structure and resilience of a tidepool fish assemblage at barbados
TRANSCRIPT

Environmental Biology of Fishes 41: 171-190,1994. 0 1994 Kluwer Academic Publishers. Printed in the Netherlands.
Structure and resilience of a tidepool fish assemblage at Barbados
Robin Mahon’ & Susan D. Mahon2 ’ Fisheries and Environmental Consulting, 48 Sunset Crest, St. James, Barbados ’ Environment, People and Information, Durants Ridge No. 3, St. James, Barbados
Received 19.10.1993 Accepted 1.3.1994
Key words: Caribbean, Community structure, Coral reef fishes, Nurseries, Stability, Recolonisation
Synopsis
Fish collections from 19 tidepools on a rock plateau at Martins Bay, on the east coast of Barbados, taken on three occasions (1981,1983 and 1987) contained 2078 individuals of 63 species. The number of species, individ- uals and total biomass increased with pool size. Partial residents, primarily juveniles of reef species, comprised 44% of species, 36% of numbers, and 26% of biomass. True and partial residents were of similar sizes. Most of the latter grow to larger sizes than those observed in the pools, indicating that the use of tidepools by fishes is size-dependent. Species richness, numbers of individuals and biomass in individual pools was positively asso- ciated with pool size. These relationships did not vary among sampling occasions. Species composition and relative abundance was also found to be similar among sampling occasions, leading to the conclusion that the tidepool assemblages are resilient and stable.
Introduction
Intertidal habitats have generally been viewed as ecotones where marine organisms encounter harsh and highly variable physical conditions, such as wave action, periodic desiccation, high temper- atures, and are also vulnerable to predation (e.g. Nybakken 1982). In the intertidal zone, tidepools provide a refuge for marine organisms which be- come concentrated in them during periods of tidal exposure. Owing to easy access, the intertidal zone has been intensively studied. Although these stud- ies have included fishes, tropical tidepools have re- ceived little attention (Chadwick 1976, Gibson 1982).
In studies of tidepools in temperate areas, local distribution and abundance of fishes appear to de- pend largely on the area and the depth of the pool, its degree of isolation from the sea and exposure to
wave action (Gibson 1969,1972, Green 1971, Yoshi- yama 1981, Bennett & Griffiths 1984). Size and the degree of isolation of pools determine the extent of stressful fluctuations such as in temperature and sa- linity. Isolation is determined primarily by the height of tidepools above mean low water.
A common feature of tidepool fish assemblages is the presence of two main groups of fishes: true residents, and partial residents (Gibson 1969). True residents are generally small, benthic fishes such as the blennies and gobies. Partial residents are pri- marily sublittoral, but occur in tidepools, particu- larly as juveniles. This pattern is supported by sev- eral studies which have used slight variations of these categories (Thompson & Lehner 1976, Chang et al. 1977, Lee 1980a, 198Ob, Grossman 1982). A third group, transients, usually comprises a small proportion of species in tidepools, and is sometimes included with the partial residents.

We investigated a tidepool fish assemblage from the east coast of Barbados, in the western tropical Atlantic, with the objectives of describing the spe- cies composition, distribution and abundance of fishes in tidepools of various sizes. We sampled the assemblage on three occasions at the same time in different years to evaluate the extent to which the observed characteristics are persistent through time.
The study area
The study area is located at Martins Bay on the east coast of Barbados. Barbados lies about 150 km east of the Lesser Antilles island arc in the eastern Ca- ribbean. A general description of rocky-shore fau- nas and habitats of Barbados is provided by Lewis (1960). Since the prevailing winds and currents are easterly, the coast on which the study area is located is exposed to heavy wave action. At many places there are eroded limestone plateaus on which tide- pools occur. These plateaus are exposed at low tide, isolating the pools. In contrast the south and west coasts are primarily sandy beaches with intermit- tent low limestone cliffs and sublittoral fringing reefs. On these sheltered coasts tidepools are scarce. Studies of rocky-shore zonation in the Ca- ribbean have not resulted in a consistent classifica- tion (John & Price 1979, Brattstrom 1980). How- ever, the slate pencil urchin, Echinometra lucunter and Sargassum spp. consistently characterise the low extreme of regular tidal exposure.
At Barbados, tides are mixed, semidiurnal with no marked inequality in amplitude (Lewis 1960). There is an amphidromic point near Barbados and consequently tidal amplitude is low (King 1975). Over an 18 month period, mean tidal range and di- urnal range were 0.7 and 1.1 m, respectively (Lewis 1960).
At Martins Bay the limestone plateau is protect- ed from the full force of the waves by headlands and by an offshore reef. The plateau is about 50 m in width at the widest point, and about 200 m in length along the shore. Shoreward it is bounded by a beach. At the seaward edge, where it drops off, the plateau is honeycombed with burrows of Echino-
metra lucunter; the surf zone of Lewis (1960). The surface of the plateau is covered with a mat of algae and dotted with depressions which become pools at low tide. The sides of the pools are usually vertical with holes and undercuts. In the pools a variety of substrates may occur: bare sand, sand with turtle grass, Thalassia testudinum, rock covered with al- gae, and limestone rubble.
The incoming tide floods the entire plateau in a period of about 30 minutes. Thus there is little vari- ation in isolation among pools. This is in contrast to areas studied by Green (1971), Gibson (1972), and Yoshiyama (1981) where tidal range was high and pool isolation varied considerably with elevation.
Methods
The fishes were collected at low tide from 19 tide- pools at Martins Bay at about the same time of year in each of three years: 17-19 June 1981,9 June 1983, 13 June 1987. Each pool was treated with an ich- thyocide, ProNoxfish, in which the active ingredient is rotenone, and was searched until no more fishes could be found. Most fishes succumbed within 5-10 minutes but some eels took up to an hour to emerge from hiding. The fishes were preserved in 4% for- maldehyde and identified using keys in Bohlke & Chaplin (1968) and Fischer (1978). Identifications were checked at the Royal Ontario Museum, To- ronto where the fishes have been deposited.
The length (L), width (W), and maximum depth (D) of each pool was measured and pool area (A = LxW) and volume (V = A x 0.5 D) were estimated. The percent of each pool covered by each substrate type - sand, sand with turtle grass, Thalassia testudi- num, rock covered with algae, and limestone rubble -was estimated by eye. Habitat diversity was esti- mated using Simpson's index, D = Cpf, where pi is the probability of encountering the ith habitat type or the proportion of that habitat type (Southwood 1978).
Each species of fish was designated as either a true or partial resident on the basis of information given by BGhlke & Chaplin (1968), Randall (1968), Fischer (1978) and references cited therein. True residents were those which could complete their
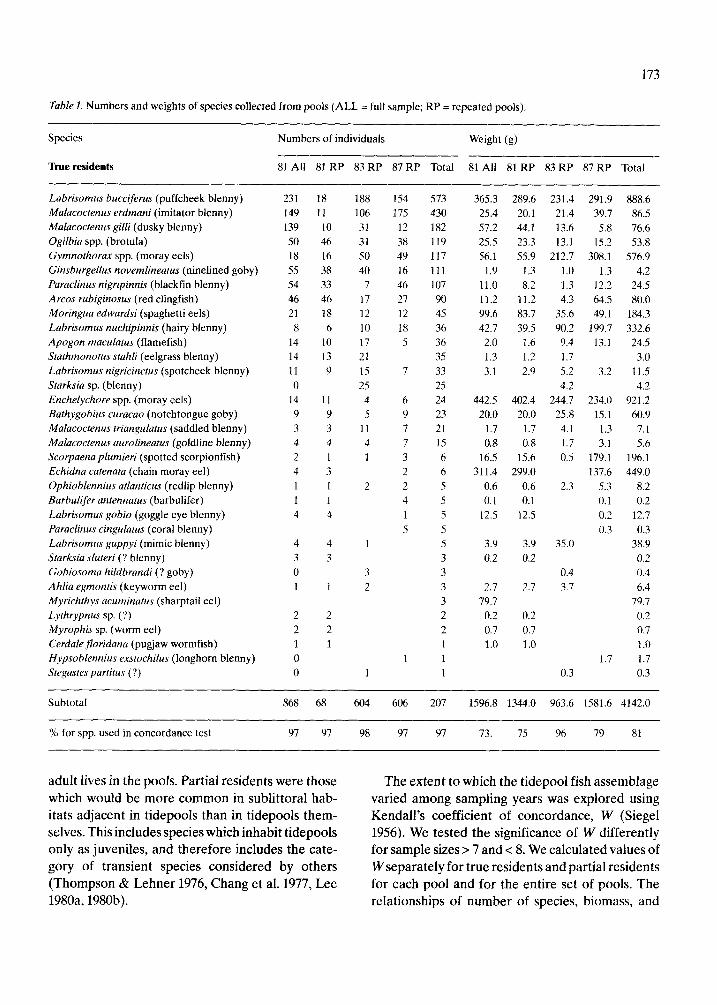
Tablel. Numbers and weights of species collected from pools (ALL =full sample; RP = repeated pools).
