the arctic fox (alopex lagopus): an opportunistic specialist
TRANSCRIPT

The arctic fox (Alopex lagopus): an opportunistic specialist
Bodil Elmhagen, Magnus Tannerfeldt, Paolo Verucci and Anders AngerbjoÈrn
Department of Zoology, Stockholm University, S-106 91 Stockholm, Sweden
(Accepted 7 July 1999)
Abstract
Reliable and abundant resources are likely to favour specialization, while unpredictable environmental
variation should favour a generalist strategy. The rodent population cycles of northern latitudes can be
seen as both predictable and unpredictable, depending on the scale in time and space. The arctic fox
Alopex lagopus is an opportunistic carnivore, but paradoxically, it seems to function as a specialist on
¯uctuating rodent Arvicolinae populations in most inland areas. We have studied the dietary response of
arctic foxes in Sweden during 5 years of varying abundance of Norwegian lemming Lemmus lemmus, and
how these changes in¯uenced the reproductive success of the foxes. The arctic fox population on mainland
Fennoscandia is threatened by extinction and the situation has deteriorated during the 1980s and 1990s
because of an absence of lemming peaks. Our results showed that in all years, lemming was the main prey
for arctic foxes, with 85% frequency of occurrence in summer faeces (scats). Bird remains (mainly
Passeriformes) were present in 34% of the scats, reindeer Rangifer tarandus in 21%, voles and shrews in 4%
and hares Lepus timidus in 2% of the scats. The occurrences of lemming, bird and larger mammal (reindeer
and hare) remains in the scats varied signi®cantly between years. Temporal variations within summer
seasons and dietary differences between sub-areas, indicated that arctic foxes fed opportunistically on the
alternative prey types. Den occupancy rates were positively correlated with lemming population densities
during the previous winter, indicating a strong numerical response. We conclude that from a functional
aspect, the arctic fox in Sweden is a lemming specialist, since lemming is the main prey and their abundance
is the best predictor of arctic fox reproductive success. Other prey are used opportunistically in relation to
their availability.
Key words: Alopex lagopus, diet, predation, numerical response, functional response
INTRODUCTION
Food availability is the most important environmentalfactor in¯uencing reproduction in mammals, func-tioning both as a proximate and ultimate cause forannual variations in reproductive output (Bronson,1989). A predator showing strong preference for oneprey species does not change its diet when the abun-dance of that species decreases, while a predator withweak preferences can switch to another type of prey(Murdoch, 1969). The degree of specialization can alsobe de®ned with regard to the function of a predator in apredator guild. The specialist predator can thus berestricted in its diet not only by preference, but alsothrough lack of ability or possibility to use other preyspecies. The generalist predator is less dependent on theavailability of one particular prey species and abandonsa declining prey type earlier than the specialist. As aconsequence, the specialist will have a more pronounced
numerical response to changes in abundance of aparticular prey species than the semi-generalist orgeneralist (Andersson & Erlinge, 1977). The specialist±generalist concepts can thus be used to analyse thestrategies or functions of different predators within aprey community (see e.g. Reid, Krebs & Kenney, 1997;Wiklund et al., 1999).
The response of a predator to changes in the avail-ability of prey depends on the predator's strategy.Andersson & Erlinge (1977) suggested that a specialistwould have a type II functional response, i.e. thenumber of killed prey would increase rapidly with preydensity. A generalist would show a slower increase,resulting in the sigmoid curve of a type III response(Holling, 1959). This could be the result of changes insearch image, foraging habitat or diet preferences(Taylor, 1984). The functional (dietary) and numericalresponses of a predator to changes in prey abundancemay also exert a strong in¯uence on the population
J. Zool., Lond. (2000) 251, 139±149 # 2000 The Zoological Society of London Printed in the United Kingdom

dynamics of the prey (Pearson, 1966; Andersson &Erlinge, 1977; Erlinge et al., 1984; Hanski, Hansson &Henttonen, 1991; Boutin, 1995). An issue under contin-uous debate is to what extent population cycles in smallrodents are regulated by predators (Henttonen et al.,1987; KorpimaÈki, Norrdahl & Rinta-Jaskari, 1991;Hanski, Turchin et al., 1993; Hanski & KorpimaÈki,1995; Sittler, 1995; Stenseth, Bjùrnstad & Falck, 1996).
The arctic fox Alopex lagopus is a circumpolar species,breeding above the tree line on the arctic tundra in NorthAmerica and Eurasia and on the alpine tundra inFennoscandia (Macpherson, 1969). Several studies haveshown that the arctic fox is a highly opportunisticomnivore (see e.g. Macpherson, 1969; Fine, 1980;Garrott, Eberhardt & Hanson, 1983; Hiruki & Stirling,1989; Prestrud, 1992; Hersteinsson & Macdonald, 1996;AngerbjoÈrn, Tannerfeldt & Erlinge, 1999; Dalerum &AngerbjoÈrn, 2000). Arctic foxes readily change their dietif the abundance of different food types varies with time(Hersteinsson, 1984; Fay & Stephenson, 1989; Stickney,1991) or when they move to another den within the sameseason (Frafjord, 1993b). There are also indications ofindividual differences in diet between foxes within thesame population (AngerbjoÈrn, Hersteinsson et al., 1994;Pond et al., 1995; AngerbjoÈrn, Stroman & Becker, 1997).
The arctic fox is found in two main habitat types,with marked differences in diets and patterns of repro-duction. Foxes living near ice-free coasts have access toboth inland prey and sea birds, seal carcasses, ®sh andinvertebrates connected to the marine environment,leading to relatively stable food availability and a moregeneralist strategy. Inland foxes are usually specialistsor semi-generalists (Reid et al., 1997), relying on ¯uctu-ating microtine rodent (Arvicolinae) populations(Braestrup, 1941; Prestrud, 1992; AngerbjoÈrn, Tanner-feldt, BjaÈrvall et al., 1995; Hersteinsson & Macdonald,1996; Dalerum & AngerbjoÈrn, 2000). This has led to theevolution of two different reproductive strategies, wherearctic foxes with a stable food supply produce relativelyfew cubs every year, while foxes depending on ¯uctu-ating prey populations have larger mean and maximumlitter sizes (Frafjord, 1993a; Tannerfeldt & AngerbjoÈrn,1998). Within each strategy, there is also variation inlitter size caused by regional changes in food abundance,affecting pre-natal mortality and cub survival (reviewedby Tannerfeldt & AngerbjoÈrn, 1998).
The arctic fox on mainland Fennoscandia is heavilydependent on ¯uctuating populations of rodents, mainlyNorwegian lemmings Lemmus lemmus, and the foxpopulation follows their prey in 3- to 5-year cycles(Finerty, 1980; AngerbjoÈrn, Tannerfeldt, BjaÈrvall et al.,1995; Kaikusalo & AngerbjoÈrn, 1995). Hunting drovethis arctic fox population close to extinction at thebeginning of the 20th century. It has not recovered,despite total protection for almost 60 years (Her-steinsson, AngerbjoÈrn et al., 1989). The situation hasbecome more severe during the 1980s and 1990s as aresult of a prolonged absence of lemming peaks. In theSwedish mountain range, the lemming population hasnot peaked since 1982 (1985 in the southernmost part).
During this period, lemming abundance has beengenerally low, with only small amplitude ¯uctuations(Lundberg, 1999). Within our study area, a 20-yearstudy on the structure of the bird community showedthat the abundance of breeding birds did not varysigni®cantly between years (Svensson, Carlsson &Liljedahl, 1984) and there is no indication of anythingbut random variation in the availability of reindeercarcasses. Feeding experiments in the ®eld have shownthat the foxes are food stressed. Winter feeding at densresulted in more breeding attempts, and more of thesewere successful than without feeding (AngerbjoÈrn,ArvidSon et al., 1991), while summer feeding led tohigher cub survival (Tannerfeldt, AngerbjoÈrn &ArvidSon, 1994).
