the abc transporter-encoding gene afr1 affects the resistance of cryptococcus neoformans to...
TRANSCRIPT

R E S E A R C H A R T I C L E
TheABC transporter-encoding geneAFR1 a¡ects the resistanceofCryptococcusneoformans tomicroglia-mediatedantifungalactivitybydelaying phagosomalmaturationCarlotta Francesca Orsi1, Bruna Colombari1, Andrea Ardizzoni1, Samuele Peppoloni1, Rachele Neglia1,Brunella Posteraro2, Giulia Morace3, Giovanni Fadda2 & Elisabetta Blasi1
1Dipartimento di Scienze di Sanita Pubblica, Universita di Modena e Reggio Emilia, Modena, Italy; 2Istituto di Microbiologia, Universita Cattolica del
Sacro Cuore, Roma, Italy; and 3Dipartimento di Sanita Pubblica, Universita degli Studi di Milano, Milan, Italy
Correspondence: Elisabetta Blasi,
Dipartimento di Scienze di Sanita Pubblica,
Universita di Modena e Reggio Emilia, Via
Campi 287, 41100 Modena, Italy. Tel.: 139
059 205 5468; fax: 139 059 205 5483;
e-mail: [email protected]
Received 25 August 2008; revised 25 October
2008; accepted 27 October 2008.
First published online 16 December 2008.
DOI:10.1111/j.1567-1364.2008.00470.x
Editor: Andrew Alspaugh
Keywords
Cryptococcus neoformans ; microglia;
phagocytosis; ABC transporter; phagosomal
maturation.
Abstract
The pathogenic yeast Cryptococcus neoformans has evolved several strategies to
survive within phagocytes. Recently, it has been demonstrated that upregulation of
the ATP binding cassette transporter-encoding gene antifungal resistance 1 (AFR1)
is important not only for determining the resistance of C. neoformans to
fluconazole but also in influencing fungal virulence. In the present study, we
showed that the fluconazole-resistant AFR1-overexpressing mutant strain was not
sensitive to microglia-mediated anticryptococcal activity, as compared with the
fluconazole-susceptible isogenic strains, the wild type and the afr1D mutant.
Interestingly, although the three strains were phagocytosed to a similar extent,
reduced acidification and delayed maturation were observed in phagosomes
containing the AFR1-overexpressing strain with respect to the others. These
findings provide the first evidence that upregulation of the AFR1 gene affects
C. neoformans–microglia interplay, adding insights to the complexity of crypto-
coccal virulence and to its unexpected link with azole resistance.
Introduction
Cryptococcus neoformans is the aetiological agent of human
cryptococcosis, an air-borne invasive opportunistic mycosis
(Casadevall & Perfect, 1998). In immunocompromised
hosts, C. neoformans often disseminates and, because of its
marked neurotropism, it invades the central nervous system,
causing meningoencephalitis that commonly relapses
despite therapy or rather may be fatal in untreated patients
(Casadevall & Perfect, 1998). Although this fungus is known
to replicate extracellularly, recent studies have shown that
C. neoformans actually behaves as a facultative intracellular
pathogen; once ingested, it is able to survive and to replicate
within phagocytes through multiple and complex strategies
(Lee et al., 1995; Mansour & Levitz, 2002; Ma et al., 2006). In
fact, in C. neoformans-infected macrophages, accumulation
of polysaccharide-filled vesicles is observed, followed by the
formation of spacious phagosomes containing viable cryp-
tococcal cells (Tucker & Casadevall, 2002). Moreover, fungal
intracellular replication occurs and may result in lysis of the
host cell; alternatively, extrusion of the cryptococcal phago-
some has been described as a novel strategy through which
both survival of the macrophage and dissemination of the
yeast cells are allowed (Tucker & Casadevall, 2002; Alvarez &
Casadevall, 2006, 2007; Ma et al., 2006).
Microglia, the brain resident phagocytes, are in charge of
locally mediating anticryptococcal defence (Kreutzberg,
1996; Zhou et al., 2007). They respond to infection with
NFkB activation followed by the release of matrix metallo-
proteinases (Alvarez & Casadevall, 2007; Kumnok et al.,
2008), reactive oxygen species and proinflammatory cyto-
kines, such as IL1a, tumor necrosis factor a (TNFa) and
MIP-2a (Jack et al., 2005; Nimmerjahn et al., 2005; Neglia
et al., 2006; Blasi et al., 2007, 1990, 1995b). Microglia may
either allow C. neoformans intracellular survival and replica-
tion (Lee et al., 1995) or exert anticryptococcal activity,
whose levels are enhanced upon exposure to IFN-g or
chloroquine (Blasi et al., 1995b; Mazzolla et al., 1997; Sale-
ppico et al., 1999). Furthermore, C. neoformans resistance to
microglia is partially reduced by yeast cell preopsonization
FEMS Yeast Res 9 (2009) 301–310 Journal compilation c� 2008 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. No claim to original Italian government works

with fresh serum or by pretreatment with protease inhibi-
tors (Blasi et al., 1992, 1995a, b; 2004; Lipovsky et al., 1997).
Thus, both the early yeast-to-microglia recognition step(s)
and the postphagocytosis intracellular events seem to be
relevant in the outcome of such pathogen–host cell inter-
play. As widely described by means of other pathogen–host
cell models, the C. neoformans-containing phagosome is
expected to undergo a multistep maturation process, even-
tually allowing the formation of an acidic compartment
(Chaka et al., 1995; Levitz et al., 1999, 1987; Wozniak et al.,
2006). This process is characterized by the gradual acquisi-
tion of early and late endosomal markers, such as Rab5,
Rab7 and Rab9, and of the late lysosome-associated mem-
brane proteins LAMP1 and LAMP2 (Meresse et al., 1999;
Zerial & McBride, 2001; Barbero et al., 2002; Rink et al.,
2005; Eskelinen, 2006; Huynh et al., 2007).
Specific virulence factors of C. neoformans, including
polysaccharide capsule (Bhattacharjee et al., 1984; Cherniak
& Sundstrom, 1994), melanin (Shaw & Kapica, 1972) or
mannitol production (Wong et al., 1990), superoxidase
dismutase (Jacobson et al., 1994), proteases (Brueske, 1986;
Blasi et al., 2004), phospholipase B and lysophospholipase
(Chen et al., 1997a, b), have been described; among all,
capsule formation and melanin production appear as crucial
elements to elude host defences at the brain level (Blasi et al.,
1992, 1995a). Recently, the antifungal resistance 1 (AFR1)
gene has been identified and characterized (Posteraro et al.,
2003; Sanguinetti et al., 2006): it encodes an ATP binding
cassette transporter protein, involved in the in vitro resis-
tance to fluconazole (Posteraro et al., 2003). We constructed
two C. neoformans strains, both derivatives of the flucona-
zole-susceptible clinical isolate BPY22, namely, the AFR1-
overexpressing (BPY445) and the afr1D(BPY444) strains
(Sanguinetti et al., 2006). Briefly, to generate BPY444, the
AFR1 gene was disrupted by homologous recombination,
and the absence of AFR1 transcripts in this strain was
determined by real-time reverse transcriptase-PCR (RT-
PCR) analysis. In contrast, BPY445 was generated by
random integration of the AFR1 gene, placed under the
control of the constitutively active promoter of the GPD1
gene, into the BPY22 genome; in this strain, AFR1 expres-
sion was about 12-fold increased as compared with that of
the BPY22 strain, as determined by real-time RT-PCR
analysis (Sanguinetti et al., 2006). When compared in a
mouse model of systemic infection, such laboratory mutants
not only retained the expected fluconazole-resistant and
-susceptible phenotypes, but, surprisingly, also differed in
the degree of virulence, BPY445 being consistently more
virulent than BPY444 or BPY22 (Sanguinetti et al., 2006). To
exclude the possibility that the BPY445 phenotype was
related to the genomic position of the overexpression
construct in this strain, we also analysed other independent
transformants, in which the overexpression construct was
located at different genomic sites, as assessed by Southern
blot analysis. The results obtained with these two strains
were fully consistent with those reported for BPY445, thus
confirming that the virulence phenotype of the three strains
was not linked to the site of insertion of the ectopic AFR1
gene (Sanguinetti et al., 2006).
