the activation of liver x receptors inhibits toll-like receptor-9-induced foam cell formation
TRANSCRIPT

ORIGINAL ARTICLE 158J o u r n a l o fJ o u r n a l o f
CellularPhysiologyCellularPhysiology
The Activation of Liver XReceptors Inhibits Toll-LikeReceptor-9-Induced Foam CellFormation
ROSALINDA SORRENTINO,* SILVANA MORELLO, SHUANG CHEN, EDUARDO BONAVITA,AND ALDO PINTO
Pharmaceutical Science Department, University of Salerno, 84084 Fisciano, Salerno, Italy
Toll-like receptors (TLRs) are related to foam cell formation (FCF), key event in the establishment/progression of atherosclerosis. Theactivation of TLR2 and TLR4 can increase FCF. The aim of this study was to evaluate the role of TLR9 in FCF. Murine macrophages weretreated with CpG-ODN, TLR9 agonist, and oxidized particles of LDL (Paz-PC) and FCF was analyzed by means of Oil Red O staining.The administration of CpG-ODN plus Paz-PC ontomacrophages increased the amount of lipid droplets, correlated to increased levels oftumor necrosis factor (TNF)-a, IFNb, and IP-10. The underlying mechanism bywhich TLR9 ligation influenced Paz-PC in the FCFwasNF-kB- and IRF7-dependent, as observed by higher levels of phosphorylated IkBa, increased nuclear translocation of the p65 subunit, lowerlevels of the total IKKa protein and higher release of interferon-dependent cytokines, such as IP-10. Liver X receptors (LXRs) regulatelipid cellular transport and negatively modulate TLR-dependent signaling pathways. Indeed, the addition of GW3965, synthetic LXRsagonist, significantly reduced FCF after CpG-ODN plus Paz-PC stimulation. In this condition, we observed decreased levels of the nucleartranslocation of the p65 subunit, related to the higher presence of LXRa into the nucleus. TNF-a, IP-10, and IFNb levels were reduced bythe administration of GW3965 following CpG-ODN and Paz-PC treatment. In conclusion, the activation of TLR9 facilitates the formationof foam cells in anNF-kB- and IRF7-dependent manner, countered by the activation of LXRs. This study further support LXRs as potentialanti-atherosclerotic target.J. Cell. Physiol. 223: 158–167, 2010. � 2010 Wiley-Liss, Inc.
Abbreviations: LXRs, liver X receptors; TLR9, toll-like receptor 9;FCF, foam cell formation.
This article was originally published on 4 Jan 2010. An error wassubsequently identified. Shuang Chen has been added as the thirdauthor in the corrected version published 25 January 2010.
Summary Sentence: The activation of TLR9 facilitates theFormation of Foam Cells in an NF-kB- and IRF7-dependentmanner, contrasted by the activation of LXRs.
Contract grant sponsor: University of Salerno (FARB 2007–2008).
*Correspondence to: Rosalinda Sorrentino, PharmaceuticalScience Department, University of Salerno, 84084 Fisciano,Salerno, Italy. E-mail: [email protected]
Received 9 October 2009; Accepted 12 November 2009
Published online in Wiley InterScience(www.interscience.wiley.com.), 4 January 2010.DOI: 10.1002/jcp.22022
Atherosclerosis, formerly considered a lipid accumulationvascular disease, is actually classified as a focal and chronicongoing inflammatory response (Michelsen and Arditi, 2006;Sorrentino and Arditi, 2009). The modification of lipoproteinsin the sub-endothelial matrix attracts diverse and multifactorialleukocytes, including monocytes, T cells, B cells, mast cells,dendritic cells (DCs), and neutrophils in the vessel wall(Michelsen and Arditi, 2006; Naiki et al., 2008; Sorrentino andArditi, 2009). The activation ofmonocytes in the subendothelialmatrix leads to the differentiation into macrophages that ingestmodified lipids and become foam cells (Naiki et al., 2008). Thisphenomenon promotes the overcoming release of a variety ofinflammatory mediators, with the resultant of a chronicinflammation.
Several outstanding studies suggested the relevance of Toll-like receptors (TLRs) in the atherosclerosis pathology, eitherusing human or animal models (Sorrentino and Arditi, 2009).TLRs recognize a wide range of bacterial molecular patterns,instructing the innate immune system to a specific adaptiveresponse (Michelsen and Arditi, 2006; Sorrentino and Arditi,2009). TLR4 and TLR2 participate to the induction andprogression of the atheroma (Michelsen et al., 2004; Michelsenand Arditi, 2006; Mullick et al., 2008; Sorrentino and Arditi,2009).
Foamcell formation (FCF) is a key event for the accumulationof lipids and it can be enhanced by the presence of bacteria orpathogen-associated molecular patterns (PAMPs) (Naiki et al.,2008; Sorrentino and Arditi, 2009). Indeed, C. pneumoniae,Gram-negative bacteria, can promote FCF in the presence ofoxidized low-density lipoprotein (LDL) (Chen et al., 2008). Theauthors related this effect to the recognition of the bacteriumby TLR2 and TLR4 (Naiki et al., 2008), that exacerbated theatheroma in a mouse model of atherosclerosis (Naiki et al.,2008). The progression/exacerbation of the atheroscleroticplaques was also observed when LPS (5), TLR4 ligand,Pam3CySK4 (Mullick et al., 2008), TLR2 ligand, or otherpathogens, such asH. influenzae, P. gingivalis, were administered
� 2 0 1 0 W I L E Y - L I S S , I N C .
into atherosclerotic-prone mice (Michelsen and Arditi, 2006).Hence, bacteria or bacteria-derived molecules, such as nucleicacids, could facilitate the formation of the atheroscleroticplaques. The recognition of PAMPs by antigen presenting cells(APCs) can promote the activation of cytoplasmic receptors,such as TLR9, other than the membrane TLR2 and TLR4. Thus,the main goal of our study was to evaluate the implication ofTLR9 in FCF in the presence of oxidized LDL particles.
