structural and functional properties of batx, a new lys49 phospholipase a2 homologue isolated from...
TRANSCRIPT

a 1770 (2007) 585–593www.elsevier.com/locate/bbagen
Biochimica et Biophysica Act
Structural and functional properties of BaTX, a new Lys49 phospholipase A2
homologue isolated from the venom of the snake Bothrops alternatus
Luis Alberto Ponce-Soto a,⁎, Bruno Lomonte b, José María Gutiérrez b, Lea Rodrigues-Simioni c,José Camillo Novello a, Sergio Marangoni a
a Biochemistry Department, Institute of Biology, Medical Sciences Faculty, State University of Campinas (UNICAMP), Campinas, SP, Brazilb Instituto Clodomiro Picado, Facultad de Microbiología, Universidad de Costa Rica, San José, Costa Rica
c Pharmacology Department, Medical Sciences Faculty, State University of Campinas (UNICAMP), Campinas, SP, Brazil
Received 4 August 2006; received in revised form 20 November 2006; accepted 21 November 2006Available online 15 December 2006
Abstract
BaTX PLA2, a K49 phospholipase A2 homologue was purified from Bothrops alternatus venom after two chromatographic steps, molecularexclusion on Superdex 75 and reverse phase HPLC on μ-Bondapack C-18. A molecular mass of 13898.71 Da was determined by MALDI-TOFmass spectrometry. The amino acid composition showed that BaTX has a high content of Lys, Tyr, Gly, Pro, and 14 half-Cys residues, typical of abasic PLA2. The complete amino acid sequence of BaTX PLA2 contains 121 residues, resulting in a calculated pI value of 8.63. This sequenceshows high identity values when compared to other K49 PLA2s isolated from the venoms of viperid snakes. Lower identity is observed incomparison to D49 PLA2s. The sequence was SLFELGKMIL QETGKNPAKS YGAYYCYCGW GGQGQPKDAT DRCCYVHKCCYKKLTGCNPK KDRYSYSWKD KTIVCGENNS CLKELCECDK AVAICLRENL NTYNKKYRYY LKPLCKKADA C. In mice, BaTXinduced myonecrosis and edema, upon intramuscular or subcutaneous injections, respectively. The LD50 of BaTX was 7 μg/g body weight, byintravenous route. In vitro, the toxin caused a potent blockade of neuromuscular transmission in young chicken biventer cervicis preparations. Theblockage 50% was achieved at a concentration of 0.03 μM: 40±0.4 min and 0.07 μM: 35±0.3 min. Moreover, this protein induced a rapidcytolytic effect upon mouse skeletal muscle myoblasts in culture. Thus, the combined structural and functional information obtained identifyBaTX as a new member of the K49 PLA2 family, which presents the typical bioactivities described for such proteins.© 2006 Elsevier B.V. All rights reserved.
Keywords: Phospholipase A2; Lys-49; Neurotoxin; Myotoxin; Snake venom; Bothrops alternatus
1. Introduction
The phospholipases A2 (PLA2; EC 3.1.1.4) are hydrolyticenzymes, which cleave the sn-2 acyl bond of glycerophos-pholipids, releasing free fatty acids and lysophospholipids [1].Secretory PLA2s occur in a large variety of biological fluidssuch as pancreatic secretions, inflammatory exudates, andsnake, arthropod and mollusc venoms. To date, such PLA2shave been classified into eleven groups, on the basis of theirprimary structure and the number and position of disulfidebonds [1].
⁎ Corresponding author. Departamento de Bioquímica, Instituto de Biología,Universidade Estadual de Campinas (UNICAMP), PO Box 6109, Zip code13083-970, Campinas, SP, Brazil. Fax: +55 19 788 6129.
E-mail address: [email protected] (L.A. Ponce-Soto).
0304-4165/$ - see front matter © 2006 Elsevier B.V. All rights reserved.doi:10.1016/j.bbagen.2006.11.015
Snake venom PLA2s constitute a puzzling group of mole-cules since, despite having a similar three-dimensional structureand highly conserved molecular regions, they display a plethoraof pharmacological activities, such as myotoxic, neurotoxic,anticoagulant, hypotensive, hemolytic, platelet aggregationinhibiting, bactericidal and pro-inflammatory activities [2].This variety of pharmacological roles derives from anaccelerated micro-evolutionary process through which a highrate of amino acid substitutions has occurred in molecularregions located mainly at the surface of these molecules [3,4].
Envenomation from the bites of Bothrops is characterized bya complex series of pathophysiological alterations. Despite thefact that these venoms are mainly characterized by their localpathological effects and by systemic hemodynamic and hemo-static disturbances, some of their isolated components affect theneuromuscular junction in vitro [5–7] an effect that has received

586 L.A. Ponce-Soto et al. / Biochimica et Biophysica Acta 1770 (2007) 585–593
little attention. The venom of Bothrops alternatus has beencharacterized as having hemorrhagic [8], phosphodiesterase [9],thrombin-like [10], thrombin inhibitory [11], and myotoxic [12]activities. However, it is necessary to purify additionalcomponents responsible for the toxic effects induced by thisvenom, and to examine the potential deleterious effects inducedby such components on the neuromuscular junction in vitro.
In this work we have isolated and characterized a novelLys49 PLA2 homologue, with the following pharmacologicalactivities: in vitro neurotoxicity on young chicken biventercervicis preparations, inflammatory activities in the footpad ofmice, in vivo myotoxicity, and in vitro cytotoxicity.
2. Materials and methods
2.1. Venom and reagents
B. alternatus venom was a gift from the Batatais serpentarium, Batatais, SP,Brazil. All chemicals and reagents used in this work were of analytical orsequencing grade.
2.2. Molecular exclusion chromatography
Venom (30 mg) was dissolved in 0.2 mM ammonium bicarbonate, pH 8.0, andfractionated on a Superdex 75 column (1×60 cm), eluted with the same buffer at aconstant flow of 0.3 ml/min, on aWaters LC650E system. Elution was monitored at280 nm, and the fractions were immediately lyophilized and stored at −20 °C.
2.3. Reverse phase (RP) HPLC
Five mg of the fraction that inhibited neuromuscular junction weredissolved in 250 μl of 0.1% (v/v) trifluoroacetic acid (TFA) (solvent A). Theresulting solution was clarified by centrifugation and the supernatant wasapplied to a μ-Bondapack C-18 column (0.78 cm×30 cm; Waters 991-PDAsystem). Proteins were eluted with a linear gradient (0–100%) of 66.5% (v/v)acetonitrile in 0.1% trifluoroacetic acid (solvent B), at a flow rate of 2 ml/min.The elution profile was monitored at 280 nm and the fractions were collected,lyophilized and stored at −20 °C.
