substrate determines asymmetrical gait dynamics in marmosets (callithrix jacchus) and squirrel...
TRANSCRIPT

Substrate Determines Asymmetrical Gait Dynamicsin Marmosets (Callithrix jacchus) and SquirrelMonkeys (Saimiri boliviensis)
Jesse W. Young*
Department of Anthropology, University of Texas at Austin, Austin, TX 78712
KEY WORDS locomotion; kinematics; kinetics; ontogeny; arboreal stability
ABSTRACT Studies of skeletal pathology indicatethat injury from falling accounts for most long bonetrauma in free-ranging primates, suggesting that prima-tes should be under strong selection to manifest morpho-logical and behavioral mechanisms that increase stabil-ity on arboreal substrates. Although previous studieshave identified several kinematic and kinetic features ofprimate symmetrical gaits that serve to increase arbo-real stability, very little work has focused on the dynam-ics of primate asymmetrical gaits. Nevertheless, asym-metrical gaits typify the rapid locomotion of most prima-tes, particularly in smaller bodied taxa. This studyinvestigated asymmetrical gait dynamics in growingmarmosets and squirrel monkeys moving on terrestrialand simulated arboreal supports (i.e., an elevated pole).Results showed that monkeys used several kinematicand kinetic adjustments to increase stability on thepole, including reducing peak vertical forces, limiting
center of mass movements, increasing substrate contactdurations, and using shorter and more frequent strides(thus limiting disruptive whole-body aerial phases).Marmosets generally showed greater adjustment to polelocomotion than did squirrel monkeys, perhaps as aresult of their reduced grasping abilities and retreatfrom the fine-branch niche. Ontogenetic increasesin body size had relatively little independent influenceon asymmetrical gait dynamics during pole locomotion,despite biomechanical theory suggesting that arborealinstability is exacerbated as body size increases relativeto substrate diameter. Overall, this study shows that 1)symmetrical gaits are not the only stable way totravel arboreally and 2) small-bodied primates utilizespecific kinematic and kinetic adjustments to increasestability when using asymmetrical gaits on arbo-real substrates. Am J Phys Anthropol 000:000–000,2008. VVC 2008 Wiley-Liss, Inc.
Studies of skeletal pathology indicate that injury fromfalling may account for the greatest incidence of longbone trauma in free-ranging primate populations(Schultz, 1944; Buikstra, 1975; Lovell, 1991; Jurmain,1997; Carter et al., 2008). Such studies suggest that pri-mates should be under strong selective pressure to mani-fest morphological and behavioral mechanisms thatincrease stability on arboreal substrates. In general, ar-boreal stability is maximized by 1) maintaining tight ad-hesion with the substrate, 2) minimizing the forcesimparted to the substrate, and 3) traveling with a rela-tively flat and low trajectory of the center of mass(COM) (Napier, 1967; Cartmill, 1985; Demes et al., 1990;Stevens, 2003). Primates display an array of morphologi-cal adaptations to maintain substrate adhesion, includ-ing: long prehensile fingers, divergent first manual andpedal rays (particularly the hallux), well-developed flexorand rotator musculature in the distal limbs, and fric-tional skin on the volar surfaces of the hands and feet(Cartmill, 1972, 1979; Grand, 1977; Lemelin, 1999; Kirket al., 2008). Additionally, prosimians and most monkeyshave long tails that can be used statically as counterbal-ances or dynamically to generate stabilizing angular mo-mentum, thereby counteracting disruptive pitching androlling moments (Larson and Stern, 2006). When walk-ing, primates also use a variety of behavioral mecha-nisms to reduce substrate forces and limit COM move-ments, including greater joint flexion, augmented sub-strate contact durations (i.e., duty factors), increasedlimb angular excursions, increased limb joint yield overa step, and longer and less frequent strides (Napier,
1967; Rose, 1973; Alexander and Maloiy, 1984; Reynolds,1987; Demes et al., 1990; Schmitt, 1999; Larson et al.,2001; Schmitt, 2003b; Stevens, 2003; Larney and Larson,2004). Collectively, these behavioral modificationsshorten the distance between the substrate and COM,thus reducing the potential for disruptive pitching androlling moments, and limiting peak forces and COM dis-placements by facilitating a more compliant gait(Schmitt, 1999).Our understanding of how primates adjust locomotor
behaviors to increase arboreal stability comes almostexclusively from studies of symmetrical gait dynamics(i.e., walking and running). In a symmetrical gait, eachforelimb is temporally paired with a hind limb and foot-falls of contralateral forelimb pairs and hind limb pairs(e.g., right and left hind limbs) are evenly spaced in time(Hildebrand, 1966; Gambaryan, 1974; Sukhanov, 1974).
Grant sponsor: L.S.B. Leakey Foundation; Grant number: 38648;Grant sponsor: Interdepartmental Doctoral Program in Anthropo-logical Sciences at Stony Brook University.
*Correspondence to: Jesse W. Young, Department of Anthropology,University of Texas at Austin, 1 University Station C3200, Austin,TX 78712, USA. E-mail: [email protected]
Received 20 May 2008; accepted 8 September 2008
DOI 10.1002/ajpa.20953Published online in Wiley InterScience
(www.interscience.wiley.com).
VVC 2008 WILEY-LISS, INC.
AMERICAN JOURNAL OF PHYSICAL ANTHROPOLOGY 000:000–000 (2008)

By contrast, during asymmetrical gaits, forelimbs arepaired together and hind limbs are paired together andthe footfalls of contralateral fore- and hind limb pairsare unevenly spaced in time (Howell, 1944; Gambaryan,1974; Hildebrand, 1977). Few studies have examined thedynamics of asymmetrical gaits in primates (but seeVilensky, 1980; Hurov, 1987; Vilensky et al., 1990;Kimura, 1992; Franz et al., 2005; Hanna et al., 2006).However, asymmetrical gaits constitute an importantelement of most primates’ locomotor repertoires, particu-larly in smaller-bodied taxa (Vilensky, 1980; Hurov,1987; Kimura, 1992; Demes et al., 1994; Arms et al.,2002; Franz et al., 2005; Schmitt et al., 2006). Asymmet-rical gaits are typically used at higher speeds and areassociated with greater substrate forces (Demes et al.,1994; Franz et al., 2005; Hanna et al., 2006), potentiallycompromising arboreal instability. Nevertheless, no pre-vious study has explicitly focused on how primates mightmodify asymmetrical gait dynamics to ensure stabilityon arboreal substrates.
SPECIFIC AIMS AND HYPOTHESES
This study focused on asymmetrical gait dynamics inontogenetic samples of common marmosets (Callithrixjacchus, L. 1758) and Bolivian (i.e., black-capped) squir-rel monkeys (Saimiri boliviensis, Geoffroy and Blainville1834) moving on terrestrial and simulated arboreal sub-strates (i.e., an elevated pole). Previous research sug-gests that asymmetrical gaits likely represent an impor-tant component of both species’ locomotor repertoire.Studies of callitrichine primates closely related to mar-mosets, such as Goeldi’s monkeys (Callimico goeldii) andtamarins (Saguinus and Leontopithecus spp.), havefound that asymmetrical gaits represent a substantialportion of the locomotor repertoire, constituting 33–50%of arboreal travel (Fleagle and Mittermeier, 1980;Garber, 1991; Rosenberger and Stafford, 1994). Addition-ally, the frequency with which asymmetrical gaits areused is inversely proportional to body size (Dagg, 1973;Hildebrand, 1977) and marmosets are among the small-est platyrrhine primates, with an average adult bodymass of 320 grams (Smith and Jungers, 1997). Similarly,although squirrel monkeys are not as small as marmo-sets, they are still among the smallest anthropoid pri-mates, with an average adult body mass of 811 g (Smithand Jungers, 1997). Fontaine (1990) found that asym-metrical gaits accounted for 16% of all quadrupedal loco-motion in semicaptive free-ranging Bolivian squirrelmonkeys.Despite belonging to the same family of platyrrhine
monkeys, the Cebidae (Horovitz et al., 1998; Opazoet al., 2006), marmosets and squirrel monkeys differ inmany aspects of their ecology, anatomy, and locomotorbehavior. Marmosets and other callitrichines are uniqueamong extant primates in having claws, rather thannails, on every digit but the hallux. Clawed digits are of-ten interpreted as an adaptation to gumnivory, enablingcallitrichines (and particularly marmosets) to grip ontolarge vertical tree trunks while feeding on tree exudates(Sussman and Kinzey, 1984; Garber, 1992; Hamrick,1998; Vinyard and Schmitt, 2004). An adaptive shiftaway from a fine-branch foraging environment, oftenunderstood to be the fundamental adaptive niche for pri-mates (Cartmill, 1972; Rasmussen, 1990; Sussman,1991; Schmitt and Lemelin, 2002), has also affectedother aspects of marmoset autopodial anatomy. Com-
pared to other small-bodied cebids, marmosets are char-acterized by relatively narrow apical pads with poorlydeveloped papillary ridges, laterally compressed terminalmanual phalanges, and a relatively short hallux markedby limited mobility and diminished intrinsic adductormusculature (Beattie, 1927; Midlo, 1934; Hamrick,1998). These derived changes in marmoset autopodialmorphology should limit stability and efficiency whenmoving and foraging in a small branch environment(Hamrick, 1998; Schmitt, 2003a). By contrast, squirrelmonkeys frequently feed and travel in a fine-branchniche (Fleagle et al., 1981; Boinski, 1989) and are char-acterized by better developed grasping morphology andabilities than callitrichines (Costello and Fragaszy, 1988;Janson and Boinski, 1992; Hamrick, 1998; but see Leme-lin and Grafton, 1998).In this study, I tested three general hypotheses. First,
I hypothesize that, marmosets and squirrel monkeysusing asymmetrical gaits on arboreal substrates shouldattempt to increase stability by decreasing vertical forcesand reducing the overall vertical displacement of theCOM. Based on previous research, I formulated andtested three predictions of how monkeys could effectthese changes. First, stance phase durations (i.e., dutyfactors) should be greater on arboreal substrates.Extending limb stance phase durations without changingvelocity necessarily reduces peak vertical force magni-tudes (McMahon et al., 1987). Second, monkeys travelingon arboreal substrates should favor limb sequence pat-terns that limit whole-body aerial phases and increasetotal contact time with the substrate during the stride.Increasing net substrate contact duration limits the ver-tical oscillation of the COM and reduces peak verticalforce by distributing impulse more evenly across thestride (Biknevicius et al., 2004; Schmitt et al., 2006).Therefore, a) the phasing between limb contacts (i.e.,lead intervals) should be greater on arboreal substrates,and b) animals should choose footfall sequences andgaits that maximize substrate contact. Third, monkeysshould use shorter and more frequent strides on arborealsubstrates. This prediction is somewhat counterintuitive,as previous research on symmetrical gaits has arguedthat primates reduce branch sway and promote arborealstability by increasing stride length and decreasingstride frequency (e.g., Demes et al., 1990). However, lon-ger strides during asymmetrical gaits are primarilyachieved with the use of extended aerial phases markedby large vertical oscillations of the COM (Dagg, 1973;Heglund et al., 1974; Gasc, 1993). Therefore, shortstrides without pronounced aerial phases shoulddecrease COM movement and increase arboreal stability.My second hypothesis was that, due to their derived
autopodial anatomy and retreat from the fine-branch for-aging niche (Sussman and Kinzey, 1984; Garber, 1992),marmosets should experience greater instability whenusing asymmetrical gaits on arboreal substrates thansquirrel monkeys. As such, kinematic and kinetic adjust-ments to arboreal locomotion should be more pronouncedamong marmosets than among squirrel monkeys. Inother words, differences between arboreal and groundlocomotion should be greater for marmosets than forsquirrel monkeys.Finally, because arboreal stability is generally compro-
mised as body size increases relative to branch diameter(Cartmill, 1985), older and larger monkeys moving onthe pole segment should experience greater challenges tostability than younger and smaller monkeys. Therefore,
2 J.W. YOUNG
American Journal of Physical Anthropology

as individuals age and grow larger, kinematic and ki-netic adjustments to locomotion on the pole shouldbecome more pronounced, whereas size-related changesduring ground locomotion should either be nonexistentor be relatively minimal.
