synergistic activation of vascular trpc6 channel by receptor and mechanical stimulation via...
TRANSCRIPT

Synergistic Activation of Vascular TRPC6 Channel byReceptor and Mechanical Stimulation via PhospholipaseC/Diacylglycerol and Phospholipase A2/�-Hydroxylase/
20-HETE Pathways
Ryuji Inoue,* Lars J. Jensen,* Zhong Jian, Juan Shi, Lin Hai, Andrew I. Lurie, Freja H. Henriksen,Max Salomonsson, Hiromitsu Morita, Yasuhiro Kawarabayashi, Masayuki Mori, Yasuo Mori, Yushi Ito
Abstract—TRPC6 is a non–voltage-gated Ca2� entry/depolarization channel associated with vascular tone regulation andremodeling. Expressed TRPC6 channel responds to both neurohormonal and mechanical stimuli, the mechanismfor which remains controversial. In this study, we examined the possible interactions of receptor and mechanicalstimulations in activating this channel using the patch clamp technique. In HEK293 cells expressing TRPC6,application of mechanical stimuli (hypotonicity, shear, 2,4,6-trinitrophenol) caused, albeit not effective bythemselves, a prominent potentiation of cationic currents (ITRPC6) induced by a muscarinic receptor agonistcarbachol. This effect was insensitive to a tarantula toxin GsMTx-4 (5 �mol/L). A similar extent of mechanicalpotentiation was observed after activation of ITRPC6 by GTP�S or a diacylglycerol analog 1-oleoyl-2-acetyl-sn-glycerol (OAG). Single TRPC6 channel activity evoked by carbachol was also enhanced by a negative pressureadded in the patch pipette. Mechanical potentiation of carbachol- or OAG-induced ITRPC6 was abolished by smallinterfering RNA knockdown of cytosolic phospholipase A2 or pharmacological inhibition of �-hydroxylation ofarachidonic acid into 20-HETE (20-hydroxyeicosatetraenoic acid). Conversely, direct application of 20-HETEenhanced both OAG-induced macroscopic and single channel TRPC6 currents. Essentially the same results wereobtained for TRPC6-like cation channel in A7r5 myocytes, where its activation by noradrenaline or Arg8vasopressin was greatly enhanced by mechanical stimuli via 20-HETE production. Furthermore, myogenicresponse of pressurized mesenteric artery was significantly enhanced by weak receptor stimulation dependently on20-HETE production. These results collectively suggest that simultaneous operation of receptor and mechanicalstimulations may synergistically amplify transmembrane Ca2� mobilization through TRPC6 activation, therebyenhancing the vascular tone via phospholipase C/diacylglycerol and phospholipase A2/�-hydroxylase/20-HETEpathways. (Circ Res. 2009;104:1399-1409.)
Key Words: mechanotransduction � Gq/11 protein–coupled receptor � Ca2� entry channel
It has widely been recognized that cellular functions areelaborately controlled by both neurohormonal activities
and mechanical stresses.1,2 For example, in the cardiovascularsystem, the contractile status of arteries is not only regulatedby autonomic nerve activities, circulating vasoactive hor-mones, and paracrinely released autacoids but is also greatlyinfluenced by mechanical stresses such as blood flow andpressure.3–5 These 2 mechanisms are expected to operate
simultaneously in vivo, but little is known about how theycould interact mutually.
A member of the canonical transient receptor potential(TRP) protein family TRPC6 (the sixth canonical subfam-ily member of TRP protein) is ubiquitously expressed invascular smooth muscle cells (VSMCs) and implicated inthe regulation of vascular tone and remodeling.6 Likemany other TRP isoforms, activation of TRPC6 channel
Original received December 21, 2008; revision received May 4, 2009; accepted May 5, 2009.From the Department of Physiology (R.I., Z.J., L.H., Y.K., M.M.), Graduate School of Medical Sciences, Fukuoka University, Japan; Department of
Biomedical Sciences (L.J.J., A.I.L., F.H.H., M.S.), University of Copenhagen, The Panum Institute, Denmark; Institute of Biomedical Engineering (Z.J.),School of Life Science and Technology, Xi’an Jiaotong University, Xi’an, China; Department of Anatomy and K. K. Leung Brain Research Centre (J.S.),the Fourth Military Medical University, Xi’an, China; Special Patient Oral Care Unit (H.M.), Kyushu University Hospital, Fukuoka, Japan; Departmentof Synthetic Chemistry and Biological Chemistry (Y.M.), Graduate School of Engineering, Kyoto University, Japan; and Research Division for LifeSciences (Y.I.), Kumamoto Health Science University, Japan.
Presented in part at the 85th Annual Meeting of the Japanese Physiological Society, Tokyo, Japan, March 25–27, 2008, and published in abstract form(J Physiol Sci. 2008;58:S9).
*Both authors contributed equally to this work.Correspondence to Ryuji Inoue, MD, PhD, Department of Physiology, Graduate School of Medical Sciences, Fukuoka University, Nanakuma 7-45-1,
Jonan-ku, Fukuoka 814-0180, Japan. E-mail [email protected]© 2009 American Heart Association, Inc.
Circulation Research is available at http://circres.ahajournals.org DOI: 10.1161/CIRCRESAHA.108.193227
1399 by guest on March 6, 2016http://circres.ahajournals.org/Downloaded from by guest on March 6, 2016http://circres.ahajournals.org/Downloaded from by guest on March 6, 2016http://circres.ahajournals.org/Downloaded from by guest on March 6, 2016http://circres.ahajournals.org/Downloaded from by guest on March 6, 2016http://circres.ahajournals.org/Downloaded from by guest on March 6, 2016http://circres.ahajournals.org/Downloaded from by guest on March 6, 2016http://circres.ahajournals.org/Downloaded from by guest on March 6, 2016http://circres.ahajournals.org/Downloaded from by guest on March 6, 2016http://circres.ahajournals.org/Downloaded from by guest on March 6, 2016http://circres.ahajournals.org/Downloaded from by guest on March 6, 2016http://circres.ahajournals.org/Downloaded from by guest on March 6, 2016http://circres.ahajournals.org/Downloaded from by guest on March 6, 2016http://circres.ahajournals.org/Downloaded from by guest on March 6, 2016http://circres.ahajournals.org/Downloaded from by guest on March 6, 2016http://circres.ahajournals.org/Downloaded from by guest on March 6, 2016http://circres.ahajournals.org/Downloaded from by guest on March 6, 2016http://circres.ahajournals.org/Downloaded from