In the present study, we investigated the role of the AFR1
phenotype in the interaction between C. neoformans and
microglia. Upon infection with the wild-type BPY22 or the
two mutants, the Afr1D and the AFR1-overexpressing
strains, microglial cells were assessed for phagocytosis,
phagosome maturation and anticryptococcal activity; NFkB
induction and cytokine response were also evaluated.
Materials and methods
Cryptococcus neoformans strains
The following strains were used: BPY22 (wild type), BPY444
(afr1D) and BPY445 (AFR1-overexpressing mutant) (Pos-
teraro et al., 2003; Sanguinetti et al., 2006). Long-term
storage was carried out in 20% glycerol at � 80 1C. Before
testing, a single colony, obtained on Sabouraud (Oxoid,
Hampshire, UK) agar from each strain, was transferred in
Sabouraud dextrose broth under shaking. When required,
staining of yeast cells was achieved by incubation with
Oregon green 488 (Levitz et al., 1999; Wozniak et al., 2006),
2 mg mL�1 in the dark at 37 1C for 1 h. After labelling, yeast
cells were washed twice with phosphate-buffered saline
(PBS), counted and suspended in Roswell Park Memorial
Institute (RPMI) medium at the desired concentration.
Microglial cell line
The previously established murine microglial cell line BV2
(Blasi et al., 1990) was maintained in RPMI 1640 medium
(Gibco, Grand Island, NY), supplemented with 10% heat-
inactivated foetal bovine serum (Defined Hyclone, Logan,
UT), gentamicin (50mg mL�1) and L-glutamine (2 mM),
hereafter referred to as complete RPMI. Cells were detached
by vigorous shaking biweekly and fresh cultures were started
at a concentration of 5� 105 mL�1, the day before the
experiment.
Reagents and chemicals
Oregon green 488 and LysoTracker DND-99 were obtained
from Molecular Probes (Eugene, OR). Uvitex 2B was
obtained from Polysciences Inc. (Warrington, PA). Poly-L-
lysine, paraformaldehyde and polyclonal antibodies against
murine Rab5, Rab7, Rab9 and LAMP2 were purchased
from Sigma (St. Louis, MO). The Chromeo 642 goat anti-
rabbit IgG, the NFkB enzyme-linked immunosorbent assay
FEMS Yeast Res 9 (2009) 301–310Journal compilation c� 2008 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. No claim to original Italian government works
302 C.F. Orsi et al.

(ELISA) kit and the nuclear extract kit were obtained from
Active Motif (CA).
Measurement of anticryptococcal activity
Yeast cells (105 mL�1 in complete RPMI, 100mL per well)
were plated in 96-well plates; then, BV2 cells were added
(106 mL�1 in complete RPMI, 100mL per well). After an
additional 2, 3 and 4 h of incubation, the plates were
vigorously shaken and Triton X-100 (0.1% final concentra-
tion) was added to each well. According to previously
described protocols (Blasi et al., 1995b), serial dilutions
from each well were made in saline and plated (triplicate
samples) on Sabouraud dextrose agar. The number of CFU
was determined after 48–72 h of incubation at room tem-
perature. Control cultures consisted of yeast cells incubated
in complete RPMI without effector cells. The results were
expressed as percent of anticryptococcal activity according
to the formula described elsewhere (Blasi et al., 1995b).
Phagocytosis assay
To strengthen attachment of BV2 cells to the wells, Lab-Tek
II chamber slides (Nalge Nunc International, Naperville, IL)
were pretreated with poly-L-lysine, 10 mg per well, for 30 min
and washed twice with PBS. BV2 cells (106 mL�1 in complete
RPMI, 100 mL per well) were seeded, incubated for 30 min at
37 1C and 5% CO2 and then infected with Oregon green
prelabelled yeast cells (5� 106 mL�1 in complete RPMI,
100 mL per well) and further incubated for 1.5 or 3 h. Then,
Uvitex 2B was added (20 mL per well) 15 min before each
end point (Levitz et al., 1987; Chaka et al., 1995; Lipovsky
et al., 1997). Cells were washed with PBS to remove non-
adherent cryptococci and fixed with 4% formaldehyde freshly
made from paraformaldehyde for 30 min at 4 1C. As pre-
viously described (Levitz et al., 1987; Chaka et al., 1995;
Lipovsky et al., 1997), Uvitex staining of fungi allows one to
discriminate between adherent (Uvitex accessible) and pha-
gocytized (i.e. internalized, Uvitex nonaccessible) yeast cells.
When visualized by epifluorescence microscopy, all the yeast
cells appeared as green (independently of the localization),
while the noninternalized yeast cells were blue stained. Finally,
merging of Oregon green and Uvitex images allowed one to
definitively exclude the extracellular yeast cells. A minimum
of 200 microglial cells per group were scored and any cell
containing one or more particles was counted as phagocytic.
Phagolysosome acidification assay
Visualization of the acid yeast-containing vesicles was per-
formed as previously described (Via et al., 1998). Briefly,
Lab-Tek II chamber slides were prepared as in Phagocytosis
assay; then, the BV2 cells were exposed to the acidotropic
dye LysoTracker DND-99 (75 nM) (Binker et al., 2007) and
infected as described above. Samples were then fixed with
4% formaldehyde and immediately examined by fluores-
cence microscopy. Acidification control groups consisted of
uninfected cells. A minimum of 200 yeast-containing pha-
gosomes were scored; the percent of acid phagolysosomes
was evaluated as the ratio between the number of Lyso-
Tracker-labelled phagosomes and the total number of yeast-
containing phagosomes.
Colocalization experiments
The procedure for immunofluorescence labelling of phago-
some membrane markers was adapted from a previously
described method (Barnewall et al., 1997). Briefly, Lab-Tek
II chamber slides were prepared as in Phagocytosis assay;
then, the BV2 cells were infected with BPY444 or BPY445
strain. At 3 h postinfection, BV2 cells were washed, fixed
with 4% formaldehyde and permeabilized for 10 min with
0.02% Triton X-100; then, primary antibodies (anti-Rab5,
Rab7, Rab9 or LAMP2) were added to each well for 1 h at
room temperature. After two washes with PBS, the second-
ary antibody was added for an additional 1 h. Samples were
washed and then observed by epifluorescence microscopy.
Determination of NFkB activation
BV2 cells (106 mL�1) were infected with BPY22, BPY444 or
BPY445 (107 mL�1; 106 mL�1) for 30 min in 25-cm2 cell
culture flasks. In parallel, BV2 cells were exposed to lipopo-
lysaccharide (1 mg mL�1) as positive controls. Then, cell and
nuclear extracts were obtained using a nuclear extract kit.
Briefly, cells were collected in the PBS/phosphatase inhibitor
solution and suspended in lysis buffer containing DTT
10 mM and a cocktail of protease inhibitors. Solubilized
proteins were then separated from cell debris by centrifuga-
tion (20 min at 14 000 g). For each sample, the protein
concentration in cytoplasmatic and nuclear fractions was
determined by Bradford assay and adjusted to the same
levels. NFkB activation was measured by the NFkB ELISA
kit, according to the manufacturer’s recommendations. The
levels of NFkB activation were measured by a Sunrise
spectrophotometer (Tecan, Salzburg, Austria) at OD450 nm.
