systematics and biogeography of eastern caribbean eleutherodactylus (anura: leptodactylidae):...
TRANSCRIPT

2217
Systematics and biogeography of Eastern Caribbean frogs (Leptodactylidae:Eleutherodactylus), with the description of a new species from Dominica
HINRICH KAISER 1 AND DAVID M. GREEN
Redpath Museum and Department of Biology, McGill University, 859 Sherbrooke Street West,Montreal, QC H3A 2K6, Canada
AND
MICHAEL SCHMID
Institut fiir Humangenetik, Universitiit Wiirzburg, Biozelllrum, Am Hubland, 97074 Wiirzburg, Germany
Received April 5, 1994Accepted October 12, 1994
KAISER, H., GREEN, D.M., and SCHMID, M. 1994. Systematics and biogeography of Eastern Caribbean frogs (Leptodactylidae: Eleutherodactylus), with the description of a new species from Dominica. Can. J. Zoo!. 72: 2217-2237.
Populations of Eleutherodactylus on the island of Dominica, West Indies, differ from other Lesser Antillean members ofthe genus by their vocalizations, morphology, sexual size dimorphism, allozymes, and chromosomes. Thus, these populationscomprise a new endemic species, closely related to E. johnstonei and E. martinicensis. The new species is most abundantin montane forest habitats at elevations of more than 500 m. Females commonly attain snout-vent lengths of more than35 mm, making them the largest Lesser Antillean Eleutherodactylus. Frogs are brown during the day, but change color to
. dark orange when active at night. This species is the ninth Eastern Caribbean Eleutherodactylus species and the fifth singleisland endemic. A phylogenetic analysis of external and internal morphological characteristics shows that Eastern CaribbeanEleutherodactylus are members of two distinct clades, one of South American origin, the other of Greater Antillean ancestry.We suggest that the present distribution of these species results from the dispersal of elements of the larger herpetofaunasfrom the Greater Antilles and South America, and that rapid divergence of the Eastern Caribbean Eleutherodactylus faunamay be continuing. The phylogenetic analysis also confirms that morphological characters of Eleutherodactylus species canbe highly homoplastic.
KAISER, H., GREEN, D.M., et SCHMID, M. 1994. Systematics and biogeography of Eastern Caribbean frogs (Leptodactylidae: Eleutherodactylus), with the description of a new species from Dominica. Can. J. Zoo!. 72 : 2217-2237.
Les populations d' Eleutherodactylus de la Dominique, dans les Antilles, different des autres membres du genre dans lesPetites Antilles par leurs cris, leur morphologie, leur dimorphisme sexuel quant 11 la taille, leurs allozymes et leurs chromosomes. Ces populations constituent une nouvelle espece endemique fortement apparentee 11 E. johnstonei et E. martinicensis.La nouvelle espece abonde surtout dans les habitats forestiers montagneux aux altitudes superieures 11 500 m. Les femellesmesurent souvent plus de 35 mm (longueur museau -event) et sont ainsi les Eleutherodacrylus les plus grands des PetitesAntilles. Les grenouilles sont brunes durant la journee, mais prennent une coloration orangee foncee lorsqu'elles sont activesla nuil. Cette esp(:ce est la neuvieme espece d'Eleutherodactylus de I'est des Antilles et la cinquieme espece endemique dansune seule lie. Une analyse phylogenetique des caracteres morphologiques internes et externes a demontre que les Eleutherodactylus de l'est des Antilles appartiennent 11 deux clades distincts, un d'origine sud-americaine, I'autre derivant d'un ancetrevivant dans les Grandes Antilles. La repartition actuelle des especes resulte probablement de la dispersion des grandesherpetofaunes des Grandes Antilles et de l'Amerique du Sud et la divergence rapide de la faune des Eleutherodactylus del'est des Antilles se fait probablement encore. L'analyse phylogcnetique a egalement revele que les caracteres morphologiquesdes especes d' Eleutherodactylus peuvent etre tres homoplastiques.
[Traduit par la Redaction]
Introduction
The neotropical frog genus Eleutherodactylus is the mostspeciose vertebrate genus, represented by over 500 describedspecies (Duellman 1993). Although some of its members havevery distinctive external features (e.g., Lynch 1975, 1980), itis often difficult to differentiate species of Eleutherodactylusmorphologically. Intraspecific variability can be considerable,yet the many species are variants, albeit sometimes extreme,of a conservative morphotype, a small brownish frog (Lynchand La Marca 1993). Previous phylogenetic analyses of relationships within Eleutherodactylus at various levels of classification have been conducted with varying degrees of success(e.g., Miyamoto 1983, 1984, 1986; Hedges 1989; Joglar1989). The use of biochemical characters in post hoc tests ofa given data set invariably improved the resolution of systematic relationships (Hedges 1989; Kaiser 1993).
Ipresent Address: Institut £iir Humangenetik, Universitiit Wiirzburg, Biozentrum, Am Hubland, 97074 Wiirzburg, Germany.
Primed in Canada I Impril11c au Canada
The 128 West Indian species of Eleutherodactylus have beenclassified (Hedges 1989) into the subgenera Eleutherodactylus(comprising 2 species groups), Euhyas (12), and Pelm'ius (2),with 70 species still unassigned to species groups withinEleutherodactylus or Euhyas. Eight species are known toinhabit the Eastern Caribbean (Table I), a region comprisingthe Lesser Antilles, Trinidad, and Tobago (Fig. 1). Many ofthese are single-island endemics whose origin and systematicrelationships are as yet unresolved. The detailed synopsis ofthese taxa by Schwartz (1967) provides few data to supportsister-group relationships or wider ranging affinities for thesetaxa. Their tentative inclusion in the Greater Antillean auriculatus section, which comprises 44 species (Hedges 1989; the"auriculatus group" of Schwartz 1969), is supported by sixcharacteristics: (1) external submandibular vocal sac in males;(2) "patch-like" vomerine teeth; (3) areolate venter; (4) highlyvocal; (5) above-ground calling sites; and (6) prominent digitaldisks. However, the phylogenetic significance of these characters is still uncertain (Hedges 1989). Hedges (1989) also
2218 CAN. J. ZOOL. VOL. 72. 1994
TABLE I. Species of Eleurherodactylus in the Eastern Caribbean; all species have been considered members of the auriculatus section, martinicensis group, martinicensis series (Hedges 1989; Schwartz 1969), with E. urichi only tentatively included by Hedges (1989)
E. bar/agnei Lynch, 1965E. euphronides (Schwartz, 1967)E. johnstonei Barbour, 1914
E. martinicensis (Dumeril and Bibron, 1841)
E. pinchoni Schwartz, 1967E. shrevei (Schwartz, 1967)E. terraebolivaris Rivero, 1961E. urichi (Boettger, 1894)
Distribution
Basse-Terre, GuadeloupeGrenadaBermuda, Cura<,:ao, Jamaica, most
Lesser Antilles, Panama, VenezuelaDominica, Guadeloupe archipelago,
Martinique, St. BarthelemyBasse-Terre, GuadeloupeSt. VincentTobago, northern South AmericaTobago, Trinidad
References
Hardy 1985; Schwartz 1967Kaiser 1993: Kaiser et al. 1994aHardy and Harris 1979; Kaiser 1992. 1993
Kaiser 1992, 1993
Hardy 1985Kaiser 1993; Kaiser et al. 1994aHardy 1982Hardy 1982; Kaiser et al. 1994a: Kaiser 1993
TABLE 2. Allozyme loci diagnostic for Eleurherodactylus amplinympha n.sp., E. johnstonei, andE. martinicensis, and electrophoretic conditions employed in their resolution
Enzyme CommissionElectrophoretic
Proteina Locusa Number" conditionsC
I. Dipeptidase (leucylalanine) PEP (LA) 3.4.13.11 12. GlyceraJdehyde-3-phosphate dehydrogenase GAPDH 1.2. I. J2 23. Glucose dehydrogenase GCDH 1.1.1.118 14. Glucose-6-phosphate isomerase GPI 5.3.1.9 25. Isocitrate dehydrogenase (2 loci) IDH 1.1.1.42 I6. L-Lactate dehydrogenase (2 loci) LDH 1.1.1.27 27. Malate dehydrogenase (2 loci) MDH I. I. 1.37 I8. Mannose-6-phosphate isomerase (2 loci) MPI 5.3.1.8 19. Peptidase-B (L-Ieucylglycylglycine) PEP (LGG) 3.4.11.4 I
10. Phosphogluconate dehydrogenase PGDH 1.1.1.44 I
aAccording to the Nomenclature Committee of the International Union of Biochemistry (1984). modifiedaccording to Murphy et al. (1990).
hAccording to the Nomenclature Committee of the International Union of Biochemistry (1984).cl, Tris-citrate, pH 8.0, 130 Y, 4 h; 2, amine citrate, pH 6.1 (Clayton and Tretiak 1972), 75 mAo 4 h.
defined a martinicensis series (22 species) based on allozymedata (but lacking morphological synapomorphies) and postulated that Lesser Antillean Eleutherodactylus, possibly inclusive of E. urichi, were members of a monophyletic martinicensis group within the martinicensis series.
A detailed survey of Eastern Caribbean taxa was conductedto test the hypothesis of monophyly for Lesser Antillean taxa,and to determine whether there was any influx into the LesserAntilles from South America, as suggested by Lescure (1983,1987). The survey led to the realization that populations athigher elevations on the island of Dominica were distinct fromE. martinicensis at the species level. This discovery nowallows a more comprehensive analysis of phylogenetic relationships among Eastern Caribbean Eleutherodactylus.
Materials and methods
CollectionFrogs were collected in the Eastern Caribbean over a 4-year period
(1990-1993). Sampling on Dominica was carried out at six localities(Appendix I) in January 1990, during the month of August in 1990,1991, and 1992, and in October 1993. A conscious effort was madeto survey as many topographically dissimilar localities as feasibleduring a total of 21 days of surveying. For biochemical comparisons,specimens of Eleutherodactylusjohnstonei, E. marlinicensis, and theDominican Eleurherodaclylus in question were captured (Appendix2) and transported Jive to the laboratory at the Redpath Museum,Montreal. All procedures with animals, including care in captivity,
conformed to guidelines established by the Canadian Council onAnimal Care (1980-1984) and were approved by the Animal CareCommittee of McGill University. Institutional abbreviations followLeviton et al. (1985).
ElectrophoresisTissue samples (liver, heart, kidney, muscle, spleen) from 204
specimens were homogenized and centrifuged, and the supernatantwas stored separately from the original tissues at -80a C. This wasdone to prevent repetitive thawing and freezing of the original sample, thus reducing the loss of enzyme activity in the sometimes minutesamples. Horizontal starch gel electrophoresis was carried out toobtain allozyme data (Table 2) following the recommendations madeby Murphy et al. (1990).
Chromosome preparationOne male and female specimen each of E. johnstonei. E. martini
censis, and the Dominican Eleutherodactylus were used for chromosome preparation. Mitotic chromosomes were prepared directly frombone marrow after in vivo colchicine treatment. All techniques usedfor the preparation of cell suspensions, hypotonic treatment, and fixation of the cells have been described previously (Schmid 1978). Fivekaryotypes from each of the six animals were prepared.
MorphometricsFor morpohometric comparisons length measurements were taken
from 720 specimens (Table 3) to the nearest 0.1 mm using a dissecting microscope with camera lucida and digitizer attachments(Numonics 2200 digitizing tablet) and supported by Jandel ScientificSigma Scan (version 3.10) software on an IBM personal computer.

KAISER ET AL. 2219
Statistical analyses were performed using Systat 5.2 software on aMacintosh LC computer.
-.:::t -..- -..\0-..'!Xl '0\ '0o . '.0 . <"1
N-N<"1<"1'D1 +I I +1 1 +1r-lri'DOO~
r-N-.t-.tr-OO~ ~
OONO('t')(""'",~
-ONONO1 +I I +1 1 +1
-.to'D--.t<"10-:'0,,-,0 "-'
t- _N -..0-..Nf'!!Xl":""';'D
- NN<"11 +1 1 +1 1 +I
r- N V') '.0 0\ -.tV)eM~~-
N~MOO('fjS
-.tOlriO'D1 +11 +11 +1~'D~-OO'D
-~-c-c
~ ~
r--.tOOr-lriN<"1O-.tO'D1 +11 +11 +1
":Nc::r-f'!N-(:!.N~-C
o ;r; 0\ c;- \0 ;;;
-.t0V')0r-1+11+11+1f'!<"1~00'DV')
...... ~-~-~
~~~~~£-.t~~~-.to
0000001+11+11+1
-.to\NN<"1!Xl("f')OOvOClON<"1<"1-.t<"1-.t
°8°8°S
o-.~("I"'J8tr)6r- <"1 '.0 -.t lr) <"1lr)Olr)OlriO0000001+11+11+1
-<"1NO\V')r---.tr-'D-r-<"1-.t<"1-.t-.t'<t'
°8°2,°8
O-.N-.--... <"1 . r- . 0\-.t .00 ·N---NNrr.1 +11 +11 +1NO~O\OOOO
'De:,<"10r-N
ClE-
EOoI.I..
zU:J
ClU:J
~
,:j>enQ:i,....E-
...l>en
100
N
t
km
~arbados
o
Lesser Antilles
_Antigua
tSI.LUCia
.~. Vincent
Bequia.-• Mustjque
oUnion Is.•
o
,GrenadaA.
~ Martinique
~@
Anguilla" 62°00'W
,~ &!>.SI. Martin ""SI. Barthelemy"!i1
Saba. .. BarbudaSI. Eustatius ~~
SI.Kitts _Nevis
Montserrat'GUadelOu~eGrande-Terre@
(J La Desirade®IlliIO@)Basse-Terre .»
• Mane-Galante@@) Les Saint~g
\\Oominica\)®o
o E. barlagnei
@) E. martinicensis
o E. amplinympha
Iffil) E. pinchoni
/;:;. E. shrevei
A. E. euphronides
Character scoring and phylogenetic analysisCharacters were identified a priori using the character lists in the
dissertations of Joglar (1986) and Ford (1989), with several novelcharacters added (Appendix 3). Character states (Appendix 4) weredetermined during a preliminary survey of specimens and adjusted asappropriate during scoring. Not all of the described character states
FIG. I. Distribution of Eleutherodactylus species in the LesserAntilles. The distribution of E. johnstonei includes all shaded islands.
VocalizationsSound recordings were made at Morne Macaque on Dominica
(August 1990) and on Guadeloupe (January. June, and August 1990).Martinique (January and August 1990), and Montserrat (August1990) using a SONY Professional Walkman WM-D3. Not all of therecorded frogs were collected. and analyses were done on the clearestcalls. Audiospectrograms were created with a Kay Elemetrics Corp.digital sonagraph 7800. Terminology for vocalizations followsDuellman and Trueb (1986). and all means reported are for 10 calls.
DiagnosisThe format of the species diagnosis follows Lynch (1979). with the
addition of the condition of the m. depressor mandibulae (see Lynch1993) and the chromosome number. Descriptions of snout shape,structure of the vocal sac, and tongue shape follow the definitionsgiven by Duellman (1970). Terminology for finger disks followsSavage (1987). Measurements are given as ranges and means ± Istandard deviation, with sample sizes, for both female and males.
MorphologySeveral morphological characters were scored from live or freshly
preserved animals. or from photographs of living specimens. Specimens of potential out-groups were obtained from several NorthAmerican herpetological collections (Appendix I). Several specimens of each species under investigation were cleared and doublestained in consultation with the protocols of Hardaway and Williams(1975), Dingerkus and Uhler (1976), Wassersug (1976), and Hankenand Wassersug (1981).

2220 CAN. J. ZOOL. VOL. 72, 1994
FIG, 2. Female holotype of Eleutherodactylus amplinympha n,sp"KU 221260, 37.8 mm SVL.
may be present in the specics considered herc; they are included inthe character descriptions to permit comparisons and to facilitate subsequcnt inclusion of other taxa into the data set we present. Somecharacters are notoriously difficult to assess when one relies exclusively on preserved specimens, even when account descriptions fromthe litcrature are taken into account. Total character variability maynot be reflected by the individuals we cxamined (e.g., tuberculation,patterning, coloration). Thus, two analyses were carried out, oneusing the complete data matrix, the other excluding characters forwhich there was any scoring uncertainty (Appendix 3). The phylogenetic analysis was accomplished using PAUP 3.1.1 (Swofford1993) with option requests for out-group rooting (in-group monophyletic with respect to out-group), both ACCTRAN and DELrRAN
optimization, and unordered characters. Since the data matrix was toolarge for branch-and-bound or exhaustive searches, heuristic searchesand bootstrapping (1000 rcpetitions) were conducted using closeststepwise addition, uncollapsed zero-length branches, the steepestdescent option, and tree bisection - reconnection branch swappingwith swapping carried out on minimal and non-minimal trees. Bothstrict and majority-rule consensus cladograms were constructed anddisplayed as phylogenetic trees and phylograms. Exploratory manualbranch swapping was carried out on the most parsimonious topologies using MacClade 3.01 (Maddison and Maddison 1992), admittingonly combinations that did not increase the number of steps in thetree(s) found by parsimony analysis.
Out-group choiceAs with any phylogenetic analysis, choice of correct out-group(s)
and appropriate characters was of paramount importance. Theprimary concern with the Eastern Caribbean Eleutherodactylus species is the taxonomic uncertainty caused by their geographic positionbetween the Eleutherodactylus-rich faunas of the Greater Antilles andSouth America. As a consequence, an out-group analysis may be confounded either by the introduction of paraphyly if the chosen outgroup is really part of the in-group, or by omission of certain key taxafrom the in-group. Whereas the second problem defies solution at thistime, owing to the unresolved relationships between South AmericanEleutherodactylus, the first problem can be alleviated by consideringthe biochemical evidence presented by Kaiser et al. (I 994b), usingthe relationships postulated therein as a working hypothesis. Thus,southern Eastern Caribbean taxa are used here as out-groups in theanalysis of relationships of Puerto Rican and northern Eastern Caribbean taxa. One species, E. fitzingeri from Panama and Costa Rica,was added to the out-group because of its proposed close affinity withthe South American E. terraebolivaris (Rivero 1961).
