the adaptor protein 14-3-3 binds to the calcium-sensing receptor and attenuates receptor-mediated...
TRANSCRIPT

Biochem. J. (2012) 441, 995–1006 (Printed in Great Britain) doi:10.1042/BJ20111277 995
The adaptor protein 14-3-3 binds to the calcium-sensing receptor andattenuates receptor-mediated Rho kinase signallingAjanthy ARULPRAGASAM*†‡, Aaron L. MAGNO*†‡, Evan INGLEY*†, Suzanne J. BROWN‡, Arthur D. CONIGRAVE§,Thomas RATAJCZAK*†‡ and Bryan K. WARD*†‡1
*Western Australian Institute for Medical Research, QEII Medical Centre, Hospital Avenue, Nedlands, WA 6009, Australia, †University of Western Australia Centre for Medical Research,University of Western Australia, Nedlands, WA 6009, Australia, ‡Department of Endocrinology and Diabetes, Sir Charles Gairdner Hospital, Hospital Avenue, Nedlands, WA 6009,Australia, and §School of Molecular Bioscience, University of Sydney, Sydney, NSW 2000, Australia
A yeast two-hybrid screen performed to identify binding partnersof the CaR (calcium-sensing receptor) intracellular tail identifiedthe adaptor protein 14-3-3θ as a novel binding partner thatbound to the proximal membrane region important for CaRexpression and signalling. The 14-3-3θ protein directly interactedwith the CaR tail in pull-down studies and FLAG-tagged CaRco-immunoprecipitated with EGFP (enhanced green fluorescentprotein)-tagged 14-3-3θ when co-expressed in HEK (humanembryonic kidney)-293 or COS-1 cells. The interaction betweenthe CaR and 14-3-3θ did not require a putative bindingsite in the membrane-proximal region of the CaR tail andwas independent of PKC (protein kinase C) phosphorylation.Confocal microscopy demonstrated co-localization of the CaRand EGFP–14-3-3θ in the ER (endoplasmic reticulum) ofHEK-293 cells that stably expressed the CaR (HEK-293/CaRcells), but 14-3-3θ overexpression had no effect on membrane
expression of the CaR. Overexpression of 14-3-3θ in HEK-293/CaR cells attenuated CaR-mediated Rho signalling, buthad no effect on ERK (extracellular-signal-regulated kinase)1/2 signalling. Another isoform identified from the library,14-3-3ζ , exhibited similar behaviour to that of 14-3-3θ withrespect to CaR tail binding, cellular co-localization and impacton receptor-mediated signalling. However, unlike 14-3-3θ , thisisoform, when overexpressed, significantly reduced CaR plasmamembrane expression. Results indicate that 14-3-3 proteinsmediate CaR-dependent Rho signalling and may modulate theplasma membrane expression of the CaR.
Key words: calcium-sensing receptor (CaR), cellular co-localization, 14-3-3 protein, protein interaction, receptorexpression, Rho signalling.
INTRODUCTION
The CaR (calcium-sensing receptor) is a class C GPCR(G-protein-coupled receptor) that is highly expressed in theparathyroid gland and renal tubules where it regulates parathyroidhormone secretion and calcium excretion respectively [1,2]. Thehuman CaR protein (1078 amino acids) is composed of a largeextracellular N-terminal ligand-binding domain (610 residues),a signature seven-transmembrane domain and an intracellularC-terminal tail (215 residues) [3]. In addition to Ca2 + , agonistsof the CaR include other di- and tri-valent cations as well aspolypeptides, polyamines and aminoglycoside antibiotics [1,2,4].The CaR is also modulated by ionic strength and pH as well asamino acids, γ -glutamyl peptides and various classes of syntheticcalcimimetics and calcilytics [4–6]. Upon ligand binding, thereceptor couples to three distinct classes of heterotrimeric G-proteins, Gq/11, Gi/o and G12/13, in a tissue- and/or ligand-specificmanner. This leads to activation of phospholipase C and releaseof Ca2 + ions from intracellular stores, activation of proteinkinases including PKC (protein kinase C), Akt and the MAPKs(mitogen-activated protein kinases) p38, ERK (extracellular-signal-regulated kinase) 1/2 and JNK (c-Jun N-terminal kinase),as well as inhibition of adenylate cyclase [2,4,7]. The CaR alsostimulates SRE (serum-response element) activity downstream of
Gq and the monomeric G-protein Rho [8], and through G12/13 thereceptor can modulate Rho-dependent phospholipase D activationand L-amino acid-induced Ca2 + mobilization [9,10] (for reviewsof CaR-mediated signalling, see [2,4,7]). Downstream of itssignalling pathways, the CaR regulates critical cellular processesincluding secretion, apoptosis, chemotactic responses, cellproliferation, cytoskeletal rearrangements, ion channel activity,the control of gene expression and cell differentiation [2,4].
The CaR intracellular tail comprises amino acids 865–1078 andcontains important determinants for cell-surface expressionand activation of signalling pathways [1,2]. The membrane-proximal region of the tail is essential for biological activitywith critical residues identified for expression between residues865 and 877, and those for signalling between residues 877 and888 [1,11,12]. The remaining tail is dispensable, as exemplifiedby the impact of a large in-frame deletion (residues 895–1075)which exhibits gain-of-function compared with the wild-type CaR[13]. There are three predicted PKC phosphorylation sites inthe CaR tail, Thr888, Ser895 and Ser915, and phosphorylation atThr888 uncouples the receptor from phosphatidylinositol-specificphospholipase C and intracellular Ca2 + release [3,14]. The CaRtail also houses the ER (endoplasmic reticulum) retention signalsRR and RKR located at amino acids 890 and 891, and 896–898 respectively [15,16]. In addition to G-protein coupling,
Abbreviations used: CaR, calcium-sensing receptor; COPI, coat protein I; DMEM, Dulbecco’s modified Eagle’s medium; EGFP, enhanced greenfluorescent protein; ER, endoplasmic reticulum; ERK, extracellular-signal-regulated kinase; FBS, fetal bovine serum; GEF, guanine-nucleotide-exchangefactor; GFP, green fluorescent protein; GPCR, G-protein-coupled receptor; GST, glutathione transferase; HEK, human embryonic kidney; HRP, horseradishperoxidase; Hsp, heat-shock protein; Luc, luciferase; MAPK, mitogen-activated protein kinase; NTA, nitrilotriacaetic acid; PDI, protein disulfide isomerase;PKA, protein kinase A; PKC, protein kinase C; PSS, physiological saline solution; RT, reverse transcription; siRNA, small interfering RNA; SRE, serum-response element; TMB, 3,3′,5,5′-tetramethylbenzidine.
1 To whom correspondence should be addressed (email [email protected]).
c© The Authors Journal compilation c© 2012 Biochemical Society
www.biochemj.org
Bio
chem
ical
Jo
urn
al

996 A. Arulpragasam and others
CaR-dependent signalling also requires interactions with bindingpartners including filamin [17,18], a 250 kDa actin-bindingprotein that anchors cell-surface receptors to the cytoskeleton[19]. Filamin is essential for CaR-mediated activation of theERK1/2 pathway [17,18] and for optimal CaR-mediated SREactivity via the Rho pathway, providing a scaffold for theGαq-dependent activation of RhoA via Rho-nucleotide-exchangefactor [8].
In addition to filamin and PKC, a number of other proteinsbind to the intracellular tail of the CaR, including the inwardlyrectifying K+ channels Kir4.1 and Kir4.2 [20], the cargo receptorp24A [21], and proteins of the ubiquitin degradation system suchas the E3 ubiquitin ligase dorfin [22] and deubiquitinating enzymeAMSH [associated molecule with the SH3 (Src homology 3)domain of STAM (signal-transducing adaptor molecule)] [23].With a view to identifying novel binding partners of the CaRtail, we embarked on a yeast two-hybrid screen using the full-length CaR tail as bait with a cDNA library derived from ahaemopoietic cell line (EMLC.1) [24], which has been usedsuccessfully in other screens [25,26]. We identified the 14-3-3adaptor protein as a binding partner of the CaR tail. The 14-3-3 proteins are ubiquitously expressed chaperones that play akey role in facilitating diverse biological processes that includethe regulation of cell signalling events and cellular trafficking[27,28]. In the present paper we show that two isoforms of 14-3-3bind to the membrane proximal region of the CaR and negativelyregulate CaR-dependent SRE activation.
MATERIALS AND METHODS
Antibodies and chemicals
The antibodies used in the present study were from the followingsources: anti-GFP (green fluorescent protein) (rabbit polyclonal),anti-(14-3-3θ ) (C-17, rabbit polyclonal), anti-(14-3-3ζ ) (C-16,rabbit polyclonal) and anti-(filamin 1) (mouse monoclonal) werefrom Santa Cruz Biotechnology; anti-FLAG (mouse monoclonal),anti-(α-tubulin) (mouse monoclonal) and HRP (horseradishperoxidise)-conjugated goat anti-mouse antibody were fromSigma–Aldrich; the anti-CaR ADD antibody (mouse monoclonal)was from Affinity Bioreagents; anti-PDI (protein disulphideisomerase) (rabbit polyclonal) was from Stressgen Bioreagents;anti-ERK1/2 (rabbit polyclonal) and HRP-conjugated goat anti-rabbit antibody were from Promega; anti-(phospho-ERK1/2)(rabbit polyclonal) was from Cell Signaling Technology andAlexa Fluor® 546 goat anti-mouse and Alexa Fluor 647 goat anti-rabbit antibodies were from Molecular Probes, Invitrogen LifeTechnologies. Chemicals were of laboratory grade and purchasedfrom Sigma–Aldrich unless otherwise specified.
Plasmids and yeast two-hybrid library
A pVP16 cDNA library of an EMLC.1 mouse pluripotenthaemopoietic cell line [24], kindly donated by Dr ShickwannTsai (Fred Hutchinson Cancer Research Center, Seattle, U.S.A.)was further amplified for use in the present study. Constructionof the library using NotI-linkered inserts and the LexA-basedscreening procedure in which reconstitution of a transcriptionaltransactivator is able to activate His and LacZ reporter genes, havebeen described previously [24,25,29]. For selection purposes, thepVP16 library vector carries the Leu gene.
The bait construct used for the library screen (pBTM116-CaR[865–1078]) was generated by PCR amplification of thehuman CaR intracellular tail (amino acids 865–1078) from atemplate described previously [30], and insertion of the CaR tail
into the unique SalI site of the bait vector pBTM116 to createa Lex-A DNA-binding fusion. This vector also carries the Trp1gene. Bait deletion constructs for use in yeast two-hybrid mappingstudies, pBTM116-CaR[865–922], pBTM116-CaR[865–898],pBTM116-CaR[899–922] and pBTM116-CaR[923–1078], weresimilarly generated.