In spite of the low lemming abundance, some foxeshave reproduced in most years. However, it is not clearwhat effect changes in lemming availability have on thecomposition of the summer diet, i.e. if foxes switch toother kinds of prey when lemming availability is low(Hersteinsson, AngerbjoÈrn et al., 1989). The aim of thisstudy, which was carried out within the Swedish ArcticFox Project, was to ®nd out to what extent arctic foxeschange their diet as a response to varying microtineabundance, and how such changes in¯uence reproductivesuccess.
METHODS
Study area
The study area was situated in the nature reserve ofVindelfjaÈllen in Swedish Lapland. It covered 1900 km2
of alpine tundra, mainly dry grass heath, meadows andwillow wetlands. The plant species diversity is lowexcept for patches with higher diversity (Rune, 1981).Below the tree line, at about 750 m a.s.l., there are birchand coniferous forests.
The study area has been surveyed yearly since 1985by the Swedish Arctic Fox Project (Hersteinsson,AngerbjoÈrn et al., 1989; AngerbjoÈrn, ArvidSon et al.,1991; Tannerfeldt, 1997). It was divided into 5 sub-areas of about equal size, separated by river valleys(Fig. 1). Sub-areas A and B were more alpine and hada harsher climate than the other sub-areas. Sub-area Cwas a medium-altitude area, with gently sloping moun-tains. The terrain in D was similar to that of C butcloser to forested valleys and lower in altitude, withonly a small part reaching over 1000 m a.s.l. E was themost varied sub-area, including both low altitude areasclose to forests and a peak at 1249 m a.s.l.
There were about 90 arctic fox dens in the study area,most of them situated at 900±1000 m a.s.l. A few denswere occupied regularly, although most have not beeninhabited since surveys began in 1985. Available preyfor arctic foxes were Norwegian lemming, ®eld voleMicrotus agrestis, bank vole Clethrionomys glareolus,grey-sided vole Clethrionomys rufocanus, shrewsSoricidae, mountain hare Lepus timidus and different
B. Elmhagen ET AL.140

kinds of birds. Ptarmigan Lagopus mutus and willowgrouse Lagopus lagopus were resident in the area whilepasserines Passeriformes, gulls and waders Charadrii-formes were present only in the summer. There werealso reindeer Rangifer tarandus in the area. Arctic foxesseldom kill reindeer but they feed from carcasses, espe-cially during the winter (Kaikusalo & AngerbjoÈrn,1995). Potential predators on the arctic foxes were redfox Vulpes vulpes, wolverine Gulo gulo, golden eagleAquila chrysaeÈtos and white-tailed eagle HaliaeeÈtusalbicilla.
Den surveys
We visited nearly all dens in the area at least once everysummer. Since we have a good knowledge of the qualityof the dens, and the only dens that were not visitedevery year were of low quality, we are con®dent that wefound almost all reproducing foxes within the studyarea. We counted the cubs at the inhabited dens. Thiswas usually done around the time of emergence in earlyJuly when the cubs were 3±4 weeks old. However, since
some cubs might die in the den before this and it is notcertain that all cubs are outside the den simultaneously,our data on litter sizes are minimum numbers (Garrottet al., 1983; AngerbjoÈrn, Tannerfeldt, BjaÈrvall et al.,1995; Tannerfeldt & AngerbjoÈrn, 1998).
In 1990 and from 1994 onwards, scats were collectedon the dens at each visit. Fresh scats were separatedfrom those of previous seasons by colour, position inthe vegetation and smell. During the period in question,juveniles only ate food brought to them by the adults, sowe did not separate scats from adult and juvenile foxes.We separated samples from each den at each `collectioninstance'. All scats were removed from the den, allowingtemporal analysis using subsequent collections. Toavoid confusion with red fox scats, we only collectedscats at dens inhabited by breeding arctic foxes. Beforeanalysis, all scats were dried in 105 8C.
Scat analysis
We analysed fresh scats from 1 collection instance perden to investigate differences in the summer diet
141Diet of Alopex lagopus
N
Fig. 1. Map of the study area with the locations of the dens included in the dietary study. For each den, the years in which the
diet was investigated are shown.

between years and sub-areas. As there are no conversionfactors for arctic fox diet, we had to assume that theproportion of remains of a prey species in the scats givesa fair representation of its proportion in the diet. Allcollections were between 9 July and 5 August. Tocompare the diet in August with that of July, we usedfresh scats from both the ®rst collection and a latercollection. In this analysis we included scats from 2 densin 1995-97 and 1 den in 1990. The scats from July werecollected between 13 and 25 July, while scats repre-senting the diet in August were collected 28±38 dayslater for each den. Scats collected in June, July andAugust at 1 den (C4) in sub-area C in 1997 were alsoanalysed. In total, 796 scats were analysed. There werecubs in all dens included in our study with the exceptionof 1997 when there was only 1 breeding den in the area.The other dens included that year were dens occupiedby fox pairs, which indicated that the foxes had tried toreproduce but failed early, presumably as a result offood shortage.
The individual scats were fragmented by hand undera magnifying glass and the proportion of fur, feathers,eggshells, insects and vegetation of the compacted dryvolume of the scat was estimated by eye. If the scatswere small or fragmented, they were combined so thatthe analysed volume in each sample was approximatelythat of 1 medium-sized scat. The fur was identi®ed asthat of reindeer, hare or small mammal using a referencecollection. Small mammals (lemmings, voles andshrews) were determined to species using the descrip-tions of molar teeth in Niethammer & Krapp (1982).The number of teeth from different rodent species in redfox scats is proportional to the ingested proportions ofthe species (Lockie, 1959). We assumed this to be truealso for arctic foxes, and when we found teeth frommore than 1 species in a scat, we used the number ofteeth from each species to determine their relativeproportions. When there were no teeth, the fur wasclassi®ed as unidenti®ed small mammal. Lemmings wereclassi®ed as juveniles when their teeth were so small thatthe shape typical of lemmings had not been developedat the masticating surface, but was discernible at theroot of the teeth. We determined bird remains to orderin the scats collected between 1994 and 1997, using thekeys of Brom (1986) and Day (1966). No attempt wasmade to further classify eggshells, insects or plantmaterial and we did not include microscopic fragmentsin the analysis.
Statistics
Frequency of occurrence is a common way to presentresults of diet analyses, but it does not take the propor-tion of different prey categories into account. Therefore,we also converted the data into whole scat equivalents,WSEs, as described by AngerbjoÈrn, Tannerfeldt &Erlinge (1999). We summarized the volume percentagesof a food category in all scats collected at the sameoccasion at a den. Thus, we could determine how many
whole scats each category corresponded to. Forinstance, if we had 1 scat containing 30% bird (feathers)and 70% lemming (fur), and another scat containing70% bird and 30% lemming, these were regarded asequivalent to 1 scat containing only lemming and 1 scatwith only bird. The total number of scats, i.e. samplesizes, thus remained the same.
As the proportions of different prey categories in ascat are interdependent (Reynolds & Aebischer, 1991),we used the data on frequency of occurrence whentesting for statistical differences between years, monthsand sub-areas in log-linear likelihood analyses (G-test).It was not possible to include more than 1 prey categoryin the same model, so we tested these separately. Sometypes of prey occurred relatively infrequently in thescats and were not represented in all years and sub-areas. In those cases we used w2 analysis instead. Chi-square was also used to test for dietary differencesbetween June, July and August at den C4 in 1997.
Effect of lemming densities on diet and reproduction
Since 1990, we have counted lemming winter nestsobserved while walking between dens in our study area.The number of lemming winter nests observed per100 km in 1990±98 was calculated and this informationused as an index for changes in lemming abundancebetween years (see e.g. Sittler, 1995). The distances eachyear varied between 130 and 940 km, totalling 3300 km.
The proportion of lemmings in the summer diet andthe relative densities of lemming as shown by thenumber of lemming nests in the preceding winter wereused to test for a functional response.