Species Numbers of individuals Weight (g)
ltue residents 81 All 81 RP 83 RP 87 RP Total 81 A11 81 RP 83 RP 87 RP Total
Labrisomus bucciferus (puffcheek blenny) Malacoctenus erdmani (imitator blenny) Malacoctenus gilli (dusky blenny) Ogilbia spp. (brotula) Gymnothorax spp. (moray eels) Ginsburgellus novemlineatus (ninelined goby) Paraclinus nigripinnis (blackfin blenny) Arcos rubiginosus (red clingfish) Moringua edwardsi (spaghetti eels) Labrisomus nuchipinnis (hairy blenny) Apogon maculatus (flamefish) Stathmonotus srahli (eelgrass blenny) Labrisomus nigricincrus (spotcheek blenny) Starksia sp. (blenny) Enchelychore spp. (moray eels) Barhygobius curacao (notchtongue goby) Malacoctenus triangulatus (saddled blenny) Malacoctenus aurolineatus (goldline blenny) Scorpaena plumieri (spotted scorpionfish) Echidna catenata (chain moray eel) Ophioblennius atlanticus (redlip blenny) Barbulifer antennatus (barbulifer) Labrisomus gobio (goggle eye blenny) Paraclinu~ cingulatus (coral blenny) Labrisomus guppyi (mimic blenny) Starksia sluteri (? blenny) Gobiosoma hddbrandi (? goby) Ahlia egmontis (keyworm eel) Myrichthys ucuminarus (sharptail eel) Lythrypnus sp. (?) Myrophis sp. (worm eel) Cerdalefloridana (pugjaw wormfish) Hypsoblennius exstochilus (longhorn blenny) Stegastes parfitus (?)
Subtotal 868 68 604 606 207 1596.8 1344.0 963.6 1581.6 4142.0
% for spp. used in concordance test 97 97 98 97 97 73. 75 96 79 81
adult lives in the pools. Partial residents were those which would be more common in sublittoral hab- itats adjacent in tidepools than in tidepools them- selves. This includes species which inhabit tidepools only as juveniles, and therefore includes the cate- gory of transient species considered by others (Thompson & Lehner 1976, Chang et al. 1977, Lee 1980a, 1980b).
The extent to which the tidepool fish assemblage varied among sampling years was explored using Kendall's coefficient of concordance, W (Siege1 1956). We tested the significance of W differently for sample sizes > 7 and < 8. We calculated values of Wseparately for true residents and partial residents for each pool and for the entire set of pools. The relationships of number of species, biomass, and
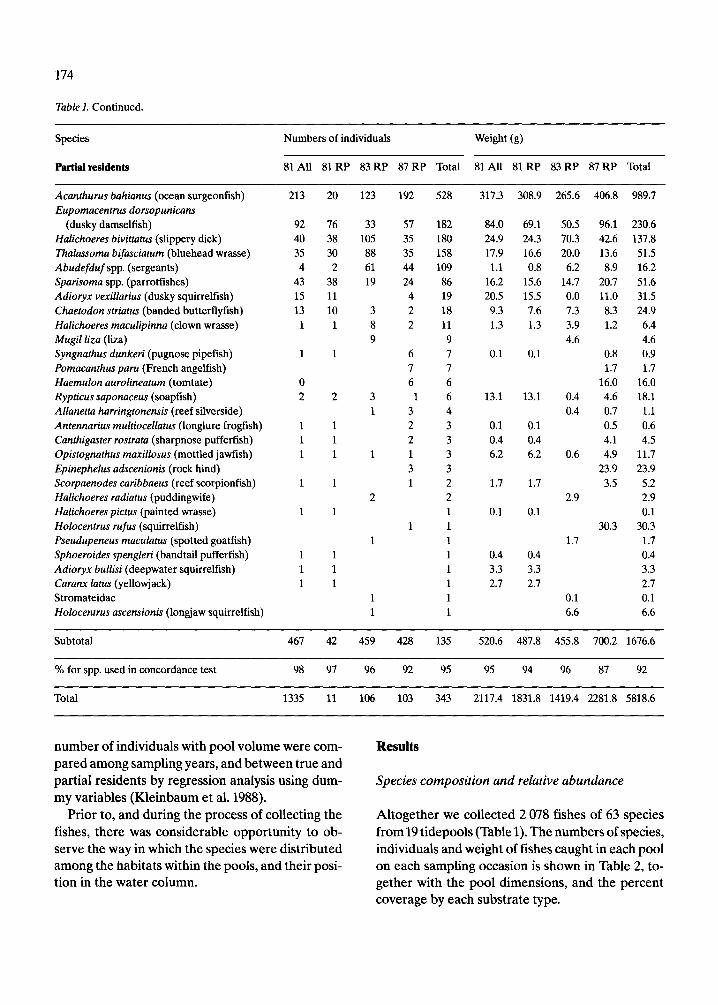
Table 1. Continued.
Species Numbers of individuals Weight (g)
Partial residents 81 All 81 RP 83 RP 87 RP Total 81 All 81 RP 83 RP 87 RP Total
Acanthurus bahianus (ocean surgeonfish) Eupornacentrus dorsopunicans
(dusky damselfish) Halichoeres bivittatus (slippery dick) Thalassoma bifmciatum (bluehead wrasse) Abudefduf spp. (sergeants) Sparisoma spp. (parrotfishes) Adioryx vexillarius (dusky squirrelfish) Chaetodon striatus (banded butterflyfish) Halichoeres maculipinna (clown wrasse) Mugil liza (ha) Syngnathus dunkeri (pugnose pipefish) Pomacanthus paru (French angelfish) Haemulon aurolineaturn (tomtate) Rypticus saponaceus (soapfish) Allanetta harringtonensis (reef silverside) Antennarius rnultiocellatus (longlure frogfish) Canthigaster rostrata (sharpnose pufferfish) Opistognathus maxillosus (mottled jawfish) Epinephelus adscenionis (rock hind) Scorpaenodes caribbaeus (reef scorpionfish) Halichoeres radiatus (puddingwife) Halichoeres pictus (painted wrasse) Holocentrus rufus (squirrelfish) Pseudupeneus maculatus (spotted goatfish) Sphoeroides spengleri (bandtail pufferfish) Adioryx bullisi (deepwater squirrelfish) Caranx latus (yellowjack) Stromateidae Holocentrus ascensionis (longjaw squirrelfish)
Subtotal 467 42 459 428 135 520.6 487.8 455.8 700.2 1676.6
% for spp. used in concordance test 98 97 96 92 95 95 94 96 87 92
Total 1335 11 106 103 343 2117.4 1831.8 1419.4 2281.8 5818.6
number of individuals with pool volume were com- pared among sampling years, and between true and partial residents by regression analysis using dum- my variables (Kleinbaum et al. 1988).
Prior to, and during the process of collecting the fishes, there was considerable opportunity to ob- serve the way in which the species were distributed among the habitats within the pools, and their posi- tion in the water column.
Results
Species composition and relative abundance
Altogether we collected 2 078 fishes of 63 species from19 tidepools (Table 1). The numbers of species, individuals and weight of fishes caught in each pool on each sampling occasion is shown in Table 2, to- gether with the pool dimensions, and the percent coverage by each substrate type.
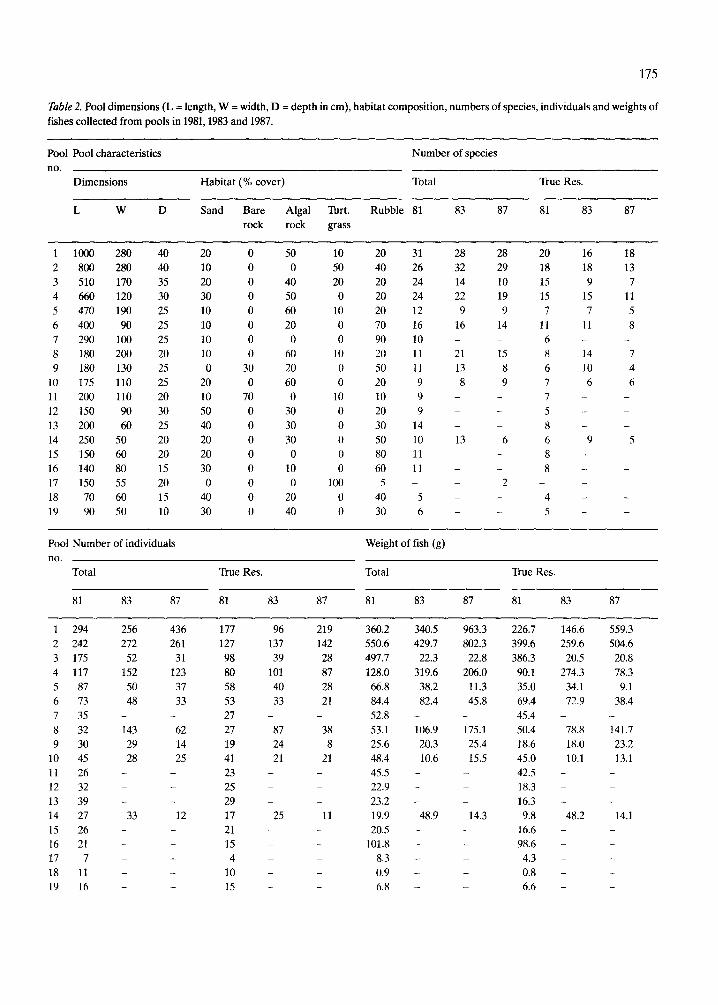
Table 2. Pool dimensions (L = length, W = width, D = depth in cm), habitat composition, numbers of species, individuals and weights of fishes collected from pools in 1981,1983 and 1987.
