pathway. Knockdown ofeither trans-Golgi Rab6 or β-COP delayed SubAB delivery in a manner similar to thatobserved in COG KD cells, suggesting that these trafficking regulators may work on thesame pathway. Block in SubAB delivery to the ER in cells that overexpress either Rab6-GTP or Arf1-GTP further underlined the importance of Rab6 and COPI for SubABtrafficking, Most importantly, simultaneous KD of either Rab6/Cog3 or β-COP/Cog3 did notchange the severity of the toxin trafficking block, supporting the notion that the COGcomplex, Rab6 and COPI regulate the same fast intra-Golgi trafficking pathway exploitedby SubAB cytotoxin. This is in good agreement with previously described interactionsbetween the COG and COPI complexes (15,23–25), as well as with the recently uncoveredepistatic interaction between Rab6 and the COG complex (17). SubAB was preferentiallyaccumulated in EEA1-positive endosomal compartment in cells that overexpress Rab6GDPmutant, but not in COG3 KD cells, indicating that Rab6 functions upstream of the COGcomplex in SubAB retrograde trafficking pathway
It is possible that all SubAB trafficking in normal cells is COG/Rab6/COPI-dependent, butin cells deficient for this major retrograde trafficking pathway, the alternative endosome toER transport is activated. The induction of routes that are normally nonexistent wasobserved before in the case of ricin trafficking in cells that express a temperature-sensitivemutant of ε-COP (37). We favour the alternative hypothesis that SubAB toxin normallytravels through the Golgi by two separate routes: fast, COG/Rab6/COPI-dependent, andslow, COG/Rab6/COPI-independent. This interpretation is supported by the toxin traffickingresults obtained in cells deficient in Qa-SNARE Syntaxin5.
SubAB trafficking block in Syntaxin5 KD cells was similar to a block observed in cells pre-treated with Brefeldin A (data not shown and (7)). In both cases, no GRP78 cleavage wasobserved after 3 hours of incubation with SubAB, indicating that Brefeldin A treatment andSyntaxin5 KD inhibit both fast and slow routes through the Golgi. Interestingly, that amongall COG complex knock-down cells the strongest inhibition of SubAB trafficking wasassociated with depletion of Cog4p, a Lobe A subunit that directly interacts with Syntaxin5(28). Syntaxin5 was implicated in both intra-Golgi (38) and early/recycling endosome to theTrans Golgi Network (TGN) (39) retrograde trafficking steps. One possibility is that SubABis delivered from endosomes to the ER via two distinct mechanisms by exploiting either thefast COG complex-dependent inter-cisternae vesicular trafficking pathway or by hopping onto the slow direct TGN to ER membrane flow (Figure 11). In specific experimentalconditions the retrograde transport of Cholera toxin from the plasma membrane to the ER
Smith et al. Page 7
Traffic. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

was shown to require the TGN, but not the Golgi complex (40). Existence of the endogenousdirect TGN-to-ER trafficking route was also proposed (41). Syntaxin5 KD most likely willeliminate both routes for SubAB delivery.
Alternatively, the complete SubAB trafficking block in Syntaxin5 KD cells is associatedwith the dual role of Syntaxin5 in both anterograde and retrograde trafficking. Indeed, someof the endogenous CD44 and the majority of the model (VSVG-GFP) anterograde cargoproteins were accumulated in the Golgi area in Syntaxin5 KD cells, while both control andCOG-deficient HeLa cells correctly delivered both proteins to the plasma membrane(Supplementary figure 2).
Depletion of several Golgi factors (Cog3, Cog4, β-COP and Syntaxin5) in HeLa cells leadsto an intense Golgi fragmentation, but the Golgi breakup observed in KD cells seemed to benot linked to the inhibition of SubAB trafficking. Indeed, the trafficking of SubAB to thefragmented Golgi continued in the presence of Nocodazole (7), and the Golgi fragmentationobserved in p115 KD and golgin84 KD cells did not interfere with cytotoxin arrival in theER.
Rather surprisingly we found that SubAB intra-Golgi trafficking does not require twopreviously described Golgi-localized coiled-coil vesicle tethers p115 and golgin-84. Recentwork from Warren lab proposed that p115-giantin and golgin-84-CASP Golgi tethers definetwo distinct sub-populations of COPI vesicles (33). Since only a sub-fraction of Golgi-derived COPI vesicles was efficiently captured by both tethering pairs, Warren labsuggested that “other tethers should identify other populations, allowing us to map the flowpatterns of resident, cargo, and recycling molecules”. Our finding clearly reinforces thisprediction, suggesting that SubAB is using a novel p115/golgin84-independent and COG/Rab6/COPI-dependent intra-Golgi trafficking pathway.
The nature of the molecule (or molecules) that carry SubAB through the retrograde Golgipathways is currently unknown. It could be a SubAB plasma membrane glycoproteinreceptor itself, analogous to a lipid-raft ganglioside GM1 that was proposed to carry Choleratoxin all the way from plasma membrane to ER (42). Alternatively, SubAB could usedistinct carriers for different trafficking steps “jumping” from one to another in sortingcompartments. While the nature of the intracellular SubAB carrier is not known at present, itis clear that SubAB retrograde trafficking is quite distinct from the trafficking of otherknown AB5 toxins. Both Cholera and Shiga toxin enter target cells via both clathrin-dependent and clathrin-independent mechanisms, while SubAB internalization is strictlyclathrin-dependent (7). Unlike Shiga toxin, SubAB trafficking does not require sortingnexins SNX1/SNX2, is more sensitive to COG complex depletion and is dependent on COPIvesicular coat.
The initial characterization of the COG/Rab6/COPI–dependent retrograde pathway mayprovide a useful framework for better understanding of intra-Golgi trafficking in mammaliancells.
MATERIALS AND METHODSReagents and antibodies
Reagents were as follows: Nocodazole (stock solution = 10 mM) (Sigma), Subtilasecytotoxin (1.49 mg/ml) (2), and Shiga-like toxin subunit B prepared as described previously(43). Antibodies used for immunofluorescence microscopy (IF) or western blotting (WB)were purchased through commercial sources, gifts from generous individual investigators, orgenerated by us. Antibodies (and their dilutions) were as follows: rabbit affinity purified
Smith et al. Page 8
Traffic. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

anti-hCog3 (WB 1:10,000 (44), hCog4 (WB 1:1000, this lab), hCog6 (WB 1:1000, this lab),hCog8 (WB 1:1000, this lab), Syntaxin 5 (WB 1:1000, this lab), p115 (IF 1:100 (45)), Rab6(WB 1:200 (Santa Cruz), Giantin (IF 1:1000 (Covance), TGN46 (IF 1:300; AdB Serotec),Vps26 (WB 1:500; Abcam); mouse monoclonal anti- β-Actin (WB 1:10,000 (Sigma), β-COP (WB 1:500 (Sigma), GAPDH (WB 1:1000 (Santa Cruz), golgin-84 (WB 1:500 (BDBiosciences), Snx1 (WB 1:1000 (BD Biosciences), Snx2 (WB 1:1000 (BD Biosciences),p115 (IF 1:100 (46)), EEA1 (IF 1:200 (BD Biosciences), P230 (IF 1:200 (BD Biosciences),GPP130 (IF 1:1000 (Alexis Biochemicals), LAMP2 (IF 1:100 (Developmental StudiesHybridoma Bank, University of Iowa), Rab7 (IF 1:200 (Cell Signaling Technology), CD44(IF 1:50; Developmental Studies Hybridoma Bank, University of Iowa), and goat anti-GRP78 (WB goat 1:1000 (Santa Cruz).