Liver X receptors (LXRs), nuclear receptors, play a pivotalrole for lipid metabolism and are master-regulators ofcholesterol metabolism (Bensinger and Tontonoz, 2008). LXRscontrol reverse cholesterol metabolism and alsocounterbalance the pro-inflammatory effects of TLRs (Castrilloet al., 2003). Of therapeutic interest, LXRs and TLRs exert anegative cross-talk onto each others’ signaling pathway(Castrillo et al., 2003; Chen et al., 2008;Naiki et al., 2008), as forexample, in the case of TLR4 activation that is negatively

T L R 9 I N D U C E S F O A M C E L L F O R M A T I O N 159
influenced by LXRs signaling (Castrillo et al., 2003; Chen et al.,2008). This effect was ascribed to the down-regulation ofNF-kB by LXRs, that mediate an anti-inflammatory pattern,suppressing TLRs signaling pathway. On the other side, theactivation of TLRs contrast LXRs-induced anti-inflammatorymechanism (Cao et al., 2007; Chen et al., 2008; Naiki et al.,2008). Indeed, stimulation of TLR3 or TLR4 by pathogen-derived ligands inhibits expression of LXR-dependent genetargets and macrophage cholesterol efflux (Castrillo et al.,2003). The cross-talk between LXRs and TLRs signalingpathways may potentially lead to understand how microbialinfection interferes with cholesterol metabolism and then leadsto cardiovascular diseases. These concepts associate the lipidmetabolic pathways with innate immune host defenses (Naikiet al., 2008), that play a fundamental role in the development ofthe atherosclerotic plaques (Naiki et al., 2008; Sorrentino andArditi, 2009). Thus, our study aimed first to evaluate theimplication of TLR9 in the FCF and then to evidentiate anycross-talk between TLR9 and LXRs signaling pathways. Here,we demonstrate that the stimulation of TLR9 with CpG-ODNfacilitated the formation of foam cells in the presence ofoxidized species of LDL (Paz-PC) via the activation of NF-kBand IRFs-dependent pathways. Interestingly, the addition ofGW3965, LXRs agonist, led to a significant reduction of lipiddroplets into TLR9-induced foam cells via the inhibition ofNF-kB. Collectively, these results first revealed the implicationof TLR9 in the formation of foam cells, and secondly, thenegative cross-talk between TLR9 and LXRs.
Materials and MethodsCell culture and reagents
Peritoneal Macrophages were harvested by C57Bl/6 mice byperitoneal lavage (0.5mMEDTA in PBS) and cultured in RPMI 1640(Cambrex Biosciences, Microtech, Naples, Italy). Macrophagiccells, RAW264.7, were cultured in Dulbecco’s Modified EagleMedium (DMEM), supplemented of 10% fetal bovine serum (FBS),100U/ml penicillin, 100U/ml streptomycin, and 2mM L-glutamine(Cambrex Biosciences, Microtech). Cells were seeded and treatedwith CpG-ODN (1mg/ml; TLR Grade 1886, Alexis, Vincibiochem,Italy), 1-palmitoyl-2-azelaoyl PC, predominant structure ofoxidized LDL (Paz-PC; 15–25–50–100mg/ml; Cayman,Vincibiochem, Italy), GW3965 (1mM, Sigma–Aldrich, Milan, Italy),neutralizing antibody for IP-10 or tumor necrosis factor (TNF)-a(Sodium Azide free-antibody; R&D Systems, Oxford, UK).
Assessment of foam cell formation
Foam cells were identified using Oil Red O staining. Peritonealmacrophages and RAW264.7 were washed twice with PBS, fixedand stained with 1% of Oil Red O solution (60% isopropanol;Sigma–Aldrich). Cells were washed three times with PBS andexamined by light microscopy (40� to 63�). Designation of amacrophage as a foam cell required-positive Oil Red O staining.Foam cells were quantified by counting the presence of lipiddroplets into the cells as identified by Oil Red O staining. Eachtreatment condition was performed in triplicate.
ELISA assays
The levels of TNF-a, IP-10/chemokine ligand 10 (CXCL10) in thecell-free supernatants were measured by means of ELISA usingcommercially availablematched antibody pairs following a protocolsupplied by the manufacturers (eBioscience, San Diego, CA; R&DSystems).
Immunoblot assay
Cells were plated at a density of 1� 106/ml in normal cultureconditions and treated as previously described. Laemmly buffersamples were loaded onto 8–12% denaturing polyacrylamide gels.
JOURNAL OF CELLULAR PHYSIOLOGY
Proteins were detected with appropriate dilutions of the followingprimary antibodies: anti-p-IkBa, anti-IkBa, and anti-IKKa (SantaCruz Biotechnology, DBA, Milan, Italy). Samples for p-IkBadetection derived from FBS-deprived cultured cells.Immunoreactive proteins were visualized by enhancedchemiluminiscence (Amersham Pharmacia Biotech,Birminghamshire, UK) and quantified by densitometry analysis(GelDoc Instrument, Bio-Rad, Milan, Italy).
Immunofluorescence technique
RAW264.7 cells were seeded (105 cells/well) 24 h before anytreatment. Paz-PC (25mg/ml) and/or CpG-ODN (1mg/ml) wereadded in a time-dependent manner, 15min up to 3 h. Cells werefixed with para-formaldehyde (4%, v/v) for 15min, permeabilizedwith PBS/Triton 0.05% for 10min and then incubated with rabbitanti-p65/Rel-A (C-20), rabbit anti-c-Rel (N-466) or goat anti-LXRa(Santa Cruz Biotechnology). Anti-rabbit or anti-goat Texas Red(Santa Cruz Biotechnology) was used as a secondary antibody andincubated for 1–2 h at room temperature. Isotype control (rabbitIgG, Santa Cruz Biotechnology) was used. Nuclear translocation ofthe above proteins was analyzed by using a fluorescencemicroscope (Carl Zeiss, Munich, Germany) by means of AxioplanImaging Programme (Carl Zeiss Axiocam Programme).