Pool Pool characteristics Number of species no.
Dimensions Habitat (% cover) Total True Res.
L W D Sand Bare Algal Turt. Rubble 81 83 87 81 83 87 rock rock grass
Pool Number of individuals Weight of fish (g) no.
Total True Res. Total True Res.
81 83 87 81 83 87 81 83 87 81 83 87

Fig. 1. The distribution of fishes in a typical tidepool in cross-section: 1 -eel, 2 -turtle grass, 3 -sand deposit, 4 - spaghetti eels, 5 -partial resident juvenile reef fishes, 6 - Echinometra Iucunrer in burrows which are inhabited by nineline goby and red clingfish, 7 - cryptic/ benthic true resident.
As observed for tidepool fish assemblages in other parts of the world, true and partial residents both contribute substantially to the numbers of spe- cies and individuals in our collections. The most common partial residents were juveniles of reef species e.g. dusky damselfish, parrotfishes, ocean surgeonfish and banded butterflyfish. These fishes are usually nektonic and conspicuous. The most common true residents were benthic, cryptic spe- cies such as the puffcheek, imitator and dusky blen- nies (Table 1).
Within the category of true residents, there were at least five species of eels, which were treated sep- arately in subsequent analyses. The uncertainty in the identity of the genera Gymnothoru~ and Echy- lychore pertains to the small specimens. In both cases, the larger individuals belonged to a single species, G. moringa and E. nigricans, respectively, and it is likely that the juveniles were predominant- ly these species.
Observations on the distribution of fishes in pools
The common partial resident juveniles swam in small schools in the middle of the pools (ocean sur- geonfish, sergeant majors) or singly at the edges un- der rock overhangs (wrasses, dusky damselfish) (Fig. 1). These species are easily visible to the casual observer. Most true resident species are distributed singly in the shallow pool margins, or on rocks and
rubble in the pools and are extremely difficult to see (blennies and gobies). This is partly because of their cryptic coloration, but also because they seek cover in crevices, holes, under ledges, and in algae very rapidly when they detect the slightest movement outside the pool. Other true resident species have very specific microhabitats within the pools. For ex- ample, spaghetti eels occur only in pools with sand deposits (Tables 1,2), red clingfish and nineline go- bies occur only in association with Echinometra lu- cunter, as described by Teytaud (1971), and moray eels inhabit holes in the sides of the pools.
Variation in assemblage composition among years
For the entire set of pools, Kendall’s W was signif- icant at the p = 0.01 and p = 0.05 levels for the true residents and partial residents respectively (Table 3). For the former, Wwas significant at the 0.05 level in four pools, whereas for the latter, W was signif- icant in only two pools. In both instances the signif- icant concordance was found in the largest pools.
The effect of pool size on assemblage similarity between pairs of samples was explored by plotting the probabilities of the Spearman rank correlation coefficients (Table 3) versus pool size (Fig. 2). The figure shows that the probability of occurrence of the calculated rs value decreases markedly with in- creasing pool size. This indicates that assemblage similarity increases with pool size.
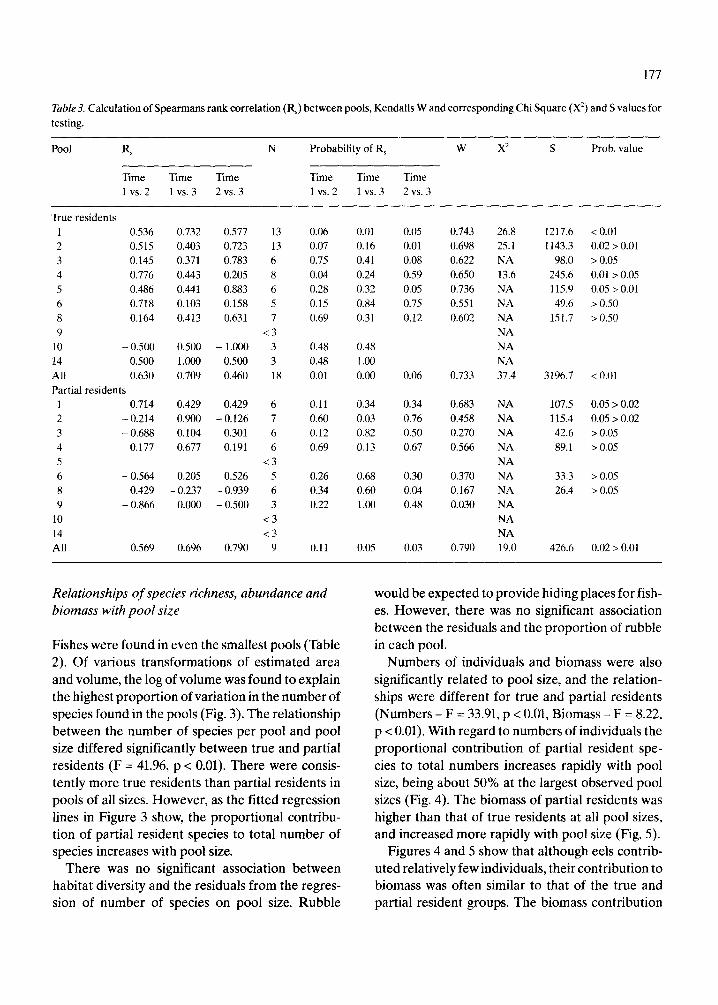
Table3. Calculation of Spearmans rank correlation (RJ between pools, Kendalls Wand corresponding Chi Square (x2) and S values for testing.
Pool Rs N Probability of R, W X2 S Prob. value
Time Time Time Time Time Time 1 vs. 2 1 vs. 3 2 vs. 3 1 vs. 2 1 vs. 3 2 vs. 3
True residents 1 0.536 2 0.515 3 0.145 4 0.776 5 0.486 6 0.718 8 0.164 9
10 - 0.500 14 0.500 All 0.630 Partial residents
1 0.714 2 - 0.214 3 - 0.688 4 0.177 5 6 - 0.564 8 0.429 9 - 0.866
10 14 All 0.569
Relationships of species richness, abundance and biomass with pool size
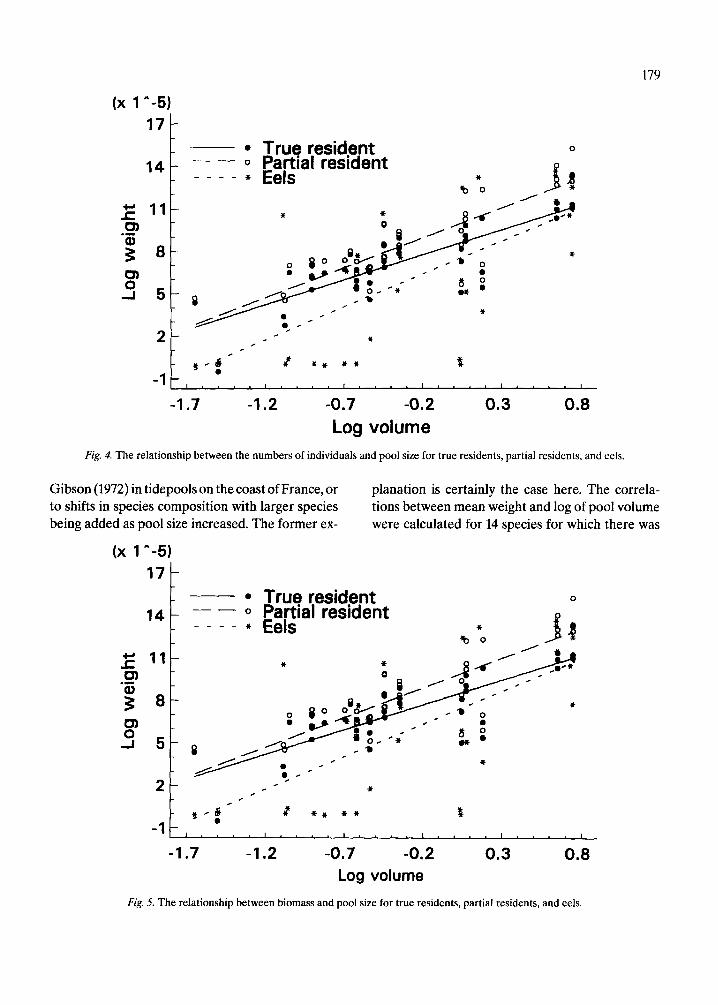
Fishes were found in even the smallest pools (Table 2). Of various transformations of estimated area and volume, the log of volume was found to explain the highest proportion of variation in the number of species found in the pools (Fig. 3). The relationship between the number of species per pool and pool size differed significantly between true and partial residents (F = 41.96, p < 0.01). There were consis- tently more true residents than partial residents in pools of all sizes. However, as the fitted regression lines in Figure 3 show, the proportional contribu- tion of partial resident species to total number of species increases with pool size.