2.4. PLA2 activity
PLA2 activity was measured using the assay described by Cho and Kézdy[13] and Holzer and Mackessy [14], adapted for 96-well plates [15]. Thestandard assay mixture contained 200 μl of buffer (10 mM Tris–HCl, 10 mMCaCl2, 100 mM NaCl, pH 8.0), 20 μl of substrate, (4-nitro-3-octanoyloxy-benzoic acid), 20 μl of water, and 20 μl of BaTX in a final volume of 260 μl.After the addition of BaTX (20 μg), the mixture was incubated for up to 40 minat 37 °C, and the absorbances at 425 nm were recorded at 10 min intervals on aSpectraMax 340 multiwell plate reader (Molecular Devices, Sunnyvale, CA).Enzyme activity, expressed as velocity of reaction (V), was calculated based onthe increase in absorbance after 20 min. The assays were done in triplicate.
2.5. SDS-PAGE
Tricine SDS-PAGE in a discontinuous gel and buffer system [16] was usedto estimate the molecular mass of the proteins, under reducing and non-reducingconditions.
2.6. MALDI-TOF mass spectrometric analysis (MS)
The molecular mass of isolated BaTX was analyzed by MALDI-TOF massspectrometry using a Voyager-DE PRO MALDI-TOF apparatus (AppliedBiosystems, Foster City, CA, USA). One μl of sample in 0.1% TFAwas mixedwith 2 μl of the matrix sinapinic acid (3,5-dimethoxy-4-hydroxycinnamic acid).
The matrix was prepared with 30% acetonitrile and 0.1% TFA and its massanalyzed under the following conditions: accelerate voltage 25 kV, the laserfixed in 2890 μJ/com2, delay 300 ns, and linear analysis mode [17].
2.7. Amino acid analysis
Amino acid analysis was performed on a Pico-Tag Analyzer (WatersSystems) as described by Heinrikson and Meredith [18]. The purified BaTXsample (30 μg) was hydrolyzed at 105 °C for 24 h, in 6 M HCl (Piercesequencing grade) containing 1% phenol (w/v). The hydrolyzates were reactedwith 20 μl of derivatization solution (ethanol:triethylamine:water:phenylisothio-cyanate, 7:1:1:1, v/v) for 1 h at room temperature, after which the PTC-aminoacids were identified and quantified by HPLC, by comparing their retentiontimes and peak areas with those from a standard amino acid mixture.
2.8. Reduction and carboxymethylation of protein anddigestion of BaTX
One mg of purified BaTX was dissolved in 6 M guanidine hydrochloridecontaining 0.4 M Tris–HCl and 2 mM EDTA, pH 8.1. The protein was reducedwith dithiotreitol and then carboxymethylated with 14C iodoacetic acid [19].Desaltingwas performed on a SephadexG-25 column in 1M acetic acid at 25 °C,and the modified protein (RC-BaTX) was lyophilized. The reduced andcarboxymethylated protein was digested with Staphylococcus aureus proteaseV8 for 17 h at 37 °C using a 1:30 enzyme:substrate molar ratio, and the reactionwas stopped by lyophilization. The RC-BaTXwas also digested with Clostripainfor 8 h at 37 °C and lyophilized again. The digested products were fractionated byRP-HPLC on a C18 μ-Bondapack column, eluted with a linear acetonitrilegradient (0–100%) in 0.1% TFA.
2.9. Amino acid sequencing
N-terminal sequencing was performed on the reduced and carboxymethylatedprotein, and the sequencing of peptide peaks in repurified digestion products wasestablished using a Procise-F automatic sequencer (Applied Biosystems). Thephenylthiohydantoin in amino acids were identified by comparing their retentiontimes with that of the 20 PTH amino acid standards. Peptides containing 14C-CM-Cys were monitored with a Beckman L-250 liquid scintillation counter.
2.10. Myotoxic activity
Groups of four CD-1 mice (18–20 g) received an intramuscular (i.m.) or anintravenous (i.v.) injection of variable amounts of BaTX, in 50 μl of PBS, in thegastrocnemius. A control group received 50 μl of PBS. At different intervals, bloodwas collected from the tail into heparinized capillary tubes, and the plasma creatinekinase (CK; EC 2.7.3.2) activity was determined by a kinetic assay (Sigma 47-UV).Activity was expressed in U/l, defined as the phosphorylation of 1 μmol of NADH/min at 25 °C. All in vivo experiments were approved by the Committee for Use andCare of Research Animals (CICUA), University of Costa Rica.
2.11. Edema-forming activity
Groups of four CD-1 mice (18–20 g) received a subcutaneous (s.c.) injection ofBaTX, in 50 μl of phosphate-buffered saline (PBS; 0.12 M NaCl, 0.04 M sodiumphosphate, pH 7.2), into the right footpad. The left footpad received 50 μl of PBS,as a control. After 3 h, mice were sacrificed by CO2 inhalation, and both feet werecut off and weighed [20]. Edema was expressed as the percent increase in weight ofthe right foot, as compared to the left one, and the minimum edema-forming dose(MED) was defined as the toxin dose inducing 30% edema.
2.12. Determination of median lethal dose (LD50)
Different amounts of BaTX (100, 120, 140 and 160 μg) in 100 μl of PBSwere injected by i.v. route, in groups of five CD-1 mice (18–20 g) under lightCO2 anesthesia [21]. A control group received a PBS injection. Deaths wererecorded after 24 h, and the median lethal dose was estimated by the Spearman–Karber method [22].
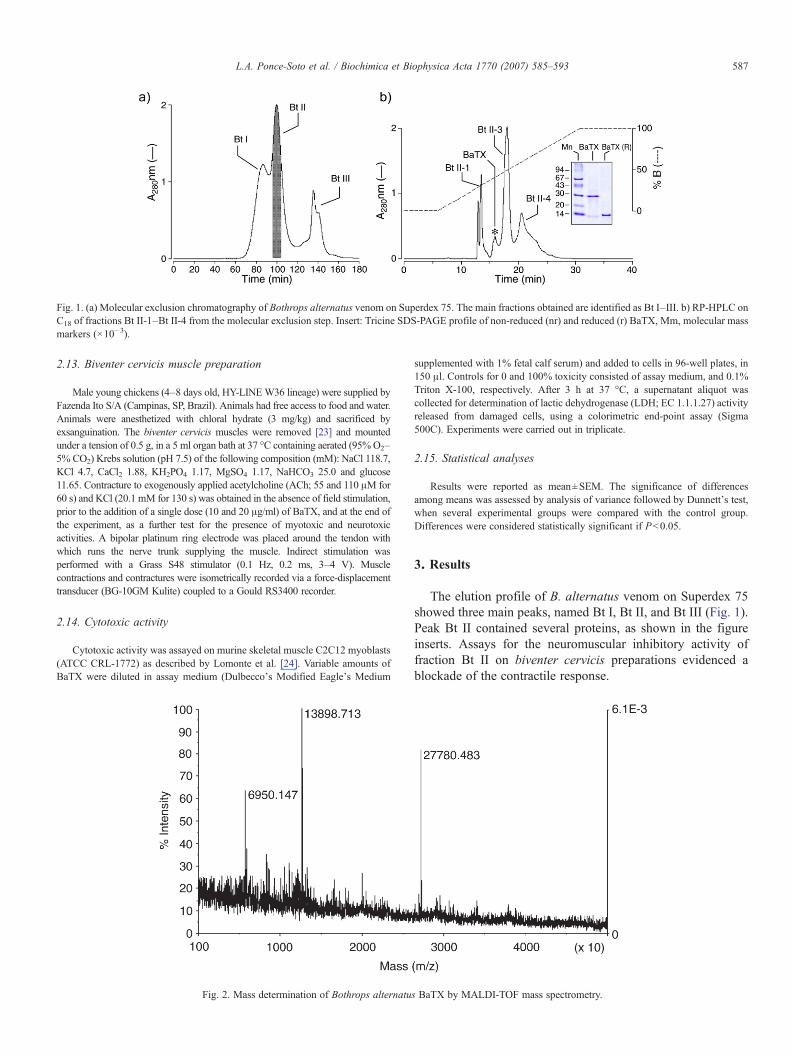
Fig. 1. (a) Molecular exclusion chromatography of Bothrops alternatus venom on Superdex 75. The main fractions obtained are identified as Bt I–III. b) RP-HPLC onC18 of fractions Bt II-1–Bt II-4 from the molecular exclusion step. Insert: Tricine SDS-PAGE profile of non-reduced (nr) and reduced (r) BaTX, Mm, molecular massmarkers (×10−3).