has been reported to occur in a polymodal fashion. Inseveral different types of blood vessels, stimulation ofphospholipase (PL)C-coupled receptors has been shown toactivate this channel via generation of diacylglycerol(DAG). In a few arteries in which the pressure-dependentautoregulation of blood flow is prominent (eg, cerebralartery), an elevation of intravascular pressure itself isthought to cause the activation of TRPC6 channel, therebyeliciting a vasoconstriction referred to as “myogenicresponse.”4,6,7
The mechanism underlying the mechanical activation ofTRPC6 channel is controversial. It was initially explainedthat DAG generated through mechanical activation of PLC7,8
secondarily activates TRPC6 channel.7 In contrast, a subse-quent study proposed that TRPC6 channel can be directlyactivated by mechanical deformation of the cell membranevia a lipid bilayer–dependent mechanism.9 The latter pro-posal was however opposed by the finding that the observedmechanosensitive cation channel in TRPC6-expressing cellmay reflect an endogenous mechanosensitive cation channelactivity in the expression system.10 Furthermore, recent inde-pendent lines of evidence have provided an alternative viewthat Gq/11 protein/PLC-coupled receptors may themselves bemechanically activated,11 and their increased cell surfacedensity may allow TRPC6 channel to activate in response tomechanical stimuli.12
The purpose of the present study, therefore, was to explorea possible explanation about the discrepant findings describedabove on the mechanosensitivity of TRPC6 channels andseek its physiological significance in native vascular tissues.As the result, we have found that TRPC6 channel exhibitsmechanosensitivity only after its receptor-mediated activa-tion, in which a metabolite of arachidonic acid (AA), 20-HETE (20-hydroxyeinosatetraenoic acid), a reportedly majorvasoconstrictive lipid mediator,13 likely plays a pivotal role.Furthermore, additional evidence has suggested that thismechanism may also contribute to the activation ofvasoconstrictor-activated cation channels in A7r5 myocytesand operate in an intact mesenteric artery under receptorstimulation that is otherwise poorly responsive to increasedintravascular pressure.
Materials and MethodsAn expanded Materials and Methods section is available in theonline data supplement at http://circres.ahajournals.org.
Cell Culture and TransfectionHuman embryonic cells HEK293 and embryonic aortic smoothmuscle myocytes A7r5 (American Type Culture Collection, Manas-sas, Va) were maintained in 10% FBS-containing DMEM. HEK cellswere used to cotransfect TRPC6 and CD8 cDNA (used as theexpression marker) plasmids with a transfection reagent SuperFect(Qiagen, Hilden, Germany). Functional evaluation was made 48 to72 hours after transfection. TRPC6-expressing cells were selectedwith bead-conjugated anti-CD8 antibody (Dynabeads M-450 CD8,Dynal Biotech).
Stealth Small Interfering RNA was transfected to HEK293 andA7r5 cells according to the instructions of the manufacturer.
ElectrophysiologyThe details of macroscopic and single channel [cell-attached (C/A),inside-out (I/O)] current recordings and data analyses have been
described by Shi et al.14 To minimize variations arising fromdifferent cell size, the magnitude of current is normalized by dividingby cell capacitance.
Shear force (�10 dyn�cm�2; see the online data supplement) wasapplied as a continuous laminar flow (�1 �L � s�1) out of a Y-tube,which was initially placed at a distance of �500 �m and quicklymoved within 20 �m from a voltage-clamped cell.
Negative pressure was applied manually by suction via patchpipette, the magnitude of which was scaled and monitored byusing a DMP-1B pneumatic transducer tester (BioTek Instru-ments, Winooski, Vt).
All experiments were performed at room temperature (22°C to 26°C).
Video Microscopic Diameter MeasurementArterial segments (second or third branches) of mesenteric arteryobtained from male Sprague–Dawley rats (Charles River, Japan)were cannulated at both ends with fine-tipped glass capillaries withnylon threads. Intraluminal pressure was controlled using a pressuremyograph.15
All procedures used for animal experiments were approved bylocal animal ethics committees of Fukuoka University and Univer-sity of Copenhagen.
SolutionsThe composition of solutions used in the present study was the sameas described previously.14 Hypotonic solutions (HTSs) were made togive a desired hypotonicity by partially replacing external NaCl withsucrose or mannitol. Na�-free external solution was made byequimolar substitution of monovalent cations with N-methyl-D-glucamine (NMDG).
ChemicalsHET0016 was kindly provided by Taisho Pharmaceuticals (Tokyo,Japan).
StatisticsAll data are expressed as means�SEM (indicated by columns/symbols and bars in each figure). Two-tailed paired and unpairedStudent’s t tests were used to evaluate statistical significance:P�0.05 (*) or P�0.01 (**). For multiple comparisons, ANOVAfollowed by Bonferroni’s t test was used.
For more details of the methods, see the online data supplement.
ResultsFour Distinct Mechanical Stimuli PotentiateReceptor-Activated TRPC6 CurrentsWe first investigated whether TRPC6 channels expressed inHEK293 cells can be directly activated by HTS.9 However, 1minute of exposure to HTS (�75 mOsm) only occasionallyactivated a small and slowly developing inward current inTRPC6-expressing HEK cells (Figure 1A). This current wasstrongly suppressed by pretreatment with DIDS(300 �mol/L) but unchanged on substitution of external Na�
and K� with NMDG (Figure 1A and Online Figure I, A andB), thus likely reflecting a noncationic conductance. Stimu-lation of intrinsic muscarinic receptors in HEK cells withcarbachol (CCh) (1 to 100 �mol/L) elicited a much noisiernonselective cationic current (reversal potential, 0.5�0.9mV; n�8) showing weak inward suppression and outwardrectification (Figure 1B), the hallmarks of TRPC6 channel.This current (hereafter designated as ITRPC6) was insensitiveto 300 �mol/L DIDS (Figure 1A and Online Figure I, C) or5 �mol/L GsMTx-4 (Figure 2A and 2B and Online Figure II),a tarantula toxin reportedly inhibiting HTS-induced inwardcurrents in TRPC6-expressing cells.9 In the rest of the present
1400 Circulation Research June 19, 2009
by guest on March 6, 2016http://circres.ahajournals.org/Downloaded from

study, to eliminate possible contribution of a slowly devel-oping HTS-induced noncationic current, we added300 �mol/L DIDS into the external solution.
In the next step, to examine whether mechanical stressaffects the receptor-mediated activation of TRPC6, we ob-served the effects of HTS on ITRPC6 after the administration ofCCh. Strikingly, the CCh-induced ITRPC6 was reversiblyenhanced in response to HTS (Figure 1A) without significantchange in the reversal potential (1.2�0.9 mV, n�8; Figure1B; not significantly different with unpaired t test from thecontrol above). In nontransfected HEK cells, however, littlecurrent was induced by CCh without discernible effects ofHTS (Figure 1A, inset). The extent of enhancement of theCCh-induced ITRPC6 became more pronounced with increas-ing hypotonicity (Figure 1C) or weaker preceding receptorstimulation (Figure 1D) but remained unchanged in the
presence of 5 �mol/L GsMTx-4 (Figure 2B, b). SimilarHTS-induced enhancement of ITRPC6 was also observed whenit was activated by intracellular perfusion of a nonhydrolyz-able GTP analog GTP�S (10 �mol/L) or a diacylglycerolanalog 1-oleoyl-2-acetyl-sn-glycerol (OAG) (10 �mol/L)(Figure 1E).
In another series of experiments, we used other means togenerate mechanical forces, shear force and a bulge-formingamphipathic agent 2,4,6-trinitrophenol (TNP). Both shearforce (�10 dyn/cm2) and TNP (500 �mol/L), which wereineffective in the absence of receptor stimulation (insets inFigure 2C, a and b), markedly enhanced the magnitude ofITRPC6 activated by CCh (1 �mol/L) or intracellularly per-fused GTP�S (10 �mol/L) (Figure 2C, a and b, and 2D).
The mechanical enhancement of receptor-activated ITRPC6
could also be reproduced at the single channel level. As
Figure 1. Hypotonic stress potentiates receptor-activated TRPC6 current (ITRPC6). Bath and pipette contained external (300 �mol/LDIDS added) and internal solutions, respectively, throughout this study, unless otherwise described. A, Representative trace showingthe effects of HTSs (�75 mOsm) on CCh-induced (10 �mol/L) ITRPC6. TRPC6-HEK indicates TRPC6-expressing HEK cell. Inset, Effectsof CCh and HTS in a nontransfected HEK cell. B, Current–voltage relationships of CCh-induced (10 �mol/L) ITRPC6 evaluated by a rampvoltage (2 seconds, �100 to 100 mV) before and after exposure to HTS (�75 mOsm). C, HTS-induced fold increase of ITRPC6. The foldincrease is defined as the ratio of peak ITRPC6 amplitudes after to just before application of HTS. D, Effect of HTS (�75 mOsm) onweakly receptor-activated ITRPC6 (1 �mol/L CCh) (a) and the fold increase of ITRPC6 by HTS (�75 mOsm) with different concentrations ofCCh (b). E, Enhancing effects of HTS (�75 mOsm) on ITRPC6 induced by intracellular perfusion of GTP�S (10 �mol/L) or OAG(10 �mol/L) via a patch pipette. *Statistically significant with paired t test (in E).
Inoue et al Chemical–Mechanical Synergy on TRPC6 Activation 1401
by guest on March 6, 2016http://circres.ahajournals.org/Downloaded from

demonstrated and summarized in Figure 3, virtually no currentswere evoked by a negative pressure up to �60 mm Hg (appliedin the patch pipette: Figure 3A, a, and left columns in Figure3B). However, after receptor activation (CCh 10 �mol/L in thepipette), the activity of �30 pS single cation channels wasincreased in a graded fashion depending on the intensity ofnegative pressure (�20 to �60 mm Hg; Figure 3A, b, and rightcolumns in Figure 3B). The enhancing effect of negativepressure on CCh-induced single TRPC6 channel activityshowed some delay in onset and outlasted the period of pressure
application. In addition, regardless of TRPC6 protein expression,application of a strong negative pressure (��80 mm Hg) oftenled to irreversible development of irregular inward currents(Online Figure III).
The Mechanism Underlying the MechanicalEnhancement of CCh-Induced ITRPC6The time course of mechanical enhancement of receptor-ac-tivated ITRPC6 (time to 50% activation and deactivation with�75 mOsm HTS: 11.8�1.4 and 13.3�1.7 seconds, respec-
Figure 2. ITRPC6 is insensitive to GsMTx-4 and potentiated by shear stress and a membrane bulging agent TNP after receptor activa-tion. A, An actual trace displaying no effect of 5 �mol/L GsMTx-4 on CCh-induced (100 �mol/L) ITRPC6. B, Current density of CCh-induced ITRPC6 (a) and fold increase in CCh-induced (1 �mol/L) ITRPC6 by HTS (�75 mmol/L) (b) in the absence and presence of5 �mol/L GsMTx-4. The relative change is defined as the ratio of ITRPC6 3 minutes after to just before application of GsMTx-4. C,Effects of 500 �mol/L TNP (a) or fluid flow shear force (b) on membrane currents induced by bath-applied CCh (1 �mol/L) (a) or intra-cellularly applied GTP�S (10 �mol/L) (b) and on those in the absence of agonists (insets). D, Fold increase of ITRPC6 by distinctmechanical stimuli. ITRPC6 was activated by either 1 �mol/L CCh or 10 �mol/L GTP�S.
Figure 3. Negative pressure potentiatessingle TRPC6 channel activity. A, Actualtraces of cell-attached recordings, in theabsence (a) and presence (b) of10 �mol/L CCh in the pipette (1 mmol/LTEA and 300 �mol/L DIDS were added toblock K� and Cl� channels). At the dou-ble bar, suction (negative pressure:�20 mm Hg) was applied in the pipette.In b, single TRPC6 channel activities (�50mV) induced by CCh was potentiated bysuction. B, Mean single TRPC6 channelcurrents (NPoi) at various negative pres-sures (0, �20, �40, �60 mm Hg) with(right columns) or without (left columns)10 �mol/L CCh in the pipette.
1402 Circulation Research June 19, 2009
by guest on March 6, 2016http://circres.ahajournals.org/Downloaded from