METHODS
Animal subjects
Research took place at two National Institutes ofHealth Primate Research Centers: the SouthwestNational Primate Research Center (SNPRC, San Anto-nio, TX) and the Center for Neotropical PrimateResearch and Resources (CNPRR, Mobile, AL). Institu-tional Animal Care and Use Committees (IACUC) atStony Brook University, the SNPRC, and the CNPRRapproved all procedures prior to the beginning of thisresearch.The marmoset sample included two young juveniles
(age range: 85–139 days; body mass range: 143–201 g),one older male juvenile (age range: 258–320 days; bodymass range: 314–349 g), and two adults (ages: 2.4 and12.2 years; mean body mass: 327 g). The two young juve-nile marmosets were studied for periods of 54 and 37days, beginning at 85 and 96 days of age, respectively.All marmosets were grouped in family units of six tonine individuals and housed in cages that were large rel-ative to body size (1.52 m 3 1.42 m 3 0.62 m) andincluded a variety of flat and ‘‘arboreal’’ supports (e.g.,dried branches and PVC pipes).Five female infant-juvenile squirrel monkeys com-
prised the squirrel monkey sample. An additional squir-rel monkey participated in two experiments but had tobe withdrawn from the study due to an unrelated tailinjury. Collectively, the squirrel monkeys ranged in agefrom 74 to 302 days and body mass from 218 to 535 g(29–71% of adult size; mean: 367 g). Participation dura-tion for individual monkeys ranged from 151 to 185 days(mean: 172.2 days; median: 179 days), beginning at agesof 74–120 days (mean: 104.6 days; median: 118 days).Monkeys lived in social groups of 15–30 individuals thatwere housed in large 1.5 m 3 2.1 m 3 4.5 m enclosureswith 35–40 linear meters of perches and substrates forfree-ranging locomotor activity.The distribution of strides in the marmoset and squir-
rel monkey datasets, organized by age group, is shownin Figure 1. Ontogenetic changes in body mass are plot-
ted against age for all monkeys in Figure 2. The distri-bution of marmoset trials was slightly skewed towardthe youngest age group. However, because the oldermale juvenile marmoset was approximately the samesize as the adults (see Fig. 2), sample composition bysize range was more homogenous.Despite the inclusion of infants and juveniles, purely
maturational effects should have little impact on theresults and conclusions of this study. Compared to otherprimates, both marmosets and squirrel monkeys aremarkedly precocial in their behavioral development.Marmosets’ full locomotor repertoire is usually presentby 3 months of age. At this time, infants begin the pro-cess of weaning, body mass starts to rapidly increase,adult-like limb proportions are reached, and synapticdensity in the visual cortex peaks (Missler et al., 1992).Squirrel monkey development proceeds on a similarpace. Infants engage in independent locomotion withinthe first month, begin foraging independently after 7weeks, are weaned by 4 months, and are completely in-dependent from their mothers by 8 months (Elias, 1977;Kaack et al., 1979; Boinski and Fragaszy, 1989; Fragaszyet al., 1991). In sum, the fast pace of marmoset andsquirrel monkey locomotor development should minimizethe influence of age per se on the kinematic and kineticparameters studied here, particularly after gait parame-ters are size-adjusted.
Fig. 1. Distribution of strides in the marmosets (A) and squirrel monkeys (B), organized by age group. White bars representground strides, whereas gray bars represent pole strides.
Fig. 2. Body mass plotted against age for all marmosets andsquirrel monkeys. Marmosets are represented by open squaresand squirrel monkeys by open circles. For ease of presentation,body masses for the two adult marmosets have been plotted onbroken axes.
3ASYMMETRICAL GAIT DYNAMICS IN CALLITHRIX AND SAIMIRI
American Journal of Physical Anthropology

Data collection
Kinematic data collection. Monkeys were filmed witha high-speed digital video camera (MotionMeter 1000,Redlake MASD, San Diego, CA) operating at 125–250Hz as they traversed a 2.75 m runway. Depending on ex-perimental condition (e.g., ground vs. pole), monkeystraversed either the flat runway floor (‘‘ground’’ condi-tion) or a 2.5 cm diameter PVC pipe elevated 10.7 cmabove the surface of the runway (‘‘pole’’ condition). Bothsubstrates were coated with a mixture of polyurethaneand nonskid paint additive (Behr Process Corporation,Santa Ana, CA) to increase traction.Prior to the beginning of each squirrel monkey experi-
ment, individuals were weighed and the skin over theapproximate center of rotation of the shoulder and thehip was shaved and marked with retro-reflective tape, aprocedure that did not require the use of anesthesia.Body weight was recorded ad libitum during marmosetexperiments with the use of a scale placed at one end ofthe runway.Video files from each marmoset and squirrel monkey
experiment were imported into the MATLAB DLT Data-viewer 2 digitizing platform (Hedrick, 2007) for coding.Individual strides were identified based on cyclic touch-downs of a reference limb. Within each stride, touch-down events for each of the four limbs were recognizedas the first frame in which the limb contacted the sub-strate. Lift-off events were recognized as the first framefollowing touchdown in which the limb did not contactthe substrate.
Kinetic data collection. Two custom-built 30.5-by-30.5cm2 triaxial force platforms (Heglund, 1981; Biewenerand Full, 1992), placed in series at the center of the run-way, were used to measure vertical forces during locomo-tion. During the simulated arboreal trials, PVC seg-ments (30.5 cm long and 2.5 cm in diameter) wereattached to each force platform via bolts secured directlyto the platform frame. Voltage outputs from channelscorresponding to each force axis were routed through aNational Instruments (Austin, TX) SC-2345 chassisand recorded using a LabView virtual instrument. Crosstalk between force channels was generally low, rangingbetween 0.3 and 3.5% without the pole segment andbetween 0.3 and 10.5% when the pole segment wasattached. Force platforms were calibrated daily followingthe recommendations of Biewener and Full (1992).Following Riskin et al. (2005), kinetic and kinematic
data were synchronized via a 3.3 V square-wave pulsegenerated by the video camera and routed separately toa bank of LEDs positioned on the back wall of therunway and to the SC-2345 chassis. This circuit wasnormally interrupted by means of a handheld switch.During each trial, the switch was briefly closed, simulta-neously illuminating the LEDs and changing the shapeof the square wave in the data file. Using this procedure,it was possible to synchronize video and kinetic data to aresolution of 4–8 ms, depending on video frame rate.Force data from each trial were imported into a cus-
tom-written MATLAB program (Mathworks, Natick,MA), where the raw voltages were corrected for crosstalk and transformed into Newtons. Force traces fromeach channel were smoothed using a zero-lag fourth-order Butterworth low-pass filter with a cut-off fre-quency of 30 Hz for the marmosets and 25 Hz for thelarger squirrel monkeys. Baseline drift during andbetween trials was corrected by sampling the average
values of unloaded periods immediately prior to and fol-lowing platform contact and subtracting these valuesfrom the force traces.
Dependent variables
Speed. For each species, I calculated average locomotorspeed from the displacement of a standard referencemarker (nose for marmoset trials and shoulder/hip forsquirrel monkey trials). Digitizing noise was correctedusing a zero-lag fourth-order Butterworth low-pass filterwith a cut-off frequency of 10 Hz. This cutoff frequencywas selected as optimal using a residual analysis proce-dure described by Winter (2005). Piecewise cubic splineinterpolation was used to interpolate over gaps of miss-ing data �10 frames (i.e., 40–80 ms, depending on framerate). After transforming raw pixel coordinates intometers by using the force platforms as calibration objects(combined length of the platforms: 0.6096 m), linearleast-squares regressions of corrected displacement dataon time were used to calculate overall speed across eachstride. Trials in which the coefficient of determination(i.e., R2) of reference marker position against time wasless than 0.99 were designated unsteady and were dis-carded. To control for differences in body mass across thesample, raw speeds were converted into Froude numbersusing the formula u/(gh)0.5, where u is the speed, g isthe gravitational acceleration (9.81 m s22), and h is thecube root of body mass (Hof, 1996; Biewener, 2003).
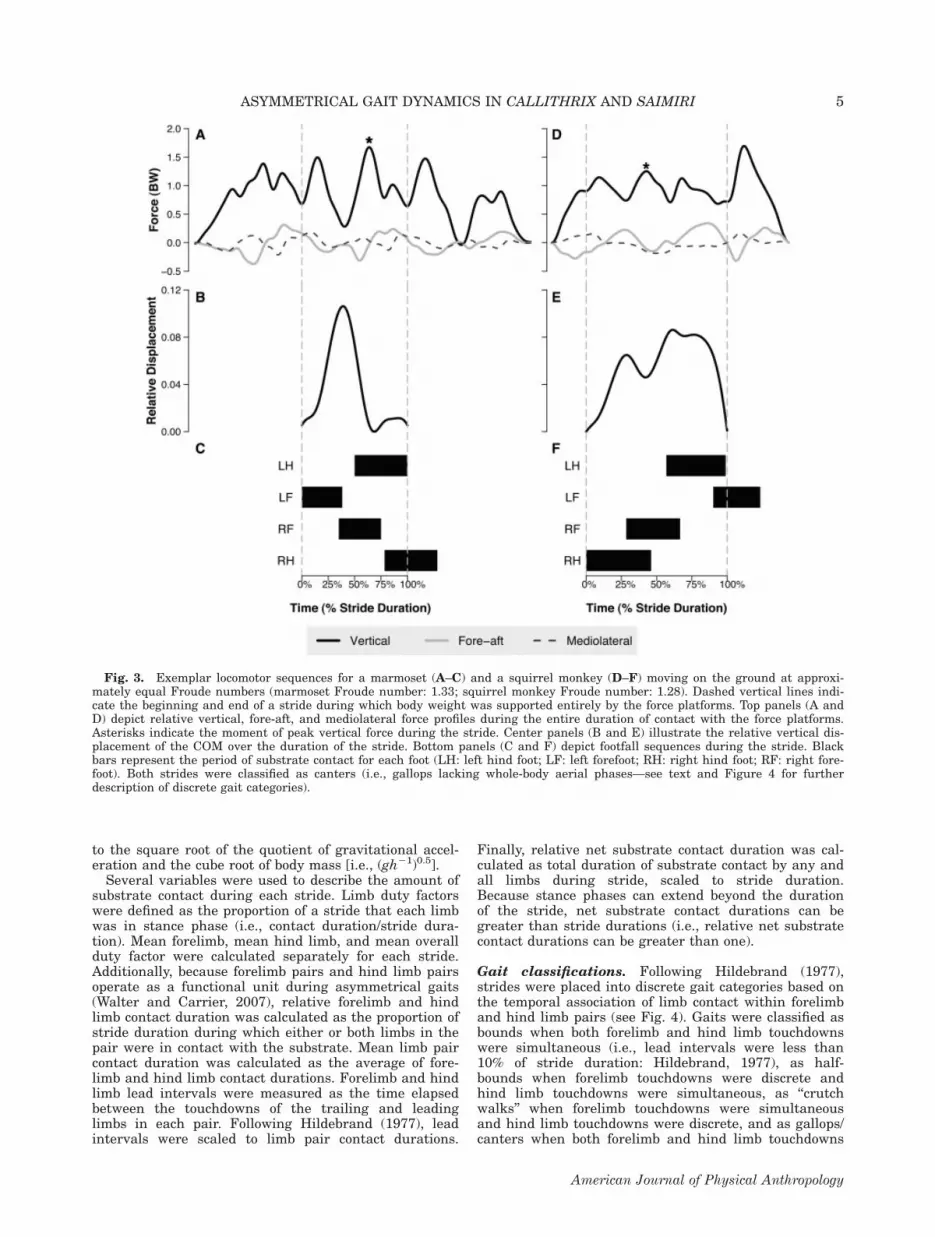
Vertical forces and COM displacements. Verticalforces and COM displacements were only measured forstrides, during which the animal’s body weight was com-pletely supported by the force platforms (see Fig. 3). Imeasured peak vertical force as the maximum verticalforce applied by the COM to the substrate during thecourse of the stride. The vertical displacements of theCOM were calculated from force data using standardmethods (Cavagna et al., 1977). The vertical force vector(minus body weight) was divided by body mass to calcu-late the instantaneous acceleration of the COM. Theacceleration vector was then integrated with respect tostride duration to obtain instantaneous velocity and thenintegrated again to obtain the instantaneous position ofthe COM. Because only steady-speed, horizontal locomo-tion was examined, the integration constant for velocitycalculations was defined such that mean vertical velocityequaled zero across the stride. The integration constantfor position calculations was set to zero. Total verticaldisplacement of the COM during each stride was quanti-fied as the net amplitude of positional changes duringthe stride. Peak vertical forces were scaled to bodyweight and COM displacement was scaled to the cuberoot of body mass to adjust for ontogenetic and interspe-cific differences in body size.
Stride variables. Stride symmetry was calculated asthe percentage of stride duration separating the touch-downs of the right and left hind limbs (Hildebrand,1977). Only strides in which symmetry was less than43.75% or greater than 56.25% were included in thedataset. Stride length was measured as the resultantdistance traversed by the reference marker from the be-ginning to the end of the stride. Stride frequency wasmeasured as the reciprocal of stride duration, wherestride duration was defined as the time in secondselapsed from the beginning to the end of the stride. Fol-lowing Hof (1996), stride length was scaled to the cuberoot of body mass, whereas stride frequency was scaled
4 J.W. YOUNG
American Journal of Physical Anthropology
to the square root of the quotient of gravitational accel-eration and the cube root of body mass [i.e., (gh21)0.5].Several variables were used to describe the amount of
substrate contact during each stride. Limb duty factorswere defined as the proportion of a stride that each limbwas in stance phase (i.e., contact duration/stride dura-tion). Mean forelimb, mean hind limb, and mean overallduty factor were calculated separately for each stride.Additionally, because forelimb pairs and hind limb pairsoperate as a functional unit during asymmetrical gaits(Walter and Carrier, 2007), relative forelimb and hindlimb contact duration was calculated as the proportion ofstride duration during which either or both limbs in thepair were in contact with the substrate. Mean limb paircontact duration was calculated as the average of fore-limb and hind limb contact durations. Forelimb and hindlimb lead intervals were measured as the time elapsedbetween the touchdowns of the trailing and leadinglimbs in each pair. Following Hildebrand (1977), leadintervals were scaled to limb pair contact durations.
Finally, relative net substrate contact duration was cal-culated as total duration of substrate contact by any andall limbs during stride, scaled to stride duration.Because stance phases can extend beyond the durationof the stride, net substrate contact durations can begreater than stride durations (i.e., relative net substratecontact durations can be greater than one).