587L.A. Ponce-Soto et al. / Biochimica et Biophysica Acta 1770 (2007) 585–593
2.13. Biventer cervicis muscle preparation
Male young chickens (4–8 days old, HY-LINEW36 lineage) were supplied byFazenda Ito S/A (Campinas, SP, Brazil). Animals had free access to food and water.Animals were anesthetized with chloral hydrate (3 mg/kg) and sacrificed byexsanguination. The biventer cervicis muscles were removed [23] and mountedunder a tension of 0.5 g, in a 5 ml organ bath at 37 °C containing aerated (95%O2–5% CO2) Krebs solution (pH 7.5) of the following composition (mM): NaCl 118.7,KCl 4.7, CaCl2 1.88, KH2PO4 1.17, MgSO4 1.17, NaHCO3 25.0 and glucose11.65. Contracture to exogenously applied acetylcholine (ACh; 55 and 110 μM for60 s) and KCl (20.1 mM for 130 s) was obtained in the absence of field stimulation,prior to the addition of a single dose (10 and 20 μg/ml) of BaTX, and at the end ofthe experiment, as a further test for the presence of myotoxic and neurotoxicactivities. A bipolar platinum ring electrode was placed around the tendon withwhich runs the nerve trunk supplying the muscle. Indirect stimulation wasperformed with a Grass S48 stimulator (0.1 Hz, 0.2 ms, 3–4 V). Musclecontractions and contractures were isometrically recorded via a force-displacementtransducer (BG-10GM Kulite) coupled to a Gould RS3400 recorder.
2.14. Cytotoxic activity
Cytotoxic activity was assayed on murine skeletal muscle C2C12 myoblasts(ATCC CRL-1772) as described by Lomonte et al. [24]. Variable amounts ofBaTX were diluted in assay medium (Dulbecco's Modified Eagle's Medium
Fig. 2. Mass determination of Bothrops alternatu
supplemented with 1% fetal calf serum) and added to cells in 96-well plates, in150 μl. Controls for 0 and 100% toxicity consisted of assay medium, and 0.1%Triton X-100, respectively. After 3 h at 37 °C, a supernatant aliquot wascollected for determination of lactic dehydrogenase (LDH; EC 1.1.1.27) activityreleased from damaged cells, using a colorimetric end-point assay (Sigma500C). Experiments were carried out in triplicate.
2.15. Statistical analyses
Results were reported as mean±SEM. The significance of differencesamong means was assessed by analysis of variance followed by Dunnett's test,when several experimental groups were compared with the control group.Differences were considered statistically significant if P<0.05.
3. Results
The elution profile of B. alternatus venom on Superdex 75showed three main peaks, named Bt I, Bt II, and Bt III (Fig. 1).Peak Bt II contained several proteins, as shown in the figureinserts. Assays for the neuromuscular inhibitory activity offraction Bt II on biventer cervicis preparations evidenced ablockade of the contractile response.
s BaTX by MALDI-TOF mass spectrometry.
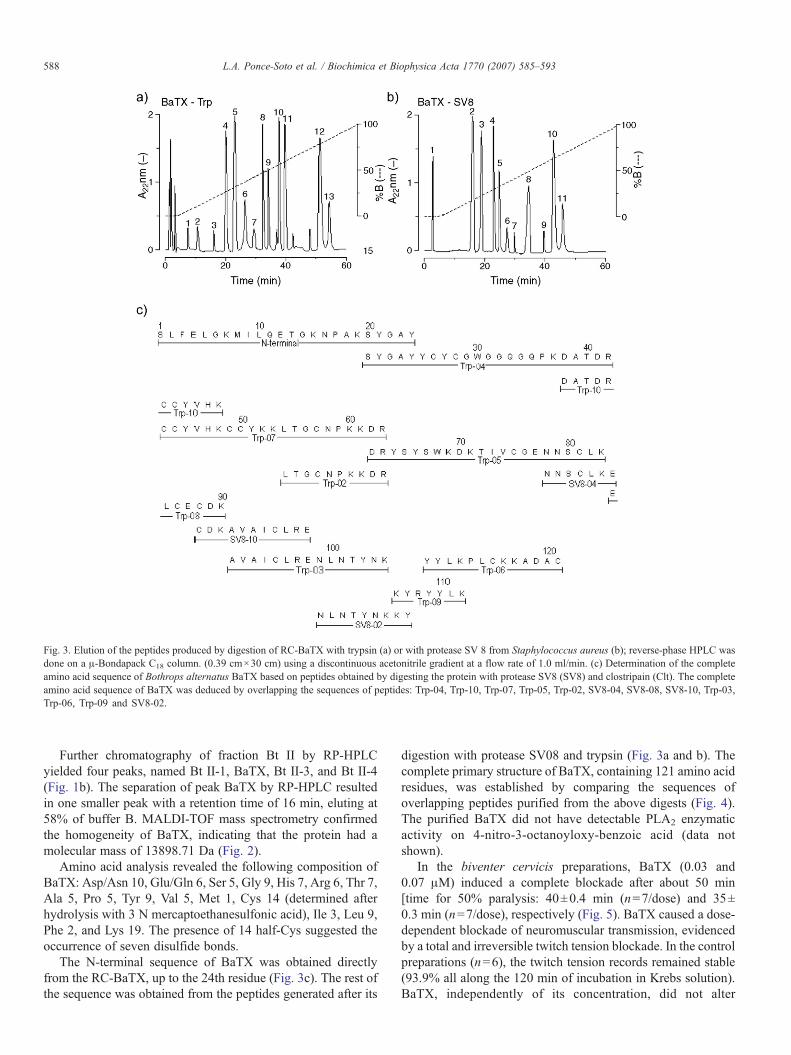
Fig. 3. Elution of the peptides produced by digestion of RC-BaTX with trypsin (a) or with protease SV 8 from Staphylococcus aureus (b); reverse-phase HPLC wasdone on a μ-Bondapack C18 column. (0.39 cm×30 cm) using a discontinuous acetonitrile gradient at a flow rate of 1.0 ml/min. (c) Determination of the completeamino acid sequence of Bothrops alternatus BaTX based on peptides obtained by digesting the protein with protease SV8 (SV8) and clostripain (Clt). The completeamino acid sequence of BaTX was deduced by overlapping the sequences of peptides: Trp-04, Trp-10, Trp-07, Trp-05, Trp-02, SV8-04, SV8-08, SV8-10, Trp-03,Trp-06, Trp-09 and SV8-02.