tively; n�15) was much slower than that expected for a“direct” bilayer-dependent activation.2 We thus reasoned thatlipid mediators, especially the metabolites of AA, may beinvolved in this enhancement, as was previously reported fora distinct TRP family member TRPV4.16 Consistent with thisspeculation, pharmacological inhibition of PLA2 with AA-COCF3 (10 �mol/L) or pBPB (200 �mol/L) (pretreated for 5
to 10 minutes; Figure 4A, b), or specific small interfering(si)RNA knockdown of cytosolic PLA2 (Figure 4A, b) almostcompletely abolished HTS- or TNP-induced enhancement ofreceptor-activated ITRPC6. A similar extent of inhibition wasalso observed with 17-ODYA and HET0016 (pretreated for 5to 10 minutes), the inhibitors for �/��-hydroxylase, whichmetabolizes AA into 20-HETE (Figure 4A, a and b). These
Figure 4. 20-HETE mediates mechanical potentiation of ITRPC6. A, Abolishing effects of HET0016 (1 �mol/L) pretreatment (5 minutes) onHTS-induced potentiation of ITRPC6 (a). Averaged fold increase of ITRPC6 by HTS (�75 mOsm) or TNP (500 nmol/L) with various drugs(left) or siRNA knockdown of cytosolic PLA2 (right). The drug concentration used (�mol/L) was 10 for AACOCF3, NDGA, indomethacin(Indo), miconazole (Micon), and 17-ODYA; 200 for pBPB; and 1 to 10 for HET0016. The drugs were pretreated for 5 to 10 minutes. Sta-tistical evaluation was made against the left-most columns as controls. Inset in b, Immunoblots of cPLA2 in HEK cells treated withsiRNA for cPLA2 (si-cPLA2) and negative control (si-con). B, Increasing effects of bath-applied 20-HETE (100 nmol/L) on ITRPC6 inducedby 10 �mol/L OAG (a) and averaged fold increase of ITRPC6 by 100 nmol/L 20-HETE (b). C, Dramatic potentiating effects of 20-HETE(100 nmol/L; applied intracellularly) on single OAG-induced (10 �mol/L) TRPC6 channel activity (�50 mV) in inside-out (I/O) patch configura-tion (a) and summary of potentiating effects of 20-HETE (100 nmol/L) on TRPC6 channel activity (NPoi) under various conditions (b).
Inoue et al Chemical–Mechanical Synergy on TRPC6 Activation 1403
by guest on March 6, 2016http://circres.ahajournals.org/Downloaded from

inhibitors were without effects on ITRPC6 when appliedacutely. In contrast, the inhibitions of cyclooxygenase, li-pooxygenase, and epoxygenase by indomethacin, NDGA,and miconazole (each 10 �mol/L), respectively, were allineffective (Figure 4A, b). These results suggest the involve-ment of 20-HETE in mechanical potentiation of receptor-ac-tivated ITRPC6. To further corroborate this idea, we nextapplied 20-HETE exogenously. As shown in Figure 4B and4C, direct application of 20-HETE (100 nmol/L) significantlyenhanced both macroscopic ITRPC6 and single TRPC6 channelcurrents induced by OAG (10 �mol/L), although applicationof 20-HETE alone failed to induce a detectable current up to10 �mol/L (left-most column in Figure 4C, b; not shown formacroscopic current). Furthermore, the enhancing effects of20-HETE were not significantly affected by the PLC inhibitorU73122 (1 �mol/L) (Figure 4B, b, and 4C, b), thus excludingthe involvement of PLC therein.
These results together suggest that activation of the cyto-solic PLA2/�-hydroxylase cascade and consequent produc-tion of 20-HETE is essential for the mechanical enhancementof receptor-activated TRPC6 current/channel activity.
Vasoconstrictor-Activated Cation Channels inA7r5 Myocytes Become Susceptible to MechanicalStimuli After Receptor ActivationTo explore the physiological significance of above-mentioned observations, we repeated the same experiments asabove in a native VSMC model A7r5 myocyte. In thismyocyte, vasoconstrictors such as noradrenaline (Norad),vasopressin, angiotensin, and endothelin were found to acti-vate TRPC6-like currents.17–19 Consistent with these previousfindings, abundant expression of TRPC6 was detected by theRT-PCR technique, and specific siRNA knockdown ofTRPC6 almost abolished Norad- or Arg8-vasopressin (AVP)-induced cation current in parallel with reduced TRPC6protein expression (Online Figure IV).19
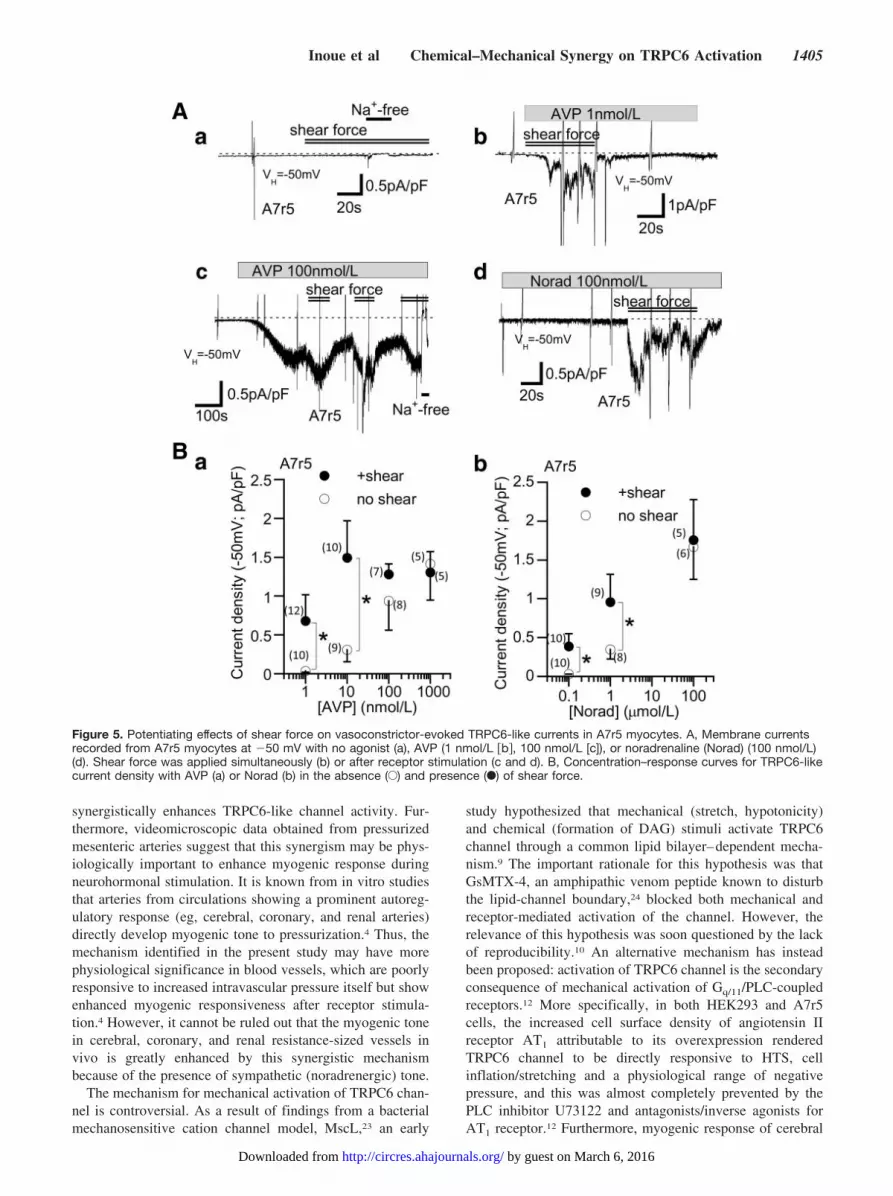
TRPC6-like currents evoked by Norad or AVP in A7r5myocytes were greatly enhanced by mechanical stimuli(HTS, shear force), whether they were applied together withor after receptor stimulation (Figures 5A, b through d, and6A, a). These mechanical stimuli were however ineffective inthe absence of receptor stimulation (300 �mol/L DIDSadded; Figure 5A, a). The mechanical enhancement ofTRPC6-like currents was more pronounced at lower concen-trations of receptor agonists. As summarized in Figure 5B, aand b, application of shear force caused a more than 10-foldleftward shift in the dose–response relationship betweenagonist concentration and the density of evoked current (openversus filled circles).
Potentiating effects of mechanical stimuli were still ob-served when TRPC6-like currents were more directly acti-vated by intracellular perfusion of GTP�S (10 �mol/L) orOAG (10 �mol/L) (Figure 6A, b, and 6C). As in the case ofexpressed TRPC6, this potentiation was greatly attenuated bypretreatment with AACOCF3 (10 �mol/L) or HET0016(10 �mol/L) and siRNA knockdown of cytosolic PLA2
(Figure 6B through 6D). In contrast, neither GsMTx-4(5 �mol/L) nor the PLC inhibitor U73122 (1 �mol/L) nor theprotein kinase C blocker GF109203X (2 �mol/L), which was
reported to inhibit the mechanical enhancement of endo-thelin-1–activated, GsMTx-4 –inhibitable single cationchannels in pulmonary and cerebral arterial myocytes,20
attenuated the mechanical potentiation of TRPC6-likecurrents (Online Figure V).
Consistent with the above macroscopic data, singleTRPC6-like channel activity in A7r5 myocyte evoked by alow concentration of AVP (3 nmol/L) (reversal potential:�2.3�1.8 mV, n�5; unitary conductance: 32.6�1.1 pS,n�5; Figure 7A, c) was also markedly enhanced by anegative pipette pressure in the range of �20 to�80 mm Hg (Figure 7A, a and b, 7C, and 7D). Further-more, this enhancement diminished after pretreatment withHET0016 (10 �mol/L) (Figure 7C and 7D). In the absenceof AVP, however, the negative pressure could inducevirtually no single channel activities (Figure 7B and opencircles in Figure 7C).
These results collectively suggest that a 20-HETE–medi-ated mechanism similar to that observed for expressedTRPC6 channel likely operates in synergistic activation ofTRPC6-like channels in A7r5 myocytes by neurohormonaland mechanical stimulations.
“Sensitized” Myogenic Response Is Mediated by20-HETE ProductionFinally, to get more physiological insight into the abovemechanism at the tissue level, we performed a videomicro-scopic diameter measurement of pressurized rat mesentericartery where expression of TRPC6 can be detected byRT-PCR (Figure 8A). Large (second or third) branches of thisartery were reported to respond poorly to elevated intravas-cular pressure in the absence of receptor stimulation.21 Asdemonstrated and summarized in Figure 8, the sustainedphase of pressure-induced diameter decrease (myogenic re-sponse) was marginal in the absence of agonist (filledtriangles in Figure 8C). However, after partial vasoconstric-tion with the �1-adrenergic agonist phenylephrine (or nor-adrenaline) and neuropeptide Y, the magnitude of myogenicresponse was greatly enhanced (Figure 8B and filled circlesin Figure 8C through 8E). This enhanced response was almostabolished by Gd3� or SKF96365 at concentrations thateffectively inhibit expressed TRPC6 channels (2 and5 �mol/L, respectively; Figure 8E)22 but are ineffective forexcess K�-induced vasoconstriction (Figure 8F). Impor-tantly, pretreatment with HET0016 (10 �mol/L) stronglycounteracted this enhanced myogenic response (open circlesin Figure 8C and 8D). These results suggest that 20-HETElikely mediates the sensitized myogenic response by �1-adrenergic receptor stimulation which involves the activationof TRPC6.
DiscussionThe present results have clearly demonstrated that expressedTRPC6 channel is not primarily activated by mechanicalstimuli. Instead, if once receptor-activated, it becomes mech-anosensitive via production of a PLA2/�-hydroxylase metab-olite 20-HETE. A similar mechanism seems also operative ina native VSMC model A7r5 myocyte, where simultaneousactivation of PLC- and PLA2-coupled signaling pathways
1404 Circulation Research June 19, 2009
by guest on March 6, 2016http://circres.ahajournals.org/Downloaded from
synergistically enhances TRPC6-like channel activity. Fur-thermore, videomicroscopic data obtained from pressurizedmesenteric arteries suggest that this synergism may be phys-iologically important to enhance myogenic response duringneurohormonal stimulation. It is known from in vitro studiesthat arteries from circulations showing a prominent autoreg-ulatory response (eg, cerebral, coronary, and renal arteries)directly develop myogenic tone to pressurization.4 Thus, themechanism identified in the present study may have morephysiological significance in blood vessels, which are poorlyresponsive to increased intravascular pressure itself but showenhanced myogenic responsiveness after receptor stimula-tion.4 However, it cannot be ruled out that the myogenic tonein cerebral, coronary, and renal resistance-sized vessels invivo is greatly enhanced by this synergistic mechanismbecause of the presence of sympathetic (noradrenergic) tone.
The mechanism for mechanical activation of TRPC6 chan-nel is controversial. As a result of findings from a bacterialmechanosensitive cation channel model, MscL,23 an early
study hypothesized that mechanical (stretch, hypotonicity)and chemical (formation of DAG) stimuli activate TRPC6channel through a common lipid bilayer–dependent mecha-nism.9 The important rationale for this hypothesis was thatGsMTX-4, an amphipathic venom peptide known to disturbthe lipid-channel boundary,24 blocked both mechanical andreceptor-mediated activation of the channel. However, therelevance of this hypothesis was soon questioned by the lackof reproducibility.10 An alternative mechanism has insteadbeen proposed: activation of TRPC6 channel is the secondaryconsequence of mechanical activation of Gq/11/PLC-coupledreceptors.12 More specifically, in both HEK293 and A7r5cells, the increased cell surface density of angiotensin IIreceptor AT1 attributable to its overexpression renderedTRPC6 channel to be directly responsive to HTS, cellinflation/stretching and a physiological range of negativepressure, and this was almost completely prevented by thePLC inhibitor U73122 and antagonists/inverse agonists forAT1 receptor.12 Furthermore, myogenic response of cerebral
Figure 5. Potentiating effects of shear force on vasoconstrictor-evoked TRPC6-like currents in A7r5 myocytes. A, Membrane currentsrecorded from A7r5 myocytes at �50 mV with no agonist (a), AVP (1 nmol/L �b�, 100 nmol/L [c]), or noradrenaline (Norad) (100 nmol/L)(d). Shear force was applied simultaneously (b) or after receptor stimulation (c and d). B, Concentration–response curves for TRPC6-likecurrent density with AVP (a) or Norad (b) in the absence (E) and presence (F) of shear force.
Inoue et al Chemical–Mechanical Synergy on TRPC6 Activation 1405
by guest on March 6, 2016http://circres.ahajournals.org/Downloaded from