Taxonomy
In the following section we describe a new species ofEleutherodactylus from forested habitats at higher elevationson Dominica, West Indies. A striking feature of these populations is the occurrence of females who attain, on occasion, abody size over twice that of males. For these distinctive populations we propose the name
Eleutherodactylus amplinympha n.sp.Dominica whistling frog, gounouj
Figs. 2 and 3
HOLOTYPE: The Natural History Museum, The University ofKansas (KU) 221259, an adult female from near FreshwaterLake, Dominica, West Indies (ca. 61°20'W, 15°20'N; elevation ca. 800 m). The specimen is one of a series collected on26 August 1992 by T.F. Sharbel and H. Kaiser.
PARATYPES: Two female topotypes (KU 221260-61), collectedon 7 August 1990 by G. Schafer, D. von Stosser, A. Werres,and H. Kaiser. Two male topotypes (KU 221262-63) and afemale topotype (KU 221264) collected on 21 August 1990 byH.M. Gray and H. Kaiser. One male topotype (KU 221265)and two female topotypes (KU 221266 -67) collected on7 August 1991 by H.M. Gray and H. Kaiser. Two males (KU221268-69) and two females (KU 221270-71) from nearEmerald Pool, elevation ca. 400 m, collected on 6 August1991 by H.M. Gray and H. Kaiser. One male (KU 221272)from 500 m SE Layou Park Estate, elevation ca. 325 m, collected on 13 January 1990 by H.H. Schwarten and H. Kaiser.Two females (KU 221273 -74) from the previous locality,collected on 8 August 1990 by G. Schafer, D. von StOsser,A. Werres, and H. Kaiser. One male (KU 221275) from theTrafalgar Falls area, elevation ca. 330 m, collected on14 January 1990 by H.H. Schwarten and H. Kaiser. Onefemale (KU 221276) from the slopes of Morne Diablotin, elevation ca. 1000 m, collected along trail on 22 August 1990 byH.M. Gray and H. Kaiser.
DiagnosisA forest-dwelling member of the auriculatus section
(Hedges 1989) with the following diagnostic features: (\) skinon dorsum coarsely shagreen with decreasing number oflarger tubercles from anterior to posterior; a fine middorsalridge extending from back of head to venter; dorsolateral foldsabsent; venter coarsely areolate between pectoral and pelvicareas; groin region coarsely areolate; (2) tympanum round,distinct, about one-third size of the orbit; supratympanic foldpresent; (3) snout trapezoid in dorsal view, rounded in profile;eye - naris distance greater than length of eye; canthus rostralis sharply angled, canthal ridge straight, with dark line;(4) interorbital distance 1. 5 - 2 times width of upper eyelid;supraocular tubercles present; cranial crests absent; (5) dentigerous processes of vomers triangular and slightly oblique,each with a single row of teeth; choanae ovoid; (6) males withexternal vocal slits and weakly bilobate subgular vocal sac;nuptial pads absent; (7) size of fingers (I = II) < IV < III,III about 1.5 times longer than I; finger disks II - IV about 1.5times wider than digits, disk I only slightly wider; subarticulartubercles round and raised; two palmar tubercles, medial oneelliptical, lateral one conical; thenar tubercle elliptical, covering base of finger I laterally; numerous supernumerary palmartubercles; (8) fingers with weak lateral fringes; (9) severaltubercles on forearm and elbow; several raised postorbitaltubercles, particularly in the area from angle of jaw to axil;
KAISER ET AL.
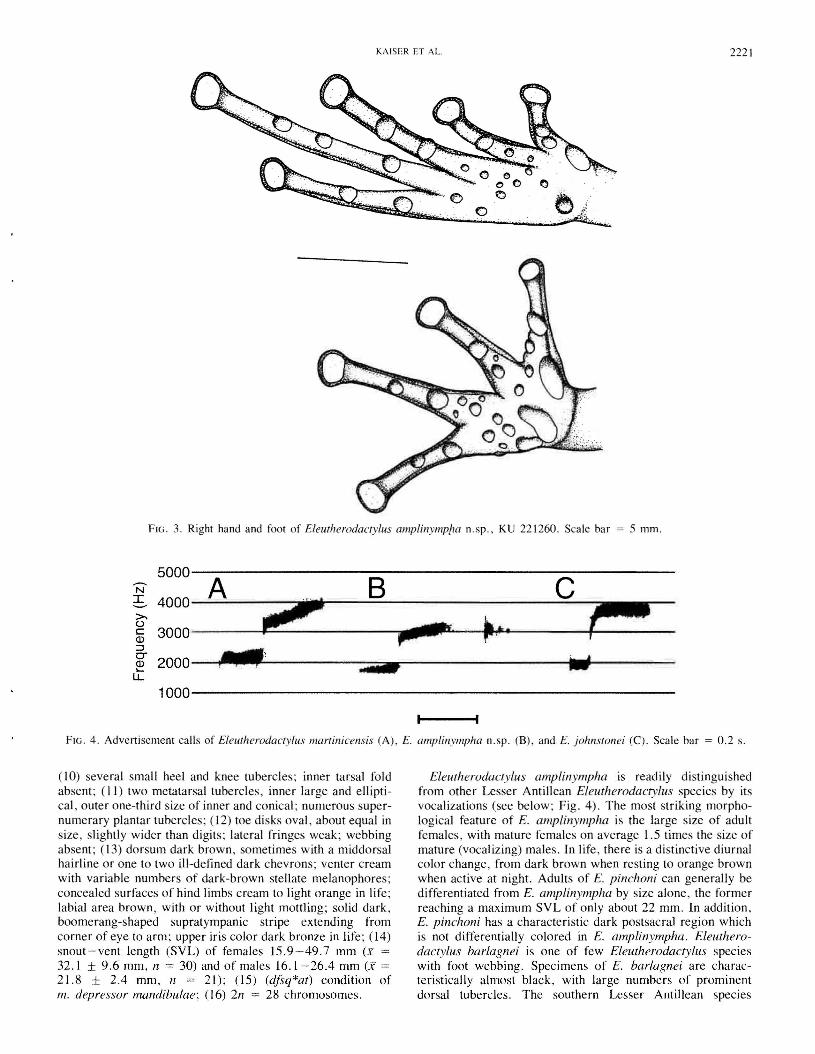
FIG. 3. Right hand and foot of Eleutherodactylus amplinymp!w n.sp., KU 221260. Scale bar = 5 mm.
5000A B C........
N:r: 4000 til'".......- ,...-->-
~~'U •c 3000· pCCD:::J
? .~CJ 2000 lidCD ! ; . •....u..
1000
2221
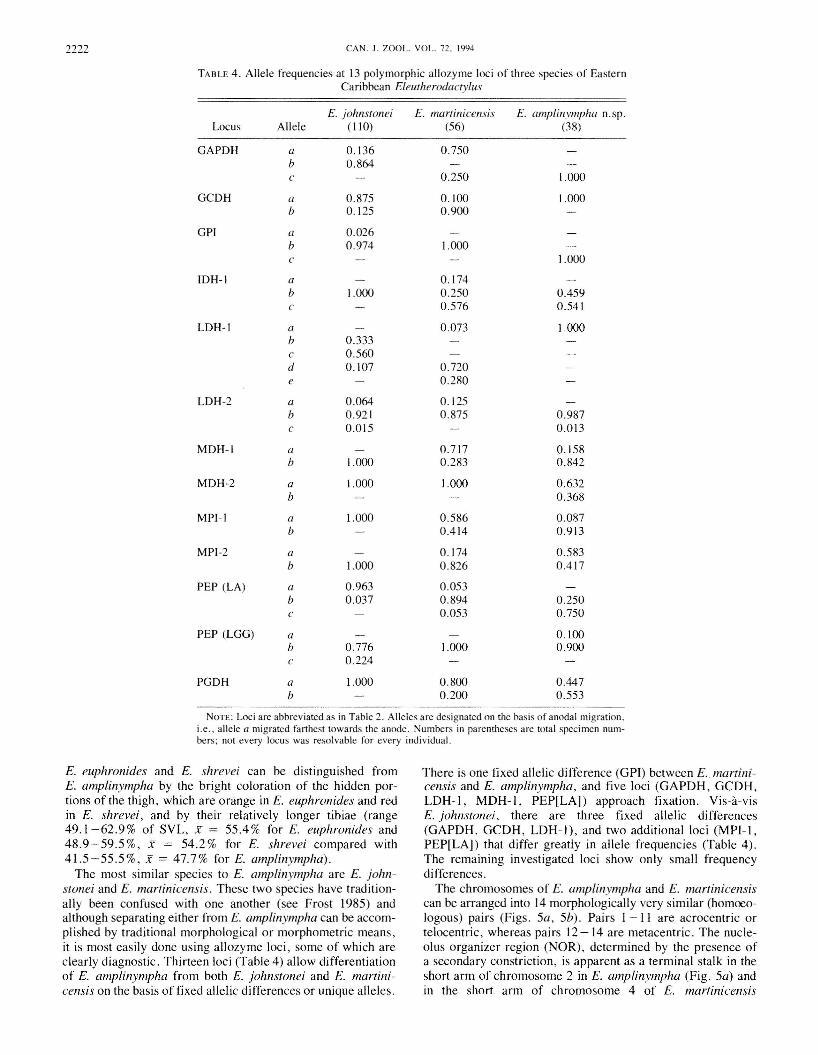
FIG. 4. Advertisement calls of Eleulherodactylus martinicensis (A), E. amplinympha n.sp. (B), and E. johnslOnei (C). Scale bar = 0.2 s.
(10) several small heel and knee tubercles; inner tarsal foldabsent; (II) two metatarsal tubercles, inner large and elliptical, outer one-third size of inner and conical; numerous supernumerary plantar tubercles; (12) toe disks oval, about equal insize, slightly wider than digits; lateral fringes weak; webbingabsent; (13) dorsum dark brown, sometimes with a middorsalhairline or one to two ill-defined dark chevrons; venter creamwith variable numbers of dark-brown stellate melanophores;concealed surfaces of hind limbs cream to light orange in life;labial area brown, with or without light mottling; solid dark,boomerang-shaped supratympanic stripe extending fromcorner of eye to arm; upper iris color dark bronze in life; (14)snout-vent length (SYL) of females 15.9-49.7 mm (x =32.1 ± 9.6 mm, n = 30) and of males 16.1-26.4 mm (x =21.8 ± 2.4 mm, n = 21); (15) (dfsq*at) condition ofm. depressor mandibulae; (16) 2n = 28 chromosomes.
Eleutherodactylus amplinympha is readily distinguishedfrom other Lesser Antillean Eleutherodactylus species by itsvocalizations (see below; Fig. 4). The most striking morphological feature of E. amplinympha is the large size of adultfemales, with mature females on average 1.5 times the size ofmature (vocalizing) males. In life, there is a distinctive diurnalcolor change, from dark brown when resting to orange brownwhen active at night. Adults of £. pinchoni can generally bedifferentiated from E. amplinympha by size alone, the formerreaching a maximum SYL of only about 22 mm. In addition,£. pinchoni has a characteristic dark postsacral region whichis not differentially colored in E. amplinympha. Eleutherodactylus barlagnei is one of few Eleutherodactylus specieswith foot webbing. Specimens of E. barlagnei are characteristically almost black, with large numbers of prominentdorsal tubercles. The southern Lesser Antillean species
2222 CAN. J. ZOOL. VOL. 72, 1994
TABLE 4. Allele frequencies at 13 polymorphic allozyme loci of three species of EasternCaribbean Eleutherodactylus
E. johnstonei E. martinicensis E. amplinympha n.sp.Locus Allele (110) (56) (38)
GAPDH a 0.136 0.750b 0.864c 0.250 1.000
GCDH a 0.875 0.100 1.000b 0.125 0.900
GPI a 0.026b 0.974 1.000c 1.000
IDH-I a 0.174b 1.000 0.250 0.459c 0.576 0.541
LDH-I a 0.073 1.000b 0.333c 0.560d 0.107 0.720e 0.280
LDH-2 a 0.064 0.125b 0.921 0.875 0.987c 0.015 0.013
MDH-I a 0.717 0.158b 1.000 0.283 0.842
MDH-2 a 1.000 1.000 0.632b 0.368
MPI-l a 1.000 0.586 0.087b 0.414 0.913
MPI-2 a 0.174 0.583b 1.000 0.826 0.417
PEP (LA) a 0.963 0.053b 0.037 0.894 0.250c 0.053 0.750
PEP (LGG) a 0.100b 0.776 1.000 0.900c 0.224
PGDH a 1.000 0.800 0.447b 0.200 0.553
NOTE: Loci are abbreviated as in Table 2. Alleles are designated on the basis of anodal migration.i.e., allele a migrated farthest towards the anode. Numbers in parentheses are total specimen num-bers; not every locus was resolvable for every individual.
E. euphronides and E. shrevei can be distinguished fromE. amplinympha by the bright coloration of the hidden portions of the thigh, which are orange in E. euphronides and redin E. shrevei, and by their relatively longer tibiae (range49.1-62.9% of SVL, X = 55.4% for E. euphronides and48.9-59.5%, X = 54.2% for E. shrevei compared with41.5-55.5%, X = 47.7% for E. amplinympha).
The most similar species to E. amplinympha are E. Johnstonei and E. martinicensis. These two species have traditionally been confused with one another (see Frost 1985) andalthough separating either from E. amplinympha can be accomplished by traditional morphological or morphometric means,it is most easily done using allozyme loci, some of which areclearly diagnostic. Thirteen loci (Table 4) allow differentiationof E. amplinympha from both E. Johnstonei and E. martinicensis on the basis offixed allelic differences or unique alleles.
There is one fixed allelic difference (GPI) between E. martinicensis and E. amplinympha, and five loci (GAPDH, GCDH,LDH-l, MDH-I, PEP[LA]) approach fixation. Vis-a-visE. Johnstonei, there are three fixed allelic differences(GAPDH, GCDH, LDH-l), and two additional loci (MPI-I,PEP[LA]) that differ greatly in allele frequencies (Table 4).The remaining investigated loci show only small frequencydifferences.
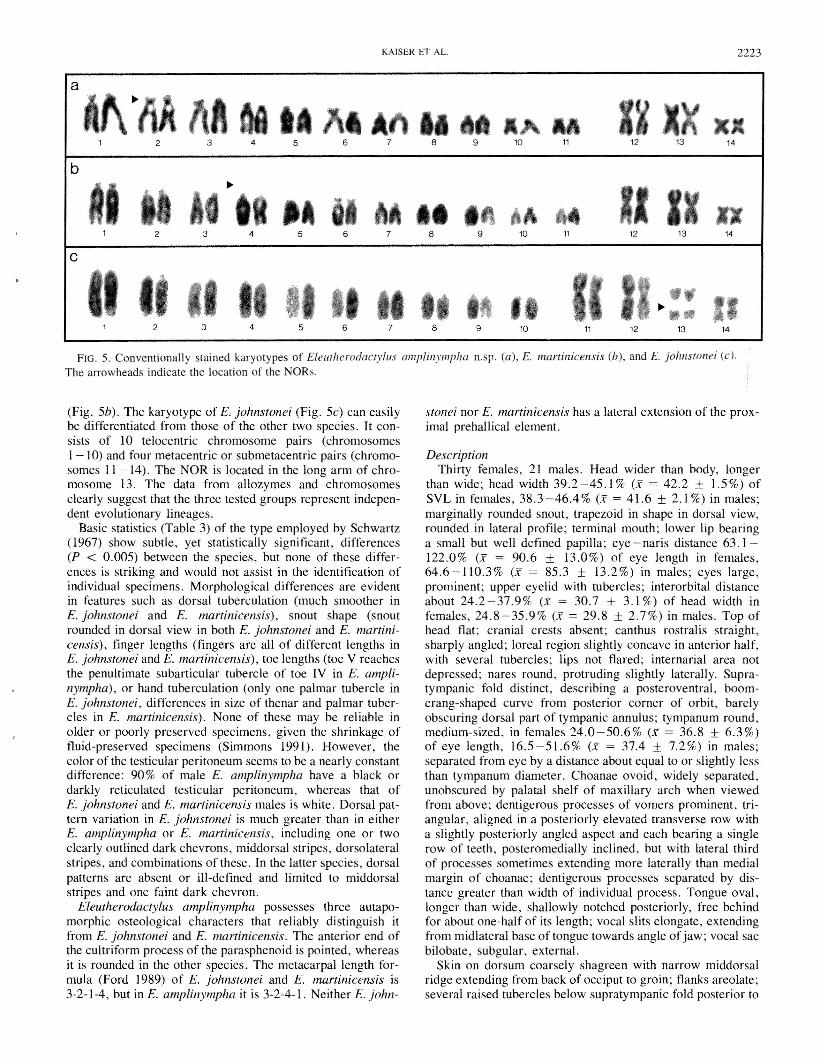
The chromosomes of E. amplinympha and E. martinicensiscan be arranged into 14 morphologically very similar (homoeologous) pairs (Figs. 5a, 5b). Pairs I -II are acrocentric ortelocentric, whereas pairs 12-14 are metacentric. The nucleolus organizer region (NOR), determined by the presence ofa secondary constriction, is apparent as a terminal stalk in theshort arm of chromosome 2 in E. amplinympha (Fig. 5a) andin the short arm of chromosome 4 of E. martinicensis

2224 CAN. J. ZOOL. VOL. 72, 1994
tympanum; several low tubercles on forearm; several smalltubercles on each knee and heel but not on tarsus; ventralposterior surface of thighs coarsely areolate. Anal openingunmodified, directed posteriorly at upper level of thighs.
Forearms moderately robust; fingers long, slender, bearingsubtruncate disks with broadly elliptical pads, relative disksizes I < II < (III = IV); relative lengths of fingers (l = II)< IV < III; number of subarticular tubercles 1-2-2-2 forfingers I - IV, respectively, subarticular tubercles round andraised; numerous supernumerary palmar tubercles; twopalmar tubercles, medial one elliptical, lateral one conical;thenar tubercle elliptical, covering base of finger I laterally;nuptial pads absent. Hind limbs moderately robust, long; heelsbroadly overlapping when hind limbs flexed at right angles tobody axis; tibia length in females 41.5-55.3% (X = 47.4 ±3.0%) of SVL, 43.8-55.5% (X = 48.2 ± 2.9%) in males.Inner tarsal fold absent; two metatarsal tubercles, inner largeand elliptical, outer one-third size of inner and conical; toeslong, slender, bearing oval disks about the size of disks onfingers III and IV; with narrow lateral fringes, without anywebbing; relative length of toes I < II < V < III < IV; number of subarticular tubercles 1-1-2-3-2 for toes I - V, respectively, subarticular tubercles round and conical; numeroussupernumerary plantar tubercles (Fig. 3).