588 L.A. Ponce-Soto et al. / Biochimica et Biophysica Acta 1770 (2007) 585–593
Further chromatography of fraction Bt II by RP-HPLCyielded four peaks, named Bt II-1, BaTX, Bt II-3, and Bt II-4(Fig. 1b). The separation of peak BaTX by RP-HPLC resultedin one smaller peak with a retention time of 16 min, eluting at58% of buffer B. MALDI-TOF mass spectrometry confirmedthe homogeneity of BaTX, indicating that the protein had amolecular mass of 13898.71 Da (Fig. 2).
Amino acid analysis revealed the following composition ofBaTX: Asp/Asn 10, Glu/Gln 6, Ser 5, Gly 9, His 7, Arg 6, Thr 7,Ala 5, Pro 5, Tyr 9, Val 5, Met 1, Cys 14 (determined afterhydrolysis with 3 N mercaptoethanesulfonic acid), Ile 3, Leu 9,Phe 2, and Lys 19. The presence of 14 half-Cys suggested theoccurrence of seven disulfide bonds.
The N-terminal sequence of BaTX was obtained directlyfrom the RC-BaTX, up to the 24th residue (Fig. 3c). The rest ofthe sequence was obtained from the peptides generated after its
digestion with protease SV08 and trypsin (Fig. 3a and b). Thecomplete primary structure of BaTX, containing 121 amino acidresidues, was established by comparing the sequences ofoverlapping peptides purified from the above digests (Fig. 4).The purified BaTX did not have detectable PLA2 enzymaticactivity on 4-nitro-3-octanoyloxy-benzoic acid (data notshown).
In the biventer cervicis preparations, BaTX (0.03 and0.07 μM) induced a complete blockade after about 50 min[time for 50% paralysis: 40±0.4 min (n=7/dose) and 35±0.3 min (n=7/dose), respectively (Fig. 5). BaTX caused a dose-dependent blockade of neuromuscular transmission, evidencedby a total and irreversible twitch tension blockade. In the controlpreparations (n=6), the twitch tension records remained stable(93.9% all along the 120 min of incubation in Krebs solution).BaTX, independently of its concentration, did not alter

Fig. 4. Amino acid alignment of Bothrops alternatus BaTX with Lys49 PLA2 homologues. P24605: Myotoxin II from Bothrops asper [33]; Q90249: bothropstoxin Ifrom Bothrops jararacussu [54], P82287: piratoxin II from Bothrops piraja [19], Q9I834: myotoxin II from Bothrops moojen [55], P58399: piratoxin I from Bothropspirajai [56], Q9IAT9: from Bothrops neuwiedi pauloensis [57], (*)Q9PVE3: myotoxin V from Bothrops asper [58], P82114: myotoxin I from Bothrops moojeni [43],Q8UVU7: from Porthidium godmani K49 [34], P81165: myotoxin II from Cerrophidion godmani [59] P04361: from Agkistrodon p. piscivorus K49 [60], P82950:myotoxin II from Atropoides nummifer [35] and Q8UVZ7: from Crotalus atrox K49 [34]. (*)Note that Q9PVE3, submitted to GenBank by Pescatori et al. (1998), ishere referred to as Bothrops aspermyotoxin V (instead of III as in the submission), to avoid repetition with the myotoxin III protein previously characterized by Kaiseret al. [61].
589L.A. Ponce-Soto et al. / Biochimica et Biophysica Acta 1770 (2007) 585–593
significantly the ACh- and KCl-induced contractures, whencompared to the control values. In the control preparations, thecontracture to ACh and KCl was stable, after a 120 min ofindirect stimulation, when compared to the control values (datanot shown).
In vitro, BaTX rapidly lysed skeletal muscle myoblasts, witha dose of 40 μg/well (18 μM) causing 87±7% cell lysis (Fig. 6).BaTX also induced moderate footpad edema, with a MED of
8±3 μg, evidencing the local increase in vascular permeability.Edema reached its highest point after 3 h, rapidly returning tobasal levels thereafter (Fig. 7).
In vivo, BaTX induced a conspicuous local myotoxic effectwhen injected by the i.m. route, but no increase in CK plasmalevels occurred after its i.v. injection. Time-course analysisshowed a maximum increase in plasma CK 2 h after i.m.injection, returning to normal by 24 h (Fig. 8). The lethal activity
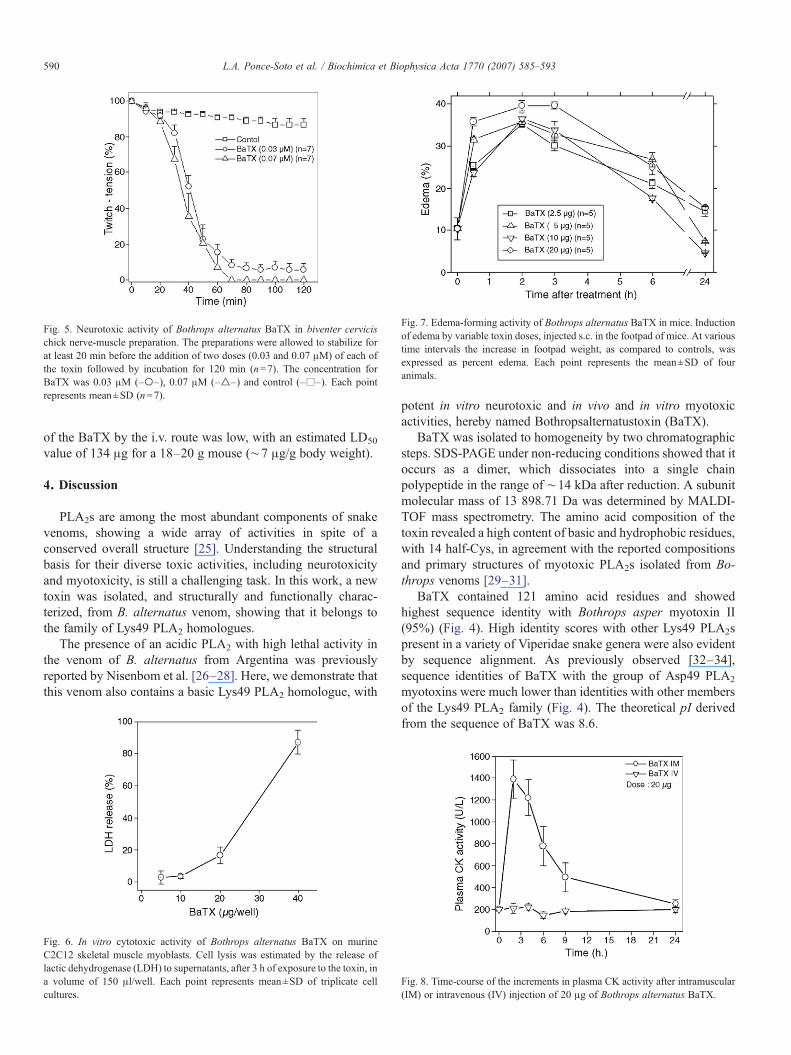
Fig. 5. Neurotoxic activity of Bothrops alternatus BaTX in biventer cervicischick nerve-muscle preparation. The preparations were allowed to stabilize forat least 20 min before the addition of two doses (0.03 and 0.07 μM) of each ofthe toxin followed by incubation for 120 min (n=7). The concentration forBaTX was 0.03 μM (–○–), 0.07 μM (–△–) and control (–□–). Each pointrepresents mean±SD (n=7).