Each sample was run in triplicate.
Cytokine measurement
BV2 cells (106 mL�1) were infected with BPY22, BPY444 or
BPY445 (107 mL�1) strain for 6 and 24 h in 24-well plates. In
parallel, BV2 cell treatment with lipopolysaccharide
(1mg mL�1) was included in the assay as positive control.
Then, the supernatants were collected and frozen at
� 80 1C. MIP-1a and TNFa levels were measured by sand-
wich ELISA according to the manufacturer’s protocol (R&D
Systems). The reaction was read as OD using a Microplate
FEMS Yeast Res 9 (2009) 301–310 Journal compilation c� 2008 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. No claim to original Italian government works
303Phagosomal maturation altered by the C. neoformans AFR1 gene
Reader (Sunrise). Experiments were repeated three to five
times and each sample was run in triplicate. Results were
expressed as pg mL�1.
Epifluorescence microscopy
Images were generated and captured with a Nikon Eclipse
90i system, equipped with Nomarski differential interfer-
ence contrast (DIC) optics. The overlapping signals of
Oregon green-labelled yeasts (green fluorescence) with
either the Uvitex 2B (blue fluorescence) or the red fluores-
cence of the LysoTracker or of the secondary antibody
Chromeo 642 were always interpreted as colocalization. At
each time point, samples were photographed with a DS-5Mc
Nikon digital camera and the photographs were then
analysed by the NIKON software program.
Statistical analysis
Statistical analysis was performed by one-way ANOVA with a
Bonferroni correction post-test or by the Student’s t-test.
The results reported in the figures and the table are the
mean� SD of replicates from a representative experiment
out of three to five performed.
Results
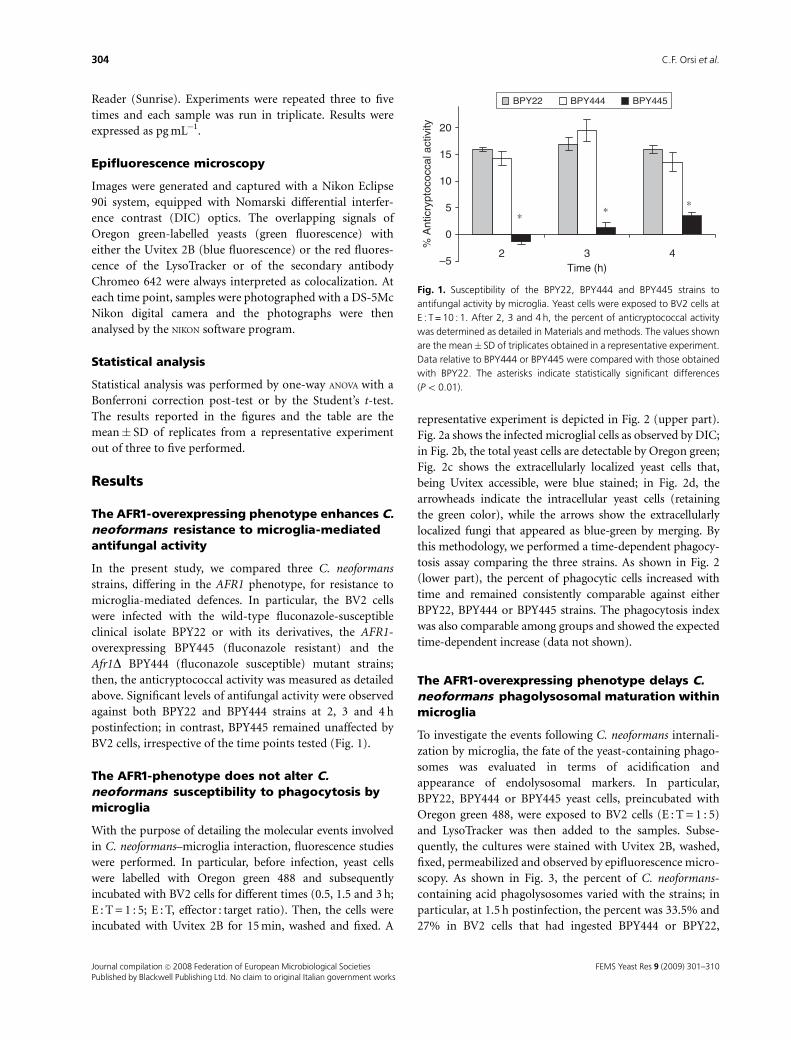
The AFR1-overexpressing phenotype enhances C.neoformans resistance to microglia-mediatedantifungal activity
In the present study, we compared three C. neoformans
strains, differing in the AFR1 phenotype, for resistance to
microglia-mediated defences. In particular, the BV2 cells
were infected with the wild-type fluconazole-susceptible
clinical isolate BPY22 or with its derivatives, the AFR1-
overexpressing BPY445 (fluconazole resistant) and the
Afr1D BPY444 (fluconazole susceptible) mutant strains;
then, the anticryptococcal activity was measured as detailed
above. Significant levels of antifungal activity were observed
against both BPY22 and BPY444 strains at 2, 3 and 4 h
postinfection; in contrast, BPY445 remained unaffected by
BV2 cells, irrespective of the time points tested (Fig. 1).
The AFR1-phenotype does not alter C.neoformans susceptibility to phagocytosis bymicroglia
With the purpose of detailing the molecular events involved
in C. neoformans–microglia interaction, fluorescence studies
were performed. In particular, before infection, yeast cells
were labelled with Oregon green 488 and subsequently
incubated with BV2 cells for different times (0.5, 1.5 and 3 h;
E : T = 1 : 5; E : T, effector : target ratio). Then, the cells were
incubated with Uvitex 2B for 15 min, washed and fixed. A
representative experiment is depicted in Fig. 2 (upper part).
Fig. 2a shows the infected microglial cells as observed by DIC;
in Fig. 2b, the total yeast cells are detectable by Oregon green;
Fig. 2c shows the extracellularly localized yeast cells that,
being Uvitex accessible, were blue stained; in Fig. 2d, the
arrowheads indicate the intracellular yeast cells (retaining
the green color), while the arrows show the extracellularly
localized fungi that appeared as blue-green by merging. By
this methodology, we performed a time-dependent phagocy-
tosis assay comparing the three strains. As shown in Fig. 2
(lower part), the percent of phagocytic cells increased with
time and remained consistently comparable against either
BPY22, BPY444 or BPY445 strains. The phagocytosis index
was also comparable among groups and showed the expected
time-dependent increase (data not shown).
The AFR1-overexpressing phenotype delays C.neoformans phagolysosomal maturation withinmicroglia
To investigate the events following C. neoformans internali-
zation by microglia, the fate of the yeast-containing phago-
somes was evaluated in terms of acidification and
appearance of endolysosomal markers. In particular,
BPY22, BPY444 or BPY445 yeast cells, preincubated with
Oregon green 488, were exposed to BV2 cells (E : T = 1 : 5)
and LysoTracker was then added to the samples. Subse-
quently, the cultures were stained with Uvitex 2B, washed,
fixed, permeabilized and observed by epifluorescence micro-
scopy. As shown in Fig. 3, the percent of C. neoformans-
containing acid phagolysosomes varied with the strains; in
particular, at 1.5 h postinfection, the percent was 33.5% and
27% in BV2 cells that had ingested BPY444 or BPY22,
–5
0
5
10
15
20
2 3 4
% A
ntic
rypt
ococ
cal a
ctiv
ity
BPY22 BPY444 BPY445
Time (h)
∗ ∗ ∗
Fig. 1. Susceptibility of the BPY22, BPY444 and BPY445 strains to
antifungal activity by microglia. Yeast cells were exposed to BV2 cells at
E : T = 10 : 1. After 2, 3 and 4 h, the percent of anticryptococcal activity
was determined as detailed in Materials and methods. The values shown
are the mean� SD of triplicates obtained in a representative experiment.