For other studies, the full-length human 14-3-3θ proteinwas recovered from the osteosarcoma cell line Saos-2 by RT(reverse transcription)–PCR, verified by sequence analysis, thencloned via the EcoRV and NotI restriction enzyme sites into thevector pcDNA3-EGFP (enhanced GFP) version 1 (supplied byProfessor Karin Eidne, Western Australian Institute for MedicalResearch, Nedlands, Australia) for expression as an N-terminallytagged EGFP fusion protein (pcDNA3-EGFP–14-3-3θ ) and intothe vector pGEX-4T-1 (GE Healthcare Life Sciences)using the SalI and NotI sites for expression as a GST (glutathionetransferase) fusion protein (pGEX-4T-1–14-3-3θ ). Untagged 14-3-3θ (pcDNA3.1–14-3-3θ ) was created by subcloning 14-3-3θinto pcDNA3.1 using the EcoRI and NotI sites. In addition, atruncated version of another 14-3-3 isoform, 14-3-3ζ , was foundto bind to the CaR in the yeast two-hybrid screen. Full-lengthhuman 14-3-3ζ was amplified from Saos-2 cells by RT–PCRand cloned into the pVP16 yeast two hybrid vector and into themammalian expression vectors pcDNA3–EGFP and pcDNA3.1essentially as described for the 14-3-3θ isoform.
The C-terminally FLAG-tagged CaR construct (pcDNA3.1-CaR–FLAG) has been described previously [30]. For cell-surface-expression assays, an extracellular domain, FLAG-tagged CaRwith the FLAG tag inserted between amino acids 371 and 372,was generated and cloned into the pcDNA3.1 vector at the KpnIand XbaI sites (pcDNA3.1-exFLAG–CaR). In order to expressHis-tagged CaR tail, the human CaR intracellular tail (aminoacids 865–1078) was cloned into the NdeI and SalI sites of thepET-28a expression vector by PCR amplification of the CaR tailfrom a pET15b-CaR tail construct using primers with engineeredNdeI and SalI sites. The SRE–Luc (luciferase) reporter plasmid,pSRE–Luc, was donated by Professor Jeffrey E. Pessin (AlbertEinstein College of Medicine, New York, U.SA.).
Yeast two-hybrid library screening, verification of positiveinteractions and deletion mapping studies
The library screen was performed using the lithium acetatemethod by sequential transformation of library plasmid DNAinto the L40 yeast strain pretransformed with the CaR tailbait, essentially as described previously [29]. Following librarytransformation, cells were amplified for 6 h, plated on to selectionmedium [84.6 mM succinic acid, 150 mM sodium hydroxide,543 nM adenine hemisulfate (pH 5.8) with agar, 2 % glucoseand 1× yeast nitrogen base/(NH4)2SO4] deficient in tryptophan,leucine, histidine, lysine and uracil [29], and examined for theappearance of cotransformants exhibiting CaR-tail-dependenttransactivation of the His reporter gene. Potential interactingclones were confirmed by examining LacZ reporter gene activityusing a β-galactosidase colony lift assay, comparing knownpositive and negative cotransformant interactions: pBTM116-CyP40[185–370] + pVP16-Hsp90[520–724] (where Hsp is heat-shock protein) and pBTM116-ARL-E1 + pVP16-Hsp90[520–724] respectively [25,29,31]. Library insert DNA from positivecolonies was amplified by PCR and categorized by size andrestriction enzyme profiling using the HaeIII site. PlasmidDNA from unique clones was ‘rescued’ by transformation intoEscherichia coli HB101 and plating on to M9 plates deficientin leucine [25,29,31]. For final verification of interaction, baitand rescued library plasmid DNA were cotransformed into
c© The Authors Journal compilation c© 2012 Biochemical Society

14-3-3 modulates calcium-sensing receptor signalling 997
yeast, plated on to selection medium and examined for Hisand LacZ reporter gene activation. Verified library inserts weresequenced and basic local alignment searches performed toestablish sequence identity. For yeast two-hybrid mapping studies,plasmid DNA of deletion constructs of the CaR tail and full-length 14-3-3 isoforms were cotransformed into yeast, plated onto selection medium, and co-transformants were examined forLacZ reporter gene activity. Times to colour development werecompared with positive and negative controls to determine therelative strength of reporter activation between each CaR taildeletion and 14-3-3.
Cells, cell culture and transfections
HEK (human embryonic kidney)-293 cells were obtained fromProfessor Karin Eidne (Western Australian Institute for MedicalResearch, Nedlands, Australia) and HEK-293 cells stablyexpressing the CaR (HEK-293/CaR) were those used in previousstudies [32]. COS-1 cells and the A7 melanoma cell line wereobtained from the A.T.C.C. and M2 cells were donated byProfessor Fumihiko Nakamura (Brigham and Women’s Hospital,Boston, MA, U.S.A.). All cells were propagated in DMEM(Dulbecco’s modified Eagle’s medium) containing 10 % FBS(fetal bovine serum) and antibiotics as described previously [30].In addition, for the HEK-293/CaR and A7 cells, G418 sulfate(Sigma–Aldrich) at 100 μg/ml and 500 μg/ml respectively, wasincluded in the medium to maintain the stable expression of CaRand filamin in these cell lines. Cell monolayers were transfectedat 40–60% confluency using LipofectamineTM 2000 (Invitrogen)as described previously [30].
Western blot analysis and co-immunoprecipitation studies
Following transfection, cells were washed, lysed in celllysis buffer [20 mM Tris/HCl (pH 6.8), 150 mM NaCl,10 mM EDTA, 1 mM EGTA and 1% (v/v) Triton X-100]containing iodoacetamide and protease inhibitors, and theextracted protein was quantified and subjected to Westernblot analysis as described previously [30]. Specifically, forco-immunoprecipitations, EGFP–14-3-3 (3 μg) and wild-typeor mutant CaR–FLAG (5 μg) were co-transfected into COS-1or HEK-293 cells and lysed 48 h later. Lysate protein (2 mg) wasthen pre-cleared with 40 μl of GammaBindTM G Sepharose beads(GE Healthcare Life Sciences) for 1 h at 4 ◦C. For EGFP–14-3-3pull-downs, pre-cleared lysate was mixed by rotation overnightat 4 ◦C with 6 μg of a rabbit polyclonal anti-GFP antibody, afterwhich the antibody–protein complex was mixed by rotation for4 hr at 4 ◦C with 40 μl of fresh Sepharose beads. The beadswere then washed six times with cell lysis buffer (with theomission of iodoacetamide) and the bound protein was elutedwith 40 μl of SDS sample buffer containing 2-mercaptoethanol.Proteins separated by SDS/PAGE (7.5% gel) were blotted andexamined for co-immunoprecipitated CaR–FLAG using anti-FLAG M2 monoclonal antibody as described previously [30].Reciprocal CaR–FLAG pull-downs were performed in essentiallythe same way, except that lysate containing 1.25 mg of proteinwas pre-cleared and mixed overnight with 5 μg of anti-FLAGM2 antibody prior to binding on to the beads. Proteins wereseparated on a PAGE gel (10 %) and, for immunodetection,a rabbit polyclonal anti-EGFP antibody (dilution 1:1000) wasused as the primary antibody. Note that the CaR is expressed intwo forms: a mature form modified with complex carbohydrates(150 kDa) and an immature form modified with high mannosecarbohydrates (130 kDa) [1,2].
In vitro pull-down studies
A GST fusion protein of 14-3-3θ was expressed in the E. coliBL21 codon ( + ) cells and purified by affinity chromatographyusing glutathione–Sepharose 4B beads (GE Healthcare LifeSciences) as described previously [33] except that the 14-3-3θprotein attached to the beads was cleaved from its GST moietyby thrombin cleavage. The CaR tail was expressed as an His6
fusion protein in the E. coli BL21 codon ( + ) cells and purifiedby affinity chromatography using Ni-NTA (Ni2 + -nitrilotriaceticacid)–agarose beads in an 8 M urea denaturation/renaturationprocedure essentially as described previously [18] with theinclusion of phosphatase inhibitors (1 mM sodium orthovanadateand 10 mM 2-glycerophosphate) as well as protease inhibitorsin the buffers. For the pull-down experiments, 32 μg of purified14-3-3θ protein was mixed by rotation overnight at 4 ◦C with anequimolar ratio of purified His6-tagged CaR tail (approximately50 μl of beads) in renaturation buffer (buffer B) [25 mMimidazole, 0.2% Triton X-100, 2.5 mM 2-mercaptoethanol,1 mM sodium orthovanadate, 10 mM 2-glycerophosphate alldissolved in PBS (pH 7.4)] after which time, beads (on ice)were washed five times in buffer B then five times in bufferB without Triton X-100. The beads were then resuspended in50 μl of SDS sample buffer containing 2-mercaptoethanol andboiled for 10 min. Samples were centrifuged and proteins in thesupernatant were separated by SDS/PAGE (12 % gel) and stainedwith Coomassie Brilliant Blue.
Site-directed mutagenesis
The pcDNA3.1-CaR–FLAG construct was used as a template forthe construction of various CaR tail mutants, pcDNA3.1-CaR–FLAG-S895A, in which the serine residue at amino acid 895was mutated to alanine; pcDNA3.1-CaR–FLAG-�RRSNVS, inwhich a putative 14-3-3 consensus binding site, encompassingamino acids 890–895, was deleted; and pcDNA3.1-CaR–FLAG-RKR/AAA in which amino acids 896–898 encompassing an ER-retention motif (RKR) were each mutated to alanine. Mutationswere generated using the QuikChange® site-directed mutagenesiskit (Stratagene). The part of the CaR tail containing the mutationand bound by the restriction enzymes SmaI and XbaI wassequenced and, once authenticated, inserted via a cassette intoSmaI- and XbaI-digested wild-type pcDNA1-CaR–FLAG [30],then subcloned into wild-type pcDNA3.1-CaR–FLAG using theunique HindIII and XbaI sites.