Transcriptional regulation of NF-kB
The transcriptional activity of NF-kB was evaluated by means ofTransAMTM Transcription Factor ELISAs Assay (Active Motif,Cambridge, UK). RAW 264.7 cells (1� 106 cells/well) were firsttreated with CpG and/or Paz-PC for 1 h and half, and then digestedto allow the separation of nuclear proteins from the cytoplasmicproteins. The ELISA-based assay was then performed following themanufacturer’s instructions.
Real-time polymerase chain reaction analysis
RAW264.7 cells were seeded in 35 cm2 plates, harvested and totalRNAwas isolated by using RNASPINMINI extraction kit accordingto the manufacturer’s instructions (GE Healthcare, Milan, Italy).Reverse transcription was performed by using first-strand cDNAsynthesis kit (GE Healthcare) followed by PCR. Thermal cyclingconditions were 15min at 958C, followed by 35 cycles of 45 sec at948C, 25 sec at 558C, 30 sec at 728C. Primer pairs were as follow:
LXRa—Forward: 50-CTCACCTCCTCAAGTCCTGC-30;Reverse: 50-GCTGTTTGGCAAAGTCAACA-30.ABCA 1—Forward: 50-CAGACCTGTAAAGGCGAAGC-30;Reverse: 50-GTCACAGGGGTGTCTCCAGT-30.IFNb—Forward: 50-CCCTATGGAGATGACGGAGA-30;Reverse: 50-AGTTGAGGACATCTCCCACG-30.b-Actin—Forward: 50-AGCCATGTACGTAGCCATCC-30;Reverse: 50-CTCTCAGCTGTGGTGGTGAA-30.
Statistical analysis
Data are reported as mean� SEM. Statistical differences wereassessed by ONE-Way Anova analysis of variance and Student’s t-test, and P values <0.05 were considered significant.
ResultsThe activation of TLR9 promotes the formation offoam cells
Accumulation of lipids leads macrophages phenotype towardsfoam cells in atherosclerosis (Chen et al., 2008). The activationof TLR2 and TLR4 was involved in FCF (Cao et al., 2007; Chenet al., 2008; Naiki et al., 2008). In order to evaluate whetherTLR9 activation was also implicated in FCF, we treatedRAW264.7 cells with Paz-PC (15–25–50–100mg/ml),predominant oxidized lipoprotein (ox-LDL) species (Podrez etal., 2000, 2002), in the presence or not of CpG-ODN (CpG,

Fig. 1. Murine macrophages (RAW264.7) were treated withmedium alone, Paz-PC (25mg/ml), CpG-ODN (1mg/ml), or bothreagents.Oil RedOstainingwasperformed.Theamountof foamcellswas reported as lipid droplets/field and derives from the count of atleast five fields of the same slide (A). CpG and Paz-PC increased theamountof lipiddroplets comparedwithPaz-PCorCpGalone (B).LPSplus Paz-PC was used as a positive control (C). Experiments wereperformed in duplicate for each treatment. Data representmeanWSEM and each experiment was performed at least five timesfor each type of experimental condition. Statistically significantdifferences are MP<0.05, MMP<0.005, MMMP<0.0005, and MMMMP<0.0001as determined by One-Way Anova analysis of variance and Student’st-test. [Colorfigurecanbeviewed in theonline issue,which is availableat www.interscience.wiley.com.]
160 S O R R E N T I N O E T A L .
type B; 1mg/ml), TLR9 agonist (Kawai and Akira, 2008).Preliminary experiments showed 25mg/ml of Paz-PC as thesub-maximal concentration to study the induction of foam cells.
The addition for 24 h of Paz-PC (25mg/ml) or CpG aloneincreased the amount of lipid droplets in RAW264.7 cells(Fig. 1A,B, ��P< 0.005 and ���P< 0.0005) compared with themedium alone (Fig. 1A,B). In contrast, the co-administration ofCpG and Paz-PC highly increased the formation of foam cellsfrom RAW264.7 cells (Fig. 1A,B, ����P< 0.0001). To confirmthese results, we isolated peritoneal macrophages fromC57Bl/6j mice and treated the cells. We observed the same effect onthe primary cells (data not shown).
LPS (0.1mg/ml), TLR4 agonist, or Pam3CSK4 (1mg/ml),TLR2 agonist, were used as positive controls. Paz-PC and LPS(Fig. 1C, P< 0.0005) or Pam3CSK4 (data not shown) increasedthe formation of foam cells.
None of the treatments altered the cell viability as tested bythe MTT assay (data not shown).
TLR9-induced foam cell formation was mediated viaNF-kB activation
TLR9-mediated signaling pathway leads to the activation ofNF-kB and interferon (IFN) regulatory factor (IRF)-7 (Kawaiand Akira, 2008). In order to understand whether the TLR9-induced formation of foam cells was NF-kB-dependent, weanalyzed the nuclear localization of p65 and/or c-Rel subunit in a
JOURNAL OF CELLULAR PHYSIOLOGY
time-dependent manner (15min up to 3 h) by means ofimmunofluorescence technique. The addition of Paz-PC did notlead to the nuclear translocation of p65 or c-Rel subunit (Fig. 2Aand data not shown, respectively), in contrast to CpG (Fig. 2Aand data not shown). The co-administration ofCpG and Paz-PCinduced the translocation of p65 subunit into the nucleus ofRAW 264.7 cells (Fig. 2A). The same results were obtained bymonitoring the nuclear localization of c-Rel (data not shown).
In order to investigate the upstream signaling that led toNF-kB activation, wewent on by analyzing the phosphorylationof IkBa and the levels of IKKa by means of Western blotting.The complex IKKa/IKKb promotes IkBa phosphorylation,thus enabling NF-kB to migrate into the nucleus (Bonizzi andKarin, 2004). As observed in Figure 2B, CpG alone increasedthe phosphorylation of IkBa (�P< 0.05), implying the nucleartranslocation of NF-kB, and confirming theimmunofluorescence experiments. The co-administration ofCpG and Paz-PC further increased the phosphorylation of IkBa(Fig. 2B, ��P< 0.001). In order to corroborate these results andto quantitatively distinguish whether the activation of NF-kBwas solely mediated by CpG, we went on to measure thetranscriptional regulation by means of a TransAM kit. Theseexperiments showed that the association of CpG and Paz-PCsignificantly increased the ability of the p65 subunit to bind aspecific nucleic sequence, compared with CpG alone(Fig. 2C, �P< 0.05; ��P< 0.001). Therewas a twofold increase inthe levels of the nuclear p65 activity when both agents wereadded compared with the addition of CpG or Paz-PC alone(Fig. 2C). Taken together these results led us to suppose thatTLR9-induced FCF was related to NF-kB activation.