Fig. 7. Edema-forming activity of Bothrops alternatus BaTX in mice. Inductionof edema by variable toxin doses, injected s.c. in the footpad of mice. At varioustime intervals the increase in footpad weight, as compared to controls, wasexpressed as percent edema. Each point represents the mean±SD of fouranimals.
590 L.A. Ponce-Soto et al. / Biochimica et Biophysica Acta 1770 (2007) 585–593
of the BaTX by the i.v. route was low, with an estimated LD50
value of 134 μg for a 18–20 g mouse (∼7 μg/g body weight).
4. Discussion
PLA2s are among the most abundant components of snakevenoms, showing a wide array of activities in spite of aconserved overall structure [25]. Understanding the structuralbasis for their diverse toxic activities, including neurotoxicityand myotoxicity, is still a challenging task. In this work, a newtoxin was isolated, and structurally and functionally charac-terized, from B. alternatus venom, showing that it belongs tothe family of Lys49 PLA2 homologues.
The presence of an acidic PLA2 with high lethal activity inthe venom of B. alternatus from Argentina was previouslyreported by Nisenbom et al. [26–28]. Here, we demonstrate thatthis venom also contains a basic Lys49 PLA2 homologue, with
Fig. 6. In vitro cytotoxic activity of Bothrops alternatus BaTX on murineC2C12 skeletal muscle myoblasts. Cell lysis was estimated by the release oflactic dehydrogenase (LDH) to supernatants, after 3 h of exposure to the toxin, ina volume of 150 μl/well. Each point represents mean±SD of triplicate cellcultures.
potent in vitro neurotoxic and in vivo and in vitro myotoxicactivities, hereby named Bothropsalternatustoxin (BaTX).
BaTX was isolated to homogeneity by two chromatographicsteps. SDS-PAGE under non-reducing conditions showed that itoccurs as a dimer, which dissociates into a single chainpolypeptide in the range of ∼14 kDa after reduction. A subunitmolecular mass of 13 898.71 Da was determined by MALDI-TOF mass spectrometry. The amino acid composition of thetoxin revealed a high content of basic and hydrophobic residues,with 14 half-Cys, in agreement with the reported compositionsand primary structures of myotoxic PLA2s isolated from Bo-throps venoms [29–31].
BaTX contained 121 amino acid residues and showedhighest sequence identity with Bothrops asper myotoxin II(95%) (Fig. 4). High identity scores with other Lys49 PLA2spresent in a variety of Viperidae snake genera were also evidentby sequence alignment. As previously observed [32–34],sequence identities of BaTX with the group of Asp49 PLA2
myotoxins were much lower than identities with other membersof the Lys49 PLA2 family (Fig. 4). The theoretical pI derivedfrom the sequence of BaTX was 8.6.
Fig. 8. Time-course of the increments in plasma CK activity after intramuscular(IM) or intravenous (IV) injection of 20 μg of Bothrops alternatus BaTX.

591L.A. Ponce-Soto et al. / Biochimica et Biophysica Acta 1770 (2007) 585–593
Primary structures of a number of Lys49 PLA2 homologueshave been determined by both protein and DNA sequencing, and arecent review on this protein family compiled at least 24 completeamino acid sequences [7]. The Lys49 PLA2 homologues presentchanges in residues that are involved in Ca2+-binding in the Asp49PLA2s: instead of the conserved Tyr28, Gly32 and Asp49, theyhave Asn28, Leu32 and Lys49 [35]. These substitutions have beenrelated to their lack of enzymatic activity, since Ca2+ is an essentialcofactor stabilizing the tetrahedral intermediate in the catalyticmechanism of PLA2s. Interestingly, the Ca+2-binding loop ofBaTX does not present the usual Asn28 and Leu32 substitutions ofLys49 PLA2-like proteins, but maintains both the Tyr28 and Gly32of the Asp49 PLA2 counterparts. Nevertheless, BaTX wascatalytically inactive in our assay. Residues forming the catalyticnetwork (His48, Tyr52, Tyr73, and Asp99) in enzymatically-activePLA2s are also intact in BaTX. Therefore, the lack of enzymaticactivity of BaTX cannot be attributed to an altered Ca+2-bindingloop, or to changes in the catalytic network residues. It remains tobe determined whether the Lys49 substitution alone is sufficient toabolish catalysis in this protein, as previously demonstrated insingle mutants of the porcine and bovine pancreatic PLA2s[36,37], and/or whether the presence of Lys122may also be relatedto this phenomenon, as proposed for piratoxin II [38].
Most of the Lys49 PLA2 homologues isolated to date formnon-covalent homodimers in solution, and the triad Glu12/Trp77/Lys80 has been described as important for dimerstabilization [36]. In agreement with these principles, BaTXconserves this amino acid triad, and forms a stable dimer.