Confocal microscopy
HEK-293/CaR cells were transfected with EGFP–14-3-3 (orEGFP alone) and 24 h later seeded on to poly-L-lysine-coatedcoverslips and incubated overnight. The next day, cells were fixedwith 4% paraformaldehyde, permeabilized with 0.2 % Triton X-100 in PBS for 30 min, then blocked (10% goat serum and 1%BSA in PBS) prior to incubation for 1 h in fresh blocking solutionwith primary antibody to detect the CaR (1 μg/ml of ADD mousemonoclonal antibody and a 1:750 dilution of a rabbit anti-PDIpolyclonal primary antibody to identify the ER). EGFP–14-3-3 was detected by the fluorescence emitted from its EGFP tag.All incubations and washes were performed at room temperature(23 ◦C). Following washing, cells were incubated for 45 min withsecondary antibody in fresh blocking solution (1:400 dilution ofgoat anti-mouse Alexa Fluor® 546 and 1:400 dilution goat anti-rabbit Alexa Fluor® 647 antibodies for detection of CaR and PDIrespectively). Cells were mounted on to glass slides using low-fade mounting medium, sealed with nail polish and examinedunder a Nikon 60× NA (numerical aperture) 1.4 oil-immersion
c© The Authors Journal compilation c© 2012 Biochemical Society

998 A. Arulpragasam and others
lens using a Bio-Rad Laboratories MRC-1000/1024 UV confocallaser-scanning microscope with excitation wavelengths of either488 nm (FITC), 543 nm (tetramethyl rhodamine isothiocyanate)or 633 nm (far-red).
ERK1/2 assay
The ERK1/2 assay was based on an assay described previously[34]. HEK-293/CaR cells in 25 cm2 flasks were transfectedwith either EGFP–14-3-3, untagged 14-3-3 or vector alone, and24 h later seeded on to poly-L-lysine-coated 24-well plates. Thefollowing day, cells were serum-starved by incubating overnightin DMEM containing 1.5 mM Ca2 + and 0.2% BSA, and thenext day for 30 min in PSS (physiological saline solution)containing 0.5 mM Ca2 + and 0.2% BSA. The cells were thenleft unstimulated or stimulated with 1, 2 or 4 mM Ca2 + in PSScontaining 0.1% BSA for 5 min. The reaction was terminated bythe addition of ice-cold PBS to the cells which were then lysedin 100 μl of MAPK lysis buffer [150 mM NaCl, 24.7 mM NaF,50 mM 2-glycerophosphate, 2 mM sodium orthovanadate, 1 mMdithiothreitol and 2 mM EDTA in 20 mM Tris/HCl (pH 7.4) with10% glycerol, 1% (v/v) Triton X-100 and protease inhibitors]and centrifuged (15800 g for 3 min at 23 ◦C), and the protein inthe supernatant quantified. Per treatment, 20 μg of protein wasseparated on a 10% PAGE gel and examined by Western blotanalysis for phosphorylated ERK1/2 using a rabbit polyclonalanti-phospho-ERK1/2 antibody at a dilution of 1:1000 for 1 hfollowed by HRP-conjugated goat anti-(rabbit IgG) at a dilution of1:10000 for 1 h. Blocking solution and diluent for the antibodiesconsisted of 5% BSA in TBS containing 0.2% Tween 20,otherwise immunodetection was as described previously [30].For examination of total ERK1/2 expression, the membranewas stripped, blocked and incubated with rabbit polyclonal anti-ERK1/2 antibody at a dilution of 1:5000 for 1 h followed byHRP-conjugated goat anti-rabbit IgG at a dilution of 1:10000 for1h with blocking solution/antibody diluent as described for thedetection of phosphorylated ERK1/2.
To examine the effect of 14-3-3ζ depletion on CaR-mediatedERK1/2 activity, HEK-293/CaR cells were transfected with avalidated predesigned Silencer Select siRNA (small interferingRNA) oligonucleotide primer specific for 14-3-3ζ (sequence 5′-GGAGGGTCGTCTCAAGTAT-3′; Ambion), at a concentrationof 20 nM using LipofectamineTM 2000, and 24h later seeded onto poly-L-lysine coated 24-well plates for the ERK1/2 assay. Toevaluate ERK1/2 activity in the presence of endogenous 14-3-3ζlevels, a Silencer Select negative control oligonucleotide primerwas used in parallel experiments. The siRNA primer specific for14-3-3ζ regularly achieved over 50 % knockdown of 14-3-3ζcompared with the control primer (Supplementary Figure S1A athttp://www.BiochemJ.org/bj/441/bj4410995add.htm).
SRE–Luc reporter assays
HEK-293/CaR cells in 25 cm2 flasks were transfected with pSRE–Luc and either untagged 14-3-3 or vector alone and 24 h latercells were harvested and grown in DMEM containing 0.5 mMCa2 + , 0.1% BSA and antibiotics overnight in poly-L-lysine-coated six-well plates. The following day, half of the cells werestimulated with 0.5 mM Ca2 + and the other half were stimulatedwith 5 mM Ca2 + for 7 h in DMEM and 0.1% BSA, after whichthe cells were incubated for 10 min on ice in 180 μl of Luclysis buffer [10% (v/v) glycerol and 0.1 % Triton X-100 in30 mM Tris/HCl (pH 7.8) containing 2 mM EDTA and 2 mMfreshly added dithiothreitol], completely lysed by freeze/thawingand pipetting, then centrifuged (15800 g for 5 min at 4 ◦C).
Protein levels in the clarified cell lysates were quantified and50 μl of lysate transferred to a 96-well Optiplate (PerkinElmerLife Sciences) and 50 μl of Luc reagent from the LuciferaseAssay System kit (Promega) was injected automatically and thenread using a POLARstar Optima microplate reader. Luc activitywas measured at 20 s intervals and readings were adjusted to Lucactivity per microgram of protein per sample. For the M2 and A7cells, the same procedure was applied except that 75 cm2 flasks ofcells were first transfected with CaR–FLAG, then 24 h later weresplit and the cells were transferred into 25 cm2 flasks for test andcontrol luciferase assays.
To examine the effect of 14-3-3ζ depletion on CaR-mediatedSRE activity, HEK-293/CaR cells were transfected with ansiRNA oligonucleotide primer specific for 14-3-3ζ (or a controloligonucleotide primer), as described in the previous section, and24 h later were transfected with pSRE–Luc. The following day, thecells were grown in DMEM containing 0.5 mM Ca2 + , 0.1% BSAand antibiotics overnight in poly-L-lysine-coated six-well platesin readiness for the Luc reporter assay performed as describedabove. The siRNA primer specific for 14-3-3ζ resulted in over50% knockdown of 14-3-3ζ compared with the control primer(Supplementary Figure S1B).
ELISA-based cell-surface-expression assay
The assay was based on that described previously [35]. HEK-293 cells were transfected with exFLAG–CaR and after a 4 hincubation seeded into poly-L-lysine-coated six-well plates. Thefollowing day, cells were transfected with either untagged 14-3-3 or vector alone and 48 h later were incubated for 90 minat 4 ◦C with DMEM containing 10% FBS and 1:2000 dilution(∼2.5 μg/ml) mouse M2 anti-FLAG antibody. Cells were thendetached by gently washing in ice-cold PBS and incubatedfor 1 h at 4 ◦C in DMEM containing 10% FBS and 1:5000dilution HRP-conjugated goat anti-(mouse IgG). The cells werethen washed and centrifuged (200 g for 2 min at 4 ◦C), andthe cell pellet was resuspended in 200 μl of TMB (3,3′,5,5′-tetramethylbenzidine) liquid peroxidase substrate for ELISA(Sigma–Aldrich) for 20 min at room temperature in the dark.Cells were centrifuged (200 g for 5 min at 23 ◦C) and 50 μl of thesupernatant was transferred (in triplicate) into a 96-well microtitreplate and the reaction stopped by adding HCl (final concentration0.5 M). Absorbance was read at wavelength of 450 nm using aPOLARstar Optima microplate reader. Background readings wereascertained using cells transfected with empty vector alone.
Densitometry and statistical analyses
For the ERK1/2 assay Western blots, bands were scanned using aScanjet 6200C scanner (Hewlett Packard) and densities measuredusing Scion Image software (Scion Corporation) with correctionfor background. Phospho-ERK1/2 values were expressed relativeto total ERK1/2 and for each experiment the phospho-ERK1/2values for 0 mM Ca2 + /empty vector were assigned a value of ‘1’and other treatment values expressed relative to this.
For ELISA-based cell surface expression and SRE activationstudies, mean absorbance and luciferase activity data wereanalysed via mixed effects models, with each experiment treatedas a random effect, and transfection and stimulation conditions asfixed effects. Graphical results are presented as means +− S.E.M.ERK1/2 densities were evaluated in a similar way, with calciumdosage and 14-3-3θ expression treated as fixed effects. Allstatistical analyses were conducted in PASW 18 and resultsassessed at the 5% significance level.
c© The Authors Journal compilation c© 2012 Biochemical Society

14-3-3 modulates calcium-sensing receptor signalling 999
Figure 1 Binding of 14-3-3θ to the membrane proximal region of the CaRintracellular tail
CaR tail deletion construct and 14-3-3θ plasmid DNA were co-transformed into yeast L40,plated on to selection plates and transformant colonies screened for protein interaction using aβ-galactosidase colony lift assay. The relative strength of reporter interaction between each CaRtail deletion and 14-3-3θ , as judged by time to first evidence of colour development comparedwith full-length CaR tail ( + + + ) is shown at the far right of each deletion construct fragment.This result was observed in three separate experiments. TMD, transmembrane domain.
RESULTS
14-3-3 protein binds to the membrane proximal region of the CaRintracellular tail
Full-length murine 14-3-3θ protein was identified as a bindingpartner for the human CaR intracellular tail (Figure 1). Theinteraction in yeast was verified by co-transformation of rescuedlibrary plasmid and CaR tail bait plasmid into L40 yeast andobservation of LacZ reporter gene activation using a colony liftassay. Amino acid sequence alignments of the murine and humanisoforms of 14-3-3θ showed a single conservative amino aciddifference at amino acid 143, with a glutamate residue in themurine sequence exchanged for aspartate in the human sequence.The near identities of the two isoforms of 14-3-3θ supported thenotion that they would bind identically to the human CaR tail,hence mapping studies using human CaR tail deletions as baitwere performed using the murine 14-3-3θ clone in LacZ reportergene assays. These studies showed that the 14-3-3θ protein boundto the proximal membrane region of the CaR tail (amino acidresidues 865–922), but not to a juxtaposed C-terminal region(amino acids 923–1078) (Figure 1). The interaction with CaRpeptide 865–922 was comparable with that with the full-lengthCaR tail, indicating that the membrane-proximal region containsthe entire binding site. Neither of the two subfragments of themembrane proximal region (amino acids 865–898 and 899–922)was positive in the LacZ reporter assays, suggesting that residuesfrom both subfragments are required for interaction with 14-3-3θ .In addition, a truncated version of another 14-3-3 isoform, 14-3-3ζ , was identified as a binding partner in the yeast two-hybridscreen. Deletion mapping studies showed that the full-lengthhuman 14-3-3ζ isoform bound to the same membrane-proximalregion of the CaR as described for 14-3-3θ (results not shown).For subsequent studies, the human DNA sequence for 14-3-3θwas recovered by RT–PCR from Saos-2 cells and cloned intobacterial and mammalian expression vectors (see the Materialsand methods section).