CoLor in preservative (n = 5/)Dorsum of head and body uniformly dark brown; 27.5 % of
specimens without any dorsal pattern, 17.6% with light middorsal hairline, 11.8% with light middorsal stripe, 21.6% withone ill-defined dark middorsal chevron, 13.7% with a lightdorsolateral area; with narrow dark interorbital bar, 7.8%having a cream interocular bar offsetting the former; dark canthal stripe; lower edge of supratympanic stripe dark brown;flanks usually dark brown, rarely lighter than middorsal area(two individuals). Dorsal surfaces of limbs dark brown, withor without 1 or 2 darker brown crossbars, sometimes offset bylighter borders (first and second numbers in parentheses arepercentages of occurrence of 1 and 2 crossbars, respectively)on forearms (91.5,3.4), thighs (64.4,5.1), shanks (91.5, 5.1,diagonal), and tarsi (74.6, 1.7); anterior surface of thighs tanand mottled, posterior surfaces tan. Venter cream to tan withsome mottling caused by differential distribution of darkbrown stellate melanophores; ventral surface of palm andfinger disks white, disk covers brown with the exception ofconspicuously darkly pigmented disk cover on finger IV; toedisks white ventrally, disk covers darkly pigmented; plantarsurfaces dark brown, sometimes offset by a medial creamhairline.
CoLor in life (n = 5)Dorsum dark brown by day, taking on a distinctly orange
hue during night activity; venter cream to tan with somedegree of mottling; hidden surfaces of thighs cream to faintorange; upper iris color bronze.
Measurements (in millimetres)Values given are for the holotype, followed by ranges, then
means ± SD in parentheses, first for 30 females, then for 21males. SVL 37.8, 15.9 -49.7 (32.1 ± 9.6), 16.1-26.4(21.8 ± 2.4); tibia length 19.3, 7.9-22.1 (15.1 ± 4.1),7.8 -13.3 (l0.5 ± 1.2); foot length 27.7, 10.6- 33.6 (21.5 ±6.3), 10.6-18.5 (14.5 ± 1.6); head width 11.8, 6.8-21.8(13.6 ± 4.3), 7.5-10.9 (9,0 ± 0.9); interorbital distance 5.1,2.2-7.6 (4,2 ± 1.4), 1.9-3.2 (2.7 ± 0.3); eye-naris dis-
tance 4.9, 1.6-6.5 (3.8 ± 1.3), 1.9-3.4 (2.6 ± 0.3); eyediameter 5.1, 2.4-6.3 (4.2 ± 1.1),2.4-3.7 (3.1 ± 0.4);tympanum diameter 1.8, 0.7-2.3 (1.5 ± 0.5), 0.6-1.6(1.1 ± 0.2).
Distribution and ecologyThe species is found only on the island of Dominica, West
Indies. It is uncertain at this time whether the population iscontinuous or fragmented, because there has been somedevelopment of broad (up to 500 m) agricultural stripsparalleling either side of the main roads traversing the island.Frogs were most abundant in the area of Morne Macaque inMorne Trois Pitons National Park (density ca. 18 frogs/m2 ;
H. Kaiser and R. Wagenseil, unpublished data). Despite theestablishment of the park, the area around Freshwater Lakehas recently suffered as a result of construction of a hydroelectric development. Tarred wooden pipes have been constructedalong a 10-15 m wide deforested and leveled corridor alongside the mountain; chemically treated wood and metal debrishas been discarded into the surrounding forest. However, theGovernment of Dominica is conscious of its natural resourcesand has established precedents (e.g., hunting seasons for theedible frog Leptodactylus faUax, restraint in construction oftourist facilities in favor of locally controlled ecotourism) inthe Lesser Antilles for responsible use and management of itsunique biota.
EleutherodactyLus amplinympha is most abundant near thetransition zone from "montane rainforest" to "elfin woodland" (sensu Nicolson 1991). This habitat is characterized bya much greater temperature variation between day and night(from as high as 25 °C to as low as IrC) than the adjacentlowland habitats, by high annual rainfall, and by nearly ubiquitous fog. During all visits to the Freshwater Lake area,H.K. encountered either rain or fog, with high gusting windsat night often preventing useable recording of vocalizations.The montane rain forest vegetation at this elevation consists offew trees (height <20 m), shrub thickets, palm brakes, andferns (Davis et a1. 1986; Nicolson 1991). The low, thicket-likeforest, frequently covered by epiphyllous hepatics or bearingmoist moss mats (Nicolson 1991), is an ideal refugium foranurans and nearly impenetrable to humans.
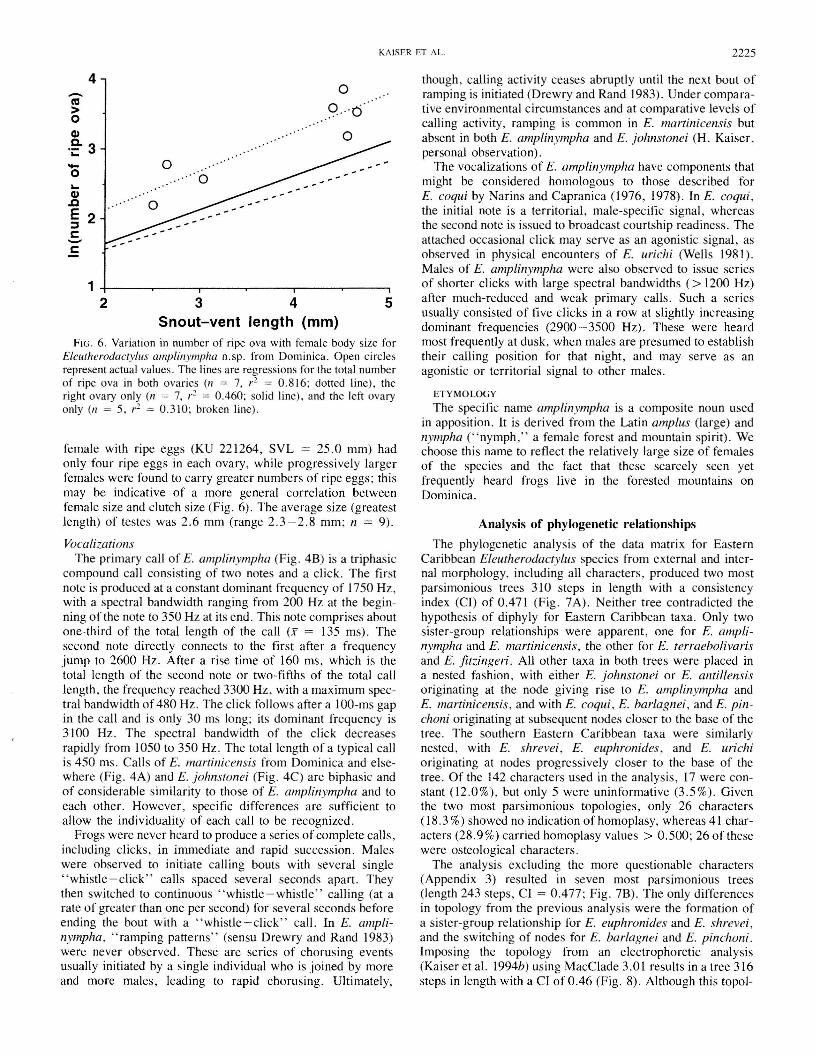
A single terrestrial egg mass was found in January 1990 ina rock crevice near Freshwater Lake. It contained 13 firmopaque eggs (estimated maximum diameter 7 mm) with anouter gelatinous layer, positioned in a three-dimensionalclump and attended by a male E. amplinympha (SVL ca.25 mm). The total size of the egg clump was about twice thatof the attending frog. Eggs were positioned on a thin mat ofground moss. After collection, no changes in egg morphologyoccurred, and dissection of several eggs showed no recognizable development (D.S. Townsend, personal communication).Seven females (SVL range 25.0-46.5 mm, X = 37.6 mm)caught during a reproductive episode, as determined by thepresence oflarge and yolky (= ripe) ovarian eggs, contained,on average, 29.3 eggs, 14.7 in the right ovary (n = 7) and14.6 in the left ovary (n = 5). The two females whose leftovary was not included in the mean calculations had few or noeggs in it; one carried 32 ripe eggs in her right ovary and4 in the left, and the other, smaller female carried 13 in theright ovary and none in the left. Such an imbalance suggeststhat these females may lay the total or partial contents of oneovary at a time, allowing them to diversify their fitness byproducing clutches fertilized by different males. The smallest
KAISER ET AL. 2223
a
JC2 3 4 5 6 7 8 9 10 11 12 13 14
b~ ,
" A2 3 4 5 6 7 8 9 10 11 12 13 14
C,.
I I I - - I'I.
I~.ll1f2 3 4 5 6 7 8 9 10 11 12 13 14
FIG. 5. Conventionally stained karyotypes of Eleutherodactyllis amplinymphll n.sp. (a), E. marrinicensis (b), and E. johnstol1ei (c).The arrowheads indicate the location of the NORs.
(Fig. 5b). The karyotype of E. johnsfonei (Fig. 5c) can easilybe differentiated from those of the other two species. It consists of 10 telocentric chromosome pairs (chromosomes1-10) and four metacentric or submetacentric pairs (chromosomes 11-14). The NOR is located in the long arm of chromosome 13. The data from allozymes and chromosomesclearly suggest that the three tested groups represent independent evolutionary lineages.
Basic statistics (Table 3) of the type employed by Schwartz(\ 967) show subtle, yet statistically significant, differences(P < 0.005) between the species, but none of these differences is striking and would not assist in the identification ofindividual specimens. Morphological differences are evidentin features such as dorsal tuberculation (much smoother inE. johnsronei and E. martinicensis), snout shape (snoutrounded in dorsal view in both E. johnstonei and E. marfinicensis) , finger lengths (fingers are all of different lengths inE. johnstonei and E. martinicensis), toe lengths (toe V reachesthe penultimate subarticular tubercle of toe IV in E. amplinympha), or hand tuberculation (only one palmar tubercle inE. johnsfonei, differences in size of thenar and palmar tubercles in E. martinicensis). None of these may be reliable inolder or poorly preserved specimens, given the shrinkage offluid-preserved specimens (Simmons 1991). However, thecolor of the testicular peritoneum seems to be a nearly constantdifference: 90% of male E. amplinympha have a black ordarkly reticulated testicular peritoneum, whereas that ofE. johnstonei and E. martinicensis males is white. Dorsal pattern variation in E. johnstonei is much greater than in eitherE. amplinympha or E. marfinicensis, including one or twoclearly outlined dark chevrons, middorsal stripes, dorsolateralstripes, and combinations of these. In the latter species, dorsalpatterns are absent or ill-defined and limited to middorsalstripes and one faint dark chevron.
Eleutherodactylus amplinympha possesses three autapomorphic osteological characters that reliably distinguish itfrom E. johnstonei and E. martinicensis. The anterior end ofthe cultriform process of the parasphenoid is pointed, whereasit is rounded in the other species. The metacarpal length formula (Ford 1989) of E. johnsronei and E. martinicensis is3-2-1-4, but in E. amplinympha it is 3-2-4-1. Neither E. john-
sronei nor E. martinicensis has a lateral extension of the proximal prehallical element.
DescriptionThirty females, 21 males. Head wider than body, longer
than wide; head width 39.2-45.1 % (x = 42.2 ± 1.5%) ofSVL in females, 38.3-46.4% (x = 41.6 ± 2.1 %) in males;marginally rounded snout, trapezoid in shape in dorsal view,rounded in lateral profile; terminal mouth; lower lip bearinga small but well defined papilla; eye - naris distance 63.1 122.0% (x = 90.6 ± 13.0%) of eye length in females,64.6-110.3% (x = 85.3 ± 13.2%) in males; eyes large,prominent; upper eyelid with tubercles; interorbital distanceabout 24.2-37.9% (x = 30.7 ± 3.1 %) of head width infemales, 24.8-35.9% (x = 29.8 ± 2.7%) in males. Top ofhead flat; cranial crests absent; canthus rostralis straight,sharply angled; loreal region slightly concave in anterior half,with several tubercles; lips not flared; internarial area notdepressed; nares round, protruding slightly laterally. Supratympanic fold distinct, describing a posteroventral, boomerang-shaped curve from posterior corner of orbit, barelyobscuring dorsal part of tympanic annulus; tympanum round,medium-sized, in females 24.0-50.6% (x = 36.8 ± 6.3%)of eye length, 16.5-51.6% (x = 37.4 ± 7.2%) in males;separated from eye by a distance about equal to or slightly lessthan tympanum diameter. Choanae ovoid, widely separated,unobscured by palatal shelf of maxillary arch when viewedfrom above; dentigerous processes of vomers prominent, triangular, aligned in a posteriorly elevated transverse row witha slightly posteriorly angled aspect and each bearing a singlerow of teeth, posteromedially inclined, but with lateral thirdof processes sometimes extending more laterally than medialmargin of choanae; dentigerous processes separated by distance greater than width of individual process. Tongue oval,longer than wide, shallowly notched posteriorly, free behindfor about one-half of its length; vocal slits elongate, extendingfrom midlateral base of tongue towards angle of jaw; vocal sacbilobate, subgular, external.
Skin on dorsum coarsely shagreen with narrow middorsalridge extending from back of occiput to groin; flanks areolate;several raised tubercles below supratympanic fold posterior to

FIG. 5. Conventionally stained karyotypes of Eleutherodactylus amplillympha n.sp. (a), E. martillicellsis (b), and E. jO!tllstollei (c).The arrowheads indicate the location of the NORs.
(Fig. 5b). The karyotype of E. johnstonei (Fig. 5c) can easilybe differentiated from those of the other two species. It consists of 10 telocentric chromosome pairs (chromosomes1-10) and four metacentric or submetacentric pairs (chromosomes 11-14). The NOR is located in the long arm of chromosome 13. The data from allozymes and chromosomesclearly suggest that the three tested groups represent independent evolutionary lineages.
Basic statistics (Table 3) of the type employed by Schwartz(1967) show subtle, yet statistically significant, differences(P < 0.005) between the species, but none of these differences is striking and would not assist in the identification ofindividual specimens. Morphological differences are evidentin features such as dorsal tuberculation (much smoother inE. johnstonei and E. martinicensis) , snout shape (snoutrounded in dorsal view in both E. johnstonei and E. martinicensis), finger lengths (fingers are all of different lengths inE. johnstonei and E. martinicensis), toe lengths (toe V reachesthe penultimate subarticular tubercle of toe IV in E. amplinympha), or hand tuberculation (only one palmar tubercle inE. johnstonei, differences in size of thenar and palmar tubercles in E. martinicensis). None of these may be reliable inolder or poorly preserved specimens, given the shrinkage offluid-preserved specimens (Simmons 1991). However, thecolor of the testicular peritoneum seems to be a nearly constantdifference: 90% of male E. amplinympha have a black ordarkly reticulated testicular peritoneum, whereas that ofE. johnstonei and E. martinicensis males is white. Dorsal pattern variation in E. johnstonei is much greater than in eitherE. amplinympha or E. martinicensis, including one or twoclearly outlined dark chevrons, middorsal stripes, dorsolateralstripes, and combinations of these. In the latter species, dorsalpatterns are absent or ill-defined and limited to middorsalstripes and one faint dark chevron.
Eleutherodactylus amplinympha possesses three autapomorphic osteological characters that reliably distinguish itfrom E. johnstonei and E. martinicensis. The anterior end ofthe cultriform process of the parasphenoid is pointed, whereasit is rounded in the other species. The metacarpal length formula (Ford 1989) of E. johnstonei and E. martinicensis is3-2-1-4, but in E. amplinympha it is 3-2-4-1. Neither E. john-
stonei nor E. martinicensis has a lateral extension of the proximal prehallical element.
DescriptionThirty females, 21 males. Head wider than body, longer
than wide; head width 39.2-45.1 % (X = 42.2 ± 1.5%) ofSVL in females, 38.3-46.4% (X = 41.6 ± 2.1 %) in males;marginally rounded snout, trapezoid in shape in dorsal view,rounded in lateral profile; terminal mouth; lower lip bearinga small but well defined papilla; eye-naris distance 63.1122.0% (x = 90.6 ± 13.0%) of eye length in females,64.6-110.3% (x = 85.3 ± 13.2%) in males; eyes large,prominent; upper eyelid with tubercles; interorbital distanceabout 24.2-37.9% (x = 30.7 ± 3.1%) of head width infemales, 24.8-35.9% (X = 29.8 ± 2.7%) in males. Top ofhead flat; cranial crests absent; canthus rostralis straight,sharply angled; loreal region slightly concave in anterior half,with several tubercles; lips not flared; internarial area notdepressed; nares round, protruding slightly laterally. Supratympanic fold distinct, describing a posteroventral, boomerang-shaped curve from posterior corner of orbit, barelyobscuring dorsal part of tympanic annulus; tympanum round,medium-sized, in females 24.0-50.6% (X = 36.8 ± 6.3%)of eye length, 16.5-51.6% (x = 37.4 ± 7.2%) in males;separated from eye by a distance about equal to or slightly lessthan tympanum diameter. Choanae ovoid, widely separated,unobscured by palatal shelf of maxillary arch when viewedfrom above; dentigerous processes of vomers prominent, triangular, aligned in a posteriorly elevated transverse row witha slightly posteriorly angled aspect and each bearing a singlerow of teeth, posteromedially inclined, but with lateral thirdof processes sometimes extending more laterally than medialmargin of choanae; dentigerous processes separated by distance greater than width of individual process. Tongue oval,longer than wide, shallowly notched posteriorly, free behindfor about one-half of its length; vocal slits elongate, extendingfrom midlateral base of tongue towards angle ofjaw; vocal sacbilobate, subgular, external.
Skin on dorsum coarsely shagreen with narrow middorsalridge extending from back of occiput to groin; flanks areolate;several raised tubercles below supratympanic fold posterior to

2224 CAN. J. ZOOL. VOL. 72, 1994
tympanum; several low tubercles on forearm; several smalltubercles on each knee and heel but not on tarsus; ventralposterior surface of thighs coarsely areolate. Anal openingunmodified, directed posteriorly at upper level of thighs.