We tested the effect of winter lemming abundance onarctic fox reproduction (number of occupied dens,number of litters, litter size) and summer diet. We alsotested the effect of summer diet on litter size. Linearregression was used for all these tests. Some dens in thearea had been provided with additional food in thewinter as part of other studies. Since winter feeding hasa positive effect on the number of occupied andbreeding dens (AngerbjoÈrn, ArvidSon et al., 1991) aswell as on litter size (AngerbjoÈrn, Tannerfeldt, BjaÈrvallet al., 1995), winter-fed dens were excluded from theseanalyses.
RESULTS
The most prominent result was the supreme dominanceof lemmings in the diet of arctic foxes, even in years oflow lemming abundance. This was true both forfrequency of occurrence and for whole scat equivalents.None the less, the remaining diet was highly varied. Thesummer diet consisted of lemmings, voles, shrews,reindeer, hare, birds, eggs, insects and vegetation(Fig. 2). Most of the vegetation was probably ingestedby accident, as it consisted of grass fragments and therewas no signi®cant variation in occurrence between
B. Elmhagen ET AL.142
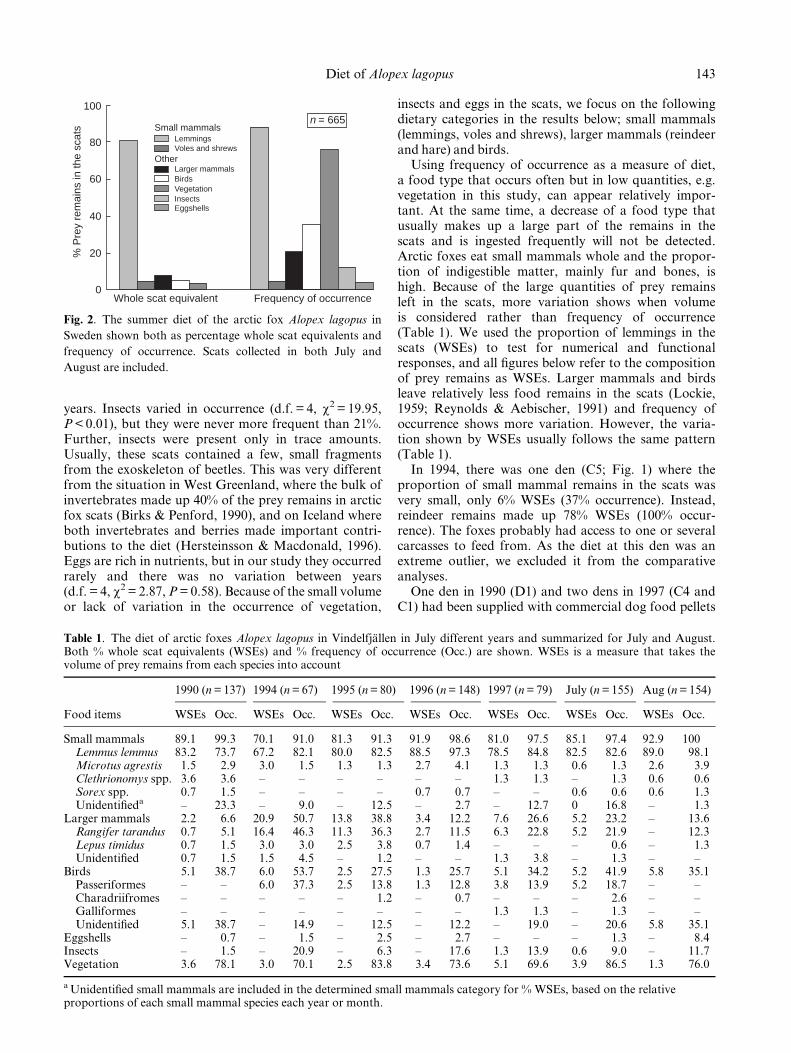
years. Insects varied in occurrence (d.f. = 4, w2 = 19.95,P < 0.01), but they were never more frequent than 21%.Further, insects were present only in trace amounts.Usually, these scats contained a few, small fragmentsfrom the exoskeleton of beetles. This was very differentfrom the situation in West Greenland, where the bulk ofinvertebrates made up 40% of the prey remains in arcticfox scats (Birks & Penford, 1990), and on Iceland whereboth invertebrates and berries made important contri-butions to the diet (Hersteinsson & Macdonald, 1996).Eggs are rich in nutrients, but in our study they occurredrarely and there was no variation between years(d.f. = 4, w2 = 2.87, P = 0.58). Because of the small volumeor lack of variation in the occurrence of vegetation,
insects and eggs in the scats, we focus on the followingdietary categories in the results below; small mammals(lemmings, voles and shrews), larger mammals (reindeerand hare) and birds.
Using frequency of occurrence as a measure of diet,a food type that occurs often but in low quantities, e.g.vegetation in this study, can appear relatively impor-tant. At the same time, a decrease of a food type thatusually makes up a large part of the remains in thescats and is ingested frequently will not be detected.Arctic foxes eat small mammals whole and the propor-tion of indigestible matter, mainly fur and bones, ishigh. Because of the large quantities of prey remainsleft in the scats, more variation shows when volumeis considered rather than frequency of occurrence(Table 1). We used the proportion of lemmings in thescats (WSEs) to test for numerical and functionalresponses, and all ®gures below refer to the compositionof prey remains as WSEs. Larger mammals and birdsleave relatively less food remains in the scats (Lockie,1959; Reynolds & Aebischer, 1991) and frequency ofoccurrence shows more variation. However, the varia-tion shown by WSEs usually follows the same pattern(Table 1).
In 1994, there was one den (C5; Fig. 1) where theproportion of small mammal remains in the scats wasvery small, only 6% WSEs (37% occurrence). Instead,reindeer remains made up 78% WSEs (100% occur-rence). The foxes probably had access to one or severalcarcasses to feed from. As the diet at this den was anextreme outlier, we excluded it from the comparativeanalyses.
One den in 1990 (D1) and two dens in 1997 (C4 andC1) had been supplied with commercial dog food pellets
143Diet of Alopex lagopus
Small mammalsLemmingsVoles and shrews
OtherLarger mammalsBirdsVegetationInsectsEggshells
n = 665100
80
60
40
20
0
% P
rey
rem
ains
in th
e sc
ats
Whole scat equivalent Frequency of occurrence
Fig. 2. The summer diet of the arctic fox Alopex lagopus in
Sweden shown both as percentage whole scat equivalents and
frequency of occurrence. Scats collected in both July and
August are included.
Table 1. The diet of arctic foxes Alopex lagopus in VindelfjaÈllen in July different years and summarized for July and August.Both % whole scat equivalents (WSEs) and % frequency of occurrence (Occ.) are shown. WSEs is a measure that takes thevolume of prey remains from each species into account
1990 (n = 137) 1994 (n = 67) 1995 (n = 80) 1996 (n = 148) 1997 (n = 79) July (n = 155) Aug (n = 154)
Food items WSEs Occ. WSEs Occ. WSEs Occ. WSEs Occ. WSEs Occ. WSEs Occ. WSEs Occ.