14-3-3θ and the CaR interact directly in pull-down studies in vitroand interact in mammalian cells
To determine whether the 14-3-3θ protein directly interacts withthe CaR tail peptide in vitro, we performed pull-down studies
Figure 2 The CaR tail and 14-3-3θ demonstrate direct interaction in anin vitro pull-down assay
14-3-3θ expressed as a GST-fusion protein was purified by affinity chromatography and cleavedof its GST moiety, and 32 μg was used in pull-down experiments with an approximatelyequimolar amount of His6-tagged CaR tail protein purified on Ni-NTA beads (see the Materials andmethods section). After washing, proteins eluted from the beads were separated by SDS/PAGE(12 % gel) and the gel was stained with Coomassie Brilliant Blue. Note that the His–CaR tailband runs slower than expected consistent with previous observations using this procedure[18]. The dividing line in the composite image signifies lanes grouped from different parts of thesame gel. The gel is representative of two separate experiments. The molecular mass is given inkDa on the left-hand side.
with purified 14-3-3θ and CaR tail proteins. Bacterially expressed14-3-3θ fused to GST was purified by affinity chromatographyusing glutathione–Sepharose 4B beads, cleaved of its GST moietyand then examined in pull-down experiments with a bacteriallyexpressed His6-tagged CaR tail protein that was purified usingNi-NTA beads. It was found that 14-3-3θ directly interacted withthe CaR tail immobilized on Ni-NTA beads (Figure 2, right-handlane) and no binding of 14-3-3θ was detected in the absence ofthe CaR tail protein (Figure 2, middle lane).
In order to determine whether the 14-3-3θ–CaR interactionobserved in yeast also occurs in mammalian cells, we performedco-immunoprecipitation studies. Human full-length 14-3-3θ ,cloned into the vector pcDNA3–EGFP for expression as anEGFP-tagged protein and pcDNA3.1 containing the full-lengthCaR for expression as a FLAG-tagged protein (CaR–FLAG),were co-expressed in COS-1 or HEK-293 cells. Lysates fromco-transfected HEK-293 cells were immunoprecipitated withanti-GFP antibody to pull down 14-3-3θ and found to co-immunoprecipitate the wild-type CaR (Figure 3A, top panel). In areciprocal experiment performed in COS-1 cells, when the lysatewas immunoprecipitated with anti-FLAG antibody to pull downthe CaR, 14-3-3θ was found to co-immunoprecipitate (Figure 3B,top panel). There was negligible non-specific binding of eitherFLAG–CaR or EGFP–14-3-3 to the GammaBindTM G Sepharosebeads used for immunoprecipitation (Figure 3A, top panel, andFigure 3B, top panel respectively). The results demonstrate thatthe CaR and 14-3-3θ interact in mammalian COS-1 and HEK-293cells. The same results were obtained in co-immunoprecipitationstudies with CaR–FLAG and EGFP–14-3-3ζ (not shown).
Binding of 14-3-3θ to the CaR does not require a 14-3-3 consensusbinding sequence in the CaR tail and is independent of PKCphosphorylation
It has been shown previously that 14-3-3 proteins bind tosome protein partners via a phosphorylated serine recognitionmotif, RSXpSXP [27]. A recognized variant, the so-calledserine-rich motif, RX1/2SX2/3S (in which at least one serineresidue is phosphorylated) [27], is present in the membrane-proximal region of the CaR tail (RRSNVS; residues 890–895) [3].Therefore we performed experiments to assess its significance in
c© The Authors Journal compilation c© 2012 Biochemical Society

1000 A. Arulpragasam and others
Figure 3 Wild-type CaR–FLAG and EGFP–14-3-3θ immunoprecipitate, buta putative 14-3-3 consensus binding site on the CaR tail is not required forCaR–14-3-3θ interaction
(A) Effect of mutating the serine phosphorylation site on the consensus binding sequence:lysate protein from HEK-293 cells, transfected with EGFP–14-3-3θ and either CaR–FLAG(wild-type) or CaR–FLAG-S895A (mutant), was immunoprecipitated with an anti-GFP antibodyto pull down EGFP–14-3-3θ and examined for co-association of wild-type or mutant CaRusing anti-FLAG antibody. (B) Effect of deleting the entire consensus binding sequence: lysateprotein from COS-1 cells transfected with EGFP–14-3-3θ and either CaR–FLAG (wild-type) orCaR–FLAG-�RRSNVS (deletion mutant), was immunoprecipitated with an anti-FLAG antibodyto pull down wild-type or mutant CaR–FLAG and examined for co-association of 14-3-3θ usinganti-GFP antibody. For both (A) and (B), the expression levels of EGFP–14-3-3θ and CaR–FLAG(wild-type and mutant) are shown in the middle and lower panels. Blots are representative ofthree separate experiments for both (A) and (B). The molecular mass is given in kDa on theleft-hand side. IP, immunoprecipitation; WB, Western blot.
14-3-3 protein binding, including the impact of CaR S895Aon the ability of 14-3-3θ to co-immunoprecipitate the CaRin HEK-293 cells (Figure 3A, top panel), as well as deletionof the entire consensus motif on the ability of mutant andwild-type CaRs to co-immunoprecipitate 14-3-3θ in COS-1cells (Figure 3B, top panel). The results demonstrated that thePKC phosphorylation site at Ser895 and the associated 14-3-3 consensus binding site in the CaR tail are not required forthe CaR–14-3-3θ interaction. Finally, we assessed the potentialsignificance of other PKC phosphorylation sites includingThr888 and Ser915 in the CaR’s 14-3-3-binding region on theability of the CaR to bind 14-3-3θ , by exposing COS-1cells to the PKC inhibitor GFX109203X (2 μM for 1 h) oractivator PMA (100 nM for 1 h) (Figures 4A and 4B respectively).Neither compound had a significant effect, indicating that theinteraction between the CaR tail and 14-3-3 is independent ofPKC phosphorylation of the receptor.
14-3-3 and the CaR co-localize in the ER in HEK-293 cells
In order to determine whether 14-3-3θ and the CaR co-localize inHEK-293 cells, we expressed EGFP–14-3-3θ in HEK-293/CaRcells and used confocal fluorescence microscopy to determinethe locations of: (i) the CaR using the anti-CaR ADD antibodyin conjunction with an Alexa Fluor® 546-labelled secondaryantibody, and (ii) EGFP–14-3-3θ via its EGFP tag. The ERwas identified using antibody against PDI in conjunction with
Figure 4 Neither PKC inhibition nor activation affects CaR–14-3-3θinteraction
(A) Effect of PKC inhibitor (GFX109203X): COS-1 cells transfected with EGFP–14-3-3θ (middlepanel) and CaR–FLAG (bottom panel) were treated 48 h later with either vehicle or 2 μMGFX109203X for 1 h prior to lysis and immunoprecipitation of protein with anti-FLAG antibodyto pull down CaR–FLAG. Co-associated 14-3-3θ was examined using anti–GFP antibody (toppanel). (B) Effect of PKC activator (PMA): COS-1 cells transfected with EGFP–14-3-3θ andCaR–FLAG were treated 48 h later with either vehicle or 100 nM PMA for 1 h prior to lysisand immunoprecipitation with anti-FLAG antibody and detection of co-associated 14-3-3θ(top panel). For both (A) and (B), the middle and bottom panels show expression levelsfor EGFP–14-3-3θ and CaR–FLAG respectively in untreated and treated lysates. Results arerepresentative of two separate experiments. The molecular mass is given in kDa on the left-handside. IP, immunoprecipitation; WB, Western blot.
an Alexa Fluor® 647-labelled secondary antibody. The CaRpredominantly localized in the ER, but was also detected inother regions of the cytoplasm and the plasma membrane insome cells (Figure 5, compare panels 1 and 2). EGFP–14-3-3θ also localized primarily in the ER in those cells in whichit was expressed (Figure 5, compare upper panels 2 and 3) andwas also detected elsewhere in the cytoplasm, unlike EGFP alonewhich predominantly localized to the nucleus (Figure 5, comparelower panels 2 and 3). Merged images derived from Figure 5(upper panels 1, 2 and 3), demonstrate co-localization of theCaR and 14-3-3θ in the ER as revealed by white fluorescence(Figure 5, upper panel 4). Although a quantitative assessmentcould not be made, 14-3-3θ appeared to have no effect on thepattern of CaR expression (Figure 5, compare CaR localizationin cells expressing and not expressing EGFP–14-3-3θ ). Theseexperiments were repeated using EGFP–14-3-3ζ in place ofEGFP–14-3-3θ with the same pattern of co-localization with theCaR observed (results not shown).
Overexpression of 14-3-3ζ (but not 14-3-3θ ) reduces CaRcell-surface expression, but not through disruption of binding to anRKR retention motif
On the basis of the results of the co-localization studies it washypothesized that 14-3-3 might play a role in post-translationalprocessing of the CaR in the ER; for example, by masking twoestablished ER-retention motifs (RR, residues 890 and 891, andRKR, residues 896–898) in the CaR tail [16,36–38]. In order todetermine whether 14-3-3 specifically binds to the RKR retentionmotif on the CaR, the RKR residues were each mutated to alanine(AAA) and the impact on binding with 14-3-3 was determined inco-immunoprecipitation experiments (Figure 6). Disrupting the
c© The Authors Journal compilation c© 2012 Biochemical Society

14-3-3 modulates calcium-sensing receptor signalling 1001
Figure 5 The CaR and EGFP–14-3-3θ co-localize in the ER in HEK-293/CaR cells
HEK-293/CaR cells were transfected with either EGFP–14-3-3θ (upper panels) or EGFP alone (lower panels) and 24 h later the cells were processed for confocal microscopy, as described in theMaterials and methods section, using the anti-CaR ADD mouse monoclonal antibody and a goat anti-mouse secondary antibody (Alexa Fluor® 546-conjugated) to localize the CaR (panels 1) andan anti-PDI rabbit polyclonal antibody and a goat anti-rabbit secondary antibody (Alexa Fluor® 647-conjugated) to locate the ER (panels 2). EGFP–14-3-3θ and EGFP alone are shown in topand bottom of panel 3 respectively. Merged images are shown in panels 4 with a particularly strong area of co-localization of CaR, PDI and EGFP–14-3-3θ evident in the upper panel (arrowed).Images are representative of four separate experiments.