artery was profoundly inhibited by the inverse agonist of theAT1 receptor, losartan.12 Although direct evidence is stilllacking, these results have been interpreted to indicate thatmembrane stretch can directly cause an agonist-independentconformational change of the receptor thereby activating thedownstream Gq/11/PLC/DAG/TRPC6 signaling cascade.11,12
Our present results have confirmed the observation thatTRPC6 channel per se is not mechanosensitive. Total insen-sitivity of receptor-activated and mechanically potentiatedTRPC6 current to GsMTx-4 further supports that altered lipidbilayer mechanics would not be a primary mechanism toactivate the channel. However, at an endogenous expressionlevel of Gq/11/PLC-coupled receptors that we used, disparateobservations were obtained. Firstly, receptor-mediated acti-vation of TRPC6 channel is a prerequisite for its responsive-ness to mechanical stimuli. Secondly, mechanical activationof TRPC6 channel likely involves the generation of a PLA2
metabolite 20-HETE rather than PLC metabolites. It has beenreported that some G protein–coupled receptors (GPCRs)show a constitutive activity being capable of activating thedownstream signaling cascades without binding of agonists,which is effectively decoupled by inverse agonists.25 If thiswere the case for the above study, the possibility must betaken into account that spontaneous activation of PLC, which
would be greatly exaggerated by the overexpression ofGPCRs, could produce a sufficient amount of DAG toactivate TRPC6 channel. Indeed, our findings that even weakreceptor activation could induce a prominent mechanosensi-tivity of TRPC6, and that TRPC6-expressing cells exhibitingan exclusively high spontaneous activity responded directlyto mechanical stimuli (Online Figure VII), may support thisspeculation. In the above study using overexpression ofGPCRs,12 however, the potential contribution of 20-HETEproduction was not tested. It is thus a remaining unresolvedissue to clarify to what extent PLC- and PLA2-mediatedsignaling pathways respectively contribute to mechanicalactivation of TRPC6 channel in physiological settings.
20-HETE was previously reported to directly activate asmall cationic current in TRPC6-overexpressing HEK cells(maximum current density: �2pA/pF at �40 mV with10 �mol/L).26 However, the concentration required to acti-vate the current (EC50�0.8 �mol/L) was much higher thanthat used in the present study (100 nmol/L), and we couldrecord little discernible current at the latter concentration(Figure 4C, b). Furthermore, the observed properties ofcationic current evoked by 20-HETE (linear current–voltagerelationship; no sensitivity to flufenamate)26 are atypical ofTRPC6 channel.22 These observations suggest that direct
Figure 6. 20-HETE production is essentialfor mechanical potentiation of TRPC6-likecurrents in A7r5 myocytes. A, HTS-induced (�75 mOsm) or shear force–in-duced enhancement of AVP-evoked (1nmol/L) (a) or OAG-evoked (10 �mol/L) (b)TRPC6-like currents. B and C, Averagedfold increase of TRPC6-like currents byHTS (�75 mOsm) (B) or shear force (C).The currents were activated by either AVP(1 nmol/L), Norad (100 nmol/L), GTP�S(10 �mol/L), or OAG (10 �mol/L) in theabsence and presence of 10 �mol/LAACOCF3 or HET0016. D, Virtually noeffects of shear force on AVP-evokedTRPC6 current after treatment with siRNAfor cytosolic PLA2 (si-cPLA2) (a) in A7r5myocytes. D, b, Averaged fold increaseinduced by shear force with negative con-trol (si-con) and si-cPLA2. Inset in b dis-plays immunoblots for PLA2 in A7r5 myo-cytes after siRNA.
1406 Circulation Research June 19, 2009
by guest on March 6, 2016http://circres.ahajournals.org/Downloaded from