Secondary anti-goat-HRP, anti-mouse-HRP, and anti-rabbit-HRP for WB were obtainedfrom Jackson ImmunoResearch Laboratories. IRDye 680 goat anti-rabbit, IRDye 700 goatanti-mouse, and IRDye 800 donkey anti-goat for WB were obtained from LI-CORbiosciences. Anti-rabbit HiLyte 488 and anti-rabbit HiLyte 555 for IF were obtained fromAnaSpec, Inc.
Cell cultureHeLa cells stably expressing GalNAcT2-GFP were cultured in DMEM/F-12 mediumsupplemented with 15 mM HEPES, 2.5 mM L-glutamine, 10% FBS, and 0.4 mg/ml G418sulfate. Additionally, the stable Cog4 KD cell line was cultured in DMEM/F-12 mediumsupplemented with 15 mM HEPES, 2.5 mM L-glutamine, 10% FBS, and 1.0 µg/mlpuromycin. GalNAcT2-GFP HeLa cells is a gift from Dr. Storrie (UAMS, Little Rock, AR).
Vero cells were grown in DMEM/F-12 (50:50) with 15 mM HEPES, 2.5 mM L-glutamine,10% fetal bovine serum, and 1% antibiotic/antimycotic (100 U/ml penicillin G, 100 g/mlstreptomycin, and 0.25 µg/ml amphotericin B). All cell culture media and sera are fromInvitrogen (Carlsbad, CA).
shRNA and siRNA-induced knockdownsTo generate a stable knockdown of Cog4, HeLa T2-GFP cells (15) were transfected with theshRNA plasmid (OriGene) (target sequenceTGACATCTTGGACCTGAAGTTCTGCATGG) using Lipofectamine 2000 according to aprotocol from the manufacturer (Invitrogen). Medium containing 1 µg ml−1 puromycin wasused to select for the plasmid. Colonies were chosen according to Cog4 down-regulation asjudged by immunoblotting.
Human Cog2 (target sequence: GGGCAGTTGATGAACGAAT), Cog4 (target sequence:GTGCTGAAATCCACCTTTA), Cog6 (target sequence: AGATATGACAAGTCGCCTA),Cog8 (target sequence: GATGATCCCTATTTCCATA), Cog3 (15), Snx1 and Snx2 (47),p115 (32), β-COP (26), Rab6 (17), golgin-84 (33), Vps26 (48) and Syntaxin 5 (49) siRNAduplexes were obtained from Dharmacon and Sigma-Proligo. Transfection was performedusing Oligofectamine (Invitrogen) following a protocol recommended by Invitrogen. Twocycles of transfections (100 nM siRNA each) were performed and cells were analyzed 72hours after the second cycle.
For Vero cells, we used RNAiMAX to transfect Cog3 and Syntaxin 5 siRNA duplexes usinga protocol from the supplier (Invitrogen).
Smith et al. Page 9
Traffic. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Plasmid preparation and transfectionPlasmids for Rab6-GDP (pRab6S21N), CFP-Arf1-Q71L, mouse Cog3 were describedpreviously (9,15,50). Mutant (siRNA-resistant) versions of Cog4, and Cog8 were generatedby GeneScript (Piscataway, NJ) using plasmids pCOD1-myc and pDOR1-myc(51).
Plasmid transfections were performed with Lipofectamine 2000 (Invitrogen) or with FuGene(Roche) according to the manufacturer’s protocol.
Immunofluorescence microscopyFor IF, cells are grown on 12 mm glass coverslips one day before transfection. 72h aftersiRNA transfection, cells are fixed and stained with antibodies as previously described(18,52). In short, cells were fixed in 4% paraformaldehyde (16% stock solution; ElectronMicroscopy Sciences). Cells were then treated with 1% Triton X-100 for one minute. Afterthat the cells were treated with 50 mM ammonium chloride for 5 min. Cells were washedwith PBS and blocked twice for 10 min with 1% BSA, 0.1% saponin in PBS. Cells werestained in primary antibody for 1 hour (1% cold fish gelatin, 0.1% saponin in PBS) at roomtemperature. Cells were washed four times with PBS and incubated for 30 min withfluorescently tagged secondary antibody (1:400 HiLyte; AnaSpec). After that cells werewashed four times with PBS, and rinsed with ddH2O. Cells were mounted on microscopeslides using Prolong® Gold antifade reagent (Invitrogen). Cells were imaged with the 63Xoil 1.4 numerical aperture (NA) objective of a LSM510 Zeiss Laser inverted microscopeoutfitted with confocal optics for image acquisition. Image acquisition is controlled withLSM510 software (Release Version 4.0 SP1).
Subtilase cytotoxin assayBoth control and knockdown GalNAcT2-GFP HeLa cells were grown on 12-well cultureplates to 70–80% of confluency. Cells were placed in 10% FBS in DMEM/F-12 (50:50)without antibiotics and antimycotics for at least one hour before the assay was performed.The subtilase cytotoxin was diluted in the same medium (without antibiotic) and warmed upto 37°C. The time course is 0-, 30-, 60-, 120-, and 180-min. Cells were incubated with thesubtilase cytotoxin for the appropriate times in the 37°C incubator with 5% CO2. Cells werethen washed off with PBS and lysed with 2% SDS warmed up to 95°C. 10 µl of cell lysateswere loaded on a 12% gel and immunoblotted with anti-GRP78 (Santa Cruz; C-20)antibody. All experiments were performed in triplicates. The blots were scanned andanalyzed with an Odyssey Infrared Imaging System (LI-COR, Lincoln, NE).
For Vero cells the concentration of SubAB was changed to 0.05 µg/ml and the incubationwas performed in a 32°C incubator.
Fluorescently-labeling of the Subtilase cytotoxin and Shiga-like toxin BSubtilase cytotoxin was labeled with HiLyte Fluor 555 (Dojindo Laboratories, Rockville,MA) and the Shiga-like Toxin B was labelled with Alexa Fluor 647 (Invitrogen) accordingto the protocols supplied by the manufacturers.
Reverse Transcriptase – Polymerase Chain Reaction (RT-PCR)Control, Cog2 KD and Cog3 KD HeLa cells were lysed and homogenized passing five timesusing a 22-gauge needle. The mRNA from control and COG2 KD cells was isolated usingthe RNeasy Plus kit (Qiagen) using their protocol. RNA samples were translated to cDNAby using multiscribe reverse transcriptase (Applied Biosystems) in a BioRad MJ MiniThermal cycler machine. The cDNAs were amplified using 25 cycles of PCR, using Taq
Smith et al. Page 10
Traffic. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

DNA polymerase and primers for COG2 and COG3. 10 µl of PCR product was loaded to a1% agarose gel and imaged with BioRad Quantity One software.
SDS-PAGE and western blottingSDS-PAGE and WB were performed as described previously (24). The blots were incubatedfirst with primary antibodies, and then with a secondary IgG antibody conjugated withIRDye 680 or IRDye 800 dyes. The blots were scanned and analyzed with an OdysseyInfrared Imaging System (LI-COR, Lincoln, NE). Alternatively, a signal was detected usingan enhanced chemiluminescent reagent for HRP from Pierce (Rockford, IL) and quantifiedusing ImageJ software (http://rsb.info.nih.gov/ij/).
Supplementary MaterialRefer to Web version on PubMed Central for supplementary material.
AcknowledgmentsWe are thankful to S. Munro, B. Storrie, E. Sztul and others who provided reagents and critical reading of themanuscript. We also would like to thank Paul Prather for entering data in the GraphPad Prism for calculating theIC50 and Imax values. Supported by grants from the NSF (MCB-0645163) and the NIH (1R01GM083144-01A1).