RKR motif had no effect on the binding affinity of either 14-3-3θor 14-3-3ζ for the CaR. Nevertheless, it is possible that binding of14-3-3 to an adjacent region of the CaR may still mask either theRKR motif or upstream RR motif leading to reduced ER retentionand forward trafficking to the cell surface [36–38]. To investigatewhether overexpression of 14-3-3 affects CaR cell-surface expres-sion, we transiently transfected exFLAG–CaR in the presence ofuntagged 14-3-3θ or 14-3-3ζ (or an empty vector control) andperformed an intact cell-surface-expression assay using an anti-FLAG antibody. Untagged 14-3-3 isoforms were used for thesestudies to avoid any interference from fusion tags. Surprisingly,overexpression of 14-3-3ζ significantly reduced CaR cell-surfaceexpression, whereas 14-3-3θ overexpression, although showinga similar trend, did not significantly reduce CaR cell-surfaceexpression (Figure 7A). Western blot analysis demonstrated anabundance of 14-3-3θ and, to a lesser extent, 14-3-3ζ overendogenously expressed levels in cells transfected in parallel withthose used for the cell surface expression studies (Figure 7B).
Altered expression of 14-3-3θ and 14-3-3ζ has no effect onCaR-mediated ERK1/2 activation
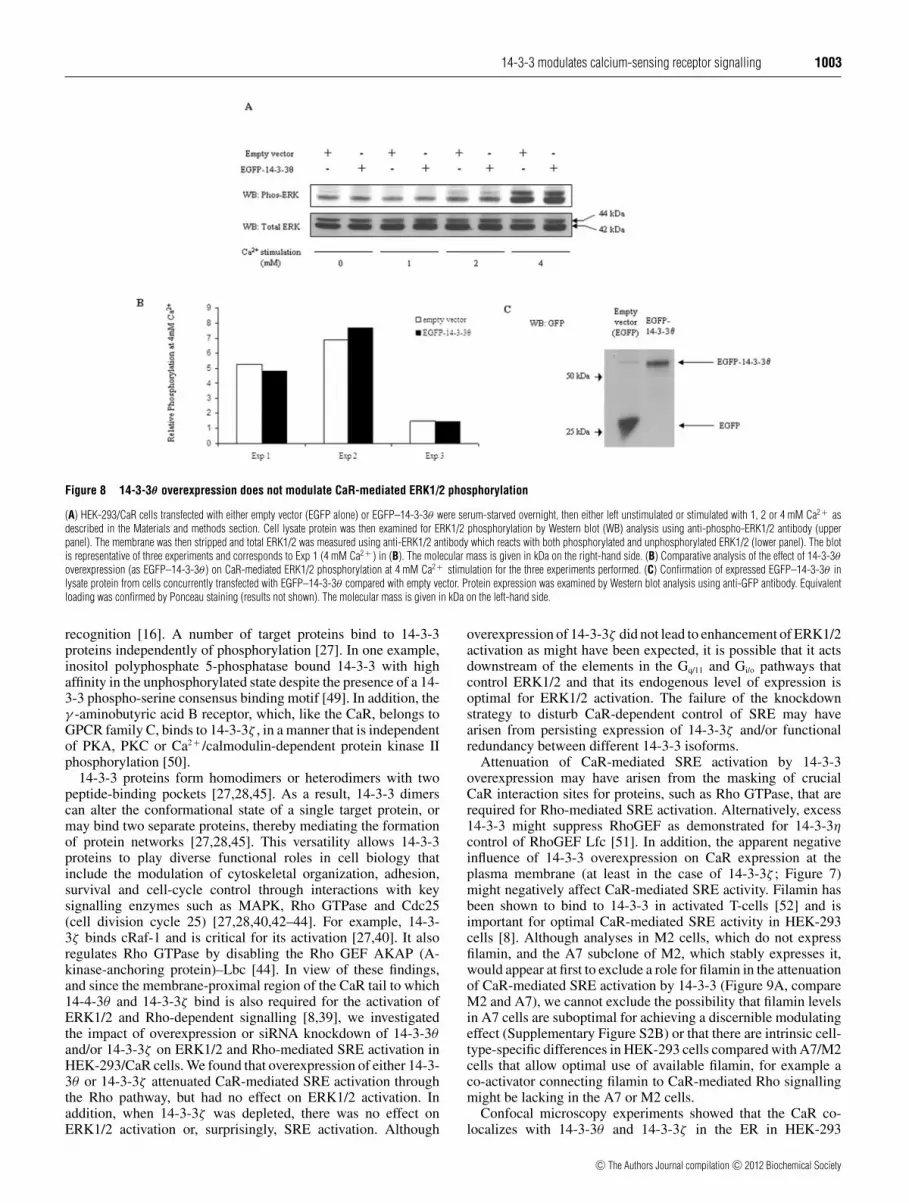
The region of the CaR tail to which 14-3-3θ bindsmediates various cell signalling events [2,4,7]. In particular,filamin-dependent ERK1/2 activation requires a predicted α-helix proximal to the membrane (residues 868–879) [39],which lies within the 14-3-3-binding domain. Therefore weinvestigated the impact of 14-3-3θ overexpression on CaR-mediated ERK1/2 activation (Figure 8). Elevated Ca2 + stimulatedERK1/2 phosphorylation as expected; however, at any givenCa2 + concentration, there was no appreciable difference inphosphorylated ERK1/2 or in total ERK1/2 between the controlcells and cells overexpressing 14-3-3θ (Figure 8A). Furthermore,when blots from all three experiments were examined bydensitometry and the data were pooled, there was no significantdifference (P < 0.05) between the mean relative phospho-ERK1/2 values of control and overexpressed 14-3-3θ at anyCa2 + concentration including that for 4 mM Ca2 + (P = 0.8346)(Figure 8B). Expression of EGFP–14-3-3θ was confirmed inHEK-293/CaR cells concurrently transfected with EGFP–14-3-3θ (Figure 8C). Since these experiments were performedwith EGFP-tagged 14-3-3θ , we repeated the experiment withoverexpressed untagged 14-3-3 to confirm that there was no
interference from the tag. As with the tagged 14-3-3θ experiments,there was no difference in phospho-ERK1/2 activity betweencontrol and overexpressed untagged 14-3-3θ (results not shown).Analogous experiments were performed with untagged 14-3-3ζwith the same outcome (results not shown). In addition, owing toa previously defined role for 14-3-3ζ in MAPK signalling [40],we investigated the effect of transient knockdown of 14-3-3ζ onCaR-mediated ERK1/2 activation in HEK-293/CaR cells using aspecific 14-3-3ζ siRNA oligonucleotide primer. No difference inCa2 + -dependent activation of ERK1/2 was observed compared toa negative siRNA oligonucleotide control (results not shown).
Overexpression of 14-3-3θ and 14-3-3ζ attenuates CaR-mediatedSRE activation, but depletion of 14-3-3ζ has no effect
Elevated Ca2 + activates SRE in CaR-expressing HEK-293 cellsvia a Rho-dependent pathway that is blocked by expression of aCaR C-terminal minigene encoding residues 906–980 [8]. Sincethis region overlaps the CaR 14-3-3θ -binding site (residues 865–922; Figure 1), we investigated whether overexpression of 14-3-3θ might modulate extracellular Ca2 + -induced SRE activationin CaR-expressing HEK-293 cells transfected with pSRE–Luc(Figure 9A, HEK-293/CaR). We used untagged isoforms of14-3–3 for these overexpression studies to avoid any possibleinterference from tags. Compared with control HEK-293/CaRcells, those in which 14-3-3θ was overexpressed exhibited agreater than 50% reduction in 5 mM Ca2 + -induced, SRE-mediated Luc expression (P < 0.05; Figure 9A, HEK-293/CaR,compare ‘I’ and ‘II’). Overexpression of 14-3-3ζ had a similarsuppressive effect (results not shown).
Since optimal CaR-mediated SRE activation is dependent uponfilamin [8], we next investigated whether the inhibitory effectof 14-3-3θ might be modulated by filamin, by comparing theimpact of elevated Ca2 + (5 mM) on SRE activation in CaRand 14-3-3θ -transfected M2 cells, which do not express filamin,with CaR and 14-3-3θ -transfected A7 cells, an M2 clone thatstably expresses filamin [41]. No difference in the effect of14-3-3θ was observed in these two cells types (Figure 9A,compare M2 and A7). A similar outcome was observed for 14-3-3ζ (results not shown). In control experiments, the expressionof filamin in A7 cells and its absence in M2 cells wasconfirmed by Western blotting (Supplementary Figure S2 athttp://www.BiochemJ.org/bj/441/bj4410995add.htm). However,
c© The Authors Journal compilation c© 2012 Biochemical Society

1002 A. Arulpragasam and others
Figure 6 Disruption of the RKR retention motif on the CaR has no effect on 14-3-3 binding
Lysate protein from COS-1 cells, transfected with EGFP–14-3-3θ or EGFP–14-3-3ζ and either CaR–FLAG (wild-type) or CaR–FLAG-RKR/AAA (mutant), was immunoprecipitated with anti-FLAGantibody to pull down wild-type or mutant CaR and examined for co-associated 14-3-3θ or 14-3-3ζ using anti-GFP antibody (top panel). Expression levels of EGFP–14-3-3 isoforms and CaR–FLAG(wild-type and mutant) in the lysates are shown in the middle and bottom panels respectively. Blots are representative of three separate experiments. The molecular mass is given in kDa on theleft-hand side. IP, immunoprecipitation; WB, Western blot.