Gait classifications. Following Hildebrand (1977),strides were placed into discrete gait categories based onthe temporal association of limb contact within forelimband hind limb pairs (see Fig. 4). Gaits were classified asbounds when both forelimb and hind limb touchdownswere simultaneous (i.e., lead intervals were less than10% of stride duration: Hildebrand, 1977), as half-bounds when forelimb touchdowns were discrete andhind limb touchdowns were simultaneous, as ‘‘crutchwalks’’ when forelimb touchdowns were simultaneousand hind limb touchdowns were discrete, and as gallops/canters when both forelimb and hind limb touchdowns
Fig. 3. Exemplar locomotor sequences for a marmoset (A–C) and a squirrel monkey (D–F) moving on the ground at approxi-mately equal Froude numbers (marmoset Froude number: 1.33; squirrel monkey Froude number: 1.28). Dashed vertical lines indi-cate the beginning and end of a stride during which body weight was supported entirely by the force platforms. Top panels (A andD) depict relative vertical, fore-aft, and mediolateral force profiles during the entire duration of contact with the force platforms.Asterisks indicate the moment of peak vertical force during the stride. Center panels (B and E) illustrate the relative vertical dis-placement of the COM over the duration of the stride. Bottom panels (C and F) depict footfall sequences during the stride. Blackbars represent the period of substrate contact for each foot (LH: left hind foot; LF: left forefoot; RH: right hind foot; RF: right fore-foot). Both strides were classified as canters (i.e., gallops lacking whole-body aerial phases—see text and Figure 4 for furtherdescription of discrete gait categories).
5ASYMMETRICAL GAIT DYNAMICS IN CALLITHRIX AND SAIMIRI
American Journal of Physical Anthropology

were discrete. Canters were classified as the subset ofgallops lacking whole-body aerial phases (Howell, 1944;Schmitt et al., 2006).
Precision of kinematic and kinetic data
To estimate precision in this dataset, 20 randomlyselected trials from each species were coded three timeson different days. For marmosets, replicate coefficientsof variation (i.e., standard deviation as a percentage ofthe mean) ranged from 2.5 to 21.7% across all variables,with an overall mean of 7.3% and a median of 4.2%. Forsquirrel monkeys, coefficients of variation ranged from1.4 to 8.3%, with a mean of 3.4% and a median of 3.5%.Forelimb and hind limb lead intervals had the highestcoefficients of variation, due to absolutely low mean val-ues. When lead intervals are excluded, all remainingvariables have coefficients of variation under 10% formarmoset trials and 5% for squirrel monkey trials.Moreover, variation due to measurement error was con-sistently minor compared to levels of biological variationwithin the experiments from which randomly sampledtrials was drawn.
Statistical analyses
Because none of the raw continuous variables exam-ined here followed a normal distribution, data were Box–Cox-transformed prior to analysis (Box and Cox, 1964).Box–Cox transformation yielded normal distributions formost, but not all, variables across the four species-by-substrate groups. Therefore, for comparisons in whichthe data for at least one of the groups were non-normallydistributed, parametric tests were supplemented by non-parametric analyses of rank-transformed data (Huitema,1980; Conover and Iman, 1981). In all cases, nonpara-metric tests corroborated parametric results, indicatingthat statistical procedures were robust to deviationsfrom normality.I used analyses of covariance (ANCOVA) and variance
(ANOVA) to test for substrate and species differences inasymmetrical gait dynamics. Where gait parameterswere significantly correlated with speed across all spe-cies-by-substrate groups, two-way ANCOVA was used,specifying substrate and species as factors and Froudenumber as the covariate. Where parameters were notcorrelated with Froude number in all conditions, orwhere regression slopes were significantly heterogeneousamong groups, the distribution of Froude numbers wascollapsed into three discrete categories: lowest valuethrough 33rd percentile, 33rd percentile through 66thpercentile, and 66th percentile through the highestvalue. Substrate and species differences were then testedusing three-way analyses of variance (ANOVA), specify-ing substrate, species, and Froude number category asfactors. Finally, I used two-way ANOVA to test for differ-ences in variables not correlated with Froude number,specifying substrate and species as factors. I tested forsignificant post-hoc substrate-related differences withinspecies and species-related differences within substratesusing tests of simple main effects, in which the meansquare between groups in each subset was tested againstthe error variance from the overall ANOVA/ANCOVA(Quinn and Keough, 2002). Finally, the significance ofthe species-by-substrate interaction term in the overallANOVA/ANCOVA was used to test for species-level dif-ferences in the magnitude of substrate-related changes.Because of the large discrepancy between squirrel
monkey and marmoset sample sizes (i.e., 156 marmosetstrides vs. 869 squirrel monkey strides, see Results sec-tion below), and the associated impacts on relative error
Fig. 4. Exemplar footfall sequences illustrating the fivetypes of asymmetrical gaits recognized in this study. Black barsdepict periods of substrate contact for each foot (abbreviationsas given in Fig. 3).
6 J.W. YOUNG
American Journal of Physical Anthropology

variances, F statistics computed during ANOVA/ANCOVA might have been too liberal. Stevens (2002)notes that whenever sample sizes differ by more than afactor of 1.5, and the variance of the larger group is sig-nificantly less than that of the smaller group, the likeli-hood of making a Type I error may be greater than thatindicated by the nominal significance level. Alternatively,if the variance of the larger group is significantly greaterthan that of the smaller group, calculated F statisticswill likely be conservative, decreasing the possibility ofmaking a Type I error. To investigate the possibility ofinflated Type I error in the current study, I used F-tests(Sokal and Rohlf, 1995) to compare sample variancesbetween species within substrates for each of the 14 de-pendent variables examined. Of the 28 comparisonsmade (i.e., 14 variables 3 two substrates), 17 indicatedthat error variances were either statistically equivalentbetween species or significantly greater in the largersquirrel monkey sample. In these cases, sample size dis-crepancies should have little effect on the interpretationof ANOVA/ANCOVA results. Nevertheless, squirrel mon-key error variances were significantly smaller than mar-moset error variances in 11 of the 28 comparisons. Thepossibility of making Type I errors in these cases mayhave been inflated over the nominal 5% level. Therefore,I used a resampling procedure (i.e., bootstrapping) toassess the potential distortion introduced by sample sizediscrepancies. First, appropriate numbers of ground andpole strides were randomly sampled from the squirrelmonkey data set to create a down-sampled squirrel mon-key dataset identical in size and composition to the mar-moset dataset. ANOVAs/ANCOVAs, including all interac-tions and associated tests of simple main effects, werethen recomputed using the full marmoset dataset andthe down-sampled squirrel monkey dataset. This processwas performed iteratively 10,000 times, and ‘‘bootstrapsupport indices,’’ calculated as the proportion of alldown-sampled tests in which P values were less than0.05, were computed for each significant comparisonfrom the original analysis. As indicated below, bootstrapsupport indices for most comparisons were high (i.e.,greater than 0.99; see Table 1 below), indicating thatmost tests were robust to discrepancies in sample size.I used G-tests of independence (Sokal and Rohlf, 1995)
to test for proportional differences in gait type betweensubstrates. Tests were performed separately for eachspecies. Standardized residuals, calculated as the differ-ences between observed and expected cell counts dividedby the square-root of the expected cell counts (Hab-erman, 1978), were compared to assess the magnitude ofsubstrate-related changes.I used a combination of multiple regression and
ANCOVA to test whether ontogenetic increases in bodymass within each species had differential effects duringground and pole locomotion. First, multiple regressionsof each parameter on Froude number and body masswere used to quantify the independent relationshipbetween gait mechanics and body size, controlling forFroude number. Second, homogeneous slope tests (i.e.,the first part of a standard ANCOVA) were used to testif patterns of size-related change differed by substrate.An iterative process was used to build the proper covari-ate-by-factor interaction terms for each homogenousslope test (Crawley, 2007). First, tests were carried outusing the fully specified model, with all interactionterms included (i.e., mass-by-Froude number, mass-by-substrate, Froude number-by-substrate, Froude number-
TABLE
1.Substra
teandsp
eciesdifferencesin
asymmetrica
lgait
dyn
amics
Variable
Callithrix
Saim
iri
Substrate
difference
Speciesdifference
Interaction
GP
GP
Callithrix
Saim
iri
GP
Froudenumber
2.26�
0.133(76)
1.95�
0.121(80)
2.05�
0.050(499)
2.00�
0.057(370)
G[
P(14.0%)[1.0000]
G5
P(2.5%)[–]
Cj[
Sb(9.3%)[0.5481]
Cj5
Sb(2.7%)[–]
Cj[
Sb[0.4232]
Relativepea
kverticalforce
2.39�
0.196(21)
1.81�
0.112(23)
1.92�
0.076(94)
1.84�
0.064(105)
G[
P(24.2%)[1.0000]
G[
P(13.2%)[0.7338]
Cj[
Sb(19.7%)[0.9735]
Cj5
Sb(8.0%)[–]
Cj[
Sb[0.3260]
RelativeverticalCOM
displacemen
t
0.245�
0.051(21)
0.132�
0.021(23)
0.184�
0.016(94)
0.120�
0.009(105)
G[
P(46.3%)[1.0000]
G[
P(34.7%)[1.0000]
Cj[
Sb(25.0%)[1.0000]
Cj5
Sb(8.7%)[–]
Cj5
Sb[–]
Forelim
bduty
factor
0.289�
0.008(76)
0.353�
0.009(80)
0.334�
0.004(499)
0.355�
0.004(370)
G\
P(19.1%)[1.0000]
G\
P(5.7%)[0.8795]
Cj\
Sb(13.5%)[1.0000]
Cj5
Sb(0.5%)[–]
Cj[
Sb[1.0000]
Hindlimbduty
factor
0.356�
0.009(76)
0.411
�0.010(80)
0.371�
0.003(499)
0.386�
0.004(370)
G\
P(14.8%)[1.0000]
G\
P(3.9%)[0.5774]
Cj\
Sb(4.1%)[0.5523]
Cj[
Sb(6.0%)[0.9876]
Cj[
Sb[0.9979]
Mea
nduty
factor
0.324�
0.007(76)
0.374�
0.009(80)
0.353�
0.003(499)
0.371�
0.004(370)
G\
P(16.4%)[1.0000]
G\
P(4.9%)[0.8477]
Cj\
Sb(8.0%)[1.0000]
Cj[
Sb(3.0%)[0.2548]
Cj[
Sb[1.0000]
Relativeforelimb
leadinterval
0.357�
0.029(76)
0.405�
0.025(80)
0.387�
0.010(499)
0.523�
0.009(370)
G\
P(12.6%)[1.0000]
G\
P(26.0%)[1.0000]
Cj5
Sb(7.8%)[–]
Cj\
Sb(22.5%)[1.0000]
Cj\
Sb[0.9986]
Relativehindlimblead
interval
0.105�
0.035(76)
0.201�
0.033(80)
0.281�
0.013(499)
0.446�
0.014(370)
G\
P(34.0%)[1.0000]
G\
P(37.1%)[1.0000]
Cj\
Sb(62.6%)[1.0000]
Cj\
Sb(55.0%)[1.0000]
Cj\
Sb[0.811
8]
Relativeforelimbcontact
duration
0.423�
0.023(76)
0.552�
0.027(80)
0.529�
0.010(499)
0.715�
0.015(370)
G\
P(22.3%)[1.0000]
G\
P(24.9%)[1.0000]
Cj\
Sb(20.9%)[1.0000]
Cj\
Sb(23.9%)[1.0000]
Cj5
Sb[–]
Relativehindlimb
contact
duration
0.382�
0.019(76)
0.483�
0.025(80)
0.498�
0.010(499)
0.688�
0.017(370)
G\
P(20.2%)[1.0000]
G\
P(27.6%)[1.0000]
Cj\
Sb(23.2%)[1.0000]
Cj\
Sb(29.8%)[1.0000]
Cj5
Sb[–]
Relativemea
ncontact
duration
0.401�
0.018(76)
0.519�
0.022(80)
0.516�
0.009(499)
0.702�
0.014(370)
G\
P(22.9%)[1.0000]
G\
P(26.4%)[1.0000]
Cj\
Sb(22.4%)[1.0000]
Cj\
Sb(26.1%)[1.0000]
Cj5
Sb[–]
Relativesu
bstrate
contact
duration
0.818�
0.028(76)
1.005�
0.026(80)
0.966�
0.010(499)
1.082�
0.012(370)
G\
P(19.3%)[1.0000]
G\
P(10.7%)[1.0000]
Cj\
Sb(15.4%)[1.0000]
Cj\
Sb(7.1%)[0.9999]
Cj[
Sb[0.7607]
Relativestridelength
0.636�
0.019(76)
0.507�
0.016(79)
0.703�
0.009(452)
0.639�
0.009(344)
G[
P(18.4%)[1.0000]
G[
P(9.2%)[0.9973]
Cj\
Sb(9.5%)[0.9998]
Cj\
Sb(20.6%)[1.0000]
Cj[
Sb[0.9970]
Relativestridefreq
uen
cy0.320�
0.010(76)
0.398�
0.011
(80)
0.285�
0.003(499)
0.318�
0.004(370)
G\
P(24.5%)[1.0000]
G\
P(10.3%)[0.9997]
Cj[
Sb(10.7%)[1.0000]
Cj[
Sb(20.0%)[1.0000]
Cj[
Sb[0.9921]
Rep
ortedvalues
wereadjusted
forbod
ysize.Mea
ns�
standard
errors
andsa
mple
sizes(inparentheses)are
reportedforea
chsp
ecieson
each
substrate
(G5
ground,P
5pole).