Forearms moderately robust; fingers long, slender, bearingsubtruncate disks with broadly elliptical pads, relative disksizes I < II < (III = IV); relative lengths of fingers (I = II)< IV < III; number of subarticular tubercles 1-2-2-2 forfingers I - IV, respectively, subarticular tubercles round andraised; numerous supernumerary palmar tubercles; twopalmar tubercles, medial one elliptical, lateral one conical;thenar tubercle elliptical, covering base of finger I laterally;nuptial pads absent. Hind limbs moderately robust, long; heelsbroadly overlapping when hind limbs flexed at right angles tobody axis; tibia length in females 41.5-55.3% (x = 47.4 ±3.0%) of SVL, 43.8-55.5% (x = 48.2 ± 2.9%) in males.Inner tarsal fold absent; two metatarsal tubercles, inner largeand elliptical, outer one-third size of inner and conical; toeslong, slender, bearing oval disks about the size of disks onfingers III and IV; with narrow lateral fringes, without anywebbing; relative length of toes I < II < V < III < IV; number of subarticular tubercles 1-1-2-3-2 for toes I - V, respectively, subarticular tubercles round and conical; numeroussupernumerary plantar tubercles (Fig. 3).
Color in preservative (n = 51)Dorsum of head and body uniformly dark brown; 27.5% of
specimens without any dorsal pattern, 17.6% with light middorsal hairline, 11.8 % with light middorsal stripe, 21.6 % withone ill-defined dark middorsal chevron, 13.7 % with a lightdorsolateral area; with narrow dark interorbital bar, 7.8 %having a cream interocular bar offsetting the former; dark canthal stripe; lower edge of supratympanic stripe dark brown;flanks usually dark brown, rarely lighter than middorsal area(two individuals). Dorsal surfaces of limbs dark brown, withor without 1 or 2 darker brown crossbars, sometimes offset bylighter borders (first and second numbers in parentheses arepercentages of occurrence of 1 and 2 crossbars, respectively)on forearms (91.5, 3.4), thighs (64.4, 5.1), shanks (91.5,5.1,diagonal), and tarsi (74.6, 1.7); anterior surface of thighs tanand mottled, posterior surfaces tan. Venter cream to tan withsome mottling caused by differential distribution of darkbrown stellate melanophores; ventral surface of palm andfinger disks white, disk covers brown with the exception ofconspicuously darkly pigmented disk cover on finger IV; toedisks white ventrally, disk covers darkly pigmented; plantarsurfaces dark brown, sometimes offset by a medial creamhairline.
Color in life (n = 5)Dorsum dark brown by day, taking on a distinctly orange
hue during night activity; venter cream to tan with somedegree of mottling; hidden surfaces of thighs cream to faintorange; upper iris color bronze.
Measurements (in millimetres)Values given are for the holotype, followed by ranges, then
means ± SD in parentheses, first for 30 females, then for 21males. SVL 37.8, 15.9-49.7 (32.1 ± 9.6), 16.1-26.4(21.8 ± 2.4); tibia length 19.3,7.9-22.1 (15.1 ± 4.1),7.8-13.3 (10.5 ± 1.2); foot length 27.7, 10.6-33.6 (21.5 ±6.3), 10.6-18.5 (14.5 ± 1.6); head width 11.8,6.8-21.8(13.6 ± 4.3), 7.5 -10.9 (9.0 ± 0.9); interorbital distance 5.1,2.2-7.6 (4.2 ± 1.4), 1.9-3.2 (2.7 ± 0.3); eye-naris dis-
tance 4.9, 1.6-6.5 (3.8 ± 1.3), 1.9-3.4 (2.6 ± 0.3); eyediameter 5.1,2.4-6.3 (4.2 ± 1.1),2.4-3.7 (3.1 ± 0.4);tympanum diameter 1.8, 0.7-2.3 (1.5 ± 0.5), 0.6-1.6(1.1 ± 0.2).
Distribution and ecologyThe species is found only on the island of Dominica, West
Indies. It is uncertain at this time whether the population iscontinuous or fragmented, because there has been somedevelopment of broad (up to 500 m) agricultural stripsparalleling either side of the main roads traversing the island.Frogs were most abundant in the area of Morne Macaque inMorne Trois Pitons National Park (density ca. 18 frogs/m2;
H. Kaiser and R. Wagenseil, unpublished data). Despite theestablishment of the park, the area around Freshwater Lakehas recently suffered as a result of construction of a hydroelectric development. Tarred wooden pipes have been constructedalong a 10-15 m wide deforested and leveled corridor alongside the mountain; chemically treated wood and metal debrishas been discarded into the surrounding forest. However, theGovernment of Dominica is conscious of its natural resourcesand has established precedents (e.g., hunting seasons for theedible frog Leptodactylus fallax, restraint in construction oftourist facilities in favor of locally controlled ecotourism) inthe Lesser Antilles for responsible use and management of itsunique biota.
Eleutherodactylus amplinympha is most abundant near thetransition zone from "montane rainforest" to "elfin woodland" (sensu Nicolson 1991). This habitat is characterized bya much greater temperature variation between day and night(from as high as 25°C to as low as Irq than the adjacentlowland habitats, by high annual rainfall, and by nearly ubiquitous fog. During all visits to the Freshwater Lake area,H.K. encountered either rain or fog, with high gusting windsat night often preventing useable recording of vocalizations.The montane rain forest vegetation at this elevation consists offew trees (height < 20 m), shrub thickets, palm brakes, andferns (Davis et al. 1986; Nicolson 1991). The low, thicket-likeforest, frequently covered by epiphyllous hepatics or bearingmoist moss mats (Nicolson 1991), is an ideal refugium foranurans and nearly impenetrable to humans.
A single terrestrial egg mass was found in January 1990 ina rock crevice near Freshwater Lake. It contained 13 firmopaque eggs (estimated maximum diameter 7 mm) with anouter gelatinous layer, positioned in a three-dimensionalclump and attended by a male E. amplinympha (SVL ca.25 mm). The total size of the egg clump was about twice thatof the attending frog. Eggs were positioned on a thin mat ofground moss. After collection, no changes in egg morphologyoccurred, and dissection of several eggs showed no recognizable development (D.S. Townsend, personal communication).Seven females (SVL range 25.0-46.5 mm, x = 37.6 nun)caught during a reproductive episode, as determined by thepresence of large and yolky (= ripe) ovarian eggs, contained,on average, 29.3 eggs, 14.7 in the right ovary (n = 7) and14.6 in the left ovary (n = 5). The two females whose leftovary was not included in the mean calculations had few or noeggs in it; one carried 32 ripe eggs in her right ovary and4 in the left, and the other, smaller female carried 13 in theright ovary and none in the left. Such an imbalance suggeststhat these females may lay the total or partial contents of oneovary at a time, allowing them to diversify their fitness byproducing clutches fertilized by different males. The smallest
KAISER ET AL. 2225
though, calling activity ceases abruptly until the next bout oframping is initiated (Drewry and Rand 1983). Under comparative environmental circumstances and at comparative levels ofcalling activity, ramping is common in E. martinicensis butabsent in both E. amplinympha and E. johnstonei (H. Kaiser,personal observation).
The vocalizations of E. amplinympha have components thatmight be considered homologous to those described forE. coqui by Narins and Capranica (1976, 1978). In E. coqui,the initial note is a territorial, male-specific signal, whereasthe second note is issued to broadcast courtship readiness. Theattached occasional click may serve as an agonistic signal, asobserved in physical encounters of E. urichi (Wells 1981).Males of E. amplinympha were also observed to issue seriesof shorter clicks with large spectral bandwidths (> 1200 Hz)after much-reduced and weak primary calls. Such a seriesusually consisted of five clicks in a row at slightly increasingdominant frequencies (2900-3500 Hz). These were heardmost frequently at dusk, when males are presumed to establishtheir calling position for that night, and may serve as anagonistic or territorial signal to other males.
ETYMOLOGY
The specific name amplinympha is a composite noun usedin apposition. It is derived from the Latin amplus (large) andnympha ("nymph," a female forest and mountain spirit). Wechoose this name to reflect the relatively large size of femalesof the species and the fact that these scarcely seen yetfrequently heard frogs live in the forested mountains onDominica.
Analysis of phylogenetic relationships
The phylogenetic analysis of the data matrix for EasternCaribbean Eleutherodactylus species from external and internal morphology, including all characters, produced two mostparsimonious trees 310 steps in length with a consistencyindex (CI) of 0.471 (Fig. 7A). Neither tree contradicted thehypothesis of diphyly for Eastern Caribbean taxa. Only twosister-group relationships were apparent, one for E. amplinympha and E. martinicensis, the other for E. terraebolivarisand E. fitzingeri. All other taxa in both trees were placed ina nested fashion, with either E. johnstonei or E. antillensisoriginating at the node giving rise to E. amplinympha andE. martinicensis, and with E. coqui, E. barlagnei, and E. pinchoni originating at subsequent nodes closer to the base of thetree. The southern Eastern Caribbean taxa were similarlynested, with E. shrevei, E. euphronides, and E. urichioriginating at nodes progressively closer to the base of thetree. Of the 142 characters used in the analysis, 17 were constant (12.0%), but only 5 were uninformative (3.5%). Giventhe two most parsimonious topologies, only 26 characters(18.3 %) showed no indication of homoplasy, whereas 41 characters (28.9%) carried homoplasy values> 0.500; 26 of thesewere osteological characters.
The analysis excluding the more questionable characters(Appendix 3) resulted in seven most parsimonious trees(length 243 steps, CI = 0.477; Fig. 7B). The only differencesin topology from the previous analysis were the formation ofa sister-group relationship for E. euphronides and E. shrevei,and the switching of nodes for E. barlagnei and E. pinchoni.Imposing the topology from an electrophoretic analysis(Kaiser et al. I994b) using MacClade 3.01 results in a tree 316steps in length with a CI of 0.46 (Fig. 8). Although this topol-
o9",6
4-Ctl>0C\)Q."i: 3-0I-C\).0E 2:::::lc:-c:
12
FIG. 6. Variation in number of ripe ova with female body size forEleutherodactylus amplinympha n.sp. from Dominica. Open circlesrepresent actual values. The lines are regressions for the total numberof ripe ova in both ovaries (n = 7, r 2 = 0.816; dotted line), theright ovary only (n = 7, r2 = 0.460; solid line), and the left ovaryonly (n = 5, r 2 = 0.310; broken line).
345Snout-vent length (mm)
female with ripe eggs (KU 221264, SVL = 25.0 mm) hadonly four ripe eggs in each ovary, while progressively largerfemales were found to carry greater numbers of ripe eggs; thismay be indicative of a more general correlation betweenfemale size and clutch size (Fig. 6). The average size (greatestlength) of testes was 2.6 mm (range 2.3-2.8 mm; n = 9).
VocalizationsThe primary call of E. amplinympha (Fig. 4B) is a triphasic
compound call consisting of two notes and a click. The firstnote is produced at a constant dominant frequency of 1750 Hz,with a spectral bandwidth ranging from 200 Hz at the beginning of the note to 350 Hz at its end. This note comprises aboutone-third of the total length of the call (X = 135 ms). Thesecond note directly connects to the first after a frequencyjump to 2600 Hz. After a rise time of 160 ms, which is thetotal length of the second note or two-fifths of the total calllength, the frequency reached 3300 Hz, with a maximum spectral bandwidth of 480 Hz. The click follows after a 100-ms gapin the call and is only 30 ms long; its dominant frequency is3100 Hz. The spectral bandwidth of the click decreasesrapidly from 1050 to 350 Hz. The total length of a typical callis 450 ms. Calls of E. martinicensis from Dominica and elsewhere (Fig. 4A) and E. johnstonei (Fig. 4C) are biphasic andof considerable similarity to those of E. amplinympha and toeach other. However, specific differences are sufficient toallow the individuality of each call to be recognized.
Frogs were never heard to produce a series of complete calls,including clicks, in immediate and rapid succession. Maleswere observed to initiate calling bouts with several single"whistle-click" calls spaced several seconds apart. Theythen switched to continuous "whistle-whistle" calling (at arate of greater than one per second) for several seconds beforeending the bout with a "whistle-click" call. In E. amplinympha, "ramping patterns" (sensu Drewry and Rand 1983)were never observed. These are series of chorusing eventsusually initiated by a single individual who is joined by moreand more males, leading to rapid chorusing. Ultimately,

2226 CAN. J. ZOOL. VOL. 72. 1994
Discussion
.~(J)c:Q)
'Q)c:0)'-
zyme data (Kaiser et a!. 1994b), but that the present speciesdiversity may be the result of multiple colonizations.
Northern Eastern Caribbean speciesTaking the conclusions from the morphological data to the
extreme, there may have been as many as four independentcolonization events in the northern Eastern Caribbean: forE. barlagnei, E. johnstonei, E. pinchoni, and the commonancestor of E. amplinympha and E. martinicensis. However,there are certain facts that strongly contradict such a scenarioof multiple colonizations. Although frogs of the genusEleutherodactylus are known for their karyological variability(e.g., Bogart 1970, 1981; DeWeese 1976) and their potentialfor rapid chromosomal change (e.g., Bogart 1991), theindependent derivation of a 2n = 28 chromosome complementfrom 2n = 24 or 26 has in all likelihood not occurred fourtimes in the Eastern Caribbean. The chromosomes of theEastern Caribbean E. johnstonei (2n = 28), for example, arevery dissimilar to those of Greater Antillean species withkaryotypes of 2n = 24, 26, or 28 (Bogart 1981, 1991; Bogartand Hedges 1995) but similar to those of other northernEastern Caribbean species (2n = 28; H. Kaiser and
FIG. 8. Phylogram from a cladistic analysis of West Indian Eleutherodactylus species (316 steps, CI = 0.460). Branch lengths are proportional to the number of character state changes along each branch; notall changes have been mapped on the tree. Solid bars are apomorphiesfor species higher in the tree; open bars indicate reversals. Numbersindicate characters as in Appendix 3. Character states are indicatedafter a colon.
..Q c: eu ,~Q) 0 eu~
(J) ..c:: 1:::: .S~ <.> eu t::>....: 5 .S ..Q eu.2:?
333 '- 45:1 E c:34 c: Q no 051 '- e 81 .....85:3 Q) 2 140:3 (J)10 ::::. ..c:: 3 120 c:
28 138:1
@ §- 4 ..c::142:1
.~21 70 C-
O..c:: Q) 34 7'(J) (J) 37:4 80
L..-~ c: 12 13 705
~140:1 138:3
118:122 140:31 34 :E:
132
6 12414
77:2 c: L..- r--
12 L..-~ '- eu13 ::J13 0- 113 5 0 54140:1 5 4 6<.> 54 9
,8 10 4
45:2 118:0 12 '33- 106 138:1 122
~<.> 2
~248 59 3 2'
77:0 9 142:2 138:0105 118:1 142:3
33:0 1634:1 33:150:1 34:051:0 50:070:0 51:171:0 70:180:0 71:185:1 eO:1106:0 85:2140:0 106:1141:0 140:2142:5 141:1
59
fitzingeri
terraebolivaris
'-----+shrevei
eu hronides
r------coqui
..-------johnstonei
I-----antillensis
amplinympha
martinicensis
A
55
B
'-------urichi
,---------+pinchoni
barlagnei
ogy results in a slightly lower CI, homoplasy is completelyeliminated for three important nonmorphological characters:vocalizations, egg tooth, and chromosome number.
General systematicsAnalysis of morphological data lends support to the hypo
thesis that Eastern Caribbean taxa do not form a monophyleticassemblage. Although the data sets from morphology and allozymes (Kaiser et a!. 1994b) differ in their ability to resolverelationships within a given tree, as indicated by lower CIvalues in the morphological analysis, there is congruence inthe main conclusion: northern Eastern Caribbean Eleutherodactylus species are members of a Greater Antillean assemblage, whereas southern Eastern Caribbean species have SouthAmerican affinities. However, indications from morphological data suggest that northern Eastern Caribbean species do notform a monophyletic group, as was strongly indicated by allo-
fitzingeri
terraebolivaris
shrevei
euphronides
'------urichi
,---------+barlagnei
,-------+pinchoni
,------coqui
..-------johnstonei
I-----anti~ens~
amplinympha
martinicensisFIG. 7. Phylogenetic trees from analyses of morphological charac
ters of West Indian Eleutherodaetylus species. (A) Strict consensustree from an analysis of 142 characters (310 steps, CI = 0.471).(B) Majority-rule consensus tree from an analysis excluding 28problematic characters (Appendix 3; 243 steps, CI = 0.477). Shadedareas highlight the only topological differences between the two trees.The numbers are bootstrap values supporting the branches.

KAISER ET AL. 2227
M. Schmid, unpublished data). This suggests a unique derivation of northern Eastern Caribbean chromosome complements. Furthermore, the occurrence of E. barlagnei andE. pinchoni in macrosympatry but microallopatry onGuadeloupe does not necessarily suggest their sequentialarrival, but habitat partitioning or niche differentiation afterin situ speciation. This suggestion is borne out by the calls ofthese frogs; whereas E. pinchoni has a high-pitched uniphasiccall that pierces the moss mats from under which it calls,E. barlagnei adds a series of loud clicks to a uniphasic call thatenables it to be heard above the din of rushing water. In thesame vein, great similarities in vocalizations as well as identical chromosome numbers support the triad of E. amplinympha, E. johnstonei, and E. maninicensis over the topologythat places E. antillensis as the sister-taxon to E. amplinymphaand E. martinicensis.
The great morphological and biochemical similarity ofE. amplinympha, E. johnstonei, and E. martinicensis is further evidence of a close phylogenetic relationship. Communality of several external morphological characteristics placesE. ampIinympha and E. martinicensis into a sister-group relationship, with E. johnstonei as the sister-taxon to that clade.Biochemical data also suggest a closer relationship betweenE. amplinympha and E. martinicensis than for either withE. johnstonei (Kaiser et al. I994b). It has been suggested thatE. martinicensis may have been imported to Dominica fromMartinique and (or) Guadeloupe by refugees during the turmoil of the French Revolution (Lescure 1983). However, noneof the specimens examined from the montane regions ofDominica are referable to that species. Thus, two biogeographical scenarios seem possible that explain establishment ofE. amplinympha on Dominica, either one conforming to current ideas about speciation (see Giddings et al. 1989; Otte andEndler 1989). In one scenario, possible multiple colonizationevents by an ancestral species, most likely from the GreaterAntilles (Schwartz 1969; Hedges 1989), succeeded in establishing island populations of Eleutherodactylus in the geologically younger Lesser Antilles. These island populationssubsequently speciated, resulting in the observed species radiation, and thus the evolution of several single-island endemics.The second scenario begins with single or multiple introductions of E. martinicensis onto Dominica, either by natural dispersal or through the agency of early Amerindian or morerecent French settlers. The established peripheral isolate(s) onDominica may have been exposed to differential selectionpressures, ultimately creating recognizable divergence at thespecies level. It is possible that additional research on Dominica may reveal pockets of introduced or remnant E. martinicensis in addition to presently known lowland populations ofthat species, and given the ease with which these frogs aretransported (Kaiser 1992), additional introductions are likely.