Data relative to BPY444 or BPY445 were compared with those obtained
with BPY22. The asterisks indicate statistically significant differences
(Po 0.01).
FEMS Yeast Res 9 (2009) 301–310Journal compilation c� 2008 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. No claim to original Italian government works
304 C.F. Orsi et al.
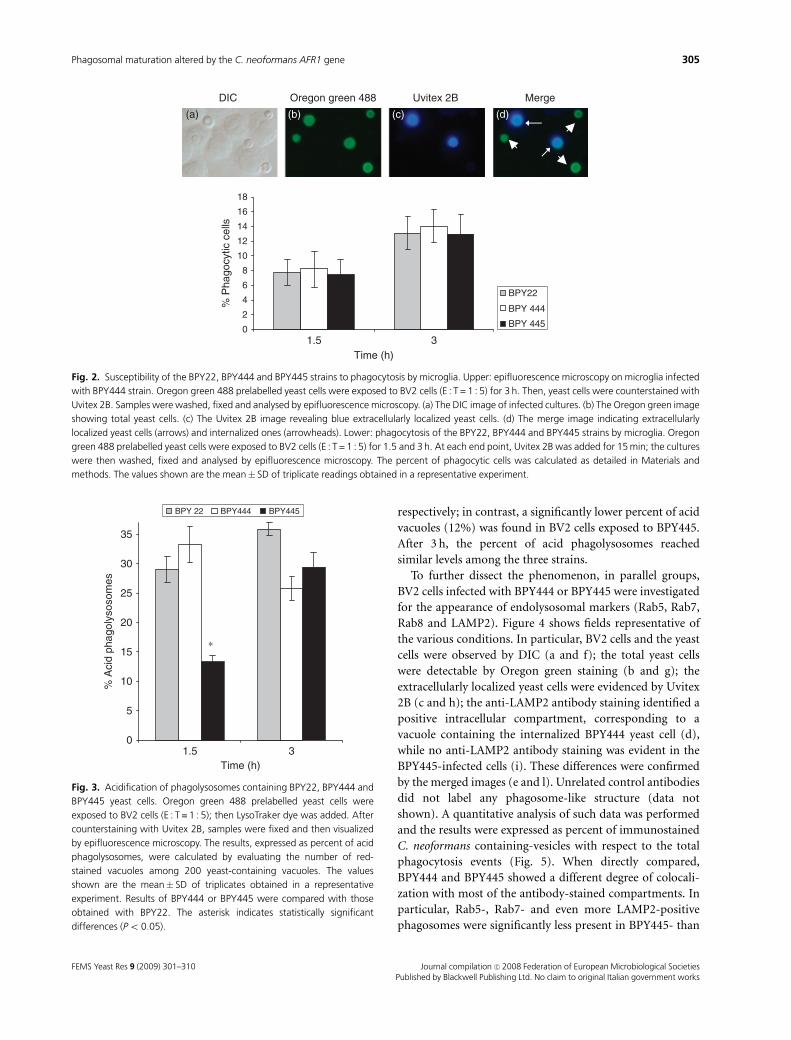
respectively; in contrast, a significantly lower percent of acid
vacuoles (12%) was found in BV2 cells exposed to BPY445.
After 3 h, the percent of acid phagolysosomes reached
similar levels among the three strains.
To further dissect the phenomenon, in parallel groups,
BV2 cells infected with BPY444 or BPY445 were investigated
for the appearance of endolysosomal markers (Rab5, Rab7,
Rab8 and LAMP2). Figure 4 shows fields representative of
the various conditions. In particular, BV2 cells and the yeast
cells were observed by DIC (a and f); the total yeast cells
were detectable by Oregon green staining (b and g); the
extracellularly localized yeast cells were evidenced by Uvitex
2B (c and h); the anti-LAMP2 antibody staining identified a
positive intracellular compartment, corresponding to a
vacuole containing the internalized BPY444 yeast cell (d),
while no anti-LAMP2 antibody staining was evident in the
BPY445-infected cells (i). These differences were confirmed
by the merged images (e and l). Unrelated control antibodies
did not label any phagosome-like structure (data not
shown). A quantitative analysis of such data was performed
and the results were expressed as percent of immunostained
C. neoformans containing-vesicles with respect to the total
phagocytosis events (Fig. 5). When directly compared,
BPY444 and BPY445 showed a different degree of colocali-
zation with most of the antibody-stained compartments. In
particular, Rab5-, Rab7- and even more LAMP2-positive
phagosomes were significantly less present in BPY445- than
0
2
4
6
8
10
12
14
16
18
% P
hago
cytic
cel
ls
BPY22
BPY 444
BPY 445
DIC
(a) (b) (c) (d)
1.5 3Time (h)
MergeUvitex 2BOregon green 488
Fig. 2. Susceptibility of the BPY22, BPY444 and BPY445 strains to phagocytosis by microglia. Upper: epifluorescence microscopy on microglia infected
with BPY444 strain. Oregon green 488 prelabelled yeast cells were exposed to BV2 cells (E : T = 1 : 5) for 3 h. Then, yeast cells were counterstained with
Uvitex 2B. Samples were washed, fixed and analysed by epifluorescence microscopy. (a) The DIC image of infected cultures. (b) The Oregon green image
showing total yeast cells. (c) The Uvitex 2B image revealing blue extracellularly localized yeast cells. (d) The merge image indicating extracellularly
localized yeast cells (arrows) and internalized ones (arrowheads). Lower: phagocytosis of the BPY22, BPY444 and BPY445 strains by microglia. Oregon
green 488 prelabelled yeast cells were exposed to BV2 cells (E : T = 1 : 5) for 1.5 and 3 h. At each end point, Uvitex 2B was added for 15 min; the cultures
were then washed, fixed and analysed by epifluorescence microscopy. The percent of phagocytic cells was calculated as detailed in Materials and
methods. The values shown are the mean� SD of triplicate readings obtained in a representative experiment.
0
5
10
15
20
25
30
35
% A
cid
phag
olys
osom
es
BPY 22 BPY444 BPY445
1.5 3Time (h)
∗
Fig. 3. Acidification of phagolysosomes containing BPY22, BPY444 and
BPY445 yeast cells. Oregon green 488 prelabelled yeast cells were
exposed to BV2 cells (E : T = 1 : 5); then LysoTraker dye was added. After
counterstaining with Uvitex 2B, samples were fixed and then visualized
by epifluorescence microscopy. The results, expressed as percent of acid
phagolysosomes, were calculated by evaluating the number of red-
stained vacuoles among 200 yeast-containing vacuoles. The values
shown are the mean� SD of triplicates obtained in a representative
experiment. Results of BPY444 or BPY445 were compared with those
obtained with BPY22. The asterisk indicates statistically significant
differences (Po 0.05).