Abbreviation List
COG conserved oligomeric Golgi complex
KD knockdown
RT-PCR Reverse Transcriptase –Polymerase Chain Reaction
SubAB subtilase cytotoxin
siRNA short interfering RNA
SNARE soluble Nethylmaleimide-sensitive fusion protein attachment protein receptor
References1. Merritt EA, Hol WG. AB5 toxins. Curr Opin Struct Biol. 1995; 5(2):165–171. [PubMed: 7648317]2. Paton AW, Srimanote P, Talbot UM, Wang H, Paton JC. A new family of potent AB(5) cytotoxins
produced by Shiga toxigenic Escherichia coli. J Exp Med. 2004; 200(1):35–46. [PubMed:15226357]
3. Paton AW, Beddoe T, Thorpe CM, Whisstock JC, Wilce MC, Rossjohn J, Talbot UM, Paton JC.AB5 subtilase cytotoxin inactivates the endoplasmic reticulum chaperone BiP. Nature. 2006;443(7111):548–552. [PubMed: 17024087]
4. Morinaga N, Yahiro K, Matsuura G, Moss J, Noda M. Subtilase cytotoxin, produced by Shiga-toxigenic Escherichia coli, transiently inhibits protein synthesis of Vero cells via degradation of BiPand induces cell cycle arrest at G1 by downregulation of cyclin D1. Cell Microbiol. 2008; 10(4):921–929. [PubMed: 18005237]
5. Yahiro K, Morinaga N, Satoh M, Matsuura G, Tomonaga T, Nomura F, Moss J, Noda M.Identification and characterization of receptors for vacuolating activity of subtilase cytotoxin. MolMicrobiol. 2006; 62(2):480–490. [PubMed: 16965518]
6. Byres E, Paton AW, Paton JC, Lofling JC, Smith DF, Wilce MC, Talbot UM, Chong DC, Yu H,Huang S, Chen X, Varki NM, Varki A, Rossjohn J, Beddoe T. Incorporation of a non-human glycanmediates human susceptibility to a bacterial toxin. Nature. 2008; 456(7222):648–652. [PubMed:18971931]
Smith et al. Page 11
Traffic. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

7. Chong DC, Paton JC, Thorpe CM, Paton AW. Clathrin-dependent trafficking of subtilase cytotoxin,a novel AB5 toxin that targets the endoplasmic reticulum chaperone BiP. Cell Microbiol. 2008;10(3):795–806. [PubMed: 18042253]
8. Amessou M, Fradagrada A, Falguieres T, Lord JM, Smith DC, Roberts LM, Lamaze C, Johannes L.Syntaxin 16 and syntaxin 5 are required for efficient retrograde transport of several exogenous andendogenous cargo proteins. J Cell Sci. 2007; 120(Pt 8):1457–1468. [PubMed: 17389686]
9. Girod A, Storrie B, Simpson JC, Johannes L, Goud B, Roberts LM, Lord JM, Nilsson T, PepperkokR. Evidence for a COP-I-independent transport route from the Golgi complex to the endoplasmicreticulum. Nat Cell Biol. 1999; 1(7):423–430. [PubMed: 10559986]
10. Lieu ZZ, Derby MC, Teasdale RD, Hart C, Gunn P, Gleeson PA. The golgin GCC88 is requiredfor efficient retrograde transport of cargo from the early endosomes to the trans-Golgi network.Mol Biol Cell. 2007; 18(12):4979–4991. [PubMed: 17914056]
11. Majoul I, Sohn K, Wieland FT, Pepperkok R, Pizza M, Hillemann J, Soling HD. KDEL receptor(Erd2p)-mediated retrograde transport of the cholera toxin A subunit from the Golgi involvesCOPI, p23, and the COOH terminus of Erd2p. J Cell Biol. 1998; 143(3):601–612. [PubMed:9813083]
12. Mallard F, Tang BL, Galli T, Tenza D, Saint-Pol A, Yue X, Antony C, Hong W, Goud B, JohannesL. Early/recycling endosomes-to-TGN transport involves two SNARE complexes and a Rab6isoform. J Cell Biol. 2002; 156(4):653–664. [PubMed: 11839770]
13. Tai G, Lu L, Wang TL, Tang BL, Goud B, Johannes L, Hong W. Participation of the syntaxin 5/Ykt6/GS28/GS15 SNARE complex in transport from the early/recycling endosome to the trans-Golgi network. Mol Biol Cell. 2004; 15(9):4011–4022. [PubMed: 15215310]
14. Wang Y, Tai GH, Lu L, Johannes L, Hong WJ, Tang BL. Trans-Golgi network syntaxin 10functions distinctly from syntaxins 6 and 16. Mol Membr Biol. 2005; 22(4):313–325. [PubMed:16154903]
15. Zolov SN, Lupashin VV. Cog3p depletion blocks vesicle-mediated Golgi retrograde trafficking inHeLa cells. Journal of Cell Biology. 2005; 168(5):747–759. [PubMed: 15728195]
16. Sandvig K, Spilsberg B, Lauvrak SU, Torgersen ML, Iversen TG, van Deurs B. Pathways followedby protein toxins into cells. Int J Med Microbiol. 2004; 293(7–8):483–490. [PubMed: 15149022]
17. Sun Y, Shestakova A, Hunt L, Sehgal S, Lupashin V, Storrie B. Rab6 regulates both ZW10/RINT-1 and conserved oligomeric Golgi complex-dependent Golgi trafficking and homeostasis.Mol Biol Cell. 2007; 18(10):4129–4142. [PubMed: 17699596]
18. Shestakova A, Zolov S, Lupashin V. COG complex-mediated recycling of Golgiglycosyltransferases is essential for normal protein glycosylation. Traffic. 2006; 7(2):191–204.[PubMed: 16420527]
19. Ungar D, Oka T, Brittle EE, Vasile E, Lupashin VV, Chatterton JE, Heuser JE, Krieger M, WatersMG. Characterization of a mammalian Golgi-localized protein complex, COG, that is required fornormal Golgi morphology and function. Journal of Cell Biology. 2002; 157(3):405–415. [PubMed:11980916]
20. Fotso P, Koryakina Y, Pavliv O, Tsiomenko AB, Lupashin VV. Cog1p plays a central role in theorganization of the yeast conserved oligomeric golgi complex. Journal of Biological Chemistry.2005; 280(30):27613–27623. [PubMed: 15932880]
21. Ungar D, Oka T, Vasile E, Krieger M, Hughson FM. Subunit architecture of the conservedoligomeric golgi complex. Journal of Biological Chemistry. 2005; 280(38):32729–32735.[PubMed: 16020545]
22. Sandvig K, van Deurs B. Membrane traffic exploited by protein toxins. Annu Rev Cell Dev Biol.2002; 18:1–24. [PubMed: 12142266]
23. Ram RJ, Li B, Kaiser CA. Identification of sec36p, sec37p, and sec38p: components of yeastcomplex that contains sec34p and sec35p. Mol Biol Cell. 2002; 13(5):1484–1500. [PubMed:12006647]
24. Suvorova ES, Duden R, Lupashin VV. The Sec34/Sec35p complex, a Ypt1p effector required forretrograde intra-Golgi trafficking, interacts with Golgi SNAREs and COPI vesicle coat proteins.Journal of Cell Biology. 2002; 157(4):631–643. [PubMed: 12011112]
Smith et al. Page 12
Traffic. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

25. Oka T, Ungar D, Hughson FM, Krieger M. The COG and COPI complexes interact to control theabundance of GEARs, a subset of Golgi integral membrane proteins. Mol Biol Cell. 2004; 15(5):2423–2435. [PubMed: 15004235]
26. Guo Y, Punj V, Sengupta D, Linstedt AD. Coat-Tether Interaction in Golgi Organization. Mol BiolCell. 2008
27. Teal SB, Hsu VW, Peters PJ, Klausner RD, Donaldson JG. An activating mutation in ARF1stabilizes coatomer binding to Golgi membranes. J Biol Chem. 1994; 269(5):3135–3138.[PubMed: 8106346]
28. Shestakova A, Suvorova E, Pavliv O, Khaidakova G, Lupashin V. Interaction of the conservedoligomeric Golgi complex with t-SNARE Syntaxin5a/Sed5 enhances intra-Golgi SNARE complexstability. J Cell Biol. 2007; 179(6):1179–1192. [PubMed: 18086915]