There was no significant association between habitat diversity and the residuals from the regres- sion of number of species on pool size. Rubble
would be expected to provide hiding places for fish- es. However, there was no significant association between the residuals and the proportion of rubble in each pool.
Numbers of individuals and biomass were also significantly related to pool size, and the relation- ships were different for true and partial residents (Numbers - F = 33.91, p < 0.01, Biomass - F = 8.22, p < 0.01). With regard to numbers of individuals the proportional contribution of partial resident spe- cies to total numbers increases rapidly with pool size, being about 50% at the largest observed pool sizes (Fig. 4). The biomass of partial residents was higher than that of true residents at all pool sizes, and increased more rapidly with pool size (Fig. 5).
Figures 4 and 5 show that although eels contrib- uted relatively few individuals, their contribution to biomass was often similar to that of the true and partial resident groups. The biomass contribution
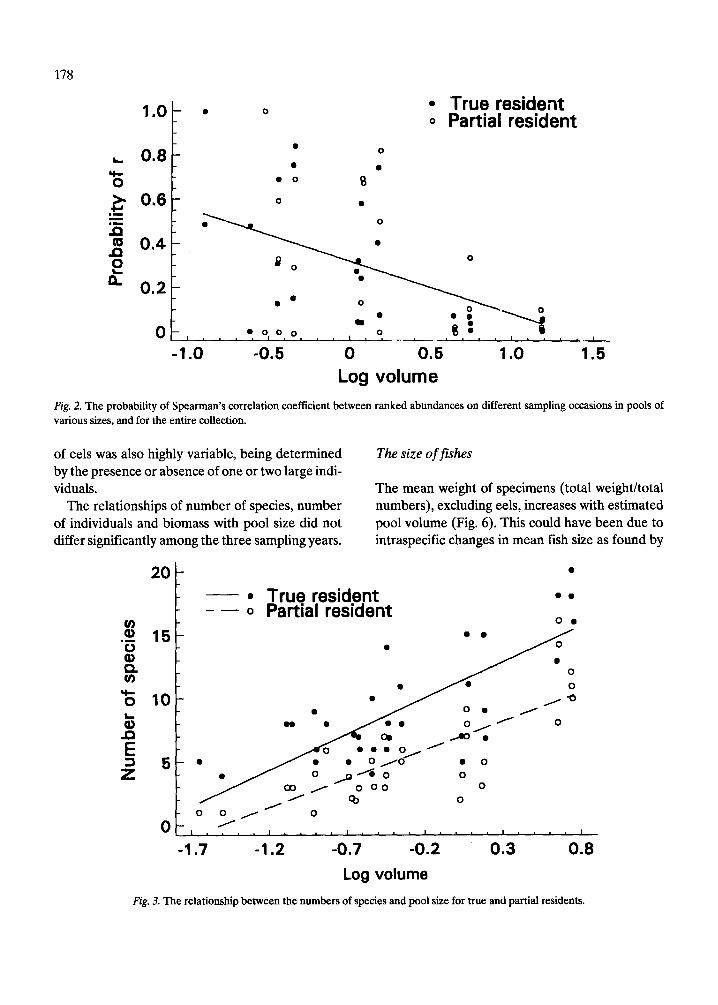
True resident 0 Partial resident
Log volume Fig. 2. The probability of Spearman's correlation coefficient between ranked abundances on different sampling occasions in pools of various sues, and for the entire collection.
of eels was also highly variable, being determined The size of fishes by the presence or absence of one or two large indi- viduals. The mean weight of specimens (total weightltotal
The relationships of number of species, number numbers), excluding eels, increases with estimated of individuals and biomass with pool size did not pool volume (Fig. 6). This could have been due to differ significantly among the three sampling years. intraspecific changes in mean fish size as found by
True resident - - 0 Partial resident
Log volume
Fig. 3. The relationship between the numbers of species and pool size for true and partial residents.
True resident O -- - - - -
Partial resident * Eels it
rb 0
Log volume Fig. 4. The relationship between the numbers of individuals and pool size for true residents, partial residents, and eels.
Gibson (1972) in tidepools on the coast of France, or planation is certainly the case here. The correla- to shifts in species composition with larger species tions between mean weight and log of pool volume being added as pool size increased. The former ex- were calculated for 14 species for which there was
Log volume
1 4 -
Fig. 5. The relationship between biomass and pool size for true residents, partial residents, and eels.
True resident O -- - - - -
Partial resident * Eels it
Jb 0
3 11- If
i t
Y
*
* it
-1
. 8 .
- t - i P * * x * 15 - , , , , , , , , , , , , , , , , , , , , , I
-1.7 -1.2 -0.7 -0.2 0.3 0.8
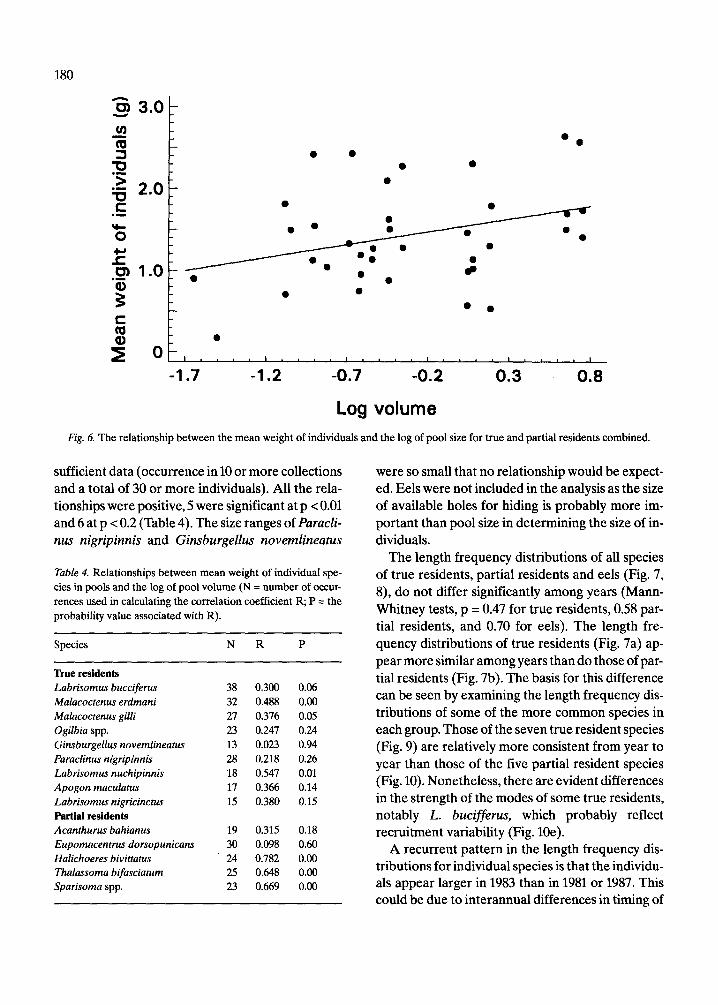
Log volume Fig. 6. The relationship between the mean weight of individuals and the log of pool size for true and partial residents combined.
sufficient data (occurrence in 10 or more collections and a total of 30 or more individuals). All the rela- tionships were positive, 5 were significant at p < 0.01 and 6 at p < 0.2 (Table 4). The size ranges of Paracli- nus nigripinnis and Ginsburgellus novemlineatus
Table 4. Relationships between mean weight of individual spe- cies in pools and the log of pool volume (N = number of occur- rences used in calculating the correlation coefficient R, P = the probability value associated with R).
Species N R P
' h e residents Labrisomus bucciferus Malacoctenus erdmani Malacoctenus gilli Ogilbia spp. Ginsburgellus novemlineatus Paraclinus nigripinnis Labrisomus nuchipinnis Apogon maculatus Labrisomus nigricinctus Partial residents Acanthurus bahianus Eupomacentrus dorsopunicans Halichoeres bivittatus Thalassoma bifasciatum Sparisoma spp.
were so small that no relationship would be expect- ed. Eels were not included in the analysis as the size of available holes for hiding is probably more im- portant than pool size in determining the size of in- dividuals.