The myotoxic activity of Lys49 PLA2 homologues is knownto depend on the membrane-destabilizing action of theirC-terminal region, as evidenced by studies using short syntheticpeptides [7] and site-directed mutagenesis [39]. BaTX sharescomplete identity at the C-terminal region 115–129 withmyotoxin II from B. asper, and this sequence (KKYR-YYLKPLCKK) has been shown to be central for the myotoxicmechanism of the latter toxin [7,40]. Given the high sequenceidentity between both proteins (95%), it is likely that BaTX andB. asper myotoxin II share a common mechanism in theinduction of muscle damage. On the other hand, BaTX wasshown in this study to exert a potent neuromuscular blockingeffect in vitro, an activity that has received little attention in thestudy of Lys49 PLA2 homologues, and for which no structuraldeterminants of action have been delineated yet. It will be ofinterest to determine which protein regions are involved in theneuromuscular blocking activity of BaTX, which appears to bemore potent than that described for other Lys49 PLA2
homologues [41–47].The conspicuous in vitro neuromuscular inhibitory effect
induced by BaTX, and by other Lys49 PLA2 homologues,contrasts with their low lethal potencies when administered i.v.or i.p. [29]. Such low lethal activity has been also described forAsp49 PLA2 myotoxins from Bothrops sp venoms [29]. Thus, itis evident that Bothrops myotoxic PLA2s, in contrast withseveral elapid and crotalid myotoxic and neurotoxic PLA2s[48], are unable to significantly affect neuromuscular junctionsin vivo upon i.v. or i.p. injections. This apparently puzzlingfinding may be related to the observation that Bothrops sp
myotoxic Lys49 PLA2 homologues are able to induce drasticlocal myotoxicity, upon i.m. injection, but are devoid ofsystemic myotoxic activity, as reflected by lack of increment inCK plasma activity after i.v. injection. The reasons behind suchlack of systemic neurotoxicity and myotoxicity may be relatedto a non-specific binding of these toxins to various tissuetargets, thus precluding their targeting to relevant pharmacolo-gical sites in distant nerve terminals or muscle cells. Thishypothesis has been presented to differentiate between locally-and systemically-acting myotoxic PLA2s [6]. In contrast withBothrops sp myotoxic PLA2s, systemically-acting PLA2s, suchas various elapid neurotoxic/myotoxic PLA2s and the crotaliddimeric PLA2 complex crotoxin, are able to bind targets inmuscle cell plasma membrane in a more specific fashion,therefore not being sequestered by irrelevant non-specificbinding to various cell types. Thus, this mechanism to explainthe lack of systemic myotoxicity in Bothrops sp Lys49 PLA2
homologues may be also applied to interpret the lack of in vivosystemic neurotoxicity of these proteins. This hypothesisremains open for future experimental testing.
In addition to in vitro neuromuscular blocking activity and invivo myotoxicity, BaTX induced cytotoxicity on myoblasts inculture and footpad edema in mice. These two activities havebeen previously described for other Lys49 PLA2 homologues[22,29]. Owing to the lack of catalytic activity of Lys49 PLA2
homologues, it is evident that the mechanism of suchtoxicological/pharmacological effects does not depend on directphospholipids hydrolysis, but instead on the action of differentmechanisms exerted by molecular regions distinct from thecatalytic site. Moreover, these proteins have been shown toinduce various other effects, such as bactericidal activity [49],apoptosis in a lymphoblastoid cell line [50], diverse pro-inflammatory activities [51], and blockade of HIV entry intocells [52]. It would be relevant to assess whether BaTX alsoexerts this variety of activities.
Acknowledgments
The authors thank Vera Lucia Bonfim, Paulo A. Baldassoand Marino Forner for general technical help. This work wassupported by FAPESP and is part of a PhD thesis by LuisAlberto Ponce-Soto.
References
[1] D.A. Six, E.A. Dennis, The expanding superfamily of phospholipase A2
enzymes: classification and characterization, Biochim. Biophys. Acta 1488(2000) 1–19.
[2] R.M. Kini, Venom Phospholipase A2 Enzymes: Structure, Function andMechanism, Wiley, Chichester, England, 1997, pp. 1–511.
[3] R.M. Kini, Y.M. Chan, Accelerated evolution and molecular surfaceof venom phospholipase A2 enzymes, J. Mol. Evol. 48 (2) (1999)125–132.
[4] D. Kordis, F. Gubensek, Bov-B long interspersed repeated DNA (LINE)sequences are present in Vipera ammodytes phospholipase A2 genesand in genomes of Viperidae snakes, Eur. J. Biochem. 246 (3) (1997)772–779.
[5] R.M. Kini, Excitement ahead: structure, function and mechanism of snakevenom phospholipase A2 enzymes, Toxicon 42 (2003) 827–840.

592 L.A. Ponce-Soto et al. / Biochimica et Biophysica Acta 1770 (2007) 585–593
[6] J.M. Gutiérrez, C.L. Ownby, Skeletal muscle degeneration induced byvenom phospholipases A2: insights into the mechanisms of local andsystemic myotoxicity, Toxicon 42 (2003) 915–931.
[7] B. Lomonte, Y. Angulo, L. Calderón, An overview of Lysine-49phospholipase A2 myotoxins from crotalid snake venoms and theirstructural determinants of myotoxic action, Toxicon 42 (2003)885–901.
[8] A.R. de Roodt, S.I. García, O.D. Salomon, L. Segre, J.A. Dolab, R.F.Funes, E.H. de Titto, Hemorrhagic activity of Bothrops venomsdetermined by two different methods and relationship with proteolyticactivity on gelatin and lethality, Toxicon 41 (8) (2003) 949–958.
[9] A.A. Valerio, A.C. Corradini, P.C. Panunto, S.M. Mello, S. Hyslop,Purification and characterization of a phosphodiesterase from Bothropsalternatus snake venom, J. Protein Chem. 21 (8) (2002) 95–503.
[10] M.B. Smolka, S. Marangoni, B. Oliveira, J.C. Novello, Purification andpartial characterization of a thrombin-like enzyme, balterobin, from thevenom of Bothrops alternatus, Toxicon 36 (1998) 1059–1063.
[11] H.C. Castro, L.S. Dutra, A.L. Oliveira-Carvalho, R.B. Zingali, Bothroal-ternin, a thrombin inhibitor from the venom of Bothrops alternatus,Toxicon 36 (12) (1998) 1903–1912.
[12] O. Acosta de Pérez, P. Koscinczuk, M.S. Negrette, P. Teibler, R. Ruiz,Effects of Bothrops alternatus venom from Argentina on muscle anddifferent organs in mice, Acta Physiol. Pharmacol. Ther. Latinoam. 46(1996) 97–102 (Spanish).
[13] W. Cho, F.J. Kezdy, Chromogenic substrates and assay of phospholipasesA2, Methods Enzymol. 197 (1991) 75–79.
[14] M. Holzer, S.P. Mackessy, An aqueous endpoint assay of snake venomphospholipase A2, Toxicon 34 (1996) 1149–1155.
[15] L.A. Ponce-Soto, M.H. Toyama, S. Hyslop, J.C. Novello, S. Marangoni,Isolation and preliminary enzymatic characterization of a novel PLA2 fromCrotalus durissus collilineatus venom, J. Protein Chem. 21 (3) (2002)131–136.