However, it is well known that TLR9-induced signalingpathway, other than NF-kB, can also promote the activation ofIRF7, that in turn induces the synthesis/release of IFN type I(Kawai and Akira, 2008). The upstream signaling of IRF7activation can derive from IKKa (14), which results activated inits phosphorylated form (Heyninck and Beyaert, 2005). Theaddition of CpG to Paz-PC reduced the amount of total IKKa inRAW264.7 cells, in contrast to Paz-PC or medium alone(Fig. 2D; �P< 0.05). A reduction of IKKa levels led us tosuppose that kinase was phosphorylated and implicated inTLR9-mediated FCF, ‘‘collaborating’’ with the ‘‘canonical’’NF-kB pathway (Kawai and Akira, 2008).
Pro-inflammatory cytokines were involved inTLR9-induced foam cell formation
The activation of the ‘‘canonical’’ NF-kB pathway leads tothe release of pro-inflammatory cytokines, such as TNF-a(Bonizzi and Karin, 2004; Naiki et al., 2008); whereas, the‘‘alternative’’ NF-kB activation pathway, which can activateIRF7 in an IKK-a-dependent manner (Kawai and Akira, 2008;Kaisho and Tanaka, 2008), leads to the production of IFNtype I (Kaisho and Tanaka, 2008). In this context, we assessedthe release of TNF-a and IP-10 (CXCL10), IFN-derivedcytokine (Kawai and Akira, 2008), during the formation of foamcells.
As expected, the addition of CpG onto RAW264.7 cellsincreased the release of TNF-a (Fig. 3A; �P< 0.05), but theco-administration with Paz-PC further enhanced the release ofthis pro-atherogenic cytokine (Fig. 3A; P< 0.005). Paz-PC alonedid not significantly alter TNF-a levels compared with themedium alone (Fig. 3A). These data confirm the hypothesesabout the implication of the ‘‘canonical’’ NF-kB activationpathway during the formation of foam cells after TLR9stimulation, since TNF-a can be considered an NF-kB-dependent cytokine.
Figure 3B shows that CpG plus Paz-PC enhanced the levels ofIP-10 compared with the medium alone (��P< 0.01). We didnot observe statistical differences between the levels of IP-10

Fig. 2. Nuclear translocationofp65/RelAsubunit1 handhalf aftertheadditionofCpG(1mg/ml)and/orPaz-PC(25mg/ml) (A).CpGandCpGplusPaz-PC increased the translocation of p65 to the nucleus at this time point (A) as demonstrated by the merge of Texas Red and DAPI-positivestainings. Rabbit IgGwas used as negative control (data not shown). Experimentswere performed in duplicate in three different time conditions.ImmunoblotanalysisofpIkBa(B)andIKKa(D).Thebargraphrepresenttheopticaldensity(OD)ratiobetweenthesignaldetectedforpIkBa(B)orIKKa (D) compared with the IkBa or Tubulin (loading control), respectively. Statistically significant differences are MP<0.05, and MMP<0.001 asdetermined by Student’s t-test. Part C: The nuclear transcription of the p65 subunit. The addition of CpG and Paz-PC on RAW264.7 cellssignificantly increased the activity ofNF-kB (C). Statistically significant differences are MP<0.05, and MMP<0.01 as determined by Student’s t-test.DatarepresentmeanWSEMandeachexperimentwasperformedat leastthreetimesforeachtypeofexperimentalcondition.[Colorfigurecanbeviewed in the online issue, which is available at www.interscience.wiley.com.]
T L R 9 I N D U C E S F O A M C E L L F O R M A T I O N 161
induced by the stimulation with CpG alone or in combinationwith Paz-PC (Fig. 3B). Paz-PC alone slightly increased IP-10release compared with medium alone (Fig. 3B; �P< 0.05). Tofurther confirm the implication of IRF7-dependent signalingpathway, we went on to analyze the mRNA expression of IFNbvia real-time PCR. CpG or Paz-PC alone tended to increase
JOURNAL OF CELLULAR PHYSIOLOGY
IFNb expression (Fig. 3C), in contrast to CpG and Paz-PC thatnoticeably augmented IFNb mRNA expression in RAW264.7cells (Fig. 3C; �P< 0.05, ��P< 0.01).
To understand the cross-talk between the release of TNF-aand IP-10, we treated the cells with a neutralizing antibody forTNF-a or IP-10, respectively. The addition of anti-IP-10

Fig. 3. Assessment of TNF-a (A) and IP-10 (B) by means of ELISA.RAW264.7 cells were treated with CpG (1mg/ml) and/or Paz-PC(25mg/ml) in the presence or not of neutralizing antibody for IP-10(anti-IP-10) (A) and for TNF-a (anti-TNF-a) (B). The release of TNF-aand IP-10 was significantly increased post the administration of CpGand Paz-PC (white bars), but the addition of neutralizing Abs forTNF-a (A, black bars) or IP-10 (B, black bars) decreased the cytokineproduction and in both type of experiments (A,B, black bars). IgG wasused as negative control for the neutralizing antibodies and did notalter the level of cytokine production, resulting similar to the controlvalues (A,B, white bars). Assessment of IFNb mRNA expression bymeansofreal-timePCR(C).Theco-administrationofCpGandPaz-PCsignificantly (MP<0.01) increased the production of IFNb (C). Datarepresent meanWSEM and each experiment was performed at leastthree times for each type of experimental condition. Statisticallysignificant differences are MP<0.05, MMP<0.01, and MMMP<0.005 asdetermined by One-Way Anova analysis of variance andStudent’s t-test.