Small mammals 89.1 99.3 70.1 91.0 81.3 91.3 91.9 98.6 81.0 97.5 85.1 97.4 92.9 100Lemmus lemmus 83.2 73.7 67.2 82.1 80.0 82.5 88.5 97.3 78.5 84.8 82.5 82.6 89.0 98.1Microtus agrestis 1.5 2.9 3.0 1.5 1.3 1.3 2.7 4.1 1.3 1.3 0.6 1.3 2.6 3.9Clethrionomys spp. 3.6 3.6 ± ± ± ± ± ± 1.3 1.3 ± 1.3 0.6 0.6Sorex spp. 0.7 1.5 ± ± ± ± 0.7 0.7 ± ± 0.6 0.6 0.6 1.3Unidenti®eda ± 23.3 ± 9.0 ± 12.5 ± 2.7 ± 12.7 0 16.8 ± 1.3
Larger mammals 2.2 6.6 20.9 50.7 13.8 38.8 3.4 12.2 7.6 26.6 5.2 23.2 ± 13.6Rangifer tarandus 0.7 5.1 16.4 46.3 11.3 36.3 2.7 11.5 6.3 22.8 5.2 21.9 ± 12.3Lepus timidus 0.7 1.5 3.0 3.0 2.5 3.8 0.7 1.4 ± ± ± 0.6 ± 1.3Unidenti®ed 0.7 1.5 1.5 4.5 ± 1.2 ± ± 1.3 3.8 ± 1.3 ± ±
Birds 5.1 38.7 6.0 53.7 2.5 27.5 1.3 25.7 5.1 34.2 5.2 41.9 5.8 35.1Passeriformes ± ± 6.0 37.3 2.5 13.8 1.3 12.8 3.8 13.9 5.2 18.7 ± ±Charadriifromes ± ± ± ± ± 1.2 ± 0.7 ± ± ± 2.6 ± ±Galliformes ± ± ± ± ± ± ± ± 1.3 1.3 ± 1.3 ± ±Unidenti®ed 5.1 38.7 ± 14.9 ± 12.5 ± 12.2 ± 19.0 ± 20.6 5.8 35.1
Eggshells ± 0.7 ± 1.5 ± 2.5 ± 2.7 ± ± ± 1.3 ± 8.4Insects ± 1.5 ± 20.9 ± 6.3 ± 17.6 1.3 13.9 0.6 9.0 ± 11.7Vegetation 3.6 78.1 3.0 70.1 2.5 83.8 3.4 73.6 5.1 69.6 3.9 86.5 1.3 76.0
a Unidenti®ed small mammals are included in the determined small mammals category for % WSEs, based on the relativeproportions of each small mammal species each year or month.
during summer as part of other studies. The pellets leftno remains in the scats so we can not tell how much ofthe diet they constituted, but there were no differencesin the occurrence of different prey remains betweenfoxes at fed and non-fed dens each year (all P-values
between 0.13 and 0.96). Further, observations in the®eld have indicated that arctic foxes prefer natural foodand continue to hunt even when using `arti®cial' foodresources (Fine, 1980; AngerbjoÈrn, Stroman & Becker,1997; M. Tannerfeldt, pers. obs.).
B. Elmhagen ET AL.144
Table 2. The separate effects of year and sub-area on the occurrence of different prey in the diet of arctic foxes Alopex lagopus ina G-test In A1.2, we have tested the occurrence of juvenile lemmings within the lemming category, not in the diet as a whole.Three-way associations are only included in the table when signi®cant. G-test could not be performed for A1.2-A4, so these weretested with w2 analysis
Effects d.f. w2 P Comment
A1.1 Model: Areas±year±lemmingsAreas±lemmings 4 4.44 0.35 Signi®cant difference in occurrence of lemmings inYear±lemmings 4 30.38 < 0.01 different years
A1.2 Juvenile lemmingsAreas±juveniles 4 1.89 0.76 No differences in the occurrence of juvenile lemmingsYear±juveniles 4 4.59 0.33
A2 Field voleAreas±®eld vole 4 5.41 0.25 No differences in the occurrences of ®eld voleYear±®eld vole 4 1.85 0.76
A3 Bank vole and grey-sided voleAreas±voles 4 4.37 0.36 No differences in the occurrence of bank vole andYear±voles 4 6.85 0.14 grey-sided vole
A4 ShrewsAreas±shrews 4 2.50 0.64 No differences in the occurrence of shrewsYear±shrews 4 1.04 0.90
B Model: Areas±year±birdsAreas±birds 4 40.91 < 0.01 Signi®cant differences in occurrence of birdsYear±birds 4 27.62 < 0.01 between sub-areas and years, but interactionThree-way interaction 16 27.88 0.033 between year and sub-area
C Model: Areas±year±larger mammalsAreas±larger mammals 4 13.52 < 0.01 Signi®cant difference in occurrence of largerYear±larger mammals 4 60.07 < 0.01 mammals between years and sub-areas
Small mammals–lemmings
Small mammals–voles and shrews
Larger mammals
BirdsLemming nests
100
80
60
40
20
0
100
80
60
40
20
0
% W
hole
sca
t equ
ival
ents
No.
of l
emm
ing
nest
s/10
0 km
1990 1994 1995 1996 1997
Yearn = 132,3 dens n = 65, 4 dens n = 78, 4 dens n = 143, 6 dens n = 74, 4 dens
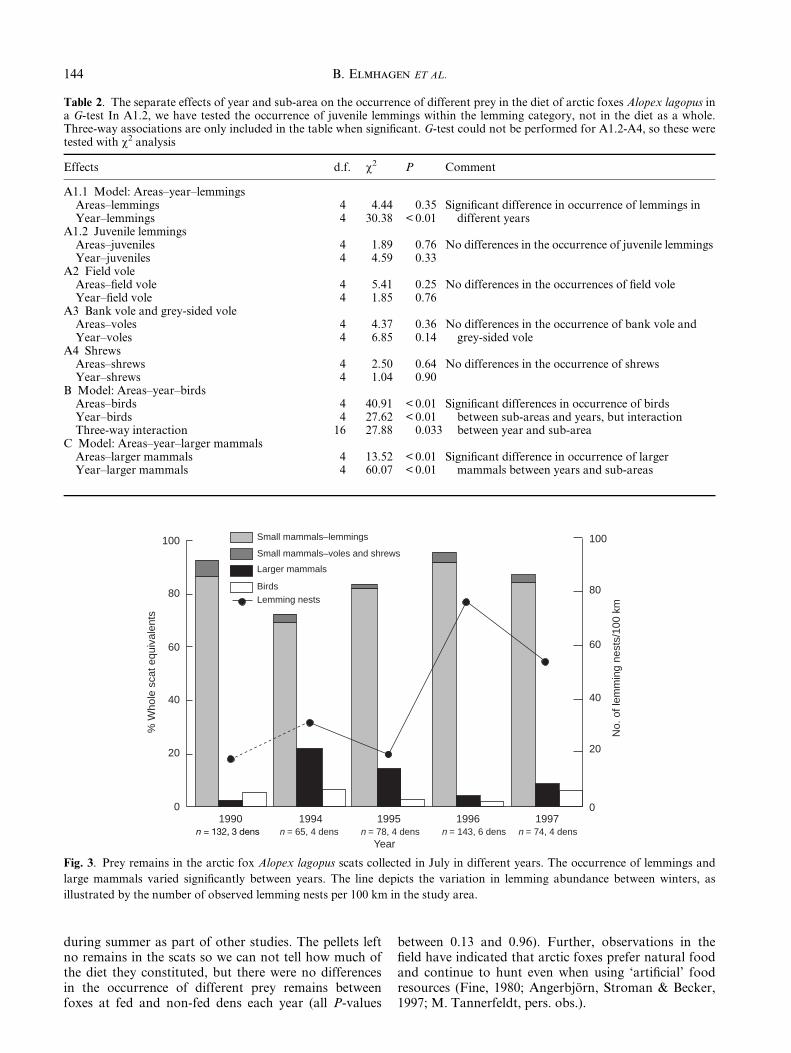
Fig. 3. Prey remains in the arctic fox Alopex lagopus scats collected in July in different years. The occurrence of lemmings and
large mammals varied signi®cantly between years. The line depicts the variation in lemming abundance between winters, as
illustrated by the number of observed lemming nests per 100 km in the study area.
n = 132, 3 dens

Lemming abundance
There were no large lemming population peaks in ourstudy area between 1990 and 1998, but changes in thenumber of lemming winter nests between years indicatedthat there were ¯uctuations in the size of the lemmingpopulation (Figs 3 & 6).