[16] H. Schägger, G. von Jagow, Tricine-sodium dodecyl sulfate-polyacry-lamide gel electrophoresis for the separation of proteins in the rangefrom 1 to 100 kDa, Anal. Biochem. 166 (2) (1987) 368–379.
[17] M.B. Smolka, H. Zhou, S. Purkayastha, R. Aebersold, Optimization of theisotope-coded affinity tag-labeling procedure for quantitative proteomeanalysis, Anal. Biochem. 297 (2001) 25–31.
[18] R.L. Heinrikson, S.C. Meredith, Amino acid analysis by reverse-phasehigh-performance liquid chromatography: precolumn derivatization withphenylisothiocyanate, Anal. Biochem. 136 (1) (1984) 65–74.
[19] M.H. Toyama, A.M. Soares, L. Wen-Hwa, I. Polikarpov, J.R. Giglio, S.Marangoni, Amino acid sequence of piratoxin-II, a myotoxic lys49phospholipase A2 homologue from Bothrops pirajai venom, Biochimie 82(2000) 245–250.
[20] M. Yamakawa, M. Nozaki, Z. Hokama, Fractionation of Sakishima-habu(Trimeresurus elegans) venom, and lethal, hemorrhagic, and edema-forming activities of the fractions, in: A. Ohsaka, K. Hayashi, Y. Sawai(Eds.), Animal, Plant and Microbial Toxins, Plenum Press, New York,1976, pp. 97–109.
[21] J.M. Gutiérrez, B. Lomonte, F. Chaves, E. Moreno, L. Cerdas,Pharmacological activities of a toxic phospholipase A isolated from thevenom of the snake Bothrops asper, Comp. Biochem. Physiol. 84C (1986)159–164.
[22] World Health Organization, Progress in the characterization of venoms andstandardization of antivenoms, WHO Offset Publication, vol. 58, 1981Geneva.
[23] B.L. Ginsborg, J. Warriner, The isolated chick biventer cervicisnerve-muscle preparation, Br. J. Pharmacol. Chemother. 15 (1960)410–4111.
[24] B. Lomonte, Y. Angulo, S. Rufini, W. Cho, J.R. Giglio, M. Ohno, J.J.Daniele, P. Geoghegan, J.M. Gutiérrez, Comparative study of thecytolytic activity of myotoxic phospholipases A2 on mouse endothelial(tEnd) and skeletal muscle (C2C12) cells in vitro, Toxicon 37 (1999)145–158.
[25] E. Valentin, G. Lambeau, What can venom phospholipases A2 tell us aboutthe functional diversity of mammalian secreted phospholipases A2,Biochimie 82 (2000) 815–831.
[26] H.E. Nisenbom, J.C. Perazzo, A.J. Monserrat, J.C. Vidal, Contribution ofphospholipase A2 to the lethal potency of Bothrops alternatus (víbora de lacruz) venom, Toxicon 24 (1986) 807–817.
[27] H.E. Nisenbom, C. Seki, J.C. Vidal, Phospholipase A2 from Bothropsalternatus (víbora de la cruz) venom. Purification and some characteristicproperties, Toxicon 24 (3) (1986) 259–272.
[28] H.E. Nisenbom, J.C. Perazzo, A.J. Monserrat, J.C. Vidal, Effect ofchemical modification with p-bromophenacyl bromide on the enzymaticand lethal properties of phospholipase A2 from Bothrops alternatus(víbora de la cruz) venom, Toxicon 26 (12) (1988) 1137–1144.
[29] J.M. Gutiérrez, B. Lomonte, Phospholipase A2 myotoxins from Bothropssnake venoms, Toxicon 33 (11) (1995) 1405–1424.
[30] J.M. Gutiérrez, B. Lomonte, Phospholipase A2 myotoxins from Bothropssnake venoms, in: R.M. Kini (Ed.), Venom Phospholipase A2 Enzymes,Structure, Function and Mechanism, John Wiley, New York, 1997,pp. 321–352.
[31] H.S. Selistre de Araujo, S.P. White, C.L. Ownby, Sequence analysis ofLys49 phospholipase A2 myotoxins: a highly conserved class of proteins,Toxicon 34 (11–12) (1996) 1237–1242.
[32] B. Francis, J.M. Gutiérrez, B. Lomonte, I.I. Kaiser, Myotoxin II fromBothrops asper (Terciopelo) venom is a lysine-49 phospholipase A2,Arch. Biochem. Biophys. 284 (1991) 352–359.
[33] I.H. Tsai, Y.H. Chen, Y.M. Wang, M.C. Tu, A.T. Tu, Purification,sequencing, and phylogenetic analyses of novel Lys-49 phospholipases A2
from the venoms of rattlesnakes and other pit vipers, Arch. Biochem.Biophys. 394 (2001) 236–244.
[34] Y. Angulo, T. Olamendi-Portugal, A. Alape-Girón, L.D. Possani, B.Lomonte, Structural characterization and phylogenetic relationships ofmyotoxin II from Atropoides (Bothrops) nummifer snake venom, a Lys49phospholipase A2 homologue, Int. J. Biochem. Cell Biol. 34 (2002)1268–1278.
[35] R.K. Arni, R.J. Ward, Phospholipase A2: a structural review, Toxicon 34(1996) 827–841.
[36] C.J. van den Bergh, A.J. Slotboom, H.M. Verheij, G.H. de Haas, The roleof aspartic acid-49 in the active site of phospholipase A2. A site-specificmutagenesis study of porcine pancreatic phospholipase A2 and therationale of the enzymatic activity of [lysine49] phospholipase A2 fromAgkistrodon piscivorus piscivorus venom, Eur. J. Biochem. 176 (1988)353–357.
[37] Y. Li, B. Yu, H. Zhu, M. Jain, M. Tsai, Phospholipase A2 engineering.Structural and functional roles of the highly conserved active site residueaspartate-49, Biochemistry 33 (1994) 14714–14722.
[38] W.H. Lee, M.T. da Silva Giotto, S. Marangoni, M.H. Toyama, I.Polikarpov, R.C Garrat, Structural basis for low catalytic activity inLys49 phospholipases A2—A hypothesis: the crystal structure of piratoxinII complexed to fatty acid, Biochemistry 40 (2001) 28–36.
[39] L. Chioato, R.J. Ward, Mapping structural determinants of biologicalactivities in snake venom phospholipases A2 by sequence analysis and sitedirected mutagenesis, Toxicon 42 (2003) 869–883.
[40] B. Lomonte, E. Moreno, A. Tarkowski, L.A. Hanson, M. Maccarana,Neutralizing interaction between heparins and myotoxin II, a Lys-49phospholipase A2 from Bothrops asper snake venom. Identification of aheparin-binding and cytolytic toxin region by the use of synthetic peptidesand molecular modeling, J. Biol. Chem. 269 (1994) 29867–29873.