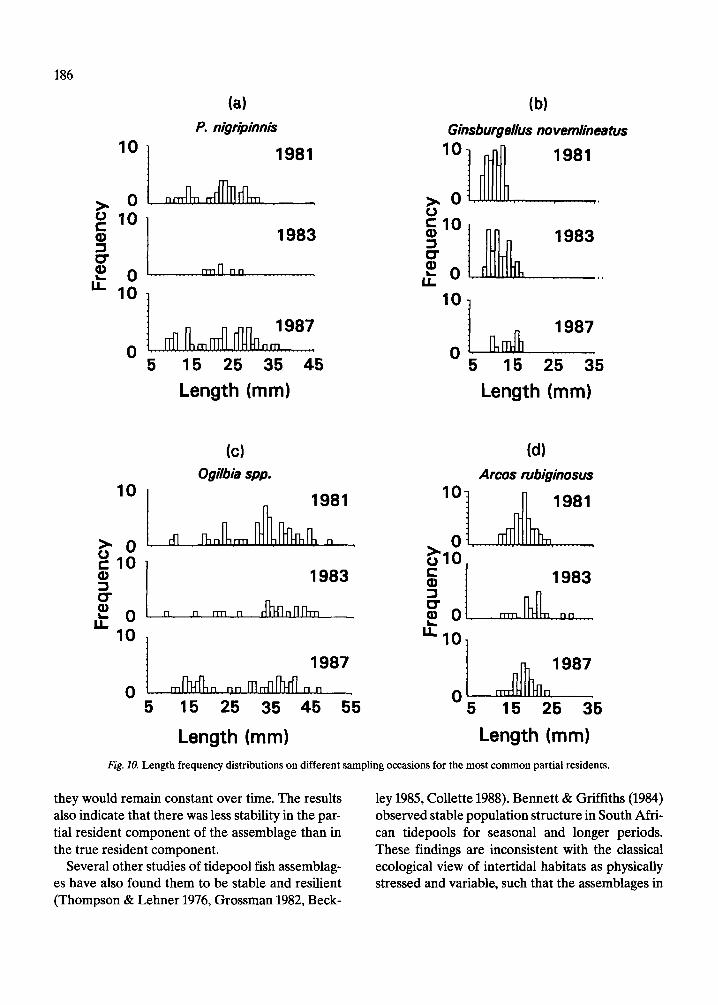
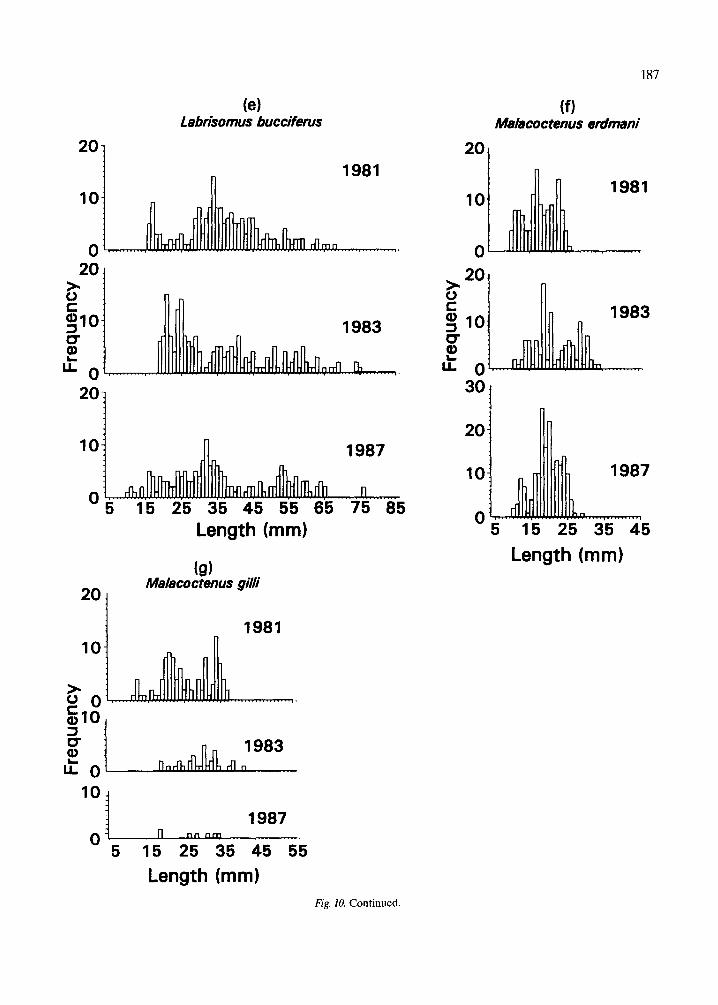
The length frequency distributions of all species of true residents, partial residents and eels (Fig. 7, 8), do not differ significantly among years (Mann- Whitney tests, p = 0.47 for true residents, 0.58 par- tial residents, and 0.70 for eels). The length fre- quency distributions of true residents (Fig. 7a) ap- pear more similar among years than do those of par- tial residents (Fig. 7b). The basis for this difference can be seen by examining the length frequency dis- tributions of some of the more common species in each group. Those of the seven true resident species (Fig. 9) are relatively more consistent from year to year than those of the five partial resident species (Fig. 10). Nonetheless, there are evident differences in the strength of the modes of some true residents, notably L. bucifferus, which probably reflect recruitment variability (Fig. 10e).
A recurrent pattern in the length frequency dis- tributions for individual species is that the individu- als appear larger in 1983 than in 1981 or 1987. This could be due to interannual differences in timing of
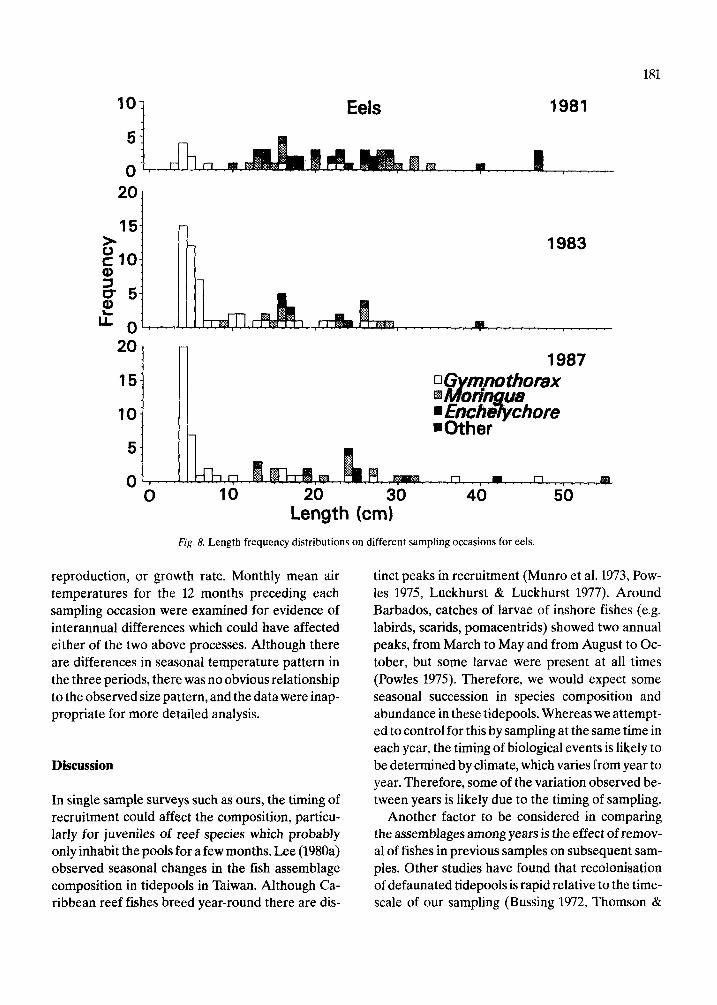
Eels
Length (cm) Fig. 8. Length frequency distributions on different sampling occasions for eels.
reproduction, or growth rate. Monthly mean air temperatures for the 12 months preceding each sampling occasion were examined for evidence of interannual differences which could have affected either of the two above processes. Although there are differences in seasonal temperature pattern in the three periods, there was no obvious relationship to the observed size pattern, and the data were inap- propriate for more detailed analysis.
Discussion
In single sample surveys such as ours, the timing of recruitment could affect the composition, particu- larly for juveniles of reef species which probably only inhabit the pools for a few months. Lee (1980a) observed seasonal changes in the fish assemblage composition in tidepools in Taiwan. Although Ca- ribbean reef fishes breed year-round there are dis-
tinct peaks in recruitment (Munro et al. 1973, Pow- les 1975, Luckhurst & Luckhurst 1977). Around Barbados, catches of larvae of inshore fishes (e.g. labirds, scarids, pomacentrids) showed two annual peaks, from March to May and from August to Oc- tober, but some larvae were present at all times (Powles 1975). Therefore, we would expect some seasonal succession in species composition and abundance in these tidepools. Whereas we attempt- ed to control for this by sampling at the same time in each year, the timing of biological events is likely to be determined by climate, which varies from year to year. Therefore, some of the variation observed be- tween years is likely due to the timing of sampling.