Figure 7 14-3-3ζ overexpression reduces CaR cell-surface expression
(A) HEK-293 cells were transfected with exFLAG–CaR in a single flask and several hourslater distributed into six-well plates. Cells were transfected the next day with pcDNA3.1 alone(1), untagged 14-3-3θ (2) or untagged 14-3-3ζ (3). Post-transfection (48 h), the cells wereexamined for cell surface expressed CaR, as described in the Materials and methods section,using anti-FLAG primary antibody, HRP-conjugated goat anti-mouse secondary antibody andcolour detection with TMB substrate. After stopping the reaction with HCl, the absorbancewas measured at 450 nm. Results are means +− S.E.M. from three separate experiments, eachperformed in triplicate. Absorbance values with different superscripts are significantly different(P<0.05). (B) Confirmation of overexpressed (i) 14-3-3θ or (ii) 14-3-3ζ by comparison of cellstransfected with vector alone ( − ) or with untagged 14-3-3θ or 14-3-3ζ ( + ). ExtracellularlyFLAG-tagged CaR-expressing cells were transfected concurrently with those used for the cellsurface expression assay and lysates were examined 48 h post-transfection by Western blot(WB) analysis using anti-14-3-3θ or anti-14-3-3ζ antibody. Dividing lines in the compositeimages in (ii) signify lanes grouped from different parts of the same gel. Blots are representativeof three experiments performed. The molecular mass is given in kDa on the left-hand side.
filamin expression in A7 cells was considerably less than thatdetected in HEK-293/CaR cells. Since 14-3-3ζ is reported toplay a key role in Rho signalling [42–44], we also examinedthe effect of transient knockdown of 14-3-3ζ on SRE reporteractivation in HEK-293/CaR cells using a specific 14-3-3ζ siRNAoligonucleotide primer. There was no difference in SRE reporteractivation compared with a negative siRNA oligonucleotidecontrol (results not shown).
DISCUSSION
In the present study, a yeast two-hybrid screen revealed that twoisoforms of the 14-3-3 adaptor protein, 14-3-3θ and 14-3-3ζ , arebinding partners of the CaR intracellular tail and EGFP-taggedforms of these two 14-3-3 isoforms were found to interact withCaR–FLAG when overexpressed in mammalian cells. Previousstudies have shown that the CaR interacts with endogenous levelsof 14-3-3 using a pan antibody to immunoprecipitate 14-3-3 [16].There are seven mammalian isoforms of 14-3-3 (β, γ , ε, η,σ , θ and ζ ), with the θ and ζ isoforms being phylogeneticallyclosely related with approximately 80 % amino acid identity[45]. It is therefore not surprising that they both bound to thesame membrane proximal region of the CaR tail, exhibitedsimilar patterns of co-localization and co-immunoprecipitationwith the CaR, and suppressed CaR-dependent activation of SRE.The 14-3-3 isoforms are differentially expressed in tissues and,consequently, appear to play distinct roles in mammalian biology[28,45]. It was from rat brain that 14-3-3θ was first cloned [46] andsubsequently mouse testis in which it plays a role in the controlof spermatogenesis via the regulation of Sertoli cell adhesion[47]. Interestingly, H-500 Leydig cell (testicular tumour) growthis controlled in part by the pituitary tumour-transforming genewhich is up-regulated by the induction of high extracellular Ca2 +
mediated by the CaR [48]. Whether 14-3-3θ binding to the CaRis required for the control of Leydig and/or Sertoli cell fate orfunction remains to be determined. The 14-3-3ζ isoform, onthe other hand, is widely expressed and plays roles in numerouscellular events [28]. For example, it is an important adaptor proteinin the MAPK and Rho family signalling pathways and regulatesthe cytoskeleton [27,40,42,43].
In many cases, 14-3-3 protein binding requires thephosphorylation of specific serine or threonine residues inconsensus binding sites on target proteins [27,28]. The locationof a 14-3-3 phospho-serine recognition motif [27] within the CaRtail-binding region prompted us to examine its significance for14-3-3θ binding. Surprisingly, neither mutation of the putativephosphorylation target Ser895 to alanine nor deletion of the entireconsensus binding site (RRSNVS; residues 890–895) impaired14-3-3θ binding. In addition, we excluded a role for PKCphosphorylation in 14-3-3θ binding to the CaR since neitherthe PKC inhibitor GFX109203X nor the PKC activator PMAinfluenced the 14-3-3θ–CaR interaction. The results suggest thatthe CaR may fall into the category of target proteins that donot require phosphorylation for 14-3-3 binding, although there isrecent evidence that the PKA site at Ser899 could influence 14-3-3
c© The Authors Journal compilation c© 2012 Biochemical Society
14-3-3 modulates calcium-sensing receptor signalling 1003
Figure 8 14-3-3θ overexpression does not modulate CaR-mediated ERK1/2 phosphorylation
(A) HEK-293/CaR cells transfected with either empty vector (EGFP alone) or EGFP–14-3-3θ were serum-starved overnight, then either left unstimulated or stimulated with 1, 2 or 4 mM Ca2 + asdescribed in the Materials and methods section. Cell lysate protein was then examined for ERK1/2 phosphorylation by Western blot (WB) analysis using anti-phospho-ERK1/2 antibody (upperpanel). The membrane was then stripped and total ERK1/2 was measured using anti-ERK1/2 antibody which reacts with both phosphorylated and unphosphorylated ERK1/2 (lower panel). The blotis representative of three experiments and corresponds to Exp 1 (4 mM Ca2 + ) in (B). The molecular mass is given in kDa on the right-hand side. (B) Comparative analysis of the effect of 14-3-3θoverexpression (as EGFP–14-3-3θ ) on CaR-mediated ERK1/2 phosphorylation at 4 mM Ca2 + stimulation for the three experiments performed. (C) Confirmation of expressed EGFP–14-3-3θ inlysate protein from cells concurrently transfected with EGFP–14-3-3θ compared with empty vector. Protein expression was examined by Western blot analysis using anti-GFP antibody. Equivalentloading was confirmed by Ponceau staining (results not shown). The molecular mass is given in kDa on the left-hand side.
recognition [16]. A number of target proteins bind to 14-3-3proteins independently of phosphorylation [27]. In one example,inositol polyphosphate 5-phosphatase bound 14-3-3 with highaffinity in the unphosphorylated state despite the presence of a 14-3-3 phospho-serine consensus binding motif [49]. In addition, theγ -aminobutyric acid B receptor, which, like the CaR, belongs toGPCR family C, binds to 14-3-3ζ , in a manner that is independentof PKA, PKC or Ca2 + /calmodulin-dependent protein kinase IIphosphorylation [50].
14-3-3 proteins form homodimers or heterodimers with twopeptide-binding pockets [27,28,45]. As a result, 14-3-3 dimerscan alter the conformational state of a single target protein, ormay bind two separate proteins, thereby mediating the formationof protein networks [27,28,45]. This versatility allows 14-3-3proteins to play diverse functional roles in cell biology thatinclude the modulation of cytoskeletal organization, adhesion,survival and cell-cycle control through interactions with keysignalling enzymes such as MAPK, Rho GTPase and Cdc25(cell division cycle 25) [27,28,40,42–44]. For example, 14-3-3ζ binds cRaf-1 and is critical for its activation [27,40]. It alsoregulates Rho GTPase by disabling the Rho GEF AKAP (A-kinase-anchoring protein)–Lbc [44]. In view of these findings,and since the membrane-proximal region of the CaR tail to which14-4-3θ and 14-3-3ζ bind is also required for the activation ofERK1/2 and Rho-dependent signalling [8,39], we investigatedthe impact of overexpression or siRNA knockdown of 14-3-3θand/or 14-3-3ζ on ERK1/2 and Rho-mediated SRE activation inHEK-293/CaR cells. We found that overexpression of either 14-3-3θ or 14-3-3ζ attenuated CaR-mediated SRE activation throughthe Rho pathway, but had no effect on ERK1/2 activation. Inaddition, when 14-3-3ζ was depleted, there was no effect onERK1/2 activation or, surprisingly, SRE activation. Although
overexpression of 14-3-3ζ did not lead to enhancement of ERK1/2activation as might have been expected, it is possible that it actsdownstream of the elements in the Gq/11 and Gi/o pathways thatcontrol ERK1/2 and that its endogenous level of expression isoptimal for ERK1/2 activation. The failure of the knockdownstrategy to disturb CaR-dependent control of SRE may havearisen from persisting expression of 14-3-3ζ and/or functionalredundancy between different 14-3-3 isoforms.
Attenuation of CaR-mediated SRE activation by 14-3-3overexpression may have arisen from the masking of crucialCaR interaction sites for proteins, such as Rho GTPase, that arerequired for Rho-mediated SRE activation. Alternatively, excess14-3-3 might suppress RhoGEF as demonstrated for 14-3-3ηcontrol of RhoGEF Lfc [51]. In addition, the apparent negativeinfluence of 14-3-3 overexpression on CaR expression at theplasma membrane (at least in the case of 14-3-3ζ ; Figure 7)might negatively affect CaR-mediated SRE activity. Filamin hasbeen shown to bind to 14-3-3 in activated T-cells [52] and isimportant for optimal CaR-mediated SRE activity in HEK-293cells [8]. Although analyses in M2 cells, which do not expressfilamin, and the A7 subclone of M2, which stably expresses it,would appear at first to exclude a role for filamin in the attenuationof CaR-mediated SRE activation by 14-3-3 (Figure 9A, compareM2 and A7), we cannot exclude the possibility that filamin levelsin A7 cells are suboptimal for achieving a discernible modulatingeffect (Supplementary Figure S2B) or that there are intrinsic cell-type-specific differences in HEK-293 cells compared with A7/M2cells that allow optimal use of available filamin, for example aco-activator connecting filamin to CaR-mediated Rho signallingmight be lacking in the A7 or M2 cells.