Percentagedifferences(calculatedrelativeto
thelarger
value)
are
show
nin
parentheses
inea
chcomparison.Bootstrapsu
pportindices
forallsignificantcomparisonsandinterac-
tion
testsare
show
nin
square
brack
ets(see
textfordetails).Boldprintindicatessignificance
(P�
0.05).
7ASYMMETRICAL GAIT DYNAMICS IN CALLITHRIX AND SAIMIRI
American Journal of Physical Anthropology
by-mass-by-substrate). Then, any nonsignificant signifi-cant interaction terms including Froude number wereremoved from the model in hierarchical order and allstatistics were recalculated. All ANOVAs/ANCOVAs andG-tests were performed using SPSS 11.0 for Macintosh(SPSS, Chicago, IL). Bootstrap analyses were performedusing the R statistical package (R Development CoreTeam, 2008).
RESULTS
A total of 1,025 strides were analyzed, including 76marmoset and 499 squirrel monkey strides on theground and 80 marmoset and 370 squirrel monkeystrides on the pole. A subset of 231 strides was availablefor kinetic analyses, including 21 marmoset and 94squirrel monkey strides on the ground and 23 marmosetand 105 squirrel monkey strides on the pole.
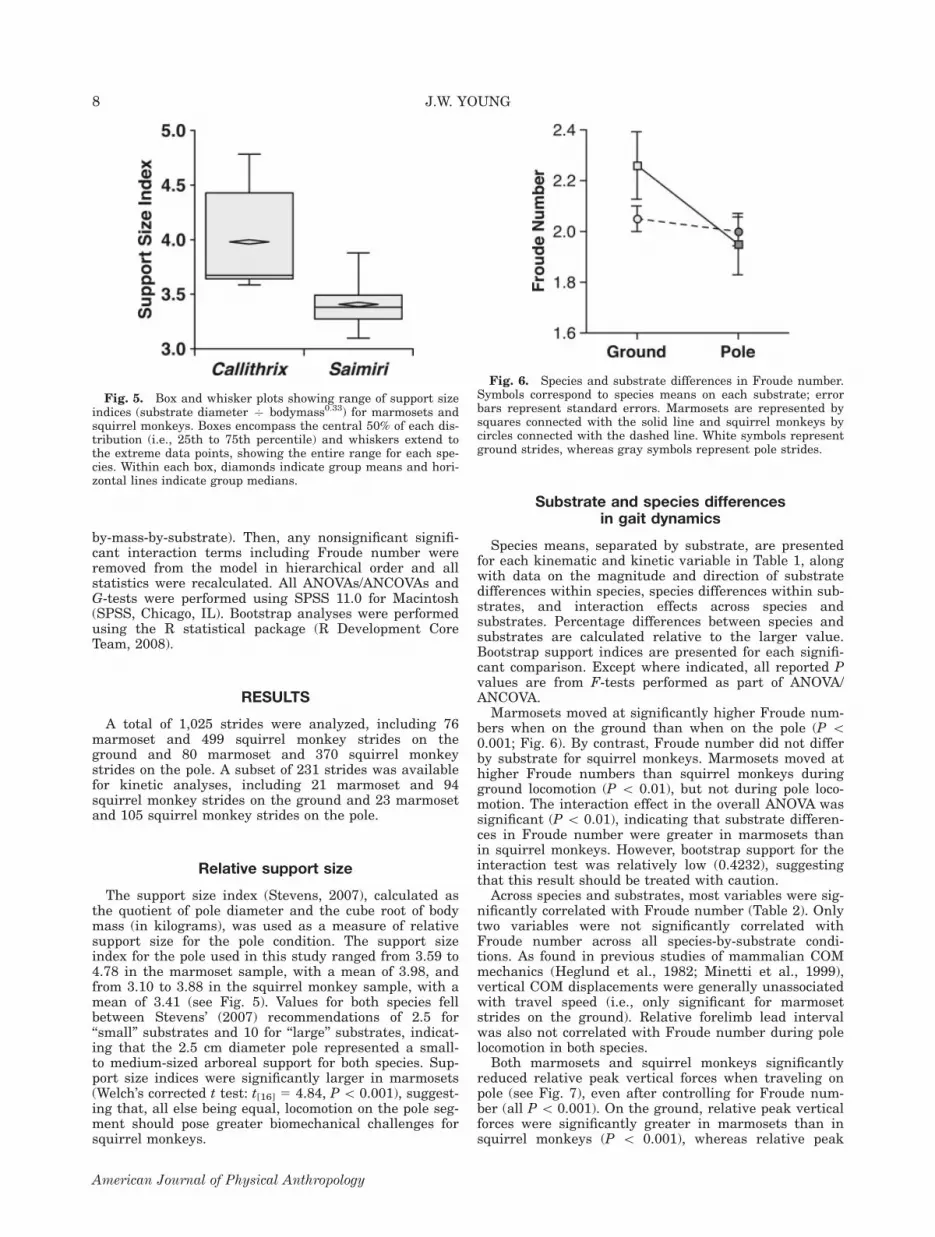
Relative support size
The support size index (Stevens, 2007), calculated asthe quotient of pole diameter and the cube root of bodymass (in kilograms), was used as a measure of relativesupport size for the pole condition. The support sizeindex for the pole used in this study ranged from 3.59 to4.78 in the marmoset sample, with a mean of 3.98, andfrom 3.10 to 3.88 in the squirrel monkey sample, with amean of 3.41 (see Fig. 5). Values for both species fellbetween Stevens’ (2007) recommendations of 2.5 for‘‘small’’ substrates and 10 for ‘‘large’’ substrates, indicat-ing that the 2.5 cm diameter pole represented a small-to medium-sized arboreal support for both species. Sup-port size indices were significantly larger in marmosets(Welch’s corrected t test: t[16] 5 4.84, P\ 0.001), suggest-ing that, all else being equal, locomotion on the pole seg-ment should pose greater biomechanical challenges forsquirrel monkeys.
Substrate and species differencesin gait dynamics
Species means, separated by substrate, are presentedfor each kinematic and kinetic variable in Table 1, alongwith data on the magnitude and direction of substratedifferences within species, species differences within sub-strates, and interaction effects across species andsubstrates. Percentage differences between species andsubstrates are calculated relative to the larger value.Bootstrap support indices are presented for each signifi-cant comparison. Except where indicated, all reported Pvalues are from F-tests performed as part of ANOVA/ANCOVA.Marmosets moved at significantly higher Froude num-
bers when on the ground than when on the pole (P \0.001; Fig. 6). By contrast, Froude number did not differby substrate for squirrel monkeys. Marmosets moved athigher Froude numbers than squirrel monkeys duringground locomotion (P \ 0.01), but not during pole loco-motion. The interaction effect in the overall ANOVA wassignificant (P \ 0.01), indicating that substrate differen-ces in Froude number were greater in marmosets thanin squirrel monkeys. However, bootstrap support for theinteraction test was relatively low (0.4232), suggestingthat this result should be treated with caution.Across species and substrates, most variables were sig-
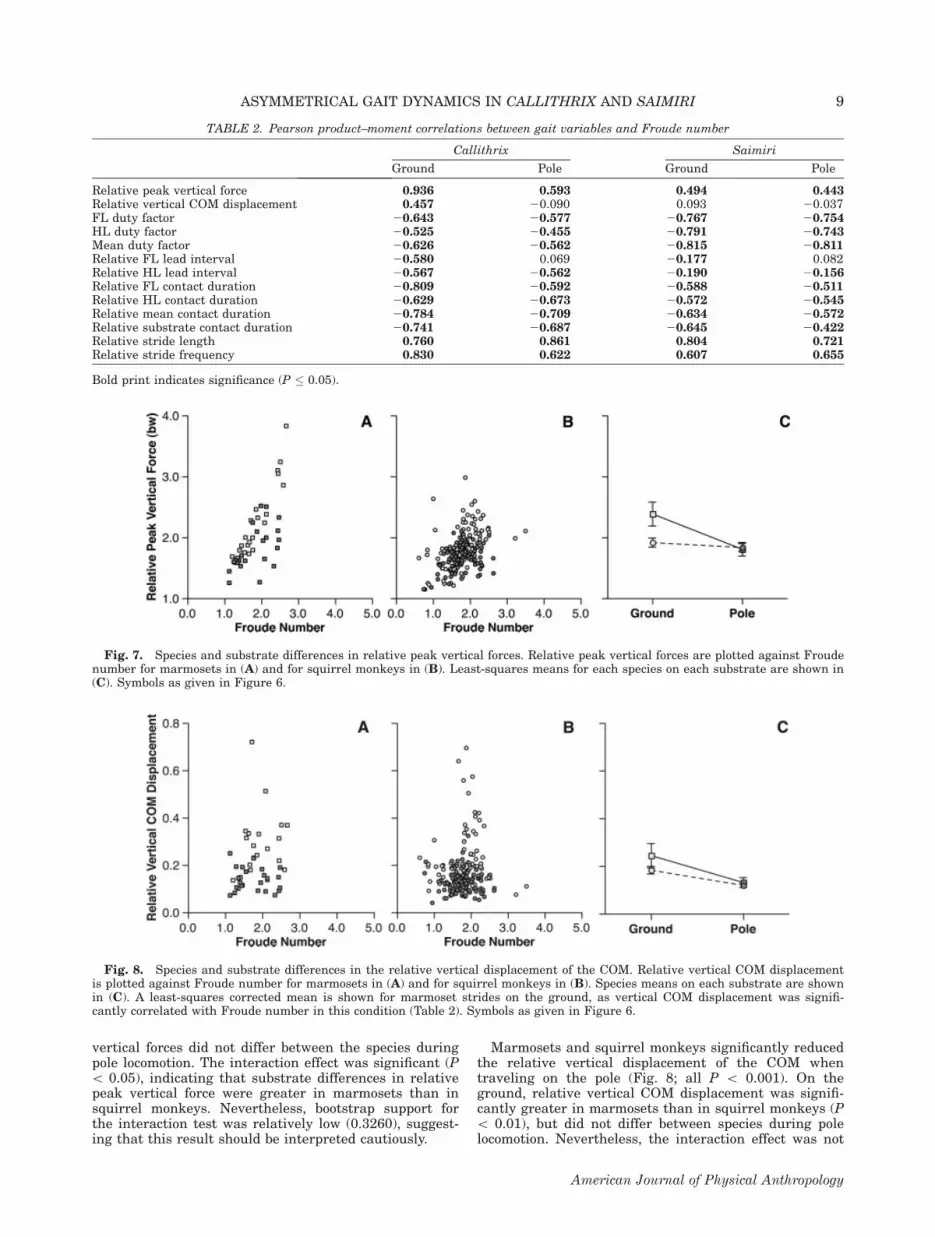
nificantly correlated with Froude number (Table 2). Onlytwo variables were not significantly correlated withFroude number across all species-by-substrate condi-tions. As found in previous studies of mammalian COMmechanics (Heglund et al., 1982; Minetti et al., 1999),vertical COM displacements were generally unassociatedwith travel speed (i.e., only significant for marmosetstrides on the ground). Relative forelimb lead intervalwas also not correlated with Froude number during polelocomotion in both species.Both marmosets and squirrel monkeys significantly
reduced relative peak vertical forces when traveling onpole (see Fig. 7), even after controlling for Froude num-ber (all P\ 0.001). On the ground, relative peak verticalforces were significantly greater in marmosets than insquirrel monkeys (P \ 0.001), whereas relative peak
Fig. 5. Box and whisker plots showing range of support sizeindices (substrate diameter 7 bodymass0.33) for marmosets andsquirrel monkeys. Boxes encompass the central 50% of each dis-tribution (i.e., 25th to 75th percentile) and whiskers extend tothe extreme data points, showing the entire range for each spe-cies. Within each box, diamonds indicate group means and hori-zontal lines indicate group medians.
Fig. 6. Species and substrate differences in Froude number.Symbols correspond to species means on each substrate; errorbars represent standard errors. Marmosets are represented bysquares connected with the solid line and squirrel monkeys bycircles connected with the dashed line. White symbols representground strides, whereas gray symbols represent pole strides.
8 J.W. YOUNG
American Journal of Physical Anthropology
vertical forces did not differ between the species duringpole locomotion. The interaction effect was significant (P\ 0.05), indicating that substrate differences in relativepeak vertical force were greater in marmosets than insquirrel monkeys. Nevertheless, bootstrap support forthe interaction test was relatively low (0.3260), suggest-ing that this result should be interpreted cautiously.