FEMS Yeast Res 9 (2009) 301–310 Journal compilation c� 2008 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. No claim to original Italian government works
305Phagosomal maturation altered by the C. neoformans AFR1 gene
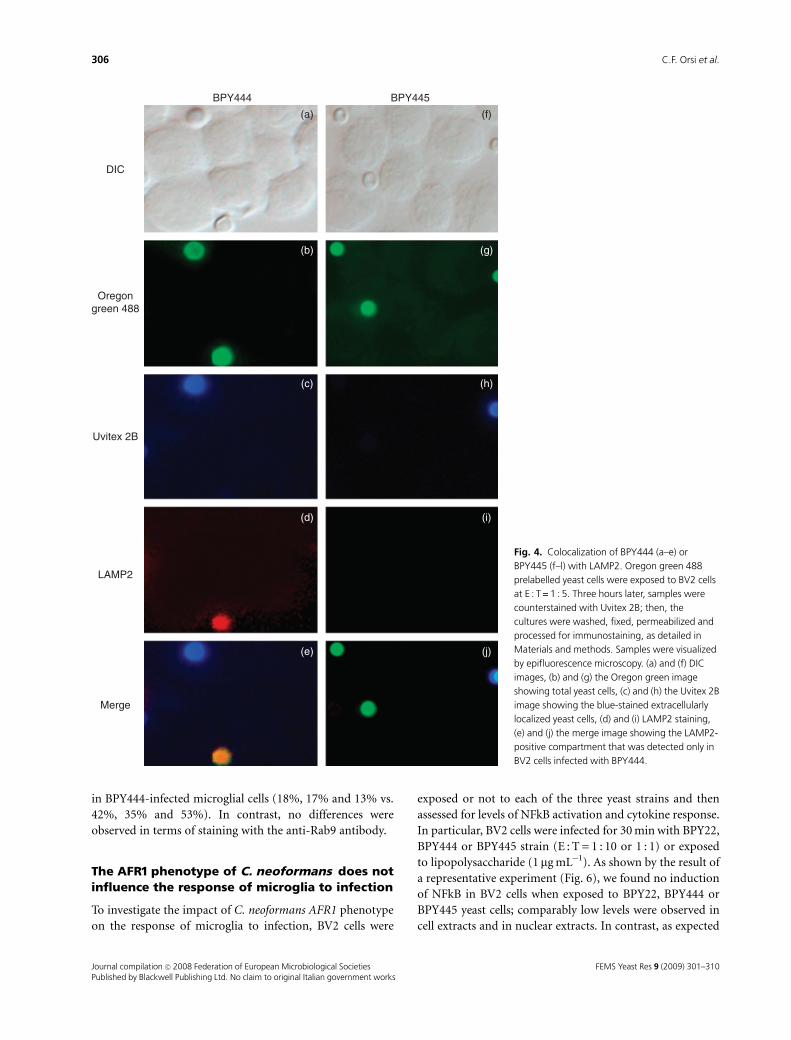
in BPY444-infected microglial cells (18%, 17% and 13% vs.
42%, 35% and 53%). In contrast, no differences were
observed in terms of staining with the anti-Rab9 antibody.
The AFR1 phenotype of C. neoformans does notinfluence the response of microglia to infection
To investigate the impact of C. neoformans AFR1 phenotype
on the response of microglia to infection, BV2 cells were
exposed or not to each of the three yeast strains and then
assessed for levels of NFkB activation and cytokine response.
In particular, BV2 cells were infected for 30 min with BPY22,
BPY444 or BPY445 strain (E : T = 1 : 10 or 1 : 1) or exposed
to lipopolysaccharide (1mg mL�1). As shown by the result of
a representative experiment (Fig. 6), we found no induction
of NFkB in BV2 cells when exposed to BPY22, BPY444 or
BPY445 yeast cells; comparably low levels were observed in
cell extracts and in nuclear extracts. In contrast, as expected
DIC
Oregongreen 488
Uvitex 2B
LAMP2
Merge
BPY444 BPY445
(a)
(b) (g)
(h)(c)
(d) (i)
(e)
(f)
(j)
Fig. 4. Colocalization of BPY444 (a–e) or
BPY445 (f–l) with LAMP2. Oregon green 488
prelabelled yeast cells were exposed to BV2 cells
at E : T = 1 : 5. Three hours later, samples were
counterstained with Uvitex 2B; then, the
cultures were washed, fixed, permeabilized and
processed for immunostaining, as detailed in
Materials and methods. Samples were visualized
by epifluorescence microscopy. (a) and (f) DIC
images, (b) and (g) the Oregon green image
showing total yeast cells, (c) and (h) the Uvitex 2B
image showing the blue-stained extracellularly
localized yeast cells, (d) and (i) LAMP2 staining,
(e) and (j) the merge image showing the LAMP2-
positive compartment that was detected only in
BV2 cells infected with BPY444.
FEMS Yeast Res 9 (2009) 301–310Journal compilation c� 2008 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. No claim to original Italian government works
306 C.F. Orsi et al.
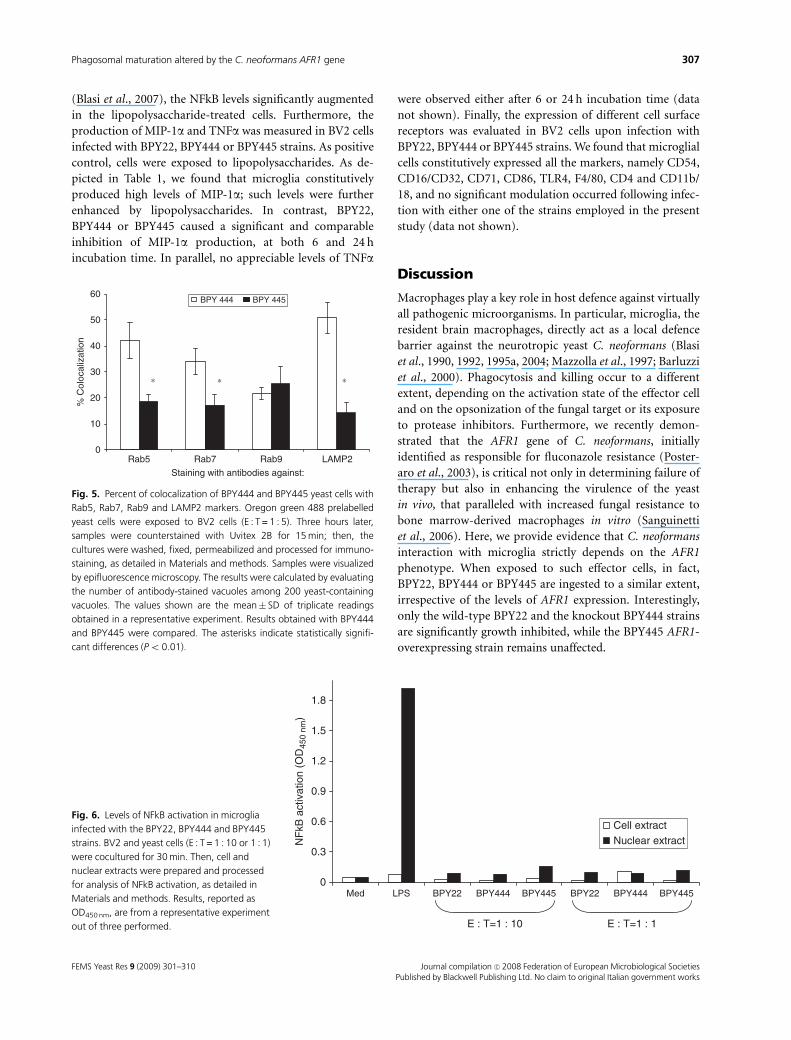
(Blasi et al., 2007), the NFkB levels significantly augmented
in the lipopolysaccharide-treated cells. Furthermore, the
production of MIP-1a and TNFa was measured in BV2 cells
infected with BPY22, BPY444 or BPY445 strains. As positive
control, cells were exposed to lipopolysaccharides. As de-
picted in Table 1, we found that microglia constitutively
produced high levels of MIP-1a; such levels were further
enhanced by lipopolysaccharides. In contrast, BPY22,
BPY444 or BPY445 caused a significant and comparable
inhibition of MIP-1a production, at both 6 and 24 h
incubation time. In parallel, no appreciable levels of TNFa
were observed either after 6 or 24 h incubation time (data
not shown). Finally, the expression of different cell surface
receptors was evaluated in BV2 cells upon infection with
BPY22, BPY444 or BPY445 strains. We found that microglial
cells constitutively expressed all the markers, namely CD54,
CD16/CD32, CD71, CD86, TLR4, F4/80, CD4 and CD11b/
18, and no significant modulation occurred following infec-
tion with either one of the strains employed in the present
study (data not shown).