Temporal variation in diet
The occurrence of remains from lemmings, birds andlarger mammals in the scats differed signi®cantlybetween years, when the effect of sub-area was con-trolled for. Lemmings always constituted the larger partof the food remains in the scats. In years when theoccurrence of small mammals decreased, largermammals occurred more often. The same relationship isevident when WSEs are considered. The overall occur-rence of birds in different years varied similarly to thelarger mammals, but there was an interaction betweensub-area and year (Table 2, Fig. 3).
The other small mammal species, voles and shrews,did not vary in occurrence between years (Table 2). Intotal, lemmings constituted 95% (WSEs) of the foodremains from the small mammals. Microtus agrestis(4%) was more common than Clethrionomys spp. (1%)while shrews only made up 0.4% of the category. Mostof the eaten birds were passerines (in total 92% of birdWSEs) while waders and ptarmigan (8%) were rare(Table 1).
Between July and August, there was a signi®cantdifference in the overall occurrence of lemmings when
the effect of year was controlled for. Further, within thelemming category, juvenile lemmings were morecommon in August. There were no signi®cant changesover the summer in the occurrence of the other smallmammals (Table 3). The occurrence of larger mammalsinteracted with both month and year. In 1995 and 1996there were less larger mammals in August, while therewere more in 1997. There was no signi®cant differencein the occurrence of bird different months (Table 3).
Temporal variation in diet at one den: a case study
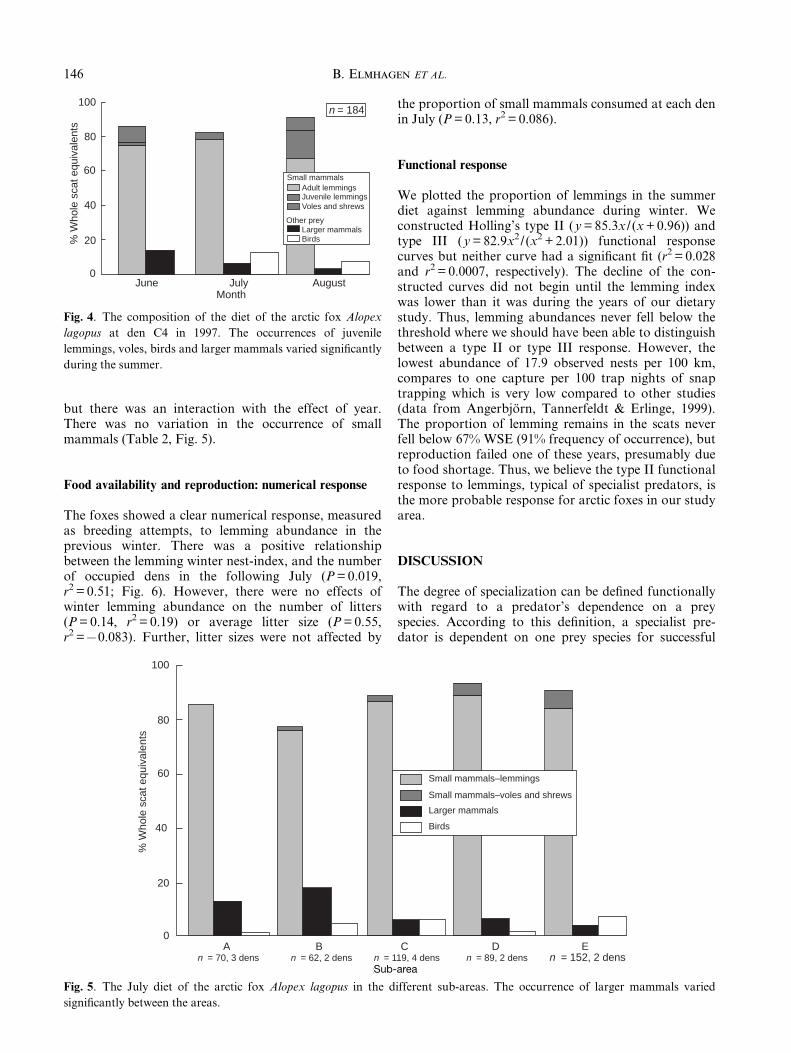
There was no variation in the overall occurrence oflemmings between June and August at den C4 in 1997(d.f. = 2, w2 = 3.19, P = 0.20), but more lemmings werejuveniles towards the end of the summer (d.f. = 2,w2 = 15.52, P < 0.01). Further, the occurrence of voles, atthis den only M. agrestis, was relatively large in June,but small in July (d.f. = 2, w2 = 15.30, P < 0.01). Birdremains peaked in July (d.f. = 2, w2 = 18.15, P < 0.01)while larger mammals decreased during the summer(d.f. = 2, w2 = 12.16, P < 0.01, Fig. 4).
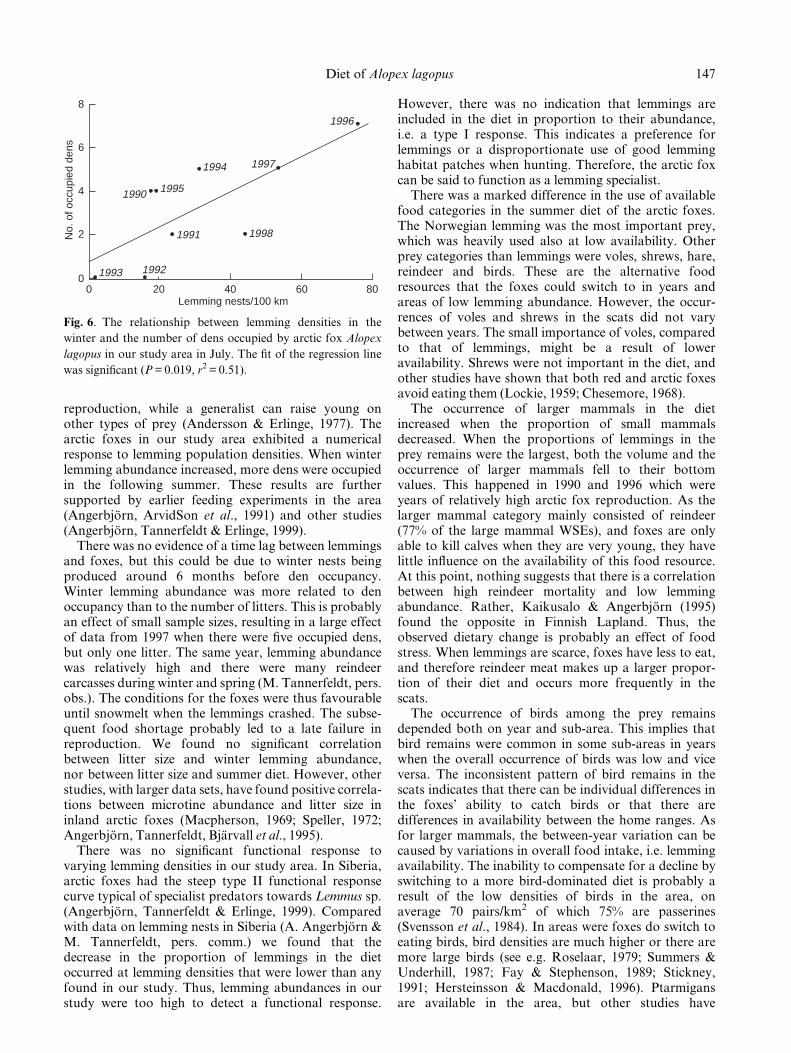
Geographical variation in diet
The diet in July varied on a geographical scale as well asa temporal. The occurrence of larger mammals variedsigni®cantly between sub-areas when controlling for theeffect of year. They were more common in scats fromthe high altitude sub-areas, A and B. Overall, birds weremore common in the scats from sub-areas B, D and E,
145Diet of Alopex lagopus
Table 3. The separate effects of month (July and August) on the diet of arctic foxes Alopex lagopus and the occurrence ofdifferent prey in the diet in a G-test, when the effect of year was controlled for. In A1.2, we have tested the occurrence of juvenilelemmings within the lemming category, not in the diet as a whole. Three-way associations are only included in the table whensigni®cant
Effects d.f. w2 P Comment
A1.1 Model: Year±month±lemmingsMonth±lemmings 1 23.93 < 0.01 Signi®cant difference in occurrence of lemmings
between monthsA1.2 Model: Year±month±juvenile lemmings
Month±juvenile lemmings 1 10.83 < 0.01 Signi®cant difference in the occurrence of juvenilelemmings
A2 Model: Year±month±®eld voleMonth±®eld vole 1 1.48 0.22 No difference in the occurrences of ®eld vole
A3 Model: Year±month±bank vole/grey-sided voleMonth±voles 1 0 0.996 No difference in the occurrence of bank vole and
grey-sided voleA4 Model: Year±month±shrews
Month±shrews 1 0.0005 0.98 No differences in the occurrence of shrewsB Model: Year±month±birds
Month±birds 1 2.07 0.15 No difference in the occurrence of birds betweenThree-way interaction 3 19.28 < 0.01 months. Signi®cant interaction between year,
month and birdC Model: Year±month±larger mammals
Month±larger mammals 1 4.74 0.029 Signi®cant difference in occurrence of largerThree-way interaction 3 12.82 < 0.01 mammals between months. Signi®cant interaction
between year, month and mammals
but there was an interaction with the effect of year.There was no variation in the occurrence of smallmammals (Table 2, Fig. 5).