Confocal microscopy experiments showed that the CaR co-localizes with 14-3-3θ and 14-3-3ζ in the ER in HEK-293
c© The Authors Journal compilation c© 2012 Biochemical Society

1004 A. Arulpragasam and others
Figure 9 14-3-3θ overexpression attenuates CaR-mediated SRE activation in HEK-293/CaR cells, but not in M2 or A7 cells
(A) HEK-293/CaR cells and CaR–FLAG-expressing M2 or A7 cells were transfected with pSRE–Luc and either empty vector (I) or untagged 14-3-3θ (II). The following day, cells were stimulated with0.5 mM or 5 mM Ca2 + then lysed and SRE activation in the lysates was measured using a luciferase reporter assay as described in the Materials and methods section. Luc activities, normalized forprotein levels, are means +− S.E.M. for three separate experiments each performed in triplicate. For each cell line, Luc values sharing the same superscript were not significantly different, whereasthose with a different superscript were significantly different (P<0.05). (B) Confirmation of overexpressed 14-3-3θ in cells transfected concurrently with those used for the luciferase reporter assaywith 14-3-3θ ( + ) compared with empty vector alone ( − ) and confirmation of equivalently expressed CaR or CaR–FLAG in HEK-293/CaR and M2 or A7 cells respectively for the two treatments.Lysates were examined 48 h following transfection by Western blot (WB) analysis using anti-14-3-3θ , anti-CaR ADD or anti-FLAG antibodies. Equivalent loading was confirmed in all cases byPonceau staining (results not shown).
cells stably expressing the CaR. Trafficking of some cell-surface proteins from the ER to the plasma membrane ispromoted by 14-3-3 proteins by masking ER-retention signalsin their binding partners [36,38]. In two examples, 14-3-3binding masks a dibasic retention motif in the KCNK3 K+
channel [37] and a tribasic RKR motif in the α-subunit ofthe KATP channel Kir6.2 [38], thereby disrupting their bindingto the vesicle COPI (coat protein I) involved in proteinretrieval in the ER. A recent study by Stepanchick et al.[16] has found that the RKR motif of the CaR (amino acids896–898) is a genuine retention motif with conversion of RKRinto AAA, leading to plasma membrane abundance. TheRKR motif forms part of an arginine-rich region (residues 890–898), which also includes a dibasic RR motif (residues 890 and891) that is regulated by phosphorylation at Ser899 by PKA.14-3-3 proteins have been shown to bind the arginine-rich regionwith binding considerably reduced when both the RR and RKRsites were converted into alanine residues, suggesting that 14-3-3binding to this region might regulate ER retention of the CaR[16]. Interestingly, we found in the present study that mutationof the RKR site alone did not reduce 14-3-3 binding (Figure 6),suggesting that the dibasic site in the arginine-rich region couldbe more important than the RKR site for 14-3-3 interaction. Thereduced plasma membrane expression of the CaR resulting fromoverexpression of 14-3-3ζ is consistent with the observations ofStepanchick et al. [16], but is contrary to the currently held viewthat 14-3-3 proteins competitively inhibit the binding of COPIto arginine-based motifs to allow release of target protein from
the ER and trafficking to the plasma membrane [36–38]. Clearly,further investigation is required to elucidate the role played by14-3-3 proteins in CaR trafficking.
The 14-3-3 isoforms play key roles in reorganization of the actincytoskeleton through their regulatory effects on Rho signalling[42–44,51] and the CaR itself initiates cytoskeletal changesthrough Rho kinase [53]. This raises the possibility that the 14-3-3 association with the CaR may be required for inducing thesecytoskeletal changes, a subject worthy of further investigation.In conclusion, the present study demonstrates that two 14-3-3isoforms, 14-3-3θ and 14-3-3ζ , bind to the membrane-proximalregion of the CaR tail, and, when overexpressed, selectivelyattenuate CaR-mediated SRE activation via the Rho pathway.In addition, 14-3-3ζ reduces plasma membrane expression of theCaR.
AUTHOR CONTRIBUTION
Bryan Ward and Thomas Ratajczak devised and supervised the project with inputfrom Ajanthy Arulpragasam, Evan Ingley and Arthur Conigrave. Experimental work wasperformed largely by Ajanthy Arulpragasam with input from Aaron Magno. Evan Ingleyamplified the cDNA library used for the yeast two-hybrid screen. Suzanne Brown performedthe statistical analyses. The paper was written by Bryan Ward with suggested commentsfrom all other authors.
ACKNOWLEDGEMENTS
We gratefully acknowledge staff at the Centre for Microscopy, Characterisation andAnalysis, University of Western Australia, for help with confocal microscopy experiments.
c© The Authors Journal compilation c© 2012 Biochemical Society

14-3-3 modulates calcium-sensing receptor signalling 1005
FUNDING
This research was supported by the National Health and Medical Research Council ofAustralia through Dora Lush Postgraduate Biomedical Research Scholarships [numbers463915 (to A. A.) and 404157 (to A. L. M.)], the Raine Foundation for Medical Research,the Sir Charles Gairdner Hospital Research Fund, the University of Western AustraliaResearch Grants Scheme and Kidney Health Australia Postgraduate Scholarship [numberB2 050607 (to A. L. M.)].
REFERENCES
1 Brown, E. M. and MacLeod, R. J. (2001) Extracellular calcium sensing and extracellularcalcium signaling. Physiol. Rev. 81, 239–297
2 Magno, A. L., Ward, B. K. and Ratajczak, T. (2011) The calcium-sensing receptor: amolecular perspective. Endocr. Rev. 32, 3–30
3 Garrett, J. E., Capuano, I. V., Hammerland, L. G., Hung, B. C., Brown, E. M., Hebert, S. C.,Nemeth, E. F. and Fuller, F. (1995) Molecular cloning and functional expression of humanparathyroid calcium receptor cDNAs. J. Biol. Chem. 270, 12919–12925
4 Brennan, S. C. and Conigrave, A. D. (2009) Regulation of cellular signal transductionpathways by the extracellular calcium-sensing receptor. Curr. Pharm. Biotechnol. 10,270–281
5 Conigrave, A. D., Quinn, S. J. and Brown, E. M. (2000) Cooperative multi-modal sensingand therapeutic implications of the extracellular Ca2 + sensing receptor. TrendsPharmacol. Sci. 21, 401–407
6 Jianxin, H. (2008) Allosteric modulators of the human calcium-sensing receptor:structures, sites of action, and therapeutic potentials. Endocr., Metab. Immune Disord.:Drug Targets 8, 192–197
7 Ward, D. T. (2004) Calcium receptor-mediated intracellular signalling. Cell Calcium 35,217–228
8 Pi, M., Spurney, R.F., Tu, Q., Hinson, T. and Quarles, L. D. (2002) Calcium-sensingreceptor activation of rho involves filamin and rho-guanine nucleotide exchange factor.Endocrinology 143, 3830–3838
9 Huang, C., Hujer, K. M., Wu, Z. and Miller, R. T. (2004) The Ca2 + -sensing receptorcouples to Gα12/13 to activate phospholipase D in Madin–Darby canine kidney cells. Am.J. Physiol. Cell Physiol. 286, C22–C30
10 Rey, O., Young, S. H., Yuan, J., Slice, L. and Rozengurt, E. (2005) Amino acid-stimulatedCa2 + oscillations produced by the Ca2 + -sensing receptor are mediated by aphospholipase C/inositol 1,4,5-triphosphate-independent pathway that requires G12, rho,filamin-A, and the actin cytoskeleton. J. Biol. Chem. 280, 22875–22882
11 Ray, K., Fan, G. F., Goldsmith, P. K. and Spiegel, A. M. (1997) The carboxyl terminus ofthe human calcium receptor. Requirements for cell-surface expression and signaltransduction. J. Biol. Chem. 272, 31355–31361
12 Bai, M., Trivedi, S. and Brown, E. M. (1998) Dimerization of the extracellularcalcium-sensing receptor (CaR) on the cell surface of CaR-transfected HEK293 cells.J. Biol. Chem. 273, 23605–23610
13 Lienhardt, A., Garabedian, M., Bai, M., Sinding, C., Zhang, Z., Lagarde, J., Boulesteix, J.,Rigaud, M., Brown, E. M. and Kottler, M. (2000) A large homozygous or heterozygousin-frame deletion within the calcium-sensing receptor’s carboxyl terminal cytoplasmic tailthat causes autosomal dominant hypocalcemia. J. Clin. Endocrinol. Metab. 85,1695–1702
14 Bai, M., Trivedi, S., Lane, C. R., Yang, Y., Quinn, S. J. and Brown, E. M. (1998) Proteinkinase C phosphorylation of threonine at position 888 in Ca2 +
o-sensing receptor (CaR)inhibits coupling to Ca2 + store release. J. Biol. Chem. 273, 21267–21275
15 Zerangue, N., Schwappach, B., Jan, Y. N. and Jan, L. Y. (1999) A new ER trafficking signalregulates the subunit stoichiometry of plasma membrane KATP channels. Neuron 22,537–548
16 Stepanchick, A., McKenna, J., McGovern, O., Huang, Y. and Breitwieser, G. E. (2010)Calcium sensing receptor mutations implicated in pancreatitis and idiopathic epilepsysyndrome disrupt an arginine-rich retention motif. Cell. Physiol. Biochem. 26,363–374
17 Awata, H., Huang, C., Handlogten, M. E. and Miller, R. T. (2001) Interaction of thecalcium-sensing receptor and filamin, a potential scaffolding protein. J. Biol. Chem. 276,34871–34879
18 Hjalm, G., MacLeod, R. J., Kifor, O., Chattopadhyay, N. and Brown, E. M. (2001)Filamin-A binds to the carboxyl-terminal tail of the calcium-sensing receptor, aninteraction that participates in CaR-mediated activation of mitogen-activated proteinkinase. J. Biol. Chem. 276, 34880–34887
19 Gorlin, J. B., Yamin, R., Egan, S., Stewart, M., Stossel, T. P., Kwiatkowski, D. J. andHartwig, J. H. (1990) Human endothelial actin-binding protein (ABP-280, nonmusclefilamin): a molecular leaf spring. J. Cell Biol. 111, 1089–1105
20 Huang, C., Sindic, A., Hill, C. E., Hujer, K. M., Chan, K. W., Sassen, M., Wu, Z., Kurachi,Y., Nielsen, S., Romero, M. F. and Miller, R. T. (2007) Interaction of the Ca2 + -sensingreceptor with the inwardly rectifying potassium channels Kir4.1 and Kir4.2 results ininhibition of channel function. Am. J. Physiol. Renal Physiol. 292, F1073–F1081