effects of 20-HETE, albeit present, would only marginallycontribute to mechanical potentiation of TRPC6 channelobserved in the present study. However, it is possible that thepreceding activation of TRPC6 via a DAG-dependent mech-anism may enhance the activating effect of 20-HETE or visaversa. This deserves further investigation.
It has been reported that reduced osmolarity or shear stresscan activate a distinct TRP subfamily member TRPV4 viageneration of a PLA2/epoxygenase metabolite 5-6-epoxyeicosatrienoic acid (EET).16 EET-mediated activationof TRPV4 in endothelial cell has been implicated in shear-induced vasodilation.5,27 Interestingly, a recent investigationrevealed that the responsiveness of TRPV4 to hypoosmolaritywas greatly enhanced (or “sensitized”) by concomitant acti-vation of P2Y receptor by ATP. This sensitization could beassigned to physical interaction between TRPV4 channel andactivated IP3 receptor via PLC-coupled receptor stimula-tion.28 Although the role of PLA2 is more primary inactivating TRPV4 than TRPC6, the above finding raises theidea that the synergism of PLC and PLA2 may be a commonbiological strategy to amplify an otherwise marginal Ca2�
mobilization caused by either neurohormonal or mechanicalstimulus alone.
In VSMCs, 20-HETE is thought to be the major AAproduct generated by cytochrome P450 CYP4A enzymesexhibiting �-hydroxylase activities, whereas vascular endo-thelial cells mainly produce EETs via CYP2C and CYP2Jenzymes.13 EETs are suggested to mediate vasodilatoryactions in both endothelium-dependent (see above) and-independent manners in part via TRPV channels.6 In humanumbilical vein, 11,12-EET is also reported to facilitate the
translocation of TRPC6 protein to caveolin-1–rich areas ofthe endothelial membrane, thereby prolonging membranehyperpolarization.29 In contrast, 20-HETE has been shown toact as a potent vasoconstrictor independently of endothelialfunction and presumably involved in the release of vasoactiveneuropeptides from sensory nerve terminals via TRPV1activation.30 Part of these effects has been ascribed toincreased voltage-dependent Ca2� influx attributable to theinhibition of a large conductance Ca2�-dependent K� chan-nel (which leads to membrane depolarization) and potentia-tion of L-type voltage-dependent Ca2� channel in VSMCs.13
20-HETE is also reported to activate protein kinase C,thereby increasing the Ca2� sensitivity of contractile machin-ery.13 Our present findings add a new important vasculartarget of 20-HETE to this repertoire, ie, TRPC6, which isexpected to serve as “depolarization” channels enhancingCa2� influx in direct and indirect (via secondary activation ofvoltage-dependent Ca2� channel or Na�/Ca2� exchanger)manners and hence increasing the vascular tone. Importantly,20-HETE has been implicated in the pathogenesis of hyper-tension in rats.31 A similar correlation with increased bloodpressure has also been suggested for the increased expressionof TRPC3, the closest homolog of TRPC6, in human essentialhypertensive patients.32 Furthermore, in the early stage ofhuman essential hypertension, hyperactivity of sympatheticnervous system and excessive activation of renin–angioten-sin–aldosterone system, both of which are expected to po-tently activate vascular TRPC6 channels, are thought to playkey roles.33 It may thus be worthy to investigate how thesemechanisms would mutually interact and contribute to thepathological dysregulation of blood pressure, particularly
Figure 7. Negative pressure greatlyenhances single AVP-induced TRPC6-likechannel activity. VH: �50 mV. Cell-attached mode. A, Enhancement of AVP-induced single channel activities by nega-tive intrapipette pressures of �20 (a) and�60 mm Hg (b) and their typical current–voltage relationship (leak-subtracted) eval-uated by a slow ramp (c). B, No channelactivity was induced by a negative pres-sure (�60 mm Hg) without agonist. C,Averaged enhancement of AVP-induced(3 nmol/L) single channel activity (NPoi) bynegative pressures in the absence orpresence of 10 �mol/L HET0016. D, Foldincrease calculated as the ratio of NPoivalues after to before application of nega-tive pressures in the absence or presenceof 10 �mol/L HET0016. HET0016 waspretreated for 10 minutes.
Inoue et al Chemical–Mechanical Synergy on TRPC6 Activation 1407
by guest on March 6, 2016http://circres.ahajournals.org/Downloaded from

with regard to the novel mechanism disclosed in this study,ie, the synergism of receptor and mechanical stimulations inTRPC6-mediated Ca2� mobilization.
Sources of FundingThis work is supported in part by grants to R.I. and L.J.J. from theJapan Society for Promotion of Sciences, the Tokyo BiochemicalResearch Foundation, the Vehicle Racing Commemorative Founda-tion, the Novo Nordisk Foundation, the Lundbeck Foundation, theDanish Heart Foundation, and the Danish Medical Research Council.
DisclosuresNone.
References1. Berridge MJ, Bootman MD, Roderick HL. Calcium signalling: dynamics,
homeostasis and remodelling. Nat Rev Mol Cell Biol. 2003;4:517–529.2. Hamill OP, Martinac B. Molecular basis of mechanotransduction in living
cells. Physiol Rev. 2001;81:685–740.3. Karaki H, Ozaki H, Hori M, Mitsui-Saito M, Amano K, Harada K,
Miyamoto S, Nakazawa H, Won KJ, Sato K. Calcium movements, dis-
Figure 8. Enhanced myogenic response of pressurized mesenteric artery by receptor stimulation. A, Expression profile of TRPC iso-forms detected by RT-PCR in single rat mesenteric arterial myocytes. B, Typical myogenic response of cannulated mesenteric arteryunder a pressure jump from 60 to 100 mm Hg in the presence of 45 nmol/L phenylephrine (Phe) and 1 nmol/L neuropeptide Y (NPY).Combined application of Norad and NPY is reported to stabilize the mesenteric arterial tone.21 HET0016 (10 �mol/L, 10 minutes) abol-ished the sustained phase of pressure-induced diameter decrease. C, Pressure–vascular tone curve in the absence (Œ) or presence of1 �mol/L phenylephrine without (F) or with (E) 10 �mol/L HET0016 pretreatment (10 minutes). The degree of vasoconstriction (vasculartone) is expressed as the difference between active and passive diameter in percentage of the diameter in Ca2�-free solution plusEGTA. Absolute changes in the outer diameter (�m) are displayed in Online Figure VI. D and E, Summary of relative change in the arte-rial outer diameter with 20 to 200 nmol/L phenylephrine/Norad plus 1 to 2 nmol/L NPY after pretreatment with 10 �mol/L HET0016 (10minutes) (D) or in the presence of 2 �mol/L Gd3� or 5 �mol/L SKF96365 (E). For D, the absolute values of outer diameter at60/100 mm Hg were 295�8/288�9, 295�8/305�9, and 347�10/363�9 �m for control, 10 �mol/L HET0016, and Ca2�-free externalsolution, respectively. F, Concentration–inhibition curves for K�-induced (75 mmol/L) diameter decrease of cannulated mesentericartery with Gd3� and SKF96365.
1408 Circulation Research June 19, 2009
by guest on March 6, 2016http://circres.ahajournals.org/Downloaded from