Another factor to be considered in comparing the assemblages among years is the effect of remov- al of fishes in previous samples on subsequent sam- ples. Other studies have found that recolonisation of defaunated tidepools is rapid relative to the time- scale of our sampling (Bussing 1972, Thomson &
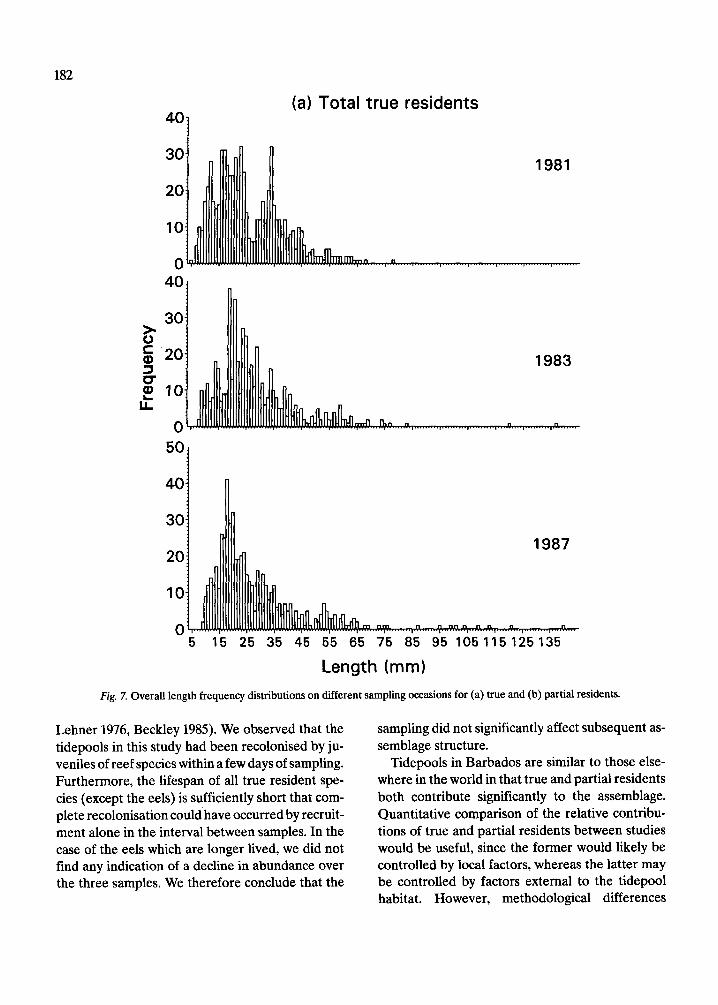
(a) Total true residents
Length (mm)
Fig. 7. Overall length frequency distributions on different sampling occasions for (a) true and (b) partial residents.
Lehner 1976, Beckley 1985). We observed that the tidepools in this study had been recolonised by ju- veniles of reef species within a few days of sampling. Furthermore, the lifespan of all true resident spe- cies (except the eels) is sufficiently short that com- plete recolonisation could have occurred by recruit- ment alone in the interval between samples. In the case of the eels which are longer lived, we did not find any indication of a decline in abundance over the three samples. We therefore conclude that the
sampling did not significantly affect subsequent as- semblage structure.
Tidepools in Barbados are similar to those else- where in the world in that true and partial residents both contribute significantly to the assemblage. Quantitative comparison of the relative contribu- tions of true and partial residents between studies would be useful, since the former would likely be controlled by local factors, whereas the latter may be controlled by factors external to the tidepool habitat. However, methodological differences
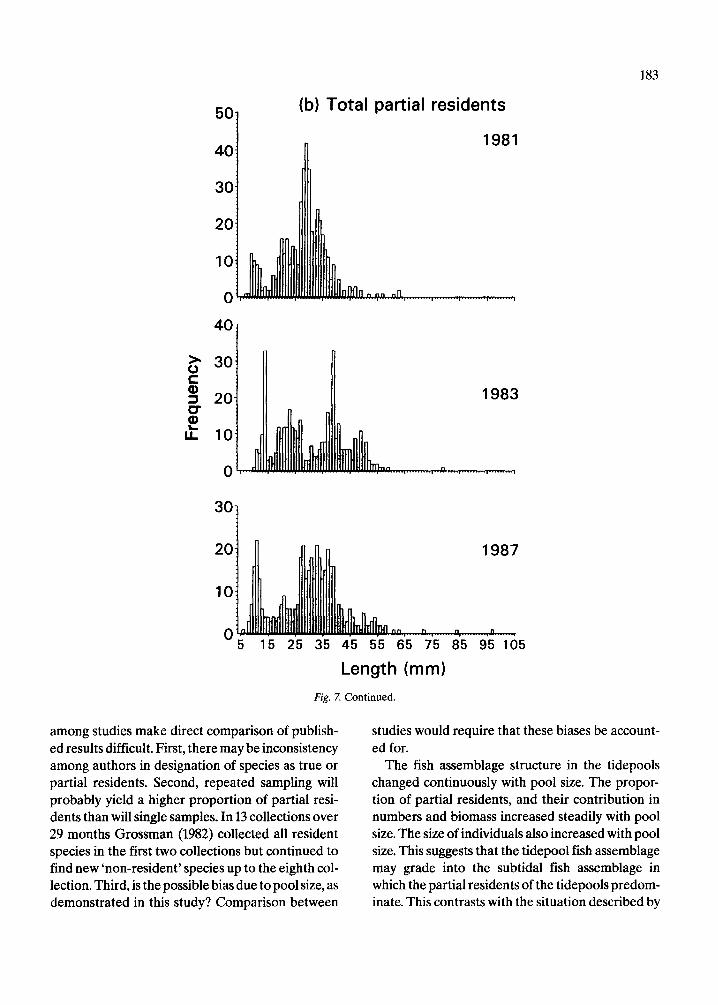
5 0 ~ (b) Total partial residents
Length (mm) Fig. 7. Continued.
among studies make direct comparison of publish- ed results difficult. First, there may be inconsistency among authors in designation of species as true or partial residents. Second, repeated sampling will probably yield a higher proportion of partial resi- dents than will single samples. In 13 collections over 29 months Grossman (1982) collected all resident species in the first two collections but continued to find new 'non-resident' species up to the eighth col- lection. Third, is the possible bias due to pool size, as demonstrated in this study? Comparison between
studies would require that these biases be account- ed for.
The fish assemblage structure in the tidepools changed continuously with pool size. The propor- tion of partial residents, and their contribution in numbers and biomass increased steadily with pool size. The size of individuals also increased with pool size. This suggests that the tidepool fish assemblage may grade into the subtidal fish assemblage in which the partial residents of the tidepools predom- inate. This contrasts with the situation described by
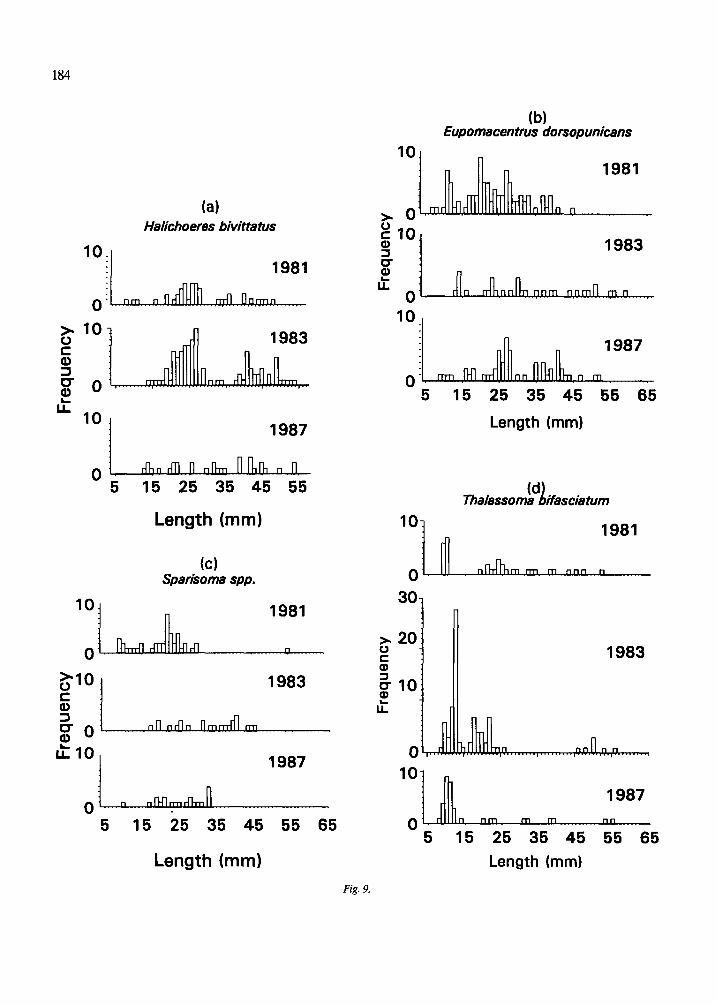
(a) Halichoeres bivittatus
Length (mm)
(c) Sparisoma spp.
(b) Eupomacentrus dorsopunicans
Length (mm)
ma,assombdli fascia tum
Length (mm) Length (mm)
Fig. 9.