The exact ranges of E. amplinympha and E. martinicensison Dominica are as yet undetermined, but the two speciesappear to be concentrated in slightly different habitats.Whereas E. amplinympha is most common at higher elevations, E. martinicensis is encountered most frequently in thelowlands. The species are sympatric for a vertical altitudinalsegment of about 50-100 m along the road to FreshwaterLake and near Emerald Pool (H. Kaiser, personal observation). The collections of Dominican specimens made byA. Schwartz are mainly from lowland populations. However,Schwartz remarked that he considered two species to bepresent, one large and one small (A. Schwartz, field notes, 19February 1962, collection of KU 282360-82; 28 February
1962, collection of KU 282241-309; 7 March 1962, collection of KU 282312 - 25), the calls of the smaller being identical with those of E. martinicensis (22 March 1961, collectionof KU 282383 -404). He also commented on the orange coloration of some frogs (23 March 1961, collection of KU282452 -76). Inspection of the KU specimens listed abovewas inconclusive as to the identities of the frogs, and Schwartzdid not detail which individuals of the collected series weredifferently colored in life or which vocalized differently. SmallE. amplinympha « 30 mm) would be difficult to identify,especially considering the effects of specimen shrinkage influid preservatives (Simmons 1991).
The morphological similarities between E. johnstonei andE. martinicensis have long caused taxonomic confusion.Although the two species can be easily distinguished in life,separation of long-preserved museum specimens is difficult orimpossible. Similarly, E. amplinympha is easily distinguishedfrom either of these species in life, yet the smaller specimensexamined are difficult to align with one species or the otherafter only 2 years in preservative. However, since both frozentissues and chromosomal preparations were retained for mostspecimens collected for this study, assignment to species bybiochemical or cytogenetic means is a reliable alternative tomorphological identification.
Southern Eastern Caribbean speciesThe phylogenetic relationships of southern Eastern Carib
bean Eleutherodactylus species are not as easily resolved.Inclusion of E. jitzingeri in the analysis gives some indicationof a closer relationship with South American taxa: all topologies (Figs. 7, 8) support the hypothesis of a close relationshipbetween E. terraebolivaris and E. jitzingeri (fide Rivero 1961;Lynch 1976). The phylogenetic position of E. euphronides andE. shrevei allies these species more closely with the E. terraebolivaris - E. jitzingeri clade than with E. urichi in two topologies (Figs. 7B, 8). The determination of sister-grouprelationships for E. urichi would require the inclusion of additional South American taxa.
Character evolutionThe morphological analysis of Eastern Caribbean Eleuthero
dactylus confirms the well-known high degree of homoplasyfound in this genus. Just as in the only other comprehensivecladistic analysis of the morphology of West Indian Eleutherodactylus (Joglar 1986, 1989), many of the studied characterswere too variable to be informative. In fact, Joglar (1986)excluded 24 of 52 characters that had CI values lower than0.200 and did not even report the CI of his analysis beforeexclusion of these characters; after exclusion the CI was0.417. Regarding these values, the present analysis comparesfavorably (41 characters of 142 with CI values <0.500; CI =0.47]). In addition, Joglar (1986) reduced the number of taxato 10 by excluding "apomorphic species," improving the CIto 0.691. This analysis does not require a reductionistapproach, since homoplasy problems could be alleviated orexplained in part by consulting allozyme data (Kaiser et al.1994b).
Among the characters used in this analysis, three distinctqualities can be identified. Osteological characters have oftenbeen considered the most reliable for phylogenetic analyses, inpart because relationships based on such characters can insome instances be verified by material from the fossil record.For Eleutherodactylus, there is very little such material, andthe scarce fossils known from the West Indies are fragmentary

2228 CAN. 1. ZOOL. VOL. 72, 1994
(e,g" Auffenberg 1958; Lynch 1966; Steadman et aL 1984;Pregill et al. 1988); the only complete fossil Eleutherodactylusis embedded in amber (Poinar and Cannatella 1987) and is oflimited comparative value, owing to its uniqueness. Thus, noevolutionary trends have been identified that could assist withdetermining the direction of osteological modifications overevolutionary time, However, there are some structures forwhich little variation has been described for anurans or inwhich variation is constrained (Duellman and Trueb 1986;Ford 1989); these may be more useful in elucidating relationships than those for which there is a high degree of variability.The phylogram (Fig, 8) is largely based on such characters,At the other extreme are those characters that display such ahigh degree of variability between species (or sometimes evenwithin species) that they do not allow any unequivocal phylogenetic inference to be drawn. These are, by and large,characters of external morphology (e,g., tuberculation, coloration, gross shape), Lastly, there are a few characters, such aschromosome number, vocalization, and developmental characteristics, whose relative stability in phylogenetic usage hasbeen valuable in a variety of other anuran families, althoughtheir degree of usefulness may vary depending on the level ofclassification and on the ecological specifics of the taxastudied,
Although there may be some degree of homoplasy even inthe more conservative characters, we consider the following tobe good indicators of an Antillean subgroup, likely synonymous with the proposed martinicensis group (Hedges 1989):(1) posterior extent of maxillary teeth to beyond maxillaquadratojugal articulation; (2) medial ramus of pterygoid narrow medially; (3) (dfsq*at) condition of the m, depressor mandibulae; (4) palatine and vomer overlap; (5) lateral sides ofhyoid plate concave; (6) alary process of hyoid plate present;(7) dorsal crest of ilium reaches sacral region; (8) two tarsalsesamoid elements present; (9) tympana round; (10) supratympanic fold weak; (II) finger II > I; (12) toe disks aboutequal in size, These observations are consistent with data forthe same characters from 14 additional Antillean species(H. Kaiser, unpublished data),
ConclusionsThis study provides further evidence that Eastern Caribbean
Eleutherodactylus species do not form a monopohyletic group,However, resolution of sister-taxon relationships is less wellsubstantiated despite the large number of characters or becauseof their relatively indiscriminate use in a computer algorithm.Unless our knowledge of the evolutionary history of morphological characteristics among Eleutherodactylus species can beimproved, by developmental studies for instance, the indiscriminate use of morphological data in isolation may preventthe formulation of meaningful hypotheses of relationships dueto homoplasy, The alternative is to use an analysis of a seconddata type, such as biochemical data, as a working hypothesis,and to conduct a careful a posterori inspection and interpretation of all characters, This is a luxury not yet available to thosestudying vertebrate fossils,
Problems with diagnostic features are all too commonamong the polytypic Eleutherodactylus species, Species descriptions of these frogs have traditionally been based entirelyon external morphological characters, often to the exclusion ofvocalizations, Considering the difficulties encountered in thisstudy with elucidating phylogenetic relationships on the basisof morphology alone, a minimal, single-data-set approach may
make accurate taxonomic decisions too tenuous to be of practical value, It is symptomatic in that respect to have several newEleutherodactylus species described each year, while othersare synonymized (e,g" Lynch and La Marca 1993), As combinations of morphological and biochemical data are beginningto influence the classification of Eleutherodactylus speciesmore and more (e,g" Miyamoto 1983, 1984, 1986; Hedges1989), taxonomic decisions will become better documentedand probably more durable, In the case of the 515 currentlyrecognized species of Eleutherodactylus, ongoing revisions ofsubgeneric and species-group classification (e,g" Lynch1986, 1989, 1993; Hedges 1989) are certain to benefit fromthe increased usage of a full, multidisciplinary systematicstoolbox,
Acknowledgments
We are grateful to C,A, George, H,M, Gray, G, Schafer,H,H, Schwarten, T,F, Sharbel, D. von Stosser, R. Wagenseil,and A. Werres for field assistance. The patience and guidanceof H,M. Reiswig, whose equipment was occupied for hourswith frog carcasses, is gratefully acknowledged, Thanks areextended to R,E, Lemon for permitting use of his sound analysis setup, and to D,J, Mountjoy for providing hints on how tomake it work for frog voices, D,S, Townsend dissected theeggs of E. amplinympha, Special thanks are given to J ,PoBogart and S,B. Hedges for sharing some insights from theirunpublished work, This study benefited greatly from discussions with A. Baez, M,W, Caldwell, R,L Carroll, eM,Dwyer, LS, Ford, LA, Lowcock, T,F, Sharbel, J,E.Simmons, and L Trueb, We thank F,R. Cook, W,E,Duellman, LS, Ford, W.R, Heyer, R,M, Rankin, J,P,Rosado, and J,E. Simmons for authorizing and preparingspecimen loans, The manuscript was improved during itsdevelopment by comments from W,E. Duellman and eM,Dwyer. Its final version was critically read by M,W, Caldwell. The fieldwork on Dominica was greatly facilitated by thelate P, Alexander, R, Wagenseil, and the staff at the ArchboldTropical Research Center. We are very grateful to A, Jamesof the Forestry and Wildlife Division, Government of Dominica, for issuing collecting and export permits and for providingassistance in clarifying local place names, Most of the workon preserved specimens was conducted by H,K. at TheNatural History Museum, The University of Kansas, and hissincere thanks go to staff and students for their support. Weexpress our gratitude to A. Rupprecht for assisting duringchromosome preparation and to G, Hesse for photographicassistance. This study was conducted as part of H,K.'s doctoral dissertation and submitted to the Faculty of GraduateStudies and Research, McGill University, MontreaL It wasfunded in part by grants and scholarships to H,K, from theHans-Kruger-Stiftung, the Studienstiftung des DeutschenVolkes, and Boehringer Ingelheim Fonds, Germany, by operating grants to D,M.G, from the Natural Sciences and Engineering Research Council of Canada, and by a grant to M,S. fromthe Deutsche Forschungsgemeinschaft (Schm 484/11-1),
Auffenberg, W. 1958. A small fossil herpetofauna from Barbuda,Leeward Islands, with the description of a new species of Hvla.Q. J. Fla. Acad. Sci. 21: 248-254.
Barbour, T. 1914. A contribution to the zoogeography of the WestIndies, with especial reference to amphibians and reptiles, Mem.Mus. Compo Zoo!. 44: 209-359.

KAISER ET AL. 2229
Barnhart, C.L., and Barnhart, R.K. 1983. The world book dictionary. World Book, Inc., Chicago.
Boenger, O. 1894. Eleutherodactylus urichi sp.nov. In A preliminarylist of reptiles and batrachians of the island of Trinidad. Edited byR.R. Mole and F.W. Urich. J. Trinidad Field Nat. Club, 2:87-89.
Bogart, J. P. 1970. Systematic problems in the amphibian familyLeptodactylidae (Anura) as indicated by karyotypic analysis.Cytogenetics, 9: 369-383.
Bogart, J.P. 1981. Chromosome studies in Smiruhillus from Cuba andEleutherodaetylus from Cuba and Puerto Rico (Anura: Leptodactylidae). R. Ont: Mus. Life Sci. Contrib. No. 129.
Bogart, J.P. 1991. The influence of life history on karyotypic evolution in frogs. In Amphibian cytogenetics and evolution. Edited byD.M. Green and S.K. Sessions. Academic Press, New York. pp.233-258.
Bogart, J.P., and Hedges, S.B. 1995. Chromosome evolution inJamaican frogs of the genus Eleutherodacrylus. J. ZooI. (Lond.).235: 9-31.
Canadian Council on Animal Care. 1980-1984. Guide to the use andcare of experimental animals. Canadian Council on Animal Care,Ottawa.
Clayton, J.W., and Tretiak, D.N. 1972. Amine citrate buffers for pHcontrol in starch gel electrophoresis. J. Fish. Res. Board Can. 29:1169-1172.
Davis, S.D., Droop, S.J.M., Gregerson, P., Henson, L., Leon, C.J.,Lamlein Villa-Lobos, J., Synge, H., and Zantovska, J. 1986.Plants in danger. What do we know? International Union for Conservation of Nature and Natural Resources, Cambridge, U. K.
DeWeese, J .E. 1976. The karyotype of Middle American frogs of thegenus Eleutherodactylus (Anura: Leptodactylidae): a case study ofthe significance of the karyologic method. Ph. D. thesis, Universityof Southern California, Los Angeles.
Dingerkus, G., and Uhler, L.D. 1976. Enzyme clearing of alcianblue stained whole small vertebrates for demonstration of cartilage. Stain Technology. 52: 229-232.
Drewry, G.E., and Rand, A.S. 1983. Characteristics of an acousticcommunity: Puerto Rican frogs of the genus Eleutherodactylus.Copeia, 1983: 941-953.
Duellman, W.E. 1970. Hylid frogs of Middle America. Univ. Kans.Mus. Nat. Hist. Monogr. NO.1.
Duellman, W.E. 1993. Amphibian species of the world: additionsand corrections. Univ. Kansas Mus. Nat. Hist. Spec. Publ. No. 21.
Duellman, W.E., and Trueb, L. 1986. Biology of the Amphibia.McGraw-Hill, New York.
Dumeril, A.M.e., and Bibron, G. 1841. Erpetologie generale.Roret, Paris.
Ford, L.S. 1989. The phylogenetic position of poison-dart frogs(Dendrobatidae): reassessment of the neobatrachian phylogenywith commentary on complex character systems. Ph.D. thesis,University of Kansas, Lawrence.
Frost, D.R. (Editor). 1985. Amphibian species of the world. A taxonomic and geographical reference. Allen Press, Inc., and TheAssociation of Systematics Collections, Lawrence, Kans.
Giddings, L. V., Kaneshiro, K. Y., and Anderson, W. W. (Editors).1989. Genetics, speciation, and the founder principle. OxfordUniversity Press, New York.
Hanken, J., and Wassersug, R.J. 1981. The visible skeleton: a newdouble-stain technique reveals the nature of the "hard" tissues.Funct. Photogr. 16: 22-26,44.
Hardaway, T. E., and Willliams, K.L. 1975. A procedure for doublestaining cartilage and bone. Br. J. Herpetol. 5: 473-474.
Hardy, J.D., Jr. 1982. Biogeography of Tobago, West Indies, withspecial reference to amphibians and reptiles: a review. Bull. Md.Herpetol. Soc. 18: 37 - 142.
Hardy, J.D., Jr. 1984. Frogs, egg teeth, and evolution: preliminarycomments on egg teeth in the genus Eleutherodactylus. Bull. Md.Herpetol. Soc. 20: I 11.
Hardy, J.D., Jr. 1985. Frog mountain: preliminary comments on the
genus Eleutherodactylus on the island of Guadeloupe, West Indies.Bull. Md. Herpetol. Soc. 21: 27-33.
Hardy, J.D., Jr., and Harris, H.S. 1979. Occurrence of the WestIndian frog, Eleutherodactylus johnstonei, in South America andon the island of Curacao. Bull. Md. Herpetol. Soc. 15: 124-133.
Hedges, S.B. 1989. Evolution and biogeography of West Indian frogsof the genus Eleutherodactylus: slow-evolving loci and the majorgroups. In Biogeography of the West Indies: past, present, andfuture. Edited by C.A. Woods. Sandhill Crane Press, Gainesville,Fla. pp. 305 - 370.
Joglar, R.L. 1986. Phylogenetic relationships of the West Indianfrogs of the genus Eleutherodactylus. Ph.D. thesis, University ofKansas, Lawrence.
Joglar, R.L. 1989. Phylogenetic relationships of the West Indianfrogs of the genus Eleutherodactylus: a morphological analysis. InBiogeography of the West Indies: past, present, and future. Editedby C.A. Woods. Sandhill Crane Press, Gainesville, Fla. pp.371-408.
Johnson, T.H. 1988. Biodiversity and conservation in the Caribbean:profiles of selected islands. Monogr. No.1, International Councilfor Bird Preservation, Cambridge, U.K.
Kaiser, H. 1992. The trade-mediated introduction of Eleutherodactylusmartinicensis (Anura: Leptodactylidae) on St. Barthelemy, FrenchAntilles, and its implications for Lesser Antillean biogeography.J. Herpetol. 26: 264-273.
Kaiser, H. 1993. Systematics and biogeography of Eastern Caribbeanfrogs. Ph.D. thesis, McGill University, Montreal.
Kaiser, H., Hardy, J.D., Jr., and Green, D.M. 1994a. The taxonomic status of Caribbean and South American frogs currentlyascribed to Eleutherodactylus urichi (Anura: Leptodactylidae).Copeia, 1994: 780-796.
Kaiser, H., Sharbel, T.F., and Green, D.M. 1994b. Systematics andbiogeography of Eastern Caribbean Eleutherodaetylus (Anura:Leptodactylidae): evidence from allozymes. Amphib.-Reptilia, 15:375-394.
Kuramoto, M. 1990. A list of chromosome numbers of anuranamphibians. Bull. Fukuoka Univ. Educ. Part 3 Nat. Sci. 39:83-127.
Lescure, J. 1983. Introductions passives et actives des reptiles etd'amphibiens dans les Antilles et les Guyanes. e. R. Soc. Biogeogr. 59: 59-70.
Lescure, J. 1987. Le peuplement en reptiles et amphibiens des PetitesAntilles. Bull. Soc. Zool. Fr. 112: 327 -342.
Leviton, A.E., Gibbs, R.H., Jr., Heal, E., and Dawson, C.E. 1985.Standards in herpetology and ichthyology: Part I. Standard symbolic codes for institutional resource collections in herpetology andichthyology. Copeia, 1985: 802-832.
Lynch, J.D. 1965. A new species of Eleutherodactylus fromGuadeloupe, West Indies. Breviora No. 220. pp. 1-7.
Lynch, J.D. 1966. The status of the tree frog, Hyla barbudensisAuffenberg , from Barbuda, British West Indies. Copeia, 1966:524-530.
Lynch, J.D. 1975. A review of the broad-headed eleutherodactylinefrogs of South America (Leptodactylidae). Occas. Pap. Mus. Nat.Hist. Univ. Kans. No. 38.
Lynch, J.D. 1976. The species groups of the South American frogsof the genus Eleutherodactylus (Leptodactylidae). Occas. Pap.Mus. Nat. Hist. Univ. Kans. No. 61.