tribution, and functions in smooth muscle. Pharmacol Rev. 1997;49:157–230.
4. Davis MJ, Hill MA. Signaling mechanisms underlying the vascularmyogenic response. Physiol Rev. 1999;79:387–423.
5. Busse R, Fleming I. Regulation of endothelium-derived vasoactiveautacoid production by hemodynamic forces. Trends Pharmacol Sci.2003;24:24–29.
6. Inoue R, Jensen LJ, Shi J, Morita H, Nishida M, Honda A, Ito Y.Transient receptor potential channels in cardiovascular function anddisease. Circ Res. 2006;99:119–131.
7. Welsh DG, Morielli AD, Nelson MT, Brayden JE. Transient receptorpotential channels regulate myogenic tone of resistance arteries. Circ Res.2002;90:248–250.
8. Osol G, Laher I, Kelley M. Myogenic tone is coupled to phospholipase Cand G protein activation in small cerebral arteries. Am J Physiol. 1993;265:H415–H420.
9. Spassova MA, Hewavitharana T, Xu W, Soboloff J, Gill DL. A commonmechanism underlies stretch activation and receptor activation of TRPC6channels. Proc Natl Acad Sci U S A. 2006;103:16586–16591.
10. Gottlieb P, Folgering J, Maroto R, Raso A, Wood TG, Kurosky A,Bowman C, Bichet D, Patel A, Sachs F, Martinac B, Hamill OP, HonoreE. Revisiting TRPC1 and TRPC6 mechanosensitivity. Pflugers Arch.2008;455:1097–1103.
11. Yasuda N, Akazawa H, Qin Y, Zou Y, Komuro I. A novel mechanism ofmechanical stress-induced angiotensin II type 1-receptor activationwithout the involvement of angiotensin II. Naunyn Schmiedebergs ArchPharmacol. 2008;377:393–399.
12. Mederos y, Schnitzler M, Storch U, Meibers S, Nurwakagari P, Breit A,Essin K, Gollasch M, Gudermann T. Gq-coupled receptors as mech-anosensors mediating myogenic vasoconstriction. EMBO J. 2008;27:3092–3103.
13. Roman RJ. P-450 metabolites of arachidonic acid in the control ofcardiovascular function. Physiol Rev. 2002;82:131–185.
14. Shi J, Mori E, Mori Y, Mori M, Li J, Ito Y, Inoue R. Multiple regulationby calcium of murine homologues of transient receptor potential proteinsTRPC6 and TRPC7 expressed in HEK293 cells. J Physiol. 2004;561:415–432.
15. Itonaga Y, Nakajima T, Morita H, Hanano T, Miyauchi Y, Ito Y, InoueR. Contribution of nifedipine-insensitive voltage-dependent Ca2� channelto diameter regulation in rabbit mesenteric artery. Life Sci. 2002;72:487–500.
16. Watanabe H, Vriens J, Prenen J, Droogmans G, Voets T, Nilius B.Anandamide and arachidonic acid use epoxyeicosatrienoic acids to acti-vate TRPV4 channels. Nature. 2003;424:434–438.
17. Jung S, Strotmann R, Schultz G, Plant TD. TRPC6 is a candidate channelinvolved in receptor-stimulated cation currents in A7r5 smooth musclecells. Am J Physiol Cell Physiol. 2002;282:C347–C359.
18. Maruyama Y, Nakanishi Y, Walsh EJ, Wilson DP, Welsh DG, Cole WC.Heteromultimeric TRPC6-TRPC7 channels contribute to argininevasopressin-induced cation current of A7r5 vascular smooth muscle cells.Circ Res. 2006;98:1520–1527.
19. Takahashi S, Lin H, Geshi N, Mori Y, Kawarabayashi Y, Takami N, MoriMX, Honda A, Inoue R. Nitric oxide-cGMP-protein kinase G pathway
negatively regulates vascular transient receptor potential channel TRPC6.J Physiol. 2008;586:4209–4223.
20. Lee HA, Baek EB, Park KS, Jung HJ, Kim JI, Kim SJ, Earm YE.Mechanosensitive nonselective cation channel facilitation byendothelin-1 is regulated by protein kinase C in arterial myocytes. Car-diovasc Res. 2007;76:224–235.
21. Chlopicki S, Nilsson H, Mulvany MJ. Initial and sustained phases ofmyogenic response of rat mesenteric small arteries. Am J Physiol HeartCirc Physiol. 2001;281:H2176–H2183.
22. Inoue R, Okada T, Onoue H, Hara Y, Shimizu S, Naitoh S, Ito Y, MoriY. The transient receptor potential protein homologue TRP6 is theessential component of vascular �1-adrenoceptor-activated Ca2�-permeable cation channel. Circ Res. 2001;88:325–332.
23. Perozo E, Kloda A, Cortes DM, Martinac B. Physical principlesunderlying the transduction of bilayer deformation forces during mech-anosensitive channel gating. Nat Struct Biol. 2002;9:696–703.
24. Suchyna TM, Tape SE, Koeppe RE Jr, Andersen OS, Sachs F, GottliebPA. Bilayer-dependent inhibition of mechanosensitive channels by neu-roactive peptide enantiomers. Nature. 2004;430:235–240.
25. Costa T, Herz A. Antagonists with negative intrinsic activity at deltaopioid receptors coupled to GTP-binding proteins. Proc Natl Acad SciU S A. 1989;86:7321–7325.
26. Basora N, Boulay G, Bilodeau L, Rousseau E, Payet MD. 20-hydroxy-eicosatetraenoic acid (20-HETE) activates mouse TRPC6 channelsexpressed in HEK293 cells. J Biol Chem. 2003;278:31709–31716.
27. Hartmannsgruber V, Heyken WT, Kacik M, Kaistha A, Grgic I,Harteneck C, Liedtke W, Hoyer J, Kohler R. Arterial response to shearstress critically depends on endothelial TRPV4 expression. PLoS ONE.2007;2:e827.
28. Fernandes J, Lorenzo IM, Andrade YN, Garcia-Elias A, Serra SA,Fernandez-Fernandez JM, Valverde MA. IP3 sensitizes TRPV4 channel tothe mechano- and osmotransducing messenger 5-6-epoxyeicosatrienoicacid. J Gen Physiol. 2008;131:i2.
29. Fleming I, Rueben A, Popp R, Fisslthaler B, Schrodt S, Sander A,Haendeler J, Falck JR, Morisseau C, Hammock BD, Busse R. Epoxyei-cosatrienoic acids regulate Trp channel dependent Ca2� signaling andhyperpolarization in endothelial cells. Arterioscler Thromb Vasc Biol.2007;27:2612–2618.
30. Scotland RS, Chauhan S, Davis C, De Felipe C, Hunt S, Kabir J, KotsonisP, Oh U, Ahluwalia A. Vanilloid receptor TRPV1, sensory C-fibers, andvascular autoregulation: a novel mechanism involved in myogenic con-striction. Circ Res. 2004;95:1027–1034.
31. Zhang F, Wang MH, Krishna UM, Falck JR, Laniado-Schwartzman M,Nasjletti A. Modulation by 20-HETE of phenylephrine-induced mes-enteric artery contraction in spontaneously hypertensive andWistar-Kyoto rats. Hypertension. 2001;38:1311–1315.
32. Liu DY, Thilo F, Scholze A, Wittstock A, Zhao ZG, Harteneck C, ZidekW, Zhu ZM, Tepel M. Increased store-operated and 1-oleoyl-2-acetyl-sn-glycerol-induced calcium influx in monocytes is mediated by transientreceptor potential canonical channels in human essential hypertension.J Hypertens. 2007;25:799–808.
33. Johnson RJ, Rodriguez-Iturbe B, Kang DH, Feig DI, Herrera-Acosta J. Aunifying pathway for essential hypertension. Am J Hypertens. 2005;18431–440.
Inoue et al Chemical–Mechanical Synergy on TRPC6 Activation 1409
by guest on March 6, 2016http://circres.ahajournals.org/Downloaded from

and Yushi ItoMax Salomonsson, Hiromitsu Morita, Yasuhiro Kawarabayashi, Masayuki Mori, Yasuo Mori
Ryuji Inoue, Lars J. Jensen, Zhong Jian, Juan Shi, Lin Hai, Andrew I. Lurie, Freja H. Henriksen,-Hydroxylase/20-HETE Pathways
ω/2Stimulation via Phospholipase C/Diacylglycerol and Phospholipase ASynergistic Activation of Vascular TRPC6 Channel by Receptor and Mechanical
Print ISSN: 0009-7330. Online ISSN: 1524-4571 Copyright © 2009 American Heart Association, Inc. All rights reserved.is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation Research
doi: 10.1161/CIRCRESAHA.108.1932272009;104:1399-1409; originally published online May 14, 2009;Circ Res.
http://circres.ahajournals.org/content/104/12/1399World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circres.ahajournals.org/content/suppl/2009/05/14/CIRCRESAHA.108.193227.DC1.htmlData Supplement (unedited) at:
http://circres.ahajournals.org//subscriptions/
is online at: Circulation Research Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer about this process is available in the
located, click Request Permissions in the middle column of the Web page under Services. Further informationEditorial Office. Once the online version of the published article for which permission is being requested is
can be obtained via RightsLink, a service of the Copyright Clearance Center, not theCirculation Researchin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on March 6, 2016http://circres.ahajournals.org/Downloaded from

CCh 100mol/LDIDS 300mol/L ruthenium red 10mol/L
-50mV
10pA/pF
100s
0
10
1min
4min
1min +DIDS 300mol/L
Curr
en
t d
en
sity (-
50
mV
; p
A/p
F)
-25 -50 -75 -100mOsm
*
*
*
**Na
+-free
-100mOsm
5pA/pF
100s
BA
C
Online Fig.I
TRPC6-HEK
TRPC6-HEK
TRPC6-HEK

VH=-50mV
CCh100mol/L
GsMTx-4 5mol/L
10pA/pF
100s
BA
Online Fig.II
TRPC6-HEK 0
0.2
0.4
0.6
0.8
1
1.2
Rela
tive c
urr
ent (
3m
in late
r)
GsMTx-4 5mol/L
CCh
(100mol/L)
GTPS
(100mol/L)
TRPC6

-20mmHg
1pA
10s
VH=-50mV
C/ACCh 10mol/LTEA 1mmol/L
DIDS 300mol/L
-100mmHg
10pA
5s
VH=-50mV
C/A
no agonist
Online Fig.III
BA
TRPC6-HEK
TRPC6-HEK

VH=-50mV
A7r5
Norad 100mol/L
0.5pA/pF
20s
-200
-100
0
100
200
300
400
500
-100 -50 0 50 100
Norad 100mol/L
NMDG
Cu
rre
nt
(pA
)
mp (mV)
VH=-50mV
A7r5
Norad 100mol/L
1pA/pF
20s
BA
C D
Online Fig.IV
------
200bp
400600800
1000
1500
A7r5
A7r5
A7r5

0
5
10
15
20
25
30
35
40agonist only
+GF109203X
+GsMTx-4
+U73122
Fo
ld in
cre
ase
Norad 0.1mol/L AVP 1nmol/L
shear
VH=-50mV
A7r5
shear force
AVP1nM + GsMTx-4 5mol/L
0.5pA/pF
20s
Online Fig.V
BA
A7r5

200
250
300
350
400
450
500
0 20 40 60 80 100
normal+PheCa-free
dia
me
ter
(m
)
Intraluminal pressure (mmHg)
rat mesenteric artery (2nd
or 3rd
)
HET0016 (10M)-pretreatedHET0016 (10µmol/L)-pretreated
1µmol/L
200
250
300
350
400
450
500
0 20 40 60 80 100
normal+PheCa-free
dia
me
ter
(m
)
Intraluminal pressure (mmHg)
rat mesenteric artery (2nd
or 3rd
)
untreated
1µmol/L
Online Fig.VI
BA