Discussion
Macrophages play a key role in host defence against virtually
all pathogenic microorganisms. In particular, microglia, the
resident brain macrophages, directly act as a local defence
barrier against the neurotropic yeast C. neoformans (Blasi
et al., 1990, 1992, 1995a, 2004; Mazzolla et al., 1997; Barluzzi
et al., 2000). Phagocytosis and killing occur to a different
extent, depending on the activation state of the effector cell
and on the opsonization of the fungal target or its exposure
to protease inhibitors. Furthermore, we recently demon-
strated that the AFR1 gene of C. neoformans, initially
identified as responsible for fluconazole resistance (Poster-
aro et al., 2003), is critical not only in determining failure of
therapy but also in enhancing the virulence of the yeast
in vivo, that paralleled with increased fungal resistance to
bone marrow-derived macrophages in vitro (Sanguinetti
et al., 2006). Here, we provide evidence that C. neoformans
interaction with microglia strictly depends on the AFR1
phenotype. When exposed to such effector cells, in fact,
BPY22, BPY444 or BPY445 are ingested to a similar extent,
irrespective of the levels of AFR1 expression. Interestingly,
only the wild-type BPY22 and the knockout BPY444 strains
are significantly growth inhibited, while the BPY445 AFR1-
overexpressing strain remains unaffected.
0
10
20
30
40
50
60
% C
oloc
aliz
atio
n
BPY 444 BPY 445
∗ ∗∗
Rab5 Rab7 Rab9 LAMP2
Staining with antibodies against:
Fig. 5. Percent of colocalization of BPY444 and BPY445 yeast cells with
Rab5, Rab7, Rab9 and LAMP2 markers. Oregon green 488 prelabelled
yeast cells were exposed to BV2 cells (E : T = 1 : 5). Three hours later,
samples were counterstained with Uvitex 2B for 15 min; then, the
cultures were washed, fixed, permeabilized and processed for immuno-
staining, as detailed in Materials and methods. Samples were visualized
by epifluorescence microscopy. The results were calculated by evaluating
the number of antibody-stained vacuoles among 200 yeast-containing
vacuoles. The values shown are the mean� SD of triplicate readings
obtained in a representative experiment. Results obtained with BPY444
and BPY445 were compared. The asterisks indicate statistically signifi-
cant differences (Po 0.01).
0
0.3
0.6
0.9
1.2
1.5
1.8
Med LPS BPY22 BPY444 BPY445 BPY22 BPY444 BPY445
NF
kB a
ctiv
atio
n (O
D45
0 nm
)
Cell extractNuclear extract
E : T=1 : 10 E : T=1 : 1
Fig. 6. Levels of NFkB activation in microglia
infected with the BPY22, BPY444 and BPY445
strains. BV2 and yeast cells (E : T = 1 : 10 or 1 : 1)
were cocultured for 30 min. Then, cell and
nuclear extracts were prepared and processed
for analysis of NFkB activation, as detailed in
Materials and methods. Results, reported as
OD450 nm, are from a representative experiment
out of three performed.
FEMS Yeast Res 9 (2009) 301–310 Journal compilation c� 2008 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. No claim to original Italian government works
307Phagosomal maturation altered by the C. neoformans AFR1 gene

These findings closely recall those previously reported
(Sanguinetti et al., 2006) showing that BPY445 displays
enhanced intracellular survival when compared with strains
BPY22 and BPY444 in bone marrow-derived macrophages;
yet, in those cells such a phenomenon is evident at 28 h,
while in microglia 2–4 h are sufficient to discriminate
between resistant and susceptible C. neoformans strains. At
the moment, it remains to be elucidated whether the
anatomical origin and/or the activation state of the two
effector populations may account for the diverse timing
observed. In any case, taken together, our present and
previous findings provide the first evidence on the relevance
of AFR1 phenotype in the intracellular fate of C. neoformans.
In this respect, the fact that two other independent transfor-
mants, having differently located the overexpressing con-
struct, exhibit behavior similar to BPY445 (Sanguinetti
et al., 2006; data not shown) argues against the relevance of
the insertion site in the phenomenon observed.
Because of the marked neurotropism of C. neoformans
and the clinical relevance of cryptococcal meningoencepha-
litis (Casadevall & Perfect, 1998), we further focused our
studies on the outcome of microglia–C. neoformans infec-
tion. As shown here, the early events occurring in such
yeast-to-effector cell interplay are not critical, because
phagocytosis as well as binding (data not shown) are
comparable with respect to each of the three strains. In
contrast, the results by epifluorescence microscopy demon-
strate that phagocytic vacuole acidification is significantly
reduced in BV2 cells engulfed with BPY445 in contrast to
BPY22- or BPY444-infected cells. In line with these findings,
the pathway of phagolysosome maturation happens to
depend on AFR1 phenotype, as demonstrated by the fact
that the early endosome marker Rab5 is detectable in
4 40% but in o 20% of the microglia infected with
BPY444 and BPY445, respectively. A similar trend is also
observed with the late marker Rab7, while no differences
occur when assessing another late maker, Rab9. The im-
paired maturation of the BPY445-containing phagolyso-
somes has been further confirmed by staining with the
anti-LAMP2 antibody, known to identify a very late marker
of phagolysosome development (Eskelinen, 2006; Huynh
et al., 2007). Our data show that LAMP2 is detected in
4 50% of the BPY444-containing phagolysosomes while
o 15% positivity is observed with BPY445. Besides provid-
ing a direct explanation for the lack of growth inhibition
observed against BPY445, these findings imply that the
differences in handling the three strains by microglia strictly
depend on the levels of maturation and acidification of the
phagocytic vacuole. Because the protein encoded by the
AFR1 gene is an efflux pump, it is reasonable to speculate
that such a phenotypical trait of C. neoformans, being
associated with reduced acidification of the phagocytosis
vacuole, is responsible for the enhanced intracellular survi-
val of the AFR1-overexpressing strain. Thus, a novel me-
chanism may be added to the ones already established by
which C. neoformans may alter phagosomal homoeostasis,
such as altered permeabilization, intraphagosomal growth
and eventually extrusion (Chaka et al., 1995; Levitz et al.,
1999; Tucker & Casadevall, 2002; Alvarez & Casadevall,
2006, 2007). On the other hand, we tend to exclude any
involvement of other cryptococcal virulence determinants in
the phenomenon described above, because no differences
have been observed between the AFR1-overexpressing
BPY445 and the parental BPY22 in terms of polysaccharide
capsule formation or other well-recognized parameters of
virulence (Sanguinetti et al., 2006).