162 S O R R E N T I N O E T A L .
(Fig. 3A; �P< 0.05) or anti-TNF-a (Fig. 3B; ��P< 0.01)decreased the release of TNF-a or IP-10, respectively, whenCpG and Paz-PC were co-administered. The same effect wasobserved for CpG or Paz-PC alone (Fig. 3A,B; �P< 0.05). These
JOURNAL OF CELLULAR PHYSIOLOGY
data led to think that both signaling pathways (NF-kB- andIRF7-dependent) collaborated for the induction of foam cellsfrom macrophages.
LXRs agonist (GW3965) reduced TLR9-inducedformation of foam cells
LXR activation interferes with TLRs signaling pathway byreducing the stimulation of NF-kB (Castrillo et al., 2003;Bensinger and Tontonoz, 2008; Chen et al., 2008). LXRs canmodulate TLR4- and TLR2-dependent signaling pathway (Chenet al., 2008).We thereforewanted to investigatewhether LXRsagonist could modulate the formation of foam cells induced byCpG in the presence of the oxidized species of LDL, Paz-PC.
The addition of GW3965 (1mM), synthetic LXRs agonist,did not alter the amount of lipid droplets when Paz-PC wasadministered alone in RAW264.7 cells (Fig. 4A,B) and inperitoneal macrophages (data not shown); whereas, itsignificantly reduced FCF when CpGwas co-administered withPaz-PC (Fig. 4A,B; ����P< 0.0001), compared with CpG plusPaz-PC (Fig. 4A,B; �P< 0.05).
To better understand whether this effect was strictlymediated by LXRs signaling pathway, we went on by analyzing adirect target implicated in this pathway, ABCA-1, membraneprotein responsible for the efflux of lipids out of the cells(Castrillo et al., 2003; Bensinger and Tontonoz, 2008). Asshown in Figure 4C, the stimulation of RAW264.7 cells withGW3965 increased ABCA-1 mRNA expression(���P< 0.0005) compared with control medium, implying thatthis protein is directly correlated to the LXRs signaling, even atthe basal levels. These results were already reported byCastrillo et al. (2003), and imply a major efflux of lipids out ofthe cells when LXRs pathway is induced. The administration ofCpG and/or Paz-PC with GW3965 did not show any differencein ABCA-1 mRNA compared with the GW3965 alone(Fig. 4C, black bars), although these levels of mRNA were stillhigher than the correspondent values when cells were treatedonly with CpG and/or Paz-PC (Fig. 4C, white bars; �P< 0.05).
To evaluate the selective activity of GW3965, we went on toanalyze the nuclear localization of LXRa, inducible isoform,mainly implicated in inflammatory conditions (Bensinger andTontonoz, 2008; Castrillo et al., 2003). Cells were treated withPaz-PC and/or CpG for 1 h and half. Figure 5A shows that thestimulation of cells with GW3965 and especially with GW3965and CpG plus Paz-PC increased the nuclear presence of LXRa.Thesameexperimentsweredoneatdifferent timepoints (15minupto3 h).Wecouldnotdetectanypositivestainingatearlier timepoints such as 15–30min or later than 2 h (data not shown).
LXRs agonist, GW3965, prevents TLR9-induced foamcell formation by blocking NF-kB activation
Castrillo et al. (2003) described that LXRs could explicate anti-inflammatory activity by inhibiting the activation of NF-kB(Castrillo et al., 2003; Bensinger and Tontonoz, 2008). Since weobserved that GW3965 reduced TLR9-mediated FCF, wewanted to evaluate whether LXRs signaling could interferewithNF-kB activation. RAW264.7 cells were treated with CpG and/or Paz-PC in the presence or not of GW3965. The addition ofGW3965 countered the nuclear translocation of the p65subunit either when CpG alone or in combination with Paz-PCwas added (Fig. 5B). In order to confirm the implication ofNF-kB, cells were harvested for TransAM assay after thetreatments as above. The addition of CpG and Paz-PC toGW3965 noticeably decreased the nuclear translocation of p65subunits (Fig. 5C; ��P< 0.01).