29. Suga K, Hattori H, Saito A, Akagawa K. RNA interference-mediated silencing of the syntaxin 5gene induces Golgi fragmentation but capable of transporting vesicles. FEBS Lett. 2005; 579(20):4226–4234. [PubMed: 16081076]
30. Sohda M, Misumi Y, Yoshimura S, Nakamura N, Fusano T, Ogata S, Sakisaka S, Ikehara Y. Theinteraction of two tethering factors, p115 and COG complex, is required for Golgi integrity.Traffic. 2007; 8(3):270–284. [PubMed: 17274799]
31. Shorter J, Beard MB, Seemann J, Dirac-Svejstrup AB, Warren G. Sequential tethering of Golginsand catalysis of SNAREpin assembly by the vesicle-tethering protein p115. J Cell Biol. 2002;157(1):45–62. [PubMed: 11927603]
32. Puthenveedu MA, Linstedt AD. Gene replacement reveals that p115/SNARE interactions areessential for Golgi biogenesis. Proc Natl Acad Sci U S A. 2004; 101(5):1253–1256. [PubMed:14736916]
33. Malsam J, Satoh A, Pelletier L, Warren G. Golgin tethers define subpopulations of COPI vesicles.Science. 2005; 307(5172):1095–1098. [PubMed: 15718469]
34. White J, Johannes L, Mallard F, Girod A, Grill S, Reinsch S, Keller P, Tzschaschel B, Echard A,Goud B, Stelzer EH. Rab6 coordinates a novel Golgi to ER retrograde transport pathway in livecells. J Cell Biol. 1999; 147(4):743–760. [published erratum appears in J Cell Biol 2000 Jan10;148(1):following table of contents]. [PubMed: 10562278]
35. Bujny MV, Popoff V, Johannes L, Cullen PJ. The retromer component sorting nexin-1 is requiredfor efficient retrograde transport of Shiga toxin from early endosome to the trans Golgi network. JCell Sci. 2007; 120(Pt 12):2010–2021. [PubMed: 17550970]
36. Utskarpen A, Slagsvold HH, Dyve AB, Skanland SS, Sandvig K. SNX1 and SNX2 mediateretrograde transport of Shiga toxin. Biochem Biophys Res Commun. 2007; 358(2):566–570.[PubMed: 17498660]
37. Llorente A, Lauvrak SU, van Deurs B, Sandvig K. Induction of direct endosome to endoplasmicreticulum transport in Chinese hamster ovary (CHO) cells (LdlF) with a temperature-sensitivedefect in epsilon-coatomer protein (epsilon-COP). J Biol Chem. 2003; 278(37):35850–35855.[PubMed: 12847103]
38. Xu Y, Martin S, James DE, Hong W. GS15 forms a SNARE complex with syntaxin 5, GS28, andYkt6 and is implicated in traffic in the early cisternae of the Golgi apparatus. Mol Biol Cell. 2002;13(10):3493–3507. [PubMed: 12388752]
39. Tai G, Lu L, Wang TL, Tang BL, Goud B, Johannes L, Hong W. Participation of the Syntaxin 5/Ykt6/GS28/GS15 SNARE Complex in Transport from the Early/Recycling Endosome to the TGN.Mol Biol Cell. 2004
40. Feng Y, Jadhav AP, Rodighiero C, Fujinaga Y, Kirchhausen T, Lencer WI. Retrograde transport ofcholera toxin from the plasma membrane to the endoplasmic reticulum requires the trans-Golginetwork but not the Golgi apparatus in Exo2-treated cells. EMBO Rep. 2004; 5(6):596–601.[PubMed: 15153932]
41. Sannerud R, Saraste J, Goud B. Retrograde traffic in the biosynthetic-secretory route: pathwaysand machinery. Curr Opin Cell Biol. 2003; 15(4):438–445. [PubMed: 12892784]
42. Lencer WI, Tsai B. The intracellular voyage of cholera toxin: going retro. Trends Biochem Sci.2003; 28(12):639–645. [PubMed: 14659695]
Smith et al. Page 13
Traffic. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

43. Tarrago-Trani MT, Jiang S, Harich KC, Storrie B. Shiga-like toxin subunit B (SLTB)-enhanceddelivery of chlorin e6 (Ce6) improves cell killing. Photochem Photobiol. 2006; 82(2):527–537.[PubMed: 16613509]
44. Suvorova ES, Kurten RC, Lupashin VV. Identification of a human orthologue of Sec34p as acomponent of the cis-Golgi vesicle tethering machinery. Journal of Biological Chemistry. 2001;276(25):22810–22818. [PubMed: 11292827]
45. Alvarez C, Fujita H, Hubbard A, Sztul E. ER to Golgi transport: Requirement for p115 at a pre-Golgi VTC stage. J Cell Biol. 1999; 147(6):1205–1222. [PubMed: 10601335]
46. Sapperstein SK, Walter DM, Grosvenor AR, Heuser JE, Waters MG. p115 is a general vesiculartransport factor related to the yeast endoplasmic reticulum to Golgi transport factor Uso1p. ProcNatl Acad Sci U S A. 1995; 92(2):522–526. [PubMed: 7831323]
47. Gullapalli A, Garrett TA, Paing MM, Griffin CT, Yang Y, Trejo J. A role for sorting nexin 2 inepidermal growth factor receptor down-regulation: evidence for distinct functions of sorting nexin1 and 2 in protein trafficking. Mol Biol Cell. 2004; 15(5):2143–2155. [PubMed: 14978220]
48. Popoff V, Mardones GA, Tenza D, Rojas R, Lamaze C, Bonifacino JS, Raposo G, Johannes L. Theretromer complex and clathrin define an early endosomal retrograde exit site. J Cell Sci. 2007;120(Pt 12):2022–2031. [PubMed: 17550971]
49. Suga K, Saito A, Tomiyama T, Mori H, Akagawa K. Syntaxin 5 interacts specifically withpresenilin holoproteins and affects processing of betaAPP in neuronal cells. J Neurochem. 2005;94(2):425–439. [PubMed: 15998293]
50. Majoul I, Straub M, Hell SW, Duden R, Soling HD. KDEL-cargo regulates interactions betweenproteins involved in COPI vesicle traffic: measurements in living cells using FRET. Dev Cell.2001; 1(1):139–153. [PubMed: 11703931]
51. Whyte JR, Munro S. The Sec34/35 Golgi Transport Complex Is Related to the Exocyst, Defining aFamily of Complexes Involved in Multiple Steps of Membrane Traffic. Developmental Cell. 2001;1(4):527–537. [PubMed: 11703943]
52. Jiang S, Storrie B. Cisternal rab proteins regulate Golgi apparatus redistribution in response tohypotonic stress. Mol Biol Cell. 2005; 16(5):2586–2596. [PubMed: 15758030]
Smith et al. Page 14
Traffic. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 1. SubAB-mediated GRP78 cleavage is inhibited in stable Cog4 KD HeLa cellsGalNAcT2-GFP HeLa cells were mock-transfected or transfected with pCog4 shRNAplasmid and maintained in selective puromycin-containing media. One month aftertransfection, the cells were solubilized in phosphate-buffered saline with 2% SDS, and thelysates were separated by SDS-PAGE and analyzed by immunoblotting with the indicatedantibodies (A) or fixed and visualized by fluorescent microscopy (B). Bar, 10 µm. Effect ofCog4 KD on SubAB-induced cleavage of GRP78. Control and Cog4 sKD cells were treatedwith SubAB (3 µg/ml) and incubated at 37°C for the time indicated. Panel C shows a blotrepresentative of three such experiments. (D) Graph of SubAB-induced GRP78 cleavage forcontrol, Cog4 sKD, and HeLa cells treated with 30 µM Nocodazole for 2 hours prior totoxin application. (E) Binding of SubAB-HF555 to control and Cog4 KD GalNAcT2-GFPHeLa cells. Cells were exposed to SubAB-Hylite 555 toxin for 20 min at 4°C, formalin fixedand viewed by laser confocal microscopy (63x oil objective). Bar, 10 µm.