In the present report, the consequences of C. neoformans–
microglia interaction have also been assessed in terms of
molecular and functional changes occurring in immune
cells, namely NFkB activation, cytokine secretion and
expression of surface markers. NFkB is known to be a central
mediator of gene transcription for a variety of cellular
responses (Li & Verma, 2002); in particular, it represents a
pivotal signal driving microglia response against infectious
agents (Neglia et al., 2006; Blasi et al., 2007). Unexpectedly,
the present results indicate that BPY22, BPY444 or BPY445
infection does not allow NFkB activation in microglial cells.
These findings are in line with (1) no changes in the
expression of surface markers by the microglial infected cells
and (2) no induction of secretory response, in terms of
TNFa production. Furthermore, the constitutively high
levels of MIP-1a production, observed in uninfected control
cells, are significantly impaired by C. neoformans infection;
the extent of the phenomenon is similar when comparing
BPY22, BPY444 or BPY445. Such inhibitory effects are in
line with the previous studies investigating the influence of
cryptococcal capsule in vitro (Barluzzi et al., 1998) or the
Table 1. Levels of MIP-1a in BV2 cells infected with BPY22, BPY 444 or
BPY445 strains
Treatment
Levels of MIP-1a
6 h 24 h
Medium 2139.0� 80.2 848.0� 80.0
Lipopolysaccharide 6579.0� 110.5� 5960.0� 118.7�
BPY22 958.0� 34.8� 327.0� 56.5�
BPY444 918.0� 28.9� 367.0� 70.0�
BPY445 960.0� 65.0� 558.0� 81.0��
BV2 cells were infected with BPY22, BPY444 or BPY445 yeast cells for 6
or 24 h. In parallel, lipopolysaccharide treatment (1 mg mL�1) was in-
cluded as positive control. Then, the supernatants were harvested and
MIP-1a levels were evaluated, as detailed in Materials and methods. The
values, expressed as picograms per millilitre, are the mean� SD of
triplicate determinations obtained in a representative experiment out of
four performed.�Po 0.001;��P = 0.02 untreated cells vs. treated cells.
FEMS Yeast Res 9 (2009) 301–310Journal compilation c� 2008 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. No claim to original Italian government works
308 C.F. Orsi et al.

role of melanin production by an in vivo model of menin-
goencephalitis (Huffnagle et al., 1995). As TNFa and MIP-
1a are crucial regulatory elements in initiating host immune
reaction (Biron et al., 1998; Maurer & von Stebut, 2004), the
present in vitro data suggest that MIP-1a production is
downregulated while TNFa is not even involved in the early
microglia–C. neoformans interplay. In any case, C. neofor-
mans immunosuppressive effects do not depend on the
AFR1 phenotype.
Although aware of the intrinsic limitations of an in vitro
model, here we provide new insights on the role of the AFR1
gene in the resistance of C. neoformans to microglia-
mediated antifungal activity, especially highlighting the
potential innate function of the AFR1 protein that has been
only lately co-opted for antifungal resistance.
Acknowledgements
We wish to acknowledge Mr A. Martino for excellent
informatic support and Prof. A. Todaro for sharing experi-
ence and providing access to the epifluorescence Eclipse 90i
Nikon microscope (recently founded by CaRiMo). Very
special thanks are also forwarded to Prof. M. Sanguinetti
(Universita Cattolica S. Cuore, Roma) for helpful sugges-
tions and a critical review of the manuscript. This work was
partially supported by the Italian grant MIUR-Programmi
di Ricerca cofinanziati 2005, no. 2005068754.
References
Alvarez M & Casadevall A (2006) Phagosome extrusion and host-
cell survival after Cryptococcus neoformans phagocytosis by
macrophages. Curr Biol 16: 2161–2165.
Alvarez M & Casadevall A (2007) Cell-to-cell spread and massive
vacuole formation after Cryptococcus neoformans infection of
murine macrophages. BMC Immunol 8: 16.
Barbero P, Bittova L & Pfeffer SR (2002) Visualization of Rab9-
mediated vesicle transport from endosomes to the trans-Golgi
in living cells. J Cell Biol 156: 511–518.
Barluzzi R, Brozzetti A, Delfino D, Bistoni F & Blasi E (1998) Role
of the capsule in microglial cell-Cryptococcus neoformans
interaction: impairment of antifungal activity but not of
secretory functions. Med Mycol 36: 189–197.
Barluzzi R, Brozzetti A, Mariucci G, Tantucci M, Neglia RG,
Bistoni F & Blasi E (2000) Establishment of protective
immunity against cerebral cryptococcosis by means of an
avirulent, non melanogenic Cryptococcus neoformans strain.
J Neuroimmunol 109: 75–86.
Barnewall RE, Rikihisa Y & Lee EH (1997) Ehrlichia chaffeensis
inclusions are early endosomes which selectively accumulate
transferrin receptor. Infect Immun 65: 1455–1461.
Bhattacharjee AK, Bennett JE & Glaudemans CP (1984) Capsular
polysaccharides of Cryptococcus neoformans. Rev Infect Dis 6:
619–624.
Binker MG, Cosen-Binker LI, Terebiznik MR et al. (2007)
Arrested maturation of Neisseria-containing phagosomes in
the absence of the lysosome-associated membrane proteins,
LAMP-1 and LAMP-2. Cell Microbiol 9: 2153–2166.
Biron CA, Cousens LP, Ruzek MC, Su HC & Salazar-Mather TP
(1998) Early cytokine responses to viral infections and their
roles in shaping endogenous cellular immunity. Adv Exp Med
Biol 452: 143–149.
Blasi E, Barluzzi R, Bocchini V, Mazzolla R & Bistoni F (1990)
Immortalization of murine microglial cells by a v-raf/v-myc
carrying retrovirus. J Neuroimmunol 27: 229–237.
Blasi E, Barluzzi R, Mazzolla R, Mosci P & Bistoni F (1992)
Experimental model of intracerebral infection with
Cryptococcus neoformans: roles of phagocytes and
opsonization. Infect Immun 60: 3682–3688.
Blasi E, Barluzzi R, Mazzolla R, Pitzurra L, Puliti M, Saleppico S &
Bistoni F (1995a) Biomolecular events involved in
anticryptococcal resistance in the brain. Infect Immun 63:
1218–1222.
Blasi E, Barluzzi R, Mazzolla R, Tancini B, Saleppico S, Puliti M,
Pitzurra L & Bistoni F (1995b) Role of nitric oxide and
melanogenesis in the accomplishment of anticryptococcal
activity by the BV-2 microglial cell line. J Neuroimmunol 58:
111–116.
Blasi E, Colombari B, Orsi CF, Pinti M, Troiano L, Cossarizza A,
Esposito R, Peppoloni S, Mussini C & Neglia R (2004) The
human immunodeficiency virus (HIV) protease inhibitor
indinavir directly affects the opportunistic fungal pathogen
Cryptococcus neoformans. FEMS Immunol Med Mic 42:
187–195.
Blasi E, Ardizzoni A, Colombari B, Neglia R, Baschieri C,
Peppoloni S & Cinco M (2007) NF-kB activation and p38
phosphorylation in microglial cells infected with Leptospira or
exposed to partially purified leptospiral lipoproteins. Microb
Pathogenesis 42: 80–87.
Brueske CH (1986) Proteolytic activity of a clinical isolate of
Cryptococcus neoformans. J Clin Microbiol 23: 631–633.
Casadevall A & Perfect J (1998) Cryptococcus neoformans. ASM
Press, Washington, DC, 541pp (hardcover).
Chaka W, Scharringa J, Verheul AF, Verhoef J, Van Strijp AG &
Hoepelman IM (1995) Quantitative analysis of phagocytosis
and killing of Cryptococcus neoformans by human peripheral
blood mononuclear cells by flow cytometry. Clin Diagn Lab
Immun 2: 753–759.