[41] D.S. Dhillon, E. Condrea, J.M. Maraganore, R.L. Heinrikson, S.Benjamin, P. Rosenberg, Comparison of enzymatic and pharmacologicalactivities of lysine-49 and aspartate-49 phospholipases A2 from Agkis-trodon piscivorus piscivorus snake venom, Biochem. Pharmacol. 36(1987) 1723–1730.
[42] A.M. Soares, R. Guerra-Sá, C.R. Borja-Oliveira, V.M. Rodrigues, L.Rodrigues-Simioni, V. Rodrigues, M.R. Fontes, B. Lomonte, J.M.Gutiérrez, J.R. Giglio, Structural and functional characterization ofBnSP-7, a Lys49 myotoxic Phospholipase A2 homologue from Bothropsneuwiedi pauloensis venom, Arch. Biochem. Biophys. 378 (2000)201–209.
[43] Y. Oshima-Franco, G.B. Leite, G.H. Silva, D.F. Cardoso, S. Hyslop, J.R.Giglio, M.A. da Cruz-Hofling, L. Rodrigues-Simioni, Neutralization of thepharmacological effects of bothropstoxin-I from Bothrops jararacussu

593L.A. Ponce-Soto et al. / Biochimica et Biophysica Acta 1770 (2007) 585–593
(jararacucu) venom by crotoxin antiserum and heparin, Toxicon 39 (2001)1477–1485.
[44] Yoko Oshima-Franco, G.B. Leite, C.A. Dal Belo, S. Hyslop, J. Prado-Franceschi, J.R. A.C.O.Cintra, M.A. Giglio, L. da Cruz-Höfling, Thepresynaptic activity of bothropstoxin-I, a myotoxin from Bothropsjararacussu snake venom, Basic. Clin. Pharmacol. Toxicol. 95 (4) (2004)175–182.
[45] C.R. Borja-Oliveira, A.M. Durigon, AC. Vallin, M.H. Toyama, C. Souccar, S.Marangoni, L. Rodrigues-Simioni, The pharmacological effect of Bothropsneuwiedii pauloensis (jararaca-pintada) snake venom on avian neuromusculartransmission, Braz. J. Med. Biol. Res. 36 (5) (2003) 617–624.
[46] V.L. Bonfim, M.H. Toyama, J.C. Novello, S. Hyslop, C.R.B. Oliveira, L.Rodrigues-Simioni, S. Marangoni, Isolation and enzymatic characteriza-tion of a basic phospholipase A2 from Bothrops jararacussu snake venom,J. Protein Chem. 20 (2001) 239–245.
[47] W.L. Cavalcante, M.D. Silva, M. Gallacci, Influence of temperature uponparalyzing and myotoxic effects of bothropstoxin-I on mouse neuromus-cular preparations, Chem.-Biol. Interact. 15 (151 (2)) (2005) 95–100.
[48] P. Rosenberg, Lethal potency of snake venom phospholipase A2 enzymes.Pitfalls to avoid in the study of correlations between enzymatic activity andpharmacological properties of phospholipase A2 enzymes, in: R.M. Kini(Ed.), Venom Phospholipase A2 Enzymes: Structure, Function andMechanism, Wiley, New York, 1997, pp. 129–154.
[49] L. Páramo, B. Lomonte, J. Pizarro-Cerdá, J.A. Bengoechea, J.P. Gorvel, E.Moreno, Bactericidal activity of Lys49 and Asp49 myotoxic phospholipasesA2 from Bothrops asper snake venom. Synthetic Lys49 myotoxin II-(115–129)-peptide identifies its bactericidal region, Eur. J. Biochem. 253(1998) 452–461.
[50] R. Mora, B. Valverde, C. Díaz, B. Lomonte, J.M. Gutiérrez, A Lys-49phospholipase A2 homologue from Bothrops asper snake venom inducesproliferation, apoptosis and necrosis in a lymphoblastoid cell line, Toxicon45 (2005) 651–660.
[51] J.P. Zuliani, C.M. Fernandes, S.R. Zamuner, J.M. Gutiérrez, C.F.P.Teixeira, Inflammatory events induced by Lys-49 and Asp-49 phospho-
lipases A2 isolated from Bothrops asper snake venom: role of catalyticactivity, Toxicon. 45 (2005) 335–346.
[52] D. Fenard, G. Lambeau, E. Valentin, J.C. Lefebvre, M. Lazdunski, A.Doglio, Secreted phospholipases A2, a new class of HIV inhibitors thatblock virus entry into host cells, J. Clin. Invest. 104 (1999) 611–618.
[54] A.C.O. Cintra, S. Marangoni, B. Oliveira, J.R. Giglio, Bothropstoxin-I:amino acid sequence and function, J. Protein Chem. 12 (1993) 57–64.
[55] A.M. Soares, V.M. Rodrigues, M.I. Homsi-Brandeburgo, M.H. Toyama,F.R. Lombardi, R.K. Arni, J.R. Giglio, A rapid procedure for theisolation of the Lys-49 myotoxin II from Bothrops moojeni (caissaca)venom: biochemical characterization, crystallization, myotoxic andedematogenic activity, Toxicon 36 (1998) 503–514.
[56] M.H. Toyama, A.M. Soares, C.A Vieira, J.C. Novello, B. Oliveira, J.R.Giglio, S. Marangoni, Amino acid sequence of piratoxin-I, a myotoxinfrom Bothrops pirajai snake venom and its biological activity afteralkylation with p-bromophenacyl bromide, J. Protein Chem. 17 (1998)713–728.
[57] V.M. Rodrigues, A.M. Soares, A.C. Mancin, M.R.M. Fontes, M.I. Homsi-Brandeburgo, J.R. Giglio, Geographic variations in the composition ofmyotoxins from Bothrops neuwiedi snake venoms: biochemical characteriza-tion and biological activity, Comp. Biochem. Physiol. 121A (1998) 215–222.
[58] M. Pescatori, A. Grasso, S. Rifini, Molecular cloning of a K-49 PLA2-likemyotoxin from the snake Botrops asper. Biology, University of Torvergata, via della ricerca scientifica. (1998) Roma 00100. Italy.
[59] M.V. de Sousa, L. Morhy, R.K. Arni, R.J. Ward, C. Díaz, J.M. Gutiérrez,Amino acid sequence of a myotoxic Lys49-phospholipase A2 homologuefrom the venom of Cerrophidion (Bothrops) godmani, Biochim. Biophys.Acta 1384 (2) (1998) 204–208.
[60] J.M. Maraganore, R.L. Heirikson, The lysine-49 phospholipase A2 fromthe venom of Agkistrodon piscivorus. Relation of structure and function toother phospholipases A2, J. Biol. Chem. 261 (1986) 4797–4804.
[61] I.I. Kaiser, J.M. Gutiérrez, D. Pulmmer, S.D. Aird, G.V. Odell, The aminoacid sequence of a myotoxic phospholipases from the venom of Bothropsasper Arch, Biochem. Biophys. 278 (1990) 319–325.