Smith et al. Page 15
Traffic. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
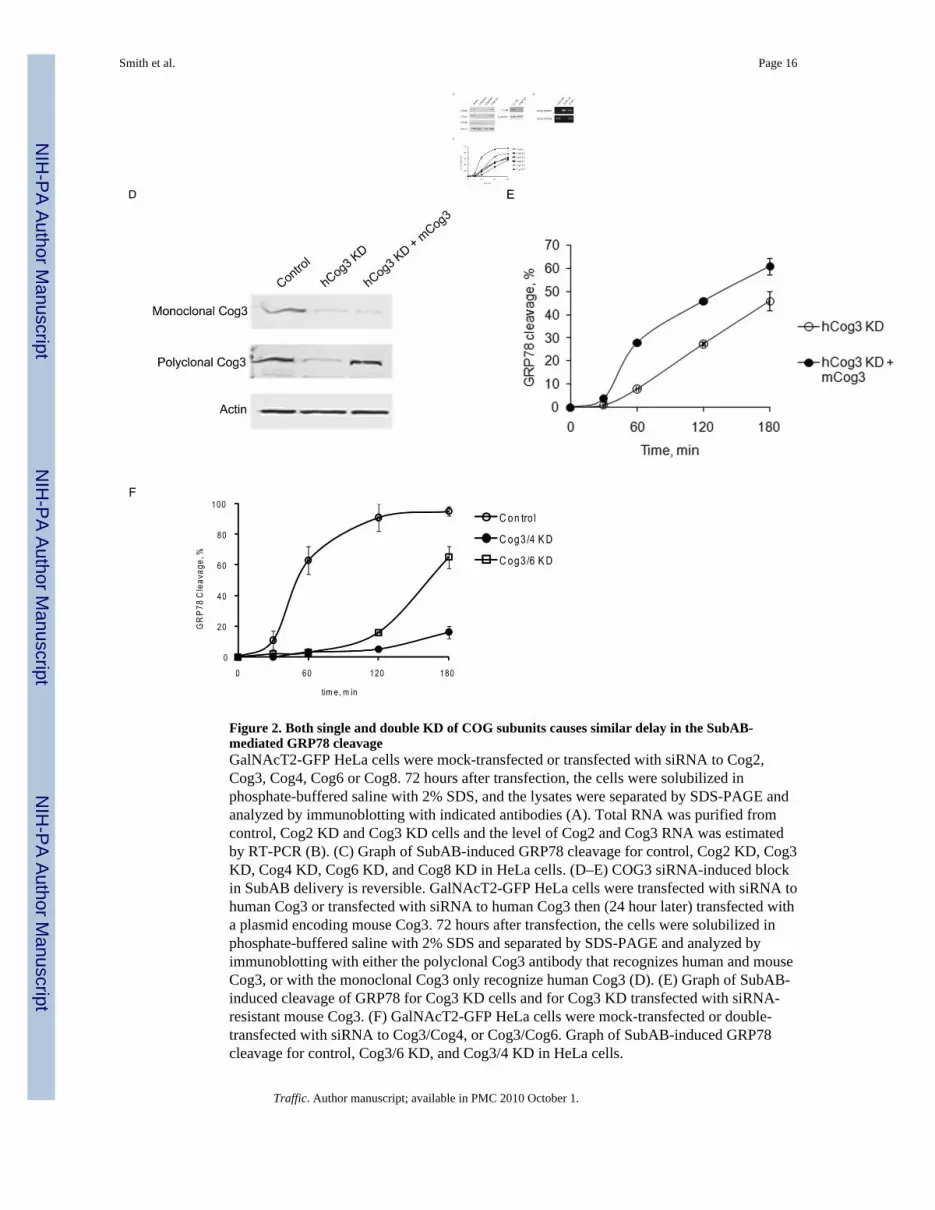
Figure 2. Both single and double KD of COG subunits causes similar delay in the SubAB-mediated GRP78 cleavageGalNAcT2-GFP HeLa cells were mock-transfected or transfected with siRNA to Cog2,Cog3, Cog4, Cog6 or Cog8. 72 hours after transfection, the cells were solubilized inphosphate-buffered saline with 2% SDS, and the lysates were separated by SDS-PAGE andanalyzed by immunoblotting with indicated antibodies (A). Total RNA was purified fromcontrol, Cog2 KD and Cog3 KD cells and the level of Cog2 and Cog3 RNA was estimatedby RT-PCR (B). (C) Graph of SubAB-induced GRP78 cleavage for control, Cog2 KD, Cog3KD, Cog4 KD, Cog6 KD, and Cog8 KD in HeLa cells. (D–E) COG3 siRNA-induced blockin SubAB delivery is reversible. GalNAcT2-GFP HeLa cells were transfected with siRNA tohuman Cog3 or transfected with siRNA to human Cog3 then (24 hour later) transfected witha plasmid encoding mouse Cog3. 72 hours after transfection, the cells were solubilized inphosphate-buffered saline with 2% SDS and separated by SDS-PAGE and analyzed byimmunoblotting with either the polyclonal Cog3 antibody that recognizes human and mouseCog3, or with the monoclonal Cog3 only recognize human Cog3 (D). (E) Graph of SubAB-induced cleavage of GRP78 for Cog3 KD cells and for Cog3 KD transfected with siRNA-resistant mouse Cog3. (F) GalNAcT2-GFP HeLa cells were mock-transfected or double-transfected with siRNA to Cog3/Cog4, or Cog3/Cog6. Graph of SubAB-induced GRP78cleavage for control, Cog3/6 KD, and Cog3/4 KD in HeLa cells.