Food availability and reproduction: numerical response
The foxes showed a clear numerical response, measuredas breeding attempts, to lemming abundance in theprevious winter. There was a positive relationshipbetween the lemming winter nest-index, and the numberof occupied dens in the following July (P = 0.019,r2 = 0.51; Fig. 6). However, there were no effects ofwinter lemming abundance on the number of litters(P = 0.14, r2 = 0.19) or average litter size (P = 0.55,r2 =70.083). Further, litter sizes were not affected by
the proportion of small mammals consumed at each denin July (P = 0.13, r2 = 0.086).
Functional response
We plotted the proportion of lemmings in the summerdiet against lemming abundance during winter. Weconstructed Holling's type II (y = 85.3x / (x + 0.96)) andtype III (y = 82.9x2/ (x2 + 2.01)) functional responsecurves but neither curve had a signi®cant ®t (r2 = 0.028and r2 = 0.0007, respectively). The decline of the con-structed curves did not begin until the lemming indexwas lower than it was during the years of our dietarystudy. Thus, lemming abundances never fell below thethreshold where we should have been able to distinguishbetween a type II or type III response. However, thelowest abundance of 17.9 observed nests per 100 km,compares to one capture per 100 trap nights of snaptrapping which is very low compared to other studies(data from AngerbjoÈrn, Tannerfeldt & Erlinge, 1999).The proportion of lemming remains in the scats neverfell below 67% WSE (91% frequency of occurrence), butreproduction failed one of these years, presumably dueto food shortage. Thus, we believe the type II functionalresponse to lemmings, typical of specialist predators, isthe more probable response for arctic foxes in our studyarea.
DISCUSSION
The degree of specialization can be de®ned functionallywith regard to a predator's dependence on a preyspecies. According to this de®nition, a specialist pre-dator is dependent on one prey species for successful
B. Elmhagen ET AL.146
Small mammalsAdult lemmingsJuvenile lemmingsVoles and shrews
Other preyLarger mammalsBirds
n = 184100
80
60
40
20
0
% W
hole
sca
t equ
ival
ents
June July AugustMonth
Fig. 4. The composition of the diet of the arctic fox Alopex
lagopus at den C4 in 1997. The occurrences of juvenile
lemmings, voles, birds and larger mammals varied signi®cantly
during the summer.
Small mammals–lemmings
Small mammals–voles and shrews
Larger mammals
Birds
100
80
60
40
20
0
% W
hole
sca
t equ
ival
ents
A B C D En = 70, 3 dens n = 62, 2 dens n = 119, 4 dens n = 89, 2 dens n = 152, 2 dens
Subarea
Fig. 5. The July diet of the arctic fox Alopex lagopus in the different sub-areas. The occurrence of larger mammals varied
signi®cantly between the areas.
Sub-area
reproduction, while a generalist can raise young onother types of prey (Andersson & Erlinge, 1977). Thearctic foxes in our study area exhibited a numericalresponse to lemming population densities. When winterlemming abundance increased, more dens were occupiedin the following summer. These results are furthersupported by earlier feeding experiments in the area(AngerbjoÈrn, ArvidSon et al., 1991) and other studies(AngerbjoÈrn, Tannerfeldt & Erlinge, 1999).
There was no evidence of a time lag between lemmingsand foxes, but this could be due to winter nests beingproduced around 6 months before den occupancy.Winter lemming abundance was more related to denoccupancy than to the number of litters. This is probablyan effect of small sample sizes, resulting in a large effectof data from 1997 when there were ®ve occupied dens,but only one litter. The same year, lemming abundancewas relatively high and there were many reindeercarcasses during winter and spring (M. Tannerfeldt, pers.obs.). The conditions for the foxes were thus favourableuntil snowmelt when the lemmings crashed. The subse-quent food shortage probably led to a late failure inreproduction. We found no signi®cant correlationbetween litter size and winter lemming abundance,nor between litter size and summer diet. However, otherstudies, with larger data sets, have found positive correla-tions between microtine abundance and litter size ininland arctic foxes (Macpherson, 1969; Speller, 1972;AngerbjoÈrn, Tannerfeldt, BjaÈrvall et al., 1995).
There was no signi®cant functional response tovarying lemming densities in our study area. In Siberia,arctic foxes had the steep type II functional responsecurve typical of specialist predators towards Lemmus sp.(AngerbjoÈrn, Tannerfeldt & Erlinge, 1999). Comparedwith data on lemming nests in Siberia (A. AngerbjoÈrn &M. Tannerfeldt, pers. comm.) we found that thedecrease in the proportion of lemmings in the dietoccurred at lemming densities that were lower than anyfound in our study. Thus, lemming abundances in ourstudy were too high to detect a functional response.
However, there was no indication that lemmings areincluded in the diet in proportion to their abundance,i.e. a type I response. This indicates a preference forlemmings or a disproportionate use of good lemminghabitat patches when hunting. Therefore, the arctic foxcan be said to function as a lemming specialist.
There was a marked difference in the use of availablefood categories in the summer diet of the arctic foxes.The Norwegian lemming was the most important prey,which was heavily used also at low availability. Otherprey categories than lemmings were voles, shrews, hare,reindeer and birds. These are the alternative foodresources that the foxes could switch to in years andareas of low lemming abundance. However, the occur-rences of voles and shrews in the scats did not varybetween years. The small importance of voles, comparedto that of lemmings, might be a result of loweravailability. Shrews were not important in the diet, andother studies have shown that both red and arctic foxesavoid eating them (Lockie, 1959; Chesemore, 1968).
The occurrence of larger mammals in the dietincreased when the proportion of small mammalsdecreased. When the proportions of lemmings in theprey remains were the largest, both the volume and theoccurrence of larger mammals fell to their bottomvalues. This happened in 1990 and 1996 which wereyears of relatively high arctic fox reproduction. As thelarger mammal category mainly consisted of reindeer(77% of the large mammal WSEs), and foxes are onlyable to kill calves when they are very young, they havelittle in¯uence on the availability of this food resource.At this point, nothing suggests that there is a correlationbetween high reindeer mortality and low lemmingabundance. Rather, Kaikusalo & AngerbjoÈrn (1995)found the opposite in Finnish Lapland. Thus, theobserved dietary change is probably an effect of foodstress. When lemmings are scarce, foxes have less to eat,and therefore reindeer meat makes up a larger propor-tion of their diet and occurs more frequently in thescats.