VH=-50mV
CCh100mol/Lshear force
20pA/pF
50s
Online Fig.VII
0
10
20
30
40
50
spont
+shear
Curr
ent density (
-50m
V; pA
/pF
)
low high
TRPC6
**
NS**
0
2
4
6
8
10
12
14spont
(+) -75mOsm
Curr
ent density (
-50m
V; pA
/pF
)
low high
NS
*
TRPC6
*
TRPC6
5pA/pF
100s
VH=-50mV
Na+-free-75mOsm
BA
DC
TRPC6-HEK
TRPC6-HEK

CIRCRESAHA/2008/193227/R2
Supplement Material
Online Fig.I Prolonged application of HTS induces a non-cationic current in
HEK293 cells.
A; a representative trace of HTS (-100mOsm)-induced inward current recorded at
-50mV, and its mean current density 1 and 4 min after exposure to -25, -50, -75 and
-100mOsm HTS. A rightmost filled column indicates the almost complete inhibition of
HTS-induced current (1min exposure) by pretreatment with 300µmol/L DIDS for
2-5min. Columns and vertical bars indicate the mean ± sem (n=5-9). C;
Receptor-activated TRPC6 current is insensitive to 300µmol/L DIDS but effectively
blocked by 10µmol/L ruthenium red.
Online Fig.II GsMTx-4 is unable to inhibit CCh- or GTPγS-induced TRPC6 current.
Recorded at -50mV. A; a typical trace showing little effect of 5µmol/Lol/L GsMTx-4 on
CCh- -induced TRPC6 current (ITRPC6). B; summary of the effect of 5µmol/L GsMTx-4
on CCh (100µmol/L)- or GTPγS (100µmol/L)-induced ITRPC6. Relative current is
calculated as the ratio of amplitudes 3min after to just before application of 5µmol/L
GsMTx-4. n=4.
Online Fig.III Outlasting effects of negative pressure on receptor-activated TRPC6
channel activity (A) and development of large irregular current by strong negative
pressure (B). Representative traces in cell-attached recording.
Online Fig.IV siRNA knockdown of TRPC6 protein abolishes Norad-induced
cationic currents in A7r5 myocytes.
A & B; an actual trace for Norad (100µmol/L)-induced cationic current (A) and its
current voltage relationship in Na+-rich and -deficient (NMDG) conditions (B). C;
mRNA expression profile of TRPC isoforms in A7r5 myocytes evaluated by RT-PCR.
D; two stealth siRNAs greatly suppressed Norad (100µmol/L)-induced TRPC6-like
cationic current with reduced expression of TRPC6 protein (inset: evaluated by
immunoblot analysis with a TRPC6-specific antibody) and. Representative of three
independent experiments.
Online Fig.V GsMTx-4 is unable to suppress shear-induced potentiation of 1nM
AVP-evoked TRPC6-like current in A7r5 myocytes.
A; shear force potently enhanced a cationic current evoked by 1nM AVP. B; summary of

- 2 -
the enhancing effects of shear force on Norad (0.1µmol/L) and AVP (1nmol/L)-evoked
cationic currents in the presence of GsMTx-4 (5µmol/L) or a PKC inhibitor
GF109203X (GF; 2µmol/L). The definition of fold increase is the same as in Fig.1. No
statistically significant difference is present among four different conditions for
shear-induced increase in Norad- and AVP-evoked cation currents. n=5-9.
Online Fig.VI 20-HETE mediates the enhancement of myogenic response by
phenylephrine in rat mesenteric artery.
A; relationships between the absolute diameter change (µmol/L) of cannulated rat
mesenteric artery (2nd
or 3rd
branch) and intraluminal pressure (mmHg) in the absence
or presence of 1µmol/L Phe or in Ca2+
-free solution (n=8). B; the same relationships
as in A in the presence of 10µmol/L HET0016 (n=8).
Online Fig.VII Spontaneously active ITRPC6 is potentiated by mechanical stimulation.
Over 200 whole-cell recordings, only 11 TRPC6-expressing HEK cells displayed direct
mechanosensitivity. A; application of HTS (-75mOsm) itself caused a slow development
of basal ITRPC6 showing a high spontaneous activity. B; summary of the effects of HTS
on spontaneously active ITRPC6. n=5 and 16 for low and high spontaneous activities,
respectively. C; representative trace of shear force-induced potentiation of
spontaneously active ITRPC6. D; summary of the effects of shear force on spontaneously
active ITRPC6. n=6 and 12 for low and high spontaneous activities, respectively.
Materials and Methods
Cell culture and transfection
Human embryonic cell line HEK293 and embryonic aortic smooth muscle cell line
A7r5 (purchased from ATCC; Manassas, VA, USA) were maintained in Dulbecco’s
modified culture medium (DMEM) containing 10% fetal bovine serum and antibiotics
(100 units/ml penicillin, streptomycin; GIBCO), under 5% CO2 with every 3-4 day
passage with 0.0025% and 0.025% trypsin/EDTA (Invitrogen, USA). Great care was
taken not to overdigest or damage the cells with trituration, which otherwise often
impaired their mechanosensitivity.
Mouse TRPC6 DNA (Gene Accession No.: NM_013838) subcloned into the pCI-neo
vector (Invitrogen, USA) was transfected together with pCI-neo-H3-CD8 (cDNA of
the T-cell antigen CD8) at the ratio of 5~10:1, to HEK cells of 50-80% confluency, with
the aid of a transfection reagent SuperFectTM
(Qiagen, Germany). CD8 was used as the

- 3 -
expression marker. Functional evaluation was made 48-72h after transfection, at the
time point when the expression of TRPC6 proteins reached the peak. TRPC6-expressing
cells were selected by using beads-conjugated anti-CD8 antibody (Dynabeads M-450
CD8, Dynal Biotech) as the expression marker15
.
To knockdown the expression of TRPC6 protein, 40-50 pmole/ml stealthTM
/siRNA
duplex (sense/antisense: 5’ to 3’); AAACCACCGUUGCAUAAAGACC/
GGUCUUUAUGCAAUCGCGGUGGUUU was transfected to 30-50% confluent A7r5
myocytes (in the presence of 2.5µl/ml lipofectamine TM
2000) according to the
manufacturer’s instruction (Invitrogen, USA). After 48-72h culture, almost no
expression of TRPC6 protein was detectable by immunoblotting. For siRNA
knockdown of cytosolic PLA2 in HEK293 (NM_024420) and A7r5 cells (NM_133551),
100pmole/ml of the following stealth TM
/siRNA duplexes (Invitrogen, USA) were
respectively used (sense/antisense: 5’ to 3’);
AUUUCGUAUGGACUAAAUUCAACCC/
GGGUUGAAUUUAGUCCAUACGAAAU, and
AUCCGAUGGACCCAACUUGCUUGGU/
ACCAAGCAAGUUGGGUCCAUCGGAU. The efficiency of the siRNA was
confirmed by immunoblotting with anti-PLA2 antibody (4-4B-3C; Santa Cruz).
Electrophysiology
Patch electrodes (3-5M) fabricated from 1.5mm boroscilicate glass capillaries and
heat-polished were used for whole-cell and single channel recordings, in conjunction
with a high-impedance low-noise patch clamp amplifier (EPC9; HEKA Electronics,
Germany) and an A/D, D/A-converter (Digidata 1200; Axon Instruments). Sampled data
low-passed filtered at 1kHz and digitized at 5kHz were analyzed by using ‘Clampfit’
v.9.2 (Axon Instruments, USA). Longer time-frame recordings (e.g. whole-cell current
traces in Fig. 1) were performed with PowerLab/400 (AD Instruments, Australia;
sampling rate: 100Hz), and analyzed with the software Chart v3.6. The magnitude of
recorded noisy currents was defined as their 5-10s averages over the time period of
concern.
To minimize variations arising from different cell size, the magnitude of current is
normalized by dividing by cell capacitance. Mean cell capacitance and resistance
(measured between -100 and -50mV with Cs-internal solution) were 28.0±2.1pF and
7.8±1.4 GΩfor HEK293 cells (n=44), and 99.9±8.5pF and 3.8±0.3GΩ (n=43) for
A7r5 myocytes, respectively.
For single channel recordings [cell-attached (C/A) and inside-out (I/O) patch