Chen SC, Muller M, Zhou JZ, Wright LC & Sorrell TC (1997a)
Phospholipase activity in Cryptococcus neoformans: a new
virulence factor? J Infect Dis 175: 414–420.
Chen SC, Wright LC, Santangelo RT, Muller M, Moran VR,
Kuchel PW & Sorrell TC (1997b) Identification of extracellular
phospholipase B, lysophospholipase, and acyltransferase
produced by Cryptococcus neoformans. Infect Immun 65:
405–411.
Cherniak R & Sundstrom JB (1994) Polysaccharide antigens of
the capsule of Cryptococcus neoformans. Infect Immun 62:
1507–1512.
FEMS Yeast Res 9 (2009) 301–310 Journal compilation c� 2008 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. No claim to original Italian government works
309Phagosomal maturation altered by the C. neoformans AFR1 gene

Eskelinen EL (2006) Roles of LAMP-1 and LAMP-2 in lysosome
biogenesis and autophagy. Mol Aspects Med 27: 495–502.
Huffnagle GB, Chen GH, Curtis JL, McDonald RA, Strieter RM &
Toews GB (1995) Down-regulation of the afferent phase of T
cell-mediated pulmonary inflammation and immunity by a
high melanin-producing strain of Cryptococcus neoformans. J
Immunol 155: 3507–3516.
Huynh KK, Eskelinen EL, Scott CC, Malevanets A, Saftig P &
Grinstein S (2007) LAMP proteins are required for fusion of
lysosomes with phagosomes. EMBO J 26: 313–324.
Jack CS, Arbour N, Manusow J, Montgrain V, Blain M, McCrea E,
Shapiro A & Antel JP (2005) TLR signaling tailors innate
immune responses in human microglia and astrocytes.
J Immunol 175: 4320–4330.
Jacobson ES, Jenkins ND & Todd JM (1994) Relationship
between superoxide dismutase and melanin in a pathogenic
fungus. Infect Immun 62: 4085–4086.
Kreutzberg GW (1996) Microglia: a sensor for pathological
events in the CNS. Trends Neurosci 19: 312–318.
Kumnok J, Ulrich R, Wewetzer K, Rohn K, Hansmann F,
Baumgartner W & Alldinger S (2008) Differential
transcription of matrix-metalloproteinase genes in primary
mouse astrocytes and microglia infected with Theiler’s murine
encephalomyelitis virus. J Neurovirol 14: 205–217.
Lee SC, Kress Y, Zhao ML, Dickson DW & Casadevall A (1995)
Cryptococcus neoformans survive and replicate in human
microglia. Lab Invest 73: 871–879.
Levitz SM, DiBenedetto DJ & Diamond RD (1987) A rapid
fluorescent assay to distinguish attached from phagocytized
yeast particles. J Immunol Methods 101: 37–42.
Levitz SM, Nong SH, Seetoo KF, Harrison TS, Speizer RA &
Simons ER (1999) Cryptococcus neoformans resides in an acidic
phagolysosome of human macrophages. Infect Immun 67:
885–890.
Li Q & Verma IM (2002) NF-kappaB regulation in the immune
system. Nat Rev Immunol 2: 725–734.
Lipovsky MM, Gekker G, Anderson WR, Molitor TW, Peterson
PK & Hoepelman AI (1997) Phagocytosis of nonopsonized
Cryptococcus neoformans by swine microglia involves CD14
receptors. Clin Immunol Immunop 84: 208–211.
Ma H, Croudace JE, Lammas DA & May RC (2006) Expulsion of
live pathogenic yeast by macrophages. Curr Biol 16:
2156–2160.
Mansour MK & Levitz SM (2002) Interactions of fungi with
phagocytes. Curr Opin Microbiol 5: 359–365.
Maurer M & von Stebut E (2004) Macrophage inflammatory
protein-1. Int J Biochem Cell Biol 36: 1882–1886.
Mazzolla R, Barluzzi R, Brozzetti A, Boelaert JR, Luna T,
Saleppico S, Bistoni F & Blasi E (1997) Enhanced resistance to
Cryptococcus neoformans infection induced by chloroquine in a
murine model of meningoencephalitis. Antimicrob Agents Ch
41: 802–807.
Meresse S, Steele-Mortimer O, Finlay BB & Gorvel JP (1999) The
Rab7 GTPase controls the maturation of Salmonella
typhimurium-containing vacuoles in HeLa cells. EMBO J 18:
4394–4403.
Neglia R, Colombari B, Peppoloni S, Orsi C, Tavanti A, Senesi S &
Blasi E (2006) Adaptive response of microglial cells to in vitro
infection by Candida albicans isolates with different genomic
backgrounds. Microb Pathog 41: 251–256.
Nimmerjahn A, Kirchhoff F & Helmchen F (2005) Resting
microglial cells are highly dynamic surveillants of brain
parenchyma in vivo. Science 308: 1314–1318.
Posteraro B, Sanguinetti M, Sanglard D, La Sorda M, Boccia S,
Romano L, Morace G & Fadda G (2003) Identification and
characterization of a Cryptococcus neoformans ATP binding
cassette (ABC) transporter-encoding gene, CnAFR1, involved
in the resistance to fluconazole. Mol Microbiol 47: 357–371.
Rink J, Ghigo E, Kalaidzidis Y & Zerial M (2005) Rab conversion
as a mechanism of progression from early to late endosomes.
Cell 122: 735–749.
Saleppico S, Boelaert JR, Omodeo Sale F, Mazzolla R, Morucci P,
Bistoni F & Blasi E (1999) Differential effects of iron load on
basal and interferon-gamma plus lipopolysaccharide enhance
anticryptococcal activity by the murine microglial cell line BV-
2. J Neuroimmunol 93: 102–107.
Sanguinetti M, Posteraro B, La Sorda M, Torelli R, Fiori B,
Santangelo R, Delogu G & Fadda G (2006) Role of AFR1, an
ABC transporter-encoding gene, in the in vivo response to
fluconazole and virulence of Cryptococcus neoformans. Infect
Immun 74: 1352–1359.
Shaw CE & Kapica L (1972) Production of diagnostic pigment by
phenoloxidase activity of Cryptococcus neoformans. Appl
Microbiol 24: 824–830.
Tucker SC & Casadevall A (2002) Replication of Cryptococcus
neoformans in macrophages is accompanied by phagosomal
permeabilization and accumulation of vesicles containing
polysaccharide in the cytoplasm. P Natl Acad Sci USA 99:
3165–3170.
Via LE, Fratti RA, McFalone M, Pagan-Ramos E, Deretic D &
Deretic V (1998) Effects of cytokines on mycobacterial
phagosome maturation. J Cell Sci 111: 897–905.
Wong B, Perfect JR, Beggs S & Wright KA (1990) Production of
the hexitol D-mannitol by Cryptococcus neoformans in vitro and
in rabbits with experimental meningitis. Infect Immun 58:
1664–1670.
Wozniak KL, Vyas JM & Levitz SM (2006) In vivo role of dendritic
cells in a murine model of pulmonary cryptococcosis. Infect
Immun 74: 3817–3824.
Zerial M & McBride H (2001) Rab proteins as membrane
organizers. Nat Rev Mol Cell Biol 2: 107–117.
Zhou Q, Gault RA, Kozel TR & Murphy WJ (2007) Protection
from direct cerebral cryptococcus infection by interferon-
gamma-dependent activation of microglial cells. J Immunol
178: 5753–5761.
FEMS Yeast Res 9 (2009) 301–310Journal compilation c� 2008 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. No claim to original Italian government works
310 C.F. Orsi et al.