Marmosets and squirrel monkeys significantly reducedthe relative vertical displacement of the COM whentraveling on the pole (Fig. 8; all P \ 0.001). On theground, relative vertical COM displacement was signifi-cantly greater in marmosets than in squirrel monkeys (P\ 0.01), but did not differ between species during polelocomotion. Nevertheless, the interaction effect was not
TABLE 2. Pearson product–moment correlations between gait variables and Froude number
Callithrix Saimiri
Ground Pole Ground Pole
Relative peak vertical force 0.936 0.593 0.494 0.443Relative vertical COM displacement 0.457 20.090 0.093 20.037FL duty factor 20.643 20.577 20.767 20.754HL duty factor 20.525 20.455 20.791 20.743Mean duty factor 20.626 20.562 20.815 20.811Relative FL lead interval 20.580 0.069 20.177 0.082Relative HL lead interval 20.567 20.562 20.190 20.156Relative FL contact duration 20.809 20.592 20.588 20.511Relative HL contact duration 20.629 20.673 20.572 20.545Relative mean contact duration 20.784 20.709 20.634 20.572Relative substrate contact duration 20.741 20.687 20.645 20.422Relative stride length 0.760 0.861 0.804 0.721Relative stride frequency 0.830 0.622 0.607 0.655
Bold print indicates significance (P � 0.05).
Fig. 7. Species and substrate differences in relative peak vertical forces. Relative peak vertical forces are plotted against Froudenumber for marmosets in (A) and for squirrel monkeys in (B). Least-squares means for each species on each substrate are shown in(C). Symbols as given in Figure 6.
Fig. 8. Species and substrate differences in the relative vertical displacement of the COM. Relative vertical COM displacementis plotted against Froude number for marmosets in (A) and for squirrel monkeys in (B). Species means on each substrate are shownin (C). A least-squares corrected mean is shown for marmoset strides on the ground, as vertical COM displacement was signifi-cantly correlated with Froude number in this condition (Table 2). Symbols as given in Figure 6.
9ASYMMETRICAL GAIT DYNAMICS IN CALLITHRIX AND SAIMIRI
American Journal of Physical Anthropology

significant, indicating that marmosets and squirrel mon-keys showed similar reductions in COM movement whentraveling on the pole.Both species significantly increased forelimb duty fac-
tor, hind limb duty factor, and mean duty factor duringlocomotion on the pole, even after controlling for theeffects of Froude number (Fig. 9; all P \ 0.001). Squirrelmonkeys displayed significantly greater forelimb, hindlimb, and mean duty factors than marmosets duringground locomotion (all P \ 0.001). By contrast, duringpole locomotion, forelimb duty factor did not significantlydiffer between species, and hind limb factors weregreater in marmosets (P \ 0.001). Mean duty factorsduring pole locomotion were also nominally greater inmarmosets (P \ 0.05). However, low bootstrap support(0.2548) suggests that this result should be viewed withcaution. Interaction effects were significant across limbs,indicating that substrate differences in duty factor wereconsistently greater in marmosets than in squirrel mon-keys (all P\ 0.001).Both species significantly increased relative forelimb
lead intervals and relative hind limb lead intervals whentraveling on the pole (Fig. 10; all P \ 0.001). Squirrelmonkey relative forelimb lead intervals were marginallygreater during locomotion on the ground (P 5 0.051) andsignificantly greater during locomotion on the pole (P \0.001). Across substrates, relative hind limb lead inter-
vals were greater in squirrel monkeys (all P \ 0.001). Incontrast to most other gait parameters examined,changes in relative lead intervals were greater in squir-rel monkeys than in marmosets (all P\ 0.05).Increases in limb duty factors and lead intervals
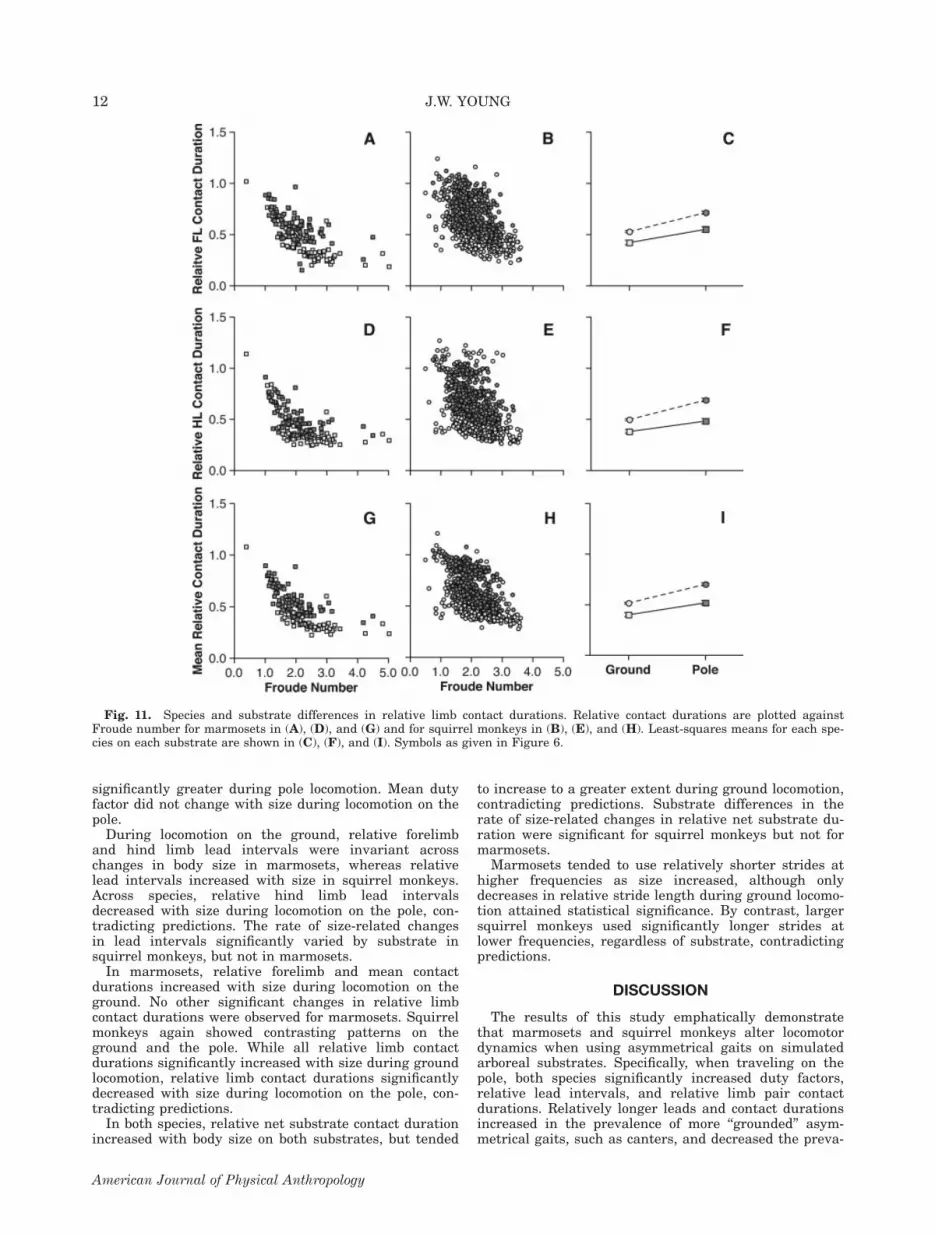
caused significant increases in relative forelimb, hindlimb, and mean limb pair contact durations during loco-motion on the pole, even after controlling for Froudenumber (Fig. 11; all P \ 0.001). Relative contact dura-tions were consistently longer in squirrel monkeys thanin marmosets, both on the ground and on the pole (allP\ 0.001). Across limbs, interaction effects were not sig-nificant, indicating that substrate-related changes inlimb contact durations were similar across species (allP[ 0.05).For both species, substrate differences in relative lead
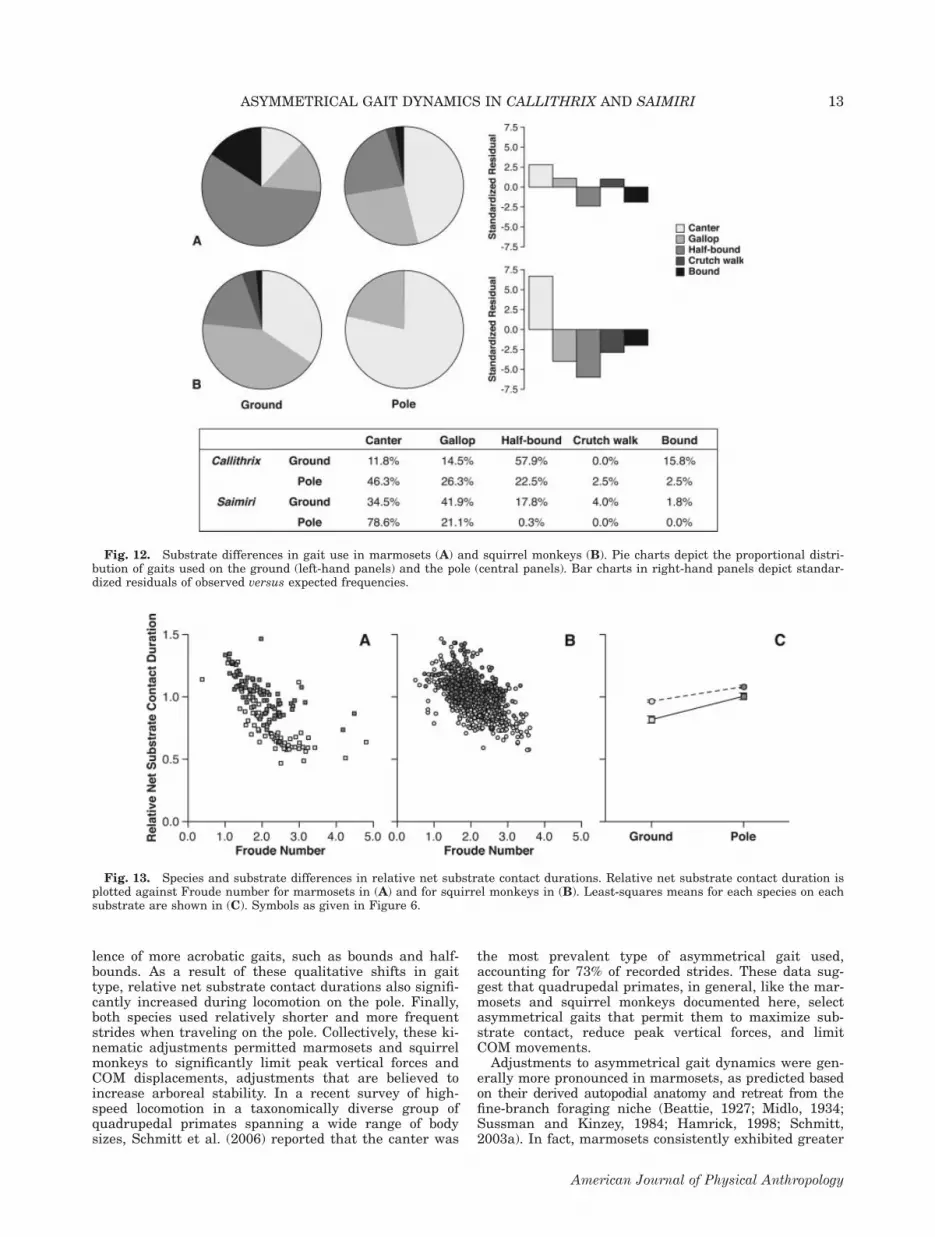
intervals and contact durations were also manifested bysignificant categorical changes in discrete gait types(Fig. 12; G-tests: all P \ 0.001). Examination of standar-dized residuals revealed that substrate differences wereprimarily due to substantial increases in the frequencyof canters used during pole locomotion. On the ground,canters accounted for 11.8% of marmoset gaits and34.5% of squirrel monkey gaits. By contrast, when ani-mals were traveling on the pole, canters accounted for49.4% of marmoset gaits and 78.6% of squirrel monkeygaits. In marmosets, an increase in the frequency of
Fig. 9. Species and substrate differences in limb duty factors. Duty factors are plotted against Froude number for marmosets in(A), (D), and (G) and for squirrel monkeys in (B), (E), and (H). Least-squares means for each species on each substrate are shownin (C), (F), and (I). Symbols as given in Figure 6.