Interestingly, the addition of GW3965 significantlydecreased the release of TNF-a but also of IP-10whenCpGandPaz-PC were co-administered (Fig. 6A,B; �P< 0.05). However,
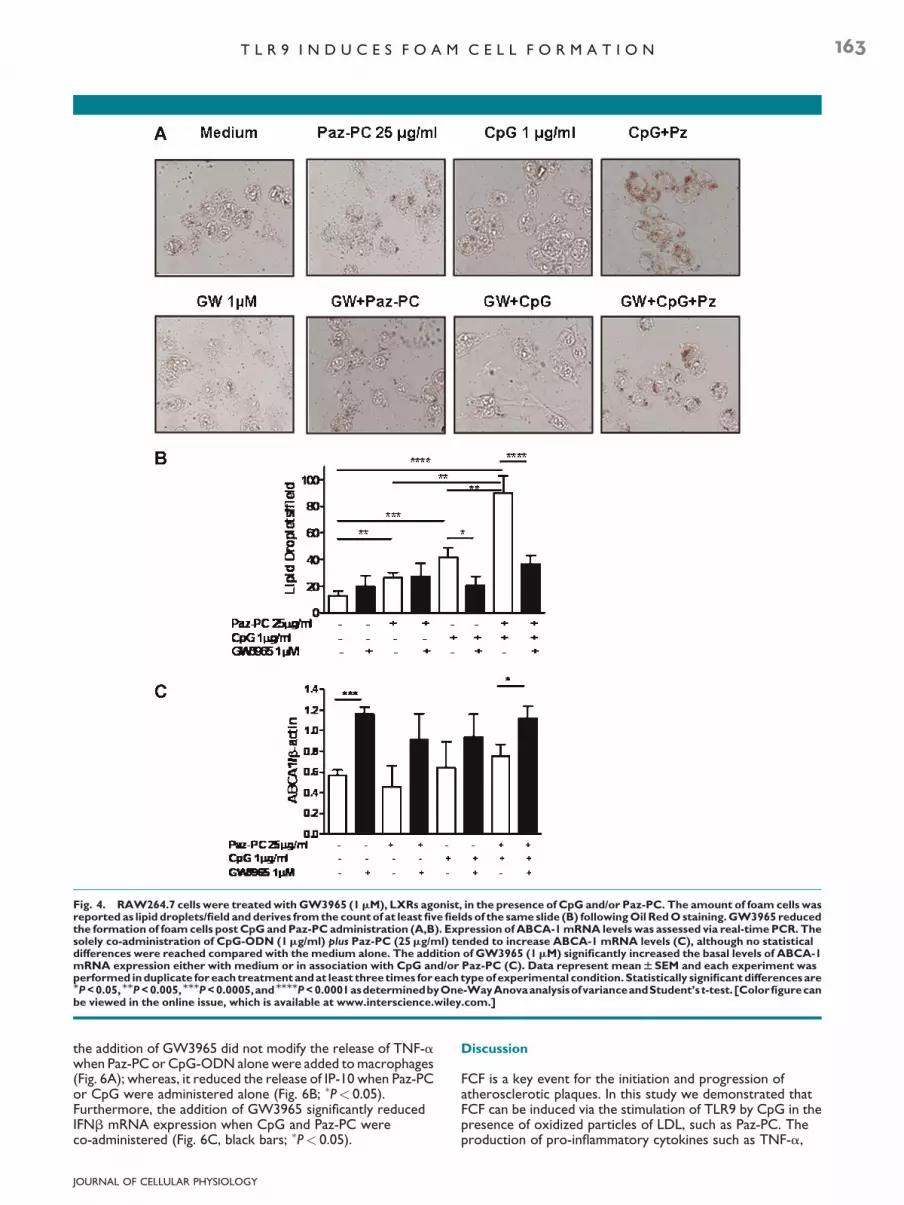
Fig. 4. RAW264.7 cells were treatedwithGW3965 (1mM), LXRs agonist, in the presence of CpG and/or Paz-PC. The amount of foam cells wasreported as lipid droplets/field andderives fromthe countof at least fivefields of the sameslide (B) followingOil RedOstaining.GW3965 reducedthe formation of foamcells post CpGandPaz-PC administration (A,B). Expression of ABCA-1mRNA levels was assessed via real-timePCR. Thesolely co-administration of CpG-ODN (1mg/ml) plus Paz-PC (25mg/ml) tended to increase ABCA-1 mRNA levels (C), although no statisticaldifferences were reached compared with the medium alone. The addition of GW3965 (1mM) significantly increased the basal levels of ABCA-1mRNA expression either with medium or in association with CpG and/or Paz-PC (C). Data represent meanWSEM and each experiment wasperformed induplicate foreach treatmentandat least three times foreach typeof experimental condition.Statistically significantdifferencesareMP<0.05,MMP<0.005,MMMP<0.0005,andMMMMP<0.0001asdeterminedbyOne-WayAnovaanalysisofvarianceandStudent’s t-test.[Colorfigurecanbe viewed in the online issue, which is available at www.interscience.wiley.com.]
T L R 9 I N D U C E S F O A M C E L L F O R M A T I O N 163
the addition of GW3965 did not modify the release of TNF-awhen Paz-PC or CpG-ODN alonewere added tomacrophages(Fig. 6A); whereas, it reduced the release of IP-10 when Paz-PCor CpG were administered alone (Fig. 6B; �P< 0.05).Furthermore, the addition of GW3965 significantly reducedIFNb mRNA expression when CpG and Paz-PC wereco-administered (Fig. 6C, black bars; �P< 0.05).
JOURNAL OF CELLULAR PHYSIOLOGY
Discussion
FCF is a key event for the initiation and progression ofatherosclerotic plaques. In this study we demonstrated thatFCF can be induced via the stimulation of TLR9 by CpG in thepresence of oxidized particles of LDL, such as Paz-PC. Theproduction of pro-inflammatory cytokines such as TNF-a,

Fig. 5. Nuclear localization of LXRa (A) or p65/RelA subunit (B) 1 h and half after the addition ofCpG (1mg/ml) and/or Paz-PC (25mg/ml) in thepresence of GW3965 (1mM), LXRa agonist. GW3965 tended to increase the translocation of LXRa (A) to the nucleus but diminished thetranslocation of p65 subunit after CpG-ODN and Paz-PC co-administration (B) as demonstrated by themerge of Texas Red and DAPI-positivestainings. Rabbit IgG were used as isotype control (A). Experiments were performed in duplicate in three different time conditions.Part C: The nuclear transcription of the p65 subunit. The addition of GW3965 toCpG andPaz-PC onRAW264.7 cells significantly decreased theactivity of NF-kB (C). n.d., not detected. Data represent meanWSEM and each experiment was performed at least three times for each typeof experimental condition. Statistically significant differences are MP<0.05, and MMP<0.01 as determined by Student’s t-test. [Color figure can beviewed in the online issue, which is available at www.interscience.wiley.com.]
164 S O R R E N T I N O E T A L .
IFNb, and IP-10 led to the formation of foam cells from murinemacrophages, implying the activation of both NF-kB- andIRF7-dependent signaling pathways. The activation of LXRsprevented the formation of foam cells by reducing the levels ofTNF-a, IFNb, and IP-10, due to the reduction of NF-kB andIRF7 activation.