Lynch, J.D. 1979. The frogs of the genus Eleutherodactylus in theAndes of southern Ecuador. Misc. Publ. Univ. Kans. Mus. Nat.Hist. No. 66.
Lynch, J.D. 1980. Two new species of earless frogs allied toEleutherodacrylus surdus (Leptodactylidae) from the Pacific slopesof the Ecuadorian Andes. Proc. BioI. Soc. Wash. 93: 327-338.
Lynch, J.D. 1986. The definition of the Middle American clade ofEleutherodactylus based on jaw musculature (Amphibia: Leptodactylidae). Herpetologica, 42: 248 - 258.
Lynch, J.D. 1989. Intrageneric relationships of mainland Eleutherodactylus (Leptodactylidae). I. A review of the frogs assigned to the

2230 CAN. J. ZOOL. VOL. 72, 1994
Eleutherodactylus discoidalis species group. Milw. Publ. Mus.Contrib. BioI. Geo!. No. 79.
Lynch, J.D. 1993. The value of the m. depressor mandibulae inphylogenetic hypotheses for Eleutherodacrylus and its allies(Amphibia: Leptodactylidae). Herpetologica, 49: 32-41.
Lynch, J.D., and La Marca, E. 1993. Synonymy and variation inEleutherodactylus bicumulus (Peters) from northern Venezuela,with a description of a new species (Amphibia: Leptodactylidae).Caribb. J. Sci. 29: 133-146.
Lynch, J.D., and Myers, C.W. 1983. Frogs of thejirzingeri groupof Eleutherodaetylus in Eastern Panama and Chocoan SouthAmerica (Leptodactylidae). Bull. Am. Mus. Nat. Hist. 175:481-572.
Maddison, W.P., and Maddison, D.R. 1992. MacClade: analysis ofphylogeny and character evolution. Version 3.01. Sinauer Associates, Sunderland, Mass.
Miyamoto, M.M. 1983. Frogs of the Eleutherodactylus rugulosusgroup: a cladistic study of allozyme, morphological and karyological data. Syst. Zoo!. 32: 109 -124.
Miyamoto, M.M. 1984. Central American frogs allied to Eleurherodacrylus cruen/us: allozyme and morphological data. J. Herpeto!.18: 256-263.
Miyamoto, M.M. 1986. Phylogenetic relationships and systematicsof the Eleurherodaetylusjitzingeri group (Anura, Leptodactylidae).Copeia, 1986: 503 - 511.
Murphy, R.W., Sites, J.W., Buth, D.G., and Haufler, C.H. 1990.Proteins 1: isozyme electrophoresis. In Molecular systematics.Edired by D.M. Hillis, and C. Moritz. Sinauer Associates, Sunderland, Mass. pp. 45-126.
Narins, P.M., and Capranica, R.R. 1976. Sexual differences in theauditory system of the treefrog Eleutherodactylus coqui. Science(Washington, D.C.), 192: 378-380.
Narins, P.M., and Capranica, R.R. 1978. Communicative significance of the two-note call of the treefrog Eleutherodactyluseoqui. J. Compo Physiol. A, 127: 1-9.
Nicolson, D.H. 1991. Flora of Dominica, part 2: Dicotyledoneae.Smithson. Contrib. Bot. No. 77.
Nomenclature Committee of the International Union of Biochemistry.1984. Enzyme nomenclature 1984. Academic Press, New York.
Nussbaum, R.A. 1982. Heterotropic bones in the hindlimbs of frogsof the families Pipidae, Ranidae and Sooglossidae. Herpetologica,38: 312 - 320.
Otte, D., and Endler, J.A. (Editors). 1989. Speciation and its consequences. Sinauer Associates Inc., Sunderland, Mass.
Peters, J.A. 1964. Dictionary of herpetology. Hafner Publishing Co.,New York.
Poinar, G.O., and Cannatella, D.C. 1987. An Upper Eocene frogfrom the Dominican Republic and its implication for Caribbeanbiogeography. Science (Washington, D.C.), 237: 1215-1216.
Pregill, G.K., Steadman, D.W., Olson, S.L., and Grady, F.V. 1988.Late Holocene fossil vertebrates from Burma Quarry, Antigua,Lesser Antilles. Smithson. Contrib. Zoo!. No. 463.
Rivero, J.A. 1961. Salientia of Venezuela. Bull. Mus. Compo Zool.126: 1-207.
Savage, J. M. 1975. Systematic and distribution of the Mexican andCentral American stream frogs related to Eleurherodactylus rugulosus. Copeia, 1975: 254-306.
Savage, J.M. 1987. Systematics and distribution of the Mexican andCentral American rainfrogs of the Eleutherodaetylus gollmerigroup (Amphibia: Leptodactylidae). Fieldiana Zoo!. (n.s.) No. 33.
Schmid, M. 1978. Chromosome banding in Amphibia. I. Constitutive heterochromatin and nucleolus organizer regions in 8ufo andHyla. Chromosoma, 66: 361-388.
Schwartz, A. 1967. Frogs of the genus Eleutherodactylus in theLesser Antilles. Stud. Fauna Curacao Other Caribb. lsI. 24: 1-62.
Schwartz, A. 1969. The Antillean Eleurherodactylus of the aurieulatus group. Stud. Fauna Curacao Other Caribb. lsI. 30: 99 -115.
Simmons, J .E. 1991. Conservation problems of fluid-preserved collections. In Natural history museums: directions for growth.Edited by P.S. Cato and C. Jones. Texas Tech University Press,Lubbock. pp. 69-89.
Steadman, D.W., Pregill, G.K., and Olson, S.L. 1984. Fossil vertebrates from Antigua, Lesser Antilles: evidence for Late Holocene human-caused extinctions in the West Indies. Proc. Nat!'Acad. Sci. U.S.A. 81: 4448-4451.
Swofford, D.L. 1993. PAUP: phylogenetic analysis using parsimony,version 3.1.1. Computer program distributed by the IllinoisNatural History Survey, Champaign.
Trueb, L. 1993. Patterns of cranial diversity. In The skull. Vo!. 2.Patterns of structural and systematic diversity. Edited by J. Hankenand B.K. Hall. University of Chicago Press, Chicago. pp. 255343.
Trueb, L., and Cloutier, R. 1991. A phylogenetic investigation of theinter- and intrarelationships of the Lissamphibia (Amphibia:Temnospondyli). In Origins of the higher groups of tetrapods: controversy and consensus. Edired by H.-P. Schultze and L. Trueb.Cornell University Press, Ithaca, N. Y. pp. 223 - 313.
Wassersug, R.J. 1976. A procedure for differential staining of cartilage and bone in whole formalin-fixed vertebrates. Strain Technol. 51: 131-134.
Wells, K.D. 1981. Territorial behavior of the frog Eleutherodaetylusuriehi in Trinidad. Copeia, 1981: 726-728.
Appendix 1
Specimens examinedThe following is a list of Eleutherodactylus specimens that were examined to determine osteological and external morpho
logical character states. Specimens were cleared and double-stained unless marked with an asterisk: these are alcoholic specimens from institutional herpetological collections or from H.K.'s collection (specimens with David M. Green (DMG) fieldtags). Additional specimens were usually available in the KU collection; comparative material was in most cases pickedrandomly from a series and numbers were not recorded. Where original distances and elevations were given in nonmetricunits by the collectors, these have been converted to SI units.
Eleutherodactvlus alricola. JAMAICA: Portland Parish, Blue Mountain Peak, AMNH 55648, 55649-50*.Eleutherodaciylus amplinympha. DOMINICA: Emerald Pool area, elev. ca. 400 m, DMG 3619-22*, 4598-99*; 500 m
SE Layou Park Estate, elev. ca. 325 m, DMG 3726, 3831 -32*, 4141-42*; Freshwater Lake area, elev. ca. 800 m, DMG3590-92*,4591*,4596-97*; Slope of Morne Diablotin along access track, elev. ca. 1000 m, DMG 4037*,4189*.
Eleurherodactylus amillensis. VIRGIN ISLANDS: St. Thomas, AMNH A 52646. Tortola, AMNH A 77502* St. John,Catherineberg, ca. 200 m, AMNH A 109414*; 800 m N, 320 m E Lameshur KU 45589. PUERTO RICO: Mayamon, AMNHA 10228*; Aibonito, AMNH A IOII8*'
Eleurherodaetylus auriculatus. CUBA: Isla de Pinos, just W Nueva Gerona, E base Sierra de las Casas, AMNH A 63278*;Isla de Pinos, 17.6 km NE Siguanea, AMNH A 63279*; Oriente, Gran Piedra, La Esperancita, 3 km SE, 16 km NE Sevilla,elev. 1065 m, AMNH A 64343-45*, KU 203372, 203373-75*.
Eleurherodactylus barlagnei. GUADELOUPE: Basse-Terre, Matouba, elev. 700 m, MCZ 35334 (holotype)*; Chutes duCarbet, along path to lower falls, elev. ca. 700 m, DMG 3738*, 3896*; Sofa'ia, Riviere Salee, end of road D19, elev. ca.300 m, DMG 3650,3745,3818; La Soufriere, 400 m W La Citerne, along road DII, elev. ca. 1200 m, DMG 4038*,4146-47*,4155*,4675; Matouba Hot Springs, elev. 1281 m, DMG 4195*.

KAISER ET AL
Eleutherodactylus cochranae. VIRGIN ISLANDS: Sl. Thomas, AMNH A 77499*, 77500, 77501 *; Sl. John, Catherine-berg, elev. ca. 200 m, A 109417*; Sl. John, Bordeaux Mountain Rd., AMNH A 109418*.
Eleutherodactylus coqui. PUERTO RICO: 4.8-8 km S EI Verde, AMNH 71998*, 71999, 72000, 72010-11 *.Eleutherodactylus eileenae. CUBA: 2.9 km S Topes de Collantes, KU 203389, 203392.Eleutherodactylus euphronides. GRENADA: Parish of Sl. Andrew, Grand Etang, AMNH 74536-44, KU 93337-38,
265429-40, MCZ 2976,43229 (holotype), UIMNH 61641-43; cable and wireless station near Mount Sl. Catherine, ca.4 km NW Paraclete, all. ca. 650 m, NMC 35009-4-7*,35010-3-10*, DMG 4149.
Eleuthrodactylusfitzingeri. PANAMA: Panama Province, Tapia, Rio Tapia, AMNH A 40680,40681-82*, nr. Altos dePaeora, E Cerro Jefe, 700-800 m, KU 107149-50. COSTA RICA: Limon, nr. Tortuguera Village, AMNH A 81466*.San Jose, La Sisica, 15 km SW Isidro del General, elev. 865 m, AMNH A 86489*.
Eleutherodactylusjohnstonei. BARBUDA: Sunset View Hotel, sea level, DMG 3633,3667-69*. GRENADA: Parish ofSt. George, Sl. Ann's Guest House, elev. ca. 60 m, NMC 35011-3-11*, 35011-18-21*. MONTSERRAT: Parish ofSl. Anthony, end of Galways Soufriere road, DMG 3380-88*. ST. KITTS: Sl. Thomas Middle Island Parish, RomneyManor, 0.8 km N Old Road Town, DMG 3094-3105*. ST. LUCIA: Sans Soucis, Castries, DMG 2850-68. ST. VINCENT: Parish of Sl. George, Kingstown, Kingstown Park Guest House, DMG 2968-81.
Eleutherodactylus karlschmidti. PUERTO RICO: EI Yunque, La mina, elev. 473 m, KU 79212.Eleutherodactylus klinikowskii. CUBA: Pinar del Rio, Cueva de Santo Tomas, 10 km N Cabezas, KU 203403-04.Eleutherodactylus leprus. GUATEMALA: El Petens, ca. 15 km NW Chinaja, elev. ca. 120 m, KU 55963.Eleutherodactylus longipes. MEXICO: Tamaulipas, Cueva de Infiernillo, KU 182345.Eleutherodactylus martinicensis. DOMINICA: Emerald Pool area, elev. ca. 400 m, DMG 4066*, 4683; 500 m SE Layou
Park Estate, elev. ca. 325 m, DMG 3744*; Freshwater Lake area, elev. ca. 800 m, DMG 4685*; Trafalgar Falls area, elev.ca. 330 m, DMG 3725*. GUADELOUPE: Basse-Terre, Chutes du Carbet, path to lower falls, elev. ca. 700 m, DMG3651-52*, 3876-77*, 3902-03*. Grande-Terre, 1.7 km S intersection of roads DI09 and N5, elev. ca. 75 m, DMG3512-13*,3553*,3660*. LA DESIRADE: 450 m N Beausejour post office, elev. ca. 100 m, DMG 3527-30*,3626-27*.MARIE-GALANTE: Les Balisiers gully, 1.5 km S Ste. Croix, elev. 76 m, DMG 3603-05*; Le Trou it Diable, elev. ca.100 m, DMG 3524-25*. MARTINIQUE: Morne Rouge, 600 m SE Montagne Pelee restaurant, along road D39, DMG3634,3826*; Deux Choux, 100 m N intersection of roads N3 and DI, DMG 3823 -24*; Deux-Terres, intersection of roadsDI5 and N4, DMG 3648-49*,3827*; 100 m below top of Morne Bigot road, DMG 3645-47*,3661-62*,3828-30*.
Eleutherodactylus pinchoni. GUADELOUPE: Basse-Terre, Chutes du Carbet, path to lower falls, elev. ca. 700 m, DMG3892-95*,3904-07*; La Soufriere, 400 m before La Citerne along road DII, elev. ca. 1200 m, DMG 4143-44*,4151;3 km W Grand Cafe, 600 ft (I ft = 0.3048 m), AMNH 74545-47*, MCZ 43231 (holotype)*, UIMNH 61647-50*.
Eleutherodactylus planirostris. BAHAMAS: Great Abaco island, Marsh Harbour, AMNH A 57619, A 57622-23*.CUBA: Las Villas, Soledad, AMNH A 61509-10*.
Eleutherodactylus richmondi. PUERTO RICO: EI Yunque peak, AMNH A 10230-31*, 10233.Eleutherodactylus shrevei. ST. VINCENT: Parish of Sl. Andrew, Lowrt [sic], elev. ca. 330 m, KU 265445-54, MCZ
43230 (holotype); Charlotte Parish, ca. 5.5 km W Orange Hill on La Soufriere summit track, elev. ca. 750 m, DMG 4695*,4698-99*, 4707, NMC 35062-4-7*, 35062-10-12*, 35062-15*; edge of Soufriere crater, elev. ca. 950 m, MCZ19814-17*, UIMNH 61644-46*.
Eleutherodaetylus terraebolivaris. COLOMBIA: USNM 144737-38*. TOBAGO: 3 mi N Mount St. George, KU265455*; Main Ridge, ca. 7 km N Roxborough, DMG 4546*, NMC 35056, 35060-1-5*, 35064-1-3*, 35064-8*,35064-10*. VENEZUELA: Rancho Grande, MCZ 31062 (holotype)*, USNM 128212-14*,167609-13*; Los Canales,USNM 1228807 -08*.
Eleutherodactylus unistrigatus. COLOMBIA: Narifio, nr. end of Laguna de la Cocha, elev. 2850 m, AMNH A 86774;7 km NE Guachual, elev. 3000 m, AMNH A 86779*. ECUADOR: Quito, Lago Cotoral, AMNH A 20442*, 20444-45*.Putamayo, Colon, elev. 2220 m, KU 168624.
Eleutherodactylus urichi. TOBAGO: Main Ridge, ca. 7 km N Roxborough, DMG 4602*, NMC 35059*,35063-1-2*;6.4 NE Pembroke, KU 265456*. TRINIDAD: N Araima Valley, DMG 4019*,4026, NMC 35061-1-8*, 35065-1-8*,35065-3 -5*; Arima Ward, Aripo Road, 3.2 km N intersection with Eastern Main Road, KU 265458*; Sl. Ann's Ward, SantaCruz Valley, 12 km N San Juan, KU 265457*.
Eleutherodactylus varleyi. CUBA: Oriente, Gran Piedra, La Esperancita, 3 km SE and 16 km NE Sevilla, elev. 1065 m,KU 203435; 3 km E Gran Piedra, KU 203438.
Appendix 2
Localities sampled for allozymesEleutherodactylus amplinympha. DOMINICA: Emerald Pool area, elev. ca. 400 m (n = 10); 500 m SE Layou Park Estate,
elev. ca. 325 m (n = 8); Freshwater Lake area, elev. ca. 800 m (n = 15); Trafalgar Falls area, elev. ca. 300 m (n = 4);Slope of Morne Diablotin along access track, elev. ca. 1000 m (n = I).
Eleutherodactylus johnstonei. ANTIGUA: Parish of Sl. Mary, end of road in Christian Valley, elev. 35 m (n = 2); Parishof Sl. Philip, Gaynor's Mill, sea level (n = 3). BARBADOS: Parish of Sl. James, Garden of Bellairs Research Institute,sea level (n = 4); Parish of Sl. Andrew, Turner's Hall Woods, at end of Sl. Simon road, elev. ca. 50 m (n = 3); Parishof Sl. John, Road to Consett Bay, 200 m from beach, sea level (n = I); Parish of Sl. Michael, Bridgetown, parking lot ofGrand Barbados Beach Hotel, sea level (n = 3). BARBUDA: Sunset View Hotel, sea level (n = 4). GRENADA: Parishof Sl. Patrick, 2.4 km SW Sauters, elev. ca. 150 m (n = 5); Parish of Sl. David, Les Avocats waterworks, elev. ca. 400 m(n = I); Parish of St. Andrew, Grand Etang Lake parking lot, elev. ca. 500 m (n = 5). GUYANA: Georgetown, courtyardof Park Hotel, sea level (n = 2). MONTSERRAT: Parish of Sl. Anthony, end of Galways Soufriere road (n = 19); Parishof St. Peter, Soldier's Ghaut, Fogarty's (n = I). NEVIS: Sl. George Gingerland Parish, Golden Rock Estate (n = 2);Sl. James Windward Parish, Nesbitt Plantation (n = 2). SABA: I km N The Gap (n = 3); I km N Windwardside beyondEnglish Quarter (n = 3); Windwardside, beginning of Mount Scenery steps (n = 2). ST. EUSTATIUS: The Quill (n = 15).ST. KITTS: Sl. Thomas Middle Island Parish. Romney Manor, 0.8 km N Old Road Town, (n = 2); Sl. Peter Basseterre
2231

2232 CAN. J. ZOOL. YOLo 72, 1994
Parish, Bayford's TV mast, I km N Ogee's (n = 2); St. John Capisterre Parish, St. George's Ghut, 0.5 km S Tabernacle(n = 2). ST. LUCIA: Sans Soucis, Castries (n = I); 3 km N Gros Islet (n = I). ST. MARTIN: Pic Paradis summit(n = 6); Terres Basses (n = 6). ST. VINCENT: Parish of St. George, Kingstown, Kingstown Park Guest House (n = 2);Parish of St. Andrew, Lowrey, 1.5 km NE Vermont (n = 2). VENEZUELA: Caracas, Sebucan, Altamira (n = 2).