10 J.W. YOUNG
American Journal of Physical Anthropology

canters was accompanied by decreases in the frequencyof half-bounds and bounds and slight increases in thefrequency of gallops and crutch walks. In squirrel mon-keys, all gaits other than canters decreased in frequencyduring pole locomotion.As predicted, a shift toward more asynchronous,
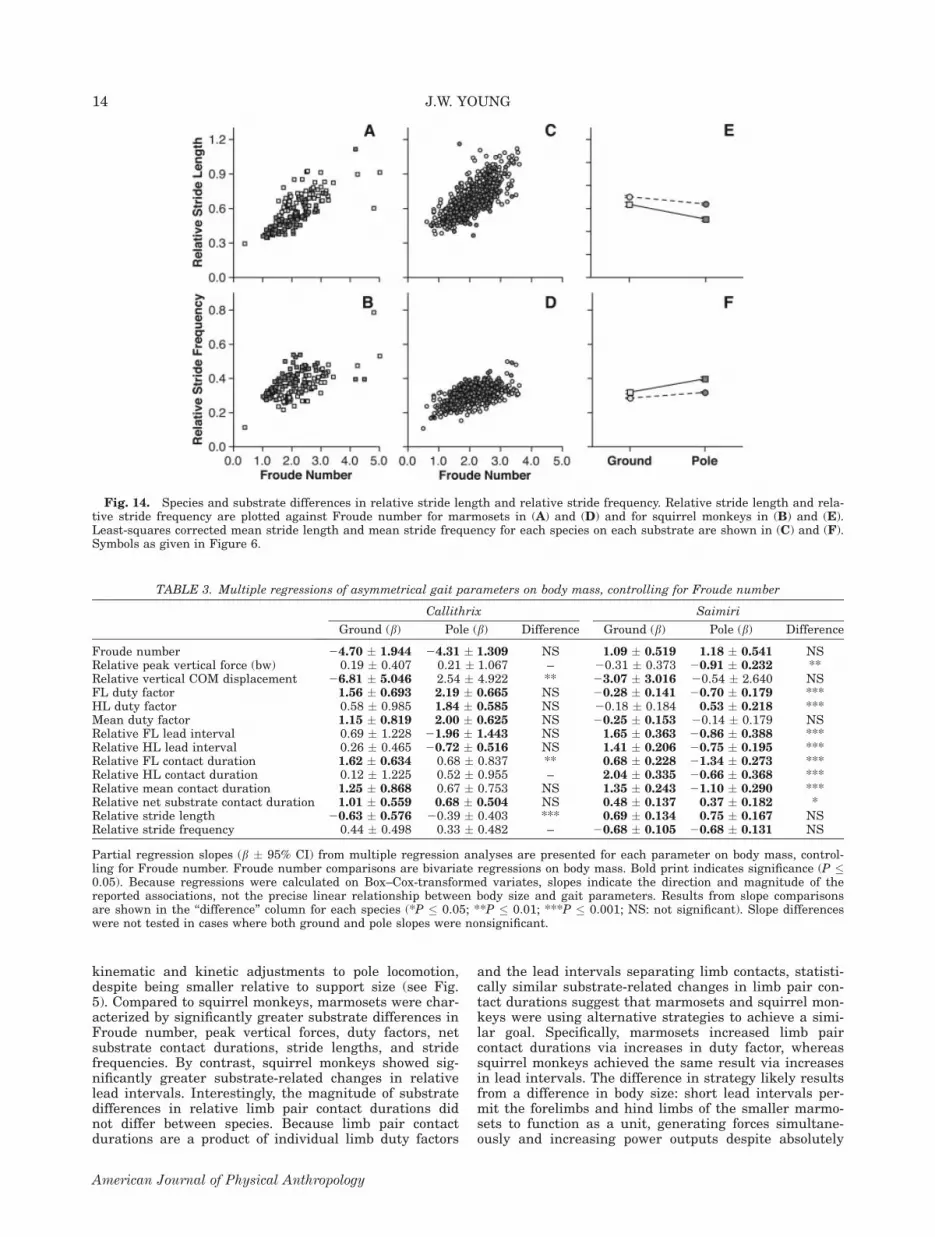
‘‘grounded’’ asymmetrical gaits, such as the canter,resulted in significant increases in relative net substratecontact duration in both species, even after controlling ofthe effects of Froude number (Fig. 13; all P \ 0.001).Relative substrate contact duration was consistentlygreater in squirrel monkeys than in marmosets, both onthe ground and on the pole (all P \ 0.001). The interac-tion effect was significant, indicating that the magnitudeof substrate differences in relative net substrate contactduration was greater in marmosets than in squirrelmonkeys (P\ 0.01).Finally, in both marmosets and squirrel monkeys, loco-
motion on the pole was associated with decreased rela-tive stride length and increased relative stride frequency,even after controlling for Froude number (Fig. 14; all P\ 0.001). On both substrates, marmosets used relativelyshorter and more frequent strides than squirrel monkeys(all P \ 0.001). Interaction effects for both variableswere significant, indicating that the magnitude of sub-strate-related change was again greater in marmosetsthan in squirrel monkeys (all P\ 0.001).
Ontogenetic changes in asymmetrical gaitdynamics
Results from multiple regression analyses of each gaitparameter on body mass, controlling for Froude number,are shown in Table 3. Across substrates, both marmosetsand squirrel monkeys showed significant size-relatedchanges in Froude number, although the direction of
change differed between species. Marmosets moved atsignificantly slower Froude numbers as body sizeincreased, whereas larger squirrel monkeys moved atsignificantly faster Froude numbers. In both species,Froude number changed with body mass at a similarrate across substrates, contradicting predictions.In marmosets, relative peak vertical forces did not sig-
nificantly vary with body size on either substrate. Aspredicted, larger squirrel monkeys significantly de-creased relative peak vertical forces when moving on thepole, but not when moving on the ground, a differencethat was statistically significant. During ground locomo-tion, both marmosets and squirrel monkeys significantlyreduced relative vertical COM displacements as bodysize increased. During pole locomotion, however, bothspecies maintained similar COM displacements acrosschanges in body size. The rate of size-related changes inrelative vertical COM displacements significantly dif-fered between substrates in marmosets, but not in squir-rel monkeys.Both species showed several significant size-related
changes in relative limb and net substrate contact pat-terns, although few of these changes accorded with pre-dictions. With the exception of hind limb duty factor dur-ing locomotion on the ground, larger marmosets usedsignificantly greater duty factors across all limbs andsubstrates. Although the rate of size-related increases induty factors tended to be greater during locomotion onthe pole, differences in slope were not statistically signif-icant. In squirrel monkeys, hind limb duty factor was in-dependent of size on the ground but significantlyincreased with size on the pole, corroborating predic-tions. Contradicting predictions, larger squirrel monkeyssignificantly decreased forelimb duty factor on both sub-strates and mean duty factor during ground locomotion.Moreover, the rate of forelimb duty factor decrease was
Fig. 10. Species and substrate differences in relative lead interval. Lead intervals are plotted against Froude number for mar-mosets in (A) and (D) and for squirrel monkeys in (B) and (E). Least-squares means for each species on each substrate are shownin (C) and (F). Uncorrected means are shown for forelimb lead intervals on the pole, as this parameter was not correlated withFroude number in either species (Table 2). Symbols as given in Figure 6.
11ASYMMETRICAL GAIT DYNAMICS IN CALLITHRIX AND SAIMIRI
American Journal of Physical Anthropology
significantly greater during pole locomotion. Mean dutyfactor did not change with size during locomotion on thepole.During locomotion on the ground, relative forelimb
and hind limb lead intervals were invariant acrosschanges in body size in marmosets, whereas relativelead intervals increased with size in squirrel monkeys.Across species, relative hind limb lead intervalsdecreased with size during locomotion on the pole, con-tradicting predictions. The rate of size-related changesin lead intervals significantly varied by substrate insquirrel monkeys, but not in marmosets.In marmosets, relative forelimb and mean contact
durations increased with size during locomotion on theground. No other significant changes in relative limbcontact durations were observed for marmosets. Squirrelmonkeys again showed contrasting patterns on theground and the pole. While all relative limb contactdurations significantly increased with size during groundlocomotion, relative limb contact durations significantlydecreased with size during locomotion on the pole, con-tradicting predictions.In both species, relative net substrate contact duration
increased with body size on both substrates, but tended
to increase to a greater extent during ground locomotion,contradicting predictions. Substrate differences in therate of size-related changes in relative net substrate du-ration were significant for squirrel monkeys but not formarmosets.Marmosets tended to use relatively shorter strides at
higher frequencies as size increased, although onlydecreases in relative stride length during ground locomo-tion attained statistical significance. By contrast, largersquirrel monkeys used significantly longer strides atlower frequencies, regardless of substrate, contradictingpredictions.
DISCUSSION
The results of this study emphatically demonstratethat marmosets and squirrel monkeys alter locomotordynamics when using asymmetrical gaits on simulatedarboreal substrates. Specifically, when traveling on thepole, both species significantly increased duty factors,relative lead intervals, and relative limb pair contactdurations. Relatively longer leads and contact durationsincreased in the prevalence of more ‘‘grounded’’ asym-metrical gaits, such as canters, and decreased the preva-
Fig. 11. Species and substrate differences in relative limb contact durations. Relative contact durations are plotted againstFroude number for marmosets in (A), (D), and (G) and for squirrel monkeys in (B), (E), and (H). Least-squares means for each spe-cies on each substrate are shown in (C), (F), and (I). Symbols as given in Figure 6.
12 J.W. YOUNG
American Journal of Physical Anthropology
lence of more acrobatic gaits, such as bounds and half-bounds. As a result of these qualitative shifts in gaittype, relative net substrate contact durations also signifi-cantly increased during locomotion on the pole. Finally,both species used relatively shorter and more frequentstrides when traveling on the pole. Collectively, these ki-nematic adjustments permitted marmosets and squirrelmonkeys to significantly limit peak vertical forces andCOM displacements, adjustments that are believed toincrease arboreal stability. In a recent survey of high-speed locomotion in a taxonomically diverse group ofquadrupedal primates spanning a wide range of bodysizes, Schmitt et al. (2006) reported that the canter was
the most prevalent type of asymmetrical gait used,accounting for 73% of recorded strides. These data sug-gest that quadrupedal primates, in general, like the mar-mosets and squirrel monkeys documented here, selectasymmetrical gaits that permit them to maximize sub-strate contact, reduce peak vertical forces, and limitCOM movements.Adjustments to asymmetrical gait dynamics were gen-
erally more pronounced in marmosets, as predicted basedon their derived autopodial anatomy and retreat from thefine-branch foraging niche (Beattie, 1927; Midlo, 1934;Sussman and Kinzey, 1984; Hamrick, 1998; Schmitt,2003a). In fact, marmosets consistently exhibited greater
Fig. 12. Substrate differences in gait use in marmosets (A) and squirrel monkeys (B). Pie charts depict the proportional distri-bution of gaits used on the ground (left-hand panels) and the pole (central panels). Bar charts in right-hand panels depict standar-dized residuals of observed versus expected frequencies.
Fig. 13. Species and substrate differences in relative net substrate contact durations. Relative net substrate contact duration isplotted against Froude number for marmosets in (A) and for squirrel monkeys in (B). Least-squares means for each species on eachsubstrate are shown in (C). Symbols as given in Figure 6.
13ASYMMETRICAL GAIT DYNAMICS IN CALLITHRIX AND SAIMIRI
American Journal of Physical Anthropology
kinematic and kinetic adjustments to pole locomotion,despite being smaller relative to support size (see Fig.5). Compared to squirrel monkeys, marmosets were char-acterized by significantly greater substrate differences inFroude number, peak vertical forces, duty factors, netsubstrate contact durations, stride lengths, and stridefrequencies. By contrast, squirrel monkeys showed sig-nificantly greater substrate-related changes in relativelead intervals. Interestingly, the magnitude of substratedifferences in relative limb pair contact durations didnot differ between species. Because limb pair contactdurations are a product of individual limb duty factors
and the lead intervals separating limb contacts, statisti-cally similar substrate-related changes in limb pair con-tact durations suggest that marmosets and squirrel mon-keys were using alternative strategies to achieve a simi-lar goal. Specifically, marmosets increased limb paircontact durations via increases in duty factor, whereassquirrel monkeys achieved the same result via increasesin lead intervals. The difference in strategy likely resultsfrom a difference in body size: short lead intervals per-mit the forelimbs and hind limbs of the smaller marmo-sets to function as a unit, generating forces simultane-ously and increasing power outputs despite absolutely
Fig. 14. Species and substrate differences in relative stride length and relative stride frequency. Relative stride length and rela-tive stride frequency are plotted against Froude number for marmosets in (A) and (D) and for squirrel monkeys in (B) and (E).Least-squares corrected mean stride length and mean stride frequency for each species on each substrate are shown in (C) and (F).Symbols as given in Figure 6.
TABLE 3. Multiple regressions of asymmetrical gait parameters on body mass, controlling for Froude number
Callithrix Saimiri
Ground (b) Pole (b) Difference Ground (b) Pole (b) Difference
Froude number 24.70 � 1.944 24.31 � 1.309 NS 1.09 � 0.519 1.18 � 0.541 NSRelative peak vertical force (bw) 0.19 � 0.407 0.21 � 1.067 – 20.31 � 0.373 20.91 � 0.232 **Relative vertical COM displacement 26.81 � 5.046 2.54 � 4.922 ** 23.07 � 3.016 20.54 � 2.640 NSFL duty factor 1.56 � 0.693 2.19 � 0.665 NS 20.28 � 0.141 20.70 � 0.179 ***HL duty factor 0.58 � 0.985 1.84 � 0.585 NS 20.18 � 0.184 0.53 � 0.218 ***Mean duty factor 1.15 � 0.819 2.00 � 0.625 NS 20.25 � 0.153 20.14 � 0.179 NSRelative FL lead interval 0.69 � 1.228 21.96 � 1.443 NS 1.65 � 0.363 20.86 � 0.388 ***Relative HL lead interval 0.26 � 0.465 20.72 � 0.516 NS 1.41 � 0.206 20.75 � 0.195 ***Relative FL contact duration 1.62 � 0.634 0.68 � 0.837 ** 0.68 � 0.228 21.34 � 0.273 ***Relative HL contact duration 0.12 � 1.225 0.52 � 0.955 – 2.04 � 0.335 20.66 � 0.368 ***Relative mean contact duration 1.25 � 0.868 0.67 � 0.753 NS 1.35 � 0.243 21.10 � 0.290 ***Relative net substrate contact duration 1.01 � 0.559 0.68 � 0.504 NS 0.48 � 0.137 0.37 � 0.182 *Relative stride length 20.63 � 0.576 20.39 � 0.403 *** 0.69 � 0.134 0.75 � 0.167 NSRelative stride frequency 0.44 � 0.498 0.33 � 0.482 – 20.68 � 0.105 20.68 � 0.131 NS
Partial regression slopes (b � 95% CI) from multiple regression analyses are presented for each parameter on body mass, control-ling for Froude number. Froude number comparisons are bivariate regressions on body mass. Bold print indicates significance (P �0.05). Because regressions were calculated on Box–Cox-transformed variates, slopes indicate the direction and magnitude of thereported associations, not the precise linear relationship between body size and gait parameters. Results from slope comparisonsare shown in the ‘‘difference’’ column for each species (*P � 0.05; **P � 0.01; ***P � 0.001; NS: not significant). Slope differenceswere not tested in cases where both ground and pole slopes were nonsignificant.