21 Stepanchick, A. and Breitwieser, G. E. (2010) The cargo receptor p24A facilitates calciumsensing receptor maturation and stabilization in the early secretory pathway. Biochem.Biophys. Res. Commun. 395, 136–140
22 Huang, Y., Niwa, J., Sobue, G. and Breitwieser, G. E. (2006) Calcium-sensing receptorubiquitination and degradation mediated by the E3 ubiquitin ligase dorfin. J. Biol.Chem.281, 11610–11617
23 Herrera-Vigenor, F., Hernandez-Garcıa, R., Valadez-Sanchez, M., Vazquez-Prado, J. andReyes-Cruz, G. (2006) AMSH regulates calcium-sensing receptor signaling throughdirect interactions. Biochem. Biophys. Res. Commun. 347, 924–930
24 Tsai, S., Bartelmez, S., Sitnicka, E. and Collins, S. (1994) Lymphohematopoieticprogenitors immortalized by a retroviral vector harboring a dominant-negative retinoicacid receptor can recapitulate lymphoid, myeloid, and erythroid development. Genes Dev.8, 2831–2841
25 Ingley, E., Williams, J. H., Walker, C. E., Tsai, S., Colley, S., Sayer, M. S., Tilbrook, P. A.,Sarna, M., Beaumont, J. G. and Klinken, S. P. (1999) A novel ADP-ribosylation like factor(ARL-6) interacts with the protein-conducting channel SEC61B subunit. FEBS Lett. 459,69–74
26 Lim, R., Winteringham, L. N., Williams, J. H., McCulloch, R. K., Ingley, I., Tiao, J. Y.,Lalonde, J., Tsai, S., Tilbrook, P. A., Sun, Y. et al. (2002) MADM, a novel adaptor proteinthat mediates phosphorylation of the 14-3-3 binding site of myeloid leukemia factor 1.J. Biol. Chem. 277, 40997–41008
27 Fu, H., Subramanian, R. R. and Masters, S. C. (2000) 14-3-3 proteins: structure, function,and regulation. Annu. Rev. Pharmacol. Toxicol. 40, 617–647
28 Aitken, A. (2006) 14-3-3 proteins: a historic overview. Sem. Cancer Biol. 16, 162–17229 Vojtek, A., Hollenberg, S. M. and Cooper, J. A. (1993) Mammalian Ras interacts directly
with the serine/threonine kinase Raf. Cell 74, 205–21430 Ward, B. K., Magno, A. L., Davis, E. A., Hanyaloglu, A. C., Stuckey, B. G. A., Burrows, M.,
Eidne, K. A., Charles, A. K. and Ratajczak, T. (2004) Functional deletion of thecalcium-sensing receptor in a case of neonatal severe hyperparathyroidism. J. Clin.Endocrinol. Metab. 89, 3721–3730
31 Carrello, A., Ingley, E., Minchin, R. F., Tsai, S. and Ratajczak, T. (1999) The commontetratricopeptide repeat acceptor site for steroid receptor-associated immunophilins andHop is located in the dimerization domain of Hsp90. J. Biol. Chem. 274, 2682–2689
32 Conigrave, A. D., Quinn, S. J. and Brown, E. M. (2000) L-Amino acid sensing by theextracellular Ca2 + -sensing receptor. Proc. Natl. Acad. Sci. U.S.A. 97, 4814–4819
33 Ward, B. K., Allan, R. K., Mok, D., Temple, S. E., Taylor, P., Dornan, J., Mark, P. J., Shaw,D. J., Kumar, P., Walkinshaw, M. D. and Ratajczak, T. (2002) A structure-based mutationalanalysis of cyclophilin 40 identifies key residues in the core tetratricopeptide repeatdomain that mediate binding to Hsp90. J. Biol. Chem. 277, 40799–40809
34 Lee, H. J., Mun, H., Lewis, N. C., Crouch, M. F., Culverston, E. L., Mason, R. S. andConigrave, A. D. (2007) Allosteric activation of the extracellular Ca2 + -sensing receptorby L-amino acids enhances ERK1/2 phosphorylation. Biochem. J. 404, 141–149
35 Tan, Y. M., Cardinal, J., Franks, A. H., Mun, H., Lewis, N., Harris, L. B., Prins, J. B. andConigrave, A. D. (2003) Autosomal dominant hypocalcemia: a novel activating mutation(E604K) in the cysteine-rich domain of the calcium-sensing receptor. J. Clin. Endocrinol.Metab. 88, 605–610
36 Shikano, S., Coblitz, B., Wu, M. and Li, M. (2006) 14-3-3 proteins: regulation ofendoplasmic reticulum localization and surface expression of membrane proteins. TrendsCell Biol. 16, 370–375
37 O’Kelly, I., Butler, M. H., Zilverberg, N. and Goldstein, S. A. N. (2002) Forward transport.14-3-3 binding overcomes retention in endoplasmic reticulum by dibasic signals. Cell111, 577–588
38 Yuan, H., Michelsen, K. and Schwappach, B. (2003) 14-3-3 dimers probe the assemblystatus of multimeric membrane proteins. Curr. Biol. 13, 638–646
39 Zhang, M. and Breitwieser, G. E. (2005) High affinity interaction with filamin A protectsagainst calcium-sensing receptor degradation. J. Biol. Chem. 280, 11140–11146
40 Fantl, W. J., Muslin, A. J., Kikuchi, A., Martin, J. A., MacNicol, A. M., Grosst, R. W. andWilliams, L. T. (1994) Activation of Raf-1 by 14-3-3 proteins. Nature 371, 612–614
41 Cunningham, C. C., Gorlin, J. B., Kwiatkowski, D. J., Hartwig, J. H., Janmey, P. A., Byers,H. R. and Stossel, T. P. (1992) Actin-binding protein requirement for cortical stability andefficient locomotion. Science 255, 325–327
42 Gohla, A. and Bokoch, G. M. (2002) 14-3-3 regulates actin dynamics by stabilizingphosphorylated cofilin. Curr. Biol. 12, 1704–1710
43 Bialkowska, K., Zaffran, Y., Meyer, S. C. and Fox, J. E. B. (2003) 14-3-3ζ mediatesintegrin-induced activation of Cdc42 and Rac. J. Biol. Chem. 278, 33342–33350
44 Diviani, D., Abuin, L., Cotecchia, S. and Pansier, L. (2004) Anchoring of both PKA and14-3-3 inhibits the Rho-GEF activity of the AKAP–Lbc signaling complex. EMBO J. 23,2811–2820
45 Wilker, E. W., Grant, R. A., Artim, S. C. and Yaffe, M. B. (2005) A structural basis for14-3-3σ functional specificity. J. Biol. Chem. 280, 18891–18898
46 Perego, L. and Berruti, G. (1997) Molecular cloning and tissue-specific expression of themouse homologue of the rat brain 14-3-3θ protein: characterization of its cellular anddevelopmental pattern of expression in the male germ line. Mol. Reprod. Dev. 47,370–379
47 Wong, E. W. P., Sun, S., Li, M. W. M., Lee, W. M. and Cheng, C. Y. (2009) 14-3-3 proteinregulates cell adhesion in the seminiferous epithelium of rat testes. Endocrinology 150,4713–4723
c© The Authors Journal compilation c© 2012 Biochemical Society

1006 A. Arulpragasam and others
48 Tfelt-Hansen, J., Schwarz, P., Terwilliger, E. F., Brown, E. M. and Chattopadhyay, N. (2003)Calcium-sensing receptor induces messenger ribonucleic acid of human securin pituitarytumor transforming gene, in rat testicular cancer. Endocrinology 144, 5188–5193
49 Campbell, J. K., Gurung, R., Romero, S., Speed, C. J., Andrews, R. K., Berndt, M. C. andMitchell, C. A. (1997) Activation of the 43 kDa inositol polyphosphate 5-phosphatase by14-3-3ζ . Biochemistry 36, 15363–15370
50 Couve, A., Kittler, J. T., Uren, J. M., Calver, A. R., Pangalos, M. N., Walsh, F. S. and Moss,S. J. (2001) Association of GABA(B) receptors and members of the 14-3-3 family ofsignaling proteins. Mol. Cell. Neurosci. 17, 317–328
51 Meiri, D., Greeve, M. A., Brunet, A., Finan, D., Wells, C. D., LaRose, J. and Rottapel, R.(2009) Modulation of Rho guanine exchange factor Lfc activity by protein kinaseA-mediated phosphorylation. Mol. Cell. Biol. 29, 5963–5973
52 Nurmi, S. M., Gahmberg, C. G. and Fagerholm, S. C. (2006) 14-3-3 proteins bind bothfilamin and αaLβ2 integrin in activated T cells. Annals N.Y. Acad. Sci. 1090, 318–325
53 Davies, S. L., Gibbons, C. E., Vizard, T. and Ward, D. T. (2006) Ca2 + -sensing receptorinduces Rho kinase-mediated actin stress fiber assembly and altered cell morphology, butnot in response to aromatic amino acids. Am. J. Physiol. Cell Physiol. 290,C1543–C1551
Received 18 July 2011/3 October 2011; accepted 19 October 2011Published as BJ Immediate Publication 19 October 2011, doi:10.1042/BJ20111277
c© The Authors Journal compilation c© 2012 Biochemical Society

Biochem. J. (2012) 441, 995–1006 (Printed in Great Britain) doi:10.1042/BJ20111277
SUPPLEMENTARY ONLINE DATAThe adaptor protein 14-3-3 binds to the calcium-sensitive receptor andattenuates receptor-mediated Rho kinase signallingAjanthy ARULPRAGASAM*†‡, Aaron L. MAGNO*†‡, Evan INGLEY*†, Suzanne J. BROWN‡, Arthur D. CONIGRAVE§,Thomas RATAJCZAK*†‡ and Bryan K. WARD*†‡1
*Western Australian Institute for Medical Research, QEII Medical Centre, Hospital Avenue, Nedlands, WA 6009, Australia, †University of Western Australia Centre for Medical Research,University of Western Australia, Nedlands, WA 6009, Australia, ‡Department of Endocrinology and Diabetes, Sir Charles Gairdner Hospital, Hospital Avenue, Nedlands, WA 6009,Australia, and §School of Molecular Bioscience, University of Sydney, Sydney, NSW 2000, Australia
Figure S1 Level of 14-3-3ζ knockdown achieved following transfection ofa siRNA oligonucleotide primer specific to 14-3-3ζ in HEK-293/CaR cells
HEK-293/CaR cells were used for experiments examining the effect of 14-3-3ζ depletion onCaR-mediated ERK1/2 activity (A) and CaR-mediated SRE activity (B). Protein expression wasexamined 48 or 72 h after transfection for ERK1/2 and SRE-Luc assays respectively, to coincidewith the time post-transfection that each assay was performed. Western blots (WB) wereperformed using an antibody specific against 14-3-3ζ . Control experiments used a validatednegative oligonucleotide primer.
Received 18 July 2011/3 October 2011; accepted 19 October 2011Published as BJ Immediate Publication 19 October 2011, doi:10.1042/BJ20111277
Figure S2 Levels of expression of filamin 1 in M2, A7, HEK-293 and HEK-293/CaR cells
Western blot (WB) analyses demonstrating the absence of filamin 1 in M2 compared withHEK-293 cells (A) and comparative levels of filamin 1 expression in A7 and HEK-293/CaR cells.Western blots were performed using equivalent amounts of protein [50 μg in (A) and 35 μg in(B)] in conjunction with an anti-filamin 1 monoclonal antibody.
1 To whom correspondence should be addressed (email [email protected]).
c© The Authors Journal compilation c© 2012 Biochemical Society