Smith et al. Page 16
Traffic. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
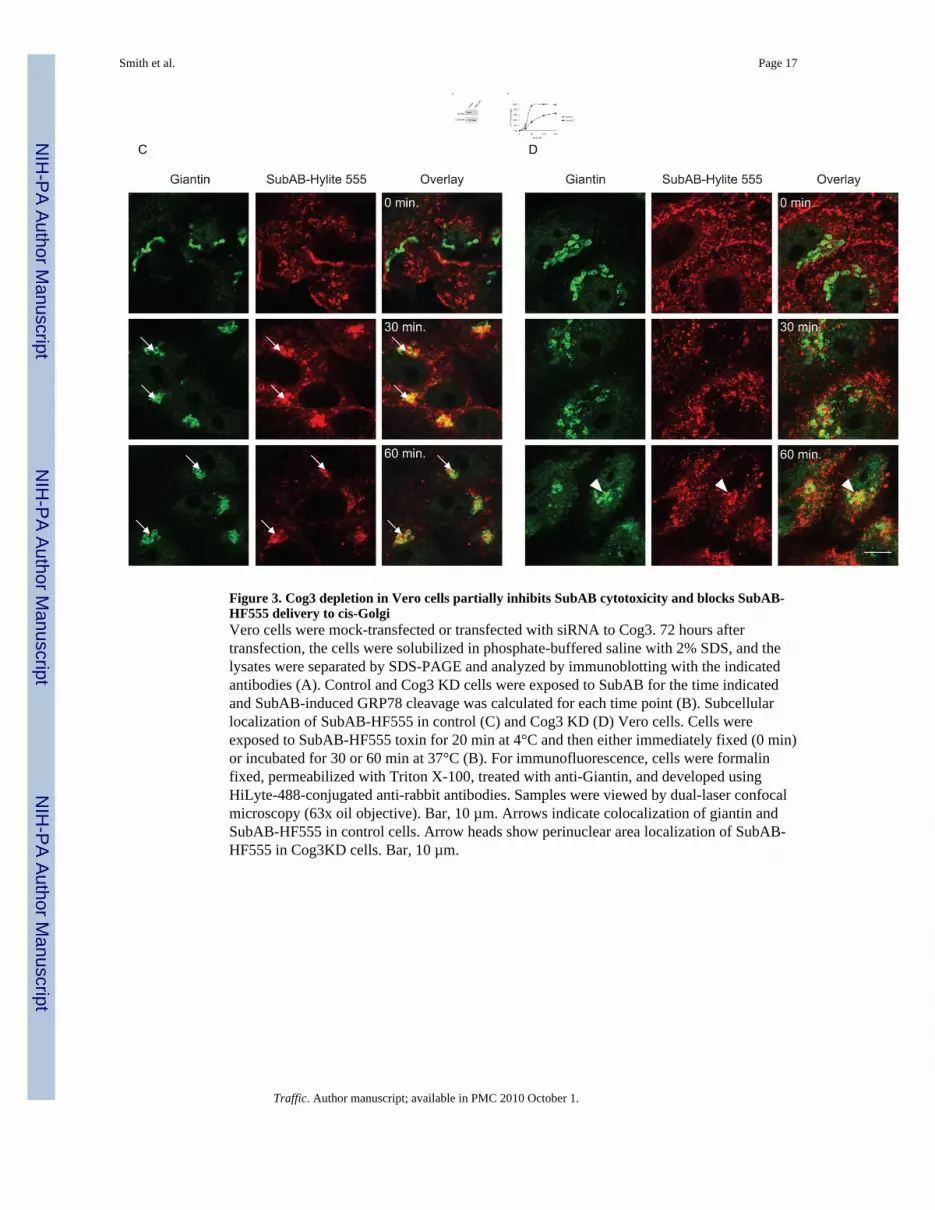
Figure 3. Cog3 depletion in Vero cells partially inhibits SubAB cytotoxicity and blocks SubAB-HF555 delivery to cis-GolgiVero cells were mock-transfected or transfected with siRNA to Cog3. 72 hours aftertransfection, the cells were solubilized in phosphate-buffered saline with 2% SDS, and thelysates were separated by SDS-PAGE and analyzed by immunoblotting with the indicatedantibodies (A). Control and Cog3 KD cells were exposed to SubAB for the time indicatedand SubAB-induced GRP78 cleavage was calculated for each time point (B). Subcellularlocalization of SubAB-HF555 in control (C) and Cog3 KD (D) Vero cells. Cells wereexposed to SubAB-HF555 toxin for 20 min at 4°C and then either immediately fixed (0 min)or incubated for 30 or 60 min at 37°C (B). For immunofluorescence, cells were formalinfixed, permeabilized with Triton X-100, treated with anti-Giantin, and developed usingHiLyte-488-conjugated anti-rabbit antibodies. Samples were viewed by dual-laser confocalmicroscopy (63x oil objective). Bar, 10 µm. Arrows indicate colocalization of giantin andSubAB-HF555 in control cells. Arrow heads show perinuclear area localization of SubAB-HF555 in Cog3KD cells. Bar, 10 µm.
Smith et al. Page 17
Traffic. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 4. Fluorescently labelled SubAB partially localizes with TGN46-positive compartments inCog3-depleted cellsVero cells were mock-transfected or transfected with siRNA to Cog3. 72 hours aftertransfection, the cells were treated with 1 µg/ml SubAB HF555 for one hour. Control (A)and Cog3 cells (B) were fixed in 4% paraformaldehyde, permeabilized with Triton X-100,stained with anti-TGN46, anti-EEA1, anti-Rab7, and anti-LAMP2, and developed usingHiLyte-488-conjugated-anti-mouse and HiLyte-488-conjugated-anti-rabbit antibodies.Samples were viewed by dual-laser confocal microscopy (63X oil objective). Bar, 10 µm.Arrows demonstrate colocalization between TGN46 and SubAB HF555.
Smith et al. Page 18
Traffic. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
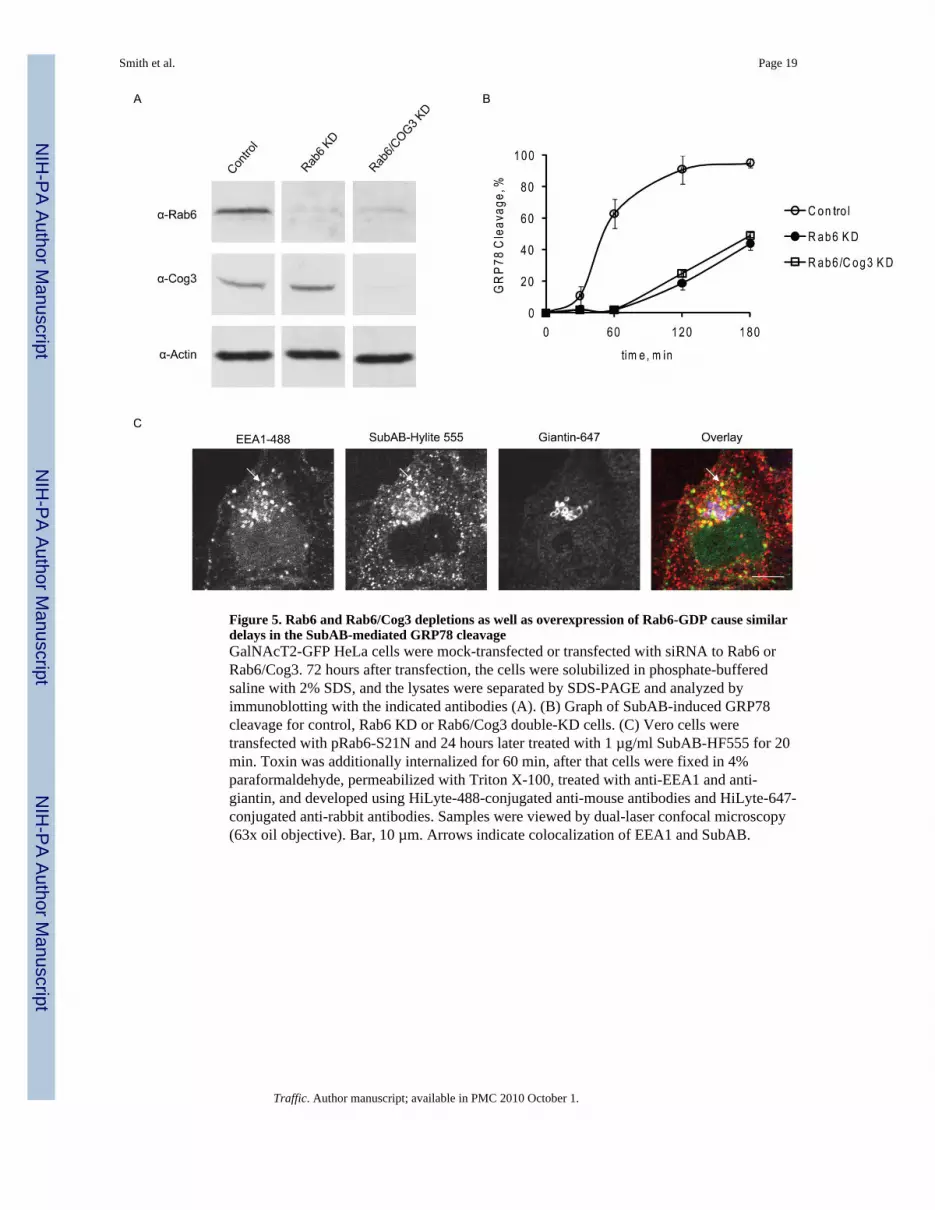
Figure 5. Rab6 and Rab6/Cog3 depletions as well as overexpression of Rab6-GDP cause similardelays in the SubAB-mediated GRP78 cleavageGalNAcT2-GFP HeLa cells were mock-transfected or transfected with siRNA to Rab6 orRab6/Cog3. 72 hours after transfection, the cells were solubilized in phosphate-bufferedsaline with 2% SDS, and the lysates were separated by SDS-PAGE and analyzed byimmunoblotting with the indicated antibodies (A). (B) Graph of SubAB-induced GRP78cleavage for control, Rab6 KD or Rab6/Cog3 double-KD cells. (C) Vero cells weretransfected with pRab6-S21N and 24 hours later treated with 1 µg/ml SubAB-HF555 for 20min. Toxin was additionally internalized for 60 min, after that cells were fixed in 4%paraformaldehyde, permeabilized with Triton X-100, treated with anti-EEA1 and anti-giantin, and developed using HiLyte-488-conjugated anti-mouse antibodies and HiLyte-647-conjugated anti-rabbit antibodies. Samples were viewed by dual-laser confocal microscopy(63x oil objective). Bar, 10 µm. Arrows indicate colocalization of EEA1 and SubAB.