Eleutherodactylus martinicensis. DOMINICA: Emerald Pool area, elev. ca. 400 m (n = 2); 500 m SE Layou Park Estate,elev. ca. 325 m (n = I); Freshwater Lake area, elev. ca. 800 m (n = I); Trafalgar Falls area, elev. ca. 330 m (n = I).GUADELOUPE: Basse-Terre, Chutes du Carbet, path to lower falls, elev. ca. 700 m (n = 4); Riviere Moreau, ca. 7 kmSW Douville, elev. ca. 300 m (/I = 2); Riviere des Vieux Habitats, Maison du Cafe, 400 m before end of road D27, elev.ca. ISO m (n = 2); Riviere Petit David, 400 m SE Les Mamelles, along road D23, elev. ca. 700 m (n = I); Sofai'a, RiviereSalee, end of road D19, elev. ca. 300 m (n = 2). Grande-Terre, 1.7 km S intersection of roads D109 and N5, elev. ca.75 m (/I = 2). LA DESIRADE: 450 m N Beausejour post office, elev. ca. 100 m (n = 5). MARIE-GALANTE: Les Balisiersgully, 1.5 km S Ste. Croix, elev. 76 m (n = 2); Le Trou aDiable, elev. ca. 100 m (n = 2). MARTINIQUE: Morne Rouge,600 m SE Montagne Pelee restaurant, along road D39 (n = I); Deux Choux, 100 m N intersection of roads N3 and DI(n = 3); Deux-Terres, intersection of roads DIS and N4 (11 = I); 100 m below top of Morne Bigot road (n = 6); Fort-deFrance, Vieux Fort Park (n = 4). ST. BARTHELEMY: St. Jean, Jean Bart Hotel (11 = 9); Lorient, Hotel La Normandie(11 = 5). TERRE-DE-HAUT: Terre-de-Haut village (n = 2).
Appendix 3List of characters alld character states
The following list contains the characters used in the phylogenetic study. Characters in each subsection are listed inanterior-posterior order and explanations are given only where clarification is required. Many of the characters used hereare identical with those used by Joglar (1986) and (or) Ford (1989) to facilitate comparisons, and detailed explanations ofthese characters (e.g., historical use, homology, variability) can be found there. Modification of characters used by theseauthors was required to deal with the specific taxa under investigation. Characters (C) used strictly as by Ford (1989) areidentified by a CF designation (e.g., CFI is Ford's character I), whereas those of Joglar (1986) have a Cl designation.Characters that were modified from the original meaning or usage by these authors include the label "m" (e.g., mCF3).Paired structures are treated in the singular unless both elements are compared or used in establishing the character. Characteruncertainties, e.g., where variability could not be ascertained or where preservation may have altered a character, are identified by the superscripted letters V and P, respectively.
Internal morphologyl. Head
A. Dermal roofing bonesI. Nasals, relative size (mCI 22): In some Eleutherodactylus species, the nasals are small, whereas in others
they cover most of the anterior region of the skull roof. Two discrete states are visible without quantifyingthe character further than relative cover. 0 = nasals covering most or all of preorbital area; I = nasals covering less than half of preorbital area.
2v Nasals, medial contact (mCFI, mCJ23): 0 = extensive contact, more than half length of nasals; I = littlecontact, less than half the length of nasals; 2 = no contact.
3. Nasals, degree of contact with frontoparietals (mCJ24): In some taxa the nasals nearly overlap the frontoparietals when examined in dorsal view, whereas in others there is a wide separation between these bones.o = nasals overlapping frontoparietals or abutting them; I = bones widely separated.
4. Frontoparietals, shape (mCF3): 0 = rectangular; I = anterior portion of frontoparietals wider than posteriorportion; 2 = posterior portion of frontoparietals wider than anterior portion.
5. Frontoparietal, anterolateral ala: In some taxa there is a lateral extension to the anterior portion of each frontoparietal. This extension is not considered in determining the state of C4. 0 = absent; I = present.
6v. Frontoparietal, fusion with elements of occiput (CI 25): 0 = not fused; I = fused, suture clearly evident;2 = fused, no suture visible.
B. Neurocranium7. Sphenethmoid, degree of ossification (mCF7): 0 = sphenethmoid divided middorsally and midventrally; I =
sphenethmoid complete ventrally, divided middorsally; 2 = sphenethmoid complete both dorsally andventrally; 3 = complete dorsally but divided midventrally.
8. Sphenethmoid, ossification of septum nasi: In some species the sphenethmoid, whether complete or not,extends anteriorly beyond the level of the nasals when examined in dorsal view. 0 = septum nasi ossifiedanteriorly underneath nasals; I = septum nasi ossified only up to level of nasals.
9. Sphenethmoid, distance to optic foramen (mCF8): 0 = distance greater than anterior-posterior diameter offoramen; I = distance less than or equal to anterior-posterior diameter of foramen.
C. Maxillary arcade10. Premaxilla, orientation of alary process (CFI2): 0 = perpendicular to horizontal plane of skull as seen in
lateral view; I = anteriorly inclined; 2 = posteriorly inclined.II. Premaxilla, size of lateral process of the pars palatina (CF13, mCJ31): This process is always present in the
species studied, but to varying degrees. In some taxa, the process is shallow or thin, whereas in others it iswide and robust. 0 = process shallow or thin; I = process wide and robust.
12. Premaxilla, orientation of lateral process of the pars palatina (CJ32): 0 = process oriented posterolaterally;I = process oriented posteromedially.
13. Maxilla, depth of pars facialis (mCFI5): This character serves two functions in assessing the features of thenasals as well as the maxilla. These features are individually difficult to compare or quantify. The pars facial is

KAISER ET AL
is expanded dorsally in all taxa studied, but to different degrees. All taxa have a preorbital process of themaxilla and a maxillary process of the nasal. This character assesses how far the lateral shelf (pars maxillaris)of the nasal curves ventrally and how much of the area is uncovered. 0 = pars facialis of maxilla and parsmaxillaris of nasal widely separated; I = pars facialis of maxilla and pars maxillaris of nasal almost touching,touching, or overlapping.
14. Maxilla, anterior "flange" of pars palatina (C F I7): 0 = absent; I = present.15. Maxilla, depth of pars palatina (CF I8, mCJ34): 0 = pars palatina shallower than deepest portion of premax
illary pars palatina; I = pars palatina as deep as or deeper than deepest portion of premaxillary pars palatina.16. Maxilla, posterior extent of maxillary teeth: Numbers of maxillary teeth vary within and between species,
so that a dentition character based on numbers is inappropriate. However, the maxillary tooth row terminateseither posteriorly to the anterior part of the maxillary -quadratojugal articulation or anteriorly to it. 0 =maxillary teeth do not extend to quadratojugal articulation; I = maxillary teeth extend beyond quadratojugalarticulation.
17. Maxilla -quadratojugal overlap (mCF22): In the taxa studied there was always some overlap betweenmaxilla and quadratojugal. A variable feature was the degree of overlap and the resulting free portion of thequadratojugal. 0 = free portion of quadratojugal less than half diameter of anterior-posterior diameter ofsubtemporal fenestra; I = free portion of quadratojugal equal to or greater than half diameter ofanterior-posterior diameter of subtemporal fenestra.
18. QuadratojugaL dorsal enlargement (CF23): 0 = dorsal enlargement of quadratojugal less than pars facialisof maxilla; I = dorsal enlargement of quadratojugal greater than or equal to pars facialis of maxilla.
D. Suspensorium19v. Pterygoid, anterior ramus (CF31): 0 = anterior ramus of pterygoid straight; I = anterior ramus of pterygoid
bowed laterally.20. Pterygoid, medial ramus: In some taxa the medial ramus of the pterygoid narrows medially and is acuminate.
In others the end is expanded or of the same size as the origin of the ramus. 0 = end expanded or of thesame size as origin of ramus; I = end of ramus not expanded, or acuminate.
21. Pterygoid, relation of anterior ramus to orbit (mCF32): In none of the specimens examined does the anteriorramus of the pterygoid reach the planum antorbitale. However, there are two distinct groups of species, somein which the free portion of the anterior ramus extends far forwards, beyond the middle of the orbit, and othersin which that portion is shorter. 0 = free portion of anterior ramus of pterygoid reaches beyond middle ororbit; I = free portion of anterior ramus of pterygoid terminates at or before middle of orbit.
22. Pterygoid, overlap with parasphenoid (MCF43): 0 = pterygoid in contact with lateral alae of parasphenoid;1 = pterygoid not in contact with lateral alae of parasphenoid.
23. Squamosal, orientation in relation to skull roof (CF24): 0 = zygomatic-otic crossbar tilted anteroventrally;I = zygomatic -otic crossbar parallel to skull roof.
24v. Squamosal, lateral profile of ventral ramus (CF25): 0 = ventral ramus straight; I = ventral ramus curved.25. Squamosal, otic ramus (mCF28, mCJ30): 0 = otic ramus absent; I = otic ramus robust; 2 = otic ramus
slender.26. Squamosal, elongation of zygomatic ramus (mCF30, mCJ28): 0 = zygomatic ramus longer than otic ramus;
I = zygomatic ramus shorter than or equal to otic ramus.27. Squamosal, structure of zygomatic ramus in lateral view (mCJ29): 0 = zygomatic ramus slender and
pointed; I = zygomatic ramus robust and expanded.28. M. depressor mandibulae, condition: This character was studied in detail by Lynch (1993), who questioned
its systematic value for the genus Eleutherodactylus and its subgenera. It is included here because Lynch(1993) studied only four West Indian taxa of the subgenus Eleutherodaetylus, one from the subgenus Euhyasand one from the subgenus Syrrhophus, all of which have the dfsq*at condition. 0 = dfvq; 1 = dfsq*at.
E. Palate29. Choana, size relative to dentigerous process of vomer: The dentigerous process sits posteroventrally on the
vomer (= prevomer of some authors). It is composed of a raised stalk or platform that bears a flattened plateof various shapes and, sometimes, teeth. It may cover part or all of the posteromedial region of the vomer.This structure has also been termed the vomerine odontophore, but that term is preoccupied in biological useage by a feature of the molluscan radula (Barnhardt and Barnhardt 1983). The measurement used for comparison is the lateral width of both dentigerous process and choana. In taxa where dentigerous processes aresecondarily lost, C34 and C36-40 are scored as "?" 0 = choana larger than dentigerous process; 1 = choanasame size as dentigerous process; 2 = choana smaller than dentigerous process; 3 = dentigerous process lostsecondarily.
30v. Choana, shape: The choanal dimensions are determined in part by the presence or absence of pre- and postchoanal processes of the vomer; however, individual characteristics of these processes are difficult to assess,and the shape of the choana provides a relationship between them. 0 = round; I = oval; 2 = triangular.
31. Vomer, anterior process (CF37): 0 = anterior process absent; I = anterior process present.32. Vomer, prechoanal process (CF39): 0 = prechoanal process absent; I = prechoanal process present.33. Vomer, postchoanal process: All taxa under investigation have the postchoanal process. The relative robust
ness of this process can be used to distinguish between two discrete groups. 0 = postchoanal process slender;I = postchoanal process robust.
34. Vomers, medial separation: 0 = narrower than greatest width of dentigerous processes; I = as wide as orwider than greatest width of dentigerous processes.
35. Vomer, width at level of anterior margin of postchoanaI process: This characteristic is one way to assess therelative size (width) of the vomer. We prefer using this character to the more general character "size of
2233

2234 CAN. J. ZOOL. VOL. 72, 1994
37.
38.
44.
45.
39.
36.
41.
43.
42.
40.
vomer" (as in CJ37) because it is compared with the size of the premaxilla, a structure reasonably uniformin size among studied taxa. 0 = width less than greatest lateral width of premaxilla; I = width equal to orgreater than greatest lateral width of premaxilla.Vomer, dentigerous process (mCF43, mCJ38-40): 0 = dentigerous process absent; I = dentigerous processpositioned medial to choana; 2 = dentigerous process extending laterally to or beyond medial margin ofchoana.Vomer, shape of dentigerous process in ventral view (mCJ39+40): 0 = triangular; I = arched or weaklyarched; 2 = round; 3 = oval; 4 = shallowly hemispherical.Vomer, orientation of dentigerous processes in ventral view: Where no specific orientation is evident, as maybe the case for rounded dentigerous processes, a "3" is scored. 0 = horizontal; I = anterolaterally toposteromedially inclined; 2 = anteromedially to posterolaterally inclined; 3 = no orientation.Vomer, distribution of vomerine teeth on dentigerous processes: The taxa under investigation usually haveseveral large teeth and often several minor toothlike projections. Considering prominent vomerine teeth only,some taxa have a row of single teeth positioned evenly at the posterior margin of the dentigerous process (seeC42), whereas others have additional teeth in a second, more irregular row. Several taxa also have a fewclumped teeth. 0 = one row of single teeth; I = more than a row of single teeth; 2 = teeth clumped.Vomerine teeth, configuration of teeth (mCF44): 0 = teeth follow posterior margin of dentigerous process;I = teeth reach posterior margin of dentigerous process but angle away from margin anteriorly; 2 = teethclumped in posteromedial corner of dentigerous process.Palatine -vomer relation: In all taxa examined, a palatine bone is present. This bone has also been called"neopalatine" (Trueb 1993) because the palatine is apparently absent in Jurassic anurans (e.g., Vierae/la).However, the use of "neopalatine" is based on Trueb and Cloutier's (1991) analysis of amphibian relationships and a parsimony argument. Given that the influence of development (e.g., delayed ossification) on taxaknown only as fossils is uncertain, we choose the least controversial nomenclature here (akin to the continuedusage of terms such as patella in birds, mammals, and reptiles; R. L. Carroll, personal communication). Adegree of overlap between the palatine and the posterior margin of the vomer can be observed in some species,with the vomer overlying the palatine when examined in ventral view. 0 = palatine and vomer separated;I = vomer overlaps palatine at least partially.Palatine, curvature: The palatine can be straight or curved depending on its relation to the orbit. 0 = palatinestraight; I = palatine bent around anterior edge of orbit.Palatine, shape of medial terminus: In some species the palatine is more prominent than in others, and itswidth at the medial terminus varies accordingly. 0 = pointed and narrow; I = not pointed, as wide as orwider than lateral portion.Parasphenoid, shape of termini of lateral alae (mCF47): 0 = pointed; I = rounded or dilated and rounded;2 = sharply angled.Parasphenoid, anterior end of cultriform process: The cultriform process of some taxa is well rounded,whereas it may be distinctly sharp and pointed, or truncate, in others. 0 = round; I = pointed; 2 = truncate.Parasphenoid, lateral borders of cultriform process (mCF45): 0 = straight; 1 = convex.Parasphenoid, length of cultriform process (mCF46): 0 = cultriform process ends before or just extends tolevel of palatine; I = cultriform process extends beyond level of palatine.
F. Mandible48v. Mento-Meckelian bones, shape in dorsal view (mCF51 + 52): 0 = straight; I = spindle-shaped.49v Angulosplenial, posterior extension (CF53): 0 = angulosplenial terminating at jaw articulation;
angulosplenial extending beyond jaw articulation.
G. Hyolaryngeal apparatus50. Hyoid plate, shape: Several taxa have straight lateral edges parallel to one another, whereas many have edges
that are laterally concave. 0 = lateral edges concave; I = lateral edges parallel.51. Hyoid plate, alary (anterolateral) process of hyoid plate (CF58, CJ44): 0 = absent; I = present.52. Hyoid plate, hyolaryngeal sinus (CF57): In some taxa the broad invagination that invades the hyoid plate (the
hyolaryngeal sinus) extends to or beyond the level of the alary process of the hyoid plate (deep); in othersit never reaches that depth (shallow). This character is coded as "?" when the alary process is absent. 0 =deep; I = shallow.
53. Hyoid plate, mineralization: "Mineralization" is used here preferentially, since we have no evidence forwhich type of mineralization is actually occurring. While in most taxa under investigation there is no evidenceof mineralization of hyoid elements, several mineralize quite distinctively. For example, E. shrevei has theposterior half of the hyoid plate mineralized, extending from the center of the plate to the posterior end, medialto the posterolateral process of the hyoid plate (= posteromedial). Conversely, E. klinikowskii has a narrowmineralized strip at each lateral edge of the hyoid plate, extending from the area midway between theanterolateral and posterolateral processes of the hyoid plate (= lateral). 0 = none; I = lateral; 2 =posteromedial.
54. Larynx (CF64 included): Simple: small larynx (~ half of hyoid plate size); cricoid ring thin or incompletewith a thin esophageal process; flaps of arytenoid cartilage poorly developed without any processes. Complex:large larynx (> half of hyoid plate size); widened cricoid ring with one esophageal process; flaps of arytenoidcartilage wide, expansive, with one or two processes. 0 = simple; I = complex.
II. BodyA. Axial skeleton
55. Vertebra II (axis), degree of expansion of lateral ends of transverse processes (mCF68): 0 = lateral greaterthan medial width; I = lateral equal to medial width.

KAISER ET AL.
56. Vertebra III, orientation of transverse processes (CF69): 0 = lateral; I = posterolateral; 2 = anterolateral.57. Vertebrae 111- IV, lengths of transverse processes (CF70): 0 = V3 longest; I = processes of same length;
2 = Vertebra IV longest.58. Vertebrae V - VIll, length of transverse processes (mCF71): 0 = subequal to width of sacral diapophyses;
I = greater than width of sacral diapophyses.59. Vertebra VIII, orientation of transverse processes (mCF74): 0 = lateral; I = anterolateral.60. Sacral diapophyses, dilation (mCF75): 0 = dilated, slightly wider than medial end; I = not dilated, cylindri
calor round.61. Coccyx, anterior process on dorsal coccygeal ridge (mCF79): 0 = expanded anteriorly; I = not expanded.