14 J.W. YOUNG
American Journal of Physical Anthropology

shorter limbs (Gambaryan, 1974; Hildebrand, 1977; Wal-ter and Carrier, 2007).Comparisons between species within each substrate
frequently demonstrated significant differences duringground locomotion but similarity during pole locomotion.For instance, when traveling on the ground marmosetsexhibited significantly greater peak vertical forces andCOM displacements and significantly shorter forelimband mean duty factors. None of these parameters signifi-cantly varied between species during pole locomotion.Lack of variation between species during locomotion onthe pole is consistent with increased stability constraintson narrow substrates, limiting kinematic and kineticflexibility despite differences in ecology and anatomy.Nevertheless, other species-level differences were con-sistent across substrates. On both the ground and thepole, squirrel monkeys used longer leads and limb con-tact durations and used a higher percentage of‘‘grounded’’ gaits, such as gallops and canters. Althoughsquirrel monkeys took longer and less frequent stridesthan marmosets, they also exhibited relatively longernet substrate contact periods, suggesting that increasesin stride length likely resulted from greater limb excur-sions, rather than an increase in potentially disruptivewhole-body aerial phases (Demes et al., 1990). Together,these data suggest that squirrel monkeys were usingmore compliant gait kinematics (McMahon et al., 1987;Schmitt, 1999), a finding consistent with the squirrelmonkeys’ greater exploitation of the fine-branch arborealniche. Compliant walking, characterized by increasedjoint excursion and substrate contact durations, hasbeen shown to be a hallmark of arboreal locomotion inprimates and other mammals (Larson et al., 2000, 2001;Larney and Larson, 2004), permitting greater arborealstability while simultaneously mitigating joint and boneloading (Schmitt, 1999). The role of compliance duringprimate asymmetrical locomotion has yet to be explored,but will surely prove to be an interesting area for futureresearch.Bootstrap results indicated that most of the docu-
mented species-level differences in asymmetrical gait dy-namics would have remained significant even if the twodatasets had been more similar in size and composition(Table 1). Nevertheless, low bootstrap support for someof the interaction tests (i.e., Froude number and relativepeak vertical force) indicates that these results shouldbe viewed with caution, given the larger size andreduced error variance of the squirrel monkey dataset.Future replication of the research presented here shouldincorporate a larger marmoset dataset to more fully cor-roborate the results of this study.Compared to the effects of substrate, size-related influ-
ences on asymmetrical gaits dynamics were more ambig-uous. Because the potential for arboreal instability isexacerbated as body size increases relative to substratediameter, I predicted that ontogenetic gains in bodymass would necessitate kinematic and kinetic adjust-ment when animals were traveling on the pole but haveless effect on ground locomotion. As predicted, squirrelmonkeys significantly decreased peak vertical forces assize increased during locomotion on the pole. By con-trast, marmosets maintained similar peak forces withontogenetic increases in body mass. Contradicting pre-dictions, both species showed significant size-relateddecreases in vertical COM movements when moving onthe ground, but not on the pole. As predicted, older andlarger marmosets tended to use greater duty factors,
particularly during pole locomotion, although slope dif-ferences between substrates were not statistically signifi-cant. By contrast, duty factors largely decreased withbody size in squirrel monkeys. Lead intervals and limbpair contact durations were generally unassociated withbody size in marmosets. In direct contradiction to mypredictions, older and larger squirrel monkeys increasedlead intervals and contact durations on the ground anddecreased lead intervals and contact durations on thepole. In both species, net substrate contact durations sig-nificantly increased with size to a greater extent whenmonkeys were traveling on the ground. Finally, amongmarmosets, relative stride lengths and frequenciestended to remain invariant across changes in body size,whereas larger squirrel monkeys used relatively longerand less frequent strides on both substrates, directlycontradicting predictions.In sum, specific size-related adjustments to gait pa-
rameters varied between species, with both species onlyexhibiting a few changes in the predicted direction. How-ever, it is unlikely that the observed kinematic and ki-netic changes in either species were a response to size-related instability per se. Marmosets tended to exhibitsimilar adjustments on both the ground and the pole, asindicated by pervasive nonsignificant mass-by-substrateinteractions. In squirrel monkeys, substrate-related dif-ferences in gait parameters frequently directly contra-dicted predictions, with greater kinematic and kinetic‘‘adjustments’’ to increases in body size evident duringground locomotion. It may be that within the range ofbody sizes encompassed in this study (143–346 g in mar-mosets and 218–535 g in squirrel monkeys), the biome-chanical challenges posed by a 2.5 cm diameter poleremained fairly constant. Indeed, the support size indexvaried from 3.56 to 4.78 for marmosets and from 3.08 to4.15 for squirrel monkeys (see Fig. 5), far less than therange of 2.5–10 recommended by Stevens (2003, 2007)for small to large supports. Future ontogenetic and com-parative studies, encompassing a wider array of bodysizes and support sizes, may find more consistent evi-dence for purely size-related adjustments to asymmetri-cal gait dynamics on arboreal substrates. Additionally,although beyond the scope of the current paper, futurestudies should incorporate the effects of allometricchanges in body shape on ontogenetic differences inasymmetrical gait mechanics (e.g., Raichlen, 2005)
SUMMARY AND CONCLUSIONS
Previous discussions of how primates manage arborealinstability have almost exclusively focused on symmetri-cal gait dynamics (e.g., Prost, 1969; Vilensky and Lar-son, 1989; Demes et al., 1990; Larson et al., 2001; Cart-mill et al., 2002; Schmitt and Lemelin, 2002; Stevens,2003; Larney and Larson, 2004; Schmitt et al., 2006).Nevertheless, asymmetrical gaits typify rapid locomotionin nearly all the major primate clades, from cheiroga-leids to great apes (e.g., Vilensky, 1980; Hurov, 1987;Kimura, 1992; Demes et al., 1994; Arms et al., 2002;Franz et al., 2005). Although few field studies have spe-cifically reported instances of galloping or bounding infree-ranging primates (but see Morbeck, 1976; Rose,1977; Fleagle and Mittermeier, 1980; Garber, 1991), thelacuna may be methodological: most studies of positionalbehavior lump asymmetrical gaits under the generalrubric of ‘‘quadrupedal running’’ or even simply ‘‘quadru-pedal locomotion,’’ suggesting the need for greater preci-
15ASYMMETRICAL GAIT DYNAMICS IN CALLITHRIX AND SAIMIRI
American Journal of Physical Anthropology

sion of behavioral categories (e.g., Hunt et al., 1996).Nevertheless, even if asymmetrical gaits are ratherinfrequent and contribute little to overall travel time,they may still account for a substantial proportion ofoverall travel distance, perhaps a more ecologically andphysiologically relevant metric when comparing locomo-tor behaviors (Fleagle and Mittermeier, 1980; Full, 1989;Arms et al., 2002).Several lines of evidence suggest that asymmetrical
gaits have represented an important component ofprimates’ locomotor repertoires throughout their evolu-tionary history. First, among noncursorial Eutherianmammals, asymmetrical gaits are typically used morefrequently as body size decreases (Dagg, 1973; Hilde-brand, 1977), and ancestral primates are most oftenreconstructed as small-bodied, arboreal quadrupeds(Cartmill, 1972; Martin, 1990; Gebo, 2004; but see Soligoand Martin, 2006). Second, recently discovered Paleo-cene and Eocene fossil postcrania indicate morphologicaladaptation to bounding locomotion among several Plesia-dapiforms (Bloch and Boyer, 2007), arguably the closestEuarchontan outgroup to Euprimates (Bloch and Boyer,2002; Silcox, 2007). Finally, several of the early eupri-mate traits associated with leaping, such as long anddorso-ventrally tall lumbar vertebral bodies, broadenedilia, shortened ischia, relatively long hind limbs andelongated tarsals (Gregory, 1920; Dagosto, 1993, 2007),could also have facilitated bounding and galloping.In sum, understanding the dynamics of primate asym-
metrical gaits may be critical to understanding primatelocomotor ecology and evolution in general. The resultsof this study illustrate that symmetrical gaits are notthe only stable way to travel arboreally and that small-bodied primates utilize specific kinematic and kineticadaptations that should increase stability when usingasymmetrical gaits on arboreal substrates.
ACKNOWLEDGMENTS
This work was greatly improved by comments fromBrigitte Demes, Audrone Biknevicius, William Jungers,Susan Larson, Christopher Ruff, Liza Shapiro, and twoanonymous reviewers. Animal research was carried outat two NIH-funded National Primate Research Centers,where I received generous help from many individuals,including the following: Bethany Brock, Heather Hyer,Donna Layne, Leigh Ann Long, Virginia Parks, Seth Pol-lack, Suzette Tardif, Lawrence Williams, and Cindy VanHook. Ty Hedrick provided software for kinematic analy-sis. Brigitte Demes and Daniel Riskin assisted with forceplate construction and Daniel Talley assisted with run-way construction.
LITERATURE CITED
Alexander RM, Maloiy GMO. 1984. Stride lengths and stridefrequencies of primates. J Zool (Lond) 202:577–582.
Arms A, Voges D, Fischer MS, Preuschoft H. 2002. Arboreallocomotion in small new-world primates. Z Morphol Anthropol83:243–263.
Beattie J. 1927. The anatomy of the common marmoset. ProcZool Soc (Lond) 27:593–718.
Biewener AA. 2003. Animal locomotion. Oxford: Oxford Univer-sity Press.
Biewener AA, Full RJ. 1992. Force platform and kinematicanalysis. In: Biewener AA, editor. Biomechanics: structuresand systems. Oxford: Oxford University Press. p 45–73.
Biknevicius AR, Mullineaux DR, Clayton HM. 2004. Groundreaction forces and limb function in tolting Icelandic horses.Equine Vet J 36:743–747.
Bloch JI, Boyer DM. 2002. Grasping primate origins. Science298:1606–1610.
Bloch JI, Boyer DM. 2007. New skeletons of Paleocene–EocenePlesiadapiformes: a diversity of arboreal positional behaviorsin early primates. In: Ravosa MJ, Dagosto M, editors. Primateorigins: adaptations and evolution. New York, NY: Springer.p 535–581.
Boinski S. 1989. The positional behavior and substrate use ofsquirrel monkeys: ecological implications. J Hum Evol 18:659–677.
Boinski S, Fragaszy DM. 1989. The ontogeny of foraging insquirrel monkeys, Saimiri oerstedi. Anim Behav 37:415–428.
Box GEP, Cox DR. 1964. An analysis of transformations. J RStat Soc Ser B 26:211–243.
Buikstra JE. 1975. Healed fractures in Macaca mulatta: age,sex and symmetry. Folia Primatol (Basel) 23:140–148.
Carter ML, Pontzer H, Wrangham R, Peterhans JK. 2008.Skeletal pathology in Pan troglodytes schweinfurthii inKibale National Park, Uganda. Am J Phys Anthropol 135:389–403.
Cartmill M. 1972. Arboreal adaptations and the origin of theOrder Primates. In: Tuttle R, editor. The functional and evolu-tionary biology of primates. Chicago: Aldine. p 97–122.
Cartmill M. 1979. The volar skin of primates: its frictional char-acteristics and their functional significance. Am J PhysAnthropol 50:497–510.
Cartmill M. 1985. Climbing. In: Hildebrand M, Bramble DM,Liem KF, Wake DB, editors. Functional vertebrate morphol-ogy. Cambridge: Harvard University Press. p 73–88.
Cartmill M, Lemelin P, Schmitt D. 2002. Support polygonsand symmetrical gaits in mammals. Zool J Linn Soc 136:401–420.
Cavagna GA, Heglund NC, Taylor CR. 1977. Mechanical workin terrestrial locomotion: two basic mechanisms for minimiz-ing energy expenditure. Am J Physiol 233:R243–R261.
Conover WJ, Iman RI. 1981. Rank transformations as a bridgebetween parametric and nonparametric statistics. Am Stat35:124–133.
Costello MB, Fragaszy DM. 1988. Prehension in Cebus and Sai-miri: I. Grip type and hand preference. Am J Primatol 15:235–245.
Crawley MJ. 2007. The R book. West Sussex, England: Wiley.Dagg AI. 1973. Gaits in mammals. Mammal Rev 3:135–154.Dagosto M. 1993. Postcranial anatomy and locomotor behavior
in Eocene primates. In: Gebo DL, editor. Postcranial adapta-tion in nonhuman primates. Dekalb, IL: Northern IllinoisUniversity Press. p 199–219.
Dagosto M. 2007. The postcranial morphotype of primates. In:Ravosa MJ, Dagosto M, editors. Primate origins: adaptationsand evolution. New York, NY: Springer. p 489–534.