Smith et al. Page 19
Traffic. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 6. β-COP and β-COP/Cog3 knockdowns along with overexpression of Arf1-Q71L causedelays in the SubAB-mediated GRP78 cleavage and block the traffic of SubABGalNAcT2-GFP HeLa cells were mock-transfected or transfected with siRNA to β-COP orβ-COP/Cog3. 72 hours after transfection, the cells were solubilized in phosphate-bufferedsaline with 2% SDS, and the lysates were separated by SDS-PAGE and analyzed byimmunoblotting with the indicated antibodies (A). (B) Graph of SubAB-induced GRP78cleavage for control, β-COP KD or β-COP/Cog3 double-KD cells. (C) Vero cells weretransfected with the CFP-Arf1-Q71L plasmid and 24 hours later treated with 1 µg/mlSubAB-HF555 for 20 min. Toxin was additionally internalized for 60 min, after that cellswere fixed in 4% paraformaldehyde, permeabilized with Triton X-100, treated with anti-giantin, and developed using HiLyte-647-conjugated anti-rabbit antibodies. Samples wereviewed by dual-laser confocal microscopy (63x oil objective). Bar, 10 µm. The asteriskindicates a control cell not transfected with CFP-Arf1-Q71L.
Smith et al. Page 20
Traffic. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 7. SubAB-mediated GRP78 cleavage is completely blocked in Syntaxin 5 KD cellsGalNAcT2-GFP HeLa cells were mock-transfected or transfected with siRNA to Syntaxin 5.72 hours after transfection, the cells were solubilized in phosphate-buffered saline with 2%SDS, and the lysates were separated by SDS-PAGE and analyzed by immunoblotting withindicated antibodies (A). Note that both long and short isoforms of Syntaxin 5 weredepleted, while the non-specific protein band (indicated with an asterisk) was not. (B) Graphof SubAB-induced GRP78 cleavage for control and Syntaxin 5 KD cells. (C) Vero cellswere transfected with siRNA to Syntaxin 5. 72 hours later cell were treated with 1 µg/mlSubAB-HF555, fixed in 4% paraformaldehyde, permeabilized with Triton X-100, treatedwith anti-P230 and anti-giantin, and developed using HiLyte-488-conjugated anti-mouseantibodies and HiLyte-647-conjugated anti-rabbit antibodies. Samples were viewed by dual-laser confocal microscopy (63x oil objective). Bar, 10 µm.
Smith et al. Page 21
Traffic. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 8. The retrograde traffic of Subtilase cytotoxin is independent of p115 and golgin-84GalNAcT2-GFP HeLa cells were mock-transfected or transfected with siRNA to COG3,p115, or golgin-84. 72 hours after transfection, the cells were solubilized in phosphate-buffered saline with 2% SDS, and the lysates were separated by SDS-PAGE and analyzedby immunoblotting with the indicated antibodies (A). A non-specific protein band on agolgin84 blot is indicated with an asterisk. Effect of protein depletion on SubAB-inducedcleavage of GRP78. Control, COG3KD, p115 KD and golgin-84 KD cells (B) were treatedwith SubAB (3 µg/ml) and incubated at 37°C for time indicated. Each lane on the graph iscalculated from three independent experiments. (C). GalNAcT2-GFP HeLa cells weremock-transfected or transfected with siRNA to Golgin84 or p115Cells. 72 hour later cellswere fixed and visualized by fluorescent microscopy. Bar, 10 µm.
Smith et al. Page 22
Traffic. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
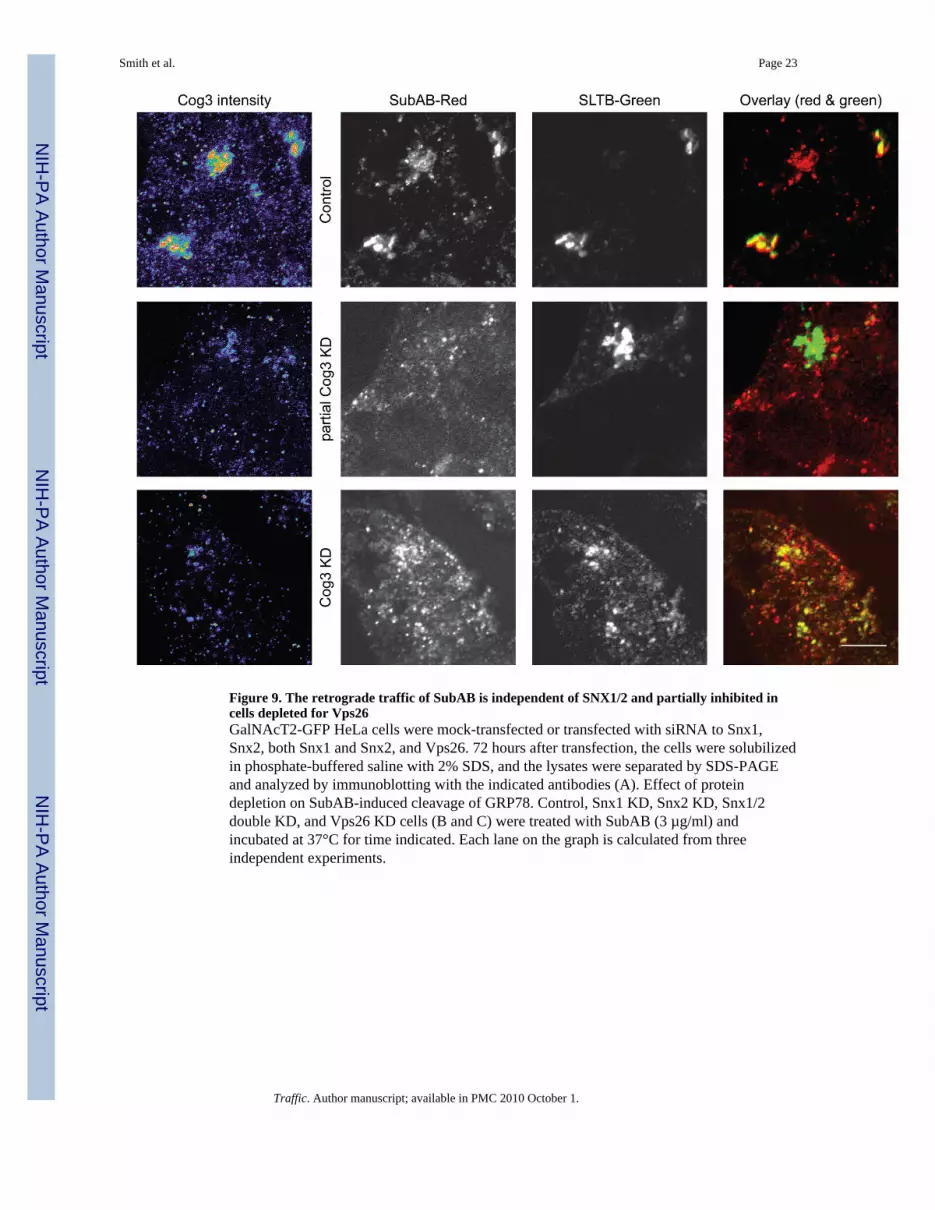
Figure 9. The retrograde traffic of SubAB is independent of SNX1/2 and partially inhibited incells depleted for Vps26GalNAcT2-GFP HeLa cells were mock-transfected or transfected with siRNA to Snx1,Snx2, both Snx1 and Snx2, and Vps26. 72 hours after transfection, the cells were solubilizedin phosphate-buffered saline with 2% SDS, and the lysates were separated by SDS-PAGEand analyzed by immunoblotting with the indicated antibodies (A). Effect of proteindepletion on SubAB-induced cleavage of GRP78. Control, Snx1 KD, Snx2 KD, Snx1/2double KD, and Vps26 KD cells (B and C) were treated with SubAB (3 µg/ml) andincubated at 37°C for time indicated. Each lane on the graph is calculated from threeindependent experiments.