B. Pectoral girdle62v. Pectoral girdle, degree of mineralization: In some taxa, there is some degree of mineralization in cartilaginous
elements of the pectoral girdle, whereas in others there is very little. 0 = most elements with mineralization;I = little mineralization.
63. Omosternum, condition (CF90): 0 = cartilaginous or partially mineralized; I = with ossified, bifurcatestyle; 2 = with ossified non-bifurcate style.
64. Clavicle, shape (mCF84): 0 = arched; I = not arched.65. Clavicle, structure: The degree of robustness of the clavicle is indicative of the degree of strengthening the
clavicle has to provide to the pectoral girdle. Whereas some taxa have relatively broad clavicles, others havethin ones. 0 = broadened laterally; I = thin throughout.
66. Coracoid, size (CF80): 0 = lateral end of coracoid wider than medial end; I = medial and lateral ends ofcoracoid equal in width; 2 = medial end of coracoid wider than lateral end.
67. Coracoid, pectoral fenestra (CF82): 0 = bordered medially by epicoracoid cartilage and coracoid; I =bordered medially by coracoid only.
68. Scapula, pars acromial is (CF97): 0 = not expanded; I = expanded.69. Sternum (CJ45): 0 = bifurcated posteriorly; I = elongated rectangular; 2 = pendulum-shaped; 3 = anchor
shaped.
C. Pelvic girdle70. Ilium, dorsal crest: The dorsal crest of the ilium reaches the sacral region in some specimens, whereas it
terminates well before the sacrum in others. 0 = reaches area of articulation with sacrum; I = does not reacharea of articulation with sacrum.
D. Forelimbs71. Terminal phalanges, shape (mCF1I7, mCJ44): The assessment of the shape of the terminal phalange consid
ers the end of finger III only, to minimize problems that may be caused by variation in the same hand. Thereare taxa with straight distal transverse processes (T-shaped) with distally bifurcated (Y-shaped), and withrounded or knoblike ("simple") termini. 0 = T-shaped; I = V-shaped; 2 = simple.
72. Prepollex, number of prepollical elements (CFI12): 0 = one element; I = two elements; 2 = threeelements.
73. Prepollex, degree of ossification: Aside from variation in the number of prepollical elements, there are alsovarying degrees of ossification. 0 = prepollical elements cartilaginous; I = some proximal ossification ofprepollical elements; 2 = complete ossification of prepollux.
74. Preaxial centrale, size in relation to postaxial centrale (CFI06): 0 = equal in size; I = preaxial half of thesize of postaxial; 2 = preaxial one-third size of postaxial.
75. Distal carpale II, fusion (CFIIO): 0 = distal carpale II present as individual bone; I = distal carpale IIfused.
76. Digital sesamoid elements (CF I13): 0 = absent; I = present.n. Metacarpals, length formula (CF I14): There is considerable variation in the relative lengths of metacarpal
elements in the taxa studied. Elements are listed in decreasing size, with integers assigned to metacarpals fromthe innermost to the outermost digit. This character is less variable, but nevertheless of interest, in the metatarsals (C86). 0 = 3-2-4-1; I = 1-3-2-4; 2 = 3-1-2-4; 3 = 3-1-4-2; 4 = 3-2-1-4; 5 = 3-4-2-1.
78. Radioulnar-carpal joint, sesamoid elements: Nussbaum (1982) investigated the presence or absence ofsesamoid bones in the hind limbs. We have found variation in both forelimb and hind limbs in the taxa studied,thus extending Nussbaum's definition to include both the manus and pes. 0 = absent; I = present.
79. Carpal sesamoid elements: 0 = none; I = one; 2 = two.
E. Hind limb80. Terminal phalanages, shape (CJ47): Assessment as for C71 above. 0 = T-shaped; I = V-shaped; 2 =
simple.81. Prehallux, number of prehallical elements (CF I22): 0 = one element; I = two elements, no expansion of
elements; 2 = two elements, with the proximal element expanded laterally.82. Prehallux, degree of ossification: Assessment as for C73 above. 0 = cartilaginous; I = proximal ossification;
2 = complete ossification.83. Distal tarsale II, fusion (C FI19): 0 = distal tarsale 11 present as individual bone; I = distal tarsale 11 fused.84. Tibiofibular-tarsal joint, sesamoid elements: See comment under C78. 0 = absent; I = present.85. Tarsal sesamoid elements: See comment under C78. 0 = none; I = one; 2 = two; 3 = three.86. Metatarsals, length formula (CFI24): See comment under cn. 0 = 4-3-5-2- I; I = 4-5-3-2-1.
External morphology
2235
1. Head87. Snout, shape in dorsal view: 0
------truncate (= trapezoid); I = rounded; 2 acuminate.

2236 CAN. J. ZOOL. VOL. 72. 1994
88. Snout, shape in lateral view: This assessment follows the examples and terms provided by Duellman (1970).o = truncate; I = round; 2 = sloping; 3 = acuminate; 4 = protruding.
89. Mouth, aspect: 0 = terminal; I = subterminaL90. Canthus rostralis, shape in dorsal view: 0 = straight; 1 = concave; 2 = convex.91. Canthus rostral is , distinctiveness: 0 = sharp; 1 = rounded.92. Loreal region, shape in frontal view: Assessment of this character approximately follows the examples in
Rivero (1961; Figs. 1w, y, z), taken just anterior to the orbit. 0 = straight: 1 = slightly angled (90° < x <110°); 2 = oblique (> 110°).
93. Tympanum, distinctiveness (mCll): 0 = distinct; I = indistinct.94. Tympanum, shape: 0 = round; 1 = ovaL95. Supratympanic fold: 0 = pronounced; 1 = weak; 2 = absent.96. Cranial crests (Cl4): 0 = absent; 1 = present.97. Vocal slits (Cl5): 0 = absent; 1 = present.98. Vocal sacs: 0 = absent; 1 = present.99. Tongue, shape: 0 = oval; 1 = rounded; 2 = triangular.
100. Tongue, shape of unattached tip: 0 = round; 1 = notched.
II. Body10 l. Foot webbing: 0 = absent; 1 = remnant; 2 = fully webbed.102. Fingers I and II, relative lengths (mCl6): 0 = I > II; 1 = I = II; 2 = II > I.103. Finger disks III and IV, shape (mCl7): 0 = absent; I = round; 2 = oval to ellipticaL104. Finger disks, size: 0 = disks II, III, and IV of same size; 1 = III and IV larger than I and II but less than
twice their size; 2 = III and IV more than twice as large as I and II.105. Finger disk I, size: 0 = much wider than digit; 1 = barely wider than digit or reduced.106. Toe disks, size: 0 = disks III and IV larger than inner disks; 1 = disks of about the same size.107. Toe disk V, size: 0 = much wider than digit; I = barely wider or reduced.108. Toe Ill, relative length: 0 = III does not reach penultimate subarticular tubercle oflV; 1 = III reaches penulti
mate subarticular tubercle of IV.109. Toe V, length: 0 = V does not extend to distal subarticular tubercle of IV; 1 = V extends to distal subarticular
tubercle of IV.IlOv. Toes IV and V, degree of fusion: 0 = proximal subarticular tubercle of V on free part of digit or right at
interdigital juncture; 1 = IV and V connected up to or beyond proximal subarticular tubercle of V.111. Nuptial pads (ClIO): 0 = absent; 1 = present.112v. Skin, consistency on dorsum (mCl20): The consistency of the skin surface is a rather puzzling character. A
great variety of descriptive terms have been used in the literature, and their use has been inconsistent. Tomake this a more reliable character, we assessed the skin areas between the back of the head and the sacralregion on the dorsum, and between the pectoral and pelvic girdles on the venter. We consulted Peters (1964)in defining terms, with one exception: in disagreement with Peters (1964), we consider "tubercle" not onlyin connection with nonglandular skin bumps (i.e., those of the hands and feet), but also in the context of anysmall raised prominence of glandular nature on the dorsal skin. It is thereby equivalent to "areola (-ae)" forthe venter. For the purposes of this study, we describe texture according to the following definitions:"smooth": no visible tubercles, bumps, prominences, or glands on skin surface; "shagreen": with varyingnumbers of tubercles, bumps, or prominences, spread over the entire surface, referring to dorsal skin only;"areolate": with varying numbers of areolae, referring to ventral skin and regions of the inner groin only.In achieving a more detailed assessment of skin texture, it is usually necessary to use one or more qualifiersin addition to the texture term. We allow the following qualifiers: "fine." with many small tubercles/areolae;'weak," with a mixture of few small and larger tubercles/areolae; "strong," with a mixture of many smalland larger tubercles/areolae; "coarse," with many larger tubercles/areolae; "sparse," with few large tubercles. 0 = smooth; 1 = finely shagreen; 2 = weakly shagreen; 3 = strongly shagreen; 4 = coarsely shagreen;5 = sparsely shagreen.
Appendix 4: TABLE AI. Data matrix for 142 morphological
1 1 2 2 3 3 4 4 5 5 6 6 75 0 5 0 5 0 5 0 5 0 5 0 5 0
amplinympha 00120 12010 11000 10011 11011 00121 10001 10000 11101 01100 10011 10020 11001 111?0antillensis 00121 12011 00010 10001 01011 00121 01101 11100 11000 01100 10001 00100 10001 21101barlagnei 00020 10110 10010 00011 10001 00100 01000 10122 00021 11010 l00?1 00021 10001 01110coqui 00121 11010 10011 10011 01001 00'120 11101 21100 00122 11110 10011 00101 10001 11101euphronides 01120 13110 10010 00000 01012 00000 11011 10000 01111 10101 0'1200 00100 11001 011?0jitzingeri 00020 12012 01000 00000 00012 10012 11100 10000 00121 11111 10011 00101 10000 11100johnstonei 00121 12010 11010 10011 11012 00110 11101 10100 10120 10110 10011 00000 10001 11101martinicensis 01120 10011 11000 10011 11012 00121 11101 10000 11020 11110 10011 01100 10001 01100pinchoni 00120 12012 10010 00011 01012 00110 11111 14100 00110 11110 10001 00111 11001 01111shrevei 00121 12010 10010 00010 00011 00021 11001 10000 01121 11001 0'1201 00100 10001 01100terraebolivaris 02121 10011 11011 00010 00001 00101 11011 10102 00011 11101 0?011 00000 10010 111?0urichi 01121 12012 10100 00011 01012 00102 11010 10122 01110 10111 0?011 00110 11001 011?0
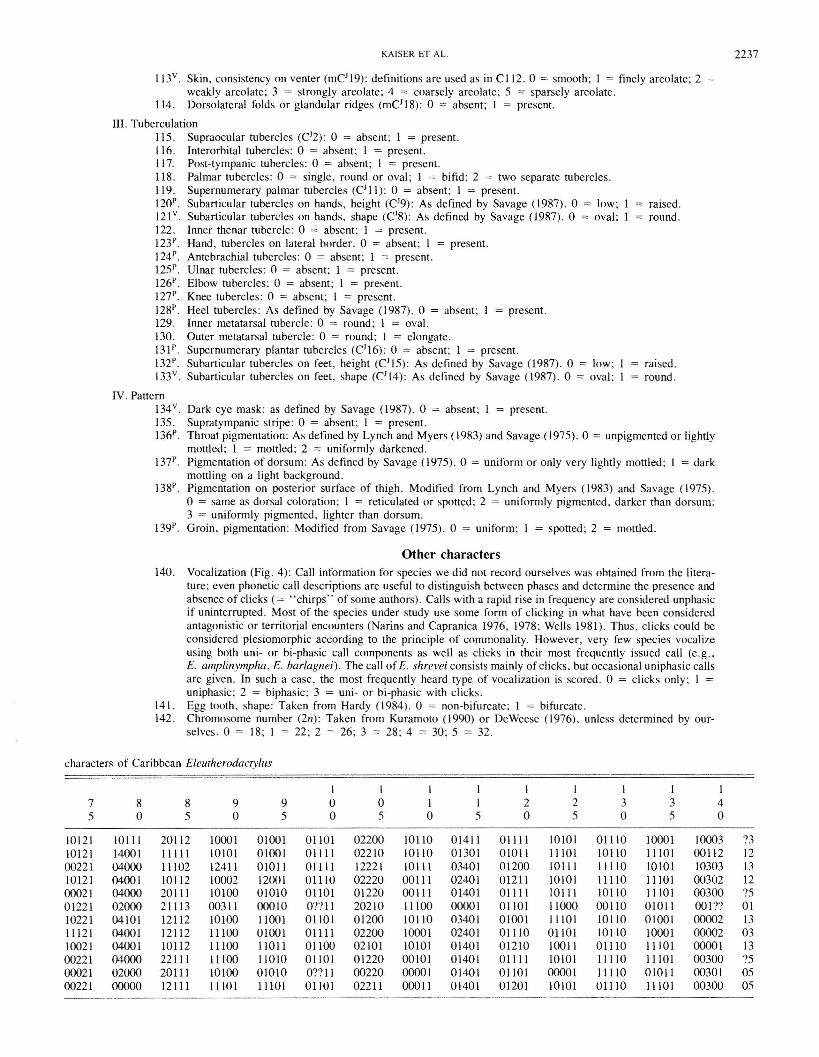
KAISER ET AL.
I13V Skin, consistency on venter (mCJI9): definitions are used as in C112. 0 = smooth; I = finely areolate; 2 =weakly areolate; 3 = strongly areolate; 4 = coarsely areolate; 5 = sparsely areolate.
114. Dorsolateral folds or glandular ridges (mCJI8): 0 = absent; 1 = present.
III. Tuberculation115. Supraocular tubercles (CJ2): 0 = absent; I = present.116. Interorbital tubercles: 0 = absent; I = present.117. Post-tympanic tubercles: 0 = absent; I = present.118. Palmar tubercles: 0 = single, round or oval; I = bifid; 2 = two separate tubercles.119. Supernumerary palmar tubercles (CJ II): 0 = absent; I = present.120P Subarticular tubercles on hands, height (CJ9): As defined by Savage (1987). 0 = low; = raised.121 v. Subarticular tubercles on hands, shape (CJ8): As defined by Savage (1987).0 = oval; = round.122. Inner thenar tubercle: 0 = absent; I = present.123P Hand, tubercles on lateral border. 0 = absent; 1 = present.124P
• Antebrachial tubercles: 0 = absent; I = present.125P
• Ulnar tubercles: 0 = absent; I = present.126P. Elbow tubercles: 0 = absent; I = present.127P Knee tubercles: 0 = absent; I = present.128P Heel tubercles: As defined by Savage (1987). 0 = absent; I = present.129. Inner metatarsal tubercle: 0 = round; I = oval.130. Outer metatarsal tubercle: 0 = round; 1 = elongate.131 P Supernumerary plantar tubercles (CJ I6): 0 = absent; I = present.132P. Subarticular tubercles on feet, height (CJI5): As defined by Savage (1987). 0 = low; = raised.133v. Subarticular tubercles on feet, shape (CJ I4): As defined by Savage (1987).0 = oval: = round.
IV. Pattern134v. Dark eye mask: as defined by Savage (1987). 0 = absent; I = present.135. Supratympanic stripe: 0 = absent; I = present.136P. Throat pigmentation: As defined by Lynch and Myers (1983) and Savage (1975). 0 = unpigmented or lightly
mottled; I = mottled; 2 = uniformly darkened.137P Pigmentation of dorsum: As defined by Savage (1975). 0 = uniform or only very lightly mottled; I = dark
mottling on a light background.138P Pigmentation on posterior surface of thigh. Modified from Lynch and Myers (1983) and Savage (1975).
o = same as dorsal coloration; I = reticulated or spotted; 2 = uniformly pigmented, darker than dorsum;3 = uniformly pigmented, lighter than dorsum.
139P• Groin, pigmentation: Modified from Savage (1975). 0 = uniform; I = spotted; 2 = mottled.
Other characters140. Vocalization (Fig. 4): Call information for species we did not record ourselves was obtained from the litera
ture; even phonetic call descriptions are useful to distinguish between phases and determine the presence andabsence of clicks (= "chirps" of some authors). Calls with a rapid rise in frequency are considered unphasicif uninterrupted. Most of the species under study use some form of clicking in what have been consideredantagonistic or territorial encounters (Narins and Capranica 1976, 1978; Wells 1981). Thus, clicks could beconsidered plesiomorphic according to the principle of commonality. However, very few species vocalizeusing both uni- or bi-phasic call components as well as clicks in their most frequently issued call (e.g.,£. amplinympha, E. barLagnei). The call of E. shrevei consists mainly of clicks, but occasional uniphasic callsare given. In such a case, the most frequently heard type of vocalization is scored. 0 = clicks only; I =uniphasic; 2 = biphasic; 3 = uni- or bi-phasic with clicks.
141. Egg tooth, shape: Taken from Hardy (1984). 0 = non-bifurcate; I = bifurcate.142. Chromosome number (2n): Taken from Kuramoto (1990) or DeWeese (1976). unless determined by our
selves. 0 = 18; I = 22; 2 = 26; 3 = 28; 4 = 30; 5 = 32.
2237
characters of Caribbean ELeutherodacryLus
I I I I I I I I I7 8 8 9 9 0 0 I I 2 2 3 3 45 0 5 0 5 0 5 0 5 0 5 0 5 0
10121 10111 20112 10001 01001 01101 02200 10110 01411 01111 10101 01110 10001 10003 ?310121 14001 11111 10101 01001 01111 02210 10110 01301 01011 11101 10110 11101 00112 1200221 04000 11102 12411 01011 01111 12221 10111 03401 01200 10111 11110 10101 10303 1310121 04001 10112 10002 12001 01110 02220 00111 02401 01211 10101 11110 11101 00302 1200021 04000 20111 10100 01010 01101 01220 00111 01401 01111 10111 10110 11101 00300 7501221 02000 21113 00311 00010 07711 20210 11100 00001 01101 11000 00110 01011 001?7 0110221 04101 12112 10100 11001 01101 01200 10110 03401 01001 11101 10110 01001 00002 1311121 04001 12112 11100 01001 01111 02200 10001 02401 01110 01101 10110 10001 00002 0310021 04001 10112 11100 11011 01100 02101 10101 01401 01210 10011 01110 11101 00001 1300221 04000 22111 11100 11010 01101 01220 00101 01401 01111 10101 11110 11101 00300 7500021 02000 20111 10100 01010 0?711 00220 00001 01401 01101 00001 11110 01011 00301 0500221 00000 12111 11101 11101 01101 02211 00011 01401 01201 10101 01110 11101 00300 05