- 4 -
configurations], sampled data were low-pass filtered at 1kHz and stored on a computer
hard disc after digitization at 5kHz. Single channel analysis was made using the
software ‘Clampfit’ v.9.2 (Axon Instruments, USA). The mean single channel current
(NPoi) was calculated for each experiment just before and after each procedure by
averaging the current amplitude over 5s with respect to the baseline as performed
previously1.
Shear force (~10dyn∙cm-2
) was applied as a continuous laminar flow (~1µl∙s-1
) out of
a Y-tube, which was initially placed at a distance of ~ 500µmol/Lol/L and quickly
moved within 20µmol/Lol/L from a voltage-clamped cell, and approximated using the
equation: τ=4µQ/πr3; τ, µ and r denote shear force, the viscosity of solution
(0.00797g∙cm-1
∙s-1
), and the inner radius of the tube (~100µmol/Lol/L), respectively.
Bathing solution was constantly perfused at a rate of 0.8 - 1ml/min into a recording
chamber, which would generate a shear force of ~0.5dyn∙cm-2
according to the equation:
τ=3µQ/wh2 where w and h denote the width (10mm) and height of chamber flow (2mm),
respectively.
Negative pressure was applied manually by suction via patch pipette, the magnitude
of which was scaled and monitored by using a DMP-1B pneumatic transducer tester
(BIO-TEK instrument, VT, USA).
All experiments were performed at room temperature (22-26˚C).
Video microscopic diameter measurement
Male Sprague Dawley rats weighing 200-300g (Charles River, Japan or Taconic,
Denmark) were exsanguinated under anesthesia with intra-peritoneal injection of
pentobarbital. After opening the abdominal cavity, arterial segments of 2nd
or 3rd
branches of mesenteric artery (300 –400µmol/L in the outer diameter recorded at 60
mm Hg) were excised, cleaned of attached fat and connective tissues, and cannulated at
both ends with fine-tipped glass capillaries and fixed tightly with nylon threads, as
performed previously2. The lumen of the segments was perfused with Krebs’ solution
(to which 10 mg BSA per ml had been added) under no-flow conditions, and
intraluminal pressure was controlled using a pressure myograph system (Living
Systems Instrumentation, Burlington, USA; or, Danish Myotechnology, Aarhus,
Denmark). The experimental chamber (model CH/1, Living Systems Instrumentation;
or DMT 120CP, Danish Myotechnology) accommodating an arterial segment was
placed on a thermo-plate (Tokai Hit, Japan) and continuously superfused at a rate of
10ml/min with Krebs solution aerated with 95%O2 and 5%CO2 (T=35-37°C; pH
7.40-7.50, both monitored in the experimental chamber), to which tested drugs were

- 5 -
added. The video images of arterial segments captured through a CCD camera under
binocular microscopy (Stemi 200-C, Zeiss, Germany; or Olympus IX71, Japan) were
measured for their diameter change every 100ms using a width analyzer (C3161,
Hamamatsu Photonics; or Myoview, Danish Myotechnology). The results were stored
on a computer hard disc after digitization (MacLab, AD Instruments, South Wales,
Australia).
We tried to remove the endothelium completely with a wire/thread/hair inserted into the
lumen of the blood vessel which was followed by the passage of air bubbles. However,
the following confirmation by acetylcholine suggested that, in more than half of trials,
albeit greatly reduced compared with control, slight vasorelaxation could still be
induced. Thus, we cannot completely exclude the possible contamination of the
contribution of endothelial cells in our myograph experiments.
The details of animal experiments were reviewed and approved by local animal
ethics committees of Fukuoka University, Kyushu University and University of
Copenhagen in advance, and all procedures used to sacrifice rats obeyed the guidelines
set by the committees.
Immunoblot
Total lysate of A7r5 myocyes or HEK293 cells (about 106 cells) was prepared in
sample buffer, and its protein concentration was determined by using the BCA protein
assay kit (Pierce). 5% (v/v) 2-mercaptoethanol and 1% (w/v) bromophenol blue were
added to the sample, and proteins were separated (30-40μg) on 10% (w/v) SDS-PAGE
and blotted to a polyvinylidene diflouride (PVDF) membrane. After blocking with 5%
(w/v) skim milk dissolved in Tween-PBS, the membrane was incubated with the
anti-TRPC6 antibody (Alomone; see the above) (at 1:100-200 dilution) overnight.
TRPC6 protein (~120kD) or PLA2 protein (~100kD) was visualized by
chemiluminescence by incubating the PVDF membrane with the secondary antibody
conjugated with horseradish peroxidase, and analyzed densitometrically.
RT-PCR
Single myocytes were dissociated from rat mesenteric arteries (2nd
or 3rd
branches) by
enzymatic treatment [incubated in 2mg/ml collagenase (type I, Worthington, Lakewood,
NJ, USA)-containing nominally Ca2+
-free external solution for 1h] and mechanical
trituration. Total RNA was extracted from single mesenteric arterial myocytes and A7r5
cells using a commercial kit (RNAeasy, Qiagen) according to the manufacturer’s
instruction, and then subjected to reverse transcription polymerase reaction using a

- 6 -
protocol; preheating at 94°C for 1min followed by 40 cycles (denaturation at 94°C for
10 seconds: annealing at 58-65°C for 30 seconds: extension at 72°C for 1 minute) and
final extension at 72°C for 10 minutes. Annealing temperatures for each primer pair
were determined using a gradient protocol.
Primer pairs used for respective rat TRP isoforms are as follows (forward/reverse: 5’ to
3’);
TRPC1(DQ839447.1: nt. 236-710): GCG TAG ATG TGC TTG GGA GAA A /GCT
CTC AGA ATT GGA TCC TCC TCT;
TRPC2 (AF136401.1: nt. 197-713) : CCA GGT GGT CCT CTG CGG AA/CAT CCT
CAC TGG CCA GCG AGA;
TRPC3 (AB022331.1: nt. 474-948): GCTGGCCAAGCTGGCCAA/
GAACACAAGCAGACCCAGGAAGA;
TRPC4 (AF421368.1: nt. 1151-1722): CCT CTC AGC ACA TCG ACA GGT /CCA
AAT ATT GAC CAA AAC AGG GA;
TRPC5 (EF672039.1: nt. 987-1511): CAA GCT TCT AAC CTG CAT GAC CA/ CCT
AAG TGG GAG TTG GCT GTG AA;
TRPC6 (A: nt. 327-1398): CAT CCC AGT GGT GCG GAA GA/ GCC TTC AAA TCT
GTC AGC TGC A;
TRPC7(XM225159.4: nt. 1907-2151): TCAACCTGTACTCCTACTACC/
GATTTCCTGATAGGAGTTGTTG.
Solutions
Pipette solution for whole-cell recording (in mM): 120 CsOH, 120 aspartate, 20 CsCl,
2 MgCl2, 10 BAPTA/4 CaCl2 (or 5 EGTA/1.5 CaCl2), 10 HEPES, 2 ATP, 0.1 GTP, 10
glucose (adjusted to pH7.2 with Tris base); External solution (in mmol/L): 140 NaCl, 5
KCl, 1 CaCl2, 1.2 MgCl2, 10 HEPES, 10 glucose (pH7.4, adjusted with Tris base).
Hypotonic solutions were made to give a desired hypotonicity (-25, -50, -75 and
-100mOsm) by reducing the external NaCl concentration, to which equi-osmolar
sucrose or mannitol was added to make the corresponding normotonic solutions. Na+
free external solution was made by equimolar substitution of monovalent cations with
N-methyl,D-glucamine (NMDG).
Pipette solution for cell-attached (C/A) and inside-out (I/O) recordings (in mmol/L):
140 NaCl, 1 tetraethylammonium-Cl, 0.3 DIDS, 1.2 MgCl2, 1 CaCl2, 10 HEPES, 10
glucose (pH 7.4, adjusted with Tris base).
Bathing solution for C/A recording (in mmol/L): 145 KCl, 2 MgCl2, 1EGTA, 10
HEPES. Bathing solution for I/O recording (in mmol/L): 120 CsOH, 120 aspartate, 20

- 7 -
CsCl, 2 MgSO4, 2 EGTA, 0.4 Ca, 10 HEPES, 2 ATP, 0.1 GTP (pH 7.2, adjusted with
Tris base).
Drugs were topically applied using a fast solution exchange device ‘Y-tube’.
Chemicals
CCh, noradrenaline, GTPγS, indomethacin, p-bromophenacyl bromide (pBPB),
miconazol, 17-octadecynoic acid (17-ODYA), and disodium
4,4'-diisothiocyanatostilbene-2,2'-disulfonate (DIDS) were purchased from
Sigma-Aldrich (USA), and AVP ([Arg8]-vasopressin), AACOCF3, nordihydroguaiaretic
acid (NDGA), U73122, GF109203X from Calbiochem-Merck (USA), and OAG,
20-HETE and 2,4,6-trinitrophenol (TNP) from Cayman (UK), and GsMTx-4 from
Peptides International. HET0016 was kindly provided by Taisho Pharmaceuticals
(Japan).
Stock solutions for drugs were made by dissolving their powders in DMSO and used
a more than 1000-fold dilution, the concentration of which did not affect the magnitude
or kinetics of TRPC6 currents significantly.
Statistics
All data are expressed as means S.E.M (indicated by columns/symbols and bars in
each figure). Two-tailed paired and unpaired Student’s t-tests were used to evaluate
statistical significance with a criterion of P <0.05 (*) or P<0.01 (**). For multiple
comparison, statistical significance was evaluated by ANOVA followed by Bonferroni’s
t-test. ‘NS’ in figures: no statistically significant difference.
References
1. Shi J, Mori E, Mori Y, Mori M, Li J, Ito Y, Inoue R. Multiple regulation by
calcium of murine homologues of transient receptor potential proteins TRPC6 and
TRPC7 expressed in HEK293 cells. J Physiol. 2004;561:415-432.
2. Itonaga Y, Nakajima T, Morita H, Hanano T, Miyauchi Y, Ito Y, Inoue R.
Contribution of nifedipine-insensitive voltage-dependent Ca2+
channel to diameter
regulation in rabbit mesenteric artery. Life Sci. 2002;72:487-500.