The occurrence of birds among the prey remainsdepended both on year and sub-area. This implies thatbird remains were common in some sub-areas in yearswhen the overall occurrence of birds was low and viceversa. The inconsistent pattern of bird remains in thescats indicates that there can be individual differences inthe foxes' ability to catch birds or that there aredifferences in availability between the home ranges. Asfor larger mammals, the between-year variation can becaused by variations in overall food intake, i.e. lemmingavailability. The inability to compensate for a decline byswitching to a more bird-dominated diet is probably aresult of the low densities of birds in the area, onaverage 70 pairs/km2 of which 75% are passerines(Svensson et al., 1984). In areas were foxes do switch toeating birds, bird densities are much higher or there aremore large birds (see e.g. Roselaar, 1979; Summers &Underhill, 1987; Fay & Stephenson, 1989; Stickney,1991; Hersteinsson & Macdonald, 1996). Ptarmigansare available in the area, but other studies have
147Diet of Alopex lagopus
8
6
4
2
0
No.
of o
ccup
ied
dens
0 20 40 60 80Lemming nests/100 km
1994
19951990
1997
1991 1998
1993 1992
1996
Fig. 6. The relationship between lemming densities in the
winter and the number of dens occupied by arctic fox Alopex
lagopus in our study area in July. The ®t of the regression line
was signi®cant (P = 0.019, r2 = 0.51).

indicated that in the summer they are dif®cult for arcticfoxes to catch (Larson, 1960; Birks & Penford, 1990).
The arctic foxes in our study area focused theirfeeding on lemmings. At the same time, they fed oppor-tunistically on other prey species, as shown by thetemporal and geographical variations in their diet. In1995, 1996 and at den C4 in 1997, the occurrence oflarge mammals decreased towards the end of thesummer. As reindeer mortality above the tree line ishigher during spring and winter (pers. obs.), this couldre¯ect a decrease in availability as most meat on oldcarcasses is gone by August. On the other hand, thefoxes at den C5 in July 1994 relied on reindeer to a largeextent. They had probably found a relatively freshcarcass to feed from. Voles were mostly used to a lowextent, but the foxes in den C4 1997 consumed ®eldvoles quite frequently in June and August. These foxesalso increased their consumption of birds in July com-pared to June. As hatching begins around this time, thisis probably a response to an increase in availability. Theoverall large percentage of passerines among theconsumed birds, in agreement with the actual composi-tion of the bird community in the area, goes further toshow the opportunistic feeding of these foxes. Theincrease in consumption of juvenile lemmings in Augustwas expected, since the proportion of juveniles in alemming population is higher in August than in July(Kalela et al., 1961).
For a prey switch to be pro®table, there must bealternative prey that become more abundant than thepreferred prey (Murdoch, 1969). In Sweden, this condi-tion is probably not ful®lled for arctic foxes, as the preybase above the tree line is very limited. When alternativeprey species are more common, as in many coastalareas, arctic foxes function as generalists. This isdifferent from, for example, wolves and kit foxes, thatdo not catch abundant, alternative prey when thedensities of their preferred prey decline (Dale, Adams &Bowyer, 1994; White, Vanderbilt White & Ralls, 1996).Arctic foxes in VindelfjaÈllen catch prey other thanlemmings when they can, but during the years of ourstudy they were not able to compensate for lowlemming abundances and breed successfully. Thus, theydo not qualify for the generalist category, but arecon®ned to a specialist strategy. The minimum averagelifespan of arctic foxes who have reached reproductiveage in our study area is 30 months (Tannerfeldt &AngerbjoÈrn, 1996.) When lemmings are cyclic, foxesborn during a lemming peak can expect around36 months of food scarcity before the next peak. Thesepredators must therefore be ready to exploit what littlediversity of food resources there is on the tundra, if theyare to survive the long years of low food abundance andbe ready to reproduce when the lemming populationincreases again. The arctic fox thus manages to combinean opportunistic feeding strategy which ensures survivalin a low productivity habitat, with a remarkable abilityto exploit lemming peaks for a large reproductiveoutput. In effect, it is an opportunistic feeder in a worldgoverned by lemmings.
Acknowledgements
The Arctic Fox Project has been ®nancially supportedby WWF Sweden, foundations at the Royal Academyof Sciences and Oscar och Lili Lamms stiftelse. ABDogman and FjaÈllraÈven AB also helped us with theirproducts. The Swedish Institute contributed to PaoloVerucci's stay in Sweden. Many thanks to all enthu-siastic, scat-collecting ®eld workers who have made theextensive den surveys possible over the years.
REFERENCES
Andersson, M. & Erlinge, S. (1977). In¯uence of predation onrodent populations. Oikos 29: 591±597.
AngerbjoÈrn, A., ArvidSon, B., NoreÂn, E. & StroÈmgren, L.(1991). The effect of winter food on reproduction in the arcticfox, Alopex lagopus: a ®eld experiment. J. Anim. Ecol. 60:705±714.
AngerbjoÈrn, A., Hersteinsson, P., LideÂn, K. & Nelson, E. (1994).Dietary variation in arctic foxes (Alopex lagopus) ± an analysisof stable carbon isotopes. Oecologia (Berl.) 99: 226±232.
AngerbjoÈrn, A., Stroman, J. & Becker, D. (1997). Home rangepatterns in arctic foxes in Sweden. J. Wildl. Res. 2: 9±14.
AngerbjoÈrn, A., Tannerfeldt, M., BjaÈrvall, A., Ericson, M., From,J. & NoreÂn, E. (1995). Dynamics of the arctic fox population inSweden. Ann. Zool. Fenn. 32: 55±68.
AngerbjoÈrn A., Tannerfeldt, M. & Erlinge, S. (1999). Predator±prey relations: arctic foxes and lemmings. J. Anim. Ecol. 68:34±49.
Birks, J. D. S. & Penford, N. (1990). Observations on the ecologyof arctic foxes Alopex lagopus in Eqalummiut Nunaat, WestGreenland. Medd. Grùnl. Biosci. 32: 1±26.
Boutin, S. (1995). Testing predator±prey theory by studying¯uctuating populations of small mammals. Wildl. Res. 22:89±100.
Braestrup, F. W. (1941). A study of the arctic fox in Greenland.Medd. Grùnl. Biosci. 131: 1±101.
Brom, T. G. (1986). Microscopic identi®cation of feathers andfeather fragments of palearctic birds. Bijdr. Dierkd. 56:181±204.
Bronson, F. H. (1989). Mammalian reproductive biology. Chicago:Chicago University Press.
Chesemore, D. L. (1968). Notes on the food habits of arctic foxesin northern Alaska. Can. J. Zool. 46: 1227±1130.
Dale, B. W., Adams, L. G. & Bowyer, R. T. (1994). Functionalresponse of wolves preying on barren-ground caribou in amultiple-prey ecosystem. J. Anim. Ecol. 63: 644±652.
Dalerum, F. & AngerbjoÈrn, A. (2000). Arctic fox (Alopex lagopus)diet in Kampelv valley, East Greenland, during a summer withlow lemming density. Arctic 53: 1±8.
Day, M. G. (1996). Identi®cation of hair and feather remains inthe gut and faeces of stoats and weasels. J. Zool. (Lond.) 148:201±217.
Erlinge, S., GoÈransson, G., HoÈgstedt, G., Jansson, G., Liberg, O.,Loman, J., Nilsson, I. N., von Schantz, T. & SylveÂn, M. (1984).Can vertebrate predators regulate their prey? Am. Nat. 123:125±133.
Fay, F. H. & Stephenson, R. O. (1989). Annual, seasonal andhabitat±related variation in feeding habits of the arctic fox(Alopex lagopus) on St. Lawrence Island, Bering Sea. Can. J.Zool. 67: 1986±1994.
Fine, H. (1980). Ecology of arctic foxes at Prudhoe Bay, Alaska.MSc thesis, University of Fairbanks, Alaska.