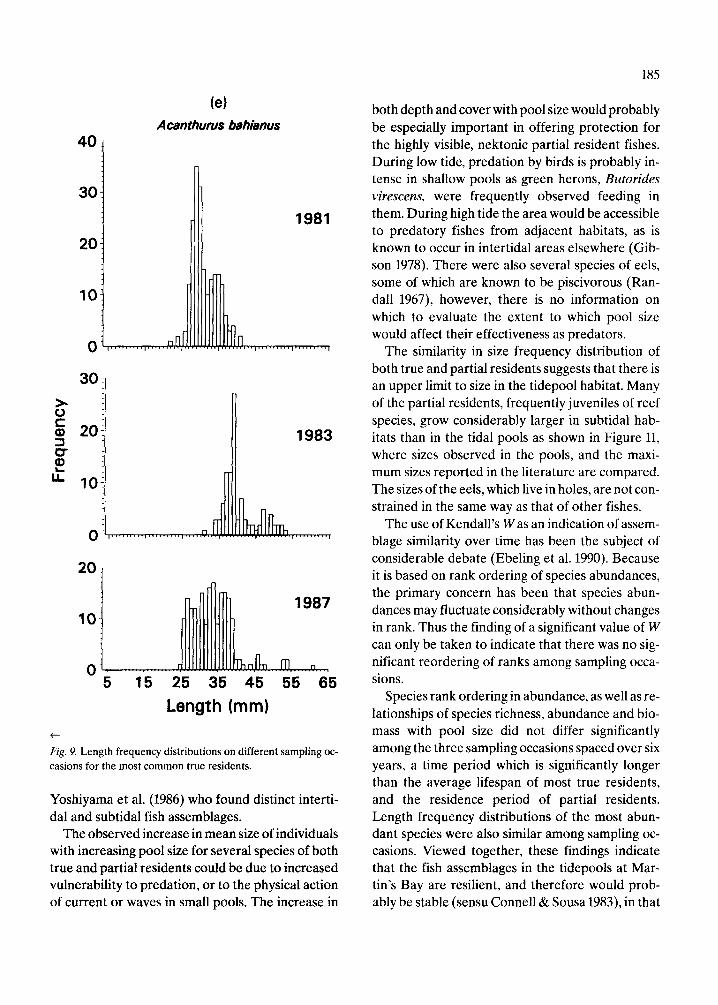
Length (mm)
Fig. 9. Length frequency distributions on different sampling oc- casions for the most common true residents.
Yoshiyama et al. (1986) who found distinct interti- dal and subtidal fish assemblages.
The observed increase in mean size of individuals with increasing pool size for several species of both true and partial residents could be due to increased vulnerability to predation, or to the physical action of current or waves in small pools. The increase in
both depth and cover with pool size would probably be especially important in offering protection for the highly visible, nektonic partial resident fishes. During low tide, predation by birds is probably in- tense in shallow pools as green herons, Butorides virescens, were frequently observed feeding in them. During high tide the area would be accessible to predatory fishes from adjacent habitats, as is known to occur in intertidal areas elsewhere (Gib- son 1978). There were also several species of eels, some of which are known to be piscivorous (Ran- dall 1967), however, there is no information on which to evaluate the extent to which pool size would affect their effectiveness as predators.
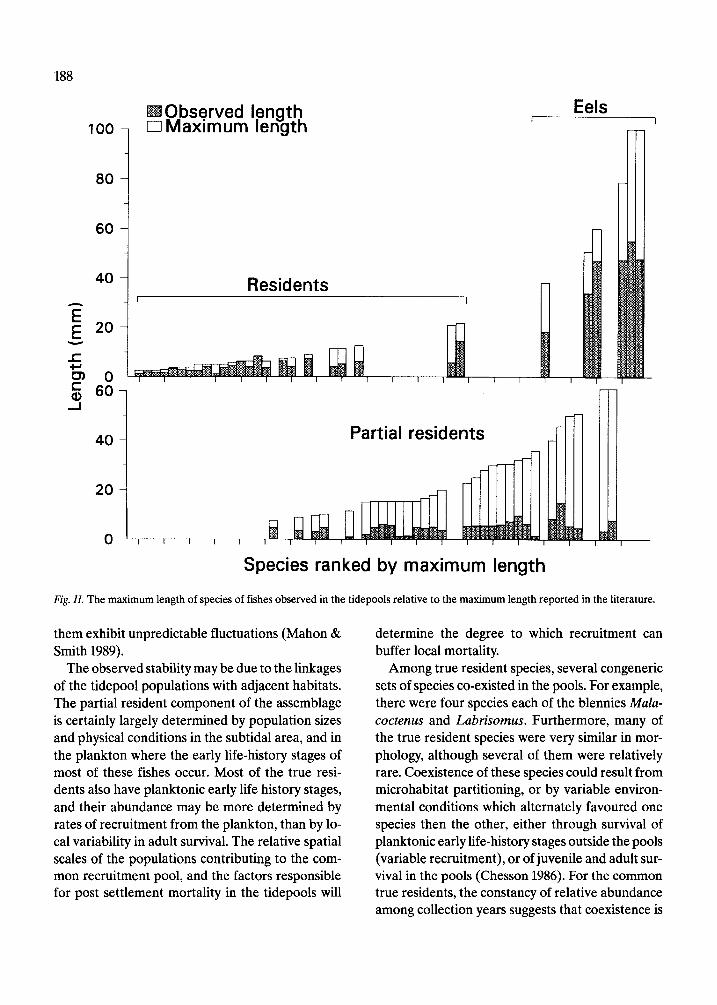
The similarity in size frequency distribution of both true and partial residents suggests that there is an upper limit to size in the tidepool habitat. Many of the partial residents, frequently juveniles of reef species, grow considerably larger in subtidal hab- itats than in the tidal pools as shown in Figure 11, where sizes observed in the pools, and the maxi- mum sizes reported in the literature are compared. The sizes of the eels, which live in holes, are not con- strained in the same way as that of other fishes.
The use of Kenda117s Was an indication of assem- blage similarity over time has been the subject of considerable debate (Ebeling et al. 1990). Because it is based on rank ordering of species abundances, the primary concern has been that species abun- dances may fluctuate considerably without changes in rank. Thus the finding of a significant value of W can only be taken to indicate that there was no sig- nificant reordering of ranks among sampling occa- sions.
Species rank ordering in abundance, as well as re- lationships of species richness, abundance and bio- mass with pool size did not differ significantly among the three sampling occasions spaced over six years, a time period which is significantly longer than the average lifespan of most true residents, and the residence period of partial residents. Length frequency distributions of the most abun- dant species were also similar among sampling oc- casions. Viewed together, these findings indicate that the fish assemblages in the tidepools at Mar- tin's Bay are resilient, and therefore would prob- ably be stable (sensu Connell & Sousa 1983), in that
(a) P. nigripinnis
Length (mm)
(c) Ogribia spp.
(b) Ginsburg ellus no vemlinea tus
Length (mm)
(d) Arcos rubiginosus
Length (mm) Length (mm) Fig. 10. Length frequency distributions on different sampling occasions for the most common partial residents.
they would remain constant over time. The results ley 1985, Collette 1988). Bennett & Griffiths (1984) also indicate that there was less stability in the par- observed stable population structure in South Afri- tial resident component of the assemblage than in can tidepools for seasonal and longer periods. the true resident component. These findings are inconsistent with the classical
Several other studies of tidepool fish assemblag- ecological view of intertidal habitats as physically es have also found them to be stable and resilient stressed and variable, such that the assemblages in (Thompson & Lehner 1976, Grossman 1982, Beck-
(el Labrisomus bucciferus
10
0
Length (mm) Length (mm)
(9) Malacoctenus gill.
Length (mm) Fig. 10. Continued.
188
100
80
60
40
20
0
-. I
BObserved length n Maximum length
Eels I I
Residents
Partial residents
Species ranked by maximum length
Fig. Il. The maximum length of species of fishes observed in the tidepools relative to the maximum length reported in the literature.
them exhibit unpredictable fluctuations (Mahon & determine the degree to which recruitment can Smith 1989). buffer local mortality.
The observed stability may be due to the linkages of the tidepool populations with adjacent habitats. The partial resident component of the assemblage is certainly largely determined by population sizes and physical conditions in the subtidal area, and in the plankton where the early life-history stages of most of these fishes occur. Most of the true resi- dents also have planktonic early life history stages, and their abundance may be more determined by rates of recruitment from the plankton, than by lo- cal variability in adult survival. The relative spatial scales of the populations contributing to the com- mon recruitment pool, and the factors responsible for post settlement mortality in the tidepools will
Among true resident species, several congeneric sets of species co-existed in the pools. For example, there were four species each of the blennies Mulu- coctenus and Labrisomus. Furthermore, many of the true resident species were very similar in mor- phology, although several of them were relatively rare. Coexistence of these species could result from microhabitat partitioning, or by variable environ- mental conditions which alternately favoured one species then the other, either through survival of planktonic early life-history stages outside the pools (variable recruitment), or of juvenile and adult sur- vival in the pools (Chesson 1986). For the common true residents, the constancy of relative abundance among collection years suggests that coexistence is

mediated by the former mechanism. The occur- rence of rare species can be accounted for by con- tinuous recolonisation of the tidepools from adja- cent habitats.