Smith et al. Page 23
Traffic. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
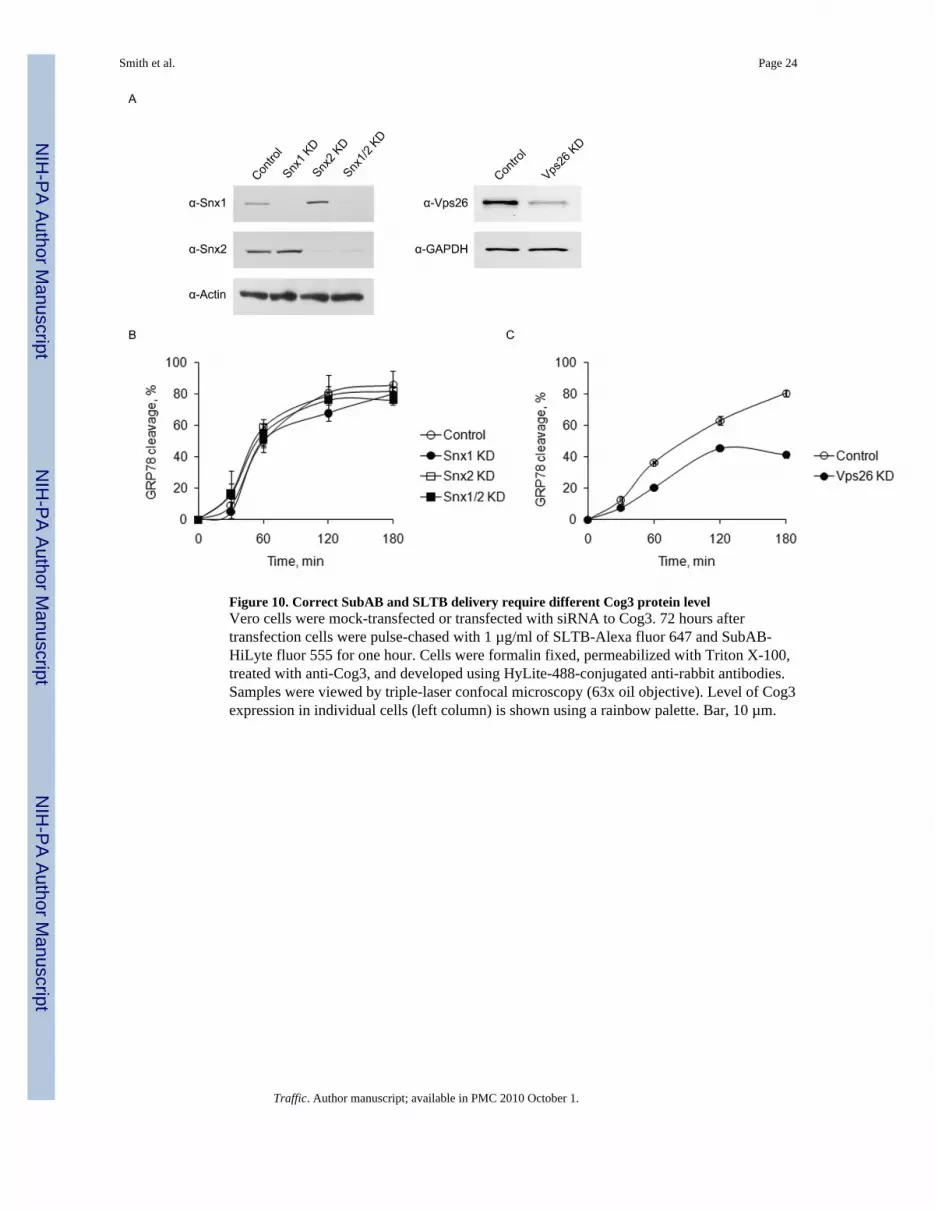
Figure 10. Correct SubAB and SLTB delivery require different Cog3 protein levelVero cells were mock-transfected or transfected with siRNA to Cog3. 72 hours aftertransfection cells were pulse-chased with 1 µg/ml of SLTB-Alexa fluor 647 and SubAB-HiLyte fluor 555 for one hour. Cells were formalin fixed, permeabilized with Triton X-100,treated with anti-Cog3, and developed using HyLite-488-conjugated anti-rabbit antibodies.Samples were viewed by triple-laser confocal microscopy (63x oil objective). Level of Cog3expression in individual cells (left column) is shown using a rainbow palette. Bar, 10 µm.
Smith et al. Page 24
Traffic. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 11. Model of retrograde transport of Subtilase cytotoxinThe solid arrows indicate the Rab6/COG complex/COPI-dependent pathway that SubABtraffics through the HeLa cell. The dotted arrow indicates that Syntaxin 5 will completelyinhibit both the Rab6/COG complex/COPI-dependent and –independent pathways.
Smith et al. Page 25
Traffic. Author manuscript; available in PMC 2010 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Smith et al. Page 26
TABLE 1Potency and efficacy of SubAB on control and single KDs in HeLa cells treated for 3hours
GalNAcT2-GFP HeLa cells were mock-transfected or transfected with siRNA to Cog2, Cog3, Cog4, Cog6,Cog8, Rab6, Syntaxin 5, β-COP, and Vps26. 72 hours after transfection, the cells were intoxicated withSubAB for 3 hours at 0-, 0.074-, 0.222-, 0.667-, 2-, and 6 µg/ml. Cells were solubilized in phosphate-bufferedsaline with 2% SDS, and the lysates were separated by SDS-PAGE with the GRP78 antibody. Results ofGRP78 cleavage by SubAB was calculated using the GraphPad Prism software to demonstrate IC50, r-squared, and Imax values.
IC50 (µg/ml) r-squared Imax (%)
Control 0.153 0.954 80
Cog2 KD 0.788 0.998 79
Cog3 KD 0.579 0.999 66
Cog4 KD 0.388 0.936 33
Cog6 KD 0.645 0.979 70
Cog8 KD 0.656 0.988 63
Rab6 KD 0.739 0.966 62
β-COP KD 3.65 0.897 37
Syntaxin 5 KD >6 No fit 0
Vps26 KD 0.417 0.996 56
Traffic. Author manuscript; available in PMC 2010 October 1.