Several clinical and animal studies have related TLRs-dependent signaling pathway to the progression andexacerbation of atherosclerosis (Michelsen et al., 2004;
JOURNAL OF CELLULAR PHYSIOLOGY
Michelsen and Arditi, 2006; Mullick et al., 2008; Naiki et al.,2008; Sorrentino and Arditi, 2009). Indeed, TLR4 andMyD88 gene deficiencies in atherosclerosis-pronehypercholesterolemic mice (ApoE�/�) have decreased plaquesize, lipid content, and expression of pro-inflammatorycytokines and chemokines such as IL-12 and MCP-1 (Michelsenet al., 2004). Mullick et al. (2008) also reported the implicationof TLR2 in the progression of atherosclerotic lesions. It wasshown that TLR2 mediated the development of the disease

Fig. 6. Assessment of TNF-a (A) and IP-10 (B) by means of ELISA.RAW264.7 cells were treated with CpG (1mg/ml) and/or Paz-PC(25mg/ml) in the presence or not of GW3965 (1mM). The additionof GW3965 decreased the release of TNF-a (A) and IP-10 (B). Datarepresent meanWSEM, NU 8 and each treatment was performed induplicate each experimental time. (C) Expression of IFNb mRNAlevels at 3 h post the addition of GW3965 to CpG and/or Paz-PC.GW3965 decreased the production of IFNb mRNA. Data representmeanWSEMandeachexperimentwasperformedat least three timesfor each type of experimental condition. Statistically significantdifferences are MP<0.05, and MMP<0.001 as determined by One-WayAnova analysis of variance and Student’s t-test.
T L R 9 I N D U C E S F O A M C E L L F O R M A T I O N 165
by explicating a fundamental role on stromal cells, such asendothelial cells (Mullick et al., 2008). Furthermore, besides thesystemic and complex implication of TLRs in the atheromaprogression, their involvement in the formation of foam cellsfrom macrophages that engulf lipids at the level of the
JOURNAL OF CELLULAR PHYSIOLOGY
sub-endothelial matrix (Sorrentino and Arditi, 2009), resultedfundamental. In very recent papers, it was showed that TLR2and TLR4 are majorly responsible for the formation of foamcells (Cao et al., 2007; Chen et al., 2008;Naiki et al., 2008), sinceTLR2�/� and TLR4�/� macrophages did not turn into foamcells.
TLRs trigger PAMPs; however, their implication in theatherosclerotic process has been related either to therecognition of endogenous molecules (Michelsen and Arditi,2006; Sorrentino and Arditi, 2009) or to bacteria, such asC. pneumoniae, that are present in the plaque of atheroscleroticpatients (Sorrentino and Arditi, 2009). In particular, Chen et al.(2008) proved that C. pneumoniae was able to increase FCF viaTLR2 and TLR4 in a MyD88-dependent and -independentmanner in the presence of ox-LDL. Our results showed thatTLR9 activation can promote the formation of foam cells. Thesedata were comparable to LPS- and Pam3CSK4-induced foamcells in the presenceof Paz-PC. LPS, TLR4 ligand, or Pam3CSK4,TLR2 ligand, plus Paz-PC were used as positive controls.Moreover, our study may lead to understand how TLRs canfacilitate or exacerbate atherosclerosis progression, sinceTLR9 triggers unmethylated CpG motifs, highly present inbacteria such as C. pneumoniae, that, as reported above, canexacerbate atherosclerotic plaques (Chen et al., 2008; Naikiet al., 2008). Although there is no direct evidence that showsthe implication of TLR9 in the atherosclerotic process, there is asubstantial amount of indirect evidence indicating that it can(Erridge et al., 2008; Lee et al., 2008). Indeed, the geneticabsence of TLR2 or TLR4, and MyD88 or TRIF inhypercholesterolemic-prone animal models showed that theplaque size was not completely abolished (Naiki et al.,2008). TLR3�/� macrophages did not develop foam cells, incontrast to TLR2 and TLR4 knockout macrophages (Chenet al., 2008). Here, in our study we speculate that TLR9 mayparticipate with TLR2 and TLR4 to the progression andexacerbation of atherosclerotic plaques, by inducing FCF. Insupport, a very recent work reported that foam cells canbe induced by TLR9 from murine macrophages (Lee et al.,2008).
TLR9 ligation leads to both the activation of NF-kB and IRF7in a MyD88-dependent pathway. In the first case, pro-inflammatory cytokines, such as TNF-a, and in the second caseIFN type I, such as IFNa or b, are produced (Kawai and Akira,2008). In our experimental conditions, TNF-a, IFNb, and IP-10,strictly correlated to IFNa/b, were significantly producedwhenCpG was added to the oxidized particles of LDL, Paz-PC. Themechanism by which murine macrophages shift theirmorphology towards foam cells were NF-kB- and IRF7-dependent when TLR9 was stimulated. The phosphorylation ofIkBa by the IKK complex leads to the nuclear translocation ofp65/Rel A subunit of NF-kB (Bonizzi and Karin, 2004). Theadministration of CpG and Paz-PC increased the nuclearactivity of p65, accompanied by the activation of IRF7 that led tothe production of IFNb and IP-10 (Fig. 7). Lee et al. (2008)supported the hypothesis that TLR9-induced FCF wasMAP3kinase-dependent. This latter study is in line with ourssince it is known that several members of the MAP3kinasefamily are involved in the phosphorylation of the IKK complex(Heyninck and Beyaert, 2005).
A very interesting finding in our study was the anti-inflammatory effect of LXRs agonist, GW3965, on TLR9-induced FCF. LXRs are nuclear receptors that arephysiologically interested in the efflux and metabolism oflipids; however, the a inducible isoform (LXRa) can exertanti-inflammatory effects (Bensinger and Tontonoz, 2008).GW3965 is a synthetic agonist for both LXRb, constitutive, andLXRa. Our data revealed that the addition of GW3965 to CpGand Paz-PC increased LXRa presence towards the nucleus,where it increased the expression of scavenger receptors

Fig. 7. Schematic for the potential mechanism by which TLR9 stimulation induces the formation of foam cells. The activation of TLR9 leads toNF-kB- and IRF7-dependent signaling pathways that lead to the synthesis of pro-inflammatory cytokines, such as TNF-a and IFNb, thus IP-10. Incontrast, the addition of GW3965 to murine macrophages-derived foam cells blocked the production of pro-inflammatory cytokines, since thecounterbalance againstNF-kBand IRF7and consequent increase of protein implicated in theeffluxof cholesterol, such asABCA-1 and scavengerreceptors. [Color figure can be viewed in the online issue, which is available at www.interscience.wiley.com.]