Demes B, Jungers WL, Nieschalk U. 1990. Size- and speed-related aspects of quadrupedal walking in slender and slowlorises. In: Jouffroy FK, Stack MH, Niemetz C, editors. Grav-ity, posture and locomotion in primates. Florence: II Sedice-simo. p 175–197.
Demes B, Larson SG, Stern JT Jr, Jungers WL, Biknevicius AR,Schmitt D. 1994. The kinetics of primate quadrupedalism:‘‘hindlimb drive’’ reconsidered. J Hum Evol 26:353–374.
Elias MF. 1977. Relative maturing of cebus and squirrelmonkeys at birth and during infancy. Dev Psychobiol 10:519–528.
Fleagle J, Mittermeier RA, Skopec AL. 1981. Differential habi-tat use by Cebus apella and Saimiri sciureus in Central Suri-nam. Primates 22:361–367.
Fleagle JG, Mittermeier RA. 1980. Locomotor behavior, bodysize, and comparative ecology of seven Surinam monkeys. AmJ Phys Anthropol 52:301–314.
Fontaine R. 1990. Positional behavior in Saimiri boliviensis andAteles geoffroyi. Am J Phys Anthropol 82:485–508.
Fragaszy DM, Baer J, Adams-Curtis L. 1991. Behavioral devel-opment and maternal care in tufted capuchins (Cebus apella)
16 J.W. YOUNG
American Journal of Physical Anthropology

and squirrel monkeys (Saimiri sciureus) from birth throughseven months. Dev Psychobiol 24:375–393.
Franz TM, Demes B, Carlson KJ. 2005. Gait mechanics oflemurid primates on terrestrial and arboreal substrates. JHum Evol 48:199–217.
Full RJ. 1989. Mechanics and energetics of terrestrial locomo-tion: bipeds to polypeds. In: Wieser W, Gnaiger E, editors.Energy transformations in cells and organisms. Stuttgart:Georg Thieme. p 175–182.
Gambaryan PP. 1974. How mammals run: anatomical adapta-tions. New York, NY: Wiley.
Garber PA. 1991. A comparative study of positional behavior inthree species of tamarin monkey. Primates 32:219–230.
Garber PA. 1992. Vertical clinging, small body size, and the evo-lution of feeding adaptations in the Callitrichinae. Am J PhysAnthropol 88:469–482.
Gasc JP. 1993. Asymmetrical gait of the Saharian rodent Mer-iones shawi shawi (Duvernoy, 1842) (Rodentia, Mammalia):a high-speed cineradiographic analyses. Can J Zool 71:790–798.
Gebo DL. 2004. A shrew-sized origin for primates. Yearb PhysAnthropol 47:40–62.
Grand TI. 1977. Body weight: its relation to tissue composition,segment distribution, and motor function. I. Interspecific com-parisons. Am J Phys Anthropol 47:211–239.
Gregory WK. 1920. On the structure and relations of Notharc-tus, an American Eocene primate. Mem Am Mus Nat Hist3:51–243.
Haberman SJ. 1978. Analysis of quantitative data. London: Aca-demic Press.
Hamrick MW. 1998. Functional and adaptive significance of pri-mate pads and claws: evidence from new world anthropoids.Am J Phys Anthropol 106:113–127.
Hanna JB, Polk JD, Schmitt D. 2006. Forelimb and hindlimbforces in walking and galloping primates. Am J Phys Anthro-pol 130:529–535.
Hedrick T. 2007. ‘‘DLT Data Viewer 2’’, Digitizing and DLT inMATLAB. http://www.unc.edu/�thedrick/software1.html.
Heglund NC. 1981. A simple design for a force-plate to measureground reaction forces. J Exp Biol 93:333–338.
Heglund NC, Cavagna GA, Taylor CR. 1982. Energetics andmechanics of terrestrial locomotion. III. Energy changes ofthe centre of mass as a function of speed and body size inbirds and mammals. J Exp Biol 97:41–56.
Heglund NC, Taylor CR, McMahon TA. 1974. Scaling stride fre-quency and gait to animal size: mice to horses. Science186:1112–1113.
Hildebrand M. 1966. Analysis of the symmetrical gaits of tetra-pods. Folia Biotheor 1–22.
Hildebrand M. 1977. Analysis of asymmetrical gaits. J Mammal58:131–156.
Hof AL. 1996. Scaling gait data to body size. Gait Posture4:222–223.
Horovitz I, Zardoya R, Meyer A. 1998. A Platyrrhine system-atics: a simultaneous analysis of molecular and morphologicaldata. Am J Phys Anthropol 106:261–281.
Howell AB. 1944. Speed in animals. New York: Hafner.Huitema BE. 1980. The analysis of covariance and its alterna-
tives. New York, NY: Wiley.Hunt KD, Cant JGH, Gebo DL, Rose MD, Walker SE, Youlatos
D. 1996. Standardized descriptions of primate locomotor andpostural modes. Primates 37:363–387.
Hurov JR. 1987. Terrestrial locomotion and back anatomy invervets (Cercopithecus aethiops) and patas monkeys (Erythro-cebus patas). Am J Primatol 13:297–311.
Janson CH, Boinski S. 1992. Morphological and behavioraladaptations for foraging in generalist primates: the case ofcebines. Am J Phys Anthropol 88:483–498.
Jurmain R. 1997. Skeletal evidence of trauma in African apes,with special reference to the Gombe chimpanzees. Primates38:1–14.
Kaack B, Walker L, Brizzee KR. 1979. The growth and develop-ment of the squirrel monkey (Saimiri sciureus). Growth43:116–135.
Kimura T. 1992. Hindlimb dominance during primate high-speed locomotion. Primates 33:465–476.
Kirk EC, Lemelin P, Hamrick MW, Boyer DM, Bloch JI. 2008.Intrinsic hand proportions of euarchontans and other mam-mals: implications for the locomotor behavior of plesiadapi-forms. J Hum Evol 55:278–299.
Larney E, Larson SG. 2004. Compliant walking in primates:elbow and knee yield in primates compared to other mam-mals. Am J Phys Anthropol 125:42–50.
Larson SG, Schmitt D, Lemelin P, Hamrick M. 2000. Unique-ness of primate forelimb posture during quadrupedal locomo-tion. Am J Phys Anthropol 112:87–101.
Larson SG, Schmitt D, Lemelin P, Hamrick M. 2001. Limbexcursion during quadrupedal walking: how do primates com-pare to other mammals? J Zool (Lond) 255:353–365.
Larson SG, Stern JT. 2006. Maintenance of above-branch bal-ance during primate arboreal quadrupedalism: coordinateduse of forearm rotators and tail motion. Am J Phys Anthropol129:71–81.
Lemelin P. 1999. Morphological correlates of substrate use indidelphid marsupials: implications for primate origins. J Zool(Lond) 247:165–175.
Lemelin P, Grafton BW. 1998. Grasping performance in Sagui-nas midas and the evolution of hand prehensility in primates.In: Strasser E, Fleagle J, Rosenberger A, McHenry H, editors.Primate locomotion: recent advances. New York, NY: PlenumPress. p 131–144.
Lovell NC. 1991. An evolutionary framework for assessing ill-ness and injury in nonhuman primates. Yearb Phys Anthropol34:117–155.
Martin RD. 1990. Primate origins and evolution: a phylogeneticreconstruction. Princeton: Princeton University Press.
McMahon TA, Valiant G, Frederick EC. 1987. Groucho running.J Appl Physiol 62:2326–2337.
Midlo C. 1934. Form of hand and foot in primates. Am J PhysAnthropol 19:337–389.
Minetti AE, Ardigo LP, Reinach E, Saibene F. 1999. The rela-tionship between mechanical work and energy expenditure oflocomotion in horses. J Exp Biol 202:2329–2338.
Missler M, Wolff JR, Rothe H, Heger W, Merker H-J, Treiber A,Scheid R, Crook GA. 1992. Developmental biology of the com-mon marmoset: proposal for a ‘‘postnatal staging’’. J Med Pri-matol 21:285–298.
Morbeck ME. 1976. Leaping, bounding and bipedalism in Colo-bus guereza: a spectrum of positional behaviors. Yearb PhysAnthropol 20:408–420.
Napier JR. 1967. Evolutionary aspects of primate locomotion.Am J Phys Anthropol 27:333–342.
Opazo J, Wildman D, Prychitko T, Johnson R, Goodman M.2006. Phylogenetic relationships and divergence times amongNew World monkeys (Platyrrhini, Primates). Mol PhylogenetEvol 40:274–280.
Prost JH. 1969. A replication study on monkey gaits. Am JPhys Anthropol 30:203–208.
Quinn GP, Keough MJ. 2002. Experimental design and data anal-ysis for biologists. Cambridge, UK: Cambridge University Press.
R Development Core Team. 2008. R: a language and environ-ment for statistical computing. Vienna, Austria: R Foundationfor Statistical Computing.
Raichlen DA. 2005. Effects of limb mass distribution on the on-togeny of quadrupedalism in infant baboons (Papio cynocepha-lus) and implications for the evolution of primate quadruped-alism. J Hum Evol 49:415–431.
Rasmussen DT. 1990. Primate origins: lessons from a neotropi-cal marsupial. Am J Primatol 22:263–277.
Reynolds TR. 1987. Stride length and its determinants inhumans, early hominids, primates, and mammals. Am J PhysAnthropol 72:101–115.
Riskin DK, Bertram JEA, Hermanson JW. 2005. Testing thehindlimb-strength hypothesis: non-aerial locomotion by Chi-roptera is not constrained by the dimensions of the femur ortibia. J Exp Biol 208:1309–1319.
Rose MD. 1973. Quadrupedalism in primates. Primates 14:337–357.
17ASYMMETRICAL GAIT DYNAMICS IN CALLITHRIX AND SAIMIRI
American Journal of Physical Anthropology

Rose MD. 1977. Positional behavior of olive baboons (Papio anu-bis) and its relationship to maintenance and social activities.Primates 18:59–116.
Rosenberger A, Stafford BJ. 1994. Locomotion in captive Leonto-pithecus and Callimico: a multimedia study. Am J PhysAnthropol 94:379–394.
Schmitt D. 1999. Compliant walking in primates. J Zool (Lond)248:149–160.
Schmitt D. 2003a. Evolutionary implications of the unusualwalking mechanics of the common marmoset (C. jacchus). AmJ Phys Anthropol 122:28–37.
Schmitt D. 2003b. Substrate size and primate forelimb mechan-ics: implications for understanding the evolution of primatelocomotion. Int J Primatol 24:1023–1036.
Schmitt D, Cartmill M, Griffin T, M., Hanna JB, Lemelin P.2006. Adaptive value of ambling gaits in primates and othermammals. J Exp Biol 209:2042–2049.
Schmitt D, Lemelin P. 2002. Origins of primate locomotion: gaitmechanics of the woolly opossum. Am J Phys Anthropol 118:231–238.
Schultz AH. 1944. Age changes and variability in gibbons. Amorphological study on a population sample of a man-likeape. Am J Phys Anthropol 2:1–129.
Silcox MT. 2007. Primate taxonomy, Plesiadapiforms, andapproaches to primate origins. In: Ravosa MJ, Dagosto M,editors. Primate origins: adaptations and evolution. NewYork, NY: Springer. p 143–178.
Smith RJ, Jungers WL. 1997. Body mass in comparative prima-tology. J Hum Evol 32:523–559.
Sokal RR, Rohlf FJ. 1995. Biometry. New York: W.H. Free-man.
Soligo C, Martin RD. 2006. Adaptive origins of primates revis-ited. J Hum Evol 50:414–430.
Stevens J. 2002. Applied multivariate statistics for the socialsciences. Mahwah, NJ: Lawrence Erlbaum.
Stevens NJ. 2003. The influence of substrate size, orientationand compliance upon prosimian arboreal quadrupedalism.Ph.D. dissertation, Stony Brook University, State Universityof New York at Stony Brook.
Stevens NJ. 2007. The effect of branch diameter on primate gaitsequence pattern. Am J Primatol 70:1–7.
Sukhanov VB. 1974. General system of symmetrical locomotionof terrestrial vertebrates and some features of movement oflower tetrapods. New Delhi: Amerind.
Sussman RW. 1991. Primate origins and the evolution of angio-sperms. Am J Primatol 23:209–223.
Sussman RW, Kinzey WG. 1984. The ecological role of the Calli-trichidae: a review. Am J Phys Anthropol 64:419–449.
Vilensky JA. 1980. Trot-gallop transition in a macaque. Am JPhys Anthropol 53:347–348.
Vilensky JA, Larson SG. 1989. Primate locomotion: utilization andcontrol of symmetrical gaits. Annu Rev Anthropol 18:17–35.
Vilensky JA, Moore-Kuhns M, Moore AM. 1990. Angular dis-placement patterns of leading and trailing limb joints duringgalloping in monkeys. Am J Primatol 22:227–239.
Vinyard C, Schmitt D. 2004. New technique for studying reac-tion forces during primate behaviors on vertical substrates.Am J Phys Anthropol 125:343–351.
Walter RM, Carrier DR. 2007. Ground forces applied by gallop-ing dogs. J Exp Biol 230:208–216.
Winter DA. 2005. Biomechanics and motor control of humanmovement. Hoboken, NJ: John Wiley & Sons, Inc.
18 J.W. YOUNG
American Journal of Physical Anthropology