Finerty, J. P. (1980). The population ecology of cycles in smallmammals. New Haven, CT: Yale University Press.
B. Elmhagen ET AL.148

Frafjord, K. (1993a). Reproductive effort in the arctic fox Alopexlagopus: a review. Norw. J. agric. Sci. 7: 301±309.
Frafjord, K. (1993b). Food habits of arctic foxes (Alopex lagopus)on the western coast of Svalbard. Arctic 46: 49±54.
Garrott, R. A., Eberhardt, L. E. & Hanson, W. C. (1983).Summer food habits of juvenile arctic foxes in northern Alaska.J. Wildl. Manage. 47: 540±545.
Hanski, I., Hansson, L. & Henttonen, H. (1991). Specialistpredators, generalist predators, and the microtine cycle.J. Anim. Ecol. 60: 353±367.
Hanski, I. & KorpimaÈki, E. (1995). Microtine rodent dynamics innorthern Europe: parameterised models for the predator±preyinteraction. Ecology 76: 840±850.
Hanski, I., Turchin, P., KorpimaÈki, E. & Henttonen, H. (1993).Population oscillations of boreal rodents: Regulation by mus-telid predators leads to chaos. Nature 364: 232±235.
Henttonen, H., Oksanen, T., Jortikka, A. & Haukisalmi, V.(1987). How much do weasels shape microtine cycles in thenorthern Fennoscandian taiga? Oikos 50: 353±365.
Hersteinsson, P. (1984). The behavioural ecology of the arctic fox(Alopex lagopus) in Iceland. DPhil. thesis, University of Oxford.
Hersteinsson, P., AngerbjoÈrn, A., Frafjord, K. & Kaikusalo, A.(1989). The arctic fox in Fennoscandia and Iceland: manage-ment problems. Biol. Conserv. 49: 67±81.
Hersteinsson, P. & Macdonald, D. W. (1996). Diet of arctic foxes(Alopex lagopus) in Iceland. J. Zool. (Lond.) 240: 457±474.
Hiruki, L. M. & Stirling, I. (1989). Population dynamics of thearctic fox, Alopex lagopus, on Banks Island, NorthwestTerritories. Can. Field-Nat. 103: 380±387.
Holling, C. S. (1959). The components of predation as revealed bya study of small- mammal predation on the European pinesaw¯y. Can. Entomol. 91: 293±320.
Kaikusalo, A. & AngerbjoÈrn, A. (1995). The arctic fox populationin Finnish Lapland during 30 years, 1964±93. Ann. Zool. Fenn.32: 69±77.
Kalela, O., Korponen, T., Lind, E. A., SkareÂn, U. & Tast, J.(1961). Seasonal change of habitat in the Norwegian lemming,Lemmus lemmus (L.). Ann. Acad. Sci.. Fenn. Ser. A 55: 5±75.
KorpimaÈki, E., Norrdahl, K. & Rinta-Jaskari, T. (1991).Responses of stoats and least weasels to ¯uctuating foodabundances: Is the low phase of the vole cycle due to mustelidpredation? Oecologia (Berl.) 88: 552±561.
Larson, S. (1960). On the in¯uence of the arctic fox, Alopexlagopus, on the distribution of arctic birds. Oikos 11: 276±305.
Lockie, J. D. (1959). The estimation on the food of foxes. J. Wildl.Manage. 23: 224±227.
Lundberg, H. (1999). Lemming cyclicity in Fennoscandia!Undergraduate thesis, Department of Zoology, StockholmUniversity.
Macpherson, A. H. (1969). The dynamics of Canadian arctic foxpopulations. Can. Wildl. Serv. Rep. Ser. 8: 1±49.
Murdoch, W. W. (1969). Switching in general predators: experi-ments on predator speci®city and stability of prey populations.Ecol. Monogr. 39: 335±354.
Niethammer, J. & Krapp, F. (1982). Handbuch der SaÈugetiereEuropas. 2/1 Nagetiere II. Wiesbaden: Akademische Verlags-gesellschaft.
Pearson, O. P. (1966). The prey of carnivores during one cycle ofmouse abundance. J. Anim. Ecol. 35: 217±233.
Pond, C. M., Mattacks, C. A., Gilmour, I., Johnston, M. A. &Pillinger, C. T. (1995). Chemical and carbon isotope composi-tion of fatty acids in adipose tissue as indicators of dietaryhistory in wild arctic foxes (Alopex lagopus) on Svalbard.J. Zool. (Lond.) 236: 611±623.
Prestrud, P. (1992). Food habits and observations of the huntingbehaviour of arctic foxes, Alopex lagopus, in Svalbard. Can.Field-Nat. 106: 225±236.
Reid, D. G., Krebs, C. J. & Kenney, A. J. (1997). Patterns ofpredation on noncyclic lemmings. Ecol. Monogr. 67: 89±108.
Reynolds, J. C. & Aebischer, N. J. (1991). Comparison andquanti®cation of carnivore diet by faecal analysis: a critique,with recommendations, based on a study of the fox Vulpesvulpes. Mammal Rev. 21: 97±122.
Roselaar, C. S. (1979). Fluctuaties i aantallen Krombekstrandlo-pers Calidris ferruginea. Watervogels 4: 202±210.
Rune, O. (1981). Floran inom VindelfjaÈllens naturreservat. Medde-lande 3. LaÈnsstyrelsen, VaÈsterbottens laÈn.
Sittler, B. (1995). Response of stoats (Mustela erminea) to a¯uctuating lemming (Dicrostonyx groenlandicus) population inNorth East Greenland: Preliminary results from a long-termstudy. Ann. Zool. Fenn. 32: 79±92.
Speller, S. W. (1972). Food ecology and hunting behavior of denningarctic foxes at Aberdeen Lake, Northwest Territories. DPhil.thesis, University of Saskatchewan.
Stenseth, N. C., Bjùrnstad, O. N. & Falck, W. C. S. (1996). Isspacing behaviour coupled with predation causing the micro-tine density cycle? A synthesis of current process-oriented andpattern-oriented studies. Proc. R. Soc. London Ser. B Biol. Sci.263: 1423±1435.
Stickney, A. (1991). Seasonal patterns of prey availability and theforaging behaviour of arctic foxes (Alopex lagopus) in a water-fowl nesting area. Can. J. Zool. 69: 2853±2859.
Summers, R. W. & Underhill, L. G. (1987). Factors related tobreeding production of Brent geese Branta b. bernicla andwaders (Charadrii) on the Taimyr peninsula. Bird Study 34:161±171.
Svensson, S., Carlsson, U. T. & Liljedahl, G. (1984). Structureand dynamics of an alpine bird community, a 20-year study.Ann. Zool. Fenn. 21: 339±350.
Tannerfeldt, M. (1997). Population ¯uctuations and life historyconsequences in the arctic fox. DPhil. thesis, University ofStockholm.
Tannerfeldt, M. & AngerbjoÈrn, A. (1996). Life history strategiesin a ¯uctuating environment: establishment and reproductivesuccess in the arctic fox. Ecography 19: 209±220.
Tannerfeldt, M. & AngerbjoÈrn, A. (1998). Fluctuating resourcesand the evolution of litter size in the arctic fox. Oikos 83:545±559.
Tannerfeldt, M., AngerbjoÈrn, A. & ArvidSon, B. (1994). Theeffect of summer feeding on juvenile arctic fox survival ± a ®eldexperiment. Ecography 17: 88±96.
Taylor, R. J. (1984). Predation. New York: Chapman & Hall.White, P. J., Vanderbilt White, C. A. & Ralls, K. (1996).
Functional and numerical responses of kit foxes to a short-termdecline in mammalian prey. Southwest. Nat. 40: 342±349.
Wiklund, C. G., AngerbjoÈrn, A., Isakson, E., KjelleÂn, N. &Tannerfeldt, M. (1999). Lemming predators on the Siberiantundra. Ambio 3: 281±286.
149Diet of Alopex lagopus