166 S O R R E N T I N O E T A L .
(i.e., CD36) or cellular lipid efflux-implicated proteins, such asABCA-1 (Castrillo et al., 2003; Bensinger and Tontonoz, 2008).
The anti-inflammatory effect of LXRs was mediated by theinhibition of NF-kB, as already reported by others (Castrilloet al., 2003; Bensinger andTontonoz, 2008). Indeed, the nucleartranslocation of p65 was diminished and the release of TNF-afrom the murine macrophages-derived foam cells was reducedafter GW3965 addition. Concomitantly, GW3965 alsodecreased the production of IP-10 and IFNb, implying thenegative cross-talk of LXRs on IRF7 signaling pathway. Insupport, Chen et al. (2008) demonstrated that IRF3 activity wasdiminished when GW3965 was added to the peritonealmacrophages pre-treated with C. pneumoniae and ox-LDL.There is no direct evidence that C. pneumoniae can trigger viaTLR9, however, peritoneal macrophages of IRF3�/� mice, inthe same study, were less susceptible to the formation of foamcells. This may strengthen our hypothesis about the implicationof IRF7 in the FCF, since IRF3 and IRF7 heterodimerize and leadto gene transcription (Kawai and Akira, 2008).
Collectively, our data showed that the ligation of TLR9 canlead to the formation of foam cells when lipids accumulate intomacrophages.MyD88-dependent signaling pathway leads to the
JOURNAL OF CELLULAR PHYSIOLOGY
activation of bothNF-kB and IRF7 upon TLR9 activation (Kawaiand Akira, 2008), that in turn promotes the synthesis of pro-inflammatory cytokines that exacerbate the atheroscleroticprocess (Fig. 7). In contrast, the activation of LXRs cannegatively modulate NF-kB and IRF7-dependent signalingpathways (Fig. 7), leading to a potential anti-atherosclerotictherapeutic.
Acknowledgments
We thank Giovanni Forte for his technical support. This workwas supported by grants from the University of Salerno (FARB2007–2008) in favor of Prof. Aldo Pinto.
Literature Cited
Bensinger SJ, Tontonoz P. 2008. Integration of metabolism and inflammation by lipid-activated nuclear receptors. Nature 454:470–477.
BonizziG, KarinM. 2004. The twoNF-kappaB activation pathways and their role in innate andadaptive immunity. Trends Immunol 25:280–288.
Cao F, Castrillo A, Tontonoz P, Re F, Byrne GI. 2007. Chlamydia pneumoniae-inducedmacrophage foam cell formation is mediated by Toll-like receptor 2. Infect Immun 75:753–759.

T L R 9 I N D U C E S F O A M C E L L F O R M A T I O N 167
Castrillo A, Joseph SB, Vaidya SA, Haberland M, Fogelman AM, Cheng G, Tontonoz P. 2003.Crosstalk between LXR and toll-like receptor signaling mediates bacterial and viralantagonism of cholesterol metabolism. Mol Cell 12:805–816.
Chen S, Sorrentino R, Shimada K, Bulut Y, Doherty TM, Crother TR, Arditi M. 2008.Chlamydia pneumoniae-induced foam cell formation requires MyD88-dependent and-independent signaling and is reciprocally modulated by liver X receptor activation.J Immunol 181:7186–7193.
Erridge C, Burdess A, Jackson AJ, Murray C, Riggio M, Lappin D, Milligan S, Webb DJ. 2008.Vascular cell responsiveness to Toll-like receptor ligands in carotid atheroma. Eur J ClinInvest 38: 713–720.
Heyninck K, Beyaert R. 2005. A20 inhibits NF-kappaB activation by dual ubiquitin-editingfunctions. Trends Biochem Sci 30:1–4.
Kaisho T, Tanaka T. 2008. Turning NF-kappaB and IRFs on and off in DC. Trends Immunol29:329–336.
Kawai T, Akira S. 2008. Toll-like receptor and RIG-I-like receptor signaling. AnnNYAcad Sci1143:1–20.
Lee JG, Lim EJ, Park DW, Lee SH, Kim JR, Baek SH. 2008. A combination of Lox-1 and Nox1regulates TLR9-mediated foam cell formation. Cell Signal 20:2266–2275. 11.
Michelsen KS, Arditi M. 2006. Toll-like receptor signaling and atherosclerosis. Curr OpinHematol 13:163–168.
JOURNAL OF CELLULAR PHYSIOLOGY
MichelsenKS,WongMH, Shah PK, ZhangW,Yano J, DohertyTM, Akira S, et al. 2004. Lack ofToll-like receptor 4 ormyeloid differentiation factor 88 reduces atherosclerosis and altersplaque phenotype in mice deficient in apolipoprotein. Proc Natl Acad Sci USA 101:10679–10684.
MullickAE, SoldauK, KiossesWB, Bell TA, Tobias PS,Curtiss LK. 2008. Increased endothelialexpression of Toll-like receptor 2 at sites of disturbed blood flow exacerbates earlyatherogenic events. J Exp Med 205:373–383.
Naiki Y, Sorrentino R, Wong MH, Michelsen KS, Shimada K, Chen S, Yilmaz A, et al. 2008.TLR/MyD88 and liver X receptor alpha signaling pathways reciprocally controlChlamydia pneumoniae-induced acceleration of atherosclerosis. J Immunol 181:7176–7185.
Podrez EA, Febbraio M, Sheibani N, Schmitt D, Silverstein RL, Hajjar DP, Cohen PA,et al. 2000. Macrophage scavenger receptor CD36 is the major receptor for LDLmodified by monocyte-generated reactive nitrogen species. J Clin Invest 105:1039–1041.
Podrez EA, Poliakov E, ShenZ, ZhangR,DengY, SunM, Finton PJ, et al. 2002. A novel family ofatherogenic oxidized phospholipids promotes macrophage foam cell formation via thescavenger receptor CD36 and is enriched in atherosclerotic lesions. J Biol Chem277:38517–38523.
Sorrentino R, Arditi M. 2009. Innate immunity, toll-like receptors, and atherosclerosis:Mouse models and methods. Methods Mol Biol 517:1–19.