Other studies of tidepool fishes have focused on resource partitioning. Cross (1981) noted that tide- pool fishes from three assemblages appeared to be partitioning the food resource. In tidepool fishes of South Africa, Bennett et al. (1983) found consider- able resource subdivision due to vertical and hori- zontal distribution, microhabitat use and morphol- ogy. They observed that the fishes frequently fed outside of the pools when the tide was in but noted that pool habitat may control density. Therefore, further studies aimed at evaluating the mechanisms of coexistence of the assemblages of fishes in our tidepools should include their activities during the flood tide.
The observation that the tidepool fish assemblag- es contain a high proportion of partial residents which are juveniles of reef species leads to the ques- tion of their importance as nurseries and reservoirs of recruitment to the adjacent reef fish assemblag- es. One possibility is that the juveniles in tidepools are merely an overflow from a glut of juveniles con- sidered typical of reefs (Smith & Tyler 1975) and that they die there. However, it is not clear that sat- uration of reef habitats by juveniles is normal (Map- stone & Fowler 1988) and the presence of juveniles inshore suggests that they may prefer these habitats as juveniles. If, as suggested by Sale (1978), living space on the reef does become available through- out the year and is obtained by whichever individu- als are available, these inshore habitats may provide a continuous supply of recruits, and play a signif- icant role in structuring fish assemblages in adja- cent subtidal habitats (Ayal & Safriel1982).
Acknowledgements
We are especially grateful to Michele and Michael Brown for their assistance with collecting the fishes. Thanks to Alan Emery who checked our identifica- tion of fishes and to the Royal Ontario Museum where this was done. We are also grateful to Wayne
Hunte, Alan Emery, and John Green for their com- ments on an early draft of the manuscript.
References cited
Ayal, Y. & U.N. Safriel. 1982. Species diversity of the coral reef - a note on the role of predation and of adjacent habitats. Bull. Mar. Sci. 32: 787-790.
Beckley, L.E. 1985. Tide-pool fishes: recolonisation after experi- mental elimination. J. Exp. Mar. Biol. Ecol. 85: 287-295.
Bennett, B., C.L. Griffiths & M-L. Penrith. 1983. The diets of littoral fish from the Cape Peninsula. S. Afr. J. Zool. 18: 343- 352.
Bennett, B.A. & C.L. Griffiths. 1984. Factors affecting the distri- bution, abundance and diversity of rock-pool fishes on the Cape Peninsula, South Africa. S. Afr. J. Zool. 19: 97-104.
Bohlke, J.E. & C.C.G. Chaplin. 1968. Fishes of the Bahamas and adjacent tropical waters. Livingston Publ., Wynnewood. 771
PP. Brattstrom, H. 1980. Rocky shore zonation in the Santa Marta
area, Colombia. Sarsia 65: 163-226. Bussing, W.A. 1972. Recolonisation of a population of supratidal
fishes at Eniwetok Atoll, Marshall Islands. Atoll Res. Bull. 154: 1-4.
Chadwick, E.M.P. 1976. A comparison of growth and abundance for tidal pool fishes in California and British Columbia. J. Fish Biol. 8: 27-34.
Chang, K.H., S.C. Lee & W.L. Wu. 1977. Fishes of reef limestone platform at Maopitou, Taiwan: diversity and abundance. Bull. Inst. Zool., Acad. Sinica 16: 9-21.
Chesson, P.L. 1986. Environmental variation and the coexistence of species. pp. 24C-256. In: J. Diamond & T.J. Case (ed.) Com- munity Ecology, Harper & Row, New York.
Collette, B.B. 1988. Resilience of the fish assemblage in New En- gland tidepools. US Fish. Bull. 84: 200-204.
Connell, J.H. & W.P. Sousa. 1983. On the evidence needed to judge ecological stability of persistence. Amer. Nat. 121: 789- 824.
Cross, J.M. 1981. Resource partitioning in three rocky intertidal fish assemblages. pp. 142-150: In: G.M. Cailllet & C.A. Simen- stad (ed.) Gutshop '81, Washington Sea Grant. University of Washington, Seattle.
Ebeling, A.W., S.J. Holbrook & R.J. Schmitt. 1990. Temporally concordant structure of a fish assemblage: bound or deter- mined. Amer. Nat. 135: 63-73.
Fischer, E. (ed.) 1978. FA0 fishery identification sheets for fish- ery purposes western central Atlantic. Vol. I-VI. Food & Agriculture Organisation, Rome.
Gibson, R.N. 1969. The biology and behaviour of littoral fish. Oceanogr. Mar. Biol. Ann. Rev. 7: 367410.
Gibson, R.N. 1972. The vertical distribution and feeding rela- tionships of intertidal fish on the Atlantic coast of France. J. Anim. Ecol. 4: 189-207.
Gibson. R.N. 1978. Lunar and tidal rhythms in fish. pp. 201-213.

In: J. Thorpe (ed.) Rhythmic Activity of Fishes, Academic Press, New York.
Gibson, R.N. 1982. Recent studies on the biology of intertidal fishes. Oceanogr. Mar. Biol. Ann. Rev. 20: 363-414.
Green, J.M. 1971. Local distribution of Oligocottus maculosus Girard and other tidepool cottids of the West Coast of Van- couver Island, British Columbia. Can. J. Zool. 49: 1111-1128.
Grossman, G.D. 1982. Dynamics and organisation of a rocky in- tertidal fish assemblage: the persistence and resilience of taxo- cene structure. Amer. Nat. 119: 611-637.
John, D.M. & J.H. Price. 1979. The marine benthos of Antigua (Lesser Antilles). I. Environment, distribution and ecology. Bot. Mar. 22: 313-326.
King, C.A.M. 1975. Introduction to physical and biological oceanography. Edward Arnold, London. 372 pp.
Kleinbaum, D.G., L.L. Kupper & K.E. Muller. 1988. Applied re- gression analysis and other multivariable methods. PWS- Kent, Boston. 718 pp.
Lee, S.C. 1980a. Intertidal fishes of the rocky pools at Lanyu (Bo- tel Tobago), Taiwan. Bull. Inst. Zool., Acad. Sinica 19: 1-13.
Lee, S.C. 1980b. Intertidal fishes of a rocky pool of the Sanhsien- tai, eastern Taiwan. Bull. Inst. Zool., Acad. Sinica 19: 19-26.
Lewis, J.B. 1960. The fauna of rocky shore of Barbados, West Indies. Can. J. Zool. 38: 391-435.
Luckhurst, B.E. & K. Luckhurst. 1977. Recruitment patterns of coral reef fishes on the fringing reef of Curacao, Netherlands Antilles. Can. J. Zool. 55: 681-689.
Mahon, R. & R.W. Smith. 1989. Demersal fish assemblages on the Scotian Shele Northwest Atlantic: spatial distribution and persistence. Can. J. Fish. Aquat. Sci. 46 (Suppl. 1): 134452.
Mapstone, B.D. & A.J. Fowler. 1988. Recruitment and the struc- ture of assemblages of fish on coral reefs. Trends in Ecology and Evolution 3: 72-77.
Munro, J.L., V.C. Gaut, R. Thompson & P.H. Reeson. 1973. The
spawning seasons of Caribbean reef fishes. J. Fish Biol. 5: 69- 84.
Nybakken, J.W. 1982. Marine biology an ecological approach. Harper & Row Publishers, New York. 446 pp.
Powles, H. 1975. Abundance, seasonality, distribution, and as- pects of the ecology of some larval fishes off Barbados. Ph.D. Thesis, McGill University, Montreal. 227 pp.
Randall, J.E. 1967. Food habits of reef fishes of the West Indies. Stud. Trop. Oceanography 5: 665-847.
Randall, J.E. 1968. Caribbean reef fishes. TFH Publications, Neptune City. 318 pp.
Sale, P.F. 1978. Coexistence of coral reef fishes - a lottery for liv- ing space. Env. Biol. Fish. 3: 85-102.
Siegel, S. 1956. Nonparametric statistics for the behavioural sci- ences. McGraw Hill, New York. 312 pp.
Smith, C.L. & J.C. 'Qler. 1975. Succession and stability in fish communities of dome-shaped patch reefs in the West Indies. Amer. Mus. Novit. 2572: 1-18.
Southwood, T.R.E. 1978. Ecological methods, with particular reference to the study of insect populations. English Language Book Society, Cambridge. 524 pp.
Teytaud, A.R. 1971. Food habits of the goby, Ginsburgellus no- vernlineatus, and the clingfish, Arcos rubiginosus, associated with echinoids in the Virgin Islands. Carib. J. Sci. 11: 41-45.
Thompson, D.A. & C.E. Lehner. 1976. Resilience of a rocky in- tertidal fish community in a physically unstable environment. J. Exp. Mar. Biol. Ecol. 22: 1-29.
Yoshiyama, R.M. 1981. Distribution and abundance patterns of rocky intertidal fishes in central California. Env. Biol. Fish. 6: 315-332.
Yoshiyama, R.M., C. Sassaman & R.N. Lea. 1986. Rocky interti- dal fish communities of California: temporal and spatial varia- tion. Env. Biol. Fish